Submitted:
16 March 2026
Posted:
19 March 2026
You are already at the latest version
Abstract
Basement membrane (BM) is an essential part of epithelial cell architecture. BM is a specialized extracellular matrix. BM assembles on the basal side outside of plasma membrane. It carries out important biological functions such as maintaining tissue shape, enabling cellular migration, and facilitating communication between cells. Defects in BM assembly or composition can cause a range of diseases. However, the regulatory mechanisms governing polarized deposition of BM are not well understood. To study this process, I use the follicular epithelium of the Drosophila melanogaster ovary as a model system. In this study, I analyzed the role of SNARE proteins in intracellular trafficking, secretion and polarized deposition of the BM. SNAREs are crucial for membrane fusion. I performed a genetic screen targeting all SNARE proteins in Drosophila. Individual knockdowns of multiple SNARE family members resulted in BM mislocalization. Three distinct mislocalization phenotypes were observed: Intracellular accumulation of BM, apical deposition of BM, or a combination of apical deposition and intracellular accumulation of BM. Taken together, these observations suggest that multiple SNARE family members contribute to the intracellular trafficking, secretion, and polarized deposition of BM proteins.
Keywords:
Drosophila
; genetic screening
; SNARE
; intracellular trafficking
; secretion
; polarized deposition
; apical-basal polarity
; basement membrane
; Crag
; Rab10
1. Introduction
Epithelia are sheets of tightly adherent cells that form selective barriers between the internal tissues and the external environment. Many essential organs, including the kidney, lung, mammary glands, and liver, contain hollow spaces lined by epithelial layers that selectively permit the exchange between different parts of the body [1]. Epithelial cells perform a variety of functions during both the embryonic and adult stages [2]. As a result, disruptions in epithelial integrity impair proper gastrulation movements and normal tissue morphogenesis [3].
The functional diversity of epithelial tissues is fundamentally dependent on the polarized architecture of epithelial cells [2]. Epithelial cells exhibit a profound apical-basal (AB) polarity [4]. A critical component of the AB polarity is the proper placement of the basement membrane (BM). A conceptual diagram of epithelial polarity and BM organization is shown in Figure 1. The BMs are sheetlike extracellular matrices found at the basal surfaces of epithelial tissues [5]. The BMs are composed of a number of proteins including Laminin, type IV collagen, Nidogen and Perlecan, etc [6]. BM facilitates a number of biological functions including maintenance of tissue shape, and tissue integrity, cellular migration, cell-cell communication, and cell-matrix adhesion [5,7]. Defects in BM assembly and composition can lead to a wide range of diseases, including skin blistering, muscular dystrophy, neuromuscular junction defects, kidney failure, and lung hemorrhage [8]. Thus, correct assembly of the BM on the basal surface is crucial.
For the proper assembly of BM, newly synthesized BM proteins need to go through polarized trafficking, secretion and deposition to accumulate at the basal surface of the epithelial cells [5]. Our knowledge about the molecular network regulating polarized secretion and deposition of BM is very little. Newly synthesized BM proteins inside of the cell need tightly regulated vesicle mediated trafficking and secretion. This vesicle-mediated transportation requires the fusion of vesicles with their target compartments [9]. The SNARE (Soluble N-ethylmaleimide-sensitive-factor Attachment Protein Receptor) protein family has an active role in this fusion process [10]. Given this SNARE is a potential candidate which plays role in polarized secretion and deposition of BM.
Fusion step of vesicle transport is mediated by SNARE protein family [11,12,13,14]. Each of the family members has conserved SNARE motif [15]. Functionally, SNAREs were classified into two categories: v-SNARE and t-SNARE [14,15]. If SNARE is localized at the donor compartment, then it is known as v-SNARE (vesicle membrane SNARE) [14,15]. If a SNARE is localized at the acceptor compartment membrane, then it is known as t-SNARE (target membrane SNARE) [14,15]. Structurally SNARE is divided into two classes: Q-SNARE and R-SNARE [15]. Q SNAREs are further separated into Qa, Qb, and Qc categories based on the specific position each helix occupies within the bundle [16]. These proteins assemble into the characteristic four-helix structure required for membrane fusion [17]. Although SNARE proteins are well characterized in membrane fusion and intracellular trafficking, their involvement in polarized BM secretion remains unknown.
To study the role of SNAREs in the polarized secretion of BM, I use the Drosophila melanogaster FE as a model system. FE is part of the Drosophila egg chamber formed inside ovary during oogenesis. The FE consists of a monolayer of somatic epithelial cells that surrounds the germline cells in the developing egg chamber [18]. FE is a classical epithelium that contains all polarity domains including apical and basal domain [2,19]. BM underlies the basal side of the FE [19]. Drosophila FE synthesizes and secretes all of the major BM proteins [5]. Thus, Drosophila FE is an excellent model system to study BM secretion.
Using this model system, I conducted a genetic screen targeting all members of the Drosophila SNARE family. My screening results show that individual knockdown of multiple SNARE family members leads to BM mislocalization. Analysis of the phenotypes revealed three distinct patterns of BM mislocalization: intracellular accumulation, apical deposition, and concurrent intracellular accumulation with apical deposition. Together, these results indicate that multiple SNARE family members are involved in regulating polarized trafficking, secretion and polarized deposition of BM.
2. Materials and Methods
Fly stocks and Genetics
Details of the Drosophila melanogaster SNARE gene family RNAi lines obtained from the Bloomington Drosophila Stock Center are provided in the Supplemental Materials. Basement membrane protein trap lines, including Vkg GFP (CC00791) and Pcan GFP (ZCL1700), were obtained from the FlyTrap collection. Tissue specific knockdowns were performed using an ovarian somatic cell–specific GAL4 driver line (kindly provided as a gift).
Screening and Phenotype Analysis
Tissue-specific knockdown of candidate genes was assessed for BM protein mislocalization. Female Drosophila melanogaster ovaries were dissected in phosphate-buffered saline (PBS), fixed in PBS containing 4% paraformaldehyde for 20 minutes at room temperature, and processed for staining using established protocols [20]. Imaging was performed using epifluorescence, confocal, and super-resolution microscopy.
3. Results
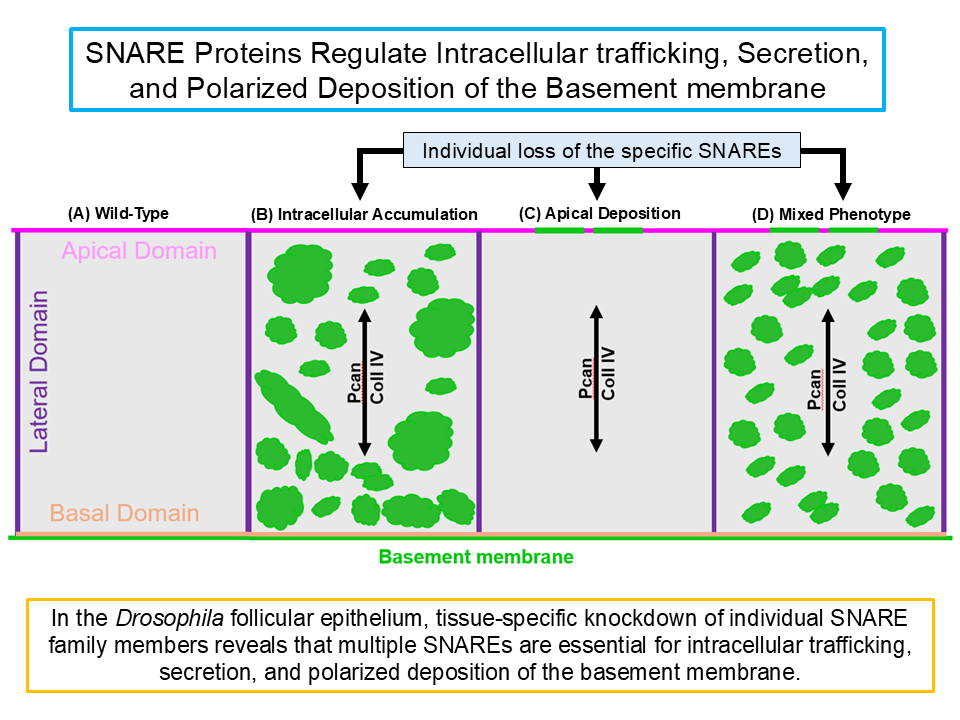
To investigate the role of SNARE family members in polarized BM secretion, a tissue-specific RNAi screen was conducted targeting all 25 Drosophila SNARE genes. In total, 35 RNAi lines were used, as several genes were targeted by more than one independent RNAi construct. The effects of individual SNARE knockdown were assessed using several GFP-tagged BM proteins as readouts. This screen identified multiple SNAREs, and their loss affected BM localization in follicular epithelial cells, resulting in three broad phenotypic classes (Fig. 2). In one class, knockdown of specific SNAREs led to intracellular accumulation of BM proteins, suggesting a role in intracellular trafficking and/or secretion of BM. In a second class, individual SNARE knockdown caused ectopic deposition of BM at the apical surface, indicating a requirement for polarized secretion. In a third class, a mixed phenotype was observed following individual SNARE knockdown, with intracellular accumulation and apical deposition of BM, suggesting roles in vesicle trafficking and in maintaining the polarity of secretion.
Figure 2.
Conceptual phenotypes associated with tissue-specific knockdown of individual SNARE family members. Tissue-specific knockdown of individual SNARE family members in Drosophila follicular epithelium shows three representative phenotypes of basement membrane (BM) mislocalization. (A) Wild-Type cell shows regular BM protein localization. (B) Representative phenotype Class I shows intracellular accumulation of BM proteins. (C) Representative phenotype Class II shows apical deposition of BM proteins. (D) Representative phenotype Class III shows both intracellular accumulation and apical deposition of BM proteins. Figures are cartoon schematics intended for conceptual illustration and do not represent raw experimental data.
Figure 2.
Conceptual phenotypes associated with tissue-specific knockdown of individual SNARE family members. Tissue-specific knockdown of individual SNARE family members in Drosophila follicular epithelium shows three representative phenotypes of basement membrane (BM) mislocalization. (A) Wild-Type cell shows regular BM protein localization. (B) Representative phenotype Class I shows intracellular accumulation of BM proteins. (C) Representative phenotype Class II shows apical deposition of BM proteins. (D) Representative phenotype Class III shows both intracellular accumulation and apical deposition of BM proteins. Figures are cartoon schematics intended for conceptual illustration and do not represent raw experimental data.

4. Discussion
Polarized secretion is a fundamental architectural feature of epithelial cells. In the Drosophila FE, newly synthesized BM proteins are targeted for secretion at the basal plasma membrane. Polarized secretion is achieved through tightly regulated intracellular trafficking mechanisms that include vesicle transport, docking to the target membrane, and subsequent membrane fusion to release cargo.
SNARE proteins function at the final stage of vesicular trafficking, where they directly mediate membrane fusion with the target compartment. In this study, depletion of individual SNARE family members resulting in mislocalization of BM proteins. The observed BM mislocalization phenotype includes intracellular accumulation of BM proteins, their ectopic deposition at the apical side, or the coexistence of both phenotypes within the same cell. These results indicate that multiple SNARE family members are involved in polarized trafficking and secretion of BM proteins.
The BM mislocalization phenotype characterized by intracellular accumulation of BM proteins suggests defects at multiple stages of the secretory pathway. These defects may arise from impaired vesicle maturation, vesicle trafficking, or disrupted fusion with intracellular organelles such as the Golgi apparatus. Alternatively, intracellular accumulation may occur even when vesicle formation, maturation and trafficking proceed normally, but loss of specific SNARE compromises vesicle fusion with the plasma membrane. In this scenario, BM cargo containing vesicles become accumulated within the cell, consistent with the well-established role of SNAREs in mediating membrane fusion.
Apical deposition of BM proteins suggests that vesicles are redirected to the incorrect membrane domain. In this scenario, a SNARE normally mediates fusion with the basal plasma membrane. Upon knockdown, vesicles accumulate, increasing the vesicle load in the cytoplasm. Although Crag activated Rab10 typically blocks apical misrouting of vesicles, excessive vesicle accumulation may overwhelm this system, allowing some vesicles to reach the apical membrane [19,21]. Fusion at the apical site may then occur via a paralogue SNARE expressed at the apical domain, resulting in apical BM deposition.
The mixed phenotype—intracellular accumulation combined with apical deposition—may reflect SNARE proteins with dual functions. One role could be upstream of the Crag/Rab10 pathway, facilitating vesicle formation, maturation, or fusion with the Golgi, so that knockdown leads to cytoplasmic accumulation. Another role could be at the plasma membrane, where paralogous SNAREs on the apical surface mediate vesicle fusion, explaining ectopic apical secretion.
Together, these observations support a conceptual model in which SNARE proteins operate at multiple stages of BM secretion: upstream in vesicle trafficking, including vesicle generation and fusion with intracellular organelles, and downstream in fusion at the basal plasma membrane. Integration with the Crag/Rab10 pathway ensures directional trafficking and domain specificity, maintaining epithelial polarity and proper BM deposition. Figure 3 illustrates the proposed conceptual model for SNARE-mediated regulation of intracellular trafficking, secretion and polarized deposition of the basement membrane in Drosophila FE. Because the available literature primarily links Stratum/Rab8 pathway to basal vesicle targeting but does not clearly address their potential roles in preventing apical secretion, these components were not incorporated into the present conceptual model [22].
Future Directions
The distinct phenotypes observed provide a framework for dissecting the molecular mechanisms of polarized BM secretion.
1. Intracellular localization studies could determine where intracellularly accumulated BM proteins reside whether in vesicles, Golgi, or ER. Immunostaining or live imaging of fluorescently tagged BM proteins may reveal trafficking dynamics and block points in the pathway.
2. Live-cell imaging could visualize vesicle transport and exocytosis in real time, helping to identify whether vesicles fail to reach the basal membrane, accumulate intracellularly, or are redirected to the apical membrane.
3. Genetic interaction analyses could probe the functional relationships between SNAREs and upstream trafficking regulators such as Crag and Rab10. Double knockdowns experiments analyzed via epistasis can reveal whether SNAREs act upstream, downstream, or in parallel with these regulators.
4. Colocalization and localization studies of individual SNARE protein with Rab10 or Crag-positive vesicles can clarify which vesicle populations each SNARE operates on, and whether paralogous SNAREs contribute to apical fusion.
5. Functional assays examining apical-basal polarity marker localization and levels, along with cell morphology, in individual SNARE deficient cells can reveal how trafficking defects impact tissue architecture.
Overall, these experiments will help define how SNARE proteins contribute to intracellular trafficking, secretion, and polarized deposition of the basement membrane, and how they functionally integrate with the existing Crag/Rab10 pathway.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. List of Drosophila melanogaster SNARE genes and corresponding RNAi lines used in this study. For each line, the Bloomington Drosophila Stock Center (BDSC) stock number and the full genotype information, as provided by the stock center, are listed.
Author Contributions
Conceptualization, N.J.; methodology, N.J.; formal analysis, N.J.; investigation, N.J.; writing—original draft preparation, N.J.; writing—review and editing, N.J.; visualization, N.J. The author has read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Access to the raw data generated in this study is currently restricted during the preprint stage due to publication considerations. The data may be made available from the corresponding author upon reasonable request. Summary data supporting the findings are provided in the figures.
Acknowledgments
Not applicable.
Conflicts of Interest
The author declares no conflicts of interest.
AI Assistance Statement
Artificial intelligence-based language tools were used for limited assistance in editing and refining the wording of portions of this manuscript. All scientific ideas, conceptual models, data interpretation, and conclusions were developed by the author.
References
- Roignot, J.; Peng, X.; Mostov, K. Polarity in mammalian epithelial morphogenesis. Cold Spring Harbor perspectives in biology 2013, 5(2), a013789. [Google Scholar] [CrossRef]
- Müller, H. A. J.; Bossinger, O. Molecular networks controlling epithelial cell polarity in development. Mechanisms of development 2003, 120(11), 1231–1256. [Google Scholar] [CrossRef] [PubMed]
- Cox, R. T.; Kirkpatrick, C.; Peifer, M. Armadillo is required for adherens junction assembly, cell polarity, and morphogenesis during Drosophila embryogenesis. The Journal of cell biology 1996, 134(1), 133–148. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W. J. Adaptation of core mechanisms to generate cell polarity. Nature 2003, 422(6933), 766–774. [Google Scholar] [CrossRef] [PubMed]
- Isabella, A. J.; Horne-Badovinac, S. Building from the ground up: basement membranes in Drosophila development. Current topics in membranes 2015, 76, 305–336. [Google Scholar] [CrossRef]
- Halfter, W.; Oertle, P.; Monnier, C. A.; Camenzind, L.; Reyes-Lua, M.; Hu, H.; Plodinec, M. New concepts in basement membrane biology. The FEBS journal 2015, 282(23), 4466–4479. [Google Scholar] [CrossRef]
- Roca-Cusachs, P.; Sunyer, R.; Trepat, X. Mechanical guidance of cell migration: lessons from chemotaxis. Current opinion in cell biology 2013, 25(5), 543–549. [Google Scholar] [CrossRef]
- Jayadev, R.; Sherwood, D. R. Basement membranes. Current Biology 2017, 27(6), R207–R211. [Google Scholar] [CrossRef]
- Hwang, I.; Robinson, D. G. Transport vesicle formation in plant cells. Current opinion in plant biology 2009, 12(6), 660–669. [Google Scholar] [CrossRef]
- Chen, Y. A.; Scheller, R. H. SNARE-mediated membrane fusion. Nature reviews Molecular cell biology 2001, 2(2), 98–106. [Google Scholar] [CrossRef]
- Söllner, T.; Whiteheart, S. W.; Brunner, M.; Erdjument-Bromage, H.; Geromanos, S.; Tempst, P.; Rothman, J. E. SNAP receptors implicated in vesicle targeting and fusion. Nature 1993, 362(6418), 318–324. [Google Scholar] [CrossRef]
- McNew, J. A.; Parlati, F.; Fukuda, R.; Johnston, R. J.; Paz, K.; Paumet, F.; Rothman, J. E. Compartmental specificity of cellular membrane fusion encoded in SNARE proteins. Nature 2000, 407(6801), 153–159. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Ahmed, M.; Melia, T. J.; Sollner, T. H.; Mayer, T.; Rothman, J. E. Fusion of cells by flipped SNAREs. Science 2003, 300(5626), 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Hong, W. SNAREs and traffic. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 2005, 1744(2), 120–144. [Google Scholar] [CrossRef] [PubMed]
- Jahn, R.; Scheller, R. H. SNAREs—engines for membrane fusion. Nature reviews Molecular cell biology 2006, 7(9), 631–643. [Google Scholar] [CrossRef]
- Bock, J. B.; Matern, H. T.; Peden, A. A.; Scheller, R. H. A genomic perspective on membrane compartment organization. Nature 2001, 409(6822), 839–841. [Google Scholar] [CrossRef]
- Hanson, P. I.; Roth, R.; Morisaki, H.; Jahn, R.; Heuser, J. E. Structure and conformational changes in NSF and its membrane receptor complexes visualized by quick-freeze/deep-etch electron microscopy. Cell 1997, 90(3), 523–535. [Google Scholar] [CrossRef]
- Horne-Badovinac, S.; Bilder, D. Mass transit: epithelial morphogenesis in the Drosophila egg chamber. Developmental dynamics: an official publication of the American Association of Anatomists 2005, 232(3), 559–574. [Google Scholar] [CrossRef]
- Denef, N.; Chen, Y.; Weeks, S. D.; Barcelo, G.; Schüpbach, T. Crag regulates epithelial architecture and polarized deposition of basement membrane proteins in Drosophila. Developmental cell 2008, 14(3), 354–364. [Google Scholar] [CrossRef]
- Ashburner, M. A laboratory manual; Cold Spring Harbor Laboratory Press: New York, NY, 1989. [Google Scholar]
- Lerner, D. W.; McCoy, D.; Isabella, A. J.; Mahowald, A. P.; Gerlach, G. F.; Chaudhry, T. A.; Horne-Badovinac, S. A Rab10-dependent mechanism for polarized basement membrane secretion during organ morphogenesis. Developmental cell 2013, 24(2), 159–168. [Google Scholar] [CrossRef]
- Devergne, O.; Sun, G. H.; Schüpbach, T. Stratum, a homolog of the human GEF Mss4, partnered with Rab8, controls the basal restriction of basement membrane proteins in epithelial cells. Cell reports 2017, 18(8), 1831–1839. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of epithelial apical-basal polarity and basement membrane assembly. A polarized epithelial cell is shown with distinct apical, lateral, and basal domains. Perlecan (Pcan), Collagen IV (Coll IV), and Laminin, are basement membrane (BM) components. The spatial organization of BM contributes to the establishment and maintenance of characteristic apical–basal polarity of epithelial cells.
Figure 1.
Schematic diagram of epithelial apical-basal polarity and basement membrane assembly. A polarized epithelial cell is shown with distinct apical, lateral, and basal domains. Perlecan (Pcan), Collagen IV (Coll IV), and Laminin, are basement membrane (BM) components. The spatial organization of BM contributes to the establishment and maintenance of characteristic apical–basal polarity of epithelial cells.

Figure 3.
Conceptual model for SNARE mediated regulation of intracellular trafficking, secretion, and polarized deposition of basement membrane (BM) proteins. BM proteins are synthesized in the endoplasmic reticulum (ER) and transported via vesicular trafficking to the Golgi apparatus, where they are packaged into secretory vesicles destined for the basal surface of epithelial cells. SNARE mediated membrane fusion contributes to multiple steps of vesicle dynamics, including vesicle biogenesis, maturation, trafficking, and fusion with the plasma membrane. The Crag/Rab10 pathway restricts BM containing vesicles transport to the apical domain. Individual loss of specific SNAREs produces three phenotypic classes: (I) intracellular accumulation of BM proteins due to defects in vesicle biogenesis, maturation, trafficking, and/or fusion with the plasma membrane; (II) apical BM deposition resulting from defects in fusion of vesicles with the basal plasma membrane, vesicle misdirection and vesicle fusion with the apical plasma membrane mediated by alternative SNARE paralogs; and (III) a mixed phenotype reflecting SNARE function at multiple stages of the trafficking pathway.
Figure 3.
Conceptual model for SNARE mediated regulation of intracellular trafficking, secretion, and polarized deposition of basement membrane (BM) proteins. BM proteins are synthesized in the endoplasmic reticulum (ER) and transported via vesicular trafficking to the Golgi apparatus, where they are packaged into secretory vesicles destined for the basal surface of epithelial cells. SNARE mediated membrane fusion contributes to multiple steps of vesicle dynamics, including vesicle biogenesis, maturation, trafficking, and fusion with the plasma membrane. The Crag/Rab10 pathway restricts BM containing vesicles transport to the apical domain. Individual loss of specific SNAREs produces three phenotypic classes: (I) intracellular accumulation of BM proteins due to defects in vesicle biogenesis, maturation, trafficking, and/or fusion with the plasma membrane; (II) apical BM deposition resulting from defects in fusion of vesicles with the basal plasma membrane, vesicle misdirection and vesicle fusion with the apical plasma membrane mediated by alternative SNARE paralogs; and (III) a mixed phenotype reflecting SNARE function at multiple stages of the trafficking pathway.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



