Submitted:
21 April 2026
Posted:
23 April 2026
You are already at the latest version
Abstract
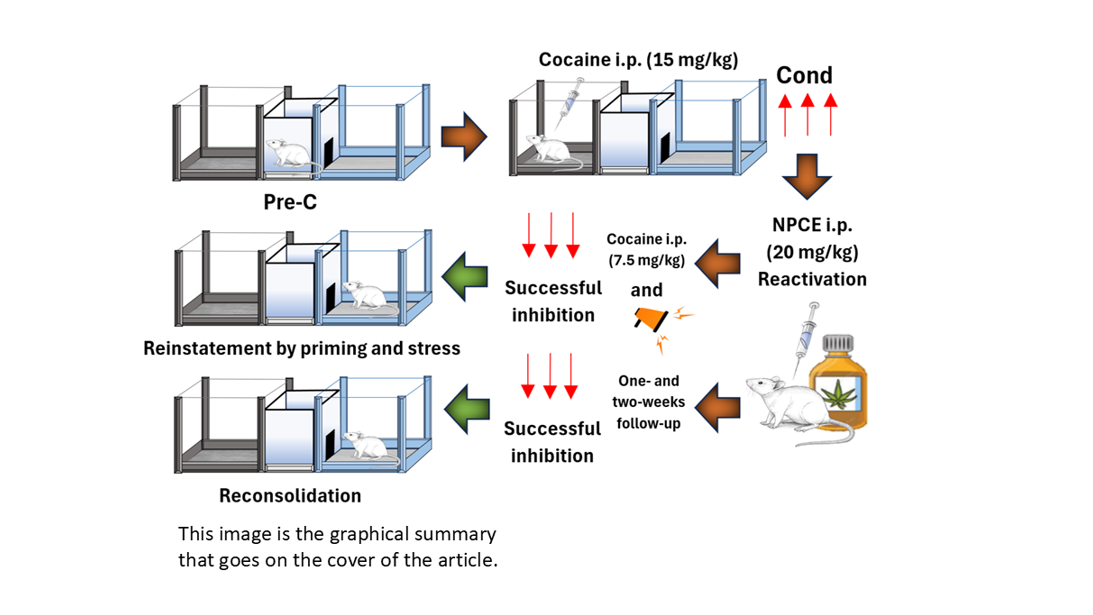
Cocaine use disorder (CUD) remains a significant global health issue, with no FDA-approved pharmacological treatments. Cannabidiol (CBD), a non-psychoactive phytocannabinoid primarily derived from Cannabis sativa L., has demonstrated promising results in preclinical research to disrupt the consolidation and retrieval of drug-associated memories, thereby reducing relapse behaviors linked to substance use disorders such as cocaine dependence. This study evaluates the effects of a non-psychoactive cannabis extract (NPCE) on the reinstatement and reconsolidation of cocaine-induced conditioned place preference (CPP) in CD1 male mice—processes that, to our knowledge, have not been previously examined. The results showed that NPCE significantly inhibited both priming -induced and stress-induced reinstatement of cocaine-induced CPP, suggesting its potential to disrupt drug-associated memories. Additionally, NPCE effectively impaired the reconsolidation of cocaine-induced CPP, suggesting an effect on memory reconfiguration lasting at least two weeks. Additionally, NPCE alone did not produce any effect on CPP acquisition. These findings underscore the potential of NPCE, in targeting memory-related mechanisms underlying cocaine addiction, specifically in the reconsolidation and reinstatement. These results indicated that NPCE may reduce relapse risk by modulating drug-reward memories, potentially through interactions with CB1 receptors and other molecular signaling pathways like serotonergic receptors. This research contributes to the growing body of evidence, which suggests that cannabinoids, particularly non-psychoactive extracts, could offer novel therapeutic options for treating CUD. Further studies are needed to explore the individual effects of other cannabinoids on cocaine dependence and to assess clinical applicability of these findings.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Animals
2.1. Chemical and NPCE
2.1. Doses
2.1. Experiments
- Pre-conditioning (Pre-C phase)
- Conditioning phase (Cond phase)
2.1.1. Experiment 1: Evaluation of NPCE in acquisition of CPP
2.1.1. Experiment 2: Evaluation of NPCE in the Reinstatement of CPP Induced by Cocaine
2.1.1. Experiment 3: Evaluation of NPCE in the Reconsolidation of CPP Induced by Cocaine
2.1. Software and Statistical Analysis
2.1. Declaration of generative AI and AI-assisted technologies in the writing process
3. Results
4. Experiment 1: Results of evaluation of NPCE in the acquisition of CPP.
5. Experiment 2: Evaluation of NPCE in the Reinstatement of CPP Induced by Cocaine
6. Experiment 3: Evaluation of NPCE in the reconsolidation of CPP induced by cocaine
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roque Bravo, R.; Faria, A.C.; Brito-Da-Costa, A.M.; Carmo, H.; Mladěnka, P.; Dias da Silva, D.; Remião, F. Cocaine: An updated overview on chemistry, detection, biokinetics, and pharmacotoxicological aspects including abuse pattern. Toxins 2022, 14, 278. [Google Scholar] [CrossRef]
- United Nations Office on Drugs and Crime. World Drug Report 2024: Key Findings and Conclusions; UNODC: Vienna, Austria, 2024. [Google Scholar]
- Viudez-Martínez, A.; García-Gutiérrez, M.S.; Medrano-Relinque, J.; Navarrón, C.M.; Navarrete, F.; Manzanares, J. Cannabidiol does not display drug abuse potential in mice behavior. Acta Pharmacol. Sin. 2019, 40, 358–364. [Google Scholar] [CrossRef]
- Paulus, V.; Billieux, J.; Benyamina, A.; Karila, L. Cannabidiol in the context of substance use disorder treatment: A systematic review. Addict. Behav. 2022, 132, 107360. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Caroba, M.E.S.; Taba, F.K.; Filev, R.; Gallassi, A.D. Evaluation of the potential use of cannabidiol in the treatment of cocaine use disorder: A systematic review. Pharmacol. Biochem. Behav. 2020, 196, 172982. [Google Scholar] [CrossRef]
- Briânis, R.C.; Iglesias, L.P.; Bedeschi, L.G.; Moreira, F.A. Effects of cannabidiol on reward contextual memories induced by cocaine in male and female mice. Acta Neuropsychiatr 2023. [Google Scholar] [CrossRef]
- Calpe-López, C.; Gasparyan, A.; Navarrete, F.; Manzanares, J.; Miñarro, J.; Aguilar, M.A. Cannabidiol prevents priming- and stress-induced reinstatement of the conditioned place preference induced by cocaine in mice. J. Psychopharmacol. 2021, 35, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Chesworth, R.; Karl, T. Cannabidiol (CBD) reduces cocaine-environment memory in mice. Pharmacol. Biochem. Behav. 2020, 199, 173065. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.R.; Takahashi, R.N. Cannabidiol disrupts the reconsolidation of contextual drug-associated memories in Wistar rats. Addict. Biol. 2017, 22, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Ledesma, J.C.; Manzanedo, C.; Aguilar, M.A. Cannabidiol prevents several behavioral alterations related to cocaine addiction in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 111, 110390. [Google Scholar] [CrossRef]
- Tzschentke, T.M. Measuring reward with the conditioned place preference (CPP) paradigm: Update of the last decade. Addict. Biol. 2007, 12, 227–462. [Google Scholar] [CrossRef]
- McKendrick, G.; Graziane, N.M. Drug-induced conditioned place preference and its practical use in substance use disorder research. Front. Behav. Neurosci. 2020, 14, 582147. [Google Scholar] [CrossRef]
- Mahmud, A.; Gallant, S.; Sedki, F.; D’Cunha, T.; Shalev, U. Effects of an acute cannabidiol treatment on cocaine self-administration and cue-induced cocaine seeking in male rats. J. Psychopharmacol. 2017, 31, 96–104. [Google Scholar] [CrossRef]
- Nedelescu, H.; Wagner, G.E.; De Ness, G.L.; Carroll, A.; Kerr, T.M.; Wang, J.; Zhang, S.; Chang, S.; Than, A.H.; Emerson, N.E.; et al. Cannabidiol produces distinct U-shaped dose-response effects on cocaine-induced conditioned place preference and recruitment of prelimbic neurons in male rats. Biol. Psychiatry Glob. Open Sci. 2022, 2, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Chesworth, R.; Yim, H.C.H.; Watt, G.; El-Omar, E.; Karl, T. Cannabidiol facilitates cocaine extinction and ameliorates cocaine-induced changes to the gut microbiome in male mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2024, 133, 111014. [Google Scholar] [CrossRef]
- Alegre-Zurano, L.; Berbegal-Sáez, P.; Luján, M.; Cantacorps, L.; Martín-Sánchez, A.; García-Baos, A.; Valverde, O. Cannabidiol decreases motivation for cocaine in a behavioral economics paradigm but does not prevent incubation of craving in mice. Biomed. Pharmacother. 2022, 148, 112708. [Google Scholar] [CrossRef] [PubMed]
- de Meneses-Gaya, C.; Crippa, J.A.; Hallak, J.E.; Miguel, A.Q.; Laranjeira, R.; Bressan, R.A.; Zuardi, A.W.; Lacerda, A.L. Cannabidiol for the treatment of crack-cocaine craving: An exploratory double-blind study. Braz. J. Psychiatry 2021, 43, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Mongeau-Pérusse, V.; Brissette, S.; Bruneau, J.; Conrod, P.; Dubreucq, S.; Gazil, G.; Stip, E.; Jutras-Aswad, D. Cannabidiol as a treatment for craving and relapse in individuals with cocaine use disorder: A randomized placebo-controlled trial. Addiction 2021, 116, 2431–2442. [Google Scholar] [CrossRef] [PubMed]
- Rizkallah, E.; Mongeau-Pérusse, V.; Lamanuzzi, L.; Castenada-Ouellet, S.A.; Stip, E.; Juteau, L.C.; Brissette, S.; Bruneau, J.; Dubreucq, S.; Jutras-Aswad, D. Cannabidiol effects on cognition in individuals with cocaine use disorder: Exploratory results from a randomized controlled trial. Pharmacol. Biochem. Behav. 2022, 216, 173376. [Google Scholar] [CrossRef]
- Murray, C.H.; Haney, M.; Foltin, R.W.; Manubay, J.; Bedi, G.; Cooper, Z.D. Smoked cannabis reduces peak cocaine plasma levels and subjective effects in a controlled drug administration study. Drug Alcohol Depend. 2023, 243, 109757. [Google Scholar] [CrossRef]
- Socías, M.E.; Kerr, T.; Wood, E.; Dong, H.; Lake, S.; Hayashi, K.; DeBeck, K.; Jutras-Aswad, D.; Montaner, J.; Milloy, M.J. Intentional cannabis use to reduce crack cocaine use in a Canadian setting: A longitudinal analysis. Addict. Behav. 2017, 72, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Labigalini, E.; Rodrigues, L.R.; Da Silveira, D.X. Therapeutic use of cannabis by crack addicts in Brazil. J. Psychoact. Drugs 1999, 31, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Wang, X.; Markowitz, J.S. In vitro inhibition of carboxylesterase 1 by major cannabinoids and selected metabolites. Drug Metab. Dispos. 2019, 47, 465–472. [Google Scholar] [CrossRef]
- Russo, E.B. Cannabis therapeutics and the future of neurology. Front. Integr. Neurosci. 2018, 12, 51. [Google Scholar] [CrossRef]
- Johnson, A.L.; Verbitsky, R.; Hudson, J.; Dean, R.; Hamilton, T.J. Cannabinoid type-2 receptors modulate terpene-induced anxiety reduction in zebrafish. Biomed. Pharmacother. 2023, 168, 115760. [Google Scholar] [CrossRef] [PubMed]
- Blasco-Benito, S.; Seijo-Vila, M.; Caro-Villalobos, M.; Tundidor, I.; Andradas, C.; García-Taboada, E.; Wade, J.; Smith, S.; Guzmán, M.; Pérez-Gómez, E.; et al. Appraising the “entourage effect”: Antitumor action of a pure cannabinoid versus a botanical drug preparation in breast cancer models. Biochem. Pharmacol. 2018, 157, 285–293. [Google Scholar] [CrossRef]
- O’Connell, B.K.; Gloss, D.; Devinsky, O. Cannabinoids in treatment-resistant epilepsy: A review. Epilepsy Behav. 2017, 70, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Parker, L.A.; Burton, P.; Sorge, R.E.; Yakiwchuk, C.; Mechoulam, R. Effect of low doses of Δ9-tetrahydrocannabinol and cannabidiol on extinction of cocaine- and amphetamine-induced conditioned place preference. Psychopharmacology 2004, 175, 360–366. [Google Scholar] [CrossRef]
- Barreto, F.L.; Lozano, M.C.; Martínez-Ramírez, J.A. Non-psychoactive cannabis extract promotes extinction and reduces reinstatement by priming dose in smoked cocaine-induced conditioned place preference. Physiol. Behav. 2025, 301, 115048. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef]
- Farrell, M.R.; Schoch, H.; Mahler, S.V. Modeling cocaine relapse in rodents: Behavioral considerations and circuit mechanisms. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 87, 33–47. [Google Scholar] [CrossRef]
- Galaj, E.; Bi, G.H.; Yang, H.J.; Xi, Z.X. Cannabidiol attenuates the rewarding effects of cocaine in rats via CB2, 5-HT1A, and TRPV1 receptor mechanisms. Neuropharmacology 2020, 167, 107740. [Google Scholar] [CrossRef]
- Rodríguez-Arias, M.; Roger-Sánchez, C.; Vilanova, I.; Revert, N.; Manzanedo, C.; Miñarro, J.; Aguilar, M.A. Effects of cannabinoid exposure during adolescence on rewarding effects of WIN 55212-2 and cocaine in mice. Neural Plast. 2016, 2016, 6481862. [Google Scholar] [CrossRef]
- Rosa, M.L.P.; Machado, C.A.; Asth, L.; Toscano, E.C.B.; da Silva Oliveira, B.; Marzano, L.A.S.; Ferreira, R.N.; Teixeira, A.L.; Moreira, F.A.; Miranda, A.S. A three-compartment apparatus alters brain cytokines and neurotrophic factors in cocaine-induced CPP. J. Neuroimmunol. 2022, 369, 577914. [Google Scholar] [CrossRef]
- Reisdorph, N.; Doenges, K.; Levens, C.; Manke, J.; Armstrong, M.; Smith, H.; Quinn, K.; Radcliffe, R.; Reisdorph, R.; Saba, L.; et al. Oral cannabis consumption and intraperitoneal THC:CBD dosing results in changes in brain and plasma neurochemicals and endocannabinoids in mice. J. Cannabis Res. 2024, 6. [Google Scholar] [CrossRef]
- Fadda, P.; Robinson, L.; Fratta, W.; Pertwee, R.G.; Riedel, G. Differential effects of THC- or CBD-rich cannabis extracts on working memory in rats. Neuropharmacology 2004, 47, 1170–1179. [Google Scholar] [CrossRef] [PubMed]
- Parker, L.A.; Gillies, T. THC-induced place and taste aversions in Lewis and Sprague–Dawley rats. Behav. Neurosci. 1995, 109, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Luján, M.; Alegre-Zurano, L.; Martín-Sánchez, A.; Cantacorps, L.; Valverde, O. CB1 receptor antagonist AM4113 reverts the effects of cannabidiol on cue- and stress-induced reinstatement of cocaine-seeking behaviour in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 113. [Google Scholar] [CrossRef]
- Bayer, H.; Stern, C.A.J.; Troyner, F.; Gazarini, L.; Guimarães, F.S.; Bertoglio, L.J. Medial prefrontal cortex mechanisms of cannabidiol-induced aversive memory reconsolidation impairments. Neuropharmacology 2022, 205. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.A.; Marshall, J.F. Molecular substrates for retrieval and reconsolidation of cocaine-associated contextual memory. Neuron 2005, 47, 873–884. [Google Scholar] [CrossRef]
- Fang, Q.; Li, F.Q.; Li, Y.Q.; Xue, Y.X.; He, Y.Y.; Liu, J.F.; Lu, L.; Wang, J.S. Cannabinoid CB1 receptor antagonist rimonabant disrupts nicotine reward-associated memory in rats. Pharmacol. Biochem. Behav. 2011, 99, 738–742. [Google Scholar] [CrossRef]
- Khodamoradi, M.; Tirgar, F.; Ghazvini, H.; Rafaiee, R.; Tamijani, S.M.S.; Karimi, N.; Yadegari, A.; Khachaki, A.S.; Akhtari, J. Role of the cannabinoid CB1 receptor in methamphetamine-induced social and recognition memory impairment. Neurosci. Lett. 2022, 779. [Google Scholar] [CrossRef]
- Luján, M.Á.; Castro-Zavala, A.; Alegre-Zurano, L.; Valverde, O. Repeated cannabidiol treatment reduces cocaine intake and modulates neural proliferation and CB1R expression in the mouse hippocampus. Neuropharmacology 2018, 143, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.L.; Zhou, S.J.; Wang, X.Y.; Liu, J.F.; Xue, Y.X.; Jiang, W.; Lu, L. Effects of cannabinoid CB1 receptor antagonist rimonabant on acquisition and reinstatement of psychostimulant reward memory in mice. Behav. Brain Res. 2011, 217, 111–116. [Google Scholar] [CrossRef]
- Guerrero-Bautista, R.; Do Couto, B.R.; Hidalgo, J.M.; Cárceles-Moreno, F.J.; Molina, G.; Laorden, M.L.; Núñez, C.; Milanés, M.V. Modulation of stress- and cocaine prime-induced reinstatement of conditioned place preference after memory extinction through dopamine D3 receptor. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 92, 308–320. [Google Scholar] [CrossRef]
- Manzoni, O.J.; Manduca, A.; Trezza, V. Therapeutic potential of cannabidiol polypharmacology in neuropsychiatric disorders. Trends Pharmacol. Sci. 2025. [Google Scholar] [CrossRef] [PubMed]
- Tronson, N.C.; Taylor, J.R. Addiction: A drug-induced disorder of memory reconsolidation. Curr. Opin. Neurobiol. 2013, 23, 573–580. [Google Scholar] [CrossRef]
- Yu, L.L.; Wang, X.Y.; Zhao, M.; Liu, Y.; Li, Y.Q.; Li, F.Q.; Wang, X.; Xue, Y.X.; Lu, L. Effects of cannabinoid CB1 receptor antagonist rimonabant in consolidation and reconsolidation of methamphetamine reward memory in mice. Psychopharmacology 2009, 204, 203–211. [Google Scholar] [CrossRef]
- Ramiro-Fuentes, S.; Ortiz, O.; Moratalla, R.; Fernandez-Espejo, E. Intra-accumbens rimonabant is rewarding but induces aversion to cocaine in cocaine-treated rats, as does in vivo accumbal cannabinoid CB1 receptor silencing: Critical role for glutamate receptors. Neuroscience 2010, 167, 205–215. [Google Scholar] [CrossRef]
- Ward, S.J.; Rosenberg, M.; Dykstra, L.A.; Walker, E.A. The CB1 antagonist rimonabant (SR141716) blocks cue-induced reinstatement of cocaine seeking and other context and extinction phenomena predictive of relapse. Drug Alcohol Depend. 2009, 105, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Schulkin, J. Addiction and stress: An allostatic view. Neurosci. Biobehav. Rev. 2019, 106, 245–262. [Google Scholar] [CrossRef]
- Ruisoto, P.; Contador, I. The role of stress in drug addiction: An integrative review. Physiol. Behav. 2019, 202, 62–68. [Google Scholar] [CrossRef]
- Shaham, Y.; Erb, S.; Stewart, J. Stress-induced relapse to heroin and cocaine seeking in rats: A review. Brain Res. Rev. 2000, 33, 13–33. [Google Scholar] [CrossRef]
- Chu, J.; Deyama, S.; Li, X.; Motono, M.; Otoda, A.; Saito, A.; Esaki, H.; Nishitani, N.; Kaneda, K. Role of 5-HT1A receptor-mediated serotonergic transmission in the medial prefrontal cortex in acute restraint stress-induced augmentation of rewarding memory of cocaine in mice. Neurosci. Lett. 2021, 743. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.; He, Y.; Suo, J.; Wu, C.; Gong, X.; Xiang, Y.; Yang, W.; Cheng, J.; Wang, Y.; Chen, W.; et al. Cannabidiol exhibits anxiolytic-like effects and antipsychotic-like effects in mice models. Neurosci. Lett. 2024, 826. [Google Scholar] [CrossRef]
- De Gregorio, D.; McLaughlin, R.J.; Posa, L.; Ochoa-Sanchez, R.; Enns, J.; Lopez-Canul, M.; Aboud, M.; Maione, S.; Comai, S.; Gobbi, G. Cannabidiol modulates serotonergic transmission and reverses both allodynia and anxiety-like behavior in a model of neuropathic pain. Pain 2019, 160, 136–150. [Google Scholar] [CrossRef]
- Zhu, J.; Hou, Y.; Li, W.; Wang, X.; Li, F.; Li, N.; Hu, Y.; Wang, X.; Ge, S.N. miR-181a expressed in the dorsal hippocampus regulates the reinstatement of cocaine CPP by targeting PRKAA1. Behav. Brain Res. 2024, 471. [Google Scholar] [CrossRef]
- Koob, G.; Kreek, M.J. Stress, dysregulation of drug reward pathways, and the transition to drug dependence. Am. J. Psychiatry 2007, 164, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Koltai, H.; Namdar, D. Cannabis phytomolecule “entourage”: From domestication to medical use. Trends Plant Sci. 2020, 25, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Yamaori, S.; Takeda, S.; Yamamoto, I.; Watanabe, K. Identification of cytochrome P450 enzymes responsible for metabolism of cannabidiol by human liver microsomes. Life Sci. 2011, 89, 165–170. [Google Scholar] [CrossRef]
- Jastrząb, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. The origin and biomedical relevance of cannabigerol. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Zagzoog, A.; Mohamed, K.A.; Kim, H.J.J.; Kim, E.D.; Frank, C.S.; Black, T.; Jadhav, P.D.; Holbrook, L.A.; Laprairie, R.B. In vitro and in vivo pharmacological activity of minor cannabinoids isolated from Cannabis sativa. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Hayduk, S.A.; Hughes, A.C.; Winter, R.L.; Milton, M.D.; Ward, S.J. Single and combined effects of cannabigerol (CBG) and cannabidiol (CBD) in mouse models of oxaliplatin-associated mechanical sensitivity, opioid antinociception, and naloxone-precipitated opioid withdrawal. Biomedicines 2024, 12. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual authors and contributors and not of MDPI or the editors. MDPI and the editors disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |





| CPP study | Experimental group (n) | Reinforcing substance | Treatment |
| Experiment 1. Cond NPCE | CNPCE (n = 10) |
NPCE 20 mg/kg | - |
| CVEH (n = 10) |
VEH (0.01 mL/g) | - | |
| Experiment 2. Reinstatement (priming dose and stress) | ECOC (NPCE+COC) (n = 10) | Cocaine 15 mg/kg | NPCE 20 mg/kg |
| CECOC (VEH+COC) (n = 10) | VEH (0.01 mL/g) | ||
| Experiment 3. Reconsolidation | RCOC (NPCE+COC) (n = 10) | NPCE 20 mg/kg | |
| CRCOC (VEH+COC) (n = 10) | VEH (0.01 mL/g) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



