Submitted:
18 February 2026
Posted:
26 February 2026
You are already at the latest version
Abstract
Colorectal cancer (CRC) remains a leading cause of cancer-related morbidity and mortality worldwide, with obesity recognized as a major modifiable risk factor. Obesity-associated CRC is characterized by systemic low-grade inflammation, altered lipid metabolism, and gut microbial dysbiosis, all of which converge to create a tumor-permissive environment. Emerging evidence implicates miR-101a, the murine homolog of human miR-101, as a key molecular mediator linking metabolic dysfunction, microbial metabolites, and epithelial homeostasis. Traditionally regarded as a tumor suppressor by repressing oncogenes such as EZH2, MCL-1, and COX-2, miR-101a appears to exhibit a paradoxical tumor-promoting role in obese colon. Elevated dietary and microbially derived ethanolamine induces miR-101a overexpression in colonic epithelial cells. This aberrant induction impairs goblet cell differentiation and reduces MUC2 mucin biosynthesis, disrupting the mucus barrier and leading to a “leaky gut” phenotype. Barrier dysfunction permits microbial translocation and endotoxemia, which activate NF-κB-driven inflammatory cascades and amplify COX-2 signaling. The resulting chronic inflammation promotes epithelial proliferation, generates mutagenic reactive oxygen species, and activates pro-survival pathways such as STAT3 and AKT, collectively driving adenoma initiation and progression. Importantly, this ethanolamine-miR-101a axis represents a novel mechanistic link between diet, the microbiota, and cancer biology. Translationally, miR-101a holds promise as a biomarker of early barrier dysfunction and CRC risk, detectable in tissue, serum, or fecal samples. Furthermore, microbiome-targeted interventions, dietary modifications, or direct inhibition of miR-101a may offer innovative therapeutic strategies. Understanding the context-dependent duality of miR-101a underscores the complexity of miRNA biology and highlights its potential as a target for preventing obesity-driven CRC.
Keywords:
ethanolamine
; obesity
; T2D
; colorectal cancer
; chronic inflammation
; microRNA
; leaky gut
1. Introduction
Colorectal cancer (CRC) remains a leading cause of cancer mortality worldwide. Widespread screening and advances in systemic treatment have broadly improved survival, the risk of CRC and cancer-related death remains elevated in certain populations. For instance, CRC incidence is rising in patients under 50 years old [1] and is substantially elevated in obese compared to non-obese individuals [2]. The underlying pathobiology of increased cancer incidence in these populations remains unclear, but recent work implicates alterations in the gut microbiome induced by a Western-style diet [3,4,5]. This review outlines a pathway linking diet and the gut microbiome to the regulation of gene expression by MicroRNAs (miRNAs) in the tumor microenvironment and may represent a novel therapeutic target in CRC.
MicroRNAs (miRNAs) are a class of small, non-coding RNAs approximately 20-24 nucleotides in length that function as critical regulators of post-transcriptional gene expression [6]. They primarily exert their effect through base-pairing with complementary sequences in the 3′ untranslated region (3′ UTR) of target messenger RNAs (mRNAs), leading to mRNA degradation or translational repression [7]. By fine-tuning protein expression, miRNAs regulate diverse cellular processes, including cell proliferation, differentiation, apoptosis, immune regulation, and stress adaptation [8,9]. Importantly, aberrant miRNA expression or activity has been recognized as a hallmark of cancer biology, contributing to both tumor initiation and progression. Some miRNAs function as tumor suppressors by restraining oncogenic pathways, while others function as oncomiRs, promoting tumor growth, metastasis, or therapeutic resistance. Notably, certain miRNAs exhibit context-dependent dual roles, functioning as tumor suppressors in one tissue or cellular environment and as tumor promoters in another [10,11,12]. miR-101a, the murine homolog of human miR-101, is encoded within conserved genomic loci and is ubiquitously expressed across several tissues, including the intestinal epithelium, liver, brain, and immune compartments [13,14,15,16,17,18]. Historically, miR-101a has been characterized as a tumor suppressor. One of its canonical targets is Enhancer of Zeste Homolog 2 (EZH2), the catalytic component of the Polycomb Repressive Complex 2 (PRC2), which mediates trimethylation of histone H3 lysine 27 (H3K27me3), leading to transcriptional silencing of tumor suppressor genes [14,19,20,21,22]. Loss or downregulation of miR-101a, therefore, results in EZH2 overexpression, enhanced histone methylation, and widespread suppression of genes critical for maintaining epithelial differentiation and genomic stability [22,23,24,25]. In prostate, breast, liver, and brain cancers, decreased expression of miR-101 has been consistently associated with increased tumor aggressiveness, angiogenesis, and poor prognosis [26,27]. Other validated targets of miR-101a include MCL-1 (myeloid cell leukemia-1), an anti-apoptotic member of the B-cell lymphoma (BCL)-2 family; cyclooxygenase-2 (COX-2), a pro-inflammatory enzyme implicated in carcinogenesis; and DNA methyltransferases (DNMT)3A/B, which regulate DNA methylation patterns in cancer [28,29]. Collectively, these studies have established miR-101a as a guardian against uncontrolled proliferation, epigenetic silencing of tumor suppressors, and evasion of apoptosis, functioning predominantly as a tumor suppressor under physiological conditions.
Despite its well-documented tumor-suppressive role, accumulating evidence suggests that miR-101a can paradoxically promote tumor growth in specific pathological contexts, particularly those characterized by metabolic dysregulation, chronic inflammation, and gut barrier dysfunction [30,31,32,33,34]. A notable example emerges in obesity and/or type-2 diabetes (T2D)-associated colorectal cancer (CRC). Obesity/T2D is characterized by systemic low-grade inflammation, altered lipid metabolism, insulin resistance, and significant changes in gut microbiome composition (dysbiosis) [35,36,37,38,39,40,41,42]. The microenvironment of CRC is saturated with pro-inflammatory lipid mediators and deficient in pro-resolving mediators, creating dysregulated, non-resolving inflammation [39,43]. These alterations collectively create a microenvironment conducive to colorectal initiation and progression. In this context, miR-101a expression has been observed to increase rather than decrease, with downstream effects that deviate markedly from its classical tumor-suppressive functions. The gut microbiota, heavily shaped by dietary patterns, is increasingly recognized as a pivotal regulator of host miRNA expression. Diets enriched in fat and red meat increase luminal ethanolamine concentrations, a membrane phospholipid-derived metabolite that serves as a carbon and nitrogen source for many gut bacteria [44,45]. Elevated intestinal ethanolamine has been identified as a critical inducer of miR-101a expression in the colonic epithelium [33,46]. Mechanistically, ethanolamine may act both directly, through epithelial signaling pathways, and indirectly, by selecting for bacterial taxa that produce metabolites influencing host gene expression [47]. Dysbiotic microbiota in obesity may amplify this effect, establishing an ethanolamine miR-101a axis that links diet, microbial metabolism, and host transcriptomic regulation.
Together, this evidence positions miR-101a as both a guardian and a potential accomplice in tumorigenesis, with its role determined by the surrounding metabolic-microbial landscape. This paradox offers a novel framework connecting obesity, gut dysbiosis, and colorectal cancer pathogenesis.
1.1. Genomic Organization and Sequence Features of the miR-101 Family
In mammals, the miR-101 family is encoded by two conserved loci that produce the same predominant mature effector strands, miR-101-3p/5p (5′-UACAGUACUGUGAUAACUGAA-3′) [48]. In humans, the precursors map to miR-101-1 on chr1p31.3 and miR-101-2 on chr9p24.1; in mice, the orthologues loci correspond to miR-101a on chr4 (negative strand; GRCm39: ~101,204,142-101,204,224) and miR-101b on chr19 (positive strand; ~29,112,679-29,112,775) [49,50,51,52,53]. Within the human genome, miR-101 derives from two primary precursors, miR-101-1 (75 bp) and miR-101-2 (79 bp), both of which are essential for its biogenesis. In mice, miR-101b is embedded within an intron of RNA Terminal Phosphate Cyclase-Like (Rcl)1, whereas miR-101a is intergenic, an arrangement that likely contributes to subtle differences in transcriptional control and co-regulation with host-gene programs [49]. Biogenesis follows the canonical microRNA processing pathway: RNA polymerase II transcription, nuclear cropping by Drosha-DiGeorge Critical Region (DGCR8), cytoplasmic cleavage by Dicer, and Argonaute loading into the RNA-Induced Silencing Complex (RISC) [54,55]. Although both arms are detectable, most tissues preferentially load the 3p arm, establishing miR-101-3p as the principal functional strand for target repression. Sequence variation introduces additional regulatory complexity. Single-nucleotide variants within the hairpin (e.g., basal junction, apical loop, dicer processing sites) can shift microprocessor/dicer efficiency, thereby affecting mature miRNA abundance and 5p/3p arm selection [56,57,58]. Variants within the seed regions (nts 2-8 of miR-101-3p) are especially consequential, as they redefine the targetome, simultaneously extinguishing canonical sites (e.g., in Enhancer of Zeste Homolog [EZH]2 or Post-Transcriptional Gene Silencing [PTGS]2/ Cyclooxygenase [COX]-2) and creating novel interactions in unrelated transcripts [59,60]. Population resources catalog multiple variants in and around miR101-1/-2; some of which have been associated with altered cancer risk, underscoring their potential functional impact [54]. Complementing miRNA-centric variation, 3′-UTR polymorphisms (“miR-eQTLs”) within target genes can gain or lose miR-101 recognition motifs, stratifying repression across individuals, tissues, and life developmental stages.
Expression atlases reveal a broad, developmentally dynamic abundance of miR-101-3p. In the mouse brain, miR-101a/b levels rise from late embryogenesis (~E16) through early postnatal stages (~P12), consistent with roles in neuronal maturation and circuit refinement [61]. In adults, miR-101 is readily detected across epithelial, stromal, and immune compartments, with absolute levels tuned by hormonal, inflammatory, and metabolic cues [53,61,62,63]. Functionally, miR-101-3p converges on regulators of chromatin state, eicosanoid/inflammatory tone, cell survival, and cytoskeletal dynamics-with repeatedly validated targets including EZH2, Myeloid Cell Leukemia (MCL)1, PTGS2/COX-2, and context-dependent effectors such as FBJ Murine Osteosarcoma Viral Oncogene (FOS), Stathmin (STMN)1, DNMT3A, Ras-related C3 botulinum toxin substrate (RAC)1, SRY (Sex-determining Region Y)-Box (SOX)9, and Cyclin-Dependent Kinase (CDK)8 [59,64,65,66,67,68,69]. These interactions typically impose anti-proliferative and anti-inflammatory constraints in epithelial tissues. However, tissue state (e.g., obesity-associated inflammation, cytokine milieu, metabolite availability) can invert net outcomes by reshaping competing RNA networks and transcriptional baselines. Aberrant expression of miR-101a exerts profound influence on genomic stability by targeting multiple classes of genes involved in DNA repair, chromatin regulation, inflammation, and barrier integrity [23,33,70,71,72]. At the level of the DNA damage response (DDR), miR-101a directly represses ataxia-telangiectasia mutated (ATM) and Protein Kinase, DNA-Activated, Catalytic Subunit (PRKDC [DNA-PKcs]), impairing double-strand break repair and thereby sensitizing epithelial cells to DNA damage and mutational accumulation [73,74]. In parallel, miR-101a regulates chromatin modifiers such as EZH2, Disruptor of Telomeric Silencing 1-Like (DOT1L), and DNMT3A/3B, leading to altered histone methylation and DNA methylation landscapes that promote epigenetic instability and oncogenic transcriptional reprogramming [28,75,76,77,78,79]. Cytoskeletal fidelity is also influenced through repression of STMN1, which disrupts microtubule dynamics and predisposes cells to aneuploidy [80,81,82]. In obesity-associated CRC, these molecular effects converge with barrier dysfunction, where miR-101a-driven defects in goblet cell differentiation and mucin secretion reduce Mucin (MUC)2 levels, permitting microbial translocation and activation of Nuclear Factor kappa-light-chain-enhancer of activated B cells (NF-κB)/ Interleukin (IL)-6/ (Signal Transducer and Activator of Transcription) STAT3 signaling [83,84,85,86,87]. This inflammatory loop sustains the production of reactive oxygen and nitrogen species (ROS/RNS), amplifying DNA damage and accelerating the clonal expansion of mutated epithelial cells. Collectively, these findings position miR-101a as a context-dependent regulator of tumorigenesis that does not directly induce point mutations but instead establishes a mutagenic microenvironment by suppressing DNA repair, promoting epigenetic deregulation, driving prostaglandin-mediated inflammation, and disrupting barrier function, ultimately promoting colorectal tumor initiation and progression.
Induction of miR-101a by Metabolic and gut barrier dysregulation
The tumor-promoting role of miR-101a is closely linked to diet and metabolites generated by the gut microbiota [33]. Recent investigations reveal that intestinal ethanolamine, a phospholipid precursor derived from dietary phosphatidylethanolamine and abundant in high-fat and high-meat diets [44,88,89,90,91,92,93], plays a central role in driving miR-101a expression. Ethanolamine is a preferential nutrient source for several gut bacterial taxa, including opportunistic pathogens and dysbiosis-associated organisms enriched during dysbiosis [94,95,96,97,98,99,100,101]. In the context of obesity and metabolic syndrome, dysbiotic microbiota increases the availability of ethanolamine within the colonic lumen [33,96,102,103,104,105]. This metabolite acts as more than a passive nutrient and functions as a signaling molecule linking diet to host transcriptional regulation [33,106]. Elevated ethanolamine stimulates miR-101a upregulation in colonic epithelial cells, thereby altering the delicate balance between epithelial homeostasis and injury responses. Unlike the canonical tumor-suppressive functions of miR-101a observed in non-obese conditions, chronic upregulation in obesity skews epithelial biology toward barrier dysfunction and downstream neoplasia as described below [33]. This metabolic induction of miR-101a positions it as a molecular bridge connecting nutrition, dysbiosis, and tumor initiation.
One of the earliest and most consequential downstream effects of miR-101a upregulation is its interference with goblet cell differentiation and mucin biosynthesis. Goblet cells are specialized epithelial cells responsible for secreting MUC2, the major structural mucin forming the intestinal mucus layer. This mucus barrier functions as the first line of defense, physically separating luminal microbes from the epithelial surface [107,108,109,110]. Overexpression of miR-101a represses transcriptional regulators that orchestrate goblet cell maturation and MUC2 production [67,111,112]. Consequently, MUC2 expression is reduced, leading to a thinner, discontinuous mucus layer. This structural defect compromises mucosal protection and allows bacteria and microbial-associated molecular patterns (MAMPs) to approach the epithelium [113,114,115,116,117]. This barrier disruption, often termed as “leaky gut”, represents a critical initiating event in CRC pathogenesis [118,119,120]. Barrier impairment creates a permissive environment for microbial translocation, allowing endotoxins, such as lipopolysaccharide (LPS), to cross into the lamina propria and the systemic circulation [121,122,123,124,125]. The resulting endotoxemia perpetuates immune activation and contributes to systemic low-grade inflammation, a hallmark of obesity-associated diseases [126,127,128,129,130,131]. The detailed schematic diagram showing obesity-driven colorectal cancer development is shown in Figure 1. Moreover, the loss of mucin alters the spatial distribution of the microbiota, promoting closer interactions between microbes and the epithelium, which further exacerbate dysbiosis [110,125,132,133,134]. Thus, miR-101a-mediated suppression of mucin biosynthesis dismantles a key protective mechanism of the gut, transforming the mucosal interface from a defensive barrier into a zone of persistent microbial challenge.
1.2. Chronic Inflammation as a Driver of Malignant Transformation via miR-101a
The epithelial barrier defects induced by miR-101a create fertile ground for sustained inflammation, which in turn may accelerate tumor initiation and progression. Microbial translocation engages pattern recognition receptors (PRRs) such as Toll-like receptors (TLRs) on epithelial and immune cells, triggering activation of the NF-κB pathway [135,136,137]. This leads to robust transcription of pro-inflammatory cytokines, including Tumor Necrosis Factor (TNF)-α, IL-6, and IL-1β, which amplify local and systemic immune responses [138,139,140]. Likewise, miR-101a overexpression directly interacts with inflammatory mediators by enhancing COX-2 expression, a critical enzyme in prostaglandin synthesis [141,142]. Elevated COX-2 activity promotes angiogenesis, inhibits apoptosis, and fosters immune evasion, thereby contributing to a tumor-permissive environment [143,144]. Furthermore, chronic inflammation stimulates the production of ROS/RNS by infiltrating immune cells [145,146,147]. These reactive ROS/RNS molecules induce DNA damage and mutagenic stress in epithelial cells, thereby accelerating the accumulation of oncogenic mutations [148,149,150,151,152,153,154]. This chronic inflammatory state not only sustains epithelial injury but also promotes cycles of injury and regenerative proliferation, further increasing the probability of malignant transformation [155,156,157,158,159]. Local immune cells experience exhaustion, and populations of anti-tumor cytotoxic T cells decline [160]. Importantly, inflammation-induced activation of STAT3 and related oncogenic pathways promotes survival signaling and resistance to apoptosis in epithelial cells, consolidating the tumor-promoting role of miR-101a in the inflamed gut [161,162,163].
This inflammatory microenvironment promotes cycles of epithelial injury and regenerative proliferation. The colon is a tissue characterized by rapid turnover, with epithelial cells replenished by stem cells located at the base of the crypts. In the presence of persistent barrier stress and inflammation, stem cells undergo hyperproliferation to repair damaged mucosa [164,165]. This process, while initially protective, increases the probability of replication errors, clonal expansion, and selection of mutant populations. Within this hyperproliferative context, canonical oncogenic pathways such as STAT3, NF-κB, COX-2/ Prostaglandin E₂ (PGE2), and Wingless Integration-1 (Wnt)/β-catenin are activated, synergizing with accumulating genetic mutations to drive the transition from normal epithelium to dysplastic adenomas [149,166,167,168]. Importantly, miR-101a acts as an upstream driver of this vicious cycle by compromising the mucus barrier and facilitating microbial translocation, thereby perpetuating the continuous activation of inflammatory and proliferative signaling pathways [33,148,169]. As adenomas develop, miR-101a continues to influence tumor progression by remodeling the tumor microenvironment. The validated gene and protein targets of miR-101a, functionally implicated in diverse disease conditions, are listed in Table 1. Persistent COX-2 activity and cytokine signaling stimulate neovascularization, ensuring adequate oxygen and nutrient supply for expanding lesions [170,171,172]. Inflammatory mediators and pathogenic bacteria remodel the extracellular matrix, weakening cell adhesion and facilitating epithelial invasion into deeper tissue layers.
1.3. Obesity Associated Colorectal Cancer Development via miR-101a
In the colon and mammary epithelium, miR-101 regulates COX-2 signaling and cytoskeletal programs, thereby linking it to barrier integrity, goblet-cell differentiation, mucus production, and epithelial restitution [32,173,174,175]. In parallel, its developmental trajectory and neuronal expression support roles in synaptic maturation and activity-dependent plasticity. Together, genomic organization, sequence features, and variant landscapes position the miR-101 family as a precision node through which diet-microbe-host signals (e.g., ethanolamine-driven transcriptional shifts) can recalibrate target accessibility and pathway flux, with implications for context-dependent phenotypes in health and disease. Chronic inflammation further recruits immunosuppressive cells, such as regulatory T cells and myeloid-derived suppressor cells, which dampen anti-tumor immunity and create a permissive environment for malignant clones [159,176]. This shift from immune surveillance to immune evasion marks a critical step in tumor progression, allowing adenomas to evolve into invasive carcinomas.
The role of miR-101a in CRC is highly context-dependent, which explains its paradoxical classification as both a tumor suppressor and a tumor promoter. In many non-obese contexts, loss of miR-101a contributes to tumorigenesis by relieving repression of oncogenes such as EZH2, MCL-1, and DNMT3A. In contrast, in the obese, dysbiotic colon, overexpression of miR-101a acts through an entirely different mechanism: weakening epithelial barrier defenses, fueling chronic inflammation, and potentiating oncogenic signaling. This duality highlights the importance of considering metabolic, microbial, and inflammatory contexts when assessing miRNA function in cancer biology. In the framework of the adenoma carcinoma sequence, miR-101a does not operate as a classical genetic driver mutation but rather as a facilitator of tumor-promoting conditions. Barrier dysfunction increases stem cell exposure to microbial ligands and mutagens, inflammation sustains proliferative and survival signaling, and microenvironmental remodeling accelerates progression. In this way, miR-101a integrates into the canonical CRC model as a non-genetic but essential regulator that increases the likelihood of malignant transformation in metabolically stressed environments.
A significant tumor-promoting effect of elevated miR-101a in this context is the disruption of mucosal barrier integrity [33]. Normally, colonic goblet cells secrete MUC2, the predominant mucin that forms the mucus layer protecting epithelial surfaces from microbial encroachment [110]. Upregulation of miR-101a interferes with goblet cell differentiation and represses the transcriptional machinery required for mucin biosynthesis. This results in decreased MUC2 production and a thinner or disrupted mucus barrier. The immediate consequence is exposure of the intestinal epithelium to luminal bacteria and their associated components, such as LPS. This condition facilitates microbial translocation into the lamina propria and systemic circulation by increasing the gut permeability [106]. The barrier defects initiated by miR-101a overexpression propagate a cycle of chronic low-grade inflammation. In summary, elevated miR-101a acts as a tumor-promoting factor in obesity associated CRC by disrupting goblet cell maturation and mucin biosynthesis, leading to MUC2 depletion and compromised mucus barrier integrity. The resulting weakening of the gut barrier permits microbial translocation and sustained low-grade inflammation, establishing a pro-carcinogenic environment.
2. Conclusions
Taken together, the evidence indicates that miR-101a serves as a central mediator of tumor initiation and progression in obesity-associated CRC. Its contribution lies not in direct oncogene activation but in ecological disruption: destabilizing mucosal defenses, perpetuating inflammation, and shaping a tumor-permissive, immune-privileged niche. By linking dietary metabolites, such as ethanolamine, to microbial dysbiosis and host epithelial regulation, miR-101a exemplifies the intricate interactions that drive cancer development beyond genetic mutations alone. Importantly, this understanding underscores the need for context-specific investigation in miRNA biology, as the same molecule may serve as a tumor suppressor in one tissue or metabolic state and a tumor promoter in another. In obesity-associated CRC, miR-101a emerges as a crucial molecular bridge linking diet, microbiota, and tumorigenesis, offering both mechanistic insight and a potential therapeutic target.
Author Contributions
S.P.M. and S.J. wrote the main manuscript text and prepared the figures. All authors (S.P.M., S.J., R.J., B.W., and H.Y.) reviewed and revised the manuscript text critically for important intellectual content.
Funding
We gratefully acknowledge the support from the Department of Defense (W81XWH-18-PRARP AZ180098), the National Institutes of Health (R56AG069676, R56AG064075, R01AG071762, R21AG072379, U01AG076928, and R21AG082164), and the Florida Department of Health (22A17) for funding provided to HY and SJ.
Data Availability Statement
No datasets were generated or analyzed in the present study.
Acknowledgments
We express our gratitude to the members of Dr. Yadav’s lab and the USF Center for Microbiome Research for their valuable contributions. We also appreciate the support and resources provided by the Microbiomes Institute, the Center of Excellence in Aging and Brain Repair, the Byrd Alzheimer’s Center, and the Department of Neurosurgery and Brain Repair at the University of South Florida Morsani College of Medicine, Tampa, FL, USA.
Conflicts of Interest
Dr. Yadav serves as Co-Founder and Chief Scientific Officer of Postbiotics Inc., MusB LLC, MusB Research LLC, and BiomAge Inc., with Dr. Jain as Co-Founder of MusB LLC and MusB Research LLC. However, their roles and the data presented in this manuscript do not currently show any conflicts of interest. All other authors declare no conflicts of interest.
References
- Akimoto, N. Rising incidence of early-onset colorectal cancer - a call to action. Nat Rev Clin Oncol 2021, 18(4), 230–243. [Google Scholar] [CrossRef] [PubMed]
- Mandic, M. Is the association of overweight and obesity with colorectal cancer underestimated? An umbrella review of systematic reviews and meta-analyses. Eur J Epidemiol 2023, 38(2), 135–144. [Google Scholar] [CrossRef]
- Arima, K. Western-Style Diet, pks Island-Carrying Escherichia coli, and Colorectal Cancer: Analyses From Two Large Prospective Cohort Studies. Gastroenterology 2022, 163(4), 862–874. [Google Scholar] [CrossRef] [PubMed]
- Lednovich, K.R. Intestinal FFA3 mediates obesogenic effects in mice on a Western diet. American Journal of Physiology-Endocrinology and Metabolism 2022, 323(3), E290–E306. [Google Scholar] [CrossRef] [PubMed]
- Lednovich, K.R. Intestinal FFA2 promotes obesity by altering food intake in Western diet-fed mice. Journal of Endocrinology 2024, 260(2), p. e230184. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet 2004, 5(7), 522–31. [Google Scholar] [CrossRef]
- Li, T. Translation suppresses exogenous target RNA-mediated microRNA decay. Nature Communications 2025, 16(1), p. 5257. [Google Scholar] [CrossRef]
- Xiao, C.; Rajewsky, K. MicroRNA Control in the Immune System: Basic Principles. Cell 2009, 136(1), 26–36. [Google Scholar] [CrossRef]
- Chaiwangyen, W. MicroRNA expression in response to environmental hazards: Implications for health. Ecotoxicology and Environmental Safety 2025, 300, 118420. [Google Scholar] [CrossRef]
- Singh, R. Dual Roles of miR-10a-5p and miR-10b-5p as Tumor Suppressors and Oncogenes in Diverse Cancers. Int J Mol Sci 2025, 26(1). [Google Scholar] [CrossRef]
- Otmani, K. The regulatory mechanisms of oncomiRs in cancer. Biomedicine & Pharmacotherapy 2024, 171, 116165. [Google Scholar] [CrossRef] [PubMed]
- Sareen, G. A new era of cancer immunotherapy: vaccines and miRNAs. Cancer Immunol Immunother 2025, 74(5), p. 163. [Google Scholar] [CrossRef]
- Chandramouli, A. MicroRNA-101 (miR-101) post-transcriptionally regulates the expression of EP4 receptor in colon cancers. Cancer Biology & Therapy 2012, 13(3), 175–183. [Google Scholar]
- Cohen, J.L. Amygdalar expression of the microRNA miR-101a and its target Ezh2 contribute to rodent anxiety-like behaviour. Eur J Neurosci 2017, 46(7), 2241–2252. [Google Scholar] [CrossRef]
- Dudakovic, A. MicroRNA-101a enhances trabecular bone accrual in male mice. Scientific Reports 2022, 12(1), p. 13361. [Google Scholar] [CrossRef]
- Liu, J.J. A novel AP-1/miR-101 regulatory feedback loop and its implication in the migration and invasion of hepatoma cells. Nucleic Acids Res 2014, 42(19), 12041–51. [Google Scholar] [CrossRef] [PubMed]
- de Sande, A.H. Cell-type-specific characterization of miRNA gene dynamics in immune cell subpopulations during aging and atherosclerosis disease development at single-cell resolution. bioRxiv 2023, 2023. 10.09.561173. [Google Scholar]
- Morita, M. Identification of microRNAs involved in acute rejection and spontaneous tolerance in murine hepatic allografts. Scientific Reports 2014, 4(1), p. 6649. [Google Scholar] [CrossRef]
- Wang, H. MiR-101 Targets the EZH2/Wnt/β-Catenin the Pathway to Promote the Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells. Scientific Reports 2016, 6(1), p. 36988. [Google Scholar] [CrossRef] [PubMed]
- Vella, S. MicroRNA-101 is repressed by EZH2 and its restoration inhibits tumorigenic features in embryonal rhabdomyosarcoma. Clinical Epigenetics 2015, 7(1), p. 82. [Google Scholar] [CrossRef]
- Xu, B. Targeting EZH2 and PRC2 dependence as novel anticancer therapy. Experimental Hematology 2015, 43(8), 698–712. [Google Scholar] [CrossRef]
- Cao, Q. Coordinated Regulation of Polycomb Group Complexes through microRNAs in Cancer. Cancer Cell 2011, 20(2), 187–199. [Google Scholar] [CrossRef] [PubMed]
- Cao, P. MicroRNA-101 negatively regulates Ezh2 and its expression is modulated by androgen receptor and HIF-1alpha/HIF-1beta. Mol Cancer 2010, 9, 108. [Google Scholar] [CrossRef]
- Smits, M. miR-101 is down-regulated in glioblastoma resulting in EZH2-induced proliferation, migration, and angiogenesis. Oncotarget 2010, 1(8). [Google Scholar] [CrossRef]
- Hillyar, C.R.T. Complex Cross-Talk between EZH2 and Mirnas Confers Hallmark Characteristics and Shapes the Tumor Microenvironment. Epigenomics 2022, 14(11), 699–709. [Google Scholar] [CrossRef]
- Gómez-Acebo, I. Role of circulating MicroRNAs in prostate cancer diagnosis and risk stratification in the MCC Spain study. Sci Rep 2025, 15(1), p. 17517. [Google Scholar] [CrossRef]
- Huang, X. SChLAP1 regulates the metastasis and apoptosis of prostate cancer partly via miR-101. Transl Androl Urol 2025, 14(6), 1782–1796. [Google Scholar] [CrossRef]
- Liu, N. MiR-101: An Important Regulator of Gene Expression and Tumor Ecosystem. Cancers (Basel) 2022, 14(23). [Google Scholar] [CrossRef] [PubMed]
- Pastena, P. Unraveling Biomarker Signatures in Triple-Negative Breast Cancer: A Systematic Review for Targeted Approaches. International Journal of Molecular Sciences 2024, 25(5), 2559. [Google Scholar] [CrossRef]
- Tessitore, A. MicroRNAs in the DNA Damage/Repair Network and Cancer. In Int J Genomics; 2014; p. 820248. [Google Scholar]
- Schnekenburger, M.; Diederich, M. Epigenetics Offer New Horizons for Colorectal Cancer Prevention. Curr Colorectal Cancer Rep 2012, 8(1), 66–81. [Google Scholar] [PubMed]
- Tâlvan, C.-D. Exploring miRNA Profiles in Colon Cancer: A Focus on miR101-3p, miR106a-5p, and miR326. Cancers 2024, 16(12), 2285. [Google Scholar] [CrossRef]
- Mishra, S.P. A mechanism by which gut microbiota elevates permeability and inflammation in obese/diabetic mice and human gut. Gut 2023, 72(10), 1848–1865. [Google Scholar] [CrossRef]
- Tarek, G. MiR-101-3p and miR-106b-5p roles in EMT pathway: prognostic and therapeutic insights for luminal breast cancer. Journal of the Egyptian National Cancer Institute 2025, 37(1), p. 43. [Google Scholar] [CrossRef]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ Res 2020, 126(11), 1549–1564. [Google Scholar] [CrossRef] [PubMed]
- Jia, X. Impact of Gut Microbiota and Microbiota-Related Metabolites on Hyperlipidemia. Front Cell Infect Microbiol 2021, 11, 634780. [Google Scholar] [CrossRef] [PubMed]
- Semo, D.; Reinecke, H.; Godfrey, R. Gut microbiome regulates inflammation and insulin resistance: a novel therapeutic target to improve insulin sensitivity. Signal Transduction and Targeted Therapy 2024, 9(1), p. 35. [Google Scholar] [CrossRef] [PubMed]
- Yarahmadi, A. Understanding the complex function of gut microbiota: its impact on the pathogenesis of obesity and beyond: a comprehensive review. Diabetology & Metabolic Syndrome 2024, 16(1), 308. [Google Scholar] [CrossRef]
- Upadhyay, G. Targeted and untargeted lipidomics with integration of liver dynamics and microbiome after dietary reversal of obesogenic diet targeting inflammation-resolution signaling in aging mice. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2024, 1869(8), 159542. [Google Scholar] [CrossRef]
- Nagpal, R.; Mishra, S.P.; Yadav, H. Unique Gut Microbiome Signatures Depict Diet-Versus Genetically Induced Obesity in Mice. International Journal of Molecular Sciences 2020, 21(10), 3434. [Google Scholar] [CrossRef]
- Glaros, S.B. Systemic and gut microbiome changes with metformin and liraglutide in youth-onset type 2 diabetes: the MIGHTY study. Gut Microbes 2025, 17(1), p. 2558071. [Google Scholar] [CrossRef]
- Dixon, S.A. The effects of prebiotics on gastrointestinal side effects of metformin in youth: A pilot randomized control trial in youth-onset type 2 diabetes. In Frontiers in Endocrinology; 2023; pp. 14–2023. [Google Scholar]
- Soundararajan, R. Integration of lipidomics with targeted, single cell, and spatial transcriptomics defines an unresolved pro-inflammatory state in colon cancer. Gut 2025, 74(4), 586–602. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J. Ethanolamine Metabolism in the Mammalian Gastrointestinal Tract: Mechanisms, Patterns, and Importance. Curr Mol Med 2017, 17(2), 92–99. [Google Scholar] [CrossRef]
- Bourdeau-Julien, I. The diet rapidly and differentially affects the gut microbiota and host lipid mediators in a healthy population. Microbiome 2023, 11(1), p. 26. [Google Scholar] [CrossRef]
- Tâlvan, C.-D. The Impact of miRNA Expression on Colon Cancer Severity, Invasiveness, and Localization. Cancers 2025, 17(7), 1091. [Google Scholar] [CrossRef]
- Rahimi-Farsi, N. Comprehensive systems biology analysis of microRNA-101-3p regulatory network identifies crucial genes and pathways in hepatocellular carcinoma. Journal of Genetic Engineering and Biotechnology 2025, 23(1), p. 100471. [Google Scholar] [CrossRef]
- Mature hsa-miR-101-3p. 2025. Available online: https://mirbase.org/mature/MIMAT0000099?utm_source=chatgpt.com.
- Zeng, X. CEBPα/miR-101b-3p promotes meningoencephalitis in mice infected with Angiostrongylus cantonensis by promoting microglial pyroptosis. Cell Communication and Signaling 2023, 21(1), p. 31. [Google Scholar] [CrossRef]
- MIR101-1 microRNA 101-1 [ Homo sapiens (human) ]. 2025. Available online: https://www.ncbi.nlm.nih.gov/gene?Cmd=DetailsSearch&Db=gene&Term=406893.
- RefSeq Gene MIR101-2. 2025.
- Perez, G. The UCSC Genome Browser database: 2025 update. Nucleic Acids Res 2025, 53(D1), D1243–d1249. [Google Scholar] [CrossRef]
- Mir101a Gene Detail. 2025.
- Cao, W. miRNASNP-v4: a comprehensive database for miRNA-related SNPs across 17 species. Nucleic Acids Research 2024, 53(D1), D1066–D1074. [Google Scholar] [CrossRef]
- SiamiGorji, S. Effects of microRNAs polymorphism in cancer progression. Med J Islam Repub Iran 2020, 34, 3. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. Genetic variations in the flanking regions of miR-101-2 are associated with increased risk of breast cancer. PLoS One 2014, 9(1), e86319. [Google Scholar] [CrossRef] [PubMed]
- Omariba, G. Genome-Wide Analysis of MicroRNA-related Single Nucleotide Polymorphisms (SNPs) in Mouse Genome. Sci Rep 2020, 10(1), p. 5789. [Google Scholar] [CrossRef]
- Miao, L. MicroRNA-101 polymorphisms and risk of head and neck squamous cell carcinoma in a Chinese population. Tumor Biology 2016, 37(3), 4169–4174. [Google Scholar] [CrossRef]
- Cao, P. MicroRNA-101 negatively regulates Ezh2 and its expression is modulated by androgen receptor and HIF-1α/HIF-1β. Molecular Cancer 2010, 9(1), p. 108. [Google Scholar] [CrossRef] [PubMed]
- Su, H. MicroRNA-101, down-regulated in hepatocellular carcinoma, promotes apoptosis and suppresses tumorigenicity. Cancer Res 2009, 69(3), 1135–42. [Google Scholar] [CrossRef]
- Lippi, G. MicroRNA-101 Regulates Multiple Developmental Programs to Constrain Excitation in Adult Neural Networks. Neuron 2016, 92(6), 1337–1351. [Google Scholar] [CrossRef]
- Kavakiotis, I. DIANA-miTED: a microRNA tissue expression database. Nucleic Acids Res 2022, 50(D1), D1055–d1061. [Google Scholar] [CrossRef]
- Panwar, B.; Omenn, G.S.; Guan, Y. miRmine: a database of human miRNA expression profiles. Bioinformatics 2017, 33(10), 1554–1560. [Google Scholar] [CrossRef]
- Su, H. MicroRNA-101, Down-regulated in Hepatocellular Carcinoma, Promotes Apoptosis and Suppresses Tumorigenicity. Cancer Research 2009, 69(3), 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. A microRNA, miR-101a, controls mammary gland development by regulating cyclooxygenase-2 expression. Differentiation 2009, 77(2), 181–187. [Google Scholar] [CrossRef] [PubMed]
- Konno, Y. MicroRNA-101 targets EZH2, MCL-1 and FOS to suppress proliferation, invasion and stem cell-like phenotype of aggressive endometrial cancer cells. Oncotarget 2014, 5(15), 6049–62. [Google Scholar] [CrossRef]
- Wang, C.Z. MiR-101: a potential therapeutic target of cancers. Am J Transl Res 2018, 10(11), 3310–3321. [Google Scholar]
- Ochs, M.J.; Steinhilber, D.; Suess, B. MicroRNA Involved in Inflammation: Control of Eicosanoid Pathway. Front Pharmacol 2011, 2, 39. [Google Scholar] [CrossRef] [PubMed]
- Li, M. MicroRNA-101 is a potential prognostic indicator of laryngeal squamous cell carcinoma and modulates CDK8. Journal of Translational Medicine 2015, 13(1), p. 271. [Google Scholar] [CrossRef]
- Mishra, R.; Singh, S.K. HIV-1 Tat C Modulates Expression of miRNA-101 to Suppress VE-Cadherin in Human Brain Microvascular Endothelial Cells. The Journal of Neuroscience 2013, 33(14), 5992–6000. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A. Hypoxia-Induced miR-101 Impairs Endothelial Barrier Integrity Through Altering VE-Cadherin and Claudin-5. Mol Neurobiol 2024, 61(3), 1807–1817. [Google Scholar] [CrossRef]
- Friedman, J.M. The Putative Tumor Suppressor microRNA-101 Modulates the Cancer Epigenome by Repressing the Polycomb Group Protein EZH2. Cancer Research 2009, 69(6), 2623–2629. [Google Scholar] [CrossRef]
- Camfield, S. Secrets of DNA-PKcs beyond DNA repair. npj Precision Oncology 2024, 8(1), 154. [Google Scholar] [CrossRef]
- Yan, D. Targeting DNA-PKcs and ATM with miR-101 sensitizes tumors to radiation. PLoS One 2010, 5(7), e11397. [Google Scholar] [CrossRef]
- Huang, Z.; Wu, X.; Li, J. miR-101 suppresses colon cancer cell migration through the regulation of EZH2. Rev Esp Enferm Dig 2021, 113(4), 255–260. [Google Scholar] [CrossRef]
- Man, X. DNMT3A and DNMT3B in Breast Tumorigenesis and Potential Therapy. Front Cell Dev Biol 2022, 10, 916725. [Google Scholar] [CrossRef] [PubMed]
- Ma, L. The Mechanism of DNA Methylation and miRNA in Breast Cancer. International Journal of Molecular Sciences 2023, 24(11), 9360. [Google Scholar] [CrossRef]
- Szczepanek, J.; Tretyn, A. MicroRNA-Mediated Regulation of Histone-Modifying Enzymes in Cancer: Mechanisms and Therapeutic Implications. Biomolecules 2023, 13(11). [Google Scholar] [CrossRef]
- Varambally, S. Genomic Loss of microRNA-101 Leads to Overexpression of Histone Methyltransferase EZH2 in Cancer. Science 2008, 322(5908), 1695–1699. [Google Scholar] [CrossRef]
- Zhu, L. MiR-101 inhibits cell proliferation and invasion of pancreatic cancer through targeting STMN1. Cancer Biomark 2018, 23(2), 301–309. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y. miR-101 inhibits autophagy and enhances cisplatin-induced apoptosis in hepatocellular carcinoma cells. Oncol Rep 2013, 29(5), 2019–24. [Google Scholar] [CrossRef]
- Sun, Q. MiR-101 sensitizes human nasopharyngeal carcinoma cells to radiation by targeting stathmin 1. Mol Med Rep 2015, 11(5), 3330–6. [Google Scholar] [CrossRef]
- Tonetti, F.R.; Eguileor, A.; Llorente, C. Goblet cells: guardians of gut immunity and their role in gastrointestinal diseases. eGastroenterology 2024, 2(3). [Google Scholar] [CrossRef] [PubMed]
- Stanforth, K.J. The MUC2 Gene Product: Polymerisation and Post-Secretory Organisation—Current Models. Polymers 2024, 16(12), 1663. [Google Scholar] [CrossRef]
- Schulz-Heddergott, R. Therapeutic Ablation of Gain-of-Function Mutant p53 in Colorectal Cancer Inhibits Stat3-Mediated Tumor Growth and Invasion. Cancer Cell 2018, 34(2), 298–314.e7. [Google Scholar] [CrossRef] [PubMed]
- Yu, H. Revisiting STAT3 signalling in cancer: new and unexpected biological functions. Nat Rev Cancer 2014, 14(11), 736–46. [Google Scholar] [CrossRef]
- Heichler, C. STAT3 activation through IL-6/IL-11 in cancer-associated fibroblasts promotes colorectal tumour development and correlates with poor prognosis. Gut 2020, 69(7), 1269–1282. [Google Scholar] [CrossRef]
- Patel, D.; Witt, S.N. Ethanolamine and Phosphatidylethanolamine: Partners in Health and Disease. In Oxid Med Cell Longev; 2017; p. 4829180. [Google Scholar]
- Sogame, R. Dietary Ethanolamine Plasmalogen from Ascidian Alleviates Chronic Hepatic Injury in Mice Treated with Continuous Acetaminophen. Applied Sciences 2025, 15(11), 5968. [Google Scholar] [CrossRef]
- Zhou, J. Ethanolamine enhances intestinal functions by altering gut microbiome and mucosal anti-stress capacity in weaned rats. British Journal of Nutrition 2018, 120(3), 241–249. [Google Scholar] [CrossRef] [PubMed]
- Everard, A. Intestinal epithelial N-acylphosphatidylethanolamine phospholipase D links dietary fat to metabolic adaptations in obesity and steatosis. Nat Commun 2019, 10(1), p. 457. [Google Scholar] [CrossRef] [PubMed]
- Elmihi, K.A. Female ethanolamine phosphate phospholyase knockout mice resisted high-fat diet-induced obesity with attenuated hepatic cholesterol deposition. Am J Physiol Gastrointest Liver Physiol 2025, 329(3), G390–g402. [Google Scholar] [CrossRef]
- Shi, Z. Dynamic regulation of gut microbiota in high-fat or normal-fed mice by dietary phosphatidylcholine and phosphatidylethanolamine. Food Bioscience 2025. 71, 107314. [Google Scholar] [CrossRef]
- Barnes, A.J. Ethanolamine metabolism through two genetically distinct loci enables Klebsiella pneumoniae to bypass nutritional competition in the gut. PLoS Pathog 2024, 20(5), e1012189. [Google Scholar] [CrossRef]
- Rowley, C.A.; Anderson, C.J.; Kendall, M.M. Ethanolamine Influences Human Commensal Escherichia coli Growth, Gene Expression, and Competition with Enterohemorrhagic E. coli O157:H7. mBio 2018, 9(5). [Google Scholar] [CrossRef]
- Thiennimitr, P. Intestinal inflammation allows Salmonella to use ethanolamine to compete with the microbiota. Proc Natl Acad Sci U S A 2011, 108(42), 17480–5. [Google Scholar] [CrossRef]
- Ormsby, M.J. Inflammation associated ethanolamine facilitates infection by Crohn’s disease-linked adherent-invasive Escherichia coli. EBioMedicine 2019, 43, 325–332. [Google Scholar] [CrossRef]
- Pacheco, A.R.; Sperandio, V. Enteric Pathogens Exploit the Microbiota-generated Nutritional Environment of the Gut. Microbiology Spectrum 2015, 3(3). [Google Scholar] [CrossRef]
- Lengfelder, I. Complex Bacterial Consortia Reprogram the Colitogenic Activity of Enterococcus faecalis in a Gnotobiotic Mouse Model of Chronic, Immune-Mediated Colitis. Front Immunol 2019, 10, 1420. [Google Scholar] [CrossRef]
- Garsin, D.A. Ethanolamine utilization in bacterial pathogens: roles and regulation. Nature Reviews Microbiology 2010, 8(4), 290–295. [Google Scholar] [CrossRef]
- Anderson, C.J. Ethanolamine Signaling Promotes Salmonella Niche Recognition and Adaptation during Infection. PLoS Pathog 2015, 11(11), e1005278. [Google Scholar]
- Fang, H. Postbiotic Impact on Host Metabolism and Immunity Provides Therapeutic Potential in Metabolic Disease. Endocrine Reviews 2024, 46(1), 60–79. [Google Scholar] [CrossRef]
- Breton, J.; Galmiche, M.; Déchelotte, P. Dysbiotic Gut Bacteria in Obesity: An Overview of the Metabolic Mechanisms and Therapeutic Perspectives of Next-Generation Probiotics. Microorganisms 2022, 10(2). [Google Scholar] [CrossRef]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunology 2017, 10(1), 18–26. [Google Scholar] [CrossRef]
- Mishra, S.P. New Horizons in Microbiota and Metabolic Health Research. The Journal of Clinical Endocrinology & Metabolism 2020, 106(2), e1052–e1059. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S. A Cascade of Microbiota-Leaky Gut-Inflammation- Is it a Key Player in Metabolic Disorders? Current Obesity Reports 2025, 14(1), p. 32. [Google Scholar] [CrossRef] [PubMed]
- Okumura, R.; Takeda, K. The role of the mucosal barrier system in maintaining gut symbiosis to prevent intestinal inflammation. Semin Immunopathol 2024, 47(1), p. 2. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X. Mucosal immune response in biology, disease prevention and treatment. Signal Transduction and Targeted Therapy 2025, 10(1), p. 7. [Google Scholar] [CrossRef]
- Song, C. Intestinal mucus components and secretion mechanisms: what we do and do not know. Experimental & Molecular Medicine 2023, 55(4), 681–691. [Google Scholar] [CrossRef]
- Mishra, S.P. Abnormalities in microbiota/butyrate/FFAR3 signaling in aging gut impair brain function. JCI Insight 2024, 9(3). [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Larsson, J.M.H.; Hansson, G.C. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host–microbial interactions. Proceedings of the National Academy of Sciences 2011, 108 (supplement_1), 4659–4665. [Google Scholar] [CrossRef]
- McKenna, L.B. MicroRNAs Control Intestinal Epithelial Differentiation, Architecture, and Barrier Function. Gastroenterology 2010, 139(5), 1654–1664.e1. [Google Scholar] [CrossRef]
- Bergstrom, K.S. Muc2 protects against lethal infectious colitis by disassociating pathogenic and commensal bacteria from the colonic mucosa. PLoS Pathog 2010, 6(5), e1000902. [Google Scholar] [CrossRef]
- Johansson, M.E.V. Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis. Gut 2014, 63(2), 281–291. [Google Scholar] [CrossRef] [PubMed]
- van der Post, S. Structural weakening of the colonic mucus barrier is an early event in ulcerative colitis pathogenesis. Gut 2019, 68(12), 2142–2151. [Google Scholar] [CrossRef]
- Jiang, Z. Proteus mirabilis exacerbates ulcerative colitis by inhibiting mucin production. Front Microbiol 2025, 16, 1556953. [Google Scholar] [CrossRef]
- Guagliano, G. The hidden piece in inflammatory bowel diseases: The significance of intestinal mucus in mediating the bacteria-cells crosstalk. Materials Today Bio 2025. 33, 101937. [Google Scholar] [CrossRef] [PubMed]
- Genua, F. The Role of Gut Barrier Dysfunction and Microbiome Dysbiosis in Colorectal Cancer Development. Front Oncol 2021, 11, 626349. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F.; Artis, D.; Becker, C. The intestinal barrier: a pivotal role in health, inflammation, and cancer. Lancet Gastroenterol Hepatol 2025, 10(6), 573–592. [Google Scholar] [CrossRef]
- Kearns, R. Gut modulation to regulate NF-κB in colorectal and gastric cancer therapy and inflammation. Cancer Immunol Immunother 2025, 74(8), p. 264. [Google Scholar] [CrossRef]
- Lei, L. Increased intestinal permeability and lipopolysaccharide contribute to swainsonine-induced systemic inflammation. Ecotoxicology and Environmental Safety 2024, 284, 116912. [Google Scholar] [CrossRef]
- He, R. Dysbiosis and extraintestinal cancers. Journal of Experimental & Clinical Cancer Research 2025, 44(1), p. 44. [Google Scholar] [CrossRef]
- Matar, A. Intestinal Barrier Impairment, Preservation, and Repair: An Update. Nutrients 2024, 16(20), 3494. [Google Scholar] [CrossRef]
- Neurath, M.F.; Artis, D.; Becker, C. The intestinal barrier: a pivotal role in health, inflammation, and cancer. The Lancet Gastroenterology & Hepatology 2025, 10(6), 573–592. [Google Scholar] [CrossRef] [PubMed]
- Dmytriv, T.R.; Storey, K.B.; Lushchak, V.I. Intestinal barrier permeability: the influence of gut microbiota, nutrition, and exercise. Front Physiol 2024, 15, 1380713. [Google Scholar] [CrossRef]
- Mazaheri-Tehrani, S. Serum lipopolysaccharide binding protein (LBP) and metabolic syndrome: a systematic review and meta-analysis. Diabetology & Metabolic Syndrome 2025, 17(1), p. 268. [Google Scholar] [CrossRef]
- Mazaheri-Tehrani, S. Serum lipopolysaccharide binding protein (LBP) and metabolic syndrome: a systematic review and meta-analysis. Diabetol Metab Syndr 2025, 17(1), p. 268. [Google Scholar] [CrossRef] [PubMed]
- Starbæk, S.M.R. Meta-inflammation and endotoxemia in a highly translational porcine model of diet-induced obesity. Lab Animal 2025. [Google Scholar] [CrossRef] [PubMed]
- Hu, J. Brd4 modulates metabolic endotoxemia-induced inflammation by regulating colonic macrophage infiltration in high-fat diet-fed mice. Communications Biology 2024, 7(1), p. 1708. [Google Scholar] [CrossRef]
- Dimeji, I.Y.; Ayodeji, A.S. Pharmacological modulation of the gut microbiota and endotoxemia: A next-generation approach to treating metabolic syndrome. ASPET Discovery 2025. 1, 100010. [Google Scholar] [CrossRef]
- Jacobson, R. Collagenolytic Enterococcus faecalis induces DDR1 signaling, proliferation and altered immune infiltrate in colorectal peritoneal metastases. Surgery Open Science 2025. 28, 65–72. [Google Scholar] [CrossRef]
- Fang, J. Slimy partners: the mucus barrier and gut microbiome in ulcerative colitis. Experimental & Molecular Medicine 2021, 53(5), 772–787. [Google Scholar] [CrossRef]
- Rath, E.; Haller, D. Intestinal epithelial cell metabolism at the interface of microbial dysbiosis and tissue injury. Mucosal Immunology 2022, 15(4), 595–604. [Google Scholar] [CrossRef]
- Mishra, P. From Dysbiosis to Diagnosis: The Role of Gut Microbiota in Breast Cancer Etiology and Management. Applied Microbiology 2025, 5(4), 134. [Google Scholar] [CrossRef]
- Mukherjee, T. The NF-κB signaling system in the immunopathogenesis of inflammatory bowel disease. Science Signaling 2024, 17(818), p. eadh1641. [Google Scholar] [CrossRef]
- Chen, L. Interactions between toll-like receptors signaling pathway and gut microbiota in host homeostasis. Immun Inflamm Dis 2024, 12(7), p. e1356. [Google Scholar] [CrossRef]
- Clarke, Thomas B. Invasive Bacterial Pathogens Exploit TLR-Mediated Downregulation of Tight Junction Components to Facilitate Translocation across the Epithelium. Cell Host & Microbe 2011, 9(5), 404–414. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, B. The role of gut microbiota, immune system, and autophagy in the pathogenesis of inflammatory bowel disease: Molecular mechanisms and therapeutic approaches. Aspects of Molecular Medicine 2024. 4, 100056. [Google Scholar] [CrossRef]
- Robles-Vera, I. Microbiota translocation following intestinal barrier disruption promotes Mincle-mediated training of myeloid progenitors in the bone marrow. Immunity 2025. 58, 2, 381–396.e9. [Google Scholar] [CrossRef]
- Lei, W. Gut microbiota-driven neuroinflammation in Alzheimer’s disease: from mechanisms to therapeutic opportunities. Front Immunol 2025, 16, 1582119. [Google Scholar] [CrossRef]
- Desind, S.Z. Long noncoding RNAs and their complex role in shaping and regulating arachidonic acid metabolism: Learning to love the (not-really) junk. Wiley Interdiscip Rev RNA 2023, p. e1828. [Google Scholar] [CrossRef]
- Hao, Y. Enforced expression of miR-101 inhibits prostate cancer cell growth by modulating the COX-2 pathway in vivo. Cancer Prev Res (Phila) 2011, 4(7), 1073–83. [Google Scholar] [CrossRef]
- Che, L. MicroRNA-101 inhibits cadmium-induced angiogenesis by targeting cyclooxygenase-2 in primary human umbilical vein endothelial cells. Biochemical Pharmacology 2021, 189, 114192. [Google Scholar] [CrossRef]
- Liu, N. MiR-101: An Important Regulator of Gene Expression and Tumor Ecosystem. Cancers 2022, 14(23), 5861. [Google Scholar] [CrossRef] [PubMed]
- Chandimali, N. Free radicals and their impact on health and antioxidant defenses: a review. Cell Death Discovery 2025, 11(1), p. 19. [Google Scholar] [CrossRef]
- Morris, G. Redox regulation of the immune response. Cellular & Molecular Immunology 2022, 19(10), 1079–1101. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, R.R. ROS signaling in innate immunity via oxidative protein modifications. Front Immunol 2024, 15, 1359600. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. Signaling pathways involved in colorectal cancer: pathogenesis and targeted therapy. Signal Transduction and Targeted Therapy 2024, 9(1), p. 266. [Google Scholar] [CrossRef]
- Shahgoli, V.K. Inflammatory bowel disease, colitis, and cancer: unmasking the chronic inflammation link. Int J Colorectal Dis 2024, 39(1), p. 173. [Google Scholar] [CrossRef]
- Farmanbar, A.; Kneller, R.; Firouzi, S. Mutational signatures reveal mutual exclusivity of homologous recombination and mismatch repair deficiencies in colorectal and stomach tumors. Scientific Data 2023, 10(1), p. 423. [Google Scholar] [CrossRef]
- Thatikonda, V. Comprehensive analysis of mutational signatures reveals distinct patterns and molecular processes across 27 pediatric cancers. Nature Cancer 2023, 4(2), 276–289. [Google Scholar] [CrossRef]
- Grzelakowska, A.; Kalyanaraman, B.; Zielonka, J. Small molecule probes for peroxynitrite detection. Redox Biochemistry and Chemistry 2024, 10, 100034. [Google Scholar] [CrossRef] [PubMed]
- Seiwert, N. Heme oxygenase 1 protects human colonocytes against ROS formation, oxidative DNA damage and cytotoxicity induced by heme iron, but not inorganic iron. Cell Death & Disease 2020, 11(9), p. 787. [Google Scholar] [CrossRef] [PubMed]
- Irrazabal, T. Limiting oxidative DNA damage reduces microbe-induced colitis-associated colorectal cancer. Nature Communications 2020, 11(1), p. 1802. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y. Inflammation in cancer: therapeutic opportunities from new insights. Molecular Cancer 2025, 24(1), p. 51. [Google Scholar] [CrossRef]
- Wen, Y. Chronic inflammation, cancer development and immunotherapy. Front Pharmacol 2022, 13, 1040163. [Google Scholar] [CrossRef]
- Tripathi, S.; Sharma, Y.; Kumar, D. Unveiling the link between chronic inflammation and cancer. Metabolism Open 2025, 25, 100347. [Google Scholar] [CrossRef]
- Zhao, H. Inflammation and tumor progression: signaling pathways and targeted intervention. Signal Transduction and Targeted Therapy 2021, 6(1), 263. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51(1), 27–41. [Google Scholar] [CrossRef]
- Chi, X. T-cell exhaustion and stemness in antitumor immunity: Characteristics, mechanisms, and implications. Front Immunol 2023, 14, 1104771. [Google Scholar] [CrossRef]
- Voshagh, Q. Investigating the association between the tissue expression of miRNA-101, JAK2/STAT3 with TNF-α, IL-6, IL-1β, and IL-10 cytokines in the ulcerative colitis patients. Immun Inflamm Dis 2024, 12(3), p. e1224. [Google Scholar] [CrossRef]
- Hu, Y.; Dong, Z.; Liu, K. Unraveling the complexity of STAT3 in cancer: molecular understanding and drug discovery. Journal of Experimental & Clinical Cancer Research 2024, 43(1), p. 23. [Google Scholar] [CrossRef]
- Rahbar Farzam, O. Interplay of miRNAs and lncRNAs in STAT3 signaling pathway in colorectal cancer progression. Cancer Cell International 2024, 24(1), p. 16. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Augenlicht, L.H. Intestinal stem cells: guardians of homeostasis in health and aging amid environmental challenges. Experimental & Molecular Medicine 2024, 56(3), 495–500. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z. Oxidative stress caused by 3-monochloro-1,2-propanediol provokes intestinal stem cell hyperproliferation and the protective role of quercetin. Ecotoxicology and Environmental Safety 2025. 291, 117851. [Google Scholar] [CrossRef]
- Good, H.J. Prostaglandin E(2) and Akt Promote Stemness in Apc Mutant Dclk1+ Cells to Give Rise to Colitis-associated Cancer. Cell Mol Gastroenterol Hepatol 2025, 19(6), p. 101469. [Google Scholar] [CrossRef]
- Cineus, R. The IL-22–oncostatin M axis promotes intestinal inflammation and tumorigenesis. Nature Immunology 2025, 26(6), 837–853. [Google Scholar] [CrossRef] [PubMed]
- Morrison, H.A. NF-κB Inducing Kinase Attenuates Colorectal Cancer by Regulating Noncanonical NF-κB Mediated Colonic Epithelial Cell Regeneration. Cellular and Molecular Gastroenterology and Hepatology 2024, 18(3). [Google Scholar] [CrossRef]
- Song, P. Wnt/β-catenin signaling pathway in carcinogenesis and cancer therapy. Journal of Hematology & Oncology 2024, 17(1), p. 46. [Google Scholar]
- Deng, B. COX2 Enhances Neovascularization of Inflammatory Tenocytes Through the HIF-1α/VEGFA/PDGFB Pathway. Front Cell Dev Biol 2021, 9, 670406. [Google Scholar] [CrossRef]
- Geindreau, M.; Bruchard, M.; Vegran, F. Role of Cytokines and Chemokines in Angiogenesis in a Tumor Context. Cancers (Basel) 2022, 14(10). [Google Scholar] [CrossRef]
- Jacobson, R.A. Enterococcus faecalis exploits the human fibrinolytic system to drive excess collagenolysis: implications in gut healing and identification of druggable targets. Am J Physiol Gastrointest Liver Physiol 2020, 318(1), G1–g9. [Google Scholar] [CrossRef]
- Strillacci, A. MiR-101 downregulation is involved in cyclooxygenase-2 overexpression in human colon cancer cells. Exp Cell Res 2009, 315(8), 1439–47. [Google Scholar] [CrossRef]
- Schaefer, J.S. MicroRNA signatures differentiate Crohn’s disease from ulcerative colitis. BMC Immunology 2015, 16(1), 5. [Google Scholar] [CrossRef] [PubMed]
- Chandramouli, A. MicroRNA-101 (miR-101) post-transcriptionally regulates the expression of EP4 receptor in colon cancers. Cancer Biol Ther 2012, 13(3), 175–83. [Google Scholar] [CrossRef] [PubMed]
- Umansky, V. The Role of Myeloid-Derived Suppressor Cells (MDSC) in Cancer Progression. Vaccines (Basel) 2016, 4(4). [Google Scholar] [CrossRef]
- Liu, J. [miR-101 inhibits the proliferation and migration of breast cancer cells via downregulating the expression of DNA methyltransferase 3a]. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2016, 32(3), 299–303. [Google Scholar]
- Wang, H. MiR-101 Targets the EZH2/Wnt/β-Catenin the Pathway to Promote the Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells. Sci Rep 2016, 6, 36988. [Google Scholar] [CrossRef]
- Vella, S. MicroRNA-101 is repressed by EZH2 and its restoration inhibits tumorigenic features in embryonal rhabdomyosarcoma. Clin Epigenetics 2015, 7(1), p. 82. [Google Scholar] [CrossRef]
- Smits, M. miR-101 is down-regulated in glioblastoma resulting in EZH2-induced proliferation, migration, and angiogenesis. Oncotarget 2010, 1(8), 710–20. [Google Scholar] [CrossRef]
- Friedman, J.M. The putative tumor suppressor microRNA-101 modulates the cancer epigenome by repressing the polycomb group protein EZH2. Cancer Res 2009, 69(6), 2623–9. [Google Scholar] [CrossRef]
- Varambally, S. Genomic loss of microRNA-101 leads to overexpression of histone methyltransferase EZH2 in cancer. Science 2008, 322(5908), 1695–9. [Google Scholar] [CrossRef]
- Chakrabarty, A. MicroRNA regulation of cyclooxygenase-2 during embryo implantation. Proc Natl Acad Sci U S A 2007, 104(38), 15144–9. [Google Scholar] [CrossRef]
- Tanaka, T. A microRNA, miR-101a, controls mammary gland development by regulating cyclooxygenase-2 expression. Differentiation 2009, 77(2), 181–7. [Google Scholar] [CrossRef]
- Liu, Y. MicroRNA-101 inhibits angiogenesis via COX-2 in endometrial carcinoma. Mol Cell Biochem 2018, 448(1-2), 61–69. [Google Scholar] [CrossRef] [PubMed]
- Che, L. MicroRNA-101 inhibits cadmium-induced angiogenesis by targeting cyclooxygenase-2 in primary human umbilical vein endothelial cells. Biochem Pharmacol 2021, 189, 114192. [Google Scholar] [CrossRef] [PubMed]
- Harati, R. Loss of miR-101-3p Promotes Transmigration of Metastatic Breast Cancer Cells through the Brain Endothelium by Inducing COX-2/MMP1 Signaling. Pharmaceuticals (Basel) 2020, 13(7). [Google Scholar] [CrossRef] [PubMed]
- Cai, Y. Eicosapentaenoic acid’s metabolism of 15-LOX-1 promotes the expression of miR-101 thus inhibits Cox2 pathway in colon cancer. Onco Targets Ther 2020, 13, 5605–5616. [Google Scholar] [CrossRef]
- Fan, M.J. Long non-coding RNA SPRY4-IT1 promotes epithelial-mesenchymal transition of cervical cancer by regulating the miR-101-3p/ZEB1 axis. Biosci Rep 2019, 39(6). [Google Scholar] [CrossRef]
- Xiong, W.C. Interplay between long noncoding RNA ZEB1-AS1 and miR-101/ZEB1 axis regulates proliferation and migration of colorectal cancer cells. Am J Transl Res 2018, 10(2), 605–617. [Google Scholar]
- Liang, H. LncRNA PTAR promotes EMT and invasion-metastasis in serous ovarian cancer by competitively binding miR-101-3p to regulate ZEB1 expression. Mol Cancer 2018, 17(1), p. 119. [Google Scholar] [CrossRef]
- Yin, L.; Wang, Y. Extracellular vesicles derived from M2-polarized tumor-associated macrophages promote immune escape in ovarian cancer through NEAT1/miR-101-3p/ZEB1/PD-L1 axis. Cancer Immunol Immunother 2023, 72(3), 743–758. [Google Scholar] [CrossRef]
- Shahverdi, M. Knockdown of Myeloid Cell Leukemia-1 by MicroRNA-101 Increases Sensitivity of A549 Lung Cancer Cells to Etoposide. Iran J Med Sci 2021, 46(4), 298–307. [Google Scholar]
- Wang, H. MALAT1/miR-101-3p/MCL1 axis mediates cisplatin resistance in lung cancer. Oncotarget 2018, 9(7), 7501–7512. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Li, Z. Overexpression of miR-101 promotes TRAIL-induced mitochondrial apoptosis in papillary thyroid carcinoma by targeting c-met and MCL-1. Oncotarget 2017, 8(65), 108665–108675. [Google Scholar] [CrossRef] [PubMed]
- Shen, W. MicroRNA-101-5p inhibits the growth and metastasis of cervical cancer cell by inhibiting CXCL6. Eur Rev Med Pharmacol Sci 2019, 23(5), 1957–1968. [Google Scholar] [PubMed]
- Chen, Q. miRNA-101-5p inhibits the growth and aggressiveness of NSCLC cells through targeting CXCL6. Onco Targets Ther 2019, 12, 835–848. [Google Scholar] [CrossRef]
- Chen, F. MicroRNA-101 Targets CXCL12-Mediated Akt and Snail Signaling Pathways to Inhibit Cellular Proliferation and Invasion in Papillary Thyroid Carcinoma. Oncol Res 2019, 27(6), 691–701. [Google Scholar] [CrossRef]
- Chen, H. Circular RNA hsa_circ_0007364 increases cervical cancer progression through activating methionine adenosyltransferase II alpha (MAT2A) expression by restraining microRNA-101-5p. Bioengineered 2020, 11(1), 1269–1279. [Google Scholar] [CrossRef]
- Wang, H.; Xiao, R.; Yang, B. MiR-101-3p Suppresses Progression of Cervical Squamous Cell Carcinoma by Targeting and Down-Regulating KPNA2. Technol Cancer Res Treat 2021, 20, p. 15330338211055948. [Google Scholar] [CrossRef]
- Li, Z. miR-101-3p sensitizes non-small cell lung cancer cells to irradiation. Open Med (Wars) 2020, 15(1), 413–423. [Google Scholar] [CrossRef]
- Chen, M. Long Noncoding RNA FAM201A Mediates the Radiosensitivity of Esophageal Squamous Cell Cancer by Regulating ATM and mTOR Expression via miR-101. Front Genet 2018, 9, 611. [Google Scholar] [CrossRef]
- Shao, N.; Song, L.; Sun, X. Exosomal circ_PIP5K1A regulates the progression of non-small cell lung cancer and cisplatin sensitivity by miR-101/ABCC1 axis. Mol Cell Biochem 2021, 476(6), 2253–2267. [Google Scholar] [CrossRef]
- Xie, F.W.; Liu, J.C. LncRNA SNHG12 regulates the miR-101-3p/CUL4B axis to mediate the proliferation, migration and invasion of non-small cell lung cancer. Kaohsiung J Med Sci 2021, 37(8), 664–674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H. Circular RNA ZFR accelerates non-small cell lung cancer progression by acting as a miR-101-3p sponge to enhance CUL4B expression. Artif Cells Nanomed Biotechnol 2019, 47(1), 3410–3416. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z. Inhibition of MicroRNA miR-101-3p on prostate cancer progression by regulating Cullin 4B (CUL4B) and PI3K/AKT/mTOR signaling pathways. Bioengineered 2021, 12(1), 4719–4735. [Google Scholar] [CrossRef] [PubMed]
- Li, K. Long non-coding RNA SNHG6 promotes the growth and invasion of non-small cell lung cancer by downregulating miR-101-3p. Thorac Cancer 2020, 11(5), 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Ding, C. Exosomal Circ-MEMO1 Promotes the Progression and Aerobic Glycolysis of Non-small Cell Lung Cancer Through Targeting MiR-101-3p/KRAS Axis. Front Genet 2020, 11, 962. [Google Scholar] [CrossRef] [PubMed]
- Li, L. MiR-101-3p inhibits EMT to attenuate proliferation and metastasis in glioblastoma by targeting TRIM44. J Neurooncol 2019, 141(1), 19–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H. Long noncoding RNA CRNDE promotes proliferation, migration and invasion in prostate cancer through miR-101/Rap1A. Neoplasma 2020, 67(3), 584–594. [Google Scholar] [CrossRef]
- Zhou, Z. MicroRNA-101 suppresses colorectal cancer progression by negative regulation of Rap1b. Oncol Lett 2020, 20(3), 2225–2231. [Google Scholar] [CrossRef]
- Jin, Q. MicroRNA-101-3p inhibits proliferation in retinoblastoma cells by targeting EZH2 and HDAC9. Exp Ther Med 2018, 16(3), 1663–1670. [Google Scholar] [CrossRef]
- Cao, K. miR-101 Inhibiting Cell Proliferation, Migration and Invasion in Hepatocellular Carcinoma through Downregulating Girdin. Mol Cells 2016, 39(2), 96–102. [Google Scholar] [CrossRef]
- Liu, Z. MicroRNA-101 suppresses migration and invasion via targeting vascular endothelial growth factor-C in hepatocellular carcinoma cells. Oncol Lett 2016, 11(1), 433–438. [Google Scholar] [CrossRef]
- Liu, P. LncRNA-MALAT1 mediates cisplatin resistance via miR-101-3p/VEGF-C pathway in bladder cancer. Acta Biochim Biophys Sin (Shanghai) 2019, 51(11), 1148–1157. [Google Scholar] [CrossRef]
- Calastri, M.C.J. Investigating VEGF. miR-145-3p, and miR-101-3p Expression in Patients with Cholangiocarcinoma. Asian Pac J Cancer Prev 2022, 23(7), 2233–2241. [Google Scholar] [CrossRef]
- Guo, X. Cancer-Associated Fibroblasts Promote Migration and Invasion of Non-Small Cell Lung Cancer Cells via miR-101-3p Mediated VEGFA Secretion and AKT/eNOS Pathway. Front Cell Dev Biol 2021, 9, 764151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. miR-101 inhibits cholangiocarcinoma angiogenesis through targeting vascular endothelial growth factor (VEGF). Am J Pathol 2013, 182(5), 1629–39. [Google Scholar] [CrossRef]
- Du, Y.L. LncRNA XIST Promotes Migration and Invasion of Papillary Thyroid Cancer Cell by Modulating MiR-101-3p/CLDN1 Axis. Biochem Genet 2021, 59(2), 437–452. [Google Scholar] [CrossRef] [PubMed]
- Si, W. The coordination between ZNF217 and LSD1 contributes to hepatocellular carcinoma progress and is negatively regulated by miR-101. Exp Cell Res 2019, 379(1), 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. MicroRNA-101-3p suppresses proliferation and migration in hepatocellular carcinoma by targeting the HGF/c-Met pathway. Invest New Drugs 2020, 38(1), 60–69. [Google Scholar] [CrossRef]
- Meng, X. Influence of miR-101 on proliferation of liver cancer cells through the MAPK/ERK signaling pathway. Oncol Lett 2020, 19(2), 1310–1316. [Google Scholar] [CrossRef]
- Saika, R. MicroRNA-101a regulates microglial morphology and inflammation. Journal of Neuroinflammation 2017, 14(1), p. 109. [Google Scholar] [CrossRef]
- Wang, L. IL-1β-Mediated Repression of microRNA-101 Is Crucial for Inflammation-Promoted Lung Tumorigenesis. Cancer Research 2014, 74(17), 4720–4730. [Google Scholar] [CrossRef]
- Ma, S. A novel regulatory loop miR-101/ANXA2/EGR1 mediates malignant characteristics of liver cancer stem cells. Carcinogenesis 2021, 42(1), 93–104. [Google Scholar] [CrossRef]
- Wu, F. MicroRNA-101-3p regulates gastric cancer cell proliferation, invasion and apoptosis by targeting PIM 1 expression. Cell Mol Biol (Noisy-le-grand) 2019, 65(7), 118–122. [Google Scholar] [CrossRef]
- Chen, L. MicroRNA-101 inhibits cell migration and invasion in bladder cancer via targeting FZD4. Exp Ther Med 2019, 17(2), 1476–1485. [Google Scholar] [CrossRef] [PubMed]
- Tao, L. HIPK3 Inhibition by Exosomal hsa-miR-101-3p Is Related to Metabolic Reprogramming in Colorectal Cancer. Front Oncol 2021, 11, 758336. [Google Scholar] [CrossRef]
- Wu, H.B. CircAPLP2 regulates the proliferation and metastasis of colorectal cancer by targeting miR-101-3p to activate the Notch signalling pathway. Am J Transl Res 2020, 12(6), 2554–2569. [Google Scholar]
- Yang, Q.; Yu, W.; Han, X. Overexpression of microRNA-101 causes anti-tumor effects by targeting CREB1 in colon cancer. Mol Med Rep 2019, 19(4), 3159–3167. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. miR-101-3p and miR-199b-5p promote cell apoptosis in oral cancer by targeting BICC1. Mol Cell Probes 2020, 52, 101567. [Google Scholar] [CrossRef] [PubMed]
- Wang, L. miR-101 promotes breast cancer cell apoptosis by targeting Janus kinase 2. Cell Physiol Biochem 2014, 34(2), 413–22. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Microbiota-Metabolite-miRNA Crosstalk Links Obesity to Colorectal Cancer Risk. Schematic representation contrasting gut ecosystem states in a healthy lean individual (left) versus an obese, type 2 diabetic (T2D) individual (right). In lean subjects, a eubiotic gut microbiota maintains balanced ethanolamine concentrations and supports efficient ethanolamine utilization pathways (Eut operon), thereby preventing overexpression of the oncogenic miR-101a. Intact mucus barrier integrity, abundant commensal taxa (e.g., Lactobacillus, Bifidobacterium, Clostridium), and regulated inflammatory responses collectively preserve epithelial homeostasis. This microenvironment is characterized by reduced pro-inflammatory signaling, proper metabolic regulation, and protection against DNA damage, preventing colorectal cancer initiation. By contrast, in obese and T2D individuals, dysbiosis results in elevated luminal ethanolamine and impaired utilization, leading to pathological miR-101a upregulation. Consequent disruption of goblet cell function and mucin production weakens the mucus barrier, allowing microbial translocation and sustained activation of inflammatory pathways. Chronic cytokine signaling fosters epithelial hyperproliferation, genomic instability, and impaired apoptosis, while simultaneous angiogenic and immunosuppressive cues create a tumor-permissive niche. These changes predispose obese and metabolically dysregulated hosts to the development of colorectal adenomas and carcinoma.
Figure 1.
Microbiota-Metabolite-miRNA Crosstalk Links Obesity to Colorectal Cancer Risk. Schematic representation contrasting gut ecosystem states in a healthy lean individual (left) versus an obese, type 2 diabetic (T2D) individual (right). In lean subjects, a eubiotic gut microbiota maintains balanced ethanolamine concentrations and supports efficient ethanolamine utilization pathways (Eut operon), thereby preventing overexpression of the oncogenic miR-101a. Intact mucus barrier integrity, abundant commensal taxa (e.g., Lactobacillus, Bifidobacterium, Clostridium), and regulated inflammatory responses collectively preserve epithelial homeostasis. This microenvironment is characterized by reduced pro-inflammatory signaling, proper metabolic regulation, and protection against DNA damage, preventing colorectal cancer initiation. By contrast, in obese and T2D individuals, dysbiosis results in elevated luminal ethanolamine and impaired utilization, leading to pathological miR-101a upregulation. Consequent disruption of goblet cell function and mucin production weakens the mucus barrier, allowing microbial translocation and sustained activation of inflammatory pathways. Chronic cytokine signaling fosters epithelial hyperproliferation, genomic instability, and impaired apoptosis, while simultaneous angiogenic and immunosuppressive cues create a tumor-permissive niche. These changes predispose obese and metabolically dysregulated hosts to the development of colorectal adenomas and carcinoma.
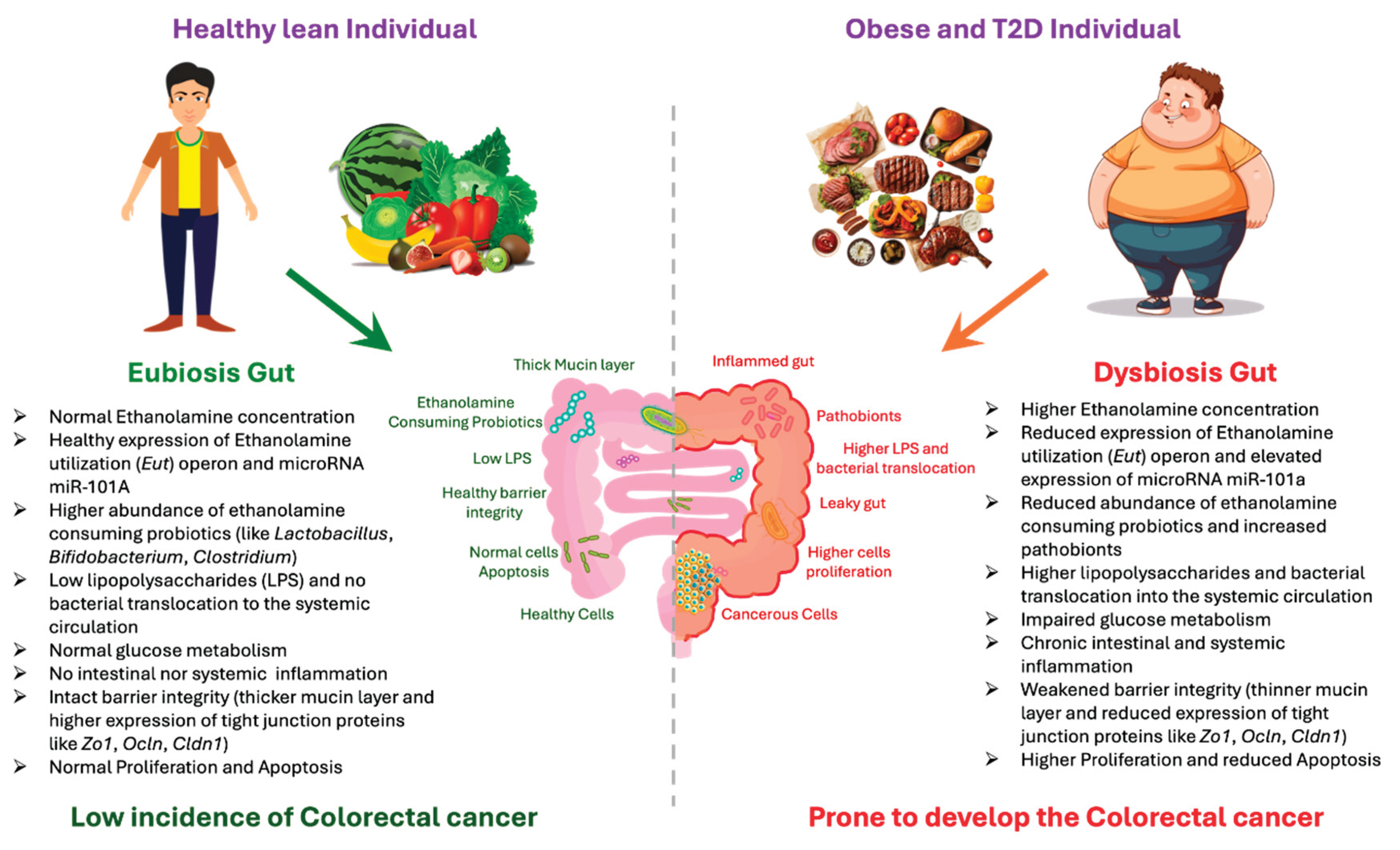

Table 1.
Validated Gene and Protein Targets of miR-101a and Their Functional Roles in Cancer and Disease Contexts.
Table 1.
Validated Gene and Protein Targets of miR-101a and Their Functional Roles in Cancer and Disease Contexts.
| Gene/Protein | Function | Ref. |
| E-cadherin | Overexpressed miR-101 suppresses DNMT3A, thereby restoring E-cadherin and inhibiting proliferation and migration in MDA-MB-231 breast cancer cells | [177] |
| EZH2 | Overexpression of miR-101a-3p in the amygdala increases anxiety-like behavior in rats by repressing Ezh2 | [14] |
| miR-101 promotes osteogenic differentiation of hBMSCs by targeting EZH2 and activating the Wnt/β-catenin pathway | [178] | |
| miR-101 is downregulated in eRMS, inversely correlated with EZH2, and its re-expression suppresses EZH2, reducing migration, clonogenicity, and tumorigenic potential of eRMS cells | [179] | |
| miR-101 suppresses Ezh2, reducing invasion and migration of prostate cancer cells, with its expression further modulated by androgen signaling and HIF-1α/β induction | [23] | |
| miR-101 downregulation drives EZH2 overexpression in GBM, and inhibiting EZH2 suppresses tumor growth, invasion, and angiogenesis both in vitro and in vivo | [180] | |
| Re-expression of miR-101 suppresses proliferation, invasion, and self-renewal of aggressive endometrial cancer cells by directly targeting EZH2, MCL-1, and FOS | [66] | |
| miR-101 is downregulated in bladder TCC, and its restoration suppresses proliferation by directly repressing EZH2, highlighting its role as a tumor suppressor. | [181] | |
| miR-101 is downregulated in CRC, and its restoration suppresses EZH2-driven migration of colorectal cancer cells. | [75] | |
| Genomic loss of miR-101 in prostate cancer reduces its repression of EZH2, leading to EZH2 overexpression and epigenetic dysregulation that drives tumor progression. | [182] | |
| PTGS2/COX2 | miR-101a directly repress COX-2 translation, with precursor overexpression reducing and antisense inhibition restoring COX-2 reporter and protein levels | [183] |
| miR-101a upregulation during mammary gland development suppresses COX-2, thereby inhibiting proliferation and modulating differentiation of mammary epithelial cells. | [184] | |
| miR-101 is downregulated in endometrial cancer, and its restoration suppresses angiogenesis and tumor growth partly through COX-2 regulation. | [185] | |
| Exogenous miR-101 directly targets COX-2, suppressing prostate cancer cell proliferation and reducing tumor growth in vitro and in vivo. | [142] | |
| miR-101 directly inhibits COX-2 translation, and its downregulation correlates with COX-2 overexpression in colorectal cancer cells and patient tissues | [173] | |
| CdCl₂ induces ER stress–driven COX-2/VEGF upregulation causing abnormal angiogenesis and cytotoxicity, which can be rescued by miR-101, siPTGS2, or COX-2 inhibition | [186] | |
| miR-101-3p suppresses breast cancer cell transmigration across the brain endothelium by downregulating COX-2/MMP1 signaling, thereby preserving junctional integrity and reducing brain metastasis potential | [187] | |
| EPA upregulates miR-101 via the 15-LOX-1 pathway, leading to COX-2 suppression and inhibiting colon cancer progression. | [188] | |
| ZEB1 | miR-101a directly targets and suppresses ZEB1, thereby inhibiting epithelial–mesenchymal transition (EMT), reducing invasion and migration, and functioning as a tumor-suppressive regulator. | [189] |
| ZEB1-AS1 promotes CRC proliferation and migration by sponging miR-101 to upregulate ZEB1, while miR-101 restoration or ZEB1-AS1 depletion suppresses these effects. | [190] | |
| miR-101 directly suppresses ZEB1 expression, thereby inhibiting epithelial-to-mesenchymal transition (EMT), reducing cell migration, and limiting metastatic potential. | [191] | |
| M2-TAM–derived EVs deliver NEAT1, which sponges miR-101-3p to upregulate ZEB1/PD-L1, thereby promoting ovarian cancer growth and inducing CD8⁺ T-cell apoptosis. | [192] | |
| MCL-1/2 | miR-101 suppresses Mcl-1, inhibits A549 lung cancer cell growth, and synergistically enhances sensitivity to etoposide by promoting apoptosis. | [193] |
| miR-101 suppresses MCL-1 by targeting its 3′-UTR, thereby promoting apoptosis and enhancing chemosensitivity. | [194] | |
| Overexpression of miR-101 sensitizes papillary thyroid carcinoma cells to TRAIL-induced apoptosis by targeting c-Met and MCL-1 and inhibiting the PI3K/AKT pathway. | [195] | |
| CXCL6 | Overexpression of miR-101-5p suppresses cervical cancer proliferation, migration, invasion, and tumor growth by directly targeting CXCL6 | [196] |
| Overexpression of miR-101-5p suppresses NSCLC growth, invasion, and metastasis by directly targeting CXCL6. | [197] | |
| CXCL12 | miR-101 is downregulated in PTC and suppresses proliferation, survival, migration, and invasion by directly targeting CXCL12 and inhibiting its downstream Akt/EMT signaling | [198] |
| MAT2A | hsa_circ_0007364 drives cervical cancer progression by sponging miR-101-5p, thereby relieving suppression of MAT2A and enhancing tumor cell proliferation, invasion, and growth. | [199] |
| KPNA2 | miR-101-3p is downregulated in cervical squamous cell carcinoma, and its overexpression suppresses tumor cell growth by directly targeting KPNA2. | [200] |
| mTOR | miR-101-3p enhances radiosensitivity of NSCLC by inhibiting the mTOR signaling pathway, reducing survival and promoting apoptosis in irradiated cells. | [201] |
| lncRNA FAM201A decreases radiosensitivity in ESCC by suppressing miR-101a, which in turn upregulates mTOR (and ATM) signaling. | [202] | |
| ABCC1 | Exosomal circ_PIP5K1A promotes NSCLC progression and cisplatin resistance by sponging miR-101a, which normally suppresses ABCC1 expression to inhibit tumor growth and enhance chemosensitivity. | [203] |
| CUL4B | miR-101a directly targets CUL4B, and its suppression by lncRNA SNHG12 enhances CUL4B expression, thereby promoting proliferation, migration, and invasion in non-small cell lung cancer. | [204] |
| circZFR promotes NSCLC progression by sponging miR-101a-3p, thereby relieving its repression of CUL4B and enhancing proliferation, migration, and invasion | [205] | |
| miR-101a directly targets CUL4B, and its overexpression suppresses prostate cancer cell proliferation, migration, and invasion while promoting apoptosis by inhibiting the PI3K/AKT/mTOR pathway | [206] | |
| CDYL | SNHG6 promotes NSCLC growth and invasion by suppressing miR-101a-3p, thereby relieving its inhibition on CDYL. | [207] |
| KRAS | miR-101a directly targets the KRAS 3′UTR, and its repression by circ-MEMO1 leads to KRAS upregulation, thereby promoting proliferation, glycolysis, and tumor growth in NSCLC. | [208] |
| TRIM44 | miR-101-3p directly targets TRIM44, suppressing EMT and thereby reducing proliferation, migration, and invasion of glioblastoma cells. | [209] |
| RAP1A/B | miR-101a suppresses Rap1A expression in prostate cancer, an effect antagonized by lncRNA CRNDE, thereby restraining proliferation, migration, and invasion. | [210] |
| miR-101a suppresses colorectal cancer progression by directly targeting Rap1b, forming a negative feedback loop that inhibits proliferation, migration, and invasion. | [211] | |
| HDAC9 | miR-101a-3p suppresses retinoblastoma cell proliferation by directly targeting HDAC9, and restoring HDAC9 reverses this anti-proliferative effect. | [212] |
| Girdin | miR-101a acts as a tumor suppressor in HCC by directly targeting Girdin, thereby inhibiting cell proliferation, migration, and invasion. | [213] |
| VEGF-C | miR-101a suppresses HCC cell migration and invasion by directly targeting VEGF-C, highlighting its role in restraining cytoskeletal remodeling and girding during metastasis. | [214] |
| miR-101a suppresses VEGF-C, and its inhibition by MALAT1 promotes cisplatin resistance in bladder cancer cells. | [215] | |
| In cholangiocarcinoma, miR-101a is underexpressed, correlating with VEGF overexpression, suggesting its loss may promote tumor progression and reduced survival. | [216] | |
| miR-101a-3p suppresses VEGFA in cancer-associated fibroblasts, thereby reducing CAF-driven EMT, invasion, and metastasis in lung cancer. | [217] | |
| miR-101a suppresses cholangiocarcinoma growth by directly targeting VEGF and indirectly repressing its transcription via COX-2 inhibition, thereby blocking angiogenesis. | [218] | |
| ZO-1 | miR-101a-3p, upregulated by ethanolamine accumulation in obesity, destabilizes ZO-1 mRNA, thereby weakening intestinal barriers and promoting gut permeability, inflammation, and metabolic dysfunction. | [33] |
| CLDN1 | miR-101a suppresses CLDN1 expression to inhibit papillary thyroid carcinoma cell migration and invasion, while XIST promotes metastasis by sponging miR-101a and restoring CLDN1. | [219] |
| ZNF217 | miR-101 suppresses ZNF217 in hepatocellular carcinoma, thereby restoring CDH1 expression and inhibiting proliferation, EMT, and invasion. | [220] |
| c-Met pathway | miR-101a sensitizes papillary thyroid carcinoma cells to TRAIL by targeting c-Met (and MCL-1), thereby inhibiting PI3K/AKT signaling and reducing apoptosis resistance. | [195] |
| c-Met pathway MAPK/ERK pathway |
miR-101a suppresses tumor growth and progression by downregulating pro-angiogenic signaling, such as VEGF-related pathways, through targeting upstream regulators like the HGF/c-MET axis. | [221] |
| miR-101a is downregulated in liver cancer, and its overexpression suppresses proliferation by targeting EZH2 and inhibiting the MAPK/ERK signaling pathway | [222] | |
| miR-101a, enriched in the brain, promotes differentiation of bone marrow cells into microglia-like cells by enhancing inflammatory responsiveness and downregulating MAPK phosphatase-1. | [223] | |
| Lin28B | In NSCLC, IL-1β suppresses miR-101, leading to Lin28B upregulation and enhanced proliferation and migration, an effect reversible by COX-2 inhibition. | [224] |
| ANXA2 | miR-101a suppresses ANXA2, thereby inhibiting ERK signaling, reducing LCSC proliferation and metastasis, and forming a regulatory miR-101/ANXA2/EGR1 loop in liver cancer. | [225] |
| SRF | miR-101a suppresses gastric adenocarcinoma cell proliferation and invasion by directly targeting SRF, thereby downregulating HOTAIR transcription. | [225] |
| PIM 1 | miR-101a suppresses PIM1 expression in gastric cancer cells, thereby inhibiting proliferation and invasion while promoting apoptosis. | [226] |
| FZD4 | miR-101a is downregulated in bladder cancer, and its restoration suppresses migration and invasion by directly targeting FZD4. | [227] |
| STMN1 | miR-101a suppresses pancreatic cancer cell proliferation and invasion by directly targeting and downregulating STMN1. | [80] |
| HIPK3 | miR-101a-3p acts as an oncomiR in colorectal cancer by targeting HIPK3, thereby enhancing cell growth, migration, glycolysis, and reducing chemosensitivity. | [228] |
| Notch1 | miR-101a suppresses NOTCH1 signaling, while CircAPLP2 promotes colorectal cancer proliferation and metastasis by sponging miR-101a and thereby reactivating the Notch pathway. | [229] |
| CREB1 | miR-101a is downregulated in colon cancer, and its overexpression suppresses proliferation and migration by directly targeting CREB1. | [230] |
| BICC1 | miR-101a is downregulated in oral cancer, and its restoration suppresses BICC1, thereby reducing cell viability and promoting apoptosis. | [231] |
| Jak2 | miR-101a suppresses proliferation and induces apoptosis in breast cancer cells by directly targeting Jak2. | [232] |
Abbreviation: ABCC1, ATP-binding cassette subfamily C member 1; AKT, protein kinase B; ANXA2, annexin A2; ATM, ataxia telangiectasia mutated; BICC1, bicaudal C homolog 1; CAF, cancer-associated fibroblast; CDH1 (E-cadherin), epithelial cadherin; CDYL, chromodomain Y-like protein; CLDN1, claudin-1; COX-2 (PTGS2), cyclooxygenase-2 (prostaglandin-endoperoxide synthase 2); CREB1, cAMP response element-binding protein 1; CRC, colorectal cancer; CRNDE, colorectal neoplasia differentially expressed; CUL4B, cullin 4B; CXCL6, C-X-C motif chemokine ligand 6; CXCL12, C-X-C motif chemokine ligand 12; DNMT3A, DNA methyltransferase 3 alpha; EMT, epithelial–mesenchymal transition; EPA, eicosapentaenoic acid; ERK, extracellular signal-regulated kinase; eRMS, embryonal rhabdomyosarcoma; EVs, extracellular vesicles; EZH2, enhancer of zeste homolog 2; FOS, FBJ murine osteosarcoma viral oncogene homolog; FZD4, frizzled class receptor 4; GBM, glioblastoma multiforme; Girdin (CCDC88A), coiled-coil domain-containing protein 88A; HDAC9, histone deacetylase 9; HGF, hepatocyte growth factor; HIF-1α/β, hypoxia-inducible factor-1 alpha/beta; HIPK3, homeodomain-interacting protein kinase 3; HOTAIR, HOX transcript antisense RNA; IL-1β, interleukin-1 beta; JAK2, Janus kinase 2; KPNA2, karyopherin subunit alpha 2; KRAS, Kirsten rat sarcoma viral oncogene homolog; Lin28B, Lin-28 homolog B; MALAT1, metastasis-associated lung adenocarcinoma transcript 1; MAPK, mitogen-activated protein kinase; MAT2A, methionine adenosyl transferase 2A; MCL-1, myeloid cell leukemia sequence 1; MMP1, matrix metalloproteinase-1; mTOR, mechanistic target of rapamycin; NEAT1, nuclear paraspeckle assembly transcript 1; NSCLC, non-small cell lung cancer; PD-L1, programmed death-ligand 1; PI3K, phosphoinositide 3-kinase; PIM1, proviral integration site for Moloney murine leukemia virus 1; RAP1A/B, Ras-related protein 1A/1B; SNHG12, small nucleolar RNA host gene 12; SRF, serum response factor; STMN1, stathmin 1; TAM, tumor-associated macrophage; TCC, transitional cell carcinoma; TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; TRIM44, tripartite motif-containing protein 44; VEGF, vascular endothelial growth factor; VEGF-C, vascular endothelial growth factor C; Wnt/β-catenin, Wingless/Integrated beta-catenin signaling pathway; XIST, X-inactive specific transcript; ZEB1, zinc finger E-box binding homeobox 1; ZEB1-AS1, ZEB1 antisense RNA 1; ZNF217, zinc finger protein 217; ZO-1 (TJP1), zonula occludens-1 (tight junction protein 1).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



