Submitted:
25 February 2026
Posted:
28 February 2026
You are already at the latest version
Abstract
Sensorineural hearing loss (SNHL) is caused by excessive noise exposure, ototoxic drugs, and aging. SNHL is frequently linked to glutamate excitotoxicity, as glutamate is the main excitatory neurotransmitter at the inner hair cell – spiral ganglion neuron synapses. However, we still do not fully understand the specific roles of different glutamate receptor subtypes both in normal signaling and in causing excitotoxic damage. The investigation of these questions demands the availability of relevant experimental models. This review aims to compare existing protocols for obtaining primary spiral ganglion neurons in vitro and to assess the utility of this model in studying glutamatergic transmission. A literature search in PubMed and Google Scholar identified 16 relevant articles in English published since 1990, when the model was first introduced. Our analysis reveals significant heterogeneity protocols in isolation and conditions of cultivation. We highlight the significant differences in glutamate concentrations when modeling excitotoxicity and the glutamate receptor agonists used to study electrophysiological properties. The most significant limitation of this model is the loss of the native microenvironment of neurons, including their dendritic and axonal contacts. Nevertheless, primary spiral ganglion neurons serve as a suitable in vitro model for investigating auditory neuron function and pathology. This in vitro model allows detailed study of the mechanisms underlying ototoxicity and otoprotection. The number of neurons and neurite length serve as reliable indicators of otoprotective effects under conditions of glutamate excitotoxicity. This work may help researchers who plan to use the primary SGNs in their laboratories, as well as those who aim to optimize their methods based on accumulated experience.
Keywords:
spiral ganglion neurons
; glutamate excitotoxicity
; primary cells culture
; sensorineural hearing loss
; neurotransmission
; сochlear synaptopathy
1. Introduction
Sensorineural hearing loss (SNHL) is the most common type of hearing impairment. It is characterized by a loss of hair cells and/or damage to the auditory nerve. In the world, more than 5% of the population suffers from hearing loss [1]. One of the major contributors to hearing loss is acoustic trauma, which can result from both sudden and prolonged exposure to loud sounds [2]. Hearing loss is often associated with the use of ototoxic drugs, such as aminoglycoside antibiotics and platinum-based chemotherapeutic agents like cisplatin and carboplatin. Other drugs known to have ototoxic effects include loop diuretics, macrolides (e.g. azithromycin), antimalarial drugs (e.g. chloroquine and hydroxychloroquine) and NSAIDs (e.g. acetylsalicylic acid) [3]. Another significant factor in hearing loss is age-related degeneration of the sound-perceiving and sound-conducting apparatus of the auditory system [4].
In mammals, degeneration of sensory hair cells (HCs) and spiral ganglion neurons (SGNs) is irreversible, and a deficit in either of them can lead to permanent hearing loss [5]. Current hearing rehabilitation approaches, such as hearing aids and cochlear implants, can only partially restore hearing function. However, the quality of sound perceived through these devices does not replicate normal hearing [6,7].
Type I spiral neurons account for 90–95% of the spiral ganglion population. They are responsible for transmitting afferent signals from the inner hair cells to the brain [8]. Excitatory transmission at the inner hair cell/type I SGN synapse (IHC/ SGN synapse) is mediated by glutamate. The presynaptic terminals of inner hair cells release glutamate onto the postsynaptic terminals of type I SGN dendrites [9,10]. The postsynaptic terminals of these neurons express all classes of glutamate receptors, notably α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs) receptors, N-methyl-D-aspartate receptors (NMDAR), kainate receptors (KARs) and metabotropic receptors (mGluRs) [11,12,13]. Since AMPARs have fast kinetics, they are believed to play a key role in neurotransmission at the IHC/SGN synapse. In contrast, NMDARs, which are characterized by their slow kinetics, are thought to be directly uninvolved in neurotransmission at this synapse [14,15,16,17]. By comparison, NMDARs play a crucial role in neurotransmission within the central nervous system. At negative membrane potentials, NMDARs experience a magnesium block. This block can be removed through an initial depolarization of the membrane, which is mediated by AMPARs [18].
AMPARs are tetrameric ligand-gated ion channels that can be composed of various combinations of GluA1, GluA2, GluA3 and GluA4 subunits in either homomeric or heteromeric arrangements. They exhibit varying degrees of permeability to sodium, potassium and calcium ions. The presence of the GluA2 subunit typically renders the channel less permeable to calcium ions. It has been noted that during cochlear development, the expression of calcium-permeable AMPARs (CP-AMPARs) lacking the GluA2 subunit is suppressed. They suggest in the mature cochlea, glutamatergic transmission is facilitated by AMPARs consisting of GluA2, GluA3 and GluA4 subunits. Current evidence suggests that afferent fibers in the mature cochlea contain both GluA2-lacking CP-AMPARs and GluA2-containing calcium-impermeable AMPAR (CI-AMPARs) [16,19,20,21].
Exposure to acute or chronic noise, cochlear ischemia, ototoxic drugs lead to excessive glutamate release from the inner hair cells into the synaptic cleft. High levels of glutamate can cause overexcitation and the destruction of type I SGN dendrites [22]. It has been demonstrated that one of the key components mediating the effects of glutamate excitotoxicity in adult animals is the CP-AMPAR [20]. Excessive excitation of type I SGN terminals leads to the accumulation of Ca²⁺ and subsequent swelling of the terminals [19]. The glutamatergic agonist induces morphological changes similar to those seen after sound overexposure. Applying antagonists of CP-AMPARs prevents excitotoxic damage to afferent terminals [19,20]. Despite data indicating the presence of CP-AMPARs in the mature cochlea, their role in the development of glutamate excitotoxicity and prevalence in the mature inner ear are not yet fully understood. By comparison, glutamate excitotoxicity in the central nervous system is primarily caused by the overactivation of NMDARs [23]. This leads to a significant increase in the concentration of intracellular calcium. Intracellular processes are initiated that result in the excessive generation of reactive oxygen species (ROS), mitochondrial damage, and the activation of caspases and other pro-apoptotic factors [24,25,26].
The role of NMDA receptors in auditory neurons is still the subject of discussion. The molecular heterogeneity of NMDARs channels provides their dual role in physiological and pathological functions [27]. Previous studies show that combined application of NMDA and glycine receptor agonists in a magnesium-free solution does not produce a significant postsynaptic potential or alter calcium concentration in auditory neurons [14,15,28,29]. However, Wang et al. (2021) revealed that in isolated auditory neurons from newborn mice, intracellular calcium levels increase in response to NMDA when D-serine is present as a co-agonist, whereas no such increase occurs with glycine. Therefore, it is reasonable to propose that D-serine plays a key role in modulating the function of NMDARs in the cochlea in both normal physiological processes and pathological states [17]. It is assumed that NMDARs contribute to noise- and salicylate-induced tinnitus. Research suggests that NMDARs may be involved in gentamycin-induced ototoxicity [30]. They also play a role in noise-induced damage to auditory neuron afferents and presbycusis [31,32,33,34]. The specific types of receptors, the downstream mechanisms that cause damage and the factors that contribute to the selective vulnerability of certain synapses are not yet fully understood.
Given the dual role of the NMDARs, it is possible that they regulate the expression of the AMPARs to modulate synaptic transmission [35,36]. They may also contribute to neuritogenesis and synaptogenesis, as well as promoting the survival of spiral ganglion neurons in the developing cochlea [37,38].
Several studies have demonstrated the presence of mGluRs in the cochlea and their functional significance in auditory processing. The eight identified mGluRs subtypes are classified into three groups: Group I (mGluR1 and mGluR5); Group II (mGluR2 and mGluR3); and Group III (mGluR4, mGluR6, mGluR7 and mGluR8). The cochlea has been found to express members of all three groups of metabotropic glutamate receptors [39,40]. However, the cochlear function of individual mGluR subtypes is still poorly defined. It is believed that mGluRs have slow kinetics. Consequently, they are not directly involved in fast excitatory neurotransmission in the cochlea but rather modulate this process [41]. Experimental data suggested that mGluRI, localized on the postsynaptic membrane, modulates the excitability of spiral ganglion neurons. This leads to an enhanced response to intense stimulation, resulting in increased cochlear vulnerability to noise-induced trauma and glutamate excitotoxicity. Therefore, mGluR1 inhibitors could be a promising therapeutic approach for treating SNHL [12].
According to Friedman et al. (2009), genetic variations in GRM7, a gene encoding mGluR7, likely influence age-related hearing impairment (ARHI) by altering susceptibility to glutamate excitotoxicity in auditory neurons [42]. It is proposed that mGluR7 receptors, localized on the presynaptic membrane, act as autoreceptors by regulating glutamate concentration in the synaptic cleft through control of its release and preventing excessive glutamate accumulation [43,44].
Glutamate excitotoxicity is widely recognized as playing a central role in the mechanisms underlying noise-induced synaptopathy [45,46,47,48]. It has long been thought that the primary factor in the pathogenesis of noise-induced damage and age-related hearing loss is the loss of hair cells and spiral ganglion neurons. Decades of research have demonstrated that cochlear nerve synapses can degenerate even when hair cells and spiral ganglion neurons remain structurally intact. Research using animal models has demonstrated that moderate noise exposure initially results in the loss of synaptic connections between inner hair cells and their associated neurons. The subsequent delayed death of neurons is linked to the loss of connectivity with inner hair cells, which provide essential neurotrophic support for maintenance of the cochlear innervation [45]. The mechanism underlying the development of noise-induced synaptopathy is not fully understood. It is believed that excessive activation of CP-AMPARs by glutamate leads to damage to cochlear ribbon synapses [19,20]. It is suggested that one mechanism of synaptopathy involves noise-induced disruption of glutamate release and uptake-related protein expression [46]. Pure-tone audiometry is unable to detect this type of hearing impairment as hearing thresholds may remain within the normal range. Current clinical diagnostic methods lack the capability to identify diffuse synaptic damage between the inner hair cells and the auditory nerve fibers. For this reason, cochlear synaptopathy is often referred to as ″hidden hearing loss. ″
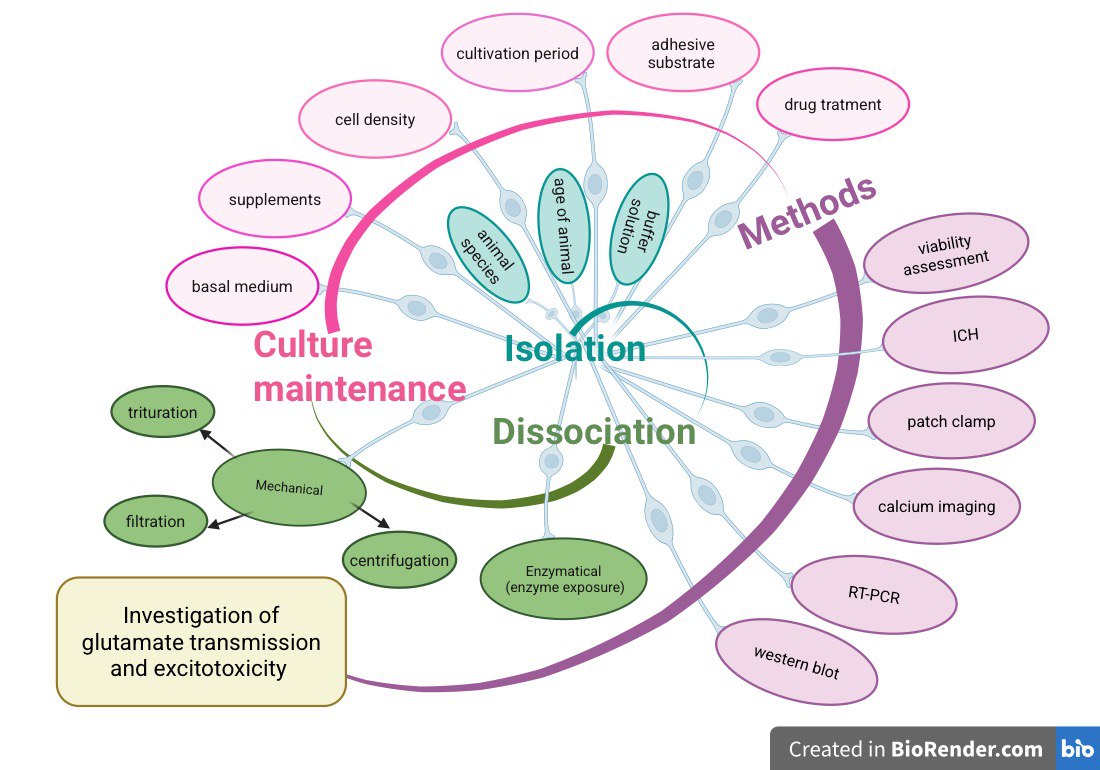
Primary neurons under in vitro conditions are a well-established experimental model, as evidenced by a significant number of studies published using primary neurons of brain, spinal cord, gastrointestinal tract, and other sources [49,50,51,52,53]. In auditory research, primary cultures of spiral ganglion cells (SGCs) have served as a valuable model for many years. This review provides examples of using SGCs isolated from the modiolar tissue of adult, neonatal, and embryonic animals to investigate glutamatergic transmission and excitotoxicity. Analysis of the presented studies can assist researchers in reproducing this model in their own laboratories, building upon the accumulated experience from previous work.
A better understanding of the function of glutamate receptors of SGNs and the mechanisms behind glutamate excitotoxicity could help us to develop new treatments for SNHL [21]. The investigation of these questions demands the availability of relevant experimental models. This review places a key focus on the methods of spiral ganglion cell isolation, dissociation, and culture since culture conditions affect the functional properties of SGNs.
2. Results
We present and analyze the main results on studying glutamatergic transmission and excitotoxicity using primary SGCs. We summarize the main parameters of isolation, dissociation, and culturing that can introduce artifacts when using this model. Our review analyzes articles from 1990 onward, when the primary spiral ganglion cell model in vitro was first used to investigate the glutamatergic system [53]. A literature search was conducted using the PubMed, Scopus, and Web of Science databases. We included in our analysis only English-language publications with accessible full text. During our analysis, we encountered a limitation related to the incomplete reporting of information in the protocols which partially restricted our analysis.
It is important to note that cell isolation from modiolar tissue results in a heterogeneous population of primary cells. This includes not only SGNs, but also other cell types, such as Schwann cells, satellite glia and fibroblasts. Therefore, in this context, cells derived from the modiolus are commonly referred to as SGCs in the literature. This review discusses studies in which primary SGCs are used as an in vitro system, either after culturing or immediately following isolation. Acutely isolated (non-cultured) cells serve as the primary model for functional assays, such as patch-clamp and calcium imaging, in the studies covered in this review. Studies investigating glutamate excitotoxicity using primary SGCs culture have typically included assessments of cellular morphology, viability, and the expression levels of pro- and anti-apoptotic factors and oxidative stress markers. Most studies on primary SGCs have used samples from newborns. This may be due to the difficulties involved in obtaining adult neurons.
The process of obtaining primary SGCs involves several stages. This analysis reviews the procedures described in various studies for obtaining primary SGCs, including the isolation of modiolus tissue and cell dissociation techniques, as well as the different methods available for culturing SGCs. Acutely isolated (non-cultured) cells either bypass the culturing step entirely or undergo only several hours of short-term culturing. Unlike cultured SGСs, acutely isolated SGСs are typically transferred directly into experimental solutions for immediate use. Existing protocols for obtaining and cultivating primary SGCs
vary across different studies. This can affect not only the survival and development of neurons but also the reproducibility of research methods utilizing these primary cultures. All procedures, including the dissection of the cochlea, preparation of reagents for cell dissociation and cultivation of primary cell cultures, must be conducted under sterile conditions. SGCs are highly sensitive to disturbances, so all steps are carried out with great care.
2.1. Cochlea Extraction and Modiolus Tissue Isolation
It is not described in the literature how methodological specifics at this stage impact on experimental outcomes in cochlear sample data in studies of glutamatergic effects. Thus, this section reviews the fundamental principles and procedures of cochlea extraction across different experimental contexts. The main parameter of this stage is the speed of cochlea isolation and surface dissection. The interval between animal decapitation and the placement of cochlear samples into the incubation or experimental medium should be kept as short as possible. These procedures require substantial practical experience and a refined technique in order to minimize the risk of damaging the sample [54].
Numerous articles describe methods for extraction and microdissection the cochlea of experimental animals at various ages [55,56,57,58,59,60,61,62,63]. However, specimens from the cochlea obtained during the prenatal, neonatal and early postnatal periods are more frequently used for isolating SGCs. The bony labyrinth of newborn rodents can easily be removed without damaging the underlying tissues because it has not yet undergone ossification. At the onset of postnatal day 3 (P3), the spiral lamina and the cartilage within the cochlea begin to ossify. Caution must be exercised when removing the bony capsule, spiral ligament, stria vascularis and organ of Corti, as neuronal cell bodies may detach from the modiolus along with the excised tissues. These procedures are performed using ultrafine forceps and microdissection knife. After P14, the removal of the completely ossified bony capsule of the bony labyrinth often damages the soft tissues of the cochlea, making it difficult to obtain intact samples for in vitro culture establishment [54,64].
The dissection of the cochlea and collection of the modiolar portion containing spiral ganglion neurons within the spiral lamina are performed in a balanced saline solution. For this purpose, ice-cold phosphate buffered solution [65,66], or Hank’s Balanced Salt Solution (HBSS) without calcium and magnesium ions are most commonly used [67,68]. Since most dissociation modes involve trypsin, the dissection solution is most often HBSS without calcium and magnesium, or PBS. Balanced buffer solutions provide a physiological environment with stable osmolarity and pH. Since most dissociation modes involve trypsin, the dissection solution is most often HBSS without calcium and magnesium, or PBS. Balanced buffer solutions provide a physiological environment with stable osmolarity and pH.
2.2. Dissociation
Modiolar tissue dissociation is achieved through a multistep procedure combining mechanical disruption, enzymatic digestion. Mechanical dissociation involves carefully pipetting the modiolar tissue to achieve a homogeneous solution, aiming for maximum cell dissociation. However, mechanical dissociation can result in cell loss, so the duration and intensity of pipetting must be carefully controlled to minimize damage. In the process of dissociation, auditory neurons lose their neurites. After cell seeding, neuritogenesis is triggered. Neurons regain their bipolar shape, but some neurons may acquire multipolar and unipolar shapes [69].
Enzymatic disaggregation of SGCs typically involves one or a combination of enzymes. The conditions of enzymatic dissociation, including the composition of the digestive enzyme solution used, duration exposure can have a significant impact on cell viability and various cellular functions. Table 1 summarizes the enzymatic dissociation protocols used in key studies investigating the glutamatergic system in primary spiral ganglion neurons.
In studies investigating the effects of glutamate on spiral ganglion neurons in primary culture, the enzymatic dissociation step mostly involves a combination of trypsin and collagenase in the digestive solution. Trypsin is the most commonly used enzyme to break down cell adhesion components. Collagenase, when used in combination with trypsin, is particularly essential for effectively isolating SGCs embedded in collagen-rich tissue of modiolus. Additionally, DNase is frequently included in the dissociation process to cleave DNA strands released from dead cells. This preventing cell aggregation and facilitating the release of individual cells into a homogeneous suspension.
Enzymatic dissociation has the potential to alter gene expression and disrupt signaling pathways in cells [70,71]. Therefore, it cannot be ruled out that the process of isolating and dissociating SGCs can significantly alter the expression profile of SGN glutamate receptors. Such changes could affect the cellular response to glutamate exposure in an experiment.
2.3. Culture Maintenance
Some factors can limit the accuracy and representativeness of results obtained from primary SGCs. To study the glutamatergic system in spiral ganglion cultures, conditions must allow the neurons to respond normally to glutamate receptor agonists and antagonists. We propose that this sensitivity depends on the expression levels and subunit composition of glutamate receptors, which are critically influenced by the culture conditions in vitro.
Primary cells of modiolus can be used in experiments immediately after isolation (acutely isolated cells) or subjected culturing. Previous studies use acutely isolated cells either right after preparing the suspension or following brief culturing [12,17,25,26,53,71]. Acutely isolated cells lack the full dendrites that mediate glutamatergic transmission in vivo. Their soma serves as a model for studying dendritic properties [28,72].
In culture, primary SGCs grown as a monolayer lack the natural microenvironment and specific intercellular signaling present in vivo [73]. This limitation alters the morphology, gene expression, and, consequently, the physiology of auditory neurons. Selecting the appropriate culture medium and coating substrate is essential for mimicking physiological conditions in vitro and ensuring successful cell cultivation.
The influence of substrate choice and culture medium composition on the sensitivity of spiral ganglion neurons to glutamatergic agonists remains uninvestigated. The influence of these factors on the functional properties of spiral ganglion neurons has not been studied. The methodological heterogeneity in culturing spiral ganglion neurons, as presented in Table 2, reflects the absence of established guidelines for substrate selection in glutamatergic system studies. However, spiral ganglion neurons demonstrate their viability when using various coating substrates, such as collagen [67], poly-L-lysine [74], poly-D-lysine [75,76], laminin [77], and poly-ornithine [17,78,79]. Commercially available cell adhesives are also utilized. Commercially available cell adhesives are also utilized [29,80,81,82]. However, poly-L-lysine is the most frequently mentioned coating substrate.
Primary cultures of spiral ganglion neurons are established using various media formulations. Research protocols examining glutamatergic effects on spiral ganglion neurons (SGNs) implement both serum-free and serum-containing culture. Standard modified Dulbecco’s Modified Eagle Medium (DMEM) and DMEM/F12 serves as the basal medium in most protocols, which is then supplemented with a range of additives such as serum, neurotrophins, growth factors, and specialized serum-free supplements B-27 and N-2 [83,84,85]. It is important to note that the antioxidant components of the B-27 supplement — such as vitamin E, vitamin E acetate, superoxide dismutase, catalase, and glutathione — enhance neuronal survival by reducing oxidative stress in cells [78]. Culture media typically contain amino acids, inorganic and organic ions, hormones, growth factors, as well as antibiotics and antifungals. These components help maintain optimal osmolarity, pH, viscosity, and the required levels of CO2 and oxygen. Primary cultures of SGCs are heterogeneous and consist not only of neurons but also other cell types, such as Schwann cells and satellite glia and fibroblasts [48,65]. In primary cell cultures derived from the cochlea of developing animals, there is an active proliferation of non-neuronal components, which poses challenges, particularly during prolonged cultivation. For this reason, most studies have only permitted short-term culture of spiral ganglion cell populations (2-5 days) [65]. Thus, the pre-experimental cultivation time is a critical parameter. The ratio neuronal/non-neuronal components in culture plays a crucial role, as the significant presence of non-neuronal cells inevitably influences the experimental outcomes. Several studies culturing SGCs mention the use of Neurobasal Medium (NBM) [75,86]. NBM is widely used to support neuronal growth and development in vitro. Initially, this culture medium was developed for the optimal cultivation of hippocampal neurons [87]. One of the advantages of NBM over other media is its ability to promote high neuronal survival rates while reducing the proliferation of glial cells in cultures [88]. Only one study investigating the effects of the glutamatergic system mentions the use of NBM supplemented with B27 for a 5-day culture period [84]. That culture exhibited a high proportion of neurons and good viability. The cell culture was predominantly composed of neurons with a normal bipolar morphology. We suggest that using neurobasal mediums could be a promising approach for optimizing the protocol to obtain primary SGCs cultures.
Cellular heterogeneity in primary cultures presents a challenge for techniques like immunoblotting. Observed changes in protein expression reflect the entire mixed population rather than neurons specifically [89]. Effective suppression of glial cell and fibroblast growth is achieved by adding the mitotic inhibitors like сytosine β-D-arabinofuranoside (Ara-C). Its mechanism of action is based on the disruption of DNA synthesis. Schwieger et al. demonstrated that the addition of 5 µM Ara-C allows for the prolongation of cultivation time up to one week and increases the ratio of neurons to non-neuronal cells [65]. When using cochleae from newborn animals, this approach may be necessary due to the presence of rapidly dividing stem cells. In the study by Xiao et al., which examined quinolinic acid-mediated effects, the authors reported using Ara-C. They noted that culture purity reached 95% SGNs [84].
The morphology of spiral ganglion neurons in vivo is typically bipolar, with dendrites connecting to inner hair cells and axons projecting to the cochlear nuclei in the medulla oblongata. SGNs require the maintenance of synaptic contacts with both their peripheral and central targets to ensure survival. Presynaptic cells can offer trophic support through the release of neurotrophic factors, neurotransmitters and by causing depolarization due to synaptic activity [79]. It is believed that the additive action of multiple neurotrophins plays a significant role in maintaining neuron survival and their normal bipolar morphology. Cochlear nucleus neurons can supply postsynaptic trophic support [90]. In vitro, spiral ganglion neurons lose afferent stimulation and trophic support from supporting cells, hair cells, and cochlear nucleus cells. To compensate for this deficit, adding neurotrophic factors to the culture medium is essential. Studies have shown that the neurotrophin family members brain derived neurotrophic factor (BDNF), neurotrophin-3 (NT3), neurotrophin-4/5 (NT-4/5) and the neuropoietic cytokines ciliary neurotrophic factor (CTNF), leukemia inhibitory factor (LIF) can significantly impact on neuronal survival and neurotogenesis in vitro [75,76,91,92,93]. Neurotrophic factors are potent modulators of the activity and expression of ion channels and receptors [94]. Therefore, using a combination of these factors is rational. It has been demonstrated that neurotrophic factors, when combined with depolarization induced by veratridine or elevated potassium concentrations in the medium, enhance the survival of spiral ganglion neurons in primary culture in an additive manner [79].
As the primary excitatory neurotransmitter, glutamate likely exerts a wide range of effects beyond neurotransmission, similar to its role in the central nervous system [95]. Neuronal maturation during development requires both trophic support and afferent-induced synaptic activity. Glutamatergic stimulation is intimately involved in these processes. After the dissociation step, where dendritic endings are lost, adding glutamate as a supplement to the culture medium can be important. We believe that moderate addition of glutamate can promote the formation of dendritic endings in vitro with functional properties similar to those in vivo in SGNs. It is clear that afferent stimulation is necessary for neuron maturation, synaptic connection formation, and neuron survival. However, there are no data on which glutamate concentration in the culture medium is suitable for these purposes. Glutamate activity largely depends on concentration and duration of exposure. Vieira et al. (2007) mentioned adding 25 μM glutamate to the culture medium, as it may have a trophic effect [75]. Both ionotropic glutamate receptors (iGluRs) and mGluRs can enhance neuronal survival and protect neurons from damage. It is important to note that glutamate stimulation furthermore plays a modeling role and affects the expression of glutamate receptors. This influences the functional response of SGNs.
Existing protocols for obtaining primary cell cultures vary across studies, which affects not only neuronal survival and development but also the reproducibility of methods using these primary cultures.
2.4. Functional Properties of the Glutamatergic System in Isolated Spiral Ganglion Neurons
The study of the glutamatergic system in isolated SGNs is conducted using two main approaches: primary cell cultures and experiments on acutely isolated (non-cultured) cells. During the isolation of dissociated cell cultures, spiral ganglion neurons are subjected to mechanical and enzymatic dissociation. This process causes unavoidable damage to the afferent nerve endings and axons of SGN. Immediately following dissociation, acutely isolated cells are rounded somata without neurites. Neurite outgrowth begins within a few hours. Thus, in contrast to acutely isolated cells, neurons in primary culture have regrown dendrites. It should be noted that in vivo neurotransmission occurs at the distal end of the dendrite in the IHC-SGN synapse. Therefore, neurons lacking dendrites or with regrown dendrites may exhibit different functional properties than those observed in vivo.
The data shown in Table 3 demonstrate a wide range of methods and recorded parameters applied in studies using glutamate receptor agonists and antagonists on primary SGNs. Research on the acutely isolated model was limited to patch-clamp techniques and functional calcium imaging employed to assess the responses to glutamate receptor agonists and antagonists. In studies using primary SGC cultures, researchers have assessed spiral ganglion neuron survival [66,67,81,85,91,96], neurite length [66,67], morphology [81,85], the expression levels of pro-apoptotic and anti-apoptotic factors [69,81,83], oxidative stress levels [81], and other parameters under glutamate excitotoxicity conditions. Consequently, when utilizing isolated SGCs model, it is important to consider that the processes of cell isolation and cultivation may lead to alterations in the expression, distribution, and subunit composition of glutamate receptors and ion channels [13]. These potential changes should be carefully accounted for in experimental design and interpretation.
Existing protocols for studying the glutamatergic system on primary cultures of SGCs predominantly utilize newborn animals. However, the subunit composition of glutamate receptors changes depending on the developmental stage, both in the central nervous system and in the peripheral nervous system. These changes directly affect the nature of neuronal responses to glutamate [97]. IHC-SGN synapses undergo maturation, acquiring their final morphological and functional characteristics during the first two postnatal weeks, with the onset of hearing occurring at postnatal day 12 (P12) in mice [98,99]. However, given that cochlear dissection is technically easier in neonates than in adults [54], most existing protocols for primary SGN cultures utilize newborn animals.
2.4.1. Acutely Isolated Cells
Using patch-clamp electrophysiology, Nakagawa et al. (1991) provided the first characterization of excitatory amino acid responses in isolated mammalian spiral ganglion neurons (SGNs). This study was among the first to show that postsynaptic responses in mammals are mediated primarily by non-NMDAR. The authors note the absence of dendrites in acutely isolated SGNs [28]. Nevertheless, the findings of this study were corroborated by in vivo investigations [14,17].
Harada et al. were the first to apply fluorescent calcium imaging to isolated SGCs specifically to study their response to L-glutamate [80]. Their critical finding was that the glutamate-induced increase in intracellular calcium ([Ca²⁺]ᵢ) was strictly dependent on the presence of neurites. Isolated SGCs without neurites did not respond to glutamate exposure. Therefore, the authors showed a dependence of SGN function on morphology under in vitro conditions. Their work also provided critical evidence supporting the hypothesis that neurotransmission at the mammalian IHC-SGN synapse is mediated by non-NMDAR. These results were obtained indirectly in the presence of extracellular Mg²⁺, which blocks NMDAR.
Shimozono et al. first demonstrated on isolated SGCs that fast excitatory synaptic transmission at the mammalian IHC–SGN synapse is mediated by non-NMDAR, using specific agonists (kainate and quisqualate). They confirmed that [Ca²⁺]ᵢ did not increase in response to NMDA and that the glutamate-induced response was not blocked by the NMDAR antagonist (APV). This provides further support for the findings of Harada et al. An important finding of this study was that non-NMDAR-mediated calcium influx is mediated primarily by voltage-gated calcium channels. In this study, patch-clamp electrophysiology and functional calcium imaging were applied in combination for the first time on isolated SGCs [29]. This work contributed to a better understanding of glutamatergic signaling in SGNs. The authors employed an SGNs isolation protocol akin to that of Nakagawa et al., who noted that neurons prepared this way lack processes. Notably, Shimozono et al. observed [Ca²⁺]ᵢ rises in response to glutamate and non-NMDAR agonists. It is unclear if this conflicts with the report by Harada et al., who found that neurites are required for glutamate-induced calcium influx. Both studies did not specify pre-experimental incubation times. The cells in the Shimozono et al. study may have regrown processes before imaging.
In addition to existing in vivo studies, experiments on acutely isolated cells have provided further insights into the role of NMDAR in SGNs. Functional calcium imaging revealed the specific modulation of excitatory NMDAR responses by D-serine, rather than by glycine. Along with in vivo data, it has been suggested that D-serine is involved in noise-induced cochlear injuries, probably through potentiation of cochlear NMDARs [17]. It is important to note that the protocol of this study also does not specify the pre-experimental incubation time and does not describe cell morphology.
2.4.2. Primary SGCs Culture
An early study by Yamaguchi et al. utilized a long-term primary culture of chick embryo SGN (5–14 days), in which neurons had the opportunity to regenerate their processes. They demonstrated that the excitatory postsynaptic potential (EPSP) is generated by NMDAR in these cells [100]. A key feature of this study was the utilization of non-mammalian neurons. Subsequent research has revealed distinct differences in neurotransmission between mammals and non-mammals. Historically, this work was the first to employ a primary culture of spiral ganglion neurons as a model system for investigating the glutamatergic system.
Subsequent studies have shown that the EPSP in mammals is mediated by AMPAR in the cochlea [14]. Chen et al., using a short-term SGCs culture, showed NMDA-dependent regulation of the number of AMPARs on the surface of SGNs from newborn mice. This was one of the first pieces of evidence that NMDARs play a modulatory role in neurotransmission in the mammalian cochlea, unlike AMPARs. The researchers confirmed these results in in vivo experiments [35,36].
Using primary SGCs culture, Peng et al. evaluated the role of metabotropic receptors. Metabotropic glutamate receptors are present in the cochlea, where they participate in auditory processing [40]. Peng et al. (2004) were among the first to demonstrate the functional role of mGluR1 in SGNs. They combined in vitro approaches, including calcium imaging and patch-clamp, with in vivo experiments [12]. The authors also revealed the protective effects of mGluR antagonists, suggesting the involvement of these receptors in the mechanisms of glutamate-induced excitotoxicity.
The study by Xiao et al. represents another example where functional calcium imaging was applied to primary cultures of SGCs [84]. The use of primary SGCs culture demonstrated the neurotoxic effect of quinolinic acid (QA) on these neurons. This finding suggests potential role of QA in the pathogenesis of SNHL associated with otitis media with effusion (OME). The authors demonstrated that quinolinic acid induces transient changes in intracellular calcium concentration in SGNs, which are mediated by NMDAR activation with functional calcium imaging. Quinolinic acid is known to play a role in various inflammatory diseases of the central nervous system. However, few studies have examined its neurotoxicity in the inner ear.
When studying primary SGCs cultures, researchers use various methods, such as fluorescence and confocal microscopy, Western blot, RT-PCR, and viability assays (e.g., MTT). The application of this model has enabled key advances in understanding glutamate excitotoxicity in the inner ear. Lefebvre et al. (1991) were the first to establish an in vitro model of glutamate excitotoxicity in mammals using primary cultures of SGNs derived from both adult and neonatal rats [91]. Using primary cultures of spiral ganglion neurons from newborn rats, Ding and colleagues demonstrated the critical role of apoptosis-inducing factor (AIF) and calpain activation in glutamate-mediated damage and apoptosis of SGNs [83,101].
This model enables examination of otoprotective compound activity. In a study conducted by Sun et al. (2021) using primary cultures of neonatal mouse spiral ganglion cells, a neuroprotective effect of natriuretic peptide was demonstrated in a model of glutamate excitotoxicity [85]. The number of neurons in the dish and the length of neurites were chosen as indicators to demonstrate the neuroprotective effect. Bai et al. demonstrated that Edaravone exerts a dose-dependent neuroprotective effect in primary SGCs culture under glutamate excitotoxicity conditions, as evidenced by improved neuronal survival and reduced apoptotic markers [81]. Using this model, Li et al. demonstrated the neuroprotective properties of vancomycin [96].
These studies demonstrate that an in vitro SGCs model can be used to simulate glutamate excitotoxicity and investigate the neuroprotective properties of compounds.
3. Discussion
The use of in vitro models is a widely adopted approach in auditory system research. The present review summarizes studies in which primary SGCs are employed as an experimental model to study glutamate transmission and to investigate the phenomenon of glutamate-induced excitotoxicity. Early studies of glutamatergic transmission in primary spiral ganglion neurons used chicken cells [28,100]. However, studies have shown a fundamental difference between birds and mammals. In chickens, NMDAR generate postsynaptic currents. In mammals, this role is performed by AMPAR [28]. Subsequent studies predominantly utilized mammalian models. Thus, when selecting an experimental animal model, it is necessary to take into account the presence of functional difference of the cells.
The process of cochlear isolation from the skull is associated with a high risk of mechanical damage to the soft tissues enclosed in the bony labyrinth. Cochlear isolation and inner ear tissue processing can be performed in experimental animals of varying ages [54]. However, neonatal mice or rats are the most common source of cochleae for creating in vitro and ex vivo models. At this age, the skull and cochlea are not yet ossified. This facilitates the isolation of the membranous labyrinth structures from the otic capsule [64]. In newborn mice, the removal of the bony cochlear capsule can be carried out without damaging the underlying tissues, including the modiolar region. After P14, the ossification of the murine labyrinthine capsule makes its removal difficult while preserving the integrity of the soft tissue structures of the cochlea [54,60,62]. Thus, microdissection of the cochlea of an adult animal presents significant technical challenges.
The functional properties of glutamatergic signaling change throughout postnatal development. This maturation is reflected in shifts in the expression profile and subunit composition of glutamate receptors, which ultimately shape the functional characteristics of SGNs. Research indicates that the period of greatest sensitivity to excitotoxicity caused by glutamate agonists is during the early stages of development. Specifically, this occurs in rats between postnatal days 9 and 12 [102].
Despite the technical challenges associated with isolating spiral ganglion neurons from adult animals, such protocols are mentioned in several studies [80,91]. Therefore, functional differences in spiral ganglion neurons at various developmental stages must be considered when investigating the role of glutamate and glutamate-induced excitotoxicity in isolated neurons.
It should be taken into account that under in vitro conditions the cell phenotype may differ from the phenotype of cells in their natural environment. Cell behavior in 2D monolayers in culture differs from that observed in vivo: morphology, receptor expression, and gene expression changes. Thus, the use of primary SGCs is associated with a number of limitations [88]. Parameters of cell isolation and cultivation protocols may substantially influence experimental outcomes. Our analysis reveals significant heterogeneity in experimental protocols for isolating SGCs, especially regarding dissociation methods and culture conditions.
Spiral ganglion neurons have a bipolar morphology in vivo. Type I SGNs form glutamatergic synaptic contacts with inner hair cells through their dendrites and transmit information to the brainstem via their axons [76]. Under in vivo conditions, the neurotransmitter is released from inner hair cells onto the distal end of the dendrite at the IHC-SGN synapse [41]. It is important to note that during the preparation of cell suspension, isolated neurons lose their processes. This limitation was noted by pioneers using patch-clamp to study mammalian glutamatergic transmission in isolated SGNs, Nakagawa et al. [28]. Based on the approach of Feltz and Rasminsky (1974) in studying neurons of the lumbar spinal ganglia, Nakagawa proposed using the spiral ganglion neuron soma and a short dendrite as a model of the primary afferent nerve terminal [72].
Enzymatic and mechanical dissociation leads to the loss of the natural cellular microenvironment and contacts with the extracellular matrix, which can trigger programmed cell death – anoikis [103]. Therefore, the interval between isolating cells from the modiolus and their attachment to a coating substrate should be minimized in time. Besides mechanical and enzymatic dissociation alters gene expression profiles [70]. It has been observed that varying dissociation protocols result in different gene expression in cultured cells [71]. It remains unclear how much the dissociation process alters functional characteristics of spiral ganglion neurons. Therefore, it is important to investigate changes in the expression levels of key glutamate neurotransmission elements following isolation. This will minimize dissociation-induced artifacts.
The loss of the natural intercellular environment requires improved approaches to culturing SGSc. This includes the use of specialized media, such as NBM, to enhance neuronal survival, as well as the use of supplements, particularly neurotrophic factors [75,86]. However, the influence of these factors on the functional properties of spiral ganglion neurons has not been studied. Therefore, the influence of culture parameters on the glutamatergic system of auditory neurons in vitro remains unclear.
Modiolus cell suspension culture contains neurons, satellite cells, Schwann cells, and fewer fibroblasts, astrocytes, and oligodendrocytes. Non-neuronal cells divide actively and overgrow the culture, accordingly long-term culture of SGCs is difficult. The cellular heterogeneity of primary SGCs cultures due to non-neuronal cells complicates data interpretation. This problem is especially severe in cultures from newborn animals. Using the mitotic inhibitor AraC was suggested as a method to improve the neuronal proportion in culture [65].
There are no standardized approaches to obtaining or using this in vitro model for studying the glutamatergic system in SGNs. The present review summarizes the accumulated experience from previous studies in the field. The resulting analysis may be useful to researchers planning to work with the model.
4. Conclusion
Isolated SGCs model continues to be actively used in experimental otolaryngology. The properties of neurons under in vitro conditions clearly differ from those in vivo. This limitation is inherent to any in vitro system in principle. This complicates the interpretation of experimental data, because the cell phenotype, including receptor expression, is inevitably altered by the cell isolation methods and artificial culture conditions. The protocols for obtaining and culturing primary SGNs demonstrate heterogeneity in existing studies, making it difficult to compare results between different studies. Despite this, there is already an understanding of how dissociation and culture conditions affect the structure and function of primary spiral ganglion neurons.
Beside some limitation, isolated SGCs model in vitro has made significant contributions to understanding the glutamatergic system and excitotoxicity mechanisms in auditory neurons. Acutely isolated SGN neurons can be used to determine signal transduction and receptor function. Cultured cells are better suited for studying calcium dysregulation and the mechanisms of glutamate excitotoxicity or otoprotection. The number of neurons in a Petri dish and also the length of neurites can serve as good indicators of the neuroprotective effect under conditions of glutamate excitotoxicity. Thus, although the lack of in vivo microenvironmental cues significantly reduces the physiological relevance of in vitro studies, the ability to manipulate spiral ganglion neurons outside the body represents an important advantage for understanding the specific mechanisms of their death and survival.
Author Contributions
Conceptualization, E.P. and Z.B.; investigation, E.P. and Z.B.; resources, A.F. and E.S.; data curation, E.S. and A.F.; writing—original draft preparation, E.P.; writing—review and editing, Z.B.; visualization, E.P.; project administration, Z.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This review only presents data that have been previously published. No new data were generated.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| 7-CKA | 7-chlorokynurenic acid |
| ACPD | (1S,3R)-aminocyclopentane-1,3-dicarboxylic acid |
| AIDA | (RS)-1-aminoindan-1,5-dicarboxylic acid |
| AIF | Apoptosis-inducing factor |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| AMPARs | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors |
| Asp | Aspartic acid |
| AraC | Cytosine β-D-arabinofuranoside |
| BDNF | Brain derived neurotrophic factor |
| CI-AMPARs | Calcium-impermeable AMPARs |
| CNQX | 6-cyano-7-nitroquinoxaline-2,3-dione |
| CP-AMPARs | Calcium-permeable AMPARs |
| DAP-5 | 2-amino-5-phosphonovalerate |
| DHPG | (S)-3,5-Dihydroxyphenylglycine |
| diCl-HQC | 6,7-dichloro-3-hydroxy-2-quinoxalinecarboxylic acid |
| DNQX | 6,7-dinitroquinoxaline-2,3-dione |
| EPSP | Excitatory postsynaptic potential |
| Glu | Glutamate |
| GRM7 | Metabotropic glutamate receptor 7 |
| GYKI 53784 | 1-(4-aminophenyl)-4-methyl-7, 8-methylenedioxy-4,5-dihydro-3-methylcarbamoyl-2,3-benzodiazepine |
| iGluRs | Ionotropic glutamate receptors |
| KA | Kainic acid |
| KARs | Kainic acid receptors |
| Kyn | Kynuranic acid |
| mGluRs | Metabotropic glutamate receptors |
| MK-801 | Dizocilpine |
| NBM | Neurobasal medium |
| NMDA | N-methyl-D-aspartate |
| NMDARs | N-methyl-D-aspartate receptors |
| NSAIDs | Nonsteroidal anti-inflammatory drug |
| NT-4/5 | Neurotrophin-4/5 |
| NT3 | Neurotrophin-3 |
| OME | Otitis media with effusion (OME) |
| QA | Quisqualic acid |
| ROS | Reactive oxygen species |
| SGCs | Spiral ganglion cells |
| SGNs | Spiral ganglion neurons |
References
- Chadha, S.; Kamenov, K.; Cieza, A. The World Report on Hearing, 2021. Bull. World Health Organ. 2021, 99, 242–242A. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, N.; Batts, S.; Stankovic, K.M. Noise-Induced Hearing Loss. J. Clin Med. 2023, 12, 2347. [Google Scholar] [CrossRef]
- Pasdelou, M.-P.; Byelyayeva, L.; Malmström, S.; Pucheu, S.; Peytavy, M.; Laullier, H.; Hodges, D.B.; Tzafriri, A.R.; Naert, G. Ototoxicity: A High Risk to Auditory Function That Needs to Be Monitored in Drug Development. Front. Mol. Neurosci. 2024, 17, 1379743. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, X.; Chai, R.; Fan, J. Progress on Mechanisms of Age-Related Hearing Loss. Front. Neurosci. 2023, 17, 1253574. [Google Scholar] [CrossRef] [PubMed]
- Bing, X. Development of the Inner Ear and Regeneration of Hair Cells after Hearing Impairment. Fundam. Res. 2025, 5, 203–214. [Google Scholar] [CrossRef]
- Blankenship, C.; Zhang, F.; Keith, R. Behavioral Measures of Temporal Processing and Speech Perception in Cochlear Implant Users. J. Am. Acad. Audiol. 2016, 27, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Lesica, N.A. Why Do Hearing Aids Fail to Restore Normal Auditory Perception? Trends Neurosci. 2018, 41, 174–185. [Google Scholar] [CrossRef]
- Barclay, M.; Ryan, A.F.; Housley, G.D. Type I vs Type II Spiral Ganglion Neurons Exhibit Differential Survival and Neuritogenesis during Cochlear Development. Neural. Dev. 2011, 6, 33. [Google Scholar] [CrossRef]
- Nayagam, B.A.; Muniak, M.A.; Ryugo, D.K. The spiral ganglion: connecting the peripheral and central auditory systems. Hear. Res. 2011, 278, 2–20. [Google Scholar] [CrossRef]
- Coate, T.M.; Kelley, M.W. Making Connections in the Inner Ear: Recent Insights into the Development of Spiral Ganglion Neurons and Their Connectivity with Sensory Hair Cells. Semin. Cell Dev. Biol. 2013, 24, 460–469. [Google Scholar] [CrossRef]
- Niedzielski, A.; Wenthold, R. Expression of AMPA, Kainate, and NMDA Receptor Subunits in Cochlear and Vestibular Ganglia. J. Neurosci. 1995, 15, 2338–2353. [Google Scholar] [CrossRef]
- Peng, B.G.; Li, Q.X.; Ren, T.Y.; Ahmad, S.; Chen, S.P.; Chen, P.; Lin, X. Group I Metabotropic Glutamate Receptors in Spiral Ganglion Neurons Contribute to Excitatory Neurotransmissions in the Cochlea. Neuroscience 2004, 123, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Reijntjes, D.O.J.; Pyott, S.J. The Afferent Signaling Complex: Regulation of Type I Spiral Ganglion Neuron Responses in the Auditory Periphery. Hear. Res. 2016, 336, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ruel, J.; Chen, C.; Pujol, R.; Bobbin, R.P.; Puel, J.L. AMPA-preferring Glutamate Receptors in Cochlear Physiology of Adult Guinea-pig. J. Physiol. 1999, 518, 667–680. [Google Scholar] [CrossRef]
- Glowatzki, E.; Fuchs, P.A. Transmitter Release at the Hair Cell Ribbon Synapse. Nat. Neurosci. 2002, 5, 147–154. [Google Scholar] [CrossRef]
- Eybalin, M.; Caicedo, A.; Renard, N.; Ruel, J.; Puel, J. Transient Ca2+ -permeable AMPA Receptors in Postnatal Rat Primary Auditory Neurons. Eur. J. Neurosci. 2004, 20, 2981–2989. [Google Scholar] [CrossRef]
- Wang, J.; Serratrice, N.; Lee, C.J.; François, F.; Sweedler, J.V.; Puel, J.-L.; Mothet, J.-P.; Ruel, J. Physiopathological Relevance of D-Serine in the Mammalian Cochlea. Front. Cell. Neurosci. 2021, 15, 733004. [Google Scholar] [CrossRef]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA Receptor Subunit Diversity: Impact on Receptor Properties, Synaptic Plasticity and Disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Sebe, J.Y.; Cho, S.; Sheets, L.; Rutherford, M.A.; Von Gersdorff, H.; Raible, D.W. Ca2+ -Permeable AMPARs Mediate Glutamatergic Transmission and Excitotoxic Damage at the Hair Cell Ribbon Synapse. J. Neurosci. 2017, 37, 6162–6175. [Google Scholar] [CrossRef]
- Hu, N.; Rutherford, M.A.; Green, S.H. Protection of Cochlear Synapses from Noise-Induced Excitotoxic Trauma by Blockade of Ca2+ -Permeable AMPA Receptors. Proc. Natl. Acad. Sci. U.S.A. 2020, 117, 3828–3838. [Google Scholar] [CrossRef]
- Walia, A.; Lee, C.; Hartsock, J.; Goodman, S.S.; Dolle, R.; Salt, A.N.; Lichtenhan, J.T.; Rutherford, M.A. Reducing Auditory Nerve Excitability by Acute Antagonism of Ca2+-Permeable AMPA Receptors. Front. Synaptic Neurosci. 2021, 13, 680621. [Google Scholar] [CrossRef]
- Steinbach, S.; Lutz, J. Glutamate Induces Apoptosis in Cultured Spiral Ganglion Explants. Biochem. Biophys. Res. Commun. 2007, 357, 14–19. [Google Scholar] [CrossRef]
- Lewerenz, J; Maher, P. Chronic Glutamate Toxicity in Neurodegenerative Diseases-What is the Evidence? Front Neurosci. 2015, 16, 469. [Google Scholar] [CrossRef]
- Pinelis, V.; Krasilnikova, I.; Bakaeva, Z.; Surin, A.; Boyarkin, D.; Fisenko, A.; Krasilnikova, O.; Pomytkin, I. Insulin Diminishes Superoxide Increase in Cytosol and Mitochondria of Cultured Cortical Neurons Treated with Toxic Glutamate. Int. J. Mol. Sci. 2022, 23, 12593. [Google Scholar] [CrossRef] [PubMed]
- Bakaeva, Z.; Goncharov, M.; Frolov, F.; Krasilnikova, I.; Sorokina, E.; Zgodova, A.; Smolyarchuk, E.; Zavadskiy, S.; Andreeva, L.; Myasoedov, N.; Fisenko, A.; Savostyanov, K. Regulatory Peptide Pro-Gly-Pro Accelerates Neuroregeneration of Primary Neuroglial Culture after Mechanical Injury in Scratch Test. Int. J. Mol. Sci. 2024, 25, 10886. [Google Scholar] [CrossRef] [PubMed]
- Shedenkova, M.; Gurianova, A.; Krasilnikova, I.; Sudina, A.; Karpulevich, E.; Maksimov, Y.; Samburova, M.; Guguchkin, E.; Nefedova, Z.; Babenko, V.; Frolov, D.; Savostyanov, K.; Fatkhudinov, T.; Goldshtein, D.; Bakaeva, Z.; Salikhova, D. Extracellular Vesicles from iPSC-Derived Glial Progenitor Cells Prevent Glutamate-Induced Excitotoxicity by Stabilising Calcium Oscillations and Mitochondrial Depolarisation. Cells 2025, 14, 1915. [Google Scholar] [CrossRef]
- Yamakura, T.; Shimoji, K. Subunit- and Site-Specific Pharmacology of the NMDA Receptor Channel. Prog. Neurobiol. 1999, 59, 279–298. [Google Scholar] [CrossRef]
- Nakagawa, T.; Komune, S.; Uemura, T.; Akaike, N. Excitatory Amino Acid Response in Isolated Spiral Ganglion Cells of Guinea Pig Cochlea. J. Neurophysiol. 1991, 65, 715–723. [Google Scholar] [CrossRef]
- Shimozono, M.; Tono, T.; Morimitsu, T.; Nakagawa, T.; Komune, S. Measurement of intracellular free Ca2+ concentration in guinea pig spiral ganglion cells. Neuroreport. 1995, 6, 421–424. [Google Scholar] [CrossRef]
- Hong, J.; Chen, Y.; Zhang, Y.; Li, J.; Ren, L.; Yang, L.; Shi, L.; Li, A.; Zhang, T.; Li, H.; et al. N-Methyl-D-Aspartate Receptors Involvement in the Gentamicin-Induced Hearing Loss and Pathological Changes of Ribbon Synapse in the Mouse Cochlear Inner Hair Cells. Neural. Plasticity 2018, 2018, 1–16. [Google Scholar] [CrossRef]
- Guitton, M.J.; Caston, J.; Ruel, J.; Johnson, R.M.; Pujol, R.; Puel, J.-L. Salicylate Induces Tinnitus through Activation of Cochlear NMDA Receptors. J. Neurosci. 2003, 23, 3944–3952. [Google Scholar] [CrossRef]
- Ruel, J.; Chabbert, C.; Nouvian, R.; Bendris, R.; Eybalin, M.; Leger, C. L.; Bourien, J.; Mersel, M.; Puel, J. L. Salicylate enables cochlear arachidonic-acid-sensitive NMDA receptor responses. J Neurosci. 2008, 28, 7313–7323. [Google Scholar] [CrossRef]
- Bing, D.; Lee, S.C.; Campanelli, D.; Xiong, H.; Matsumoto, M.; Panford-Walsh, R.; Wolpert, S.; Praetorius, M.; Zimmermann, U.; Chu, H.; Knipper, M.; Rüttiger, L.; Singer, W. Cochlear NMDA Receptors as a Therapeutic Target of Noise-Induced Tinnitus. Cell Physiol. Biochem. 2015, 35, 1905–1923. [Google Scholar] [CrossRef]
- Tang, X.; Zhu, X.; Ding, B.; Walton, J.P.; Frisina, R.D.; Su, J. Age-Related Hearing Loss: GABA, Nicotinic Acetylcholine and NMDA Receptor Expression Changes in Spiral Ganglion Neurons of the Mouse. Neuroscience 2014, 259, 184–193. [Google Scholar] [CrossRef]
- Chen, Z.; Kujawa, S.G.; Sewell, W.F. Auditory Sensitivity Regulation via Rapid Changes in Expression of Surface AMPA Receptors. Nat. Neurosci. 2007, 10, 1238–1240. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Peppi, M.; Kujawa, S.G.; Sewell, W.F. Regulated Expression of Surface AMPA Receptors Reduces Excitotoxicity in Auditory Neurons. J. Neurophysiol. 2009, 102, 1152–1159. [Google Scholar] [CrossRef]
- Sanchez, J.T.; Ghelani, S.; Otto-Meyer, S. From Development to Disease: Diverse Functions of NMDA-Type Glutamate Receptors in the Lower Auditory Pathway. Neuroscience 2015, 285, 248–259. [Google Scholar] [CrossRef]
- Zhang-Hooks, Y.; Agarwal, A.; Mishina, M.; Bergles, D.E. NMDA Receptors Enhance Spontaneous Activity and Promote Neuronal Survival in the Developing Cochlea. Neuron 2016, 89, 337–350. [Google Scholar] [CrossRef]
- Klotz, L.; Wendler, O.; Frischknecht, R.; Shigemoto, R.; Schulze, H.; Enz, R. Localization of Group II and III Metabotropic Glutamate Receptors at Pre- and Postsynaptic Sites of Inner Hair Cell Ribbon Synapses. FASEB J. 2019, 33, 13734–13746. [Google Scholar] [CrossRef]
- Lu, Y. Metabotropic Glutamate Receptors in Auditory Processing. Neuroscience 2014, 274, 429–445. [Google Scholar] [CrossRef]
- Kleinlogel, S.; Oestreicher, E.; Arnold, T.; Ehrenberger, K.; Felix, D. Metabotropic Glutamate Receptors Group I Are Involved in Cochlear Neurotransmission. Neuroreport. 1999, 10, 1879–1882. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.A.; Van Laer, L.; Huentelman, M.J.; Sheth, S.S.; Van Eyken, E.; Corneveaux, J.J.; Tembe, W.D.; Halperin, R.F.; Thorburn, A.Q.; Thys, S.; et al. GRM7 Variants Confer Susceptibility to Age-Related Hearing Impairment. Hum. Mol. Genet. 2009, 18, 785–796. [Google Scholar] [CrossRef]
- Klotz, L.; Enz, R. MGluR7 Is a Presynaptic Metabotropic Glutamate Receptor at Ribbon Synapses of Inner Hair Cells. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Ma, X.; Guo, J.; Fu, Y.; Shen, C.; Jiang, P.; Zhang, Y.; Zhang, L.; Yu, Y.; Fan, J.; Chai, R. G Protein-Coupled Receptors in Cochlea: Potential Therapeutic Targets for Hearing Loss. Front. Mol. Neurosci. 2022, 15, 1028125. [Google Scholar] [CrossRef] [PubMed]
- Liberman, M.C.; Kujawa, S.G. Cochlear Synaptopathy in Acquired Sensorineural Hearing Loss: Manifestations and Mechanisms. Hear. Res. 2017, 349, 138–147. [Google Scholar] [CrossRef]
- Ma, K.; Zhang, A.; She, X.; Yang, H.; Wang, K.; Zhu, Y.; Gao, X.; Cui, B. Disruption of Glutamate Release and Uptake-Related Protein Expression After Noise-Induced Synaptopathy in the Cochlea. Front. Cell Dev. Biol. 2021, 9, 720902. [Google Scholar] [CrossRef]
- Moverman, D.J.; Liberman, L.D.; Kraemer, S.; Corfas, G.; Liberman, M.C. Ultrastructure of Noise-Induced Cochlear Synaptopathy. Sci.Rep. 2023, 13, 19456. [Google Scholar] [CrossRef]
- Wang, M.; Xu, L.; Han, Y.; Wang, X.; Chen, F.; Lu, J.; Wang, H.; Liu, W. Regulation of Spiral Ganglion Neuron Regeneration as a Therapeutic Strategy in Sensorineural Hearing Loss. Front. Mol. Neurosci. 2022, 14, 829564. [Google Scholar] [CrossRef]
- Safina, D.R.; Surin, A.M.; Pinelis, V.G.; Kostrov, S.V. Effect of Neurotrophin-3 Precursor on Glutamate-induced Calcium Homeostasis Deregulation in Rat Cerebellum Granule Cells. J Neurosci Res. 2015, 93, 1865–1873. [Google Scholar] [CrossRef]
- Schonkeren, S.L.; Küthe, T.T.; Idris, M.; Bon-Frauches, A.C.; Boesmans, W.; Melotte, V. The Gut Brain in a Dish: Murine Primary Enteric Nervous System Cell Cultures. Neurogastroenterol. Motil. 2022, 34, e14215. [Google Scholar] [CrossRef]
- Vargova, I.; Kriska, J.; Kwok, J.C.F.; Fawcett, J.W.; Jendelova, P. Long-Term Cultures of Spinal Cord Interneurons. Front. Cell. Neurosci. 2022, 16, 827628. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, A.; Milivojev, N.; Hosmann, A.; Duvigneau, J.C.; Szabo, C.; Törö, G.; Rauter, L.; Vaglio-Garro, A.; Mkrtchyan, G.V.; Trofimova, L.; et al. Oxoglutarate Dehydrogenase Complex Controls Glutamate-Mediated Neuronal Death. Redox Biol. 2023, 62, 102669. [Google Scholar] [CrossRef] [PubMed]
- Cramer, T.M.L.; Tyagarajan, S.K. Protocol for the Culturing of Primary Hippocampal Mouse Neurons for Functional in Vitro Studies. STAR Protoc. 2024, 5, 102991. [Google Scholar] [CrossRef]
- Meas, S.J.; Nishimura, K.; Scheibinger, M.; Dabdoub, A. In Vitro Methods to Cultivate Spiral Ganglion Cells, and Purification of Cellular Subtypes for Induced Neuronal Reprogramming. Front. Neurosci. 2018, 12, 822. [Google Scholar] [CrossRef]
- Szabó, Z.; Harasztosi, C.; Szûcs, G.; Sziklai, I.; Rusznák, Z. A Detailed Procedure and Dissection Guide for the Isolation of Spiral Ganglion Cells of the Guinea Pig for Electrophysiological Experiments. Brain Res. Brain Res. Protoc. 2003, 10, 139–147. [Google Scholar] [CrossRef]
- Parker, M.; Brugeaud, A.; Edge, A.S.B. Primary Culture and Plasmid Electroporation of the Murine Organ of Corti. J. Vis. Exp. 2010, 1685. [Google Scholar]
- Grant, L.; Yi, E.; Goutman, J.D.; Glowatzki, E. Postsynaptic Recordings at Afferent Dendrites Contacting Cochlear Inner Hair Cells: Monitoring Multivesicular Release at a Ribbon Synapse. J. Vis. Exp. 2011, 2442. [Google Scholar]
- Montgomery, S.C.; Cox, B.C. Whole Mount Dissection and Immunofluorescence of the Adult Mouse Cochlea. J. Vis. Exp. 2016, 53561. [Google Scholar]
- May-Simera, H. Evaluation of Planar-Cell-Polarity Phenotypes in Ciliopathy Mouse Mutant Cochlea. J. Vis. Exp. 2016, 53559. [Google Scholar]
- Landegger, L.D.; Dilwali, S.; Stankovic, K.M. Neonatal Murine Cochlear Explant Technique as an In Vitro Screening Tool in Hearing Research. J. Vis. Exp. 2017, 55704. [Google Scholar]
- Sakamoto, A.; Kuroda, Y.; Kanzaki, S.; Matsuo, K. Dissection of the Auditory Bulla in Postnatal Mice: Isolation of the Middle Ear Bones and Histological Analysis. J. Vis. Exp. 2017, 55054. [Google Scholar]
- Fang, Q.-J.; Wu, F.; Chai, R.; Sha, S.-H. Cochlear Surface Preparation in the Adult Mouse. J. Vis. Exp. 2019, 10, 3791/60299. [Google Scholar]
- Ballesteros, A.; Swartz, K.J. Dextran Labeling and Uptake in Live and Functional Murine Cochlear Hair Cells. J. Vis. Exp. 2024, 10.3791/60769. [Google Scholar]
- Hahnewald, S.; Roccio, M.; Tscherter, A.; Streit, J.; Ambett, R.; Senn, P. Spiral Ganglion Neuron Explant Culture and Electrophysiology on Multi Electrode Arrays. J. Vis. Exp. 2016, 54538. [Google Scholar] [CrossRef] [PubMed]
- Schwieger, J.; Esser, K.-H.; Lenarz, T.; Scheper, V. Establishment of a Long-Term Spiral Ganglion Neuron Culture with Reduced Glial Cell Number: Effects of AraC on Cell Composition and Neurons. J. Neurosci. Methods 2016, 268, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Zhou, X.; Shi, L.; Xia, M.; Wang, Y.; Guo, N.; Hu, H.; Zhang, P.; Yang, H.; Zhu, F.; et al. Shikonin Attenuates Cochlear Spiral Ganglion Neuron Degeneration by Activating Nrf2-ARE Signaling Pathway. Front. Mol. Neurosci. 2022, 15, 829642. [Google Scholar] [CrossRef]
- Zhai, S.-Q.; Wang, D.-J.; Wang, J.-L.; Han, D.-Y.; Yang, W.-Y. Basic Fibroblast Growth Factor Protects Auditory Neurons and Hair Cells from Glutamate Neurotoxicity and Noise Exposure. Acta Otolaryngol. 2004, 124, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Anacker, A.; Esser, K.-H.; Lenarz, T.; Paasche, G. Purification of Fibroblasts From the Spiral Ganglion. Front. Neurol. 2022, 13, 877342. [Google Scholar] [CrossRef]
- Lie, M.; Grover, M.; Whitlon, D.S. Accelerated Neurite Growth from Spiral Ganglion Neurons Exposed to the Rho Kinase Inhibitor H-1152. Neuroscience 2010, 169, 855–862. [Google Scholar] [CrossRef]
- Machado, L.; Relaix, F.; Mourikis, P. Stress Relief: Emerging Methods to Mitigate Dissociation-Induced Artefacts. Trends Cell Biol. 2021, 31, 888–897. [Google Scholar] [CrossRef]
- Uniken Venema, W.T.C.; Ramírez-Sánchez, A.D.; Bigaeva, E.; Withoff, S.; Jonkers, I.; McIntyre, R.E.; Ghouraba, M.; Raine, T.; Weersma, R.K.; Franke, L.; et al. Gut Mucosa Dissociation Protocols Influence Cell Type Proportions and Single-Cell Gene Expression Levels. Sci. Rep 2022, 12, 9897. [Google Scholar] [CrossRef] [PubMed]
- Feltz, P.; Rasminsky, M. A Model for the Mode of Action of GABA on Primary Afferent Terminals: Depolarizing Effects of GABA Applied Iontophoretically to Neurones of Mammalian Dorsal Root Ganglia. Neuropharmacology 1974, 13, 553–563. [Google Scholar] [CrossRef]
- Sun, G.; Liu, W.; Fan, Z.; Zhang, D.; Han, Y.; Xu, L.; Qi, J.; Zhang, S.; Gao, B.T.; Bai, X.; et al. The Three-Dimensional Culture System with Matrigel and Neurotrophic Factors Preserves the Structure and Function of Spiral Ganglion Neuron In Vitro. Neural. Plasticity 2016, 2016, 1–15. [Google Scholar]
- Zuo, W.-Q.; Hu, Y.-J.; Yang, Y.; Zhao, X.-Y.; Zhang, Y.-Y.; Kong, W.; Kong, W.-J. Sensitivity of Spiral Ganglion Neurons to Damage Caused by Mobile Phone Electromagnetic Radiation Will Increase in Lipopolysaccharide-Induced Inflammation in Vitro Model. J. Neuroinflammation 2015, 12, 105. [Google Scholar] [CrossRef]
- Vieira, M.; Christensen, B.L.; Wheeler, B.C.; Feng, A.S.; Kollmar, R. Survival and Stimulation of Neurite Outgrowth in a Serum-Free Culture of Spiral Ganglion Neurons from Adult Mice. Hear. Res. 2007, 230, 17–23. [Google Scholar] [CrossRef]
- Whitlon, D.S.; Ketels, K.V.; Coulson, M.T.; Williams, T.; Grover, M.; Edpao, W.; Richter, C.P. Survival and Morphology of Auditory Neurons in Dissociated Cultures of Newborn Mouse Spiral Ganglion. Neuroscience 2006, 138, 653–662. [Google Scholar] [CrossRef]
- Kaiser, O.; Paasche, G.; Stöver, T.; Ernst, S.; Lenarz, T.; Kral, A.; Warnecke, A. TGF-Beta Superfamily Member Activin A Acts with BDNF and Erythropoietin to Improve Survival of Spiral Ganglion Neurons in Vitro. Neuropharmacology 2013, 75, 416–425. [Google Scholar] [CrossRef]
- Schwieger, J.; Warnecke, A.; Lenarz, T.; Esser, K.-H.; Scheper, V. Neuronal Survival, Morphology and Outgrowth of Spiral Ganglion Neurons Using a Defined Growth Factor Combination. PLoS One 2015, 10, e0133680. [Google Scholar] [CrossRef]
- Hegarty, J.L.; Kay, A.R.; Green, S.H. Trophic Support of Cultured Spiral Ganglion Neurons by Depolarization Exceeds and Is Additive with That by Neurotrophins or cAMP and Requires Elevation of [Ca2+ ]i within a Set Range. J. Neurosci. 1997, 17, 1959–1970. [Google Scholar] [CrossRef]
- Harada, N.; Han, D.-Y.; Komeda, M.; Yamashita, T. Glutamate-Induced Intracellular Ca2+ Elevation in Isolated Spiral Ganglion Cells of the Guinea Pig Cochlea. Acta Otolaryngol. 1994, 114, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Zhang, C.; Chen, A.; Liu, W.; Li, J.; Sun, Q.; Wang, H. Protective Effect of Edaravone on Glutamate-Induced Neurotoxicity in Spiral Ganglion Neurons. Neural. Plasticity 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Han, D.-Y.; Yamashita, T.; Harada, N.; Kumazawa, T. Calcium Mobilization in Isolated Cochlear Spiral Ganglion Cells of the Guinea Pig. Acta Otolaryngol. 1993, 113, 26–29. [Google Scholar] [CrossRef]
- Ding, Z.-J.; Chen, X.; Tang, X.-X.; Wang, X.; Song, Y.-L.; Chen, X.-D.; Wang, J.; Wang, R.-F.; Mi, W.-J.; Chen, F.-Q.; et al. Apoptosis-Inducing Factor and Calpain Upregulation in Glutamate-Induced Injury of Rat Spiral Ganglion Neurons. Mol. Med. Rep. 2015, 12, 1685–1692. [Google Scholar] [CrossRef]
- Xiao, H.; Yang, C.; He, Y.; Zheng, N. Neurotoxicity of Quinolinic Acid to Spiral Ganglion Cells in Rats. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2010, 30, 397–402. [Google Scholar] [CrossRef]
- Sun, F.; Zhou, K.; Tian, K. Y.; Zhang, X. Y.; Liu, W.; Wang, J.; Zhong, C. P.; Qiu, J. H.; Zha, D. J. Atrial Natriuretic Peptide Promotes Neurite Outgrowth and Survival of Cochlear Spiral Ganglion Neurons In vitro Through NPR-A/cGMP/PKG Signaling. Front Cell Dev Biol. 2021, 9, 681421. [Google Scholar] [CrossRef]
- Wright, T.; Gillespie, L.N.; O’Leary, S.J.; Needham, K. Firing Frequency and Entrainment Maintained in Primary Auditory Neurons in the Presence of Combined BDNF and NT3. Sci. Rep. 2016, 6, 28584. [Google Scholar] [CrossRef] [PubMed]
- Brewer, G.J. Serum-free B27/Neurobasal Medium Supports Differentiated Growth of Neurons from the Striatum, Substantia Nigra, Septum, Cerebral Cortex, Cerebellum, and Dentate Gyrus. J. Neurosci. Res. 1995, 42, 674–683. [Google Scholar] [CrossRef]
- Xie, C.; Markesbery, W.R.; Lovell, M.A. Survival of Hippocampal and Cortical Neurons in a Mixture of MEM+ and B27-Supplemented Neurobasal Medium. Free Radic. Biol. Med. 2000, 28, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.H.; Ngwainmbi, J.; Grider, J.R.; Dewey, W.L.; Akbarali, H.I. An In-Vitro Preparation of Isolated Enteric Neurons and Glia from the Myenteric Plexus of the Adult Mouse. J. Vis. Exp. 2013, 50688. [Google Scholar] [PubMed]
- Vincent, P.F.Y.; Young, E.D.; Edge, A.S.B.; Glowatzki, E. Auditory Hair Cells and Spiral Ganglion Neurons Regenerate Synapses with Refined Release Properties in Vitro. Proc. Natl. Acad. Sci. U.S.A. 2024, 121, e2315599121. [Google Scholar] [CrossRef]
- Lefebvre, P.P.; Weber, T.; Leprince, P.; Rigo, J.-M.; Delre’e, P.; Rogister, B.; Moonen, G. Kainate and NMDA Toxicity for Cultured Developing and Adult Rat Spiral Ganglion Neurons: Further Evidence for a Glutamatergic Excitatory Neurotransmission at the Inner Hair Cell Synapse. Brain Res. 1991, 555, 75–83. [Google Scholar] [CrossRef]
- Zheng, J.; Stewart, R.; Gao, W. Neurotrophin-4/5 Enhances Survival of Cultured Spiral Ganglion Neurons and Protects Them from Cisplatin Neurotoxicity. J. Neurosci. 1995, 15, 5079–5087. [Google Scholar] [CrossRef]
- Bailey, E.M.; Green, S.H. Postnatal Expression of Neurotrophic Factors Accessible to Spiral Ganglion Neurons in the Auditory System of Adult Hearing and Deafened Rats. J. Neurosci. 2014, 34, 13110–13126. [Google Scholar] [CrossRef]
- Needham, K.; Nayagam, B.A.; Minter, R.L.; O’Leary, S.J. Combined Application of Brain-Derived Neurotrophic Factor and Neurotrophin-3 and Its Impact on Spiral Ganglion Neuron Firing Properties and Hyperpolarization-Activated Currents. Hear. Res. 2012, 291, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Balazs, R. Trophic Effect of Glutamate. Curr. Top. Med. Chem. 2006, 6, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zou, Q.; Zhang, J. Vincamine exerts protective effect on spiral ganglion neurons in endolymphatic hydrops guinea pig models. Am. J. Transl. Res. 2018, 10, 3650–3663. [Google Scholar]
- McDonald, J. W.; Johnston, M. V. Physiological and pathophysiological roles of excitatory amino acids during central nervous system development. Brain Res. Brain Res. Rev. 1990, 15, 41–70. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.C.; Appler, J.M.; Houseman, E.A.; Goodrich, L.V. Developmental Profiling of Spiral Ganglion Neurons Reveals Insights into Auditory Circuit Assembly. J. Neurosci. 2011, 31, 10903–10918. [Google Scholar] [CrossRef]
- Lu, J.; Liu, H.; Lin, S.; Li, C.; Wu, H. Electrophysiological Characterization of Acutely Isolated Spiral Ganglion Neurons in Neonatal and Mature Sonic Hedgehog Knock-in Mice. Neurosci. Lett. 2020, 714, 134536. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Ohmori, H. Voltage-gated and Chemically Gated Ionic Channels in the Cultured Cochlear Ganglion Neurone of the Chick. J. Physiol. 1990, 420, 185–206. [Google Scholar] [CrossRef]
- Ding, Z. J.; Chen, X.; Tang, X. X.; Wang, X.; Song, Y. L.; Chen, X. D.; Mi, W. J.; Wang, J.; Lin, Y.; Chen, F. Q.; Qiu, J. H. Calpain inhibitor PD150606 attenuates glutamate induced spiral ganglion neuron apoptosis through apoptosis inducing factor pathway in vitro. PloS One. 2015, 10, e0123130. [Google Scholar] [CrossRef] [PubMed]
- Carricondo, F.; Bartolomé, M. V.; Vicente-Torres, M. A.; Fernández-Pacheco, P.; Rodríguez, T.; Gil-Loyzaga, P. Sensitivity to glutamate neurotoxicity in different developmental periods of the rat cochlea. Adv. Otorhinolaryngol. 2002, 59, 91–95. [Google Scholar] [PubMed]
- Frisch, S.; Francis, H. Disruption of Epithelial Cell-Matrix Interactions Induces Apoptosis. J. Cell Biol. 1994, 124, 619–626. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Conditions for enzymatic dissociation included in protocols for studying glutamatergic system on primary SGCs. Missing data are presented by the symbol “NA”.
Table 1.
Conditions for enzymatic dissociation included in protocols for studying glutamatergic system on primary SGCs. Missing data are presented by the symbol “NA”.
| Study | Animal species and age | Enzyme type | Enzyme concentration | units | Duration [min] | Temperature [˚C] |
|---|---|---|---|---|---|---|
|
Yamaguchi and Ohmori, 1990 [100] |
Chick embryo (16-19 embryonic days) |
Collagenase Papain |
0.3 10 |
units/ml units/ml |
30 25 |
37 |
|
Lefebvre et al., 1991 [91] |
Rats (P5) |
Trypsin Collagenase DNAse |
0.1 0.l 0.01 |
% % % |
25 | 37 |
| Adult rats | Collagenase Trypsin |
5 0.1 |
% % |
20 17 |
||
| Nakagawa et al., 1991 [28] |
Adult guinea pigs |
Collagenase or Dispase |
0,5 500 |
mg/ml IU/ml |
30-60 30 |
31 |
| Chickens from 2 to 5 wk post-hatch of either sex | Collagenase Trypsin type IX or Dispase |
1.0 0.5 1000 |
mg/ml mg/ml IU/ml |
30 30 60-90 |
||
| Harada et al., 1994 [80] |
Adult guinea pigs | Collagenase | 0.1 | mg/ml | 30 | NA |
| Shimozono et al., 1995 [29] |
Adult guinea pigs | Collagenase or Dispase |
0.5 500 |
mg/ml IU/ml |
30-60 30 |
31 |
| Peng et al., 2004 [12] |
Mice (P0, adult) |
Collagenase type IV Trypsin |
0.5 2.5 |
mg/ml mg/ml |
30 + 30 |
On ice 37 |
| Zhai et al., 2004 [67] |
Mice (P3) |
Collagenase Trypsin |
0.25 0.25 |
% % |
25 | 37 |
| Chen et al., 2007 [35] |
Mice (P6-8) |
Collagenase type IV Trypsin |
0.5 2.5 |
mg/ml mg/ml |
25 | 37 |
| Chen et al., 2009 [36] |
Mice (P3-5) |
Collagenase type IV Trypsin |
0.5 2.5 |
mg/ml mg/ml |
25 | 37 |
| Xiao et al., 2010 [84] |
Rats (P3-7) |
Trypsin | NA | NA | 8 + 6 |
37 low temperature |
| Ding et al., 2015 [101] |
Rats (P0-3) |
Trypsinase | 0.125 | % | 15 | 37 |
|
Bai et al., 2016 [81] |
Rats (P<5) |
Trypsinase | 0.125 | % | 15 | 37 |
|
Li et al., 2018 [96] |
Rats (embryonic day-18) |
Trypsin | 1.25 | mg/ml | 10 | 37 |
| Sun et al., 2021 [85] |
Rats (P3) |
Collagenase type IV Trypsin |
0.1 0.25 |
% % |
20 | 37 |
| Wang et al., 2021 [17] |
Rats (P1-P4) |
Collagenase type IV Trypsin |
0.5 2.5 |
mg/ml mg/ml |
60 | 37 |
Table 2.
Key methodological parameters of primary spiral ganglion cell cultures used in glutamate signaling research. Note: Missing data are indicated by “NA”.
Table 2.
Key methodological parameters of primary spiral ganglion cell cultures used in glutamate signaling research. Note: Missing data are indicated by “NA”.
| Study | Animal species and age | Long-term/short term culture | Coating substrate | Medium and supplements | Pre- experimental cultivation time/ Maximum incubation period |
|---|---|---|---|---|---|
| Yamaguchi and Ohmori, 1990 [100] |
Chick embryo (16-19 embryonic days) |
Long-term culture | Collagen Poly-D-lysine |
DMEM+F12 calf serum |
5-14 days/5-14 days |
| Short- term culture | Poly-D-lysine/ concanavalin A |
DMEM+F12 calf serum |
<1 day/ <1 day | ||
| Lefebvre et al., 1991 [91] |
Rats (P5) |
Long- term culture | Poly-ornithine; Laminin; |
DMEM+ N1 cocktail: bovine insulin, progesterone, putrescine, transferrin, selenium |
5 days/6 days |
| Adult rats | Long-term culture | Poly-ornithine; Laminin; Astrocyte-conditioned medium |
DMEM+ N1 cocktail: bovine insulin, progesterone, putrescine, transferrin, selenium |
3 days/4 days | |
| Peng et al., 2004 [12] |
Mice (P0, adult) |
Long-term culture | NA |
DMEM+ FBS penicillin streptomycin |
2 days/ 2 days and more |
| Zhai et al., 2004 [67] |
Mice (P3) |
Long-term culture | Rat-tail collagen | Not fully specified | 24 h/14 days |
| Chen et al., 2007 [35] |
Mice (P6-P8) |
Short-term culture | Gelatin |
DMEM+ F12 FBS, horse serum, NT-3, BDNF, penicillin and streptomycin |
15-18 h/ 15-18 h + ~1 h |
| Chen et al., 2009 [36] |
Mice (P3-P5) |
Short-term culture | Poly-L-ornithine |
DMEM+ F12 FBS, horse serum, NT-3, BDNF, B-27 supplement, penicillin and streptomycin |
15-18 h/15-18 h+ ~24h |
| Xiao et al., 2010 [84] |
Rats (P3-7) |
Long-term culture | NA |
NBM B27 AraC |
72 h/96 h |
| Ding et al., 2015 [101] |
Rats (P1) |
Long-term culture | NA |
DMEM B27, BDNF, penicillin |
24 h/72 h |
| Ding et al., 2015 [83] |
Rats (P0-3) |
Long-term culture | NA |
DMEM B27, BDNF, penicillin |
NA |
| Bai et al., 2016 [81] |
Rats (<P5) |
Short-term culture | Poly-L-lysine |
DMEM 10% FBS |
NA |
| Li et al., 2018 [96] |
Rats, (embryonic day-18) | Short-term culture | Poly-L-lysine |
DMEM 10% FBS |
NA |
| Sun et al., 2021 [85] |
Rats (P3) |
Long-term culture | Poly-L-lysine | NA |
4h/5 days |
Table 3.
Summary of Experimental Studies on Glutamate Receptor Agonists and Antagonists in Spiral Ganglion Neurons. Note: Missing data are indicated by “NA”.
Table 3.
Summary of Experimental Studies on Glutamate Receptor Agonists and Antagonists in Spiral Ganglion Neurons. Note: Missing data are indicated by “NA”.
| Study | Animal species and age | Methods | Applications | Glu receptors agonists (concentration) | Glu receptors antagonists (concentration) |
|
|---|---|---|---|---|---|---|
| Yamaguchi and Ohmori, 1990 [100] |
Chick embryo (16-19 embryonic days) | Patch-clamp | Measurement of ionic currents | Glu (30 µM) KA (100 µM) Asp (100 µM) NMDA (100 µM) |
APV (100 µM) | |
| Fluorescent microscopy |
Cell morphology | |||||
| Lefebvre et al., 1991 [91] |
Rats (P5); |
Immunohistochemical staining | Survival assessment; Cell morphology |
Glu 10-8 - 10-3 M NMDA 10-8 - 10-3 M Kainic acid 10-8 - 10-3 M Quisqualic acid 10-8 - 10-3 M |
DAP-5 (10-9 -10-3 M) Kyn (10-9-10-6 M) |
|
| Adult rats |
Glu (10-4 M) NMDA (10-4 M) Kainic acid (10-4 M) Quisqualic acid (10-4 M) |
DAP-5 (10-4 M) Kyn (10-4 M) |
||||
| Nakagawa et al., 1991 [28] |
Adult guinea pigs; |
Patch-clamp |
Measurement of ionic currents |
Glu (3х10-6 - 10-2 M) QA (3x10-7- 3x10-4 M) KA (3х10-6-3х10-4 M) Asp (3x10-3 M) NMDA (3x10-3 M) |
CNQX (10-8-10-5 M); DNQX (10-8-10-5 M); diCl-HQC (10-7-10-4 M); Kyn (3x10-6-3x10-3 M); APV (3х10-5 M) |
|
|
Phase-contrast microscopy |
Cell morphology | |||||
| Chickens from 2 to 5 wk post-hatch | Glu (3х10-4 M) QA (3х10-5 M) KA (10-4 M) Asp (10-3 M) NMDA (3x10-3 M) |
APV (3х10-5 M) | ||||
| Harada et al., 1994 [80] |
Adult guinea pigs | Calcium imaging | Intracellular calcium [Ca²⁺]i concentration measurement | Glu (100 µM) | NA | |
|
Phase-contrast microscopy |
Cell morphology | |||||
| Shimozono et al., 1995 [29] |
Adult guinea pigs | Patch-clamp | Measurement of ionic currents | Glu (10, 50, 100 µM) KA (100 µM); NMDA (100 µM); QA (100 µM) |
APV (NA); |
|
| Calcium imaging | Intracellular calcium [Ca²⁺]i concentration measurement | |||||
| Peng et al., 2004 [12] |
Mice (P0, adult) |
Patch-clamp | Measurement of ionic currents | KA (100 µM); Glu (300 µM) DHPG (100 µM); ACPD (100 µM) |
AIDA (300 µM) DNQX (200 µM) |
|
| Calcium imaging | Intracellular calcium [Ca²⁺]i concentration measurement | |||||
| Zhai et al., 2004 [67] |
Mice (P3) |
Histochemical staining | Survival assessment; Neurite length measurement |
Glu (20 mM) | NA | |
| Chen et al., 2007 [35] |
Mice (P6-P8) |
Immunofluorescence staining | Assessment of surface GluR2 receptors expression levels; Viability assessment |
Glu (20 µM) AMPA (20 µM) NMDA (20 µM) |
DNQX (20 µM) APV (50 µM) |
|
| Chen et al., 2009 [36] |
Mice (P3-P5) |
Immunofluorescence staining | Assessment of surface GluR2 receptors expression levels; Viability assessment |
AMPA (20 µM) NMDA (20 µM) AMPA (300 µM) NMDA (300 µM) |
NA | |
| Xiao et al., 2010 [84] |
Rats (P3-7) |
Fluorescent microscopy |
Cell morphology; Detection of apoptosis |
Quinolinic acid (100; 1000 µM/L) |
MK-801 (20 µM/L) | |
| Laser confocal microscopy | Intracellular calcium [Ca²⁺]i concentration measurement | |||||
| Ding et al., 2015 [101] |
Rats (P1) |
Stereoscopic microscopy |
Cell morphology |
Glu (20 µM) | NA | |
|
Immunofluorescence staining; |
AIF distribution |
|||||
| TUNEL assay | Apoptotic cells counting | |||||
| Western Blot | Assessment of AIF and calpain expression Levels |
|||||
| RT-PCR | Assessment of mRNA expression of AIF, calpain and caspase-3 |
|||||
| Ding et al., 2015 [83] |
Rats (P0-3) |
Immunofluorescence staining | AIF distribution |
Glu (10 mM, 20 mM, 40 mM) | NA | |
| RT-PCR | Assessment of mRNA expression of AIF, calpain and caspase-3 |
|||||
| Bai et al., 2016 [81] |
Rats (P<5) |
MTT assay | Cell viability assessment |
Glu (2 mM) | NA | |
| Trypan blue staining | Cell viability assessment |
|||||
| Ho.33342 and Propidium iodide double staining | Detection of apoptosis and necrosis | |||||
| Spectrophotometry | Detection of GSH content, SOD activity, MDA level | |||||
| Western Blot | Assessment of AKT, p-AKT, Bax, Bcl-2 genes expression levels | |||||
| Li et al., 2018 [96] |
Rats embryo (18 embryonic days) | MTT assay |
Cell viability assessment | Glu (2 mM) | NA | |
| Ho.33342 and Propidium iodide double staining | Detection of apoptosis and necrosis | |||||
| Sun et al., 2021 [85] |
Rats (P3) | Immunofluorescence staining | Neuronal number and neurite length measurement |
Glu (100 µM) | NA | |
| Wang et al., 2021 [17] |
Rats (P1-P4) |
Calcium imaging | Intracellular calcium [Ca²⁺]i concentration measurement | AMPA (100 µM) Glu (100 µM) NMDA (100 µM) Glu (100 µM) + D-serine (100 µM) |
7-CKA (50 µM) D-APV (50 µM) GYKI 53784 (40 µM) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



