Submitted:
06 May 2026
Posted:
07 May 2026
You are already at the latest version
Abstract
Blood can clot into anomalous, fibrinolysis-resistant forms that arise from prothrombotic seeding areas, including damaged cellular debris and membrane-derived surfaces, giving rise to what we have termed fibrinaloid microclot complexes (colloquially: microclots).Their proteolytic resistance is due to the fact that they are amyloid in nature, and they can also entrap inhibitors of proteolysis. They consist of a variety of proteins besides the expected fibrin, and are highly enriched for other amyloidogenic proteins (in contrast to normal clots, whose proteome largely reflects the soluble plasma proteome). They also contain DNA in the form of neutrophil extracellular traps (NETs). Importantly, fibrinaloid microclot complexes are heterogeneous structures comprising multiple phenotypic forms, including those that nucleate and grow on cellular debris such as damaged membranes, microparticles, and immune-derived material. These debris-associated complexes act as catalytic scaffolds that recruit fibrin(ogen) and inflammatory molecules, thereby amplifying amyloidogenic transformation and prothrombotic activity. Fibrinaloid microclot complexes have been reported in a widening range of chronic inflammatory and thrombo-inflammatory diseases in which they have been sought, and are highly enriched for amyloidogenic proteins. Additionally, the thrombi extracted from ischaemic stroke also contain proteins in an amyloid form, implying that such macroclots can form via the accretion of microclots that already contain amyloid. We here show that these microclots exhibit a classical ‘apple-green’ birefringence when stained with the dye Congo red. The urgent task now is to find means of inhibiting the transition to amyloid forms during the clotting process.
Keywords:
amyloid
; microclots
; inflammation
; oxidative stress
; microcirculation
Image for thumbnail

Introduction
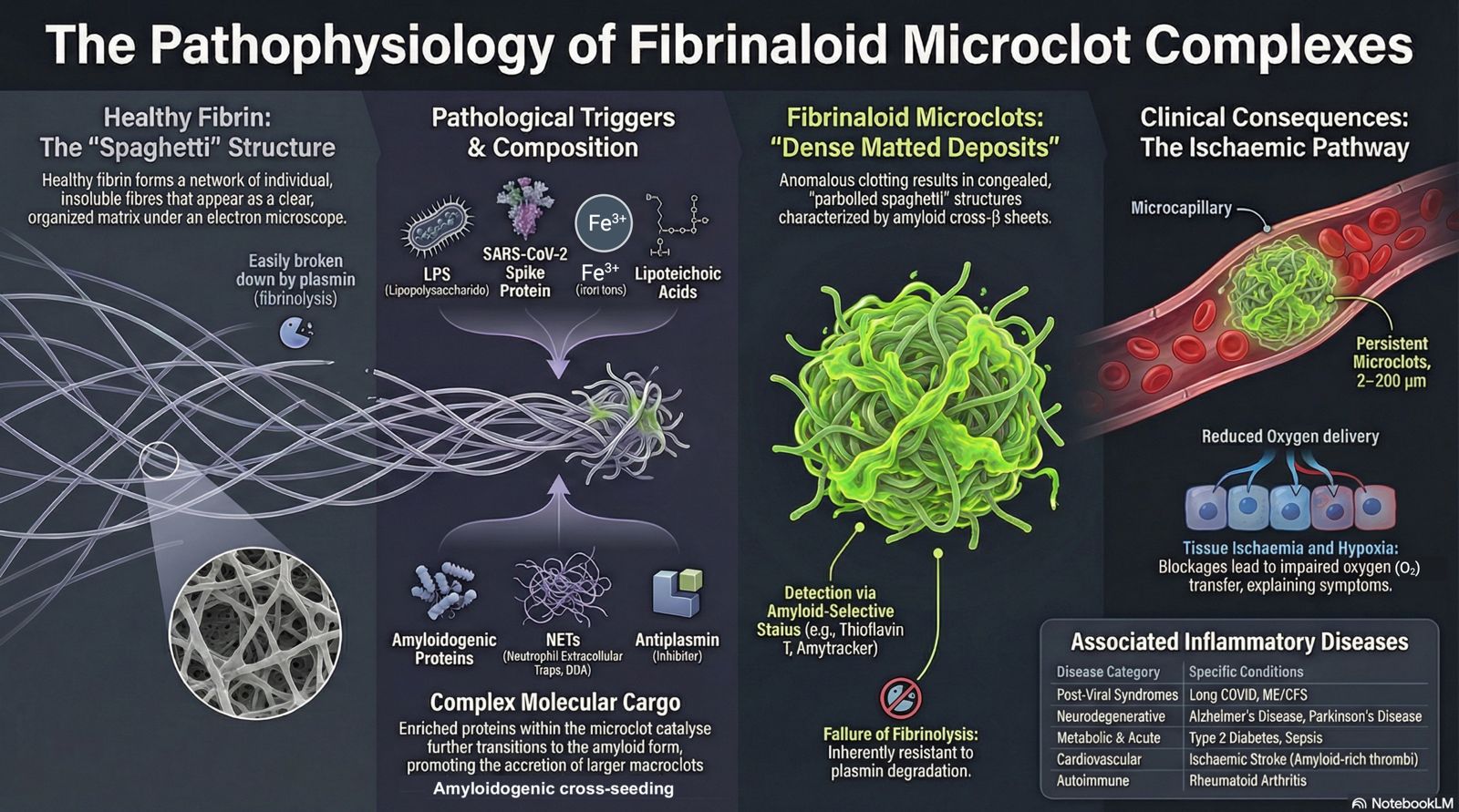
Blood clotting is traditionally understood as a thrombin-driven process in which fibrin polymers form structured networks that are susceptible to regulated fibrinolysis; however, under inflammatory conditions, clotting can also originate from prothrombotic seeding areas that give rise to structurally distinct, fibrinolysis-resistant assemblies. In healthy haemostasis these fibres form branching networks whose architecture is influenced by fibrinogen concentration, thrombin generation, flow and cross-linking. In the electron microscope these appear like spaghetti [1,2].
However, blood can also clot into anomalous forms that are significantly resistant to fibrinolysis and that in the electron microscope appear instead as ‘dense matted deposits’ that take on the appearance of parboiled spaghetti that has been allowed to congeal (e.g. [3,4,5]).
It transpired that that this form was in fact amyloid in nature [6], involving the kind of cross-b sheets seen in prion diseases and classical amyloidoses [7]. These observations suggest that pathological clotting does not simply reflect dysregulated thrombin activity, but rather emerges from surface-driven nucleation processes in which damaged biological interfaces act as catalytic scaffolds for amyloidogenic fibrin(ogen) assembly. We have referred to these microclot complexes as fibrinaloid, and they typically possess a diameter in the range 2-200 mm. They also contain neutrophil extracellular traps (NETs [8]). Importantly, these complexes represent heterogeneous phenotypic forms, reflecting their origins on distinct seeding substrates such as membrane debris, microparticles, NET-derived nucleoprotein scaffolds, and plasma protein aggregates.
The fibrinaloid microclot complexes are typically observed using fluorogenic amyloid stains such as thioflavin T [9,10,11] or the oligothiophene ‘Amytracker’ dyes [12,13,14]. As is well known, amyloids are much more resistant to normal proteolysis than the soluble forms formed at the ribosome. Figure 1 shows examples of representative microclot complexes stained with Thioflavin T and detected with imaging flow cytometry. Together, these findings define a mechanistically distinct mode of clot formation in which amyloidogenic, surface-seeded complexes precede and potentially drive the development of larger, clinically relevant thrombotic structures.
Fibrinaloid microclot complexes have now been reported across a widening range of inflammatory and thrombo-inflammatory conditions [16] (Table 1), including acute COVID-19, Long COVID, sepsis, type 2 diabetes, Parkinson’s disease, rheumatoid arthritis and Alzheimer-related syndromes, with support coming both from our own studies and from independent groups. However, while the presence of fibrinaloid microclot complexes has been observed across these diverse conditions, detailed mechanistic characterization of their phenotypes and their origins from specific prothrombotic seeding areas has not been uniformly established.
To date, such phenotypic resolution, including the identification of distinct seeding substrates such as cellular debris, microparticles, and nucleoprotein complexes, has been most clearly demonstrated in COVID-19, Long COVID, and type 2 diabetes. In contrast, in other disease settings, fibrinaloid microclots have largely been described at the level of presence, morphology, or proteomic enrichment, without explicit discrimination of their underlying phenotypic diversity or seeding mechanisms.
The thrombi causing ischaemic stroke also display amyloid [17,18], and the fibrinaloid microclots can be formed using just purified fibrinogen, thrombin, and a suitable trigger molecule [6,19].
Several triggers of amyloidogenic clotting have been observed experimentally, including ferric ions [4,54,55,56], bacterial lipopolysaccharide (LPS) [6], lipoteichoic acids [14], and the SARS-CoV-2 spike protein [57]. These have all been found to trigger the amyloidogenic clotting, often at minuscule levels [6].
The SARS-CoV-2 spike protein is itself highly amyloidogenic [58,59], its amyloidogenicity (in different strains of SARS-CoV-2) being related to the virulence of the strains [60]. Taken together, these findings are consistent with the view that clot amyloidogenesis can lie on an important disease pathway.
It is recognised that thioflavin T, while seen as a gold standard amyloid stain [11,61], is not entirely specific for amyloid [37]. To this end, we have also employed the more selective oligothiophene ‘Amytracker’ dyes, again showing that they stained fibrinaloid microclot complexes (e.g. [12,13,14,62]). Another ‘classical’ amyloid dye is Congo Red, that shows an ‘apple-green’ birefringence when illuminated with polarised light (e.g. [63,64,65], cf. [66]). We here show, for the first time, fibrinaloid microclots assessed in this way with Congo Red.
Figure 2 shows images of a fibrinaloid microclot created in vitro (as per [6,67], stained with the classical amyloid dye Congo Red [68] and assessed using a polarising microscope as described in Methods, below. It displays the classical ‘apple green’ birefringence, providing further evidence for the amyloid nature of the microclots.
In Long COVID, increased FMC burdens have been reported in platelet-poor plasma by more than one group [23,24,25,26], although with inter-individual variability and continuing debate over assay standardisation and interpretation. These observations are consistent with the broader concept that persistent thrombo-inflammation, endothelial activation and impaired fibrinolysis may contribute to symptom burden in at least a subset of patients. Within this framework, microcirculatory impairment provides one plausible route by which diverse symptoms may arise, although the degree of causal contribution is still being defined.
The effect of these insoluble FMCs is potentially to partially block or interfere with blood flow in microcapillaries (the ‘microcirculation’) and hence O2 transfer to tissues. Taking Long COVID as an example, such a blockage can straightforwardly and self-consistently explain a great many symptoms that accompany this highly variable syndrome, including fatigue [27], post-exertional malaise [28], autoimmunity [29], atrial fibrillation [69], fibromyalgia [70], POTS [71], and tinnitus [72].
Emerging evidence also indicates that FMCs represent a spectrum of structurally and mechanistically distinct entities rather than a single homogeneous clotting product. While earlier work has focused primarily on fibrin(ogen)-derived amyloid formation, it is now clear that a substantial proportion of these complexes originate from prothrombotic seeding areas, including cellular debris, membrane fragments, microparticles, and nucleoprotein immune structures. These debris-associated complexes are fundamentally different from classical fibrin networks in that they do not form through thrombin-driven polymerisation, do not integrate into organised fibrin fibre architectures, and are largely resistant to conventional plasmin-mediated fibrinolysis. In parallel, a related but distinct process occurs in which soluble inflammatory and amyloidogenic molecules interact with fibrinogen and, upon thrombin activation, become incorporated into fibrin networks, giving rise to the dense matted deposits previously described. These observations support the existence of at least two overlapping but mechanistically distinct pathways: (i) surface-seeded formation of debris-associated fibrinaloid complexes, and (ii) thrombin-mediated formation of structurally altered fibrin networks incorporating inflammatory components. This distinction is critical, as conventional coagulation and fibrinolysis assays are likely to capture only the latter, thereby underestimating or entirely missing the presence of debris-associated fibrinaloid structures and contributing to ongoing discrepancies in the literature.
We have used the Amylogram program [73,74] to assess the amyloidogenicity of individual proteins (the output is a score between 0 and 1, with anything over ca 0.75 being significantly and experimentally amyloidogenic [75]. The proteome of normal thrombi [76] largely reflects that of the soluble plasma proteome. However, the proteomes of the various fibrinaloid microclot complexes are very different, with the proteins that are enriched in them being themselves highly amyloidogenic [77,78,79]. Table 2 shows proteins enriched in the microclots, a dataset [77] to which we have added their Amylogram scores. Each of these exceeds 0.75, some by a considerable margin. It is likely that this is caused by cross-seeding of these amyloidogenic proteins catalysing the transition to the amyloid form, and that the macroclots in ischaemic stroke and other cardiovascular diseases can form via accretion of these microclots. Of course their amyloid nature again straightforwardly explains their resistance to the normal fibrinolysis that would be catalysed by plasmin. A pictorial summary is given in Figure 3.
Although the broader disease-spanning framework has been developed largely through our own work, recent reports from independent groups in sepsis and Long COVID indicate that the phenomenon is reproducible beyond a single laboratory context. At the same time, the field would benefit greatly from further methodological standardisation, head-to-head comparison of assays, and additional external replication across disease settings.
In particular, the next stage is to develop methods that are not simply anticoagulating (though such can be efficacious [80]), but that can inhibit the transition to the amyloid form.
Methods
To create and analyse fibrinaloid microclots with Congo Red in vitro, 30 ng/mL of bacterial LPS were added to purified FBG at 2.6 mg/mL. To this, Congo Red was added at 0.3 mM, followed by 7U of Thrombin. All concentrations stated are final. Images were taken on an Axio Observer.Z1 (Zeiss, Jena, Germany) microscope using a halogen lamp (HAL100) and a polariser filter with a GXCAM-3 (GT Vision, Wickhambrook, UK). Images were acquired with the software GXCam version 7.3. All images were taken with a Plan-APOCHROMAT FLUAR 20x 0.75 DIC and saved in .tif format. The methods and data for Figure 1 were obtained using imaging flow cytometry, the image being taken from a CC-BY 4.0 publication [15].
Conclusions
FMCs have now been reported across a widening range of inflammatory and thrombo-inflammatory conditions (Kell and Pretorius 2018a) (Table 1), including acute COVID-19, Long COVID, sepsis, type 2 diabetes, Parkinson’s disease, rheumatoid arthritis and Alzheimer-related syndromes, with support coming both from our own studies and from independent groups. However, while the presence of fibrinaloid microclot complexes has been observed across these diverse conditions, detailed mechanistic characterization of their phenotypes and their origins from specific prothrombotic seeding areas has not been uniformly established, although our group has identified the need for better phenotyping [36]. To date, such phenotypic resolution, including the identification of distinct seeding substrates such as cellular debris, microparticles, and nucleoprotein complexes, has been most clearly demonstrated in COVID-19, Long COVID, and type 2 diabetes. In contrast, in other disease settings, fibrinaloid microclots have largely been described at the level of presence, morphology, or proteomic enrichment, without explicit discrimination of their underlying phenotypic diversity or seeding mechanisms.
This distinction is critical, as it indicates that FMCs are not a single pathological entity but a spectrum of mechanistically distinct, surface-seeded assemblies, and that resolving their phenotypic diversity and seeding origins is essential for understanding their role in disease pathogenesis, as well as for developing targeted diagnostic and therapeutic strategies.
Author Contributions
Conceptualization, DBK and EP; Resources, DBK and EP; Data generation for Figure 2 JMG; Writing – Original Draft Preparation, DBK; Writing – Review & Editing, all authors; Visualization, EP; Funding Acquisition, DBK and EP.
Funding
We thank the Balvi Foundation for financial support. The content and findings reported and illustrated are the sole deduction, view and responsibility of the researchers and do not reflect the official position and sentiments of the funders. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
The authors gratefully acknowledge Marco Marcello of the LIV-SRF Centre for Cell Imaging https://www.liverpool.ac.uk/research/facilities/shared-research-facilities/centre-for-cell-imaging/ for assistance with the polarised light microscopy.
Declaration of Competing Interests
EP holds a patent application for the use of fluorescence microcopy in microclot imaging.
References
- Weisel, J. W.; Litvinov, R. I. Fibrin Formation, Structure and Properties. Subcell. Biochem. 2017, 82, 405–456. [Google Scholar] [CrossRef]
- Pretorius, E.; Page, M. J.; Mbotwe, S.; Kell, D. B. Lipopolysaccharide-binding protein (LBP) can reverse the amyloid state of fibrin seen or induced in Parkinson’s disease. PlosOne 2018, 13, e0192121. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Oberholzer, H. M.; van der Spuy, W. J.; Meiring, J. H. The changed ultrastructure of fibrin networks during use of oral contraception and hormone replacement. J. Thromb. Thrombolysis 2010, 30, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Vermeulen, N.; Bester, J.; Lipinski, B.; Kell, D. B. A novel method for assessing the role of iron and its functional chelation in fibrin fibril formation: the use of scanning electron microscopy. Toxicol. Mech. Methods 2013, 23, 352–359. [Google Scholar] [CrossRef]
- Swanepoel, A. C.; Visagie, A.; de Lange, Z.; Emmerson, O.; Nielsen, V. G.; Pretorius, E. The clinical relevance of altered fibrinogen packaging in the presence of 17beta-estradiol and progesterone. Thromb. Res. 2016, 146, 23–34. [Google Scholar] [CrossRef]
- Pretorius, E.; Mbotwe, S.; Bester, J.; Robinson, C. J.; Kell, D. B. Acute induction of anomalous and amyloidogenic blood clotting by molecular amplification of highly substoichiometric levels of bacterial lipopolysaccharide. J. R Soc. Interface 2016, 123, 20160539. [Google Scholar] [CrossRef]
- Kell, D. B.; Pretorius, E. Proteins behaving badly. Substoichiometric molecular control and amplification of the initiation and nature of amyloid fibril formation: lessons from and for blood clotting. Progr Biophys. Mol. Biol. 2017, 123, 16–41. [Google Scholar] [CrossRef] [PubMed]
- Thierry, A. R.; Usher, T.; Sanchez, C.; Turner, S.; Venter, C.; Pastor, B.; Waters, M.; Thompson, A.; Mirandola, A.; Pisareva, E.; Prevostel, C.; Laubscher, G. J.; Kell, D. B.; Pretorius, E. Circulating microclots are structurally associated with Neutrophil Extracellular Traps and their amounts are elevated in Long COVID patients. J. Med. Virol. 2025, 97, e70613. [Google Scholar] [CrossRef]
- Biancalana, M.; Koide, S. Molecular mechanism of Thioflavin-T binding to amyloid fibrils. Biochim Biophys. Acta 2010, 1804, 1405–1412. [Google Scholar] [CrossRef]
- Gade Malmos, K.; Blancas-Mejia, L. M.; Weber, B.; Buchner, J.; Ramirez-Alvarado, M.; Naiki, H.; Otzen, D. ThT 101: a primer on the use of thioflavin T to investigate amyloid formation. Amyloid 2017, 24, 1–16. [Google Scholar] [CrossRef]
- Xue, C.; Lin, T. Y.; Chang, D.; Guo, Z. Thioflavin T as an amyloid dye: fibril quantification, optimal concentration and effect on aggregation. R Soc. Open Sci. 2017, 4, 160696. [Google Scholar] [CrossRef] [PubMed]
- de Waal, G. M.; Engelbrecht, L.; Davis, T.; de Villiers, W. J. S.; Kell, D. B.; Pretorius, E. Correlative Light-Electron Microscopy detects lipopolysaccharide and its association with fibrin fibres in Parkinson's Disease, Alzheimer's Disease and Type 2 Diabetes Mellitus. Sci. Rep. 2018, 8, 16798. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Page, M. J.; Engelbrecht, L.; Ellis, G. C.; Kell, D. B. Substantial fibrin amyloidogenesis in type 2 diabetes assessed using amyloid-selective fluorescent stains. Cardiovasc Diabetol. 2017, 16, 141. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Page, M. J.; Hendricks, L.; Nkosi, N. B.; Benson, S. R.; Kell, D. B. Both lipopolysaccharide and lipoteichoic acids potently induce anomalous fibrin amyloid formation: assessment with novel Amytracker™ stains. J. R Soc. Interface 2018, 15, 20170941. [Google Scholar] [CrossRef]
- Turner, S.; Laubscher, G. J.; Khan, M. A.; Kell, D. B.; Pretorius, E. Accelerating discovery: A novel flow cytometric method for detecting fibrin(ogen) amyloid microclots using long COVID as a model. Heliyon 2023, 9, e19605. [Google Scholar] [CrossRef]
- Kell, D. B.; Pretorius, E. No effects without causes. The Iron Dysregulation and Dormant Microbes hypothesis for chronic, inflammatory diseases. Biol. Rev. 2018, 93, 1518–1557. [Google Scholar] [CrossRef]
- Grixti, J. M.; Chandran, A.; Pretorius, J. H.; Walker, M.; Sekhar, A.; Pretorius, E.; Kell, D. B. The clots removed from ischaemic stroke patients by mechanical thrombectomy are amyloid in nature. medRxiv 2024. [Google Scholar] [CrossRef]
- Grixti, J. M.; Chandran, A.; Pretorius, J. H.; Walker, M.; Sekhar, A.; Pretorius, E.; Kell, D. B. Amyloid presence in acute ischemic stroke thrombi: observational evidence for fibrinolytic resistance. Stroke 2025, 56, e165–e167. [Google Scholar] [CrossRef]
- Grixti, J. M.; Theron, C. W.; Salcedo-Sora, J. E.; Pretorius, E.; Kell, D. B. Automated microscopic measurement of fibrinaloid microclots and their degradation by nattokinase, the main natto protease. J. Exp. Clin. Appl. Chin. Med. 2024, 5, 30–55. [Google Scholar] [CrossRef]
- Bunch, C. M.; Moore, E. E.; Moore, H. B.; Neal, M. D.; Thomas, A. V.; Zackariya, N.; Zhao, J.; Zackariya, S.; Brenner, T. J.; Berquist, M.; Buckner, H.; Wiarda, G.; Fulkerson, D.; Huff, W.; Kwaan, H. C.; Lankowicz, G.; Laubscher, G. J.; Lourens, P. J.; Pretorius, E.; Kotze, M. J.; Moolla, M. S.; Sithole, S.; Maponga, T. G.; Kell, D. B.; Fox, M.; Gillespie, L.; Khan, R. Z.; Mamczak, C. N.; March, R.; Macias, R.; Bull, B. S.; Walsh, M. M. Immuno-thrombotic Complications of COVID-19: Implications for Timing of Surgery and Anticoagulation. Front Surg. 2022, 9, 889999. [Google Scholar] [CrossRef]
- Grobler, C.; Maphumulo, S. C.; Grobbelaar, L. M.; Bredenkamp`, J.; Laubscher, J.; Lourens, P. J.; Steenkamp, J.; Kell, D. B.; Pretorius, E. COVID-19: The Rollercoaster of Fibrin(ogen), D-dimer, von Willebrand Factor, P-selectin and Their Interactions with Endothelial Cells, Platelets and Erythrocytes. Int. J. Mol. Sci. 2020, 21, 5168. [Google Scholar] [CrossRef]
- Pretorius, E.; Venter, C.; Laubscher, G. J.; Lourens, P. J.; Steenkamp, J.; Kell, D. B. Prevalence of readily detected amyloid blood clots in ‘unclotted’ Type 2 Diabetes Mellitus and COVID-19 plasma: A preliminary report. Cardiovasc Diabetol. 2020, 19, 193. [Google Scholar] [CrossRef] [PubMed]
- Dalton, C. F.; de Oliveira, M. I. R.; Stafford, P.; Peake, N.; Kane, B.; Higham, A.; Singh, D.; Jackson, N.; Davies, H.; Price, D.; Duncan, R.; Tattersall, N.; Barnes, A.; Smith, D. P. Increased fibrinaloid microclot counts in platelet-poor plasma are associated with Long COVID. medRxiv 2024, 2004.24305318. [Google Scholar] [CrossRef]
- Okuducu, Y. K.; Boribong, B.; Ellett, F.; Hajizadeh, S.; VanElzakker, M.; Haas, W.; Pillai, S.; Fasano, A.; Irimia, D.; Yonker, L. Evidence Circulating Microclots and Activated Platelets Contribute to Hyperinflammation Within Pediatric Post Acute Sequala of COVID. Am. J. Respir. Crit. Care Med. 2024, 209, A2247. [Google Scholar]
- Irimia, D.; Gill, K.; Alvarez-Carcamo, B.; Steifman, C.; Swank, Z.; Walt, D.; VanElzakker, M.; Yonker, L. Quantification of fibrinaloid clots in plasma from pediatric Long COVID patients using a microfluidic assay. Research 2025. [Google Scholar] [CrossRef]
- Steifman, C. B.; Alvarez-Carcamo, B.; Verma, S.; McCarthy, R.; Guthrie, L. B.; Gill, K. K.; Swank, Z.; Walt, D. R.; Grabowski, E. F.; Fasano, A.; VanElzakker, M. B.; Irimia, D.; Yonker, L. M. Endovascular profiles linked to neutrophil activation in children and young adults with long COVID. Pediatr. Res. 2026. [Google Scholar] [CrossRef]
- Kell, D. B.; Laubscher, G. J.; Pretorius, E. A central role for amyloid fibrin microclots in long COVID/PASC: origins and therapeutic implications. Biochem J. 2022, 479, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Kell, D. B.; Pretorius, E. The potential role of ischaemia-reperfusion injury in chronic, relapsing diseases such as rheumatoid arthritis, long COVID and ME/CFS: evidence, mechanisms, and therapeutic implications. Biochem J. 2022, 479, 1653–1708. [Google Scholar] [CrossRef] [PubMed]
- Kell, D. B.; Pretorius, E. Are fibrinaloid microclots a cause of autoimmunity in Long Covid and other post-infection diseases? Biochem J. 2023, 480, 1217–1240. [Google Scholar] [CrossRef]
- Kruger, A.; Vlok, M.; Turner, S.; Venter, C.; Laubscher, G. J.; Kell, D. B.; Pretorius, E. Proteomics of fibrin amyloid microclots in Long COVID/ Post-Acute Sequelae of COVID-19 (PASC) shows many entrapped pro-inflammatory molecules that may also contribute to a failed fibrinolytic system. Cardiovasc Diabetol. 2022, 21, 190. [Google Scholar] [CrossRef]
- Pretorius, E.; Vlok, M.; Venter, C.; Bezuidenhout, J. A.; Laubscher, G. J.; Steenkamp, J.; Kell, D. B. Persistent clotting protein pathology in Long COVID/ Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. Cardiovasc Diabetol. 2021, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Venter, C.; Laubscher, G. J.; Kotze, M. J.; Oladejo, S.; Watson, L. R.; Rajaratnam, K.; Watson, B. W.; Kell, D. B. Prevalence of symptoms, comorbidities, fibrin amyloid microclots and platelet pathology in individuals with Long COVID/ Post-Acute Sequelae of COVID-19 (PASC). Cardiovasc Diabetol. 2022, 21, 148. [Google Scholar] [CrossRef]
- Turner, S.; Khan, M. A.; Putrino, D.; Woodcock, A.; Kell, D. B.; Pretorius, E. Long COVID: pathophysiological factors and abnormal coagulation. Trends Endocrinol. Metab. 2023, 34, 321–344. [Google Scholar] [CrossRef]
- Booyens, R. M.; Vlok, M.; Bester, C.; Hira, R.; Khan, M. A.; Kell, D. B.; Raj, S. R.; Pretorius, E. Post-translational modifications within fibrinaloid microclot complexes distinguish Pre-COVID-19 Postural Orthostatic Tachycardia Syndrome (POTS), Long COVID, and Long COVID-POTS and reveal disease-specific molecular pathways. bioRxiv 2025, 2025, 2029.696828. [Google Scholar] [CrossRef]
- Kruger, A.; Joffe, D.; Lloyd-Jones, G.; Khan, M. A.; Salamon, S.; Laubscher, G. J.; Putrino, D.; Kell, D. B.; Pretorius, E. Vascular Pathogenesis in Acute and Long COVID: Current Insights and Therapeutic Outlook. Semin Thromb. Hemost. 2025, 51, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.; Kell, L.; Slaghekke, A.; Wust, R. C.; Fielding, B. C.; Kell, D. B.; Pretorius, E. Virus-induced endothelial senescence as a cause and driving factor for ME/CFS and long COVID: mediated by a dysfunctional immune system. Cell Death Dis. 2026, 17, 16. [Google Scholar] [CrossRef]
- Thierry, A. R.; Usher, T.; Sanchez, C.; Turner, S.; Venter, C.; Pastor, B.; Waters, M.; Thompson, A.; Mirandola, A.; Pisareva, E.; Prevostel, C.; Laubscher, G. J.; Kell, D. B.; Pretorius, E. Circulating Microclots Are Structurally Associated With Neutrophil Extracellular Traps and Their Amounts Are Elevated in Long COVID Patients. J. Med. Virol. 2025, 97, e70613. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Naidoo, C. A.; Usher, T. J.; Kruger, A.; Venter, C.; Laubscher, G. J.; Khan, M. A.; Kell, D. B.; Pretorius, E. Increased Levels of Inflammatory and Endothelial Biomarkers in Blood of Long COVID Patients Point to Thrombotic Endothelialitis. Semin Thromb. Hemost. 2024, 50, 288–294. [Google Scholar] [CrossRef]
- Schofield, J.; Abrams, S. T.; Jenkins, R.; Lane, S.; Wang, G.; Toh, C. H. Microclots, as defined by amyloid-fibrinogen aggregates, predict risks of disseminated intravascular coagulation and mortality. Blood Adv. 2024, 8, 2499–2508. [Google Scholar] [CrossRef]
- Kell, D. B.; Pretorius, E. To what extent are the terminal stages of sepsis, septic shock, SIRS, and multiple organ dysfunction syndrome actually driven by a toxic prion/amyloid form of fibrin? Semin Thromb. Hemost. 2018, 44, 224–238. [Google Scholar] [CrossRef]
- Grixti, J. M.; Chandran, A.; Pretorius, J.-H.; Walker, M.; Sekhar, A.; Pretorius, E.; Kell, D. B. The clots removed from ischaemic stroke patients by mechanical thrombectomy are amyloid in nature. medRxiv 2024, 2024, 2001.24316555. [Google Scholar] [CrossRef]
- Grixti, J. M.; Chandran, A.; Pretorius, J. H.; Walker, M.; Sekhar, A.; Pretorius, E.; Kell, D. B. Amyloid Presence in Acute Ischemic Stroke Thrombi: Observational Evidence for Fibrinolytic Resistance. Stroke 2025, 56, e165–e167. [Google Scholar] [CrossRef]
- Pretorius, E.; Bester, J.; Vermeulen, N.; Alummoottil, S.; Soma, P.; Buys, A. V.; Kell, D. B. Poorly controlled type 2 diabetes is accompanied by significant morphological and ultrastructural changes in both erythrocytes and in thrombin-generated fibrin: implications for diagnostics. Cardiovasc Diabetol. 2015, 134, 30. [Google Scholar] [CrossRef]
- Pretorius, E.; Bester, J. Viscoelasticity as a measurement of clot structure in poorly controlled type 2 diabetes patients: towards a precision and personalized medicine approach. Oncotarget 2016, 7, 50895–50907. [Google Scholar] [CrossRef]
- Soma, P.; Pretorius, E. Interplay between ultrastructural findings and atherothrombotic complications in type 2 diabetes mellitus. Cardiovasc Diabetol. 2015, 14, 96. [Google Scholar] [CrossRef]
- Adams, B.; Nunes, J. M.; Page, M. J.; Roberts, T.; Carr, J.; Nell, T. A.; Kell, D. B.; Pretorius, E. Parkinson’s disease: a systemic inflammatory disease accompanied by bacterial inflammagens. Front Ag. Neurosci. 2019, 11, 210. [Google Scholar] [CrossRef]
- Grobler, C.; van Tongeren, M.; Gettemans, J.; Kell, D.; Pretorius, E. Alzheimer-type dementia: a systems view provides a unifying explanation of its development. J. Alz Dis. 2023, 91, 43–70. [Google Scholar] [CrossRef]
- Itzhaki, R. F.; Lathe, R.; Balin, B. J.; Ball, M. J.; Braak, H.; Bearer, E. L.; Bullido, M. J.; Carter, C.; Clerici, M.; Cosby, S. L.; Del Tredici, K.; Field, H.; Fulop, T.; Grassi, C.; Griffin, W. S. T.; Haas, J.; Hudson, A. P.; Kamer, A.; Kell, D. B.; Licastro, F.; Letenneur, L.; Lövheim, H.; Mancuso, R.; Miklossy, J.; Otth, C.; Palamara, A. T.; Perry, G.; Preston, C.; Pretorius, E.; Strandberg, T.; Tabet, N.; Taylor-Robinson, S. D.; Whittum-Hudson, J. A. Microbes and Alzheimer's Disease. J. Alzheimers Dis. 2016, 51, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Bester, J.; Kell, D. B. A bacterial component to Alzheimer-type dementia seen via a systems biology approach that links iron dysregulation and inflammagen shedding to disease. J. Alzheimers Dis. 2016, 53, 1237–1256. [Google Scholar] [CrossRef] [PubMed]
- van Vuuren, M. J.; Nell, T. A.; Carr, J. A.; Kell, D. B.; Pretorius, E. Iron dysregulation and inflammagens related to oral and gut health are central to the development of Parkinson’s disease. Biomolecules 2021, 11, 30. [Google Scholar] [CrossRef]
- Pretorius, E.; Akeredolu, O.-O.; Soma, P.; Kell, D. B. Major involvement of bacterial components in rheumatoid arthritis and its accompanying oxidative stress, systemic inflammation and hypercoagulability. Exp. Biol. Med. 2017, 242, 355–373. [Google Scholar] [CrossRef] [PubMed]
- Nunes, J. M.; Kruger, A.; Proal, A.; Kell, D. B.; Pretorius, E. The occurrence of hyperactivated platelets and fibrinaloid microclots in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Pharmaceuticals 2022, 15, 931. [Google Scholar] [CrossRef]
- Nunes, J. M.; Kell, D. B.; Pretorius, E. Cardiovascular and haematological pathology in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): a role for Viruses. Blood Rev. 2023, 60, 101075. [Google Scholar] [CrossRef]
- Pretorius, E.; Vermeulen, N.; Bester, J.; Lipinski, B. Novel use of scanning electron microscopy for detection of iron-induced morphological changes in human blood. Microsc. Res. Tech. 2013, 76, 268–271. [Google Scholar] [CrossRef]
- Pretorius, E.; Bester, J.; Vermeulen, N.; Lipinski, B.; Gericke, G. S.; Kell, D. B. Profound morphological changes in the erythrocytes and fibrin networks of patients with hemochromatosis or with hyperferritinemia, and their normalization by iron chelators and other agents. PLoS ONE 2014, 9, e85271. [Google Scholar] [CrossRef]
- Pretorius, E.; Kell, D. B. Diagnostic morphology: biophysical indicators for iron-driven inflammatory diseases. Integr. Biol. 2014, 6, 486–510. [Google Scholar] [CrossRef] [PubMed]
- Grobbelaar, L. M.; Venter, C.; Vlok, M.; Ngoepe, M.; Laubscher, G. J.; Lourens, P. J.; Steenkamp, J.; Kell, D. B.; Pretorius, E. SARS-CoV-2 spike protein S1 induces fibrin(ogen) resistant to fibrinolysis: implications for microclot formation in COVID-19. Biosci. Rep. 2021, 41, BSR20210611. [Google Scholar] [CrossRef] [PubMed]
- Nyström, S.; Hammarström, P. Amyloidogenesis of SARS-CoV-2 Spike Protein. J. Amer Chem. Soc. 2022, 144, 8945–8950. [Google Scholar] [CrossRef]
- Westman, H.; Hammarström, P.; Nyström, S. SARS-CoV-2 Spike Protein Amyloid Fibrils Impair Fibrin Formation and Fibrinolysis. Biochemistry 2025, 64, 4818–4829. [Google Scholar] [CrossRef]
- Grobbelaar, L. M.; Kruger, A.; Venter, C.; Burger, E. M.; Laubscher, G. J.; Maponga, T. G.; Kotze, M. J.; Kwaan, H. C.; Miller, J. B.; Fulkerson, D.; Huff, W.; Chang, E.; Wiarda, G.; Bunch, C. M.; Walsh, M. M.; Raza, S.; Zamlut, M.; Moore, H. B.; Moore, E. E.; Neal, M. D.; Kell, D. B.; Pretorius, E. Relative hypercoagulopathy of the SARS-CoV-2 Beta and Delta variants when compared to the less severe Omicron variants is related to TEG parameters, the extent of fibrin amyloid microclots, and the severity of clinical illness. Semin Thromb. Haemost. 2022, 48, 858–868. [Google Scholar] [CrossRef]
- Biancalana, M.; Makabe, K.; Koide, A.; Koide, S. Molecular mechanism of thioflavin-T binding to the surface of beta-rich peptide self-assemblies. J. Mol. Biol. 2009, 385, 1052–1063. [Google Scholar] [CrossRef]
- Pretorius, E.; Page, M. J.; Hendricks, L.; Nkosi, N. B.; Benson, S. R.; Kell, D. B. Both lipopolysaccharide and lipoteichoic acids potently induce anomalous fibrin amyloid formation: assessment with novel Amytracker™ stains. bioRxiv Prepr. bioRxiv 2017, 143867. [Google Scholar] [CrossRef]
- Elghetany, M. T.; Saleem, A.; Barr, K. The congo red stain revisited. Ann. Clin. Lab Sci. 1989, 19, 190–195. [Google Scholar]
- Howie, A. J. Green (or apple-green) birefringence" of Congo red-stained amyloid. Amyloid 2015, 22, 205–206. [Google Scholar] [CrossRef] [PubMed]
- Yakupova, E. I.; Bobyleva, L. G.; Vikhlyantsev, I. M.; Bobylev, A. G. Congo Red and amyloids: history and relationship. Biosci. Rep. 2019. [Google Scholar] [CrossRef]
- Howie, A. J. Anomalous colours, not interference colours or 'apple-green birefringence', in Congo red-stained amyloid. Amyloid 2024, 31, 356–357. [Google Scholar] [CrossRef]
- Grixti, J. M.; Theron, C. W.; Salcedo-Sora, J. E.; Pretorius, E.; Kell, D. B. Automated microscopic measurement of fibrinaloid microclots and their degradation by nattokinase, the main natto protease. J. Exp. Clin. Appl. Chin. Med. 2024, 5, 30–55. [Google Scholar] [CrossRef]
- Dapson, R. W. Amyloid from a histochemical perspective. A review of the structure, properties and types of amyloid, and a proposed staining mechanism for Congo red staining. Biotech. Histochem. 2018, 93, 543–556. [Google Scholar] [CrossRef]
- Kell, D. B.; Lip, G. Y. H.; Pretorius, E. Fibrinaloid Microclots and Atrial Fibrillation. Biomedicines 2024, 12, 891. [Google Scholar] [CrossRef]
- Kell, D. B.; Pretorius, E. Some potential roles of fibrin amyloid ('fibrinaloid') microclots in fibromyalgia syndrome. Int. J. Adv. Med. Clin. Ther. 2026. [Google Scholar] [CrossRef]
- Kell, D. B.; Khan, M. A.; Kane, B.; Lip, G. Y. H.; Pretorius, E. Possible role of fibrinaloid microclots in Postural Orthostatic Tachycardia Syndrome (POTS): focus on Long COVID. J. Pers. Med. 2024, 14, 170. [Google Scholar] [CrossRef]
- Kell, D. B.; Pretorius, E. Potential roles of fibrinaloid microclot complexes in inhibiting the cochlear microcirculation during the development of tinnitus. Preprints 2025, 2025081557. [Google Scholar] [CrossRef]
- Burdukiewicz, M.; Sobczyk, P.; Rödiger, S.; Duda-Madej, A.; Mackiewicz, P.; Kotulska, M. Amyloidogenic motifs revealed by n-gram analysis. Sci. Rep. 2017, 7, 12961. [Google Scholar] [CrossRef]
- Szulc, N.; Burdukiewicz, M.; Gąsior-Głogowska, M.; Wojciechowski, J. W.; Chilimoniuk, J.; Mackiewicz, P.; Šneideris, T.; Smirnovas, V.; Kotulska, M. Bioinformatics methods for identification of amyloidogenic peptides show robustness to misannotated training data. Sci. Rep. 2021, 11, 8934. [Google Scholar] [CrossRef] [PubMed]
- Kell, D. B.; Doyle, K. M.; Salcedo-Sora, J. E.; Sekhar, A.; Walker, M.; Pretorius, E. AmyloGram reveals amyloidogenic potential in stroke thrombus proteomes. Biochem J. 2025, 482, 1689–1706. [Google Scholar] [CrossRef]
- Ząbczyk, M.; Stachowicz, A.; Natorska, J.; Olszanecki, R.; Wiśniewski, J. R.; Undas, A. Plasma fibrin clot proteomics in healthy subjects: relation to clot permeability and lysis time. J. Proteom. 2019, 208, 103487. [Google Scholar] [CrossRef] [PubMed]
- Kell, D. B.; Pretorius, E. Proteomic evidence for amyloidogenic cross-seeding in fibrinaloid microclots. Int. J. Mol. Sci. 2024, 25, 10809. [Google Scholar] [CrossRef] [PubMed]
- Kell, D. B.; Pretorius, E. The proteome content of blood clots observed under different conditions: successful role in predicting clot amyloid(ogenicity). Molecules 2025, 30, 668. [Google Scholar] [CrossRef]
- Kell, D. B.; Doyle, K. M.; Salcedo-Sora, E.; Sekhar, A.; Walker, M.; Pretorius, E. AmyloGram reveals amyloidogenic potential in stroke thrombus proteomes. Biochem J. 2025, 482, BCJ20253317. [Google Scholar] [CrossRef]
- Wright, C.; Kell, D. B.; Pretorius, E.; Putrino, D. Treatment of Long Covid with enoxaparin. Cardiopulm. Phys. Ther. J. 2025, 36, 70–73. [Google Scholar] [CrossRef]
Figure 1.
Fibrinaloid microclot complexes stained with Thioflavin T (ThT) as detected in platelet poor plasma (PPP) using imaging flow cytometry. Image taken from the CC-BY 4.0 publication [15].
Figure 1.
Fibrinaloid microclot complexes stained with Thioflavin T (ThT) as detected in platelet poor plasma (PPP) using imaging flow cytometry. Image taken from the CC-BY 4.0 publication [15].

Figure 2.
A fibrinaloid microclot created using purified fibrinogen and thrombin as in Methods, stained with 0.3 mM Congo Red, and imagied in a polarising microcope as in Methods. The classical ‘apple-green’ birefringence is observed in particular around the periphery of the microclot.
Figure 2.
A fibrinaloid microclot created using purified fibrinogen and thrombin as in Methods, stained with 0.3 mM Congo Red, and imagied in a polarising microcope as in Methods. The classical ‘apple-green’ birefringence is observed in particular around the periphery of the microclot.

Figure 3.
A summary of fibrinaloid microclot complexes. This figure (and also the graphical abstract) was generated on 23/4/2026 with the assistance of the artificial intelligence tool NotebookLM (https://notebooklm.google/), using author-provided text and figure concepts; all scientific content, interpretation, and final design decisions were verified and approved by the authors.
Figure 3.
A summary of fibrinaloid microclot complexes. This figure (and also the graphical abstract) was generated on 23/4/2026 with the assistance of the artificial intelligence tool NotebookLM (https://notebooklm.google/), using author-provided text and figure concepts; all scientific content, interpretation, and final design decisions were verified and approved by the authors.


Table 1.
(above). Summary of representative evidence for fibrinaloid microclot complexes across selected disease settings. The table distinguishes disease context, study provenance, material analysed, analytical modality, and the type of supporting evidence. It is intended as an evidence map rather than an exhaustive systematic review.
Table 1.
(above). Summary of representative evidence for fibrinaloid microclot complexes across selected disease settings. The table distinguishes disease context, study provenance, material analysed, analytical modality, and the type of supporting evidence. It is intended as an evidence map rather than an exhaustive systematic review.
| Condition | Representative independent studies | Representative Kell/Pretorius studies | Sample/material | Readout | Amyloid-selective stain | Proteomics | NET association | Clinical link |
|---|---|---|---|---|---|---|---|---|
| Acute COVID-19 | [20] | [21,22] | PPP/plasma | microscopy / staining | yes | limited | variable | coagulopathy |
| Long COVID | [23,24,25,26] | [15,27,28,29,30,31,32,33,34,35,36,37,38] | PPP/plasma | IFC / microscopy / proteomics | yes | yes | yes | symptom burden |
| Sepsis | [39] | Presaged in [40] | plasma | amyloid-fibrinogen aggregates | yes | no | not central | DIC / mortality |
| Stroke thrombi | — | [41,42] | retrieved thrombi | amyloid staining / proteomics | yes | yes | not central | fibrinolysis resistance |
| T2D | — | [12,13,22,43,44,45] | PPP/fibrin | microscopy / stains | yes | limited | no | abnormal clotting |
| Parkinson’s / Alzheimer’s | — | [2,12,46,47,48,49,50] | plasma/fibrin | correlative LM/EM, stains | yes | limited | no | chronic inflammation |
| Rheumatoid arthritis | — | [51] | plasma/fibrin | EM | no | no | no | chronic inflammation |
| ME/CFS | [28,36,52,53] | Plasma/fibrin | Microscopy /stains | Yes | Yes | No | chronic inflammation |
Table 2.
Proteins enriched or lowered in fibrinaloid microclots relative to normal clots [77], with their Amylogram scores.
Table 2.
Proteins enriched or lowered in fibrinaloid microclots relative to normal clots [77], with their Amylogram scores.
| Protein | Uniprot ID | Amylogram score |
|---|---|---|
| Adiponectin | Q15848 | 0.833 |
| Kallikrein | P03952 | 0.769 |
| LBLC1/BNIB1/BNIFB1/LPLUNC1 | Q8TDL5 | 0.918 |
| Platelet factor 4 | P02776 | 0.778 |
| Periostin | Q15063 | 0.914 |
| Thrombospondin-1 | P07996 | 0.863 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



