Submitted:
28 February 2026
Posted:
04 March 2026
You are already at the latest version
Abstract
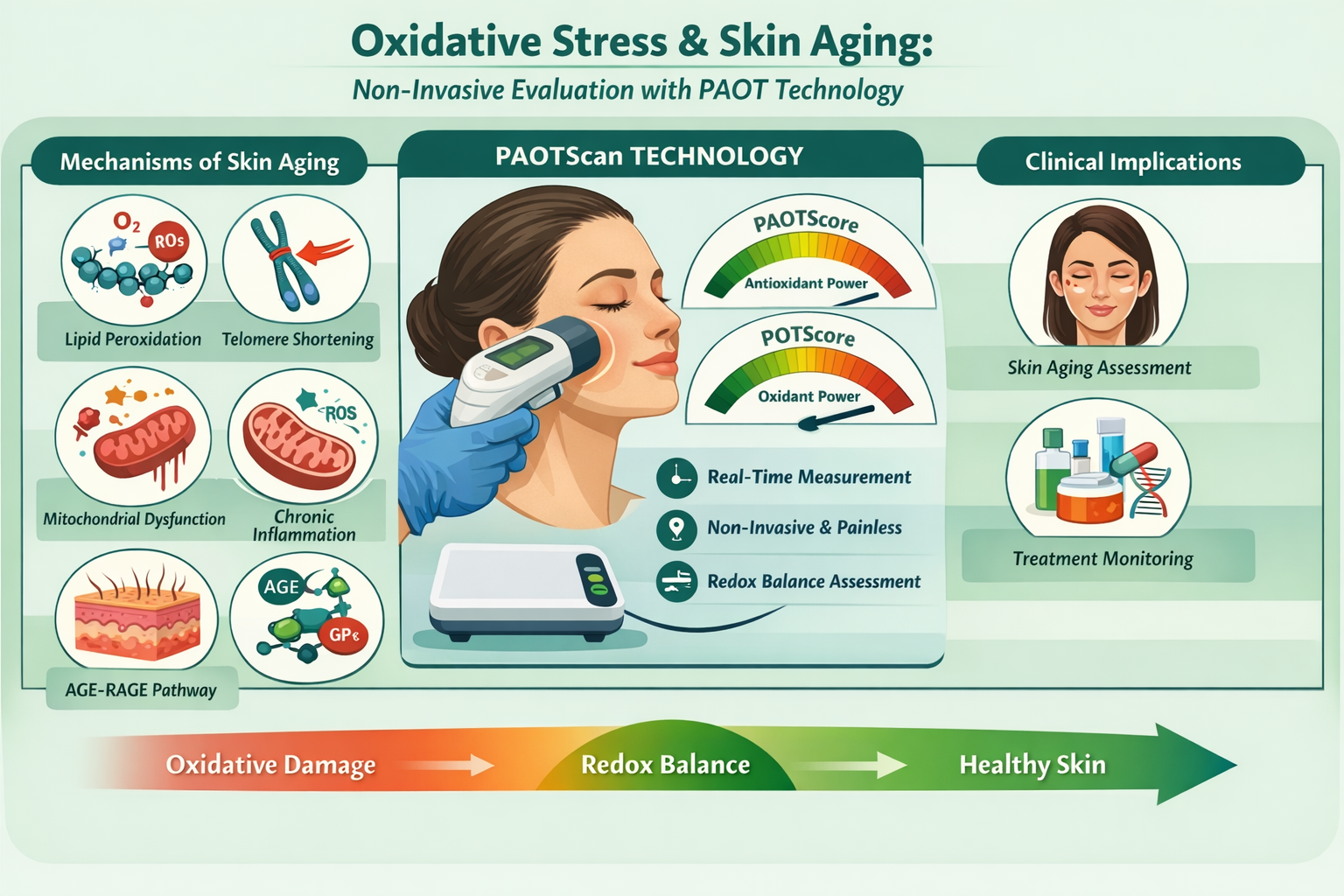
Oxidative stress accelerates skin aging by causing damage to DNA, lipids and proteins in skin cells. Exacerbated by environmental factors such as UV rays, pollution and smoking, it leads to loss of elasticity, the appearance of wrinkles, cutaneous dryness, pigmentation spots and a general weakening of the skin's natural defenses. These changes, accumulated over time, are responsible for premature or accelerated aging. Against this backdrop, PAOTScan is an innovative, non-invasive technology for assessing the oxidative state of the skin in real time. The device is based on a patch equipped with microelectrodes and an electrochemical gel. It provides two essential scores: the PAOTScore, which measures the skin's Total Antioxidant Power to neutralize free radicals, and the POTScore (Total Oxidant Power), which quantifies the level of oxidants. These data provide a detailed analysis of the skin's redox balance, enabling early detection of imbalances responsible for premature aging. The PAOTScan not only measures the speed of skin ageing, but also enables prevention and treatment strategies to be adapted. It is therefore an invaluable tool in dermatology and dermato-cosmetics, making it easier to monitor the effectiveness of anti-aging interventions and helping to preserve the skin's health and youthfulness.
Keywords:
oxidative stress
; antioxidants
; oxidants
; cellular damage
; accelerated aging
; skin pathologies
1. Introduction
Oxidative stress is defined as an imbalance between the production of reactive oxygen species (ROS) and the ability of antioxidant systems to neutralize them. ROS include free radicals and other reactive oxygen derivatives, which are produced naturally during cellular metabolism or in response to external stimuli such as exposure to ultraviolet radiation, pollution or tobacco. When ROS production exceeds antioxidant capacity, this leads to molecular alterations affecting DNA, lipids and proteins. This oxidative damage impairs cellular and tissue functionality, contributing to premature aging and the onset of various pathologies [1,2].
Oxidative stress is implicated in many age-related diseases, such as cardiovascular disease, neurodegenerative pathologies (such as Alzheimer's and Parkinson's disease) and certain cancers[3]. In addition, it accelerates the process of cellular senescence by shortening telomeres, the chromosomal structures essential for protecting DNA during cell division. This deterioration of telomeres, aggravated by oxidative aggression, promotes cell cycle arrest and the progressive loss of tissue regeneration capacity [4,5].
Given these complex mechanisms, accurate and regular assessment of oxidative status is crucial. This not only makes it possible to monitor oxidative imbalances, but also to identify preventive strategies likely to slow down the effects of biological aging. In this context, PAOTScan technology represents a major innovation. It relies on non-invasive electrochemical measurement to analyze the oxidative state of the skin in real time. By quantifying imbalances between oxidants (POTScore) and antioxidants (PAOTScore)[6], this tool provides an estimate of the speed of biological aging, based on complementary parameters such as hydration levels and skin pH. The aim of this review is to present the underlying mechanisms of oxidative stress in skin aging, highlighting the impact of free radicals on the skin's cellular structures. It also explores the role of antioxidant biomarkers in protecting against this damage, as well as the contribution of innovative technologies, such as PAOTScan, in measuring the speed of aging in real time and identifying the imbalances responsible for premature aging. Finally, the review highlights the importance of these assessments in adapting dermatological care and lifestyle habits to preserve the skin's youthfulness and health
2. Biological Aging and Oxidative Stress
Radical Theory of Aging
The radical theory of aging, formulated by Denham Harman in 1956, proposes that reactive oxygen species (ROS), such as free radicals, are responsible for a progressive accumulation of damage at the cellular level, thus contributing to aging. According to this theory, ROS are generated during normal metabolic processes, notably mitochondrial respiration, and can interact with biological macromolecules, leading to irreversible modifications [7].
- Cellular alterations linked to aging
- Damage to DNA and telomere function
Oxidative stress induces significant damage to telomeres, directly affecting their structure and integrity. Telomeres, made up of guanine-rich sequences, are particularly sensitive to attack by reactive oxygen species (ROS), notably through the formation of oxidative lesions such as 8-oxoguanine [8,9]. This damage impairs telomeric DNA protection and repair mechanisms, exacerbating telomere length loss over cell divisions [8].
Oxidative stress also compromises the activity of telomerase, the enzyme responsible for telomere maintenance. The incorporation of oxidized bases, such as oxodGTP, blocks telomere elongation, preventing proper regeneration [8]. This dysfunction accelerates telomere shortening, leading to premature cellular ageing, particularly in skin tissue where rapidly renewing cells such as keratinocytes and fibroblasts are affected [9]. As a result, oxidative stress contributes significantly to skin ageing through integrated mechanisms affecting cell regeneration capacity as well as genetic stability.
- Lipid peroxidation
Oxidative stress induces lipid peroxidation, a key process in skin ageing. Under the effect of free radicals such as hydroxyls and hydroperoxyls, polyunsaturated lipids in cell membranes are attacked, generating lipid radicals that propagate a chain reaction of degradation [10,11]. This peroxidation produces reactive compounds such as isoprostanes, malondialdehyde (MDA) and 4-hydroxynonenal (4-HNE), which can covalently bind to DNA, proteins and phospholipids, disrupting essential cellular processes [12,13].
These alterations compromise the integrity of cell membranes, reducing their fluidity and protective function, thus weakening the skin's natural defenses against environmental aggressors such as UV rays [10,12]. Furthermore, the chronic accumulation of this oxidative damage leads to persistent inflammation, accelerating cellular senescence and reducing the regenerative capacity of skin tissue [11,13].
In response to these stresses, compensatory mechanisms such as the activation of transcription factors (NF-κB, Nrf2) are activated, but their repair capacity is often exceeded under conditions of chronic stress [12]. Thus, lipid peroxidation contributes directly to premature skin aging, underscoring the importance of preventive approaches aimed at neutralizing
- The AGE-RAGE axis and aging
Advanced glycation end-products (AGEs), resulting from the non-enzymatic reaction between reducing sugars and proteins, lipids or nucleic acids, accumulate over time, a phenomenon amplified by oxidative stress [14,15]. These AGEs interact with their specific receptor, RAGE (Receptor for Advanced Glycation End-products), activating a cascade of pro-inflammatory and pro-oxidative reactions in tissues [2,16]. This activation leads to increased production of ROS (reactive oxygen species) via the NADPH oxidase enzyme, promoting a vicious circle of oxidative damage [15].
At cutaneous level, EFAs alter the structure of the extracellular matrix (ECM), notably by modifying collagen and elastin, reducing tissue suppleness and integrity [15,17]. These modifications compromise cellular repair mechanisms and accelerate fibroblast senescence, limiting the skin's regenerative capacity [2]. In addition, AGE-RAGE interaction activates signaling pathways such as NF-κB and MAPK, amplifying chronic inflammation and disrupting the natural antioxidant response [16,17].
The progression of these mechanisms is associated with premature aging, manifested by loss of elasticity, the appearance of wrinkles and a skin more vulnerable to environmental aggressors [2,15]. The management of oxidative stress and the reduction of EFAs, through
- Mitochondrial dysfunction and aging
Cellular aging is closely linked to mitochondrial dysfunction, mainly due to increased oxidative stress over time. Mitochondria, as the main sources of ROS production via the respiratory chain, are particularly vulnerable to these reactive oxygen species [18,19]. Electron leakage at complexes I and III generates superoxide (O₂--), which can be transformed into other radicals, such as hydrogen peroxide and the hydroxyl radical, causing damage to mitochondrial components [19,20].
This damage particularly affects mitochondrial DNA (mtDNA), which, due to its proximity to ROS production sites and limited repair mechanisms, undergoes frequent mutations and deletions [19,21]. These alterations compromise the performance of OXPHOS complexes, reducing the efficiency of ATP production and increasing electron leakage, thus intensifying oxidative stress in an amplification loop [18]. This energetic deterioration affects cells with high metabolic demand, notably skin and muscle cells, limiting their regenerative capacity [19,20].
Lipid peroxidation of mitochondrial inner membranes, initiated by ROS, generates toxic compounds such as 4-hydroxynonenal (4-HNE), which impair the fusion and fission processes required to eliminate dysfunctional mitochondria by mitophagy [18,19]. These dysfunctions also disrupt cell renewal mechanisms by activating pro-apoptotic pathways, notably through the release of cytochrome c, triggering programmed cell death [18].
These processes contribute to the accumulation of senescent cells in tissues, reducing their ability to repair and renew themselves. In skin tissue, this translates into a loss of elasticity, reduced hydration and an altered protective barrier, favoring the visible signs of aging [18]. Interventions aimed at regulating ROS production, such as enhancing mitochondrial antioxidant systems or modulating mitophagy processes, could represent promising strategies for slowing aging and preserving tissue regeneration [19,20].
- Enzyme deregulation
Superoxide Dismutase (SOD) is an essential antioxidant enzyme, playing a key role in cellular defense against oxidative stress. It catalyzes the dismutation of superoxide anion (O2.-) into hydrogen peroxide (H2O2) and oxygen (O2) [22,23,24]. Under conditions of intense oxidative stress, such as excessive exposure to UV rays or pollution, or during chronic inflammatory processes, the production of free radicals exceeds the neutralizing capacity of SOD [25]. This oxidative overload leads to inhibition of SOD activity by several mechanisms. For example, excess H2O2 can promote oxidative modifications of SOD, thus reducing its enzymatic efficiency. Free radicals can also oxidize specific residues within the SOD structure, notably amino acids such as tyrosine, thus impairing its function [22]. This deregulation of SOD leads to a vicious circle in which reduced enzyme activity results in an accumulation of superoxide anion, intensifying oxidative stress [26]. This causes direct damage to membrane lipids, notably through their peroxidation, to structural proteins (collagen, elastin) and to the DNA of skin cells [27]. Clinically, this translates into a loss of elasticity, the appearance of deep wrinkles, skin slackening and increased sensitivity to environmental aggressions, accentuating the skin's vulnerability to damage.
Catalase (CAT) is also an antioxidant enzyme that catalyzes the breakdown of hydrogen peroxide into water and oxygen, limiting the accumulation of these potentially toxic ROS [28,29,30]. Located mainly in peroxisomes, catalase plays an essential role in the detoxification of ROS produced during metabolic processes such as the β-oxidation of fatty acids [29]. This function maintains a healthy cellular redox balance and prevents oxidative damage to cellular components, including membrane lipids, proteins and DNA [29]. However, under conditions of intense oxidative stress, catalase activity can be significantly impaired. For example, oxidative stress induced by environmental contaminants such as bisphenol A (BPA) has been shown to dramatically reduce catalase activity in human liver cells, compromising H2O2 detoxification capacity and amplifying oxidative stress [31]. In addition, during cellular aging, the import capacity of catalase into peroxisomes is often reduced, leading to accumulation of H2O2 in the cytosol and exacerbating oxidative stress-related cellular damage[32]. This deregulation of catalase creates a vicious circle in which the accumulation of H2O2 promotes the formation of more reactive free radicals, such as the hydroxyl radical (OH-) via the Fenton reaction, leading to widespread oxidative damage to membrane lipids, structural proteins and cellular DNA [28]. In the context of skin aging, this altered catalase activity contributes to increased lipid peroxidation, collagen and elastin degradation, and increased skin sensitivity to external aggressors [32,33]. As in the case of SOD, this will manifest clinically as loss of elasticity, the appearance of deep wrinkles and sagging skin, accelerating the visible signs of cutaneous aging.
Glutathione Peroxidase (GPx) is an essential antioxidant enzyme that plays a key role in protecting cells against oxidative stress. It is part of a family of proteins including both selenoproteins (GPx1 to GPx4, as well as GPx6) and cysteine-based proteins (GPx5, GPx7, GPx8) [34,35,36]. Its main purpose is to neutralize the damaging effects of free radicals by converting hydrogen peroxide (H2O2) and various organic hydroperoxides into less toxic compounds, such as water and alcohols, using reduced glutathione (GSH) as a cofactor [37]. Among the members of this family, GPx4 stands out for its unique ability to target complex lipid hydroperoxides, thereby protecting cell membranes against lipid peroxidation and preventing ferroptosis, a form of cell death regulated by lipid oxidation [35,38]. Excessive ROS accumulation can lead to oxidation of cysteine or selenocysteine residues in the GPx structure, reducing its enzymatic efficiency and disturbing the cellular redox balance [37]. This deregulation leads to a vicious circle in which reduced GPx activity promotes the accumulation of lipid peroxides, intensifying oxidative stress and causing cellular damage, notably to membrane lipids, structural proteins such as collagen and elastin, and DNA [39].
- Oxidative stress promotes inflammation
Oxidative stress promotes chronic inflammation by inducing the production of pro-inflammatory cytokines such as interleukin-6 (IL6), interleukin 1-b (IL1b) and tumor necrosis factor alpha (TNFa) [40,41,42]. This increase in pro-inflammatory cytokines creates a vicious circle in which inflammation induces increased ROS formation, promoting oxidative stress [41,43]. For example, exposure to microplastics induces splenic inflammation in Japanese quail via activation of the p38 MAPK and TNF pathways, leading to a significant increase in ROS and a decrease in the activity of antioxidant enzymes such as superoxide dismutase (SOD) and catalase (CAT) [43]. Similarly, heme can induce programmed macrophage necrosis by promoting autocrine TNF production and ROS generation, illustrating the complex interplay between oxidative stress and inflammation [44]. This ROS-fed inflammatory loop leads to significant tissue damage. Prolonged exposure to ultraviolet (UV) radiation has been shown to generate significant stress, promoting the release of pro-inflammatory cytokines such as IL6 and IL1b, and reducing the expression of antioxidant enzymes such as MnSOD [45]. In internal tissues, ROS accumulation can disrupt the endothelial barrier and increase vascular permeability, initiating irreversible cell damage [40]. This aggravates chronic inflammatory diseases and contributes to the acceleration of skin ageing, characterized by thickening of the epidermis and degradation of the collagen network.
- Dysbiosis and inflammation favor ROS generation
An imbalance in the gut microbiota, or dysbiosis, greatly favors increased oxidative stress and damage to the skin barrier. Dysbiosis leads to disruption of intestinal permeability, often referred to as "leaky gut", allowing the translocation of bacterial components such as lipopolysaccharides (LPS) into the systemic circulation [46,47]. These pro-inflammatory molecules activate the immune system, leading to increased production of pro-inflammatory cytokines such as IL6 and TNFa, promoting a pro-oxidant environment and contributing to a chronic inflammatory state known as "inflammaging" [47,48]. This systemic inflammation directly alters skin barrier function via the gut-skin axis. Indeed, intestinal dysbiosis influences keratinocyte differentiation and reduces the production of ceramides, lipids essential for maintaining skin barrier integrity [49,50]. Studies have shown that the intestinal microbiota can disrupt skin cell cohesion and moisture retention, notably by reducing levels of skin barrier proteins such as loricrin, keratin 10 and desmoglein 1 [49]. In addition, some pathogenic bacteria produce metabolites such as phenol and p-cresol, which can penetrate the skin, disrupt epidermal differentiation and reduce skin hydration [51]. When the cutaneous barrier is impaired, the skin becomes more permeable to external agents, accentuating local oxidative stress and skin damage. This contributes to premature skin ageing, characterized by dryness, loss of elasticity, the appearance of wrinkles and increased vulnerability to external aggressors [50].

Table 1.
Comparison of different oxidative stress markers: biological effects and age-related variations.
Table 1.
Comparison of different oxidative stress markers: biological effects and age-related variations.
| Biomarkers | Impact | Tendance |
| 8-oxodG | DNA damages | Increase |
| MDA (lipid peroxidation) | Oxidative stress markers | Increase |
| Telomere length | Cellular senescence indicator | Decrease |
| PAOTScore | Total Antioxidant Power | Decrease |
| SOD | Antioxidant enzyme, ROS detoxification | Decrease |
| Catalase | Conversion of H2O2 into H2O and O2 | Decrease |
| GPx | Peroxide reduction, cell protection | Decrease |
| Pro-inflammatory cytokines (IL6, TNFa, etc.) | Pro-inflammatory cytokines | Increase |
| Bacteria of the intestinal microbiota | Modulation of inflammation and oxidative stress | Dysbiosis |
- Pro-oxidant effects of vitamins.
Although antioxidants are generally recognized for their protective role against oxidative stress, several studies have shown that they can also exhibit pro-oxidant behavior, particularly at high concentrations, in the presence of transition metals, or depending on their chemical structure. This paradox is well illustrated by flavonoids which, despite their ability to scavenge reactive oxygen species (ROS), can generate free radicals when oxidized into reactive quinones. These quinones are capable of damaging DNA or initiating the production of superoxide through the reduction of metals such as Cu²⁺ or Fe³⁺ [52]. A similar phenomenon has been observed with vitamin C: although it is a powerful antioxidant, it can reduce Fe³⁺ or Cu²⁺ to Fe²⁺ or Cu⁺, thereby facilitating the Fenton reaction and the formation of highly reactive hydroxyl radicals [53,54]. These effects are generally dose-dependent: at low concentrations, antioxidants exert a protective effect, whereas at high doses or in pro-oxidant environments (e.g., high pH or excess metals), they promote oxidation.
This dual behavior is now being exploited in certain anticancer strategies, where the pro-oxidant effect of polyphenols is used to induce apoptosis in tumor cells through a targeted increase in ROS levels [55]. It has also been demonstrated that red wine polyphenols, although antioxidant in vitro, slightly increase ROS levels in erythrocytes, thereby activating an adaptive response of endogenous antioxidant systems such as glutathione and enzymes of the pentose phosphate pathway [56].
In a dermatological context, these ambivalent properties highlight the importance of proper formulation and controlled dosage. When applied topically, certain antioxidants may act as pro-oxidants under the influence of UV radiation, skin pH, or traces of metals present in sweat or environmental pollution. This can lead to local ROS production, promoting premature skin aging or inflammatory reactions. Conversely, at low concentrations and under controlled conditions, these compounds may stimulate endogenous skin defenses and enhance the resilience of the epidermal barrier against oxidative stress.
- PAOTScan to measure oxidative stress
PAOTScan is an innovative, patented technology for rapid, non-invasive, real-time measurement of oxidative stress. The technique is based on the application of a patch containing microelectrodes to the skin, combined with a gel enriched with an electrochemical mediator (Figure 1). This device detects electrochemical variations due to reactions between oxidants and antioxidants present on the skin's surface.
Figure 1.
Enzymatic pathways of oxidative stress detoxification: role of superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx) (Yu et al. 2010 [24]).
Figure 1.
Enzymatic pathways of oxidative stress detoxification: role of superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx) (Yu et al. 2010 [24]).

The PAOTScan generates two key scores for assessing oxidative balance: the PAOTscore and the POTscore. In addition, the tool can identify other biomarkers, including vitamin C, vitamin E and glutathione. It also takes into account clinical parameters such as skin hydration and associated pH, as well as the speed of cellular aging.
- The PAOTscore
Total Antioxidant Power reflects the skin's antioxidant capacity, i.e. its ability to neutralize ROS. A high score indicates a high level of antioxidants such as vitamin C, vitamin E and glutathione, which play a key role in protecting against cellular damage.
Antioxidants reduce the impact of free radicals on cells by limiting the oxidation of lipids, proteins and DNA. Consequently, a high PAOTScore is synonymous with good antioxidant defense and reduced susceptibility to oxidative stress-related pathologies
The PAOT Score is calculated as follows:
 Where EP0 is the electrochemical potential at time 0, EPmax is the maximum electrochemical potential, EP10 is the electrochemical potential at time 10 mins [6].
Where EP0 is the electrochemical potential at time 0, EPmax is the maximum electrochemical potential, EP10 is the electrochemical potential at time 10 mins [6].
- The POTscore
Total Oxidant Power quantifies the level of oxidants present on the skin. These oxidants, mainly free radicals, are capable of damaging cells if not neutralized by antioxidants. An elevated POTScore reveals an oxidative imbalance, indicating an increased risk of oxidative stress and, by extension, cell damage.
The POT score is calculated as follows:

Where EP0 is the electrochemical potential at time 0, EPmax is the maximum electrochemical potential, EP10 is the electrochemical potential at time 10 mins.
The protocol followed has already been described by Pincemail.J et al. (2019).
- Markers observed by PAOTScan
Vitamin C (ascorbic acid) plays an essential role in dermato-cosmetology, notably for its anti-aging and regenerative properties on the skin. It is a powerful antioxidant capable of neutralizing free radicals, thus limiting oxidative damage caused by UV rays and other environmental aggressions [57,58]. By stimulating collagen synthesis, it improves skin elasticity and firmness, while reducing the appearance of wrinkles [57,59,60].
Its role in cosmetology is not limited to collagen production; it also acts as a cofactor in collagen hydroxylation, a key step in the maturation of collagen fibers. This strengthens the structure of the dermis and helps prevent skin slackening [61]. In addition, vitamin C helps to lighten pigmentation spots by inhibiting tyrosinase, a key enzyme in melanogenesis, making it effective in the treatment of hyperpigmentation [61]. Regular topical application of vitamin C, in stabilized form, significantly increases collagen levels and improves overall skin texture, reducing the visible signs of aging [62].
Vitamin E, mainly in the form of alpha-tocopherol, is a powerful fat-soluble antioxidant that protects cell membranes against lipid peroxidation. This mechanism prevents damage to lipids, proteins and DNA caused by free radicals, particularly in fat-rich tissues such as skin [63,64]. Vitamin E is able to neutralize free radicals generated by various environmental factors, such as UV rays. By reducing oxidative stress, it prevents the signs of skin ageing, including the premature appearance of wrinkles and loss of elasticity [63]. By stabilizing cell membranes, vitamin E promotes the repair of damaged tissue and enhances skin regeneration. Its anti-inflammatory action, by inhibiting the NF-κB pathway, reduces the production of pro-inflammatory mediators such as prostaglandins and cytokines, which is beneficial in limiting redness, irritation and skin inflammation [63]. Vitamin E works in synergy with vitamin C to reinforce antioxidant defense. Once oxidized, vitamin E is regenerated by vitamin C, enabling it to maintain its effectiveness in the fight against free radicals [64].
Glutathione (GSH) is a major intracellular antioxidant that protects cells against oxidative stress by neutralizing reactive oxygen species (ROS). It plays a part in antioxidant defense mechanisms by maintaining cellular redox balance. Its role is crucial in preventing oxidative damage to lipids, proteins and DNA [65]. In UV-induced skin damage, glutathione acts synergistically with the enzyme GPX4 (glutathione peroxidase 4) to reduce lipid peroxides. This protects keratinocytes against ferroptotic necrosis, a form of cell death linked to the accumulation of iron and peroxidized lipids. This mechanism is essential for preserving skin integrity under the effects of sun exposure [66,67]. Certain substances, such as resveratrol or NMN (nicotinamide mononucleotide), can boost glutathione production by activating the Nrf2 pathway. This improves the skin's endogenous antioxidant defences and reduces the accumulation of toxic products resulting from lipid peroxidation [66,67].
The pH of the skin is generally between 4 and 6, corresponding to a slightly acidic environment known as “acid mantle”. This acidity comes mainly from the fatty acids in sebum and the organic acids in sweat [68,69]. The hydrolipidic film thus formed plays a key role in protection against external aggression. Skin pH regulates the activity of several essential enzymes, notably those involved in the desquamation process and lipid renewal. An acid pH maintains the balance between proteases and their inhibitors, ensuring the stability of the skin barrier [68]. Skin acidity inhibits the growth of many pathogenic micro-organisms, while promoting a healthy microbiota. This antimicrobial function is essential for preventing skin infections such as atopic dermatitis [69]. As pH becomes alkaline, the risk of bacterial and fungal proliferation increases, compromising skin integrity [70].
With age, the skin's pH tends to rise, leading to a reduced capacity for cell regeneration and impaired production of protective lipids [68]. This change favors the appearance of signs of premature aging, such as dryness, loss of elasticity and wrinkles [70]. Skin acidity plays an essential role in maintaining skin barrier homeostasis [71].
The stratum corneum is the outermost layer of the epidermis and plays a crucial role in regulating hydration. It contains corneocytes surrounded by a lipid matrix, which form a barrier to excessive water loss (TEWL) [72]. Hydration directly influences the flexibility and cohesion of stratum corneum cells, as well as the enzymatic activities essential for desquamation [73]. Significant variations in hydration can be observed with age, where a decrease in sebum production and water retention capacity is noted [74]. Dehydrated skin becomes stiffer and less elastic, increasing susceptibility to cracking and infection [74]. Chronic moisture loss can lead to skin pathologies such as eczema and atopic dermatitis [69,73]. As we age, the skin's ability to maintain its hydration diminishes, accentuating signs of ageing such as wrinkles and loss of elasticity. Maintaining a good level of hydration is essential to slow down these processes and preserve the skin's appearance and health [73].
PAOTscan can accurately detect and quantify various oxidative stress parameters, and compare them with a reference interval. These values can be used to interpret the results obtained with the PAOTscan, situating the patient's condition in relation to normal physiological values [6].
Table 2.
Highlighting reference ranges of oxidative stress markers (Pincemail.J et al. 2019).
| Parameter | Human Reference interval |
| Antioxydants | |
| vitamin C (µg/mL) | 6.0 - 15 |
| vitamin E as a-tocopherol (µg/mL) | 8.6 - 19.2 |
| γ-tocopherol (µg/mL) | 0.39 - 2.42 |
| β-carotene (µg/mL) | 0.06 - 0.68 |
| thiol proteins (µM) | 314 - 516 |
| reduced glutathione (µM) | 717 - 1110 |
| oxidized glutathione (µM) | 0.96 - 10 |
| glutathione peroxidase (UI/g Hb) | 20 - 56 |
| Uric acid | >70 |
| Trace elements | |
| copper (mg/mL) | 0.70 - 1.1 |
| zinc (mg/mL) | 0.70 - 1.20 |
| selenium (µg/mL) | 73 - 110 |
| Biomarkers of lipid peroxidation | |
| lipid peroxides (µM) | 0 - 432 |
| oxidized LDL (ng/mL) | 28 - 70 |
| antibobies against oxidized LDL | 200 - 600 |
| Sources of ROS production | |
| copper/zinc ratio | 1 - 1.17 |
| myeloperoxidase | 27 - 72 |
| PAOT | |
| plasma | 1.42 - 36.78 |
| skin | 7.86 - 62.91 |
| saliva | 1.52 - 14.14 |
| urine | 42.85 - 104.63 |
| creatinine standardized urine | 6.23 - 121.96 |
- The aging speed
PAOTScan measures the speed of skin aging based on a set of key markers, including pH, hydration and skin antioxidant levels (Jones et al., 2019). These indicators provide a detailed assessment of the state of the skin barrier and its resistance to external aggressors. The biological age of the skin, a reflection of an individual's overall physiological state, results from the interaction between genetic, environmental and behavioral factors [75].
Studies show that high concentrations of antioxidants in the skin are correlated with better resistance to ageing, while low levels accentuate skin vulnerability, favoring the appearance of age-related signs [76]. Over time, antioxidant levels decrease progressively, weakening the skin's regenerative capacity and increasing its sensitivity to free radical-induced damage (Packer et al., 1995). Conversely, the POT score increases with age, reflecting an accumulation of uncompensated oxidative damage and a progressive alteration of the skin's defense mechanisms. These factors contribute to premature skin ageing and the appearance of clinical signs such as wrinkles, loss of elasticity and pigmentation spots [77].
The PAOTScan assessment not only determines the skin's antioxidant resources, but also anticipates its predisposition to premature ageing (Figure 2). The system is based on a comparison of the results obtained with adapted reference standards [77], and a marked drop in PAOTScore indicates an acceleration in the skin's ageing process.
Figure 2.
Photograph of skin oxidative stress analysis using PAOTScan technology. More details on the protocol and results of this technology have been described by Pincemail J. et al.(2019).
Figure 2.
Photograph of skin oxidative stress analysis using PAOTScan technology. More details on the protocol and results of this technology have been described by Pincemail J. et al.(2019).

Figure 3.
Kinetic curve of electrochemical potential changes during the reaction of antioxidants and oxidants with oxidized and reduced forms of iron, at the skin surface and using PAOTScan (Pincemail.J et al. 2019).
Figure 3.
Kinetic curve of electrochemical potential changes during the reaction of antioxidants and oxidants with oxidized and reduced forms of iron, at the skin surface and using PAOTScan (Pincemail.J et al. 2019).

Figure 4.
correlation between PAOT score and skin ageing by age group.

Vitamins C, E and glutathione (GSH) play an essential role as antioxidants, contributing to cellular protection against damage caused by oxidative stress (Farris, 2014). In order to quantify their influence, a corrective coefficient is applied according to their concentration within the skin. The equation used to adjust this influence is written as follows:
- -
- VitCmeasured, VitEmeasured and GSHmeasured represent the measured concentrations of vitamins C, E and glutathione in the skin.
- -
- VitCopt, VitEopt and GSHopt correspond to the optimal levels expected for these molecules.
The evolution of oxidative stress over time can be used to assess the rate of skin aging (Berneburg et al., 2000). This parameter is calculated by measuring the evolution of total antioxidant power (PAOTScore) over a given period. The corresponding equation is as follows:

- -
- PAOTT and PAOT0H represent the values of total antioxidant power measured respectively after a certain time T and at the initial time.
- -
- T is the duration of the measurement.
- -
- Antiox factor is the previously defined coefficient, adjusting for the impact of vitamins and glutathione on antioxidant protection.

- -
- -Phototypefacteur / TypePeaufacteur introduces a correction taking into account the subject's skin type and phototype, in order to refine the assessment of skin aging (Krtmann et al., 2017).
- -
- Accuracy of skin ageing assessment
The inclusion of additional parameters, such as phototype and hydration level, enhances the accuracy of the estimate of the skin's biological age [78]. This integration makes it possible to identify individuals at increased risk of premature aging, and to adapt preventive or corrective strategies accordingly.
Finally, this algorithmic approach offers a robust and objective method for assessing the rate of skin aging. It is particularly useful for dermatological diagnosis and for monitoring the efficacy of anti-aging treatments, paving the way for personalized interventions based on reliable quantitative data [78].
- Skin parameters influencing results
When assessing cutaneous oxidative stress using methods such as PAOT® measurement, a number of skin parameters need to be taken into account to ensure the reliability and accurate interpretation of the results obtained. Indeed, factors such as skin pigmentation, texture, elasticity and hydration can not only influence the penetration of the signals used for measurement, but also directly modulate the skin's redox state by affecting the production and neutralization of reactive oxygen species (ROS). The integration of these parameters makes it possible to improve the accuracy of PAOT measurement results and therefore to adapt dermatological recommendations and care [75,78].
- Phototype
Skin pigmentation, influenced by the distribution of eumelanin and pheomelanin, plays a key role in protection against oxidative stress. Eumelanin, predominant in darker skins, effectively absorbs UV radiation, reducing the formation of free radicals and protecting DNA from oxidative damage [79]. In contrast, pheomelanin, more abundant in lighter skins, promotes the production of reactive oxygen species (ROS) under UVA and UVB irradiation, accentuating oxidative stress [80,81]. This difference explains why individuals with eumelanin-rich skin often have higher PAOT scores, reflecting a better resistance to oxidative stress. This difference explains why individuals with eumelanin-rich skin often have higher PAOT scores, reflecting better intrinsic antioxidant capacity. These variations in photoprotection directly influence the measurement of oxidative stress, and must be taken into account when interpreting results obtained on different skin phototypes.
- Elasticity
Skin elasticity is a key parameter influencing the measurement of oxidative stress, as it reflects the integrity and functionality of the dermis. With age, collagen and elastin synthesis declines, leading to a progressive loss of the skin's mechanical properties and favoring the appearance of wrinkles and skin sagging [82]. This decrease is associated with altered fibroblast metabolism and increased collagen degradation, contributing to a more fragile skin that is less able to resist oxidative aggression [83]. Variations in elasticity must therefore be taken into account when interpreting oxidative stress scores, particularly in elderly populations or those with altered skin metabolism.
- Texture
Skin texture is a key parameter to consider when measuring oxidative stress by PAOT. Chronic exposure to UVA and UVB induces major structural alterations, notably degradation of collagen and elastin fibers, leading to loss of firmness and accentuation of skin irregularities [84]. In addition, UV rays reduce concentrations of carotenoids such as lycopene and β-carotene, which play a role in photoprotection and the neutralization of free radicals [77]. These changes potentially influence the scattering of optical signals used for PAOT measurement, due to reflectance variations by structurally altered skin. Skin marked by wrinkles and irregularities could therefore present biased values, requiring an adapted interpretation of the results obtained. So, taking skin texture into account is essential to ensure reliable, reproducible measurements of oxidative stress.
- Comparison of PAOTScan with other current techniques
PAOTScan is positioned as an innovative alternative to conventional techniques for underlining the importance of a combined strategy asuring oxidative stress, offering a rapid, non-invasive, real-time method. Unlike biochemical assays, which require biological samples to achieve high precision, PAOTScan simplifies the procedure while avoiding discomfort for the patient. Compared with Raman spectroscopy, which also shares the advantage of non-invasiveness but has limited sensitivity, PAOTScan provides a complementary approach well suited to routine clinical monitoring. However, like other surface techniques, it remains limited to the analysis of coats, which are more exposed to external aggression.
Table 3.
Highlighting reference ranges of oxidative stress markers (Pincemail.J et al. 2019).
| Method | Principle | Advantages | disadvantages |
| Raman Spectroscopy | Detection of oxidative biomarkers | Non-invasive | Limited sensitivity (Ruffien-Ciszak et al., 2008) |
| Biochemical assay | MDA and 8-oxodG assay | High precision | Requires sampling (Ayala et al., 2014) |
| PAOTScan | Real-time electrochemical analysis | Fast and non-invasive | No Specific species |
- Antioxidant protection strategies
To effectively combat cutaneous oxidative stress, several antioxidant protection strategies can be implemented. Vitamins C and E are particularly effective. Vitamin C neutralizes free radicals and stimulates collagen production [61], while vitamin E protects lipid membranes [63]. Glutathione (GSH), an endogenous antioxidant, plays a key role in regulating intracellular oxidative stress, offering additional protection to the skin [65]. Polyphenols, notably resveratrol, also contribute to this defense by activating the Nrf2 pathway, although their efficacy varies from medium to high according to studies [67]. Finally, sun protection (SPF) remains essential for blocking UV rays, and is considered essential for preventing oxidative stress-induced skin damage [85]. Each approach has its own specific strengths, underlining the importance of a combined strategy for optimal skin protection.
Table 4.
Effectiveness of antioxidant approaches in limiting cutaneous oxidative stress.
| Strategy | Strategy | Efficiency |
|---|---|---|
| Vitamin C | Free radical neutralization, collagen stimulation | High |
| Vitamin C | Protection of lipid membranes | High |
| Glutathione | Regulation of intracellular oxidative stress | High |
| Polyphenols (resveratrol) | Activation of the Nrf2 pathway | Medium to High |
| Sun protection (SPF) | UV blocking | Essential |
5. Conclusions
Skin aging is a complex process influenced by intrinsic and extrinsic factors, among which oxidative stress plays a central role. The accumulation of oxidative damage to cellular macromolecules, notably DNA, lipids and proteins, leads to progressive alteration of skin structure and function. The radical theory of aging, combined with mechanisms such as lipid peroxidation, telomere shortening and mitochondrial dysfunction, explains how oxidative stress contributes not only to premature aging, but also to accelerating the rate of skin aging.
PAOTScan technology is positioned as an innovative tool for non-invasive, real-time assessment of the oxidative state of the skin. By providing a precise analysis of the skin's redox balance via PAOT (Total Antioxidant Power) and POT (Total Oxidant Power) scores, this device makes it possible to estimate the speed at which the skin is undergoing the effects of biological aging. This dynamic approach enables early detection of oxidative imbalances and quantification of their impact on the speed of skin aging. By adapting prevention and treatment strategies to the speed of aging, we can act more effectively to slow down the process.
In addition, the use of antioxidant biomarkers such as vitamins C and E, and glutathione, strengthens the skin's ability to combat the deleterious effects of free radicals. By neutralizing these reactive oxygen species, these antioxidants help not only to reduce cellular damage, but also to stabilize the rate of skin ageing by maintaining an optimal redox balance.
In conclusion, the integration of advanced measurement methods such as PAOTScan with targeted antioxidant strategies represents a promising approach to preserving skin health and youthfulness in the face of oxidative aggression. By enabling a precise assessment of the speed of aging, this technology opens up new perspectives in preventive dermatology and cosmetology, offering personalized solutions for controlled and delayed skin aging. However, a combined approach including both precise biochemical biomarkers and adapted antioxidant strategies appears to be the most effective way of not only slowing down premature ageing, but also controlling the speed of cutaneous ageing.
Author Contributions
Conceptualization, M.K, GL,PJ, J.P,S.M participated to the paper publication. M.K, GL,PJ, J.P,S.M.; writing—original draft preparation, M.K, S.M ; writing—review
Funding
This research received no external funding
Institutional Review Board Statement
The study was conducted in accordance with the Decla-ration of Helsinki, and approved by the Ethics Committee of Comité de Protection de Personnes of CHU Liège (protocol code 177/2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Chaudhary, P.; Janmeda, P.; Docea, A.O.; Yeskaliyeva, B.; Abdull Razis, A.F.; Modu, B.; et al. Oxidative stress, free radicals and antioxidants: potential crosstalk in the pathophysiology of human diseases. Front Chem. 2023, 11, 1158198. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.P. Oxidative Stress in Health and Disease. Biomedicines 2023, 11, 2925. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; et al. Oxidative stress, aging, and diseases. Clin Interv Aging 2018, 13, 757-72. [Google Scholar] [CrossRef]
- Pole, A.; Dimri, M.; P Dimri G; Department of Biochemistry and Molecular Medicine; School of Medicine and Health Sciences; The George Washington University; Washington DC; USA. Oxidative stress, cellular senescence and ageing. AIMS Mol Sci. 2016, 3, 300-24. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.P.; Sulaiman Rahman, H. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef]
- Joël, P.; Mouna-Messaouda, K.; Jean-Paul, C.B.; Jean-Olivier, D.; Smail, M. Electrochemical Methodology for Evaluating Skin Oxidative Stress Status (SOSS). Diseases 2019, 7, 40. [Google Scholar] [CrossRef]
- Denham, H. Free-Radical Theory of Aging 1994, 717, 1-15.
- Ahmed, W.; Lingner, J. Impact of oxidative stress on telomere biology. Differentiation 2018, 99, 21-7. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195-214. [Google Scholar] [CrossRef]
- Lasch, J.; Schonfelder, U.; Walke, M.; Zellmer, S.; Beckert, D. Oxidative damage of human skin lipids Dependence of lipid peroxidation on sterol concentration.
- Villalón-García, I.; Álvarez-Córdoba, M.; Povea-Cabello, S.; Talaverón-Rey, M.; Villanueva-Paz, M.; Luzón-Hidalgo, R.; et al. Vitamin E prevents lipid peroxidation and iron accumulation in PLA2G6-Associated Neurodegeneration. Neurobiol Dis. 2022, 165, 105649. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid Med Cell Longev. 2014, 2014, 1-31. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Zavadskiy, S.P.; Astakhov, D.V.; Terentiev, A.A. Lipid peroxidation: Reactive carbonyl species, protein/DNA adducts, and signaling switches in oxidative stress and cancer. Biochem Biophys Res Commun. 2023, 687, 149167. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Q.; Zhang, D.D.; Wang, Y.N.; Tan, Y.Q.; Yu, X.Y.; Zhao, Y.Y. AGE/RAGE in diabetic kidney disease and ageing kidney. Free Radic Biol Med. 2021, 171, 260-71. [Google Scholar] [CrossRef] [PubMed]
- Senatus, L.M.; Schmidt, A.M. The AGE-RAGE Axis: Implications for Age-Associated Arterial Diseases. Front Genet. 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D.; Nicklett, E.J.; Ferrucci, L. Does Accumulation of Advanced Glycation End Products Contribute to the Aging Phenotype? J Gerontol A Biol Sci Med Sci. 2010, 65A, 963-75. [Google Scholar] [CrossRef]
- Vitorakis, N.; Piperi, C. Pivotal role of AGE-RAGE axis in brain aging with current interventions. Ageing Res Rev. 2024, 100, 102429. [Google Scholar] [CrossRef]
- Kung, H.C.; Lin, K.J.; Kung, C.T.; Lin, T.K. Oxidative Stress, Mitochondrial Dysfunction, and Neuroprotection of Polyphenols with Respect to Resveratrol in Parkinson’s Disease. [CrossRef]
- Kudryavtseva, A.V.; Krasnov, G.S.; Dmitriev, A.A.; Alekseev, B.Y.; Kardymon, O.L.; Sadritdinova, A.F.; et al. Mitochondrial dysfunction and oxidative stress in aging and cancer. Oncotarget 2016, 7, 44879-905. [Google Scholar] [CrossRef]
- Peoples, J.N.; Saraf, A.; Ghazal, N.; Pham, T.T.; Kwong, J.Q. Mitochondrial dysfunction and oxidative stress in heart disease. Exp Mol Med. 2019, 51, 1-13. [Google Scholar] [CrossRef]
- Hayakawa, M.; Hattori, K.; Sugiyama, S.; Ozawa, T. Age-associated oxygen damage and mutations in mitochondrial DNA in human hearts. Biochem Biophys Res Commun. 1992, 189, 979-85. [Google Scholar] [CrossRef]
- Bafana, A.; Dutt, S.; Kumar, A.; Kumar, S.; Ahuja, P.S. The basic and applied aspects of superoxide dismutase. J Mol Catal B Enzym. 2011, 68, 129-38. [Google Scholar] [CrossRef]
- Doran, M.L.; Knee, J.M.; Wang, N.; Rzezniczak, T.Z.; Parkes, T.L.; Li, L.; et al. Metabolomic analysis of oxidative stress: Superoxide dismutase mutation and paraquat induced stress in Drosophila melanogaster. Free Radic Biol Med. 2017, 113, 323-34. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yin, Y.; Zhu, J.; Huang, X.; Luo, Q.; Xu, J.; et al. A modulatory bifunctional artificial enzyme with both SOD and GPx activities based on a smart star-shaped pseudo-block copolymer. Soft Matter 2010, 6, 5342. [Google Scholar] [CrossRef]
- Buonocore, G.; Perrone, S.; Tataranno, M.L. Oxygen toxicity: chemistry and biology of reactive oxygen species. Semin Fetal Neonatal Med. 2010, 15, 186-90. [Google Scholar] [CrossRef]
- Bernard, K.E.; Parkes, T.L.; Merritt, T.J.S. A Model of Oxidative Stress Management: Moderation of Carbohydrate Metabolizing Enzymes in SOD1-Null Drosophila melanogaster. In PLoS ONE; Oliveira, PL, Ed.; 2011; Volume 6. [Google Scholar]
- Alfadda, A.A.; Sallam, R.M. Reactive Oxygen Species in Health and Disease. J Biomed Biotechnol. 2012, 2012, 1-14. [Google Scholar] [CrossRef]
- Terlecky, S.R.; Koepke, J.I.; Walton, P.A. Peroxisomes and aging. Biochim Biophys Acta BBA - Mol Cell Res. 2006, 1763, 1749-54. [Google Scholar] [CrossRef]
- Schrader, M.; Fahimi, H.D. Peroxisomes and oxidative stress. Biochim Biophys Acta BBA - Mol Cell Res. 2006, 1763, 1755-66. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; He, F.; Li, X.; Li, Y.; Huo, C.; Wang, H.; et al. Response pathways of superoxide dismutase and catalase under the regulation of triclocarban-triggered oxidative stress in Eisenia foetida: Comprehensive mechanism analysis based on cytotoxicity and binding model. Sci Total Environ. 2023, 854, 158821. [Google Scholar] [CrossRef]
- Piao, X.; Liu, Z.; Li, Y.; Yao, D.; Sun, L.; Wang, B.; et al. Investigation of the effect for bisphenol A on oxidative stress in human hepatocytes and its interaction with catalase. Spectrochim Acta A Mol Biomol Spectrosc. 2019, 221, 117149. [Google Scholar] [CrossRef] [PubMed]
- Koepke, J.I.; Nakrieko, K.; Wood, C.S.; Boucher, K.K.; Terlecky, L.J.; Walton, P.A.; et al. Restoration of Peroxisomal Catalase Import in a Model of Human Cellular Aging. Traffic 2007, 8, 1590-600. [Google Scholar] [CrossRef] [PubMed]
- Selvaratnam, J.; Robaire, B. Overexpression of catalase in mice reduces age-related oxidative stress and maintains sperm production. Exp Gerontol. 2016, 84, 12-20. [Google Scholar] [CrossRef] [PubMed]
- Chu, F.F.; Esworthy, R.S.; Chu, P.G.; Longmate, J.A.; Huycke, M.M.; Wilczynski, S.; et al. Bacteria-Induced Intestinal Cancer in Mice with Disrupted Gpx1 and Gpx2 Genes. Cancer Res. 2004, 64, 962-8. [Google Scholar] [CrossRef]
- Flohé, L.; Toppo, S.; Orian, L. The glutathione peroxidase family: Discoveries and mechanism. Free Radic Biol Med. 2022, 187, 113-22. [Google Scholar] [CrossRef] [PubMed]
- Herbette, S.; Labrouhe, D.T.D.; Drevet, J.R.; Roeckel-Drevet, P. Transgenic tomatoes showing higher glutathione peroxydase antioxidant activity are more resistant to an abiotic stress but more susceptible to biotic stresses. Plant Sci. 2011, 180, 548-53. [Google Scholar] [CrossRef]
- Pei, J.; Pan, X.; Wei, G.; Hua, Y. Research progress of glutathione peroxidase family (GPX) in redoxidation. Front Pharmacol. 2023, 14, 1147414. [Google Scholar] [CrossRef]
- Bersuker, K.; Hendricks, J.M.; Li, Z.; Magtanong, L.; Ford, B.; Tang, P.H.; et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature 2019, 575, 688-92. [Google Scholar] [CrossRef]
- Kipp, A.; Banning, A.; Brigelius-Flohé, R. Activation of the glutathione peroxidase 2 (GPx2) promoter by β-catenin. bchm 2007, 388, 1027-33. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid Redox Signal. 2014, 20, 1126-67. [Google Scholar] [CrossRef]
- Liu, K.L.; Sun, T.Z.; Yang, Y.; Gao, Q.X.; Tu, L.M.; Yu, J.Y.; et al. Blockade of PVN neuromedin B receptor alleviates inflammation via the RAS/ROS/NF-κB pathway in spontaneously hypertensive rats. Brain Res Bull. 2025, 220, 111180. [Google Scholar] [CrossRef]
- Chen, A.; Huang, H.; Fang, S.; Hang, Q. ROS: A “booster” for chronic inflammation and tumor metastasis. Biochim Biophys Acta BBA - Rev Cancer 2024, 1879, 189175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, Y.; Jing, L.; Zhao, H. Microplastics induced inflammation in the spleen of developmental Japanese quail (Coturnix japonica) via ROS-mediated p38 MAPK and TNF signaling pathway activation1. Environ Pollut. 2024, 341, 122891. [Google Scholar] [CrossRef]
- Fortes, G.B.; Alves, L.S.; De Oliveira, R.; Dutra, F.F.; Rodrigues, D.; Fernandez, P.L.; et al. Heme induces programmed necrosis on macrophages through autocrine TNF and ROS production. Blood 2012, 119, 2368-75. [Google Scholar] [CrossRef]
- Bang, E.; Kim, D.H.; Chung, H.Y. Protease-activated receptor 2 induces ROS-mediated inflammation through Akt-mediated NF-κB and FoxO6 modulation during skin photoaging. Redox Biol. 2021, 44, 102022. [Google Scholar] [CrossRef]
- Fransen, F.; Van Beek, A.A.; Borghuis, T.; Aidy, S.E.; Hugenholtz, F.; Van Der Gaast – De Jongh, C.; et al. Aged Gut Microbiota Contributes to Systemical Inflammaging after Transfer to Germ-Free Mice. Front Immunol. 2017, 8, 1385. [Google Scholar] [CrossRef]
- Thevaranjan, N.; Puchta, A.; Schulz, C.; Naidoo, A.; Szamosi, J.C.; Verschoor, C.P.; et al. Age-Associated Microbial Dysbiosis Promotes Intestinal Permeability, Systemic Inflammation, and Macrophage Dysfunction. Cell Host Microbe 2017, 21, 455–466.e4. [Google Scholar] [CrossRef] [PubMed]
- Millman, J.F.; Kondrashina, A.; Walsh, C.; Busca, K.; Karawugodage, A.; Park, J.; et al. Biotics as novel therapeutics in targeting signs of skin ageing via the gut-skin axis. Ageing Res Rev. 2024, 102, 102518. [Google Scholar] [CrossRef]
- Gao, T.; Li, Y.; Wang, X.; Tao, R.; Ren, F. Bifidobacterium longum 68S mediated gut-skin axis homeostasis improved skin barrier damage in aging mice. Phytomedicine 2023, 120, 155051. [Google Scholar] [CrossRef]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J Dermatol Sci. 2017, 85, 152-61. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.;; Chen, Z.M.; Fan, X.Y.; Jin, Y.L.; Li, X.; Wu, S.R.; et al. Gut–Brain–Skin Axis in Psoriasis: A Review. Dermatol Ther. 2021, 11, 25-38. [Google Scholar] [CrossRef] [PubMed]
- Hodnick, W.F.; Milosavljevi, E.B. ELECTROCHEMISTRY OF FLAVONOIDS. [CrossRef] [PubMed]
- Carr, A.; Frei, B. Does vitamin C act as a pro-oxidant under physiological conditions? FASEB J 1999, 13, 1007-24. [Google Scholar] [CrossRef]
- Prasad, A.; Rathi, D.; Sedlářová, M.; Manoharan, R.R.; Průdková, E.; Pospíšil, P. Differential effects of ascorbic acid on monocytic cell morphology and protein modification: Shifting from pro-oxidative to antioxidant properties. Biochem Biophys Rep. 2024, 37, 101622. [Google Scholar] [CrossRef] [PubMed]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem Pharmacol. 2015, 98, 371-80. [Google Scholar] [CrossRef]
- Tedesco, I.; Spagnuolo, C.; Russo, G.L.; Russo, M.; Cervellera, C.; Moccia, S. The Pro-Oxidant Activity of Red Wine Polyphenols Induces an Adaptive Antioxidant Response in Human Erythrocytes. Antioxidants 2021, 10, 800. [Google Scholar] [CrossRef]
- DePhillipo, N.N.; Aman, Z.S.; Kennedy, M.I.; Begley, J.P.; Moatshe, G.; LaPrade, R.F. Efficacy of Vitamin C Supplementation on Collagen Synthesis and Oxidative Stress After Musculoskeletal Injuries: A Systematic Review. Orthop J Sports Med. 2018, 6, 2325967118804544. [Google Scholar] [CrossRef]
- Darr, D.; Combs, S.; Dunston, S.; Manning, T.; Pinnell, S. Topical vitamin C protects porcine skin from ultraviolet radiation-induced damage. Br J Dermatol 1992, 127, 247-53. [Google Scholar] [CrossRef]
- Phillips, C.L.; Combs, S.B.; Pinnell, S.R. Effects of Ascorbic Acid on Proliferation and Collagen Synthesis in Relation to the Donor Age of Human Dermal Fibroblasts. J Invest Dermatol 1994, 103, 228-32. [Google Scholar] [CrossRef]
- Dulinska-Molak, I.; Pasikowska-Piwko, M.; Dębowska, R.; Święszkowski, W.; Rogiewicz, K.; Eris, I. Determining the effectiveness of vitamin C in skin care by atomic force microscope. Microsc Res Tech. 2019, 82, 1430-7. [Google Scholar] [CrossRef]
- Boo, Y.C. Ascorbic Acid (Vitamin C) as a Cosmeceutical to Increase Dermal Collagen for Skin Antiaging Purposes: Emerging Combination Therapies. Antioxidants 2022, 11, 1663. [Google Scholar] [CrossRef] [PubMed]
- Pullar, J.; Carr, A.; Vissers, M. The Roles of Vitamin C in Skin Health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef] [PubMed]
- Rychter, A.M.; Hryhorowicz, S.; Słomski, R.; Dobrowolska, A.; Krela-Kaźmierczak, I. Antioxidant effects of vitamin E and risk of cardiovascular disease in women with obesity – A narrative review. Clin Nutr. 2022, 41, 1557-65. [Google Scholar] [CrossRef]
- Higgins, M.; Izadi, A.; Kaviani, M. Antioxidants and Exercise Performance: With a Focus on Vitamin E and C Supplementation. Int J Environ Res Public Health 2020, 17, 8452. [Google Scholar] [CrossRef]
- Harris, I.S.; Treloar, A.E.; Inoue, S.; Sasaki, M.; Gorrini, C.; Lee, K.C.; et al. Glutathione and Thioredoxin Antioxidant Pathways Synergize to Drive Cancer Initiation and Progression. Cancer Cell. 2015, 27, 211-22. [Google Scholar] [CrossRef]
- Feng, Z.; Qin, Y.; Huo, F.; Jian, Z.; Li, X.; Geng, J.; et al. NMN recruits GSH to enhance GPX4-mediated ferroptosis defense in UV irradiation induced skin injury. Biochim Biophys Acta BBA - Mol Basis Dis. 2022, 1868, 166287. [Google Scholar] [CrossRef]
- Soeur, J.; Eilstein, J.; Léreaux, G.; Jones, C.; Marrot, L. Skin resistance to oxidative stress induced by resveratrol: From Nrf2 activation to GSH biosynthesis. Free Radic Biol Med. 2015, 78, 213-23. [Google Scholar] [CrossRef]
- Gunathilake, R.; Schurer, N.Y.; Shoo, B.A.; Celli, A.; Hachem, J.P.; Crumrine, D.; et al. pH-Regulated Mechanisms Account for Pigment-Type Differences in Epidermal Barrier Function. J Invest Dermatol 2009, 129, 1719-29. [Google Scholar] [CrossRef]
- Chikakane, K.; Takahashi, H. Measurement of skin pH and its significance in cutaneous diseases. Clin Dermatol 1995, 13, 299-306. [Google Scholar] [CrossRef]
- Ruffien-Ciszak, A.; Baur, J.; Gros, P.; Questel, E.; Comtat, M. Electrochemical microsensors for cutaneous surface analysis: Application to the determination of pH and the antioxidant properties of stratum corneum. IRBM 2008, 29(2-3), 162-70. [Google Scholar] [CrossRef]
- Agache, P. Physiologie de la peau et explorations fonctionnelles cutanées. 1ère; Tec & Doc Lavoisier, 2000. [Google Scholar]
- Proksch, E.; Brandner, J.M.; Jensen, J. The skin: an indispensable barrier. Exp Dermatol 2008, 17, 1063-72. [Google Scholar] [CrossRef] [PubMed]
- Boncheva, M.; De Sterke, J.; Caspers, P.J.; Puppels, G.J. Depth profiling of Stratum corneum hydration in vivo : a comparison between conductance and confocal Raman spectroscopic measurements. Exp Dermatol 2009, 18, 870-6. [Google Scholar] [CrossRef]
- Chicharro-Luna, E.; Zúnica-García, S.; Martinez-Algarra, C.; Gracia-Sánchez, A. Age-related variations in stratum corneum hydration in the foot. Maturitas 2024, 189, 108104. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.; Makrantonaki, E. Hormonal therapy of intrinsic aging. Rejuvenation Research 2012, 15, 302-12. [Google Scholar] [CrossRef] [PubMed]
- Hussen, N.H.A.; Abdulla, S.K.; Ali, N.M.; Ahmed, V.A.; Hasan, A.H.; Qadir, E.E. Role of antioxidants in skin aging and the molecular mechanism of ROS: A comprehensive review. Asp Mol Med. 2025, 5, 100063. [Google Scholar] [CrossRef]
- Schagen, S.K.; Zampeli, V.A.; Makrantonaki, E.; Zouboulis, C.C. Discovering the link between nutrition and skin aging. Dermatoendocrinol 2012, 4, 298-307. [Google Scholar] [CrossRef]
- Vierkötter, A.; Krutmann, J. Environmental influences on skin aging and ethnic-specific manifestations. Dermatoendocrinol 2012, 4, 227-31. [Google Scholar] [CrossRef]
- Solano, F. Photoprotection and Skin Pigmentation: Melanin-Related Molecules and Some Other New Agents Obtained from Natural Sources. Molecules 2020, 25, 1537. [Google Scholar] [CrossRef]
- Takeuchi, S.; Zhang, W.; Wakamatsu, K.; Ito, S.; Hearing, V.J.; Kraemer, K.H.; et al. Melanin acts as a potent UVB photosensitizer to cause an atypical mode of cell death in murine skin. Proc Natl Acad Sci. 2004, 101, 15076-81. [Google Scholar] [CrossRef]
- Tanaka, H.; Yamashita, Y.; Umezawa, K.; Hirobe, T.; Ito, S.; Wakamatsu, K. The Pro-Oxidant Activity of Pheomelanin is Significantly Enhanced by UVA Irradiation: Benzothiazole Moieties Are More Reactive than Benzothiazine Moieties. Int J Mol Sci. 2018, 19, 2889. [Google Scholar] [CrossRef]
- Pu, S.Y.; Huang, Y.L.; Pu, C.M.; Kang, Y.N.; Hoang, K.D.; Chen, K.H.; et al. Effects of Oral Collagen for Skin Anti-Aging: A Systematic Review and Meta-Analysis. Nutrients 2023, 15, 2080. [Google Scholar] [CrossRef]
- Nomoto, T.; Iizaka, S. Effect of an Oral Nutrition Supplement Containing Collagen Peptides on Stratum Corneum Hydration and Skin Elasticity in Hospitalized Older Adults: A Multicenter Open-label Randomized Controlled Study. Adv Skin Wound Care 2020, 33, 186-91. [Google Scholar] [CrossRef] [PubMed]
- Addor, F.A.S. Antioxidants in dermatology. An Bras Dermatol 2017, 92, 356-62. [Google Scholar] [CrossRef] [PubMed]
- Berneburg, M.; Plettenberg, H.; Krutmann, J. Photoaging of human skin. Photodermatol Photoimmunol Photomed. 2000, 16, 239-44. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



