Submitted:
26 May 2026
Posted:
27 May 2026
You are already at the latest version
Abstract
Hantaviruses are emerging rodent-borne pathogens that pose increasing global public health concerns due to their association with hemorrhagic fever with renal syndrome (HFRS) and hantavirus cardiopulmonary syndrome (HCPS), both of which can result in substantial morbidity and mortality. Environmental change, climate variability, urbanization, and land-use transformation are increasingly recognized as critical drivers of hantavirus emergence and transmission. This review summarizes current evidence regarding hantavirus virology, epidemiology, pathogenesis, clinical manifestations, diagnostics, surveillance systems, prevention strategies, and One Health preparedness approaches. Emphasis is placed on the influence of climate change and ecological disruption on rodent reservoir dynamics and spillover risk, as well as major surveillance and diagnostic gaps in tropical and Caribbean regions where hantavirus circulation may be underrecognized. Advances in molecular diagnostics, genomic surveillance, vaccine development, monoclonal antibody therapies, and climate-based early warning systems are also discussed. Existing evidence highlights the importance of integrated One Health surveillance systems that combine human, animal, and environmental monitoring to improve early detection and outbreak preparedness. Strengthening laboratory capacity, ecological surveillance, regional collaboration, and public health infrastructure will be essential for reducing the global burden of hantavirus infections and improving preparedness for future zoonotic disease threats.
Keywords:
hantavirus
; HFRS
; HCPS
; virology
; disease surveillance
; emerging infectious diseases
; zoonoses
; rodent-borne pathogens
; climate change
; pathogenesis
; One Health
1. Introduction
The emergence and re-emergence of zoonotic infectious diseases continue to threaten global health, specifically in the setting of climate change, ecological disruption, urbanization, and increasing human mobility [1]. Among these pathogens, hantaviruses are an important group of rodent-borne viruses belonging to the family Hantaviridae that can cause severe and potentially fatal disease in humans [2]. Since the recognition of hantavirus cardiopulmonary syndrome (HCPS) in the southwestern United States in 1993, growing evidence of hantavirus circulation across Asia, Europe, and the Americas has increased concern regarding their epidemic potential and public health impact [3,4]. Hemorrhagic fever with renal syndrome (HFRS) accounts for thousands of hospitalizations annually, notably in Asia [5], while HCPS remains associated with high mortality rates that frequently exceed 30% in the Americas [6,7].
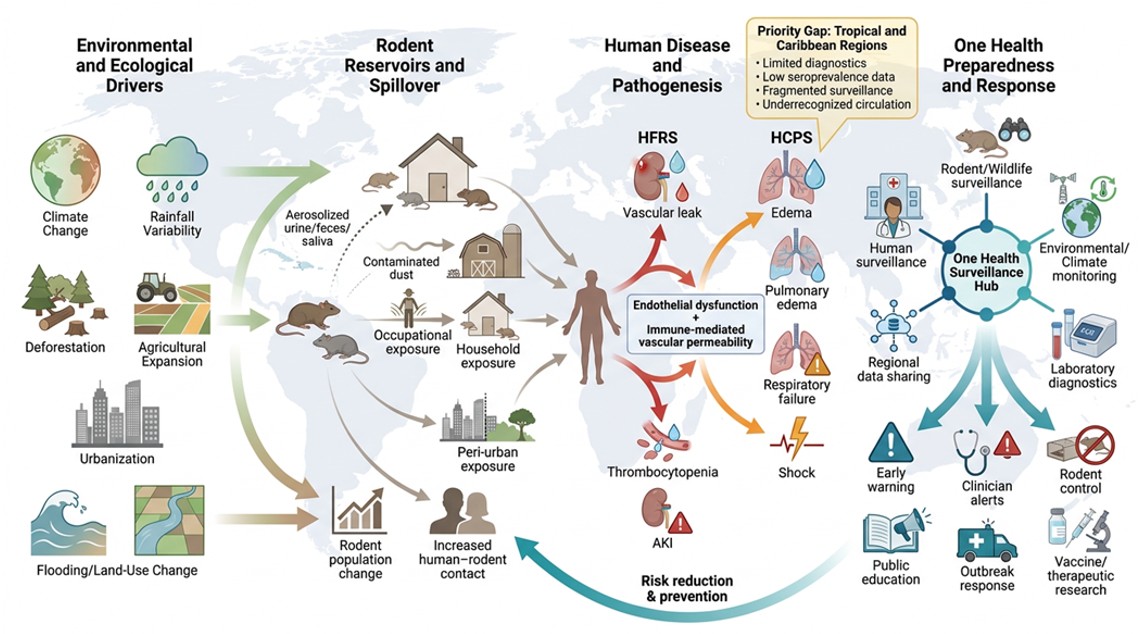
Hantaviruses are maintained in nature through persistent infection of specific rodent reservoir hosts, which shed virus in urine, feces, and saliva without developing significant disease [8]. Human infection most commonly occurs through inhalation of aerosolized viral particles from contaminated rodent excreta, although rodent bites and rare cases of person-to-person transmission, primarily involving Andes virus, have also been reported [9]. Consequently, hantavirus epidemiology is intricately linked to rodent ecology, environmental conditions, and patterns of human exposure [10].
Environmental and anthropogenic changes, such as climate variability, deforestation, urbanization, agricultural expansion, and land-use change, are increasingly influencing hantavirus transmission by altering rodent ecology and human–rodent interactions [11,12]. Climate-related events such as El Niño Southern Oscillation (ENSO) cycles and altered rainfall patterns have been linked to increased hantavirus incidence in endemic regions [13,14,15]. These concerns are especially relevant in tropical and subtropical regions, including parts of Latin America and many island nations in the Caribbean, where favorable ecological conditions and limited surveillance infrastructure may contribute to underdiagnosis [16]. This may occur because hantavirus infections can overlap clinically with other endemic febrile illnesses, such as dengue, leptospirosis, malaria, and rickettsial infections [17,18].
Furthermore, the coronavirus disease 2019 (COVID-19) pandemic highlighted the consequences of delayed pathogen detection, inadequate surveillance, and limited outbreak preparedness for emerging infectious diseases [19]. These lessons emphasize the need for integrated surveillance systems, improved diagnostic infrastructure, rapid data sharing, and coordinated public health responses [20]. In this context, a One Health approach, integrating human, animal, and environmental surveillance may enhance early detection of hantavirus circulation and strengthen preparedness, particularly in under-surveilled and resource-limited settings [21,22].
This review summarizes current evidence regarding hantavirus virology, epidemiology, pathogenesis, clinical manifestations, diagnostics, surveillance strategies, and prevention. Emphasis is placed on the environmental and ecological drivers of hantavirus emergence, surveillance and preparedness gaps in tropical and Caribbean regions, and the role of integrated One Health approaches in strengthening global preparedness and outbreak response.
2. Virology and Classification
2.1. Viral Structure and Genome Organization
Hantaviruses are enveloped, negative-sense, single-stranded RNA viruses belonging to the family Hantaviridae, order Bunyavirales [23]. The viral genome consists of three segments: small (S), medium (M), and large (L), encoding the nucleocapsid protein (N), the glycoprotein precursor (GPC, which is cleaved into Gn and Gc envelope glycoproteins), and the RNA-dependent RNA polymerase (L protein), respectively [23]. The nucleocapsid protein is highly immunogenic and serves as the primary target for serological diagnostics, while the glycoproteins mediate viral entry and are targets for neutralizing antibodies [24].
2.2. Old World and New World Hantaviruses
Hantaviruses are classified into Old World and New World lineages based on their geographic distribution and phylogenetic relationships [23]. Old World hantaviruses, mostly found in Asia and Europe, are associated with HFRS and include Hantaan virus (HTNV) [25], Seoul virus (SEOV) [26], Puumala virus (PUUV) [27], and Dobrava-Belgrade virus (DOBV) [28]. These viruses are carried by rodents of the family Muridae, with each virus typically associated with a specific rodent host [29].
New World hantaviruses, found in the Americas, are associated with HCPS and include Sin Nombre virus (SNV) [30], Andes virus (ANDV) [31], and numerous other species [32]. These viruses are primarily carried by rodents of the family Cricetidae, subfamily Sigmodontinae [29]. The Andes virus is notable for being the only hantavirus with documented person-to-person transmission [33]. Major reservoir relationships and conceptual evolutionary associations among hantaviruses are shown in Figure 1.
2.3. Host Specificity and Viral Persistence
Each hantavirus species typically exhibits a high degree of host specificity, with persistent infection in a single primary rodent reservoir species [6]. The virus establishes chronic infection in the reservoir host without causing apparent disease, facilitating long-term viral shedding in urine, feces, and saliva [34]. This host-virus co-evolution has resulted in distinct geographic distributions of hantavirus species that mirror the distributions of their rodent hosts [29]. The major hantaviruses, their reservoirs, clinical syndrome, and geographic distribution are shown in Table 1.
3. Global and Regional Epidemiology
3.1. Global Burden and Distribution
Hantavirus infections represent a notable global public health burden, with an estimated 150,000–200,000 hospitalizations annually, predominantly due to HFRS in Asia [35]. China accounts for the majority of global HFRS cases, with tens of thousands of cases reported annually [36,37]. The Republic of Korea, Russia, and several European countries also report substantial HFRS incidence [38]. In the Americas, HCPS cases are reported from Canada to southern Argentina, with the highest incidence in rural and peri-urban areas where human-rodent contact is frequent [9]. The global distribution of major hantaviruses and associated surveillance gaps is summarized in Figure 2.
3.2. Hemorrhagic Fever with Renal Syndrome (HFRS) Distribution
HFRS is endemic in Asia and Europe, with the highest burden in China, where Hantaan and Seoul viruses are the predominant causative agents [39]. The disease exhibits distinct seasonal patterns, with peak incidence typically occurring in late autumn and early winter, coinciding with increased rodent activity and agricultural harvests [40,41]. Puumala virus causes a milder form of HFRS known as nephropathia epidemica (NE) in Northern and Central Europe, with thousands of cases reported annually [42,43]. Climate variables, rodent population dynamics, and land-use changes have been associated with regional variations in HFRS incidence and transmission patterns [44].
3.3. Hantavirus Cardiopulmonary Syndrome (HCPS) Distribution
HCPS was first recognized in the southwestern United States in 1993 during an outbreak caused by Sin Nombre virus [30]. Since then, HCPS cases have been reported throughout the Americas, with notable endemic areas in Argentina, Chile, Brazil, Paraguay, and the United States [8,45]. The disease exhibits high case fatality rates, typically ranging from 20–32% [46,47], with some outbreaks reporting rates exceeding 40–50% [48,49]. Environmental and climatic factors, including El Niño-associated rainfall variability and land-use change, have been linked to increased HCPS transmission risk in several endemic regions [13,14].
3.4. Underrepresented Regions: Tropical and Caribbean Contexts
Despite the presence of diverse rodent populations and suitable ecological conditions, tropical and Caribbean regions remain significantly understudied with respect to hantavirus surveillance [16,50]. Limited seroprevalence studies and sporadic case reports suggest potential hantavirus circulation in these areas, but the true burden remains unknown due to inadequate diagnostic capacity, limited surveillance infrastructure, and low clinical awareness [51,52]. The Caribbean region, specifically, represents a major knowledge gap because systematic hantavirus surveillance remains largely absent despite the presence of both endemic and introduced rodent populations [53].
4. Pathogenesis and Immunopathology
4.1. Viral Entry and Cellular Tropism
Hantaviruses enter human cells chiefly through interactions between viral glycoproteins (Gn and Gc) and cellular receptors, including integrins [54]. The primary cellular targets are vascular endothelial cells, where viral replication occurs without direct cytopathic effects [55]. This endothelial tropism is central to the pathogenesis of both HFRS and HCPS, as endothelial dysfunction leads to increased vascular permeability and the characteristic clinical manifestations of these diseases [56,57].
4.2. Immune Response and Immunopathology
The pathogenesis of hantavirus disease is largely immune-mediated rather than directly cytopathic [58]. Following infection, both innate and adaptive immune responses are activated, with the production of pro-inflammatory cytokines, chemokines, and cellular immune responses [59]. While these responses are essential for viral control, excessive or dysregulated immune activation contributes to disease severity [58]. Neutralizing antibodies targeting the viral glycoproteins play a critical role in protection and recovery [60]. Studies have demonstrated that early and robust neutralizing antibody responses are associated with better clinical outcomes [60]. Conversely, delayed or inadequate antibody responses may contribute to severe disease [61]. T cell responses, especially CD8+ T cells, are also crucial for viral clearance but may contribute to immunopathology when dysregulated [62]. Key mechanisms contributing to hantavirus pathogenesis are summarized in Figure 3.
4.3. Vascular Permeability and Organ Dysfunction
The hallmark of hantavirus disease is increased vascular permeability resulting from endothelial dysfunction [17]. In HFRS, this manifests as capillary leakage in the kidneys, leading to acute kidney injury, proteinuria, and hematuria [63]. The disease typically progresses through five phases: febrile, hypotensive, oliguric, diuretic, and convalescent [64]. Severe cases may develop hemorrhagic complications, shock, and multi-organ failure [65].
In HCPS, vascular permeability primarily affects the pulmonary vasculature, leading to non-cardiogenic pulmonary edema, respiratory failure, and cardiogenic shock [66]. The rapid progression from initial symptoms to respiratory failure and shock is characteristic of HCPS and contributes to its high mortality rate [67]. Myocardial dysfunction, likely mediated by immune mechanisms and cytokine storm, further complicates the clinical picture [68]. The clinical progression and major pathophysiologic differences between HFRS and HCPS are illustrated in Figure 4.
4.4. Host Genetic Factors
Host genetic factors, specifically human leukocyte antigen (HLA) polymorphisms, have been associated with susceptibility to and severity of hantavirus disease [69]. Certain HLA alleles have been linked to increased risk of severe disease, while others appear protective [69]. Understanding these genetic determinants may inform risk stratification and personalized approaches to clinical management [70].
5. Environmental and Ecological Drivers
5.1. Climate Change and Hantavirus Emergence
Climate change is increasingly recognized as a major driver of hantavirus emergence by influencing rodent population dynamics, geographic distribution, and transmission risk [11,14,15]. Temperature changes, precipitation patterns, and ENSO events have been associated with increased hantavirus incidence and may support the development of climate-based early warning systems [71,72]. Climate-related expansion of rodent reservoirs into new habitats may further increase the risk of hantavirus spread to previously unaffected regions [13]. The interactions between climate variability, reservoir ecology, and hantavirus emergence are illustrated in Figure 5.
5.2. Land Use Change and Agricultural Expansion
Anthropogenic land-use changes, such as deforestation, agricultural expansion, and urbanization, can alter rodent habitats and increase human exposure to hantaviruses [14,41,71]. Studies from Brazil and China have linked environmental modification and agricultural intensification to increased hantavirus transmission risk and shifts in disease epidemiology [72,73].
5.3. Urbanization and Peri-Urban Transmission
Urbanization represents an increasingly vital driver of hantavirus transmission, especially for Seoul virus, which is carried by the Norway rat (Rattus norvegicus), a highly adaptable urban rodent [26,74]. Studies have documented Seoul virus circulation in urban rat populations in multiple countries, with sporadic human cases reported [75,76]. Long-term studies of urban rodent populations have revealed persistent hantavirus circulation in protected urban areas, with seroprevalence rates varying seasonally in association with rodent population dynamics and weather patterns [77]. These findings highlight the need for urban rodent control measures and surveillance in cities where human-rat contact is frequent [78].
5.4. Biodiversity and Dilution Effects
The relationship between biodiversity and hantavirus risk is complex, as higher biodiversity may reduce transmission through dilution effects, whereas biodiversity loss may favor reservoir species and increase transmission risk [79]. Studies suggest that biodiversity, climate, and socioeconomic factors interact to influence hantavirus emergence and should be considered in prevention strategies [80].
6. Clinical Manifestations and Diagnosis
6.1. Clinical Presentation of HFRS
HFRS typically presents with sudden onset of fever, headache, back pain, abdominal pain, and gastrointestinal symptoms [81,82]. The disease progresses through characteristic phases: febrile (3–7 days), hypotensive (hours to 3 days), oliguric (3–7 days), diuretic (days to weeks), and convalescent (weeks to months) [83]. Laboratory findings include thrombocytopenia, proteinuria, hematuria, and elevated creatinine [82]. Imaging findings in HFRS commonly include renal enlargement, perirenal fluid accumulation, and retroperitoneal edema, with hemorrhagic manifestations observed in severe disease [84]. The severity of HFRS varies by viral species, with Hantaan and Dobrava-Belgrade viruses typically causing more severe disease than Puumala virus [85].
6.2. Clinical Presentation of HCPS
HCPS typically begins with a prodromal phase characterized by fever, myalgia, headache, and gastrointestinal symptoms lasting 3–6 days [86]. This is followed by rapid onset of respiratory distress, pulmonary edema, and cardiogenic shock [87]. Characteristic laboratory findings include thrombocytopenia, hemoconcentration, elevated lactate dehydrogenase, and immunoblastic lymphocytes [8]. Chest imaging reveals bilateral interstitial infiltrates and pleural effusions [8]. The rapid progression from initial symptoms to respiratory failure necessitates early recognition and intensive supportive care [87]. Survivors typically experience complete recovery, though convalescence may be prolonged [86]. The clinical differences between the two syndromes are shown in Table 2.
6.3. Diagnostic Methods
Early and accurate diagnosis of hantavirus infection is critical for appropriate clinical management and public health response [88,89]. Diagnostic approaches include serological methods, molecular detection, and immunohistochemistry [17,90].
6.3.1. Serological Diagnostics
Serological testing is the primary diagnostic approach for hantavirus infections, with Enzyme-linked immunosorbent assay (ELISA)-based detection of IgM and IgG antibodies commonly used for initial screening [17]. Confirmatory methods, namely immunofluorescence assays (IFA), Western blot, and microneutralization tests, can improve specificity and allow species-level identification; however, cross-reactivity among hantavirus species may complicate interpretation in regions where multiple strains co-circulate [90].
6.3.2. Molecular Diagnostics
Reverse transcription polymerase chain reaction (RT-PCR) enables direct detection of hantavirus RNA and is extremely useful for early diagnosis before antibody seroconversion [88]. Real-time RT-PCR assays targeting conserved viral regions provide rapid and highly sensitive detection, although their utility is limited by the rapid decline in viral RNA after symptom onset [89].
6.3.3. Point-of-Care and Field-Deployable Diagnostics
A proposed diagnostic workflow for suspected hantavirus infection is presented in Figure 6, and laboratory approaches for hantavirus detection are detailed in Table 3. Point-of-care (POC) diagnostics are increasingly crucial for hantavirus detection in resource-limited and outbreak settings [91]. Emerging approaches, comprising RT-LAMP and lateral flow immunoassays, offer rapid and field-deployable testing, although challenges related to sensitivity and specificity remain [92]. Ideal assays should provide rapid, accurate, and affordable detection with minimal equipment requirements [93].
6.4. Diagnostic Challenges in Tropical and Caribbean Settings
Diagnostic capacity for hantavirus infections in tropical and Caribbean regions faces multiple challenges [53]. Limited availability of validated serological assays, lack of molecular diagnostic infrastructure, absence of reference laboratories, and limited awareness among healthcare providers all contribute to underdiagnosis [96]. Additionally, the potential circulation of uncharacterized hantavirus species in these regions may limit the performance of existing diagnostic assays developed for well-characterized species [38]. Strengthening laboratory infrastructure and improving clinician awareness are essential for enhancing early detection and surveillance capacity in these settings [53]. Differential diagnoses of hantavirus infection in tropical settings are listed in Table 4.
7. Surveillance and Early Warning Systems
7.1. Current Surveillance Approaches
Hantavirus surveillance systems vary widely in scope and sophistication across different regions [98]. In countries with high disease burden, such as China and the Republic of Korea, national surveillance systems capture case data, conduct seroprevalence surveys, and monitor rodent populations [82,83]. These systems have generated valuable long-term datasets that have informed understanding of disease epidemiology and risk factors.
In contrast, many tropical and Caribbean countries lack systematic hantavirus surveillance, relying instead on passive case detection and sporadic research studies [16]. This surveillance gap limits understanding of disease burden, circulating viral species, and risk factors in these regions.
7.2. Integrated One Health Surveillance
The One Health approach integrates human, animal, and environmental surveillance systems to improve hantavirus monitoring, risk assessment, and outbreak preparedness [21]. Human surveillance focuses on clinical case detection and laboratory confirmation, while rodent surveillance evaluates reservoir distribution, population dynamics, and viral prevalence [29]. Environmental surveillance incorporates climate variability, land-use change, and ecological factors associated with transmission risk [40]. The integration of these surveillance streams improves early detection of changing transmission patterns and supports targeted public health interventions. The interconnected surveillance framework and data flows are illustrated in Figure 7.
7.3. Climate-Based Early Warning Systems
Associations between climate variables and hantavirus incidence have prompted the development of climate-based early warning systems that integrate environmental data, rodent surveillance, and predictive models to identify periods of increased transmission risk [11,13]. Studies in China and the Americas suggest that temperature, precipitation, and El Niño patterns may help forecast outbreaks and support public health preparedness [15].
7.4. Surveillance Priorities for Tropical and Caribbean Regions
Strengthening hantavirus surveillance in tropical and Caribbean regions requires improved epidemiological research, diagnostic capacity, healthcare awareness, and regional collaboration [12,17]. Integrating hantavirus monitoring into existing infectious disease surveillance systems may provide a practical and cost-effective approach to improving preparedness [12]. Major surveillance gaps and preparedness priorities are summarized in Figure 8, while surveillance challenges and proposed solutions are outlined in Table 5.
8. Prevention and Control
8.1. Prevention of Human Exposure
Primary prevention of hantavirus focuses on reducing human exposure to infected rodent excreta through rodent-proofing, sanitation, safe cleanup practices, use of personal protective equipment, and public education [6,47]. Enhanced precautions are crucial in high-risk occupational and outdoor settings, including farming, forestry, construction, and military activities [38].
8.2. Rodent Control
Rodent control is an important strategy for reducing hantavirus transmission risk, although long-term effectiveness may be limited by ecological and logistical challenges [99]. Habitat modification, sanitation measures, trapping, and targeted rodent reduction programs are commonly used to reduce human exposure in high-risk settings [71]. The use of rodenticides may be effective in some contexts but raises concerns regarding environmental impact and non-target species exposure [100]. Integrated pest management (IPM) strategies that combine environmental sanitation, habitat reduction, surveillance, and selective rodent control are generally recommended for sustainable prevention efforts [101]. Urban rodent control is essential for preventing Seoul virus transmission associated with Norway rats in densely populated cities [78].
8.3. Vaccines
No hantavirus vaccine is presently licensed outside China and the Republic of Korea, where inactivated vaccines against Hantaan and Seoul viruses have shown limited and short-lived immunity [87,102]. Several vaccine platforms, such as DNA, recombinant protein, viral vector, virus-like particle, and mRNA vaccines, are under investigation, with preclinical studies demonstrating promising protective immune responses and potential for rapid multivalent vaccine development [103,104].
8.4. Therapeutics
There are currently no FDA-approved treatments for hantavirus infection, and management remains largely supportive, including dialysis for HFRS and mechanical ventilation or ECMO for severe HCPS cases [105,106]. Ribavirin may reduce mortality in HFRS when administered early, although evidence remains limited and benefits have not been demonstrated for HCPS [17,102]. Emerging immunotherapies, for example neutralizing monoclonal antibodies targeting viral glycoproteins, have shown promising protective effects in preclinical studies [103,107].
9. Research Priorities and Future Directions
9.1. Surveillance and Diagnostic Innovation
Advancing surveillance and diagnostic capabilities is essential for improving hantavirus detection and outbreak preparedness, especially in underrepresented tropical and Caribbean regions. Priority areas include the development of genomic surveillance platforms, rapid point-of-care diagnostics, digital surveillance systems, and standardized reporting frameworks [88,90]. Artificial intelligence, machine learning approaches, and environmental monitoring tools may further improve outbreak prediction and risk mapping by integrating climate, ecological, and epidemiological data [89,91,108]. Collectively, these innovations may strengthen early detection and public health response capacity [109].
9.2. Ecological and Environmental Research
Understanding hantavirus emergence requires integrated ecological, environmental, and epidemiological research. Key priorities include ecological niche modeling, longitudinal rodent surveillance, and studies evaluating the effects of climate change, biodiversity loss, land-use change, and environmental disruption on reservoir ecology and transmission risk [110]. Incorporating hantavirus risk assessments into environmental and land-use planning may help reduce future spillover risk [71].
9.3. Pathogenesis and Immunology
Improving understanding of hantavirus pathogenesis and host immune responses is important for the development of vaccines and therapeutics. Key research priorities include identifying mechanisms of endothelial dysfunction, vascular permeability, and immune-mediated injury associated with severe disease [56]. Studies investigating protective immunity, host genetic susceptibility, and improved animal models may further support the development of targeted therapies and vaccine strategies [69,70].
9.4. Vaccines and Therapeutics
Current research priorities for hantavirus vaccines and therapeutics focus on developing broadly protective and rapidly deployable countermeasures. Major areas of investigation include multivalent vaccines, mRNA vaccine platforms, monoclonal antibody therapies, and antiviral agents targeting viral replication or entry pathways [102,103]. Immunomodulatory therapies aimed at reducing excessive inflammatory responses may also improve outcomes in severe hantavirus disease [86,87].
9.5. Capacity Building in Underrepresented Regions
Addressing hantavirus surveillance and research gaps in tropical and Caribbean regions requires sustained investment in laboratory infrastructure, workforce training, and regional collaboration [53]. Priority efforts include strengthening diagnostic and genomic surveillance capacity, improving outbreak investigation expertise, and establishing collaborative regional networks for data sharing and coordinated response activities [96]. Targeted funding and South–South collaboration may further support sustainable preparedness and surveillance development in under-resourced regions [111,112].
10. One Health Framework for Hantavirus Preparedness
10.1. Integrating Human, Animal, and Environmental Health
The One Health framework recognizes the interconnected relationship between human, animal, and environmental health and is highly relevant to hantavirus preparedness [113]. Effective preparedness requires collaboration among public health agencies, veterinary services, environmental scientists, and wildlife experts to support integrated surveillance and coordinated outbreak response [114]. Shared surveillance systems that combine human case data, rodent ecology, and environmental monitoring may improve early detection of emerging transmission hotspots and support timely interventions.
10.2. Lessons from COVID-19 for Hantavirus Preparedness
The COVID-19 pandemic highlighted the importance of rapid surveillance, decentralized diagnostic capacity, international collaboration, and transparent public health communication for emerging infectious disease preparedness [115]. These lessons emphasize the need for integrated surveillance systems, rapid data sharing, and flexible public health infrastructure to strengthen hantavirus preparedness and outbreak response [19,115].
10.3. Building Resilient Health Systems
Strengthening health systems for emerging infectious disease preparedness requires sustained investment in surveillance, laboratory diagnostics, outbreak response capacity, and workforce development [24]. Flexible public health infrastructure and coordinated partnerships among governments, academic institutions, and international organizations are essential for effective preparedness and response [12]. Resilient and adaptable health systems will be critical for responding to future hantavirus outbreaks and other emerging zoonotic threats. One Health preparedness strategies are shown in Table 6.
11. Current Knowledge Gaps and Limitations
Important gaps remain in hantavirus surveillance, diagnostics, and disease recognition, most notably in tropical and resource-limited regions where infections may be underdiagnosed due to overlap with other endemic febrile illnesses and limited access to molecular and serological testing [12,17]. In addition, the ecological drivers of hantavirus emergence, including climate change, land-use change, and biodiversity loss, remain incompletely understood [13,14,15,41].
Despite progress in vaccine and therapeutic research, no widely available vaccine or specific antiviral therapy currently exists [102,103,104]. Variability in surveillance systems, diagnostic methods, and case definitions further limits comparisons across regions and may underestimate the global disease burden, highlighting the need for strengthened surveillance, standardized reporting, and integrated One Health approaches [21,22].
12. Conclusions
Hantaviruses remain critical emerging zoonotic pathogens with significant global health implications, primarily in regions where environmental change, urbanization, and climate variability are altering reservoir ecology and increasing opportunities for human exposure. The pathogenesis of hantavirus infections is driven largely by immune-mediated endothelial dysfunction and vascular permeability, leading to severe clinical syndromes including HFRS and HCPS. Despite advances in understanding hantavirus virology, immunopathology, diagnostics, and reservoir ecology, major gaps persist in surveillance infrastructure, diagnostic capacity, and epidemiological data. These gaps are especially evident in tropical and Caribbean regions, where hantavirus transmission may be underrecognized.
Recent progress in molecular diagnostics, genomic surveillance, vaccine platforms, monoclonal antibody therapies, and climate-informed risk modeling provides vital opportunities to improve early detection, preparedness, and outbreak response. However, sustained investment in laboratory infrastructure, ecological surveillance, workforce training, and regional collaboration remains essential. Integrated One Health approaches that combine human, animal, and environmental surveillance will be critical for strengthening global preparedness and reducing the impact of future hantavirus outbreaks and other emerging zoonotic threats.
Author Contributions
Conceptualization, M.E.R. and P.V.A.K.R.; methodology, M.E.R. and P.V.A.K.R.; validation, P.V.A.K.R.; investigation, M.E.R. and P.V.A.K.R.; resources, M.E.R. and P.V.A.K.R.; writing—original draft preparation, M.E.R., N.C. and P.V.A.K.R.; writing—review and editing, M.E.R., N.C. and P.V.A.K.R.; visualization, M.E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding. The APC was funded by St. George’s University School of Medicine, St. George, Grenada.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data was generated. All cited references are publicly available in PubMed or other indexed databases.
Acknowledgments
During the preparation of this manuscript, the author used FigureLabs (www.figurelabs.com) to generate figures, and OpenAI’s ChatGPT (GPT-5.5, 2026) for proofreading assistance. After using these tools/service, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ANDV | Andes virus |
| COVID-19 | Coronavirus disease 2019 |
| DOBV | Dobrava-Belgrade virus |
| ECMO | Extracorporeal membrane oxygenation |
| ELISA | Enzyme-linked immunosorbent assay |
| ENSO | El Niño Southern Oscillation |
| FDA | Food and Drug Administration |
| HCPS | Hantavirus cardiopulmonary syndrome |
| HFRS | Hemorrhagic fever with renal syndrome |
| HLA | Human leukocyte antigen |
| HTNV | Hantaan virus |
| IFA | Immunofluorescence assay |
| LAMP | Loop-mediated isothermal amplification |
| LNV | Laguna Negra virus |
| MNT | Microneutralization test |
| NE | Nephropathia epidemica |
| NHP | Non-human primates |
| POC | Point-of-care |
| PUUV | Puumala virus |
| RT-PCR | Reverse transcription polymerase chain reaction |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| SEOV | Seoul virus |
| SNV | Sin Nombre virus |
References
- Wang, S.; Li, W.; Wang, Z.; Yang, W.; Li, E.; Xia, X.; Yan, F.; Chiu, S. Emerging and Reemerging Infectious Diseases: Global Trends and New Strategies for Their Prevention and Control. Signal Transduct. Target. Ther. 2024, 9, 223. [Google Scholar] [CrossRef]
- Taylor, S.L.; Schmaljohn, C.S.; Williams, E.P.; Jonsson, C.B. Pathogenicity and Virulence of Rodent-Borne Orthohantaviruses. Virulence 2025, 16, 2553784. [Google Scholar] [CrossRef]
- Ksiazek, T.G.; Peters, C.J.; Rollin, P.E.; Zaki, S.; Nichol, S.; Spiropoulou, C.; Morzunov, S.; Feldmann, H.; Sanchez, A.; Khan, A.S. Identification of a New North American Hantavirus That Causes Acute Pulmonary Insufficiency. Am. J. Trop. Med. Hyg. 1995, 52, 117–123. [Google Scholar] [CrossRef]
- Song, J.-N.; Chen, D.; Wang, L.-M.; Jiang, H. Global Threats and Regional Trends: Navigating the Complex Landscape of Human Orthohantavirus Infections. Infectious Dis. Immunity 9900. [CrossRef]
- Korva, M.; Rus, K.R.; Pavletič, M.; Saksida, A.; Knap, N.; Jelovšek, M.; Smrdel, K.S.; Jakupi, X.; Humolli, I.; Dedushaj, J.; et al. Characterization of Biomarker Levels in Crimean-Congo Hemorrhagic Fever and Hantavirus Fever with Renal Syndrome. Viruses 2019, 11. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef]
- Armien, B.; Pascale, J.M.; Muñoz, C.; Mariñas, J.; Núñez, H.; Herrera, M.; Trujillo, J.; Sánchez, D.; Mendoza, Y.; Hjelle, B.; et al. Hantavirus Fever without Pulmonary Syndrome in Panama. Am. J. Trop. Med. Hyg. 2013, 89, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Vial, P.A.; Ferrés, M.; Vial, C.; Klingström, J.; Ahlm, C.; López, R.; Le Corre, N.; Mertz, G.J. Hantavirus in Humans: A Review of Clinical Aspects and Management. Lancet Infect. Dis. 2023, 23, e371–e382. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Valdebenito, C.; Calvo, M.; Vial, C.; Mansilla, R.; Marco, C.; Palma, R.E.; Vial, P.A.; Valdivieso, F.; Mertz, G.; Ferrés, M. Person-to-Person Household and Nosocomial Transmission of Andes Hantavirus, Southern Chile, 2011. Emerg. Infect. Dis. 2014, 20, 1629–1636. [Google Scholar] [CrossRef] [PubMed]
- Guterres, A.; de Lemos, E.R.S. Hantaviruses and a Neglected Environmental Determinant. One Health 2018, 5, 27–33. [Google Scholar] [CrossRef]
- Guo, J.; Semenza, J.C.; Ecke, F.; Rizzoli, A.; Dagostin, F.; Ulrich, R.G.; Sjödin, H.; Treskova, M.; Rocklöv, J. A Pan-European Assessment of Multi-Sector Drivers of Human Hantavirus Risk: Climate, Biodiversity, and Socio-Economic Factors as Key Determinants. Env. Res. 2026, 298, 124282. [Google Scholar] [CrossRef]
- Bhushan, K.; Kanwar, S. The Negative Sense RNA Hantavirus – A Threat to the Modern World. Asian Pac. J. Health Sci. 2020, 7, 42–47. [Google Scholar] [CrossRef]
- Prist, P.R.; Uriarte, M.; Fernandes, K.; Metzger, J.P. Climate Change and Sugarcane Expansion Increase Hantavirus Infection Risk. PLoS Negl. Trop. Dis. 2017, 11, e0005705. [Google Scholar] [CrossRef] [PubMed]
- Muylaert, R.L.; Sabino-Santos, G.J.; Prist, P.R.; Oshima, J.E.F.; Niebuhr, B.B.; Sobral-Souza, T.; de Oliveira, S.V.; Bovendorp, R.S.; Marshall, J.C.; Hayman, D.T.S.; et al. Spatiotemporal Dynamics of Hantavirus Cardiopulmonary Syndrome Transmission Risk in Brazil. Viruses 2019, 11. [Google Scholar] [CrossRef]
- Bai, Y.; Xu, Z.; Lu, B.; Sun, Q.; Tang, W.; Liu, X.; Yang, W.; Xu, X.; Liu, Q. Effects of Climate and Rodent Factors on Hemorrhagic Fever with Renal Syndrome in Chongqing, China, 1997-2008. PLoS ONE 2015, 10, e0133218. [Google Scholar] [CrossRef]
- Douglas, K.O.; Payne, K.; Sabino-Santos, G.J.; Agard, J. Influence of Climatic Factors on Human Hantavirus Infections in Latin America and the Caribbean: A Systematic Review. Pathogens 2021, 11. [Google Scholar] [CrossRef]
- Mattar, S.; Guzmán, C.; Figueiredo, L.T. Diagnosis of Hantavirus Infection in Humans. Expert Rev. Anti Infect. Ther. 2015, 13, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Lerma, L.; Mattar, S.; Contreras, V.; Miranda, J.; Tique, V.; Rodríguez, V.; Rodriguez, D.; Lopez, S.; Rojas-Gulloso, A. Hantavirus and Leptospira Are Important Causes of Nonspecific Acute Febrile Syndrome, Meta, Colombia. Travel Med. Infect. Dis. 2025, 64, 102800. [Google Scholar] [CrossRef]
- Williams, B.A.; Jones, C.H.; Welch, V.; True, J.M. Outlook of Pandemic Preparedness in a Post-COVID-19 World. npj Vaccines 2023, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Maccaro, A.; Audia, C.; Stokes, K.; Masud, H.; Sekalala, S.; Pecchia, L.; Piaggio, D. Pandemic Preparedness: A Scoping Review of Best and Worst Practices from COVID-19. Healthcare 2023, 11. [Google Scholar] [CrossRef]
- Desvars-Larrive, A.; Vogl, A.E.; Puspitarani, G.A.; Yang, L.; Joachim, A.; Käsbohrer, A. A One Health Framework for Exploring Zoonotic Interactions Demonstrated through a Case Study. Nat. Commun. 2024, 15, 5650. [Google Scholar] [CrossRef]
- Berjaoui, S.; Puglia, I.; Caporale, M.; Pinoni, C.; Salvaggiulo, A.; Gatta, G.; Sepashvili, M.; Di Donato, G. Hantaviruses in the One Health Era: Strengthening Surveillance before the next Spillover. Vet. Ital. 2026, 62. [Google Scholar] [CrossRef]
- Bradfute, S.B.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Laenen, L.; Tischler, N.D.; Maes, P. ICTV Virus Taxonomy Profile: Hantaviridae 2024. J. Gen. Virol. 2024, 105. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; McFadden, E.; Slough, M.M.; Stone, E.T.; Berrigan, J.; Mittler, E.; Hatzakis, K.; Hinkley, T.; Kain, H.S.; Ke, Z.; et al. High-Resolution in Situ Structures of Hantavirus Glycoprotein Tetramers. Cell 2026, 189, 2731–2747.e15. [Google Scholar] [CrossRef] [PubMed]
- Khaiboullina, S.F.; Morzunov, S.P.; St Jeor, S.C. Hantaviruses: Molecular Biology, Evolution and Pathogenesis. Curr. Mol. Med. 2005, 5, 773–790. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-D.; Guo, W.-P.; Wang, W.; Zou, Y.; Hao, Z.-Y.; Zhou, D.-J.; Dong, X.; Qu, Y.-G.; Li, M.-H.; Tian, H.-F.; et al. Migration of Norway Rats Resulted in the Worldwide Distribution of Seoul Hantavirus Today. J. Virol. 2012, 86, 972–981. [Google Scholar] [CrossRef]
- Brummer-Korvenkontio, M.; Vaheri, A.; Hovi, T.; von Bonsdorff, C.H.; Vuorimies, J.; Manni, T.; Penttinen, K.; Oker-Blom, N.; Lähdevirta, J. Nephropathia Epidemica: Detection of Antigen in Bank Voles and Serologic Diagnosis of Human Infection. J. Infect. Dis. 1980, 141, 131–134. [Google Scholar] [CrossRef]
- Klempa, B.; Schütt, M.; Auste, B.; Labuda, M.; Ulrich, R.; Meisel, H.; Krüger, D.H. First Molecular Identification of Human Dobrava Virus Infection in Central Europe. J. Clin. Microbiol. 2004, 42, 1322–1325. [Google Scholar] [CrossRef]
- Herbreteau, V.; Henttonen, H.; Yoshimatsu, K.; Gonzalez, J.-P.; Suputtamongkol, Y.; Hugot, J.-P. Hantavirus Coevolution with Their Rodent Hosts. Encycl. Infect. Dis. Mod. Methodol. 2006, 243–264. [Google Scholar]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.; Rollin, P.E.; Ksiazek, T.G.; Feldmann, H.; Sanchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic Identification of a Hantavirus Associated with an Outbreak of Acute Respiratory Illness. Science 1993, 262, 914–917. [Google Scholar] [CrossRef]
- López, N.; Padula, P.; Rossi, C.; Lázaro, M.E.; Franze-Fernández, M.T. Genetic Identification of a New Hantavirus Causing Severe Pulmonary Syndrome in Argentina. Virology 1996, 220, 223–226. [Google Scholar] [CrossRef]
- Young, J.C.; Mills, J.N.; Enria, D.A.; Dolan, N.E.; Khan, A.S.; Ksiazek, T.G. New World Hantaviruses. Br. Med. Bull. 1998, 54, 659–673. [Google Scholar] [CrossRef]
- Wells, R.M.; Sosa Estani, S.; Yadon, Z.E.; Enria, D.; Padula, P.; Pini, N.; Mills, J.N.; Peters, C.J.; Segura, E.L. An Unusual Hantavirus Outbreak in Southern Argentina: Person-to-Person Transmission? Hantavirus Pulmonary Syndrome Study Group for Patagonia. Emerg. Infect. Dis. 1997, 3, 171–174. [Google Scholar] [CrossRef]
- Easterbrook, J.D.; Klein, S.L. Immunological Mechanisms Mediating Hantavirus Persistence in Rodent Reservoirs. PLoS Pathog. 2008, 4, e1000172. [Google Scholar] [CrossRef]
- Watson, D.C.; Sargianou, M.; Papa, A.; Chra, P.; Starakis, I.; Panos, G. Epidemiology of Hantavirus Infections in Humans: A Comprehensive, Global Overview. Crit. Rev. Microbiol. 2014, 40, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Z.; Zou, Y.; Fu, Z.F.; Plyusnin, A. Hantavirus Infections in Humans and Animals, China. Emerg. Infect. Dis. 2010, 16, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, S.; Yin, W.; Liang, M.; Li, J.; Zhang, Q.; Feng, Z.; Li, D. Epidemic Characteristics of Hemorrhagic Fever with Renal Syndrome in China, 2006-2012. BMC Infect. Dis. 2014, 14, 384. [Google Scholar] [CrossRef] [PubMed]
- Manigold, T.; Vial, P. Human Hantavirus Infections: Epidemiology, Clinical Features, Pathogenesis and Immunology. Swiss Med. Wkly. 2014, 144, w13937. [Google Scholar] [CrossRef]
- Jiang, H.; Zheng, X.; Wang, L.; Du, H.; Wang, P.; Bai, X. Hantavirus Infection: A Global Zoonotic Challenge. Virol. Sin. 2017, 32, 32–43. [Google Scholar] [CrossRef]
- Tian, H.-Y.; Yu, P.-B.; Luis, A.D.; Bi, P.; Cazelles, B.; Laine, M.; Huang, S.-Q.; Ma, C.-F.; Zhou, S.; Wei, J.; et al. Changes in Rodent Abundance and Weather Conditions Potentially Drive Hemorrhagic Fever with Renal Syndrome Outbreaks in Xi’an, China, 2005-2012. PLoS Negl. Trop. Dis. 2015, 9, e0003530. [Google Scholar] [CrossRef]
- Tian, H.; Yu, P.; Bjørnstad, O.N.; Cazelles, B.; Yang, J.; Tan, H.; Huang, S.; Cui, Y.; Dong, L.; Ma, C.; et al. Anthropogenically Driven Environmental Changes Shift the Ecological Dynamics of Hemorrhagic Fever with Renal Syndrome. PLoS Pathog. 2017, 13, e1006198. [Google Scholar] [CrossRef]
- Tscherne, A.; Guardado-Calvo, P.; Clark, J.J.; Krause, R.; Krammer, F.
- Koroknai, A.; Nagy, A.; Nagy, O.; Csonka, N.; Zsichla, L.; Szomor, K.; Takács, M. Human Hantavirus Infections in Hungary (2018–2025): Epidemiology, Molecular Detection Across Clinical Sample Types, and Phylogenetic Analysis. Viruses 2026, 18, 366. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zou, L.-X. Spatiotemporal Analysis and Forecasting Model of Hemorrhagic Fever with Renal Syndrome in Mainland China. Epidemiol. Infect. 2018, 146, 1680–1688. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, S.L.M.; Whitesell, A.; Mobley, M.; Talundzic, E.; Shedroff, E.; Cossaboom, C.M.; Messenger, S.; Deldari, M.; Bhatnagar, J.; Estetter, L.; et al. Human Orthohantavirus Disease Prevalence and Genotype Distribution in the U.S., 2008–2020: A Retrospective Observational Study. Lancet Reg. Health – Am. 2024, 37. [Google Scholar] [CrossRef]
- Alonso, D.O.; Iglesias, A.; Coelho, R.; Periolo, N.; Bruno, A.; Córdoba, M.T.; Filomarino, N.; Quipildor, M.; Biondo, E.; Fortunato, E.; et al. Epidemiological Description, Case-Fatality Rate, and Trends of Hantavirus Pulmonary Syndrome: 9 Years of Surveillance in Argentina. J. Med. Virol. 2019, 91, 1173–1181. [Google Scholar] [CrossRef]
- Martínez Valeria, P.; Di Paola Nicholas; Alonso Daniel, O.; Pérez-Sautu Unai; Bellomo, Carla M.; Iglesias Ayelén, A.; Coelho Rocio, M.; López Beatriz; Natalia, Periolo; Larson Peter, A.; et al. “Super-Spreaders” and Person-to-Person Transmission of Andes Virus in Argentina. N. Engl. J. Med. 2020, 383, 2230–2241. [Google Scholar] [CrossRef]
- Figueiredo, L.T.M.; Moreli, M.L.; de-Sousa, R.L.M.; Borges, A.A.; de-Figueiredo, G.G.; Machado, A.M.; Bisordi, I.; Nagasse-Sugahara, T.K.; Suzuki, A.; Pereira, L.E.; et al. Hantavirus Pulmonary Syndrome, Central Plateau, Southeastern, and Southern Brazil. Emerg. Infect. Dis. 2009, 15, 561–567. [Google Scholar] [CrossRef]
- Mendes, W.S.; da Silva, A.A.M.; Aragão, L.F.C.; Aragão, N.J.L.; Raposo, M. de L.; Elkhoury, M.R.; Suzuky, A.; Ferreira, I.B.; de Sousa, L.T.; Pannuti, C.S. Hantavirus Infection in Anajatuba, Maranhao, Brazil. Emerg. Infect. Dis. 2004, 10, 1496–1498. [Google Scholar] [CrossRef]
- Castel, G.; Filippone, C.; Tatard, C.; Vigan, J.; Dobigny, G. Role of Seaports and Imported Rats in Seoul Hantavirus Circulation, Africa. Emerg. Infect. Dis. 2023, 29, 20–25. [Google Scholar] [CrossRef]
- Witkowski, P.; Leendertz, S.A.J.; Auste, B.; Akoua-Koffi, C.; Schubert, G.; Klempa, B.; Muyembe-Tamfum, J.-J.; Karhemere, S.; Leendertz, F.; Krüger, D. Human Seroprevalence Indicating Hantavirus Infections in Tropical Rainforests of Côte d’Ivoire and Democratic Republic of Congo. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Douglas, K.O.; Samuels, T.A.; Iheozor-Ejiofor, R.; Vapalahti, O.; Sironen, T.; Gittens-St Hilaire, M. Serological Evidence of Human Orthohantavirus Infections in Barbados, 2008 to 2016. Pathogens 2021, 10. [Google Scholar] [CrossRef]
- Rodríguez, Á.; Couto, P.; Acevedo, A.; Herrera, B.A.; Astudillo, O.; Avaro, M.; Badillo, G.B.; Bruno, A.; Bustos, P.; Cerpa, M.; et al. Strengthening the Surveillance and Response to Public Health Events With a One Health Approach: A Perspective From 12 Countries in Latin America and the Caribbean. J. Infect. Dis. 2025, 231, S108–S113. [Google Scholar] [CrossRef]
- Mittler, E.; Dieterle, M.E.; Kleinfelter, L.M.; Slough, M.M.; Chandran, K.; Jangra, R.K. Hantavirus Entry: Perspectives and Recent Advances. Adv. Virus Res. 2019, 104, 185–224. [Google Scholar] [CrossRef]
- Ermonval, M.; Baychelier, F.; Tordo, N. What Do We Know about How Hantaviruses Interact with Their Different Hosts? Viruses 2016, 8, 223. [Google Scholar] [CrossRef]
- Taylor, S.L.; Wahl-Jensen, V.; Copeland, A.M.; Jahrling, P.B.; Schmaljohn, C.S. Endothelial Cell Permeability during Hantavirus Infection Involves Factor XII-Dependent Increased Activation of the Kallikrein-Kinin System. PLoS Pathog. 2013, 9, e1003470. [Google Scholar] [CrossRef]
- Krautkrämer, E.; Grouls, S.; Hettwer, D.; Rafat, N.; Tönshoff, B.; Zeier, M. Mobilization of Circulating Endothelial Progenitor Cells Correlates with the Clinical Course of Hantavirus Disease. J. Virol. 2014, 88, 483–489. [Google Scholar] [CrossRef]
- Saavedra, F.; Díaz, F.E.; Retamal-Díaz, A.; Covián, C.; González, P.A.; Kalergis, A.M. Immune Response during Hantavirus Diseases: Implications for Immunotherapies and Vaccine Design. Immunology 2021, 163, 262–277. [Google Scholar] [CrossRef]
- Klingström, J.; Smed-Sörensen, A.; Maleki, K.T.; Solà-Riera, C.; Ahlm, C.; Björkström, N.K.; Ljunggren, H.G. Innate and Adaptive Immune Responses against Human Puumala Virus Infection: Immunopathogenesis and Suggestions for Novel Treatment Strategies for Severe Hantavirus-Associated Syndromes. J. Intern Med. 2019, 285, 510–523. [Google Scholar] [CrossRef]
- Rissanen, I.; Krumm, S.A.; Stass, R.; Whitaker, A.; Voss, J.E.; Bruce, E.A.; Rothenberger, S.; Kunz, S.; Burton, D.R.; Huiskonen, J.T.; et al. Structural Basis for a Neutralizing Antibody Response Elicited by a Recombinant Hantaan Virus Gn Immunogen. mBio 2021, 12, e0253120. [Google Scholar] [CrossRef]
- Iglesias, A.A.; Períolo, N.; Bellomo, C.M.; Lewis, L.C.; Olivera, C.P.; Anselmo, C.R.; García, M.; Coelho, R.M.; Alonso, D.O.; Dighero-Kemp, B.; et al. Delayed Viral Clearance despite High Number of Activated T Cells during the Acute Phase in Argentinean Patients with Hantavirus Pulmonary Syndrome. eBioMedicine 2022, 75. [Google Scholar] [CrossRef]
- Kilpatrick, E.D.; Terajima, M.; Koster, F.T.; Catalina, M.D.; Cruz, J.; Ennis, F.A. Role of Specific CD8+ T Cells in the Severity of a Fulminant Zoonotic Viral Hemorrhagic Fever, Hantavirus Pulmonary Syndrome. J. Immunol. 2004, 172, 3297–3304. [Google Scholar] [CrossRef]
- Bui-Mansfield, L.T.; Cressler, D.K. Imaging of Hemorrhagic Fever with Renal Syndrome: A Potential Bioterrorism Agent of Military Significance. Mil. Med. 2011, 176, 1327–1334. [Google Scholar] [CrossRef]
- Bruno, M.; Sheer, R.; Reed, C.; Schepart, A.; Nair, R.; Simmons, J.D. Clinical Characteristics and Health Care Resource Use of Patients at Risk for Wild-Type Transthyretin Amyloid Cardiomyopathy Identified by Machine Learning Model. J. Manag. CARE Spec. Pharm. 2023, 29, 530–540. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, X.-Y.; Liu, X.-J.; Zhang, M.; Sun, Y.-T.; Ning, X.; Xu, J.; Bu, C.-Y. Case Report: Multiple Organ Failure Caused by Hemorrhagic Fever with Renal Syndrome. Am. J. Trop. Med. Hyg. 2023, 109, 101–104. [Google Scholar] [CrossRef]
- Llah, S.T.; Mir, S.; Sharif, S.; Khan, S.; Mir, M.A. Hantavirus Induced Cardiopulmonary Syndrome: A Public Health Concern. J. Med. Virol. 2018, 90, 1003–1009. [Google Scholar] [CrossRef]
- Chang, B.; Crowley, M.; Campen, M.; Koster, F. Hantavirus Cardiopulmonary Syndrome. Semin Respir. Crit. Care Med. 2007, 28, 193–200. [Google Scholar] [CrossRef]
- Bondu, V.; Schrader, R.; Gawinowicz, M.A.; McGuire, P.; Lawrence, D.A.; Hjelle, B.; Buranda, T. Elevated Cytokines, Thrombin and PAI-1 in Severe HCPS Patients Due to Sin Nombre Virus. Viruses 2015, 7, 559–589. [Google Scholar] [CrossRef]
- Ma, Y.; Yuan, B.; Yi, J.; Zhuang, R.; Wang, J.; Zhang, Y.; Xu, Z.; Zhang, Y.; Liu, B.; Wei, C.; et al. The Genetic Polymorphisms of HLA Are Strongly Correlated with the Disease Severity after Hantaan Virus Infection in the Chinese Han Population. Clin. Dev. Immunol. 2012, 2012, 308237. [Google Scholar] [CrossRef]
- Saxenhofer, M.; Labutin, A.; White, T.A.; Heckel, G. Host Genetic Factors Associated with the Range Limit of a European Hantavirus. Mol. Ecol. 2022, 31, 252–265. [Google Scholar] [CrossRef]
- Muschetto, E.; Cueto, G.R.; Cavia, R.; Padula, P.J.; Suárez, O.V. Long-Term Study of a Hantavirus Reservoir Population in an Urban Protected Area, Argentina. Ecohealth 2018, 15, 804–814. [Google Scholar] [CrossRef]
- Xiao, H.; Tian, H.-Y.; Gao, L.-D.; Liu, H.-N.; Duan, L.-S.; Basta, N.; Cazelles, B.; Li, X.-J.; Lin, X.-L.; Wu, H.-W.; et al. Animal Reservoir, Natural and Socioeconomic Variations and the Transmission of Hemorrhagic Fever with Renal Syndrome in Chenzhou, China, 2006-2010. PLoS Negl. Trop. Dis. 2014, 8, e2615. [Google Scholar] [CrossRef]
- Pinto Junior, V.L.; Hamidad, A.M.; Albuquerque Filho, D. de O.; dos Santos, V.M. Twenty Years of Hantavirus Pulmonary Syndrome in Brazil: A Review of Epidemiological and Clinical Aspects. J. Infect. Dev. Ctries. 2014, 8, 137–142. [Google Scholar] [CrossRef]
- McELHINNEY, L.M.; Marston, D.A.; Pounder, K.C.; Goharriz, H.; Wise, E.L.; Verner-Carlsson, J.; Jennings, D.; Johnson, N.; Civello, A.; Nunez, A.; et al. High Prevalence of Seoul Hantavirus in a Breeding Colony of Pet Rats. Epidemiol. Infect. 2017, 145, 3115–3124. [Google Scholar] [CrossRef]
- Knust, B.; Brown, S.; de St. Maurice, A.; Whitmer, S.; Koske, S.E.; Ervin, E.; Patel, K.; Graziano, J.; Morales-Betoulle, M.E.; House, J.; et al. Seoul Virus Infection and Spread in United States Home-Based Ratteries: Rat and Human Testing Results From a Multistate Outbreak Investigation. J. Infect. Dis. 2020, 222, 1311–1319. [Google Scholar] [CrossRef]
- Combs, M.; Byers, K.A.; Ghersi, B.M.; Blum, M.J.; Caccone, A.; Costa, F.; Himsworth, C.G.; Richardson, J.L.; Munshi-South, J. Urban Rat Races: Spatial Population Genomics of Brown Rats (Rattus Norvegicus) Compared across Multiple Cities. Proc. Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Himsworth, C.G.; Parsons, K.L.; Jardine, C.; Patrick, D.M. Rats, Cities, People, and Pathogens: A Systematic Review and Narrative Synthesis of Literature Regarding the Ecology of Rat-Associated Zoonoses in Urban Centers. Vector Borne Zoonotic Dis. 2013, 13, 349–359. [Google Scholar] [CrossRef]
- Alburkat, H.; Smura, T.; Bouilloud, M.; Pradel, J.; Anfray, G.; Berthier, K.; Dutra, L.; Loiseau, A.; Niamsap, T.; Olander, V.; et al. Evolution and Genetic Characterization of Seoul Virus in Wild Rats Rattus Norvegicus from an Urban Park in Lyon, France 2020–2022. PLoS Neglected Trop. Dis. 2024, 18, e0012142. [Google Scholar] [CrossRef]
- Suzán, G.; Marcé, E.; Giermakowski, J.T.; Mills, J.N.; Ceballos, G.; Ostfeld, R.S.; Armién, B.; Pascale, J.M.; Yates, T.L. Experimental Evidence for Reduced Rodent Diversity Causing Increased Hantavirus Prevalence. PLoS ONE 2009, 4, e5461. [Google Scholar] [CrossRef]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of Species Diversity on Disease Risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef]
- Tkachenko, E.; Dzagurova, T.; Galieva, G.; Ivanis, V.; Kurashova, S.; Tkachenko, P.; Balkina, A.; Trankvilevsky, D.; Ishmukhametov, A. Clinical Manifestations of Hemorrhagic Fever with Renal Syndrome, Various Nosologic Forms and Issues of Hantavirus Infections Terminology. Viruses 2025, 17. [Google Scholar] [CrossRef]
- Park, M.E.; Kim, D.Y.; Seo, J.-W.; Yun, N.R.; Lee, Y.M.; Kim, C.M.; Kim, D.-M. Analysis of Clinical and Laboratory Profiles of Patients Hospitalized with Hemorrhagic Fever with Renal Syndrome in Southwestern South Korea. Am. J. Trop. Med. Hyg. 2025, 112, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.-X.; Chen, M.-J.; Sun, L. Haemorrhagic Fever with Renal Syndrome: Literature Review and Distribution Analysis in China. Int. J. Infect. Dis. 2016, 43, 95–100. [Google Scholar] [CrossRef]
- Paakkala, A.; Dastidar, P.; Ryymin, P.; Huhtala, H.; Mustonen, J. Renal MRI Findings and Their Clinical Associations in Nephropathia Epidemica: Analysis of Quantitative Findings. Eur. Radiol. 2005, 15, 968–974. [Google Scholar] [CrossRef]
- Kraft, L.; Latus, J. Hantavirus Infections—Clinical Recognition and Diagnostic Challenges. Lancet Reg. Health-Eur. 2026, 101728. [Google Scholar] [CrossRef]
- Singh, P.; Talwar, P.; Palaniyandi, R. Hantavirus Pulmonary Syndrome (HPS): A Concise Review Based on Current Knowledge and Emerging Concept. J. Appl. Pharm. Sci. 2014, 4, 122–130. [Google Scholar] [CrossRef]
- Brocato, R.L.; Hooper, J.W. Progress on the Prevention and Treatment of Hantavirus Disease. Viruses 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Geeraedts, F.; Wevers, M.; Bosma, F.; de Boer, M.; Brinkman, J.N.; Delsing, C.; GeurtsvanKessel, C.; Rockx, B.; van der Zanden, A.; Laverman, G.D. Use of a Diagnostic Puumala Virus Real-Time RT-PCR in an Orthohantavirus Endemic Region in the Netherlands. Microbiol. Spectr. 2024, 12, e0381323. [Google Scholar] [CrossRef]
- Niskanen, S.; Jääskeläinen, A.; Vapalahti, O.; Sironen, T. Evaluation of Real-Time RT-PCR for Diagnostic Use in Detection of Puumala Virus. Viruses 2019, 11, 661. [Google Scholar] [CrossRef]
- Hoornweg, T.E.; Zutt, I.; De Vries, A.; Maas, M.; Hoogerwerf, M.N.; Avšič-Županc, T.; Korva, M.; Reimerink, J.H.J.; Reusken, C.B.E.M. Development of a Comparative European Orthohantavirus Microneutralization Assay With Multi- Species Validation and Evaluation in a Human Diagnostic Cohort. Front. Cell. Infect. Microbiol. 2020, 10, 580478. [Google Scholar] [CrossRef]
- Zeng, Y.; Feng, Y.; Zhao, Y.; Zhang, X.; Yang, L.; Wang, J.; Gao, Z.; Zhang, C. An HFman Probe-Based Multiplex Reverse Transcription Loop-Mediated Isothermal Amplification Assay for Simultaneous Detection of Hantaan and Seoul Viruses. Diagnostics 2022, 12, 1925. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wei, Q.; Hu, Q.; Wen, T.; Xue, L.; Li, S.; Zeng, X.; Shi, F.; Jiao, Y.; Zhou, L. Rapid Detection of Severe Fever with Thrombocytopenia Syndrome Virus (SFTSV) Total Antibodies by up-Converting Phosphor Technology-Based Lateral-Flow Assay. Luminescence 2019, 34, 162–167. [Google Scholar] [CrossRef]
- Adedokun, G.; Alipanah, M.; Fan, Z.H. Sample Preparation and Detection Methods in Point-of-Care Devices towards Future at-Home Testing. Lab Chip 2024, 24, 3626–3650. [Google Scholar] [CrossRef] [PubMed]
- Mattar, S.; Guzmán, C.; Figueiredo, L.T. Diagnosis of Hantavirus Infection in Humans. Expert Rev. Anti-Infect. Ther. 2015, 13, 939–946. [Google Scholar] [CrossRef]
- Geeraedts, F.; Wevers, M.; Bosma, F.; Boer, M.D.; Brinkman, J.N.; Delsing, C.; GeurtsvanKessel, C.; Rockx, B.; Van Der Zanden, A.; Laverman, G.D. Use of a Diagnostic Puumala Virus Real-Time RT-PCR in an Orthohantavirus Endemic Region in the Netherlands. Microbiol. Spectr. 2024, 12, e03813-23. [Google Scholar] [CrossRef] [PubMed]
- Writer, J.V.; Kelley, P.W.; Boisson, E.V.; Hospedales, J. Caribbean Public Health Laboratory Surveillance Project: A Department of Defense-Sponsored Humanitarian Mission. Mil. Med. 2003, 168, 843–848. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef]
- Xu, B.; Yin, Q.; Ren, D.; Mo, S.; Ni, T.; Fu, S.; Zhang, Z.; Yan, T.; Zhao, Y.; Liu, J.; et al. Scientometric Analysis of Research Trends in Hemorrhagic Fever with Renal Syndrome: A Historical Review and Network Visualization. J. Infect. Public Health 2025, 18, 102647. [Google Scholar] [CrossRef]
- Tian, H.; Stenseth, N.C. The Ecological Dynamics of Hantavirus Diseases: From Environmental Variability to Disease Prevention Largely Based on Data from China. PLoS Negl. Trop. Dis. 2019, 13, e0006901. [Google Scholar] [CrossRef]
- Tosh, D.G.; Shore, R.F.; Jess, S.; Withers, A.; Bearhop, S.; Ian Montgomery, W.; McDonald, R.A. User Behaviour, Best Practice and the Risks of Non-Target Exposure Associated with Anticoagulant Rodenticide Use. J. Environ. Manag. 2011, 92, 1503–1508. [Google Scholar] [CrossRef]
- Brown, L.M.; Laco, J. Rodent Control and Public Health: A Description of Local Rodent Control Programs. J. Env. Health 2015, 78, 28–29. [Google Scholar]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and Therapeutics Against Hantaviruses. Front Microbiol. 2019, 10, 2989. [Google Scholar] [CrossRef]
- Hooper, J.W.; Brocato, R.L.; Kwilas, S.A.; Hammerbeck, C.D.; Josleyn, M.D.; Royals, M.; Ballantyne, J.; Wu, H.; Jiao, J.; Matsushita, H.; et al. DNA Vaccine-Derived Human IgG Produced in Transchromosomal Bovines Protect in Lethal Models of Hantavirus Pulmonary Syndrome. Sci. Transl. Med. 2014, 6, 264ra162. [Google Scholar] [CrossRef]
- Qamar, M.T.U.; Ahmad, S.; Khan, A.; Wei, D. Editorial: Immunotherapeutics Development against Hantaviruses. Front Immunol. 2024, 15, 1377137. [Google Scholar] [CrossRef]
- Arancibia Hernandez, F.; Fritz, R.; Sepúlveda, C.; Gavilán, J.; Espinoza, C.; Ossandon, E.; Concha, M.; Iturra, S. Impact of Early ECMO Support on Survival of Patients with Hantavirus Cardiopulmonary Syndrome in Chile. Eur. Respir. Soc. 60, 122. [CrossRef]
- Hong, Jiang; Changxing, Huang; Xuefan, Bai; Fuchun, Zhang; Bingliang, Lin; Shiwen, Wang; Zhansheng, Jia; Jingjun, Wang; Jing, Liu; Shuangsuo, Dang; et al. Expert Consensus on the Prevention and Treatment of Hemorrhagic Fever with Renal Syndrome. Infect. Dis. Immun. 2022, 02, 224–232. [Google Scholar] [CrossRef]
- Engdahl, T.B.; Binshtein, E.; Brocato, R.L.; Kuzmina, N.A.; Principe, L.M.; Kwilas, S.A.; Kim, R.K.; Chapman, N.S.; Porter, M.S.; Guardado-Calvo, P.; et al. Antigenic Mapping and Functional Characterization of Human New World Hantavirus Neutralizing Antibodies. Elife 2023, 12. [Google Scholar] [CrossRef]
- Zhang, T.; Rabhi, F.; Chen, X.; Paik, H.; MacIntyre, C.R. A Machine Learning-Based Universal Outbreak Risk Prediction Tool. Comput. Biol. Med. 2024, 169, 107876. [Google Scholar] [CrossRef] [PubMed]
- Worsley-Tonks, K.E.L.; Bender, J.B.; Deem, S.L.; Ferguson, A.W.; Fèvre, E.M.; Martins, D.J.; Muloi, D.M.; Murray, S.; Mutinda, M.; Ogada, D.; et al. Strengthening Global Health Security by Improving Disease Surveillance in Remote Rural Areas of Low-Income and Middle-Income Countries. Lancet Glob. Health 2022, 10, e579–e584. [Google Scholar] [CrossRef]
- Danforth, M.E.; Messenger, S.; Buttke, D.; Weinburke, M.; Carroll, G.; Hacker, G.; Niemela, M.; Andrews, E.S.; Jackson, B.T.; Kramer, V.; et al. Long-Term Rodent Surveillance after Outbreak of Hantavirus Infection, Yosemite National Park, California, USA, 2012. Emerg. Infect. Dis. 2020, 26, 560–567. [Google Scholar] [CrossRef]
- Gongora, V.; Trotman, M.; Thomas, R.; Max, M.; Zamora, P.A.; Lepoureau, M.T.F.; Phanord, S.; Quirico, J.; Douglas, K.; Pegram, R.; et al. The Caribbean Animal Health Network: New Tools for Harmonization and Reinforcement of Animal Disease Surveillance. Ann. N Y Acad. Sci. 2008, 1149, 12–15. [Google Scholar] [CrossRef]
- Belay, E.D.; Kile, J.C.; Hall, A.J.; Barton-Behravesh, C.; Parsons, M.B.; Salyer, S.; Walke, H. Zoonotic Disease Programs for Enhancing Global Health Security. Emerg. Infect. Dis. 2017, 23, S65–70. [Google Scholar] [CrossRef]
- Kelly, T.R.; Karesh, W.B.; Johnson, C.K.; Gilardi, K.V.K.; Anthony, S.J.; Goldstein, T.; Olson, S.H.; Machalaba, C.; Mazet, J.A.K. One Health Proof of Concept: Bringing a Transdisciplinary Approach to Surveillance for Zoonotic Viruses at the Human-Wild Animal Interface. Prev. Vet. Med. 2017, 137, 112–118. [Google Scholar] [CrossRef]
- Buregyeya, E.; Atusingwize, E.; Nsamba, P.; Musoke, D.; Naigaga, I.; Kabasa, J.D.; Amuguni, H.; Bazeyo, W. Operationalizing the One Health Approach in Uganda: Challenges and Opportunities. J. Epidemiol. Glob. Health 2020, 10, 250–257. [Google Scholar] [CrossRef]
- Lau, D.T.; Sosa, P.; Dasgupta, N.; He, H. Impact of the COVID-19 Pandemic on Public Health Surveillance and Survey Data Collections in the United States. Am. J. Public Health 2021, 111, 2118–2121. [Google Scholar] [CrossRef]
Figure 1.
Conceptual evolutionary and reservoir host relationships of major hantaviruses. Schematic illustrating the evolutionary relationships and predominant reservoir host associations of major Old World hantaviruses (blue), including Hantaan (HTNV), Seoul (SEOV), Dobrava-Belgrade (DOBV), and Puumala (PUUV) viruses, and New World hantaviruses (red/orange), including Sin Nombre (SNV), Andes (ANDV), Laguna Negra (LNV), and Choclo viruses. Major rodent reservoirs and additional shrew-, mole-, and bat-associated hantavirus-like viruses are shown to highlight the ecological diversity and host-linked evolution of hantaviruses. The figure emphasizes the complex patterns of reservoir adaptation, divergence, and spillover.
Figure 1.
Conceptual evolutionary and reservoir host relationships of major hantaviruses. Schematic illustrating the evolutionary relationships and predominant reservoir host associations of major Old World hantaviruses (blue), including Hantaan (HTNV), Seoul (SEOV), Dobrava-Belgrade (DOBV), and Puumala (PUUV) viruses, and New World hantaviruses (red/orange), including Sin Nombre (SNV), Andes (ANDV), Laguna Negra (LNV), and Choclo viruses. Major rodent reservoirs and additional shrew-, mole-, and bat-associated hantavirus-like viruses are shown to highlight the ecological diversity and host-linked evolution of hantaviruses. The figure emphasizes the complex patterns of reservoir adaptation, divergence, and spillover.

Figure 2.
Global distribution of major pathogenic hantaviruses associated with human disease, highlighting the distinct geographic patterns of Old World hantaviruses linked to hemorrhagic fever with renal syndrome (HFRS) and New World hantaviruses associated with hantavirus cardiopulmonary syndrome (HCPS/HPS). Seoul virus (SEOV) is depicted as globally distributed due to its association with urban rats and international trade. Gray hatched regions indicate areas with limited surveillance and possible underrecognized circulation, including parts of tropical Africa, the Caribbean, tropical Latin America, and South and Southeast Asia. Distribution patterns are approximate and emphasize the focal and ecologically heterogeneous nature of hantavirus transmission.
Figure 2.
Global distribution of major pathogenic hantaviruses associated with human disease, highlighting the distinct geographic patterns of Old World hantaviruses linked to hemorrhagic fever with renal syndrome (HFRS) and New World hantaviruses associated with hantavirus cardiopulmonary syndrome (HCPS/HPS). Seoul virus (SEOV) is depicted as globally distributed due to its association with urban rats and international trade. Gray hatched regions indicate areas with limited surveillance and possible underrecognized circulation, including parts of tropical Africa, the Caribbean, tropical Latin America, and South and Southeast Asia. Distribution patterns are approximate and emphasize the focal and ecologically heterogeneous nature of hantavirus transmission.

Figure 3.
Mechanisms of hantavirus pathogenesis. An overview of the major mechanisms underlying hantavirus disease following inhalation of aerosolized rodent excreta. Hantaviruses initially infect endothelial cells through integrin-associated viral entry pathways. Early innate immune responses include type I interferon signaling, macrophage and monocyte activation, dendritic cell antigen presentation, and initiation of antiviral inflammatory pathways. Adaptive immune responses involve CD4+ helper T-cell activation, CD8+ cytotoxic T-cell responses, and neutralizing antibody production, which contribute to viral control but may also amplify vascular inflammation. Progression to severe disease is characterized by cytokine amplification and immune-mediated endothelial dysfunction involving inflammatory mediators such as TNF-α, IL-6, IFN-γ, IL-1β, chemokines, and regulatory cytokine responses including IL-10. These immune processes contribute to endothelial barrier dysfunction, complement activation, thrombocytopenia, platelet–endothelial interactions, coagulation dysregulation, and increased vascular permeability with plasma extravasation and tissue edema. Disease severity reflects the balance between antiviral immunity and immune-mediated vascular injury.
Figure 3.
Mechanisms of hantavirus pathogenesis. An overview of the major mechanisms underlying hantavirus disease following inhalation of aerosolized rodent excreta. Hantaviruses initially infect endothelial cells through integrin-associated viral entry pathways. Early innate immune responses include type I interferon signaling, macrophage and monocyte activation, dendritic cell antigen presentation, and initiation of antiviral inflammatory pathways. Adaptive immune responses involve CD4+ helper T-cell activation, CD8+ cytotoxic T-cell responses, and neutralizing antibody production, which contribute to viral control but may also amplify vascular inflammation. Progression to severe disease is characterized by cytokine amplification and immune-mediated endothelial dysfunction involving inflammatory mediators such as TNF-α, IL-6, IFN-γ, IL-1β, chemokines, and regulatory cytokine responses including IL-10. These immune processes contribute to endothelial barrier dysfunction, complement activation, thrombocytopenia, platelet–endothelial interactions, coagulation dysregulation, and increased vascular permeability with plasma extravasation and tissue edema. Disease severity reflects the balance between antiviral immunity and immune-mediated vascular injury.

Figure 4.
Dual-panel schematic comparing the clinical progression and major pathophysiologic features of hemorrhagic fever with renal syndrome (HFRS)-Panel A, and hantavirus cardiopulmonary syndrome (HCPS)-Panel B.. Both syndromes share common mechanisms involving endothelial infection, immune-mediated inflammation, thrombocytopenia, and increased vascular permeability leading to capillary leak and organ dysfunction.
Figure 4.
Dual-panel schematic comparing the clinical progression and major pathophysiologic features of hemorrhagic fever with renal syndrome (HFRS)-Panel A, and hantavirus cardiopulmonary syndrome (HCPS)-Panel B.. Both syndromes share common mechanisms involving endothelial infection, immune-mediated inflammation, thrombocytopenia, and increased vascular permeability leading to capillary leak and organ dysfunction.

Figure 5.
Climate change and hantavirus emergence. Conceptual overview of how climate variability, environmental change, and reservoir host ecology influence hantavirus emergence within a One Health framework. The figure illustrates the effects of El Niño Southern Oscillation (ENSO)-related rainfall variability on hantavirus cardiopulmonary syndrome (HCPS) in Argentina, climate-sensitive drivers of hemorrhagic fever with renal syndrome (HFRS) incidence in China, and potential shifts in reservoir habitat suitability under future climate change scenarios.
Figure 5.
Climate change and hantavirus emergence. Conceptual overview of how climate variability, environmental change, and reservoir host ecology influence hantavirus emergence within a One Health framework. The figure illustrates the effects of El Niño Southern Oscillation (ENSO)-related rainfall variability on hantavirus cardiopulmonary syndrome (HCPS) in Argentina, climate-sensitive drivers of hemorrhagic fever with renal syndrome (HFRS) incidence in China, and potential shifts in reservoir habitat suitability under future climate change scenarios.

Figure 6.
Diagnostic algorithm for suspected hantavirus infections. Clinical flowchart outlining the evaluation of suspected hantavirus infection based on epidemiologic exposure, clinical presentation, laboratory findings, and confirmatory testing. The algorithm differentiates between hemorrhagic fever with renal syndrome (HFRS) and hantavirus cardiopulmonary syndrome (HCPS), while incorporating common differential diagnoses with overlapping clinical features.
Figure 6.
Diagnostic algorithm for suspected hantavirus infections. Clinical flowchart outlining the evaluation of suspected hantavirus infection based on epidemiologic exposure, clinical presentation, laboratory findings, and confirmatory testing. The algorithm differentiates between hemorrhagic fever with renal syndrome (HFRS) and hantavirus cardiopulmonary syndrome (HCPS), while incorporating common differential diagnoses with overlapping clinical features.

Figure 7.
One Health surveillance framework for hantavirus preparedness and early warning. Integrated surveillance outputs support targeted public health interventions, including rodent control, clinician alerts, healthcare preparedness, and community education, particularly in under-surveilled regions.
Figure 7.
One Health surveillance framework for hantavirus preparedness and early warning. Integrated surveillance outputs support targeted public health interventions, including rodent control, clinician alerts, healthcare preparedness, and community education, particularly in under-surveilled regions.

Figure 8.
Surveillance gaps and preparedness priorities for hantavirus detection in Caribbean and tropical regions. Regional map illustrating variations in hantavirus surveillance capacity across the Caribbean basin and selected tropical regions of Central and South America. Areas with documented hantavirus activity and regions with limited surveillance infrastructure are highlighted to emphasize the potential for underrecognized transmission. The figure also identifies priority areas for strengthening laboratory capacity, rodent surveillance, clinician awareness, and environmental monitoring.
Figure 8.
Surveillance gaps and preparedness priorities for hantavirus detection in Caribbean and tropical regions. Regional map illustrating variations in hantavirus surveillance capacity across the Caribbean basin and selected tropical regions of Central and South America. Areas with documented hantavirus activity and regions with limited surveillance infrastructure are highlighted to emphasize the potential for underrecognized transmission. The figure also identifies priority areas for strengthening laboratory capacity, rodent surveillance, clinician awareness, and environmental monitoring.


Table 1.
Major hantaviruses, reservoirs, clinical syndrome, and geographic distribution.
| Virus | Lineage | Primary Reservoir | Syndrome | Geographic Distribution |
|---|---|---|---|---|
| Hantaan (HTNV) | Old World | Apodemus agrarius | HFRS (severe) | China, Korea, Russia |
| Seoul (SEOV) | Old World | Rattus norvegicus | HFRS (mild) | Global (urban) |
| Puumala (PUUV) | Old World | Myodes glareolus | NE/HFRS (mild) | Scandinavia, W. Europe |
| Dobrava-Belgrade (DOBV) | Old World | Apodemus flavicollis | HFRS (severe) | Balkans, C. Europe |
| Sin Nombre (SNV) | New World | Peromyscus maniculatus | HCPS | North America |
| Andes (ANDV) | New World | Oligoryzomys longicaudatus | HCPS | South America |
| Laguna Negra (LNV) | New World | Calomys laucha | HCPS | Paraguay, Bolivia |
| Choclo | New World | Oligoryzomys fulvescens | HCPS | Panama, Central America |
Note: Adapted from [29].
Table 2.
Clinical differences between HFRS and HCPS.
| Feature | HFRS | HCPS |
|---|---|---|
| Primary organ | Kidney | Lungs/Cardiovascular |
| Clinical phases | 5 (febrile, hypotensive, oliguric, diuretic, convalescent) | 2 (prodromal, cardiopulmonary) |
| Key manifestations | AKI, hemorrhage, hypotension | Pulmonary edema, respiratory failure |
| Thrombocytopenia | Yes | Yes (severe) |
| Mortality | 0.5–15% | 30–50% |
| Associated viruses | HTNV, PUUV, SEOV, DOBV | SNV, ANDV, Choclo, Laguna Negra |
| Geographic focus | Asia, Europe | Americas |
Table 3.
Laboratory diagnostic approaches for hantavirus detection.
| Method | Specimen | Timing | Sensitivity | Specificity | BSL Required |
|---|---|---|---|---|---|
| IgM ELISA | Serum | Acute (≥3 days) | 85–95% | 90–95% | BSL-2 |
| IgG ELISA | Serum | Acute/convalescent | 90–98% | 90–98% | BSL-2 |
| RT-PCR | Blood, tissue | Early acute | 70–90% | >95% | BSL-2 (post-extraction) |
| Immunohistochemistry | Tissue | Any (autopsy) | 80–95% | >95% | BSL-2 |
| Viral culture | Blood, tissue | Acute phase | Variable | 100% | BSL-3 |
| Rapid lateral flow | Whole blood | Acute phase | 70–85% | 85–95% | BSL-2 |
Table 4.
Differential diagnosis of hantavirus infection in tropical settings.
| Condition | Shared Features | Distinguishing Features | Key Test |
|---|---|---|---|
| Dengue fever | Fever, thrombocytopenia, myalgia | Rash, NS1 antigen, no renal failure | NS1 ELISA, RT-PCR |
| Leptospirosis | Fever, renal failure, myalgia | Jaundice, conjunctival suffusion | MAT, IgM ELISA |
| Malaria | Fever, thrombocytopenia, myalgia | Cyclic fever, splenomegaly | Blood smear, RDT |
| Influenza | Fever, myalgia, respiratory | No renal failure, no thrombocytopenia | Rapid influenza test |
| COVID-19 | Fever, respiratory, myalgia | SARS-CoV-2 exposure, anosmia | RT-PCR |
| Rickettsial disease | Fever, headache, thrombocytopenia | Rash, eschar, tick exposure | Serology, PCR |
| Hantavirus | Fever, thrombocytopenia, renal/pulmonary | Rodent exposure, pulmonary edema, AKI | IgM ELISA, RT-PCR |
Table 5.
Surveillance challenges and proposed solutions in Caribbean and tropical regions.
| Challenge | Impact | Proposed Solution |
|---|---|---|
| Limited molecular diagnostics | Missed cases, delayed outbreak detection | Regional RT-PCR platform sharing, training programs |
| No baseline seroprevalence data | Unknown disease burden | Population-based serosurveys |
| Clinical overlap with dengue/leptospirosis | Systematic misdiagnosis | Multiplex diagnostic panels, clinician education |
| No rodent surveillance | Undetected reservoir activity | Integrated rodent trapping and molecular testing |
| Fragmented reporting systems | Delayed outbreak recognition | Syndromic surveillance integration |
| Limited biosafety infrastructure | Inability to manage BSL-3 specimens | Biosafety capacity building, regional reference labs |
| Extreme weather events | Post-disaster exposure spikes | Disaster-response hantavirus surveillance protocols |
Table 6.
One Health preparedness strategies for hantavirus surveillance.
| Sector | Key Activities | Expected Outcomes |
|---|---|---|
| Human health | Case surveillance, clinician training, syndromic systems | Improved case detection, reduced diagnostic delays |
| Veterinary/wildlife | Rodent trapping, viral testing, species mapping | Reservoir identification, early outbreak warning |
| Environmental | GIS mapping, climate modeling, environmental sampling | Risk area identification, predictive modeling |
| Laboratory | RT-PCR capacity, serology, biosafety, reference labs | Rapid confirmation, quality-assured diagnostics |
| Public health policy | Regional networks, data sharing, standardized protocols | Coordinated outbreak response, harmonized surveillance |
| Research | Seroprevalence studies, genomic surveillance, vaccine development | Evidence base, preparedness tools |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



