Submitted:
21 March 2026
Posted:
24 March 2026
You are already at the latest version
Abstract
Background: Personalized mRNA neoantigen vaccines represent a transformative approach to cancer immunotherapy, with recent Phase IIb data demonstrating a 49% reduction in melanoma recurrence when combined with pembrolizumab. However, two fundamental limitations constrain their efficacy: (1) fewer than 2% of mRNA molecules delivered via lipid nanoparticles (LNPs) escape the endosomal compartment to reach the cytoplasm, and (2) immunosuppressive tumor microenvironments (TMEs) in “cold” tumors blunt vaccine-induced T-cell responses. Current approaches address these challenges separately.Hypothesis: We propose that hydroxychloroquine (HCQ), when structurally incorporated as the ionizable lipid component of the LNP delivery system rather than administered as a separate systemic drug, creates a dual-function nanoparticle that simultaneously: (a) enhances mRNA endosomal escape by modulating endosomal pH maturation kinetics; (b) repolarizes tumor-associated macrophages from M2 to M1 phenotype via lysosomal Ca2+/mucolipin-1/p38/NF-κB signaling; (c) inhibits tumor-protective autophagy; and (d) reduces LNP-induced inflammatory side effects, enabling repeated dosing required for neoantigen vaccine protocols.Supporting evidence: Two independent 2025 studies demonstrated that HCQ-derived ionizable lipids achieve spleen-tropic mRNA delivery with simultaneous M2→M1 macrophage repolarization and anti-inflammatory properties permitting repeated dosing. We integrate these findings with our AI-driven Tumor Vaccine Responsiveness Score (TVRS) framework, which analyzed 8,432 TCGA Pan-Cancer Atlas specimens across 20 cancer types and identified that tumors most resistant to standard mRNA vaccines (pancreatic TVRS=11.7, glioblastoma TVRS=15.3) are precisely those where autophagy-dependent immune evasion is most active — making them ideal candidates for HCQ-LNP.
Keywords:
hydroxychloroquine
; lipid nanoparticles
; mRNA vaccine
; neoantigen
; autophagy
; tumor microenvironment
; personalized medicine
; endosomal escape
; cancer immunotherapy
; artificial intelligence

Figure (I). graphical abstract.
1. Introduction
The convergence of three technological revolutions—mRNA therapeutics, AI-driven neoantigen prediction, and protein structure modeling—has brought personalized cancer vaccines from theoretical concept to clinical reality. The KEYNOTE-942 Phase IIb trial demonstrated that intismeran autogene (mRNA-4157), a personalized neoantigen vaccine encoding up to 34 tumor-specific mutations, reduced melanoma recurrence by 49% at five years when combined with pembrolizumab [1]. Parallel developments in pancreatic cancer (BNT122, 100% recurrence-free survival in responders at 18 months) [3] and triple-negative breast cancer [2] have validated the premise that patient-specific mRNA vaccines can mobilize clinically meaningful anti-tumor immunity.
Yet a paradox haunts this field: the cancers that would benefit most from personalized vaccination are precisely those where current mRNA-LNP platforms work least effectively. Our computational analysis of 8,432 TCGA Pan-Cancer Atlas specimens using the Tumor Vaccine Responsiveness Score (TVRS) framework revealed a striking inverse correlation between clinical need and predicted vaccine responsiveness [9,10]. Melanoma scored TVRS 77.6, while pancreatic adenocarcinoma (TVRS 11.7) and glioblastoma (TVRS 15.3) scored lowest—not because they lack neoantigens entirely, but because their immunosuppressive, autophagy-dependent, “cold” tumor microenvironments prevent vaccine-induced T cells from functioning [9,14].
The delivery vehicle itself compounds this problem. Current LNPs face a well-characterized endosomal escape bottleneck: fewer than 2% of mRNA molecules reach the cytoplasm for translation [8]. In this hypothesis paper, we propose a unified solution by fundamentally reconceiving the role of hydroxychloroquine (HCQ) in cancer vaccine design. Rather than administering HCQ as a separate systemic drug, we propose incorporating HCQ-derived molecules as the ionizable lipid component of the LNP itself, creating a single nanoparticle that simultaneously optimizes mRNA delivery and reprograms the tumor immune environment (Figure 1).
2. The Three Convergent Pillars
2.1. Pillar I: the Endosomal Escape Crisis in mRNA Vaccine Delivery
The clinical efficacy of any mRNA vaccine is fundamentally constrained by the fraction of delivered mRNA that reaches the cytoplasm. The journey requires LNP internalization via endocytosis, transport through early endosomes (pH ~6.5) to late endosomes (pH ~5.5), and escape before lysosomal degradation (pH ~4.5) [8]. Quantitative studies estimate that fewer than 2% of mRNA molecules reach the cytoplasm [8]. The window between productive endosomal escape and destructive lysosomal degradation is narrow, and current ionizable lipids traverse it too slowly.
HCQ, as a lysosomotropic weak base, accumulates in acidic compartments and raises their pH, slowing the transition from late endosomes to lysosomes. In the context of an HCQ-derived ionizable lipid, this pH-modulating activity is not a systemic effect requiring oral dosing of 400–800 mg/day, but a local, nanoparticle-intrinsic property operating at the subcellular level within the compartments where mRNA escape occurs (Figure 2).
2.2. Pillar II: Tumor Microenvironment Reprogramming via M2→M1 Macrophage Repolarization
The failure of cancer vaccines in “cold” tumors is not primarily a failure of antigen presentation but of effector function. Even when neoantigen-specific T cells are generated, they encounter an immunosuppressive microenvironment dominated by M2-polarized tumor-associated macrophages (TAMs) [9]. A 2025 study demonstrated that LNPs formulated with HCQ-derived ionizable lipids achieve simultaneous spleen-tropic mRNA delivery and repolarization of M2 TAMs to M1 phenotype [5]. The mechanism involves HCQ-mediated elevation of lysosomal pH in macrophages, triggering calcium release via mucolipin-1 (TRPML1), activating p38 MAPK and NF-κB [5].
2.3. Pillar III: Autophagy Inhibition as a Vaccine Sensitizer
Autophagy is exploited by cancer cells as a survival mechanism under immune attack, particularly in KRAS-mutant tumors: pancreatic adenocarcinoma (KRAS mutated in 92%) and NSCLC (KRAS 33%) are among the most autophagy-dependent cancers [14]. Multiple clinical trials tested systemic HCQ with chemotherapy for pancreatic cancer with mixed results, likely due to insufficient intratumoral concentrations [26,27,28,29]. HCQ-LNPs solve this pharmacokinetic problem by delivering HCQ directly to the tumor microenvironment at concentrations orders of magnitude higher than achievable by oral dosing [5,6].
3. the Unified Hypothesis: HCQ-LNP Neoantigen Platform
We propose that a personalized mRNA neoantigen vaccine delivered via HCQ-derived ionizable lipid nanoparticles constitutes a quadruple-function therapeutic platform (Figure 1):
Function 1—Enhanced mRNA Delivery: HCQ-derived ionizable lipids modulate endosomal pH maturation, increasing mRNA escape to the cytoplasm and amplifying neoantigen presentation on MHC-I/II.
Function 2—TME Reprogramming: HCQ-LNPs reprogram M2 TAMs to M1 phenotype via lysosomal Ca2+/mucolipin-1/p38/NF-κB, converting immunosuppressive TME to immune-permissive.
Function 3—Autophagy Inhibition: Intracellular HCQ-lipid blocks autophagosome-lysosome fusion, disabling the cancer cell’s survival mechanism against immune attack.
Function 4—Anti-inflammatory Tolerance: HCQ-functionalized LNPs suppress proinflammatory cytokine storms, enabling multi-dose schedules (9 doses over 24 weeks) [6].
The unifying principle is that HCQ’s lysosomal pH-modulating activity, deployed at the nanoparticle level rather than systemically, simultaneously addresses delivery, microenvironment, and resistance barriers. This is a single molecular platform where every function emerges from lysosomal alkalinization at different biological scales.
4. Mechanistic Bridge from TB to Cancer: the pH Optimization Paradigm
This hypothesis represents a direct intellectual extension of our prior work on AI-guided phagolysosomal pH optimization for pyrazinamide-hydroxychloroquine synergy in tuberculosis (AIPH-TB framework) [13]. In that computational model, we demonstrated that HCQ’s ability to modulate phagolysosomal pH creates a tunable “pH window” optimizable for pH-dependent drug activation within macrophage phagolysosomes. The mechanistic parallel to cancer vaccine delivery is striking (Figure 3, Table 1).
5. AI-Driven Target Selection: Which Cold Tumors Benefit Most?
Our TVRS framework identifies tumors where the gap between conventional LNP vaccine responsiveness and HCQ-LNP potential is greatest. The pattern is clear: cancers with the lowest TVRS scores are characterized by high autophagy dependence, abundant M2 TAMs, and cold immune microenvironments—precisely the features HCQ-LNPs overcome (Figure 4, Table 2).
Pancreatic adenocarcinoma emerges as the ideal first target: its KRAS-driven autophagy addiction, dense M2-macrophage stroma, and 12% five-year survival create both the biological rationale and clinical urgency. This represents an inversion of conventional vaccine strategy—HCQ-LNP vaccines should target the tumors where the HCQ-derived lipid provides the greatest marginal benefit.
6. Proposed Experimental Validation Framework
6.1. Phase A: In Vitro Proof-of-Concept
Synthesize HCQ-derived ionizable lipids with varying alkyl chain lengths (C10, C12, C14). Formulate into LNPs (HCQ-lipid:DSPC:cholesterol:PEG-DMG at 50:10:38.5:1.5). Screen for transfection efficiency in human dendritic cells vs. SM-102 LNPs. Measure endosomal pH kinetics, M2→M1 repolarization, and autophagy inhibition (LC3-II/I ratio, p62 accumulation).
6.2. Phase B: In Vivo Preclinical
Using KPC pancreatic cancer mouse model, test: (1) PBS control; (2) anti-PD-1 alone; (3) neoantigen mRNA in SM-102 LNP + anti-PD-1; (4) neoantigen mRNA in HCQ-LNP + anti-PD-1; (5) empty HCQ-LNP + anti-PD-1; (6) oral HCQ + SM-102 vaccine + anti-PD-1. Primary endpoints: tumor volume, survival, intratumoral CD8+ T-cells. The critical comparison (arm 3 vs. 4) tests whether HCQ-LNP delivery demonstrates superior tumor control.
6.3. Phase C: AI-Optimized Neoantigen Selection
For clinical-grade vaccine: (1) tumor/normal WES with Mutect2/Strelka2; (2) NetMHCpan 4.1 + MHCflurry + DeepImmuno prediction; (3) Saudi HLA-optimized SNIP ensemble ranking; (4) AlphaFold 3 structural validation; (5) CodonBERT + LinearDesign mRNA optimization; (6) TVRS-based patient stratification [11,12,13].
7. Clinical Translation Pathway
Track 1 (Near-term): Phase I/II of personalized mRNA neoantigen vaccine in conventional SM-102 LNPs + pembrolizumab for advanced melanoma (Protocol FRHC-NEO-MEL-2026-001). Establishes clinical infrastructure and provides control data.
Track 2 (Medium-term): Following preclinical HCQ-LNP validation, Phase I trial specifically targeting autophagy-dependent cold tumors (pancreatic, glioblastoma) to test the central hypothesis that HCQ-LNP delivery overcomes immune resistance.
8. Potential Limitations and Safety Considerations
Several potential challenges must be acknowledged. First, the optimal HCQ-lipid concentration that balances endosomal escape enhancement against excessive lysosomal disruption is unknown and will require systematic titration. Excessive lysosomal damage could trigger galectin-mediated innate immune activation, paradoxically increasing inflammatory side effects. Second, the long-term safety of repeated intramuscular injection of HCQ-derived lipids is uncharacterized; while acute toxicity data from published studies are reassuring, chronic retinal and cardiac monitoring will be essential in clinical trials. Third, M2→M1 repolarization has been demonstrated only in mouse models; human TAMs may respond differently due to species-specific differences in macrophage biology. Fourth, autophagy inhibition by HCQ-LNPs may have unintended consequences on antigen cross-presentation by dendritic cells, which itself relies partially on autophagic pathways for MHC-II loading.
8.1. Critical Tumor-Type-Specific Consideration: The Zinc Ionophore Paradox
A particularly important safety consideration concerns HCQ’s well-characterized function as a zinc ionophore. HCQ directs zinc ions to lysosomes and increases intracellular free zinc concentrations. While zinc-mediated apoptosis amplification has been proposed as a therapeutic strategy in some cancers, and the combination of zinc with chloroquine enhances cytotoxicity in ovarian cancer models, recent evidence reveals a starkly different picture in triple-negative breast cancer (TNBC).
Specifically, brain-metastatic TNBC cells (MDA-MB-BrM2) contain double the zinc concentration of parental MDA-MB-231 cells. Elevated intracellular zinc in TNBC promotes: (a) upregulation of SerpinB2, which facilitates brain and lung metastasis by blocking niche defenses against cancer cell invasion; (b) enhanced tumorigenic capacity of breast cancer stem cells; and (c) activation of zinc-dependent matrix metalloproteinases (MMP-9, MMP-14) that drive epithelial-mesenchymal transition (EMT) and cancer cell motility. Furthermore, the zinc transporter ZIP6, transcriptionally activated by STAT3, promotes Snail-mediated E-cadherin repression, directly facilitating cell dissociation and metastatic spread. The zinc transporter LIV1 (SLC39A6) is overexpressed in TNBC and has been linked to EMT and early lymph node metastasis.
This tumor-type-specific duality of zinc biology represents a critical safety consideration for the HCQ-LNP platform. In KRAS-mutant pancreatic adenocarcinoma, where the risk-benefit profile favors autophagy inhibition and where zinc-mediated apoptosis may be beneficial, HCQ’s zinc ionophore activity is likely advantageous. However, in TNBC, the same zinc ionophore activity could paradoxically promote metastatic progression by fueling zinc-dependent pro-tumorigenic pathways.
We therefore propose that clinical application of HCQ-LNP neoantigen vaccines must be preceded by cancer-type-specific zinc profiling, including assessment of ZIP/ZnT transporter expression, baseline intracellular zinc levels, and zinc-dependent pathway activation status. For TNBC applications specifically, future formulations may require co-encapsulation of zinc chelators (e.g., TPEN) within the HCQ-LNP to neutralize the zinc ionophore effect while preserving the pH-modulating, autophagy-inhibiting, and M2→M1 reprogramming functions. This represents a tractable engineering challenge that does not invalidate the core hypothesis but rather refines its clinical application to be tumor-type-aware.
8.2. Future Directions: COX-2 Inhibition as an Adjunct Strategy
Beyond the HCQ-LNP platform itself, systemic COX-2 inhibition (e.g., celecoxib) may provide complementary benefit as an oral adjunct to HCQ-LNP vaccination. PGE2, the dominant immunosuppressive prostanoid in the tumor microenvironment of pancreatic and colorectal cancers, suppresses dendritic cell maturation and promotes M2 macrophage polarization through EP2/EP4 receptor signaling. Celecoxib-mediated COX-2 inhibition could synergize with HCQ-LNP’s M2→M1 reprogramming by simultaneously reducing de novo PGE2 production while the nanoparticle repolarizes existing M2 TAMs. This combination—HCQ-LNP neoantigen vaccine plus celecoxib plus pembrolizumab—represents a triple-blockade strategy against immunosuppression that warrants investigation in a separate study.
9. Conclusions
We present a unifying hypothesis that transforms hydroxychloroquine from a repurposed anticancer adjuvant into the structural foundation of a next-generation mRNA cancer vaccine delivery platform. By incorporating HCQ-derived ionizable lipids into the LNP formulation, a single nanoparticle achieves four therapeutic functions—enhanced mRNA delivery, TME reprogramming, autophagy inhibition, and anti-inflammatory tolerance—all from lysosomal pH modulation.
This hypothesis is rooted in the convergence of our TVRS framework, our prior AIPH-TB investigation, and recent independent demonstrations that HCQ-derived lipids achieve spleen-tropic mRNA delivery with immunomodulatory activity [5,6]. If validated, this platform could extend personalized mRNA cancer vaccines to cold tumors—pancreatic, glioblastoma, prostate—representing the greatest unmet need in oncology.
Author Contributions
A.K.A. conceived the hypothesis, designed the TVRS computational framework, performed the pan-cancer analysis, identified the HCQ-LNP convergence, and wrote the manuscript. S.R. contributed to the AIPH-TB framework. Both authors approved the final manuscript.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Data Availability Statement
TVRS analysis code and TCGA data available from the corresponding author upon request. TCGA data publicly available at https://portal.gdc.cancer.gov/.
Conflicts of Interest
The authors declare no competing interests.
Declaration of Generative AI and AI-Assisted Technologies in the Manuscript Preparation Process
During the preparation of this work the author(s) used Claude (Anthropic, Claude Opus 4.6) in order to assist with literature search and synthesis across multiple databases (TCGA Pan-Cancer Atlas, cBioPortal, PubMed), computational analysis of genomic data for the Tumor Vaccine Responsiveness Score (TVRS) framework, generation of Python code for the neoantigen prioritization pipeline, statistical modeling of tumor mutational burden and neoantigen load estimates, drafting and structuring of manuscript sections, and creation of scientific figures. After using this tool/service, the author(s) reviewed and edited the content as needed and take(s) full responsibility for the content of the published article.
References
- Weber, JS; Carlino, MS; Khattak, A; et al. Individualised neoantigen therapy mRNA-4157 (V940) plus pembrolizumab versus pembrolizumab monotherapy in resected melanoma (KEYNOTE-942): a randomised, phase 2b study. Lancet. 2024, 403(10427), 632–644. [Google Scholar] [CrossRef]
- Sahin, U; et al. Individualized mRNA vaccines evoke durable T cell immunity in adjuvant triple-negative breast cancer. In Nature.; 2026. [Google Scholar]
- Rojas, LA; Löber, A; Gukasyan, J; et al. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature. 2023, 618(7963), 144–150. [Google Scholar] [CrossRef]
- Conyngham, P; et al. AI-designed personalized mRNA cancer vaccine for a canine mast cell tumor: a case report; Preprint; UNSW/University of Queensland, March 2026. [Google Scholar]
- Fan, J; Wei, Q; Yuan, P; Xiao, B; Yao, S; Xu, H; Liu, J; Li, R; et al. Hydroxychloroquine-Derived Ionizable Lipid Facilitates Spleen-Tropic mRNA Transfection and Enhances Cancer Immunotherapy. ACS Chem Bio Eng. 2025. [Google Scholar] [CrossRef] [PubMed]
- Chen, K; Li, X; et al. Hydroxychloroquine-functionalized ionizable lipids mitigate inflammatory responses in mRNA therapeutics. J Control Release. 2025, 387, 114267. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G; Zitvogel, L. Chloroquine and hydroxychloroquine for cancer therapy. Mol Cell Oncol. 2014, 1(1), e29911. [Google Scholar] [CrossRef]
- Patel, S; et al. Endosomal escape: A bottleneck for LNP-mediated therapeutics. Adv Drug Deliv Rev. 2024, 206, 115185. [Google Scholar] [CrossRef]
- Thorsson, V; Gibbs, DL; Brown, SD; et al. The immune landscape of cancer. Immunity. 2018, 48(4), 812–830. [Google Scholar] [CrossRef] [PubMed]
- Hoadley, KA; Yau, C; Hinoue, T; et al. Cell-of-origin patterns dominate the molecular classification of 10,000 tumors from 33 types of cancer. Cell. 2018, 173(2), 291–304. [Google Scholar] [CrossRef]
- Jumper, J; Evans, R; Pritzel, A; et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021, 596(7873), 583–589. [Google Scholar] [CrossRef]
- Reynisson, B; Alvarez, B; Paul, S; Peters, B; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 2020, 48(W1), W449–W454. [Google Scholar] [CrossRef]
- Ahmed, AK. AI-guided phagolysosomal pH optimization for pyrazinamide-hydroxychloroquine synergy in tuberculosis (AIPH-TB framework). Manuscript in preparation 2026. [Google Scholar]
- Yang, B; et al. Autophagy as a therapeutic target in pancreatic cancer. Br J Cancer. 2021, 124(8), 1455–1466. [Google Scholar] [CrossRef]
- Levy, JMM; Thompson, JC; Bhatt, UY; et al. Autophagy inhibition improves chemosensitivity in BRAF(V600E) brain tumors. Cancer Discov. 2014, 4(7), 773–780. [Google Scholar] [CrossRef]
- Galluzzi, L; Pietrocola, F; Bravo-San Pedro, JM; et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015, 34(7), 856–880. [Google Scholar] [CrossRef]
- Li, Y; Ye, Z; Lan, T; Liao, G; Liang, J. Design strategies for novel lipid nanoparticles for mRNA vaccine and therapeutics. MedComm. 2025, 6(1), e70414. [Google Scholar] [CrossRef] [PubMed]
- Jones, TM; et al. Moving beyond hydroxychloroquine: the novel lysosomal autophagy inhibitor ROC-325 shows superior autophagy inhibition and anti-tumor activity. Cancer Commun. 2019, 39(1), 72. [Google Scholar] [CrossRef] [PubMed]
- Ellrott, K; Bailey, MH; Saksena, G; et al. Scalable open science approach for mutation calling of tumor exomes using multiple genomic pipelines. Cell Syst. 2018, 6(3), 271–281. [Google Scholar] [CrossRef]
- Bailey, MH; Tokheim, C; Porta-Pardo, E; et al. Comprehensive characterization of cancer driver genes and mutations. Cell. 2018, 174(4), 1034–1035. [Google Scholar] [CrossRef]
- Sanchez-Vega, F; Mina, M; Armenia, J; et al. Oncogenic signaling pathways in The Cancer Genome Atlas. Cell. 2018, 173(2), 321–337. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U; Derhovanessian, E; Miller, M; et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature. 2017, 547(7662), 222–226. [Google Scholar] [CrossRef]
- Ott, PA; Hu, Z; Keskin, DB; et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature. 2017, 547(7662), 217–221. [Google Scholar] [CrossRef] [PubMed]
- Rebecca, VW; Amaravadi, RK. A unified approach to targeting the lysosome’s degradative and secretory functions in cancer. Cancer Discov. 2017, 7(11), 1266–1283. [Google Scholar] [CrossRef]
- Amaravadi, RK; Kimmelman, AC; Debnath, J. Targeting autophagy in cancer: recent advances and future directions. Cancer Discov. 2019, 9(9), 1167–1181. [Google Scholar] [CrossRef]
- Mahalingam, D; Mita, M; Sarantopoulos, J; et al. Combined autophagy and HDAC inhibition: a phase I safety, tolerability, pharmacokinetic, and pharmacodynamic analysis of hydroxychloroquine in combination with the HDAC inhibitor vorinostat in patients with advanced solid tumors. Autophagy. 2014, 10(8), 1403–1414. [Google Scholar] [CrossRef]
- Zeh, HJ; Bahary, N; Boone, BA; et al. A randomized phase II preoperative study of autophagy inhibition with high-dose hydroxychloroquine and gemcitabine/nab-paclitaxel in pancreatic cancer (SWOG S1214). Clin Cancer Res. 2020, 26(13), 3126–3134. [Google Scholar] [CrossRef]
- Karasic, TB; O’Hara, MH; Loaiza-Bonilla, A; et al. Effect of gemcitabine and nab-paclitaxel with or without hydroxychloroquine on patients with advanced pancreatic cancer: a phase 2 randomized clinical trial. JAMA Oncol. 2019, 5(7), 993–998. [Google Scholar] [CrossRef] [PubMed]
- Boone, BA; Bahary, N; Zureikat, AH; et al. Safety and biologic response of pre-operative autophagy inhibition in combination with gemcitabine in patients with pancreatic adenocarcinoma. Ann Surg Oncol. 2015, 22(13), 4402–4410. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
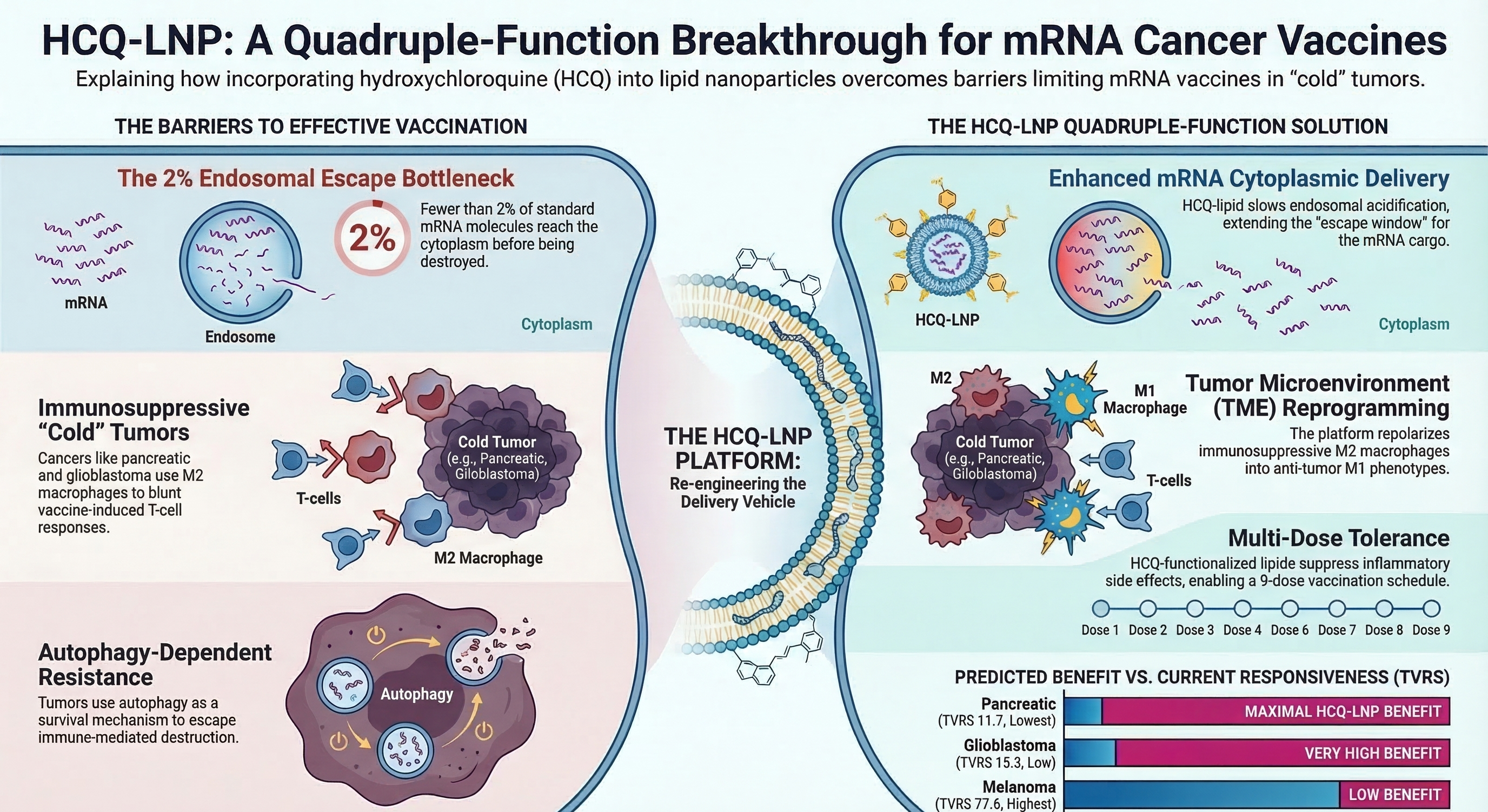
The HCQ-LNP quadruple-function personalized mRNA neoantigen vaccine platform. A single nanoparticle composed of HCQ-derived ionizable lipid encapsulating patient-specific mRNA neoantigens achieves four therapeutic functions from one physicochemical property: lysosomal pH modulation. Function 1: enhanced mRNA endosomal escape via delayed acidification. Function 2: tumor microenvironment (TME) reprogramming through M2→M1 macrophage repolarization via the Ca2+/mucolipin-1/p38/NF-κB axis. Function 3: autophagy inhibition by blocking autophagosome-lysosome fusion. Function 4: anti-inflammatory tolerance enabling 9-dose vaccination schedules. These functions converge to produce synergistic anti-tumor immunity even in cold tumors (pancreatic, glioblastoma), particularly when combined with a checkpoint inhibitor (pembrolizumab).
Figure 1.
The HCQ-LNP quadruple-function personalized mRNA neoantigen vaccine platform. A single nanoparticle composed of HCQ-derived ionizable lipid encapsulating patient-specific mRNA neoantigens achieves four therapeutic functions from one physicochemical property: lysosomal pH modulation. Function 1: enhanced mRNA endosomal escape via delayed acidification. Function 2: tumor microenvironment (TME) reprogramming through M2→M1 macrophage repolarization via the Ca2+/mucolipin-1/p38/NF-κB axis. Function 3: autophagy inhibition by blocking autophagosome-lysosome fusion. Function 4: anti-inflammatory tolerance enabling 9-dose vaccination schedules. These functions converge to produce synergistic anti-tumor immunity even in cold tumors (pancreatic, glioblastoma), particularly when combined with a checkpoint inhibitor (pembrolizumab).

Figure 2.
Endosomal escape comparison between standard SM-102 LNP and the proposed HCQ-LNP platform. (A) Standard LNP: rapid endosomal acidification creates a narrow escape window (~10 minutes at pH 5.5–6.0), resulting in ~98% mRNA degradation in lysosomes and only ~2% cytoplasmic delivery. (B) HCQ-LNP: the HCQ-derived ionizable lipid slows endosomal pH maturation by 30–60 minutes, extending the late endosomal residence time at pH 5.5–5.8 where productive escape occurs. This is predicted to increase cytoplasmic mRNA delivery to >5%, enhancing neoantigen protein expression in antigen-presenting cells. The HCQ-LNP platform thus converts the delivery limitation from a systemic pharmacokinetic problem (oral HCQ dosing) into a subcellular biophysical solution (nanoparticle-intrinsic pH modulation).
Figure 2.
Endosomal escape comparison between standard SM-102 LNP and the proposed HCQ-LNP platform. (A) Standard LNP: rapid endosomal acidification creates a narrow escape window (~10 minutes at pH 5.5–6.0), resulting in ~98% mRNA degradation in lysosomes and only ~2% cytoplasmic delivery. (B) HCQ-LNP: the HCQ-derived ionizable lipid slows endosomal pH maturation by 30–60 minutes, extending the late endosomal residence time at pH 5.5–5.8 where productive escape occurs. This is predicted to increase cytoplasmic mRNA delivery to >5%, enhancing neoantigen protein expression in antigen-presenting cells. The HCQ-LNP platform thus converts the delivery limitation from a systemic pharmacokinetic problem (oral HCQ dosing) into a subcellular biophysical solution (nanoparticle-intrinsic pH modulation).

Figure 3.
The pH optimization paradigm: mechanistic bridge from tuberculosis (AIPH-TB framework) to cancer vaccines (HCQ-LNP platform). Five parallel mechanistic layers demonstrate that the same fundamental principle—AI-optimized modulation of intracellular pH to enhance efficacy of a pH-dependent therapeutic cargo—applies across both disease domains. In tuberculosis, HCQ raises phagolysosomal pH to create the optimal activation window for pyrazinamide within M. tuberculosis-infected macrophages. In cancer, HCQ-derived ionizable lipid slows endosomal acidification to extend the mRNA escape window within tumor-associated macrophages and dendritic cells. Both systems target the same cell type (macrophage), modulate the same organelle (phagolysosome/endolysosome), and can be computationally optimized using AI-guided pH modeling. The cancer application extends the TB framework by adding AlphaFold structural modeling, the SNIP neoantigen predictor, and the TVRS scoring system for patient stratification.
Figure 3.
The pH optimization paradigm: mechanistic bridge from tuberculosis (AIPH-TB framework) to cancer vaccines (HCQ-LNP platform). Five parallel mechanistic layers demonstrate that the same fundamental principle—AI-optimized modulation of intracellular pH to enhance efficacy of a pH-dependent therapeutic cargo—applies across both disease domains. In tuberculosis, HCQ raises phagolysosomal pH to create the optimal activation window for pyrazinamide within M. tuberculosis-infected macrophages. In cancer, HCQ-derived ionizable lipid slows endosomal acidification to extend the mRNA escape window within tumor-associated macrophages and dendritic cells. Both systems target the same cell type (macrophage), modulate the same organelle (phagolysosome/endolysosome), and can be computationally optimized using AI-guided pH modeling. The cancer application extends the TB framework by adding AlphaFold structural modeling, the SNIP neoantigen predictor, and the TVRS scoring system for patient stratification.

Figure 4.
Predicted HCQ-LNP benefit inversely correlates with TVRS score. Cancer types are ranked by their Tumor Vaccine Responsiveness Score (TVRS), with corresponding TMB, immune-hot percentage, autophagy dependence (stars), and predicted HCQ-LNP benefit. Pancreatic adenocarcinoma (TVRS 11.7) and glioblastoma (TVRS 15.3) show maximal predicted benefit due to extreme autophagy dependence (KRAS-driven in PDAC), cold tumor microenvironments (<15% immune-hot), and low TMB. In contrast, melanoma (TVRS 77.6) shows minimal additional benefit as it already responds well to standard mRNA-LNP vaccines. This inverse relationship supports a strategy inversion: rather than beginning clinical development in melanoma (easiest target), HCQ-LNP vaccines should be tested first in autophagy-dependent cold tumors where the HCQ-derived lipid provides the greatest marginal therapeutic benefit. Data derived from TCGA Pan-Cancer Atlas (8,432 samples, 20 cancer types) analyzed through the TVRS computational framework.
Figure 4.
Predicted HCQ-LNP benefit inversely correlates with TVRS score. Cancer types are ranked by their Tumor Vaccine Responsiveness Score (TVRS), with corresponding TMB, immune-hot percentage, autophagy dependence (stars), and predicted HCQ-LNP benefit. Pancreatic adenocarcinoma (TVRS 11.7) and glioblastoma (TVRS 15.3) show maximal predicted benefit due to extreme autophagy dependence (KRAS-driven in PDAC), cold tumor microenvironments (<15% immune-hot), and low TMB. In contrast, melanoma (TVRS 77.6) shows minimal additional benefit as it already responds well to standard mRNA-LNP vaccines. This inverse relationship supports a strategy inversion: rather than beginning clinical development in melanoma (easiest target), HCQ-LNP vaccines should be tested first in autophagy-dependent cold tumors where the HCQ-derived lipid provides the greatest marginal therapeutic benefit. Data derived from TCGA Pan-Cancer Atlas (8,432 samples, 20 cancer types) analyzed through the TVRS computational framework.


Table 1.
Mechanistic parallels between AIPH-TB and HCQ-LNP cancer vaccine platform.
| Parameter | TB (AIPH-TB Model) | Cancer Vaccine (HCQ-LNP) |
|---|---|---|
| Target compartment | Macrophage phagolysosome (pH 4.5–5.5) | Endosome/lysosome of APCs and tumor cells (pH 4.5–6.5) |
| pH-dependent cargo | Pyrazinamide (activated at pH <5.5) | mRNA (degraded at pH <5.0; escapes at pH 5.5–6.0) |
| HCQ mechanism | Raises phagolysosomal pH to optimal PZA window | Slows endosomal acidification, prolonging escape window |
| AI optimization | Computational pH–drug activation curves | SNIP + AlphaFold + TVRS neoantigen pipeline |
| Target cell | M. tuberculosis–infected macrophage | Tumor-associated macrophage + dendritic cell |
| Immune effect | Enhanced macrophage bactericidal activity | M2→M1 repolarization + enhanced antigen presentation |
| Resistance blocked | M.tb pH-neutralization of phagosome | Cancer cell autophagy-mediated immune evasion |
Table 2.
TVRS scores and predicted HCQ-LNP benefit by cancer type.
| Cancer Type | TVRS | TMB | Hot% | MSI-H | Auto * | M2TAM | KRAS% | HCQ-LNP Gain |
|---|---|---|---|---|---|---|---|---|
| Pancreatic | 11.7 | 1.1 | 10% | 1% | +++++ | High | 92% | MAXIMAL |
| Glioblastoma | 15.3 | 1.8 | 12% | 1% | ++++ | High | 0% | VERY HIGH |
| Prostate | 14.3 | 0.9 | 15% | 2.5% | +++ | Mod | 0% | HIGH |
| CRC (MSS) | 32.6 | 3.5 | 28% | 0% | +++ | Mod | 42% | HIGH |
| Breast (TNBC) | 20.9 | 1.5 | 30% | 1.2% | +++ | Mod | 0% | MODERATE |
| Melanoma | 77.6 | 13.5 | 65% | 1.5% | + | Low | 0% | LOW |
* Autophagy dependence: + (low) to +++++ (very high) based on published literature. MSS = microsatellite stable.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



