Submitted:
16 February 2026
Posted:
26 February 2026
You are already at the latest version
Abstract
Parkinson’s disease (PD) is a progressive neurodegenerative disorder character-ized by α-synuclein aggregation and degeneration of nigrostriatal dopaminergic neu-rons. Increasing evidence implicates gut microbiota (GM) dysbiosis as a potential con-tributor to PD pathophysiology through bidirectional gut–brain interactions. This narrative review integrates recent taxonomic, functional, metabolomic, im-munological, and interventional evidence linking GM alterations to basal ganglia dys-function. Across multiple cohorts, PD is consistently associated with reduced abun-dance of short-chain fatty acid (SCFA)-producing taxa (e.g., Faecalibacterium, Roseburia) and enrichment of mucin-degrading and pro-inflammatory organisms (e.g., Akkermansia). Shotgun metagenomic studies further reveal alterations in vitamin biosynthesis pathways, carbohydrate metabolism, and microbial network architecture. Metabolomic and Mendelian randomization analyses implicate SCFAs, branched-chain amino acids, bile acids, and GABA metabolism in modulating intestinal permeability, immune activation, and dopaminergic vulnerability. Experimental mi-crobiota transplantation models support mechanistic links involving intestinal barrier disruption, Toll-like receptor signaling, microglial priming, and potential vagal prop-agation of α-syn pathology. Emerging microbiome-targeted strategies—including dietary modulation, probi-otics, prebiotics, fecal microbiota transplantation, and interventions targeting bacterial levodopa metabolism—demonstrate early translational promise. However, inter-cohort heterogeneity and limited longitudinal data highlight the need for precision microbi-ome stratification and prodromal studies. Overall, current multi-level evidence supports gut dysbiosis as a biologically plausible contributor to PD pathogenesis and a promising target for adjunctive nutri-tional and pharmacomicrobiomic interventions.
Keywords:
Parkinson’s disease
; gut microbiota
; microbiota–gut–brain axis
; α-synuclein
; basal ganglia
; short-chain fatty acids
; neuroinflammation
; metabolomics
; fecal microbiota transplantation
; precision nutrition
1. Introduction
Parkinson’s disease (PD) is the second most prevalent neurodegenerative disorder worldwide and is characterized by progressive degeneration of dopaminergic neurons in the substantia nigra pars compacta and intracellular aggregation of misfolded α-synuclein (α-syn) [1,2]. Traditionally conceptualized as a central nervous system disorder, PD is now increasingly viewed as a multisystem disease with significant peripheral involvement [1].
Gastrointestinal dysfunction, particularly constipation, often precedes motor manifestations by up to two decades, suggesting the existence of a prodromal enteric phase [3,6,7]. Braak’s hypothesis proposes a caudo-rostral propagation of misfolded α-syn from the enteric nervous system (ENS) to the dorsal motor nucleus of the vagus and subsequently to midbrain structures [8,9]. Emerging imaging and neuropathological evidence further supports the existence of a “body-first” PD subtype in which peripheral pathology may precede central neurodegeneration [10,11].
The gut microbiota (GM), consisting of trillions of microorganisms, plays essential roles in epithelial barrier integrity, immune modulation, metabolic regulation, and neurotransmitter synthesis [3,12,13]. Dysbiosis, defined as compositional and functional imbalance within microbial communities, has been repeatedly observed in PD across geographically diverse populations [14,15]. However, variability in diet, medication exposure, sequencing platforms, and analytical pipelines has contributed to inter-cohort heterogeneity [8,16].
Recent large-scale meta-analyses integrating 16S rRNA and shotgun metagenomic datasets demonstrate relatively consistent depletion of short-chain fatty acid (SCFA)-producing bacteria and enrichment of mucin-degrading taxa in PD [17,18]. Functional analyses further identify alterations in microbial vitamin biosynthesis pathways, carbohydrate-active enzyme networks, and metabolic gene co-expression patterns [19,20,21,22]. Machine learning-based cross-cohort analyses confirm reproducible functional alterations while underscoring limited cross-study portability [13,23,24].
Beyond compositional shifts, metabolomic profiling and Mendelian randomization approaches implicate microbial-derived metabolites—including SCFAs, bile acids, branched-chain amino acids (BCAAs), and gamma-aminobutyric acid (GABA), as potential mediators of immune activation and dopaminergic vulnerability [25,26,27,28]. Experimental transplantation of PD-associated microbiota into murine models induces intestinal barrier disruption, immune dysregulation, microglial activation, and dopaminergic neuronal loss, supporting mechanistic plausibility [29,30,31].
Collectively, these findings suggest that gut dysbiosis may represent a biologically relevant contributor, rather than a mere epiphenomenon, to basal ganglia dysfunction in PD [14,32], as illustrated in Figure 1. Importantly, microbiome-targeted interventions, including dietary modulation, probiotics, prebiotics, fecal microbiota transplantation, and strategies addressing microbial levodopa metabolism, are emerging as potential adjunctive therapeutic approaches [16,25,33,34].
This review integrates taxonomic, functional, metabolomic, immunological, and translational evidence to delineate mechanistic pathways linking gut microbiota alterations to nigrostriatal degeneration and to critically evaluate emerging microbiome-based therapeutic strategies.
2. Materials and Methods
2.1. Review Design and Literature Search Strategy
This work was conducted as a structured narrative review aimed at integrating recent mechanistic and translational evidence regarding the gut microbiota–brain axis in Parkinson’s disease (PD).
A literature search was performed in PubMed, Web of Science, and Scopus for articles published between January 2022 and January 2026. The following search terms were used in various Boolean combinations: “Parkinson’s disease” AND “gut microbiota” OR “microbiota–gut–brain axis” OR “shotgun metagenomics” OR “metabolomics” OR “Mendelian randomization” OR “fecal microbiota transplantation” OR “levodopa metabolism”.
The search focused on human observational studies, interventional trials, mechanistic animal models, and multi-omics analyses. Non-English publications, conference abstracts, editorials without primary data, and isolated case reports were excluded.
Priority was given to: large-scale meta-analyses, multi-country shotgun metagenomic datasets, multi-omics integrative studies, mendelian randomization analyses, randomized controlled trials, mechanistic microbiota transplantation models.
The objective was to synthesize high-quality, recent evidence rather than exhaustively catalog all published studies.
2.2. Data Categorization and Thematic Integration
Eligible studies were categorized into six mechanistic domains reflecting major pathways linking gut dysbiosis to nigrostriatal degeneration: taxonomic alterations in gut microbiota, functional pathway disruptions (shotgun metagenomics and metatranscriptomics), microbial metabolite alterations, intestinal barrier dysfunction and immune activation, α-synuclein propagation and neuroinflammation, microbiome-targeted therapeutic strategies.
Studies were assigned to domains based on their primary mechanistic focus. Where overlap existed (e.g., metabolomics combined with immune profiling), findings were integrated across relevant sections.
Taxonomic reproducibility was evaluated by comparing 16S rRNA and shotgun datasets across geographically distinct cohorts, with particular attention to findings replicated in multi-cohort meta-analyses and machine learning-based cross-study assessments.
Functional pathway alterations were extracted from metagenomic and network-based analyses, emphasizing gene ontology enrichment, metabolic pathway shifts, and co-expression network changes. Causal inference regarding microbial metabolites was examined using Mendelian randomization (MR) studies and experimental transplantation models.
Mechanistic plausibility was further supported by germ-free, antibiotic-treated, and microbiota-transplanted animal models demonstrating immune activation, barrier dysfunction, or dopaminergic neurodegeneration.
Given the narrative design, no formal risk-of-bias assessment or meta-analytic pooling was performed.
3. Results
3.1. Reproducible Taxonomic Signatures
Multiple cohort studies and recent meta-analyses have identified recurrent patterns of gut microbiota alterations in Parkinson’s disease (PD), although effect sizes and specific taxa vary across populations and analytical platforms [34].
Large-scale shotgun sequencing meta-analyses integrating international datasets report a relative depletion of short-chain fatty acid (SCFA)-producing species, including Faecalibacterium prausnitzii and Roseburia intestinalis, alongside enrichment of mucin-degrading taxa such as Akkermansia muciniphila [35,36,37]. These findings have been partially corroborated by 16S rRNA-based meta-analyses, which also describe reductions in butyrate-producing genera (Faecalibacterium, Roseburia, Coprococcus) and relative increases in taxa such as Akkermansia and Bilophila [38,39,40,41].
Although absolute taxonomic concordance is not universal, depletion of SCFA-producing bacteria represents one of the most frequently replicated observations across geographically distinct cohorts [14,21]. Enrichment of mucin-degrading organisms has likewise been repeatedly described, though the functional implications may depend on host context, dietary patterns, and disease stage [23,31].
A recent machine learning-based meta-analysis comprising 4,489 samples from 22 case–control studies demonstrated that microbiome-based classifiers achieved moderate discriminatory performance within individual cohorts (mean AUC ≈ 0.72), but cross-cohort portability was limited [8,45,46]. These findings underscore substantial geographic, dietary, medication-related, and methodological heterogeneity. Nevertheless, relative depletion of SCFA-producing taxa emerged as a reproducible feature across multiple analytical frameworks [42].
Importantly, taxonomic shifts alone do not establish causality. However, the recurrent reduction of butyrate-producing bacteria and relative enrichment of mucin-degrading and pro-inflammatory taxa provide a biologically plausible substrate for downstream alterations in epithelial barrier function, immune signaling, and microbial metabolite production [39,40].
The principal taxa most consistently reported across meta-analyses and multi-cohort studies are summarized in Table 1.
3.2. Loss of Microbial Functional Diversity
Beyond compositional shifts, Parkinson’s disease (PD) is increasingly characterized by functional alterations within the gut microbiome, as revealed by shotgun metagenomic and multi-omics analyses [20,38,56,57].
Several international meta-analyses report reduced abundance of genes involved in riboflavin and biotin biosynthesis pathways in PD-associated microbiota [16,58,59,60,61]. Given the role of these vitamins in mitochondrial metabolism and redox homeostasis, such alterations may influence host metabolic resilience, although direct causal links remain to be established [62,63,64,65]. Reductions in vitamin biosynthesis pathways have also been correlated with decreased levels of fecal short-chain fatty acids (SCFAs) and polyamines in some cohorts, suggesting coordinated metabolic perturbations [66,67,68,69,70].
Functional pathway analyses further identify decreased representation of carbohydrate-active enzymes (CAZymes), potentially limiting the microbial capacity to ferment complex polysaccharides into SCFAs [71,72,73,74,75]. Alterations in secondary bile acid biosynthesis pathways have also been reported, indicating potential disruption of gut–liver and gut–brain signaling networks [76,77].
In parallel, enrichment of genes associated with xenobiotic metabolism, including pesticide and solvent biotransformation pathways, has been observed in certain PD cohorts, raising hypotheses regarding microbiota-mediated modulation of environmental risk factors [78,79,80,81].
Network-based analyses provide additional insight into systems-level reorganization of microbial communities. Co-expression and gene co-abundance network studies have demonstrated reduced connectivity and altered hub gene structure in PD samples, suggesting decreased functional integration within microbial ecosystems [24,43,59,60,61,62].
Rather than indicating complete “collapse,” these findings reflect disrupted interspecies metabolic interactions and reduced network robustness. Notably, reduced gene expression diversity and altered cross-feeding interactions between commensal taxa have been described in PD cohorts [63,64,65,66].
Collectively, these data indicate that PD-associated dysbiosis extends beyond taxonomic imbalance to encompass alterations in microbial metabolic capacity and ecological network architecture. Such functional reorganization may influence epithelial barrier integrity, immune signaling, and metabolite production, thereby contributing to downstream neuroinflammatory processes explored in subsequent sections [67,68,69,70].
The principal microbial functional pathways reported across shotgun metagenomic studies are summarized in Table 2.
3.3. Microbial Metabolites and Dopaminergic Vulnerability
3.3.1. Short-Chain Fatty Acids (SCFAs)
Short-chain fatty acids (SCFAs), primarily acetate, propionate, and butyrate, are produced through bacterial fermentation of dietary fiber and play important roles in epithelial integrity, immune modulation, and host metabolism [24,25,26,27,83].
Several observational studies report reduced fecal concentrations of SCFAs in patients with Parkinson’s disease (PD), particularly butyrate, although effect sizes vary across cohorts [29,84,85,86].
Experimental studies suggest that butyrate supports epithelial tight junction expression, mucus production, and regulatory immune responses. Therefore, diminished SCFA availability may contribute to increased intestinal permeability and low-grade inflammation, although direct causative evidence in human PD remains limited [59,87,88,89].
In addition to local intestinal effects, SCFAs can influence peripheral immune cells and microglial maturation through histone deacetylase inhibition and G-protein–coupled receptor signaling [90]. Preclinical models demonstrate that SCFA depletion or supplementation can modulate neuroinflammatory responses; however, translation to human dopaminergic neurodegeneration requires further longitudinal investigation [91].
3.3.2. Branched-Chain Amino Acids (BCAAs)
Branched-chain amino acids (BCAAs)—isoleucine, leucine, and valine—have emerged as metabolites of interest in PD-related microbiome research [13,25,26,92,93].
Mendelian randomization (MR) analyses suggest that genetically predicted higher circulating levels of certain BCAAs, particularly isoleucine, are associated with a lower risk of PD [29,80,94,95,96]. While MR strengthens causal inference by leveraging genetic instruments, these findings remain subject to assumptions regarding pleiotropy and instrument validity [91,95]. Thus, the results should be interpreted as supportive rather than definitive evidence of causality.
Specific microbial genera, including Coprococcus, have been associated with modulation of circulating isoleucine levels, suggesting a potential microbiota–metabolite–disease axis [35,97]. Mechanistically, BCAAs may influence mitochondrial function, neurotransmitter synthesis, and systemic metabolic signaling, though their precise contribution to dopaminergic vulnerability remains incompletely defined [95,96,97,98].
3.3.3. Gamma-Aminobutyric Acid (GABA)
SCFAs are small molecules that regulate synthesis of glutamic acids, glutamine and
GABA in human cells. GABA is produced from glutamate in the presence of glutamic acid decarboxylase (GAD) enzyme. This enzyme requires pyridoxal-5′-phosphate (PLP) as a cofactor. The enzyme undergoes a cycle where it alternates between the active holoenzyme and an inactive apoenzyme form through a PLP-dependent mechanism, allowing it to regulate GABA synthesis. Glutamic acid decarboxylase presents two forms (GAD65 and GAD67) which are regulated by specific genes [92,94]. GAD65 and GAD67 belong to thegroup II pyridoxal 5′-phosphate (PLP)-dependent enzymes of fold type I, which comprise two other evolutionarily related human enzymes: aromatic l-amino acid decarboxylase (synonym: l-dopa decarboxylase) and histidine decarboxylase.
Expression of these enzymes in the brain is responsible for the synthesis of the biogenic amines GABA, dopamine, serotonin, and histamine which are implicated in a wide range of biological activities from central homeostatic functions to cognitive phenomena [90]. Propionate, acetate, and butyrate can influence the pH level in biosynthesis of GABA, because the GAD65 is sensitive to different factors such as phosphate-base compounds, pH, magnesium and PLP [93].
Recent meta-analytic evidence has identified enrichment of GABA-consuming bacterial species, including Evtepia gabavorous, in PD cohorts [99,100,101].
GABA is the principal inhibitory neurotransmitter within basal ganglia circuits; however, peripheral and central GABA pools are largely compartmentalized. While gut-derived GABA does not readily cross the blood–brain barrier, alterations in microbial GABA metabolism may influence host physiology indirectly through enteric nervous system signaling, vagal pathways, or immune-mediated mechanisms [97].
Experimental studies suggest that microbiota-derived GABA can modulate stress responses and neuroimmune signaling, but direct evidence linking microbial GABA metabolism to nigrostriatal dysfunction in humans remains preliminary. Therefore, dysregulation of microbial GABA pathways should be considered a plausible but not yet fully validated contributor to PD pathophysiology [100,102].
3.3.4. Xenobiotic and Environmental Pathways
Shotgun metagenomic analyses in certain PD cohorts report enrichment of microbial genes involved in xenobiotic metabolism, including pathways associated with pesticide and solvent biotransformation [34,97,103].
Given established epidemiological associations between pesticide exposure and PD risk, these findings raise the hypothesis that gut microbiota may influence host susceptibility to environmental toxins [104]. Microbial communities can both detoxify and bioactivate xenobiotics, potentially modifying systemic exposure profiles [105].
However, current evidence remains associative, and direct demonstration that microbiota-mediated xenobiotic metabolism alters dopaminergic neurodegeneration in humans is lacking. This area warrants targeted mechanistic and longitudinal investigation [88.102].
The principal microbial-derived metabolites implicated in Parkinson’s disease pathogenesis and their proposed mechanistic roles are summarized in Table 3.
3.4. Intestinal Barrier Dysfunction and Immune Activation
Accumulating evidence suggests that alterations in gut microbiota composition and metabolic output in Parkinson’s disease (PD) may be associated with impaired intestinal barrier integrity and immune activation [2,16,78].
Several clinical studies report increased intestinal permeability and elevated markers of mucosal inflammation in subsets of PD patients [110,111]. Reductions in short-chain fatty acid (SCFA)-producing bacteria, together with enrichment of mucin-degrading taxa, may compromise epithelial tight junction expression and mucus layer maintenance, thereby facilitating increased exposure of the lamina propria to luminal antigens [112,113]. However, the magnitude and stage-dependence of barrier dysfunction remain heterogeneous across cohorts.
Elevated circulating lipopolysaccharide (LPS) levels and increased expression of Toll-like receptor (TLR)-related inflammatory markers have been described in some PD populations, suggesting low-grade systemic endotoxemia [114,115,116]. Activation of TLR signaling pathways can promote nuclear factor κB (NF-κB)-mediated cytokine production, contributing to peripheral inflammatory priming. Nonetheless, direct demonstration of sustained LPS-driven neuroinflammation in human PD remains limited [117].
Preclinical models provide additional mechanistic insight. Transplantation of PD-associated microbiota into germ-free or antibiotic-treated mice has been shown to induce intestinal barrier alterations, increased T helper 17 (Th17) responses, and systemic immune activation [90,118]. In these models, peripheral immune activation is accompanied by microglial reactivity and exacerbation of dopaminergic neuronal vulnerability [119,120].
Microglial priming may represent a key interface between peripheral immune perturbations and central neurodegeneration. Peripheral cytokines and immune mediators can influence microglial phenotype through humoral signaling or neural pathways, potentially lowering the threshold for inflammatory responses within basal ganglia circuits [121]. However, translating findings from animal models to human disease requires cautious interpretation, as direct longitudinal evidence linking gut-derived immune activation to progressive nigrostriatal degeneration in patients is still emerging [122].
Collectively, current data support a model in which microbiota-associated barrier dysfunction and immune signaling constitute plausible intermediates connecting gut dysbiosis to neuroinflammatory processes in PD, while acknowledging that definitive causal pathways remain to be fully delineated [123].
3.5. α-Synuclein Propagation and Basal Ganglia Dysfunction
One influential framework linking gut dysbiosis to central neurodegeneration is the “body-first” hypothesis of Parkinson’s disease (PD), which proposes that misfolded α-synuclein (α-syn) pathology may originate in the enteric nervous system (ENS) and propagate to the brain via the vagus nerve [124].
Neuropathological observations demonstrating α-syn aggregates in enteric neurons and the dorsal motor nucleus of the vagus have supported this model [10,11,125]. Epidemiological studies have reported that truncal vagotomy may be associated with reduced PD risk, although findings are not entirely consistent across cohorts and effect sizes are modest [11,125]. Importantly, accumulating clinical and imaging data suggest the existence of both “body-first” and “brain-first” PD subtypes, indicating that enteric initiation is unlikely to represent a universal mechanism [126].
Experimental models provide mechanistic insight into potential propagation pathways. In rodent studies, injection of misfolded α-syn into the intestinal wall or exposure to pro-inflammatory gut conditions can induce caudo-rostral spread of α-syn pathology to the dorsal motor nucleus and, subsequently, to midbrain dopaminergic regions [43,121,126,127]. However, many of these models rely on supraphysiological α-syn expression or direct fibril inoculation, and therefore may not fully recapitulate sporadic human PD [125,126].
Gut dysbiosis and intestinal inflammation may contribute indirectly to α-syn aggregation. Pro-inflammatory signaling, oxidative stress, and microbial metabolite alterations can promote α-syn misfolding within enteric neurons in preclinical systems [43,121,126].
In parallel, systemic immune activation may prime microglia, lowering the threshold for neuroinflammatory responses within basal ganglia circuits.
Rather than implying a singular linear cascade, current evidence supports a bidirectional and potentially self-reinforcing model: gut-derived immune perturbations and altered microbial metabolites may promote α-syn aggregation and neuroinflammation, while evolving central neurodegeneration may further influence autonomic and gastrointestinal function [128,129].
3.6. Therapeutic Implications
Growing recognition of gut microbiota involvement in Parkinson’s disease (PD) has stimulated interest in microbiome-targeted therapeutic strategies. Current approaches range from dietary modulation to microbial transplantation and pharmacomicrobiomic interventions, though levels of clinical evidence vary substantially [101,104,121].
3.6.1. Diet-Based Interventions
Diet is a major determinant of microbial composition and metabolic output. Higher Healthy Eating Index (HEI)-2015 scores and increased dietary fiber intake have been associated with greater abundance of SCFA-producing bacteria in PD cohorts [129,130]. Reduced consumption of added sugars has been linked to lower relative abundance of pro-inflammatory taxa [131].
Mediterranean-style dietary patterns, characterized by high intake of fiber, polyphenols, and unsaturated fats, have been associated with microbial profiles enriched in SCFA-producing organisms and reduced lipopolysaccharide biosynthesis pathways [132].
While observational data and short-term interventional studies suggest potential benefits on gastrointestinal symptoms and inflammatory markers, robust evidence for disease-modifying effects remains limited. Longitudinal randomized trials are required to determine whether dietary modulation can influence neurodegenerative trajectories [131,132].
3.6.2. Probiotics and Prebiotics
Probiotic and prebiotic supplementation has primarily been investigated for management of constipation and gastrointestinal dysfunction in PD. Randomized controlled trials demonstrate that certain probiotic formulations can improve stool frequency and reduce inflammatory markers [39,133].
Prebiotic supplementation has been shown to increase SCFA-producing bacteria and fecal SCFA levels in small pilot studies [20]. However, strain specificity, dosage, treatment duration, and long-term effects remain heterogeneous across trials. Evidence supporting direct effects on motor progression or dopaminergic neurodegeneration is currently insufficient [20,39].
Thus, while probiotics and prebiotics represent promising adjunctive strategies for symptom management and microbiota modulation, their role in altering disease progression has yet to be established.
3.6.3. Fecal Microbiota Transplantation (FMT)
Fecal microbiota transplantation (FMT) aims to restore microbial diversity and metabolic balance through transfer of donor microbiota. Preliminary open-label studies and small randomized trials report improvements in both motor and non-motor symptoms following FMT in selected PD patients [112,134,135].
However, sample sizes remain small, study designs heterogeneous, and follow-up durations limited. The durability of microbiota engraftment, optimal donor selection criteria, and long-term safety profile require further clarification. At present, FMT should be considered an experimental intervention within controlled research settings rather than a standard therapeutic option [112,134,135,136].
3.6.4. Pharmacomicromodulation
Emerging evidence highlights bidirectional interactions between gut microbes and PD pharmacotherapy. Certain intestinal bacteria express tyrosine decarboxylases capable of converting levodopa to dopamine in the gut lumen, thereby reducing systemic drug bioavailability [112,136].
Strategies targeting bacterial levodopa metabolism—including selective decarboxylase inhibition or engineered probiotic strains—represent innovative approaches to optimizing pharmacokinetics [137]. Preclinical studies demonstrate proof-of-concept improvements in motor performance in animal models; however, clinical translation remains at an early stage [138,139,140,141].
Pharmacomicrobiomic modulation may ultimately enable personalized therapeutic strategies integrating microbial profiling with medication optimization [142].
A summary of microbiome-targeted interventions and current evidence levels is provided in Table 4.
4. Limitations and Future Directions
Despite substantial progress in delineating gut microbiota alterations in Parkinson’s disease (PD), several limitations constrain current interpretation and translational application.
First, most available studies are cross-sectional, limiting the ability to infer temporal directionality. It remains unclear whether gut dysbiosis precedes neurodegeneration, emerges as a consequence of autonomic dysfunction and altered gastrointestinal motility, or reflects bidirectional interactions. Reverse causality is a plausible explanation for at least part of the observed microbial alterations.
Second, medication exposure represents a major confounder. Levodopa, catechol-O-methyltransferase inhibitors, monoamine oxidase-B inhibitors, anticholinergics, and proton pump inhibitors may independently influence microbial composition and metabolic output. Disentangling disease-specific signatures from medication-induced shifts remains challenging.
Third, dietary patterns, which strongly shape microbial ecology, vary across cohorts and are often insufficiently controlled. Geographic and cultural heterogeneity further contribute to limited cross-study portability of microbiome classifiers.
Fourth, most studies rely on 16S rRNA sequencing, which provides genus-level resolution but limited functional insight. While shotgun metagenomics improves pathway-level characterization, strain-level functional heterogeneity remains underexplored. Opposing metabolic effects may occur within the same genus, underscoring the need for higher-resolution strain profiling and metatranscriptomic validation.
Fifth, mechanistic extrapolation from animal models requires caution. Although microbiota transplantation and α-syn propagation studies provide proof-of-concept evidence, experimental systems frequently involve supraphysiological α-syn expression, direct fibril inoculation, or germ-free conditions that may not fully replicate sporadic human PD.
Future research should prioritize longitudinal prodromal cohorts to clarify temporal dynamics and identify early microbial signatures preceding motor onset. Integration of host genomics, metabolomics, immune profiling, and microbiome data through systems biology approaches may enable identification of patient-specific microbial–host interaction networks.
Strain-level functional characterization, causal mediation analyses, and standardized reporting frameworks will be essential to improve reproducibility. Ultimately, precision microbiome stratification—rather than universal microbial targets—may represent the most realistic path toward disease-modifying microbiota-based interventions.
5. Conclusions
Emerging multi-level evidence from taxonomic profiling, shotgun metagenomics, metabolomics, Mendelian randomization analyses, and experimental models supports a biologically plausible framework in which gut microbiota alterations may influence key pathways implicated in Parkinson’s disease (PD). Recurrent depletion of short-chain fatty acid–producing taxa, functional metabolic reorganization, intestinal barrier perturbation, immune activation, and potential α-synuclein propagation together suggest that the gut–brain interface represents a relevant axis in PD pathophysiology.
However, substantial heterogeneity persists across cohorts, disease stages, and methodological platforms. Current data do not support a singular or universal microbial signature, nor do they establish definitive causality between dysbiosis and nigrostriatal degeneration. Rather, gut microbiota alterations likely interact dynamically with host genetics, immune responses, environmental exposures, and pharmacotherapy within a bidirectional system.
Translational strategies—including dietary modulation, probiotic and prebiotic supplementation, fecal microbiota transplantation, and pharmacomicrobiomic optimization of levodopa metabolism—demonstrate early promise but require larger, longitudinal, and mechanistically informed clinical trials.
Future research should focus on prodromal cohorts, strain-level functional resolution, and integrated host–microbiome multi-omics modeling. Precision microbiome stratification may ultimately enable individualized interventions that complement conventional dopaminergic therapies and potentially modify disease trajectories.
While microbiota modulation is unlikely to represent a standalone cure, it may emerge as a biologically rational component of comprehensive, systems-based management of Parkinson’s disease.
Author Contributions
Conceptualization, M.I., A.B. and E.R.; methodology, M.I., and A.B.; software, M.I.; validation, M.I., A.B. and E.R.; formal analysis, M.I.; investigation, A.B.; resources, M.I., A.B., and E.R.; data curation, M.I. and E.R.; writing—original draft preparation, A.B.; writing—review and editing, M.I.; visualization, M.I. and E.R.; supervision, E.R.; project administration, E.R.; funding acquisition, M.I., A.B. and E.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bai, H.; Ma, W.; Zhu, L.; Lu, Y.; Fan, J.; Chen, M.; Huang, C. Updates on Parkinson’s Disease. Neuropsychiatr. Dis. Treat. 2025, Volume 21, 1945–1953. [Google Scholar] [CrossRef]
- Cannon, T.; Gruenheid, S. Microbes and Parkinson’s Disease: From Associations to Mechanisms. Trends Microbiol. 2022, 30, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Abbas, K.; Mustafa, M.; Usmani, N.; Habib, S. Microbiome-Based Therapies for Parkinson’s Disease. Front. Nutr. 2024, 11, 1496616. [Google Scholar] [CrossRef]
- Benvenuti, L.; Di Salvo, C.; Bellini, G.; Seguella, L.; Rettura, F.; Esposito, G.; Antonioli, L.; Ceravolo, R.; Bernardini, N.; Pellegrini, C.; et al. Gut-Directed Therapy in Parkinson’s Disease. Front. Pharmacol. 2024, 15, 1407925. [Google Scholar] [CrossRef] [PubMed]
- Iordache, M.P.; Buliman, A.; Costea-Firan, C.; Gligore, T.C.I.; Cazacu, I.S.; Stoian, M.; Teoibaș-Şerban, D.; Blendea, C.-D.; Protosevici, M.G.-I.; Tanase, C.; et al. Immunological and Inflammatory Biomarkers in the Prognosis, Prevention, and Treatment of Ischemic Stroke: A Review of a Decade of Advancement. Int. J. Mol. Sci. 2025, 26, 7928. [Google Scholar] [CrossRef] [PubMed]
- Ai, P.; Xu, S.; Yuan, Y.; Xu, Z.; He, X.; Mo, C.; Zhang, Y.; Yang, X.; Xiao, Q. Targeted Gut Microbiota Modulation Enhances Levodopa Bioavailability and Motor Recovery in MPTP Parkinson’s Disease Models. Int. J. Mol. Sci. 2025, 26, 5282. [Google Scholar] [CrossRef]
- Alfonsetti, M.; Castelli, V.; d’Angelo, M. Are We What We Eat? Impact of Diet on the Gut–Brain Axis in Parkinson’s Disease. Nutrients 2022, 14, 380. [Google Scholar] [CrossRef]
- Beisembayeva, M.; Shayakhmetova, Y.; Muratbekova, S.; Li, I.; Yakupov, E.; Kaiyrzhanov, R.; Grigolashvili, M. A Comparative Analysis of the Gut Microbiota in Patients With Parkinson’s Disease Worldwide: A Systematic Review. Int. J. Clin. Pract. 2025, 2025, 5511146. [Google Scholar] [CrossRef]
- Camacho, M.; Greenland, J.C.; Daruwalla, C.; Scott, K.M.; Patel, B.; Apostolopoulos, D.; Ribeiro, J.; O’Reilly, M.; Hu, M.T.; Williams-Gray, C.H. The Profile of Gastrointestinal Dysfunction in Prodromal to Late-Stage Parkinson’s Disease. Npj Park. Dis. 2025, 11, 123. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.; Baek, J.W.; Jung, K.-Y.; Lee, Y.; Koh, A.; Kim, H.-J. Enrichment of Gut-Derived Metabolites in a Parkinson’s Disease Subtype with REM Sleep Behavior Disorder. Npj Park. Dis. 2025, 11, 189. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Xie, R.; Feng, Y.; Zhang, M.-N.; He, L.; Yang, B.; Wang, H.-G.; Yang, X.-Z. Gut Microbiota Helps Identify Clinical Subtypes of Parkinson’s Disease. Mil. Med. Res. 2024, 11, 42. [Google Scholar] [CrossRef]
- Abou Izzeddine, N.; Ahmad, K.; Bacha, C.; Jabbour, M.; Najjar, M.; Salhab, S.; Ghadieh, H.E.; Kanaan, A.; Azar, S.; Khattar, Z.A.; et al. The Microbial Guardians: Unveiling the Role of Gut Microbiota in Shaping Neurodegenerative Disease. IBRO Neurosci. Rep. 2025, 19, 17–37. [Google Scholar] [CrossRef]
- Agostini, D.; Bartolacci, A.; Rotondo, R.; De Pandis, M.F.; Battistelli, M.; Micucci, M.; Potenza, L.; Polidori, E.; Ferrini, F.; Sisti, D.; et al. Homocysteine, Nutrition, and Gut Microbiota: A Comprehensive Review of Current Evidence and Insights. Nutrients 2025, 17, 1325. [Google Scholar] [CrossRef] [PubMed]
- Lima, I.S.; Pêgo, A.C.; Martins, A.C.; Prada, A.R.; Barros, J.T.; Martins, G.; Gozzelino, R. Gut Dysbiosis: A Target for Protective Interventions against Parkinson’s Disease. Microorganisms 2023, 11, 880. [Google Scholar] [CrossRef]
- Pfaffinger, J.M.; Hays, K.E.; Seeley, J.; Ramesh Babu, P.; Ryznar, R. Gut Dysbiosis as a Potential Driver of Parkinson’s and Alzheimer’s Disease Pathogenesis. Front. Neurosci. 2025, 19, 1600148. [Google Scholar] [CrossRef]
- Ayten, Ş.; Bilici, S. Modulation of Gut Microbiota Through Dietary Intervention in Neuroinflammation and Alzheimer’s and Parkinson’s Diseases. Curr. Nutr. Rep. 2024, 13, 82–96. [Google Scholar] [CrossRef]
- Nishiwaki, H.; Ueyama, J.; Ito, M.; Hamaguchi, T.; Takimoto, K.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Mori, H.; Kurokawa, K.; et al. Meta-Analysis of Shotgun Sequencing of Gut Microbiota in Parkinson’s Disease. Npj Park. Dis. 2024, 10, 106. [Google Scholar] [CrossRef]
- Shalash, A.; Ezzeldin, S.; Hashish, S.; Salah, Y.; Dawood, N.L.; Moustafa, A.; Salama, M. Gut Microbial Shifts toward Inflammation in Parkinson’s Disease: Insights from Pilot Shotgun Metagenomics Egyptian Cohort. J. Park. Dis. 2025, 15, 1540–1543. [Google Scholar] [CrossRef]
- Nakhal, M.M.; Yassin, L.K.; Alyaqoubi, R.; Saeed, S.; Alderei, A.; Alhammadi, A.; Alshehhi, M.; Almehairbi, A.; Al Houqani, S.; BaniYas, S.; et al. The Microbiota–Gut–Brain Axis and Neurological Disorders: A Comprehensive Review. Life 2024, 14, 1234. [Google Scholar] [CrossRef]
- Bedarf, J.R.; Romano, S.; Heinzmann, S.S.; Duncan, A.; Traka, M.H.; Ng, D.; Segovia-Lizano, D.; Simon, M.-C.; Narbad, A.; Wüllner, U.; et al. A Prebiotic Dietary Pilot Intervention Restores Faecal Metabolites and May Be Neuroprotective in Parkinson’s Disease. Npj Park. Dis. 2025, 11, 66. [Google Scholar] [CrossRef]
- Department of Nutrition and Dietetics, Faculty of Health Sciences, Üsküdar University, Istanbul, Turkey.; Atak, E.S.; Yıldız, D.; Department of Nutrition and Dietetics, Faculty of Health Sciences, Üsküdar University, Istanbul, Turkey.; Kocatürk, R.R.; Department of Nutrition and Dietetics, Faculty of Health Sciences, Üsküdar University, Istanbul, Turkey.; Temizyürek, A.; Department of Physiology, School of Medicine, Koç University, Istanbul, Turkey.; Özcan, Ö.Ö.; Department of Molecular Neuroscience, Health Sciences Institute, Üsküdar University, Istanbul, Turkey.; et al. Therapeutic Targets of Probiotics in Parkinson Disease: A Systematic Review of Randomized Controlled Trials. Basic Clin. Neurosci. J. 2024, 15, 165–174. [CrossRef]
- Xiromerisiou, G.; Marogianni, C.; Androutsopoulou, A.; Ntavaroukas, P.; Mysiris, D.; Papoutsopoulou, S. Parkinson’s Disease, It Takes Guts: The Correlation between Intestinal Microbiome and Cytokine Network with Neurodegeneration. Biology 2023, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Wirbel, J.; Ansorge, R.; Schudoma, C.; Ducarmon, Q.R.; Narbad, A.; Zeller, G. Machine Learning-Based Meta-Analysis Reveals Gut Microbiome Alterations Associated with Parkinson’s Disease. Nat. Commun. 2025, 16, 4227. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, S.A.; Chaudhary, S.; Rawat, S. Understanding Parkinson’s Disease: Current Trends and Its Multifaceted Complications. Front. Aging Neurosci. 2025, 17, 1617106. [Google Scholar] [CrossRef]
- Choe, U. Role of Dietary Fiber and Short-Chain Fatty Acids in Preventing Neurodegenerative Diseases through the Gut-Brain Axis. J. Funct. Foods 2025, 129, 106870. [Google Scholar] [CrossRef]
- Hegelmaier, T.; Duscha, A.; Desel, C.; Fuchs, S.; Shapira, M.; Amidror, S.; Shan, Q.; Stangl, G.I.; Hirche, F.; Kempa, S.; et al. Supplementation with Short-Chain Fatty Acids and a Prebiotic Improves Clinical Outcome in Parkinson’s Disease: A Randomized Double-Blind Prospective Study. Sci. Rep. 2025, 16, 315. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Cheng, G.; Hardy, M. Gut Microbiome, Short-Chain Fatty Acids, Alpha-Synuclein, Neuroinflammation, and ROS/RNS: Relevance to Parkinson’s Disease and Therapeutic Implications. Redox Biol. 2024, 71, 103092. [Google Scholar] [CrossRef]
- Rusu, E.; Necula, L.G.; Neagu, A.I.; Alecu, M.; Stan, C.; Albulescu, R.; Tanase, C.P. Current Status of Stem Cell Therapy: Opportunities and Limitations. Turk. J. Biol. 2016, 40, 955–967. [Google Scholar] [CrossRef]
- Eslami, M.; Adampour, Z.; Fadaee Dowlat, B.; Yaghmayee, S.; Motallebi Tabaei, F.; Oksenych, V.; Naderian, R. A Novel Frontier in Gut–Brain Axis Research: The Transplantation of Fecal Microbiota in Neurodegenerative Disorders. Biomedicines 2025, 13, 915. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Gao, H.; Wang, Y.; Xiang, Y. Exploring the Role of Gut Microbiota in Parkinson’s Disease: Insights from Fecal Microbiota Transplantation. Front. Neurosci. 2025, 19, 1574512. [Google Scholar] [CrossRef]
- Nabil, Y.; Helal, M.M.; Qutob, I.A.; Dawoud, A.I.A.; Allam, S.; Haddad, R.; Manasrah, G.M.; AlEdani, E.M.; Sleibi, W.; Faris, A.; et al. Efficacy and Safety of Fecal Microbiota Transplantation in the Management of Parkinson’s Disease: A Systematic Review. BMC Neurol. 2025, 25, 291. [Google Scholar] [CrossRef]
- Denman, C.R.; Park, S.M.; Jo, J. Gut-Brain Axis: Gut Dysbiosis and Psychiatric Disorders in Alzheimer’s and Parkinson’s Disease. Front. Neurosci. 2023, 17, 1268419. [Google Scholar] [CrossRef]
- Blendea, C.-D.; Khan, M.T.; Stoian, M.; Gligore, T.C.I.; Cuculici, Ștefan; Stanciu, I.L.; Protosevici, M.G.-I.; Iordache, M.; Buliman, A.; Costea-Firan, C.; et al. Advances in Minimally Invasive Treatments for Prostate Cancer: A Review of the Role of Ultrasound Therapy and Laser Therapy. Balneo PRM Res. J. 2025, 16, 827–827. [Google Scholar] [CrossRef]
- Jin, Y.; Wang, L.; Lin, R.; He, J.; Liu, D.; Liu, Y.; Deng, Y. Synergistic Effects of Plant Polysaccharides and Probiotics: A Novel Dietary Approach for Parkinson’s Disease Intervention. Pharmaceuticals 2026, 19, 157. [Google Scholar] [CrossRef]
- Miyaue, N.; Yamamoto, H.; Liu, S.; Ito, Y.; Yamanishi, Y.; Ando, R.; Suzuki, Y.; Mogi, M.; Nagai, M. Association of Enterococcus Faecalis and Tyrosine Decarboxylase Gene Levels with Levodopa Pharmacokinetics in Parkinson’s Disease. Npj Park. Dis. 2025, 11, 49. [Google Scholar] [CrossRef]
- De Sciscio, M.; Bryant, R.V.; Haylock-Jacobs, S.; Day, A.S.; Pitchers, W.; Iansek, R.; Costello, S.P.; Kimber, T.E. Faecal Microbiota Transplant in Parkinson’s Disease: Pilot Study to Establish Safety & Tolerability. Npj Park. Dis. 2025, 11, 203. [Google Scholar] [CrossRef]
- Chui, Z.S.W.; Chan, L.M.L.; Zhang, E.W.H.; Liang, S.; Choi, E.P.H.; Lok, K.Y.W.; Tun, H.M.; Kwok, J.Y.Y. Effects of Microbiome-Based Interventions on Neurodegenerative Diseases: A Systematic Review and Meta-Analysis. Sci. Rep. 2024, 14, 9558. [Google Scholar] [CrossRef]
- Christopher, C.; Morgan, K.; Tolleson, C.; Trudell, R.; Fernandez-Romero, R.; Rice, L.; Abiodun, B.; Vickery, Z.; Jones, K.; Woodall, B.; et al. Specific Bacterial Taxa and Their Metabolite, DHPS, May Be Linked to Gut Dyshomeostasis in Patients with Alzheimer’s Disease, Parkinson’s Disease, and Amyotrophic Lateral Sclerosis. Nutrients 2025, 17, 1597. [Google Scholar] [CrossRef] [PubMed]
- Forero-Rodríguez, J.; Zimmermann, J.; Taubenheim, J.; Arias-Rodríguez, N.; Caicedo-Narvaez, J.D.; Best, L.; Mendieta, C.V.; López-Castiblanco, J.; Gómez-Muñoz, L.A.; Gonzalez-Santos, J.; et al. Changes in Bacterial Gut Composition in Parkinson’s Disease and Their Metabolic Contribution to Disease Development: A Gut Community Reconstruction Approach. Microorganisms 2024, 12, 325. [Google Scholar] [CrossRef]
- Li, Z.; Liang, H.; Hu, Y.; Lu, L.; Zheng, C.; Fan, Y.; Wu, B.; Zou, T.; Luo, X.; Zhang, X.; et al. Gut Bacterial Profiles in Parkinson’s Disease: A Systematic Review. CNS Neurosci. Ther. 2023, 29, 140–157. [Google Scholar] [CrossRef]
- Soto-Avellaneda, A.; Prigent, A.; Meyerdirk, L.; Schautz, N.; Pospisilik, J.A.; Brundin, L.; Henderson, M.X. Helicobacter Pylori Infection and α-Synuclein Pathology Drive Parallel Neurodegenerative Pathways in the Substantia Nigra. J. Neuroinflammation 2025, 22, 293. [Google Scholar] [CrossRef]
- Rust, C.; Van Den Heuvel, L.L.; Bardien, S.; Carr, J.; Pretorius, E.; Seedat, S.; Hemmings, S.M.J. Association between the Relative Abundance of Butyrate-Producing and Mucin-Degrading Taxa and Parkinson’s Disease. Neuroscience 2025, 576, 149–154. [Google Scholar] [CrossRef]
- Balsamo, J.M.; Yan, Y.; Thai, D.; Cologna, S.M.; Bess, E.N. Multiomic Analysis Reveals Molecular Pathways Associated with Intestinal Aggregation of α-Synuclein. ACS Chem. Biol. 2026, 21, 83–95. [Google Scholar] [CrossRef]
- Jia, Y.; Zhang, Y.; Tai, X.; Zhao, T.; Zhang, H.; Zhou, H. Research Progress on Natural Products in Regulating the Gut Microbiota in Parkinson’s Disease. Front. Pharmacol. 2025, 16, 1667694. [Google Scholar] [CrossRef]
- Bai, F.; You, L.; Lei, H.; Li, X. Association between Increased and Decreased Gut Microbiota Abundance and Parkinson’s Disease: A Systematic Review and Subgroup Meta-Analysis. Exp. Gerontol. 2024, 191, 112444. [Google Scholar] [CrossRef] [PubMed]
- Clasen, F.; Yildirim, S.; Arıkan, M.; Garcia-Guevara, F.; Hanoğlu, L.; Yılmaz, N.H.; Şen, A.; Celik, H.K.; Neslihan, A.A.; Demir, T.K.; et al. Microbiome Signatures of Virulence in the Oral-Gut-Brain Axis Influence Parkinson’s Disease and Cognitive Decline Pathophysiology. Gut Microbes 2025, 17, 2506843. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-X.; Sun, N.-Q.; Mo, S.-J. Rhapontin Activating Nuclear Factor Erythroid 2-Related Factor 2 to Ameliorate Parkinson’s Disease-Associated Gastrointestinal Dysfunction. World J. Gastroenterol. 2026, 32. [Google Scholar] [CrossRef]
- Barbu, L.A.; Vasile, L.; Cercelaru, L.; Șurlin, V.; Mogoantă, S.-Ștefaniță; Mogoș, G.F.R.; Țenea Cojan, T.S.; Mărgăritescu, N.-D.; Iordache, M.P.; Buliman, A. Aggressiveness in Well-Differentiated Small Intestinal Neuroendocrine Tumors: A Rare Case and Narrative Literature Review. J. Clin. Med. 2025, 14, 5821. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xue, L.; Zhang, M.; Shen, P.; Zhao, W.; Tong, Q.; Wu, S.; Dai, W.; Yang, X.; Wang, H. Colonoscopic Fecal Microbiota Transplantation for Mild-to-Moderate Parkinson’s Disease: A Randomized Controlled Trial. Brain. Behav. Immun. 2025, 130, 106086. [Google Scholar] [CrossRef]
- Xie, L.; Chen, D.; Zhu, X.; Cheng, C. Efficacy and Safety of Probiotics in Parkinson’s Constipation: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2023, 13, 1007654. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Pang, D.; Shang, H. Akkermansia Muciniphila: A Double-Edged Sword in Life-Stage-Specific Nutritional Modulation of Parkinson’s Disease via the Gut-Brain Axis. Microbiol. Res. 2026, 305, 128436. [Google Scholar] [CrossRef]
- Buliman, A.; Iordache, M.P.; Protosevici, M.I.; Tanase, C. Therapeutic Strategies Targeting Anti- CD47 Therapies in Glioblastoma Multiforme: Lead or Dead End? J. Cell. Mol. Med. 2025, 29, e70889. [Google Scholar] [CrossRef]
- Albani, G.; Chellamuthu, V.R.; Morlacchi, L.; Zirone, F.; Youssefi, M.; Giardini, M.; Chao, Y.-X.; Tan, E.-K.; Albani, S. Gut Microbiota and Dopamine: Producers, Consumers, Enzymatic Mechanisms, and In Vivo Insights. Bioengineering 2025, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Marzouk, N.H.; Rashwan, H.H.; El-Hadidi, M.; Ramadan, R.; Mysara, M. Proinflammatory and GABA Eating Bacteria in Parkinson’s Disease Gut Microbiome from a Meta-Analysis Perspective. Npj Park. Dis. 2025, 11, 145. [Google Scholar] [CrossRef] [PubMed]
- Camberos-Barraza, J.; Guadrón-Llanos, A.M.; De La Herrán-Arita, A.K. The Gut Microbiome-Neuroglia Axis: Implications for Brain Health, Inflammation, and Disease. Neuroglia 2024, 5, 254–273. [Google Scholar] [CrossRef]
- Buliman, A.; Iordache, M.P.; Protosevici, M.G.-I.; Coroescu, M.-M.; Oncioiu, I.; Popa, M.-L.; Bondar, A.-C. What’s Left When All Is Gone? Limitations of the ABCD2 Score in Transient Ischemic Attacks. Balneo PRM Res. J. 2025, No. 4. [Google Scholar] [CrossRef]
- Cannas, F.; Kopeć, K.K.; Zuddas, N.; Cesare Marincola, F.; Arcara, G.; Loi, M.; Mussap, M.; Fanos, V. Parkinson’s Disease Through the Lens of Metabolomics: A Targeted Systematic Review on Human Studies (2019–2024). J. Clin. Med. 2025, 14, 6277. [Google Scholar] [CrossRef]
- Buliman, A.; Chiotoroiu, A.L.; Panchici, T.-A.; Cintacioiu, D.; Parasca, S.V.; Boiangiu, I.C.; Iordache, M.P. Modified Meek Technique Using Pre-Folded Polyamide Gauzes:A 10-Patient Case Series with Extensive Burns. Ind. Textila 2025, 76, 731–736. [Google Scholar] [CrossRef]
- Bolen, M.L.; Buendia, M.; Shi, J.; Staley, H.; Kachergus, J.M.; Efron, P.A.; Park, G.; Nagpal, R.; Alvarez, S.D.; Xue, Q.-S.; et al. Spatial Single-Cell Multiomics Reveals Peripheral Immune Dysfunction in Parkinson’s and Inflammatory Bowel Disease. Npj Park. Dis. 2026, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Chen, Q.; Zhang, Y.; Asakawa, T. Multidirectional Associations between the Gut Microbiota and Parkinson’s Disease, Updated Information from the Perspectives of Humoral Pathway, Cellular Immune Pathway and Neuronal Pathway. Front. Cell. Infect. Microbiol. 2023, 13, 1296713. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Liu, Y.; Yang, B.; Yin, P.; Guan, C.; Fang, Y.; Yang, L.; Zan, K.; Cui, G.; et al. Alterations in Gut Microbiota and Plasma Metabolites: A Multi-Omics Study of Mild Cognitive Impairment in Parkinson’s Disease. Front. Neurosci. 2025, 19, 1667331. [Google Scholar] [CrossRef]
- Roy, S.; Chattopadhyay, D.; Choudhury, L.; Ghosh, S.; Mahajan, A.A. Harnessing the Power of Faecal Microbiota Transplantation: Optimizing Neuroimmune Function for Improved Treatment of Parkinson’s, Alzheimer’s, and Multiple Sclerosis. Discov. Neurosci. 2025, 20, 21. [Google Scholar] [CrossRef]
- Shafieinouri, M.; Hong, S.; Lee, P.S.; Grant, S.M.; Khani, M.; Dadu, A.; Schumacher Schuh, A.F.; Makarious, M.B.; Sandon, R.; Simmonds, E.; et al. Gut-Brain Nexus: Mapping Multimodal Links to Neurodegeneration at Biobank Scale. Sci. Adv. 2025, 11, eadu2937. [Google Scholar] [CrossRef]
- Villette, R.; Ortís Sunyer, J.; Novikova, P.V.; Aho, V.T.E.; Petrov, V.A.; Hickl, O.; Busi, S.B.; De Rudder, C.; Kunath, B.J.; Heintz-Buschart, A.; et al. Integrated Multi-Omics Highlights Alterations of Gut Microbiome Functions in Prodromal and Idiopathic Parkinson’s Disease. Microbiome 2025, 13, 200. [Google Scholar] [CrossRef] [PubMed]
- Constantinoiu, S.; Cochior, D. Severe Acute Pancreatitis - Determinant Factors and Current Therapeutic Conduct. Chirurgia (Bucur.) 2018, 113, 385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lai, Y.; Darweesh, S.K.L.; Bloem, B.R.; Forsgren, L.; Hansen, J.; Katzke, V.A.; Masala, G.; Sieri, S.; Sacerdote, C.; et al. Gut Microbial Metabolites and Future Risk of Parkinson’s Disease: A Metabolome-Wide Association Study. Mov. Disord. 2025, 40, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Wang, H.; Song, J. Gut-Brain Axis Modulation in Remote Rehabilitation of Parkinson’s Disease: Reconstructing the Fecal Metabolome and Nigral Network Connectivity. Front. Neurol. 2025, 16, 1644490. [Google Scholar] [CrossRef] [PubMed]
- Villette, R.; Novikova, P.V.; Laczny, C.C.; Mollenhauer, B.; May, P.; Wilmes, P. Human Gut Microbiome Gene Co-Expression Network Reveals a Loss in Taxonomic and Functional Diversity in Parkinson’s Disease. Npj Biofilms Microbiomes 2025, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Buliman, A.; Iordache, M.P.; Bondar, A.-C.; Protosevici, M.G.-I.; Oncioiu, I.; Popa, M.-L. From Basic Blood Counts to Functional Outcomes: Neutrophil– Lymphocyte Ratio as a Predictor of Disability in Acute and Subacute Ischemic Stroke. Balneo PRM Res. J. 2025, No. 4. [Google Scholar] [CrossRef]
- Fan, W.; Li, K.; Wang, R.; Chen, R.; Liu, Y.; Yang, Z.; Zhao, N.; Yan, J. Vasoactive Intestinal Peptide: A Neuropeptide That Plays an ImportantRole in Parkinson’s Disease. Curr. Neuropharmacol. 2025, 23. [Google Scholar] [CrossRef]
- Bondar, A.-C.; Iordache, M.P.; Coroescu, M.; Buliman, A.; Rusu, E.; Budișteanu, M.; Tanase, C. Unlocking the Sugar Code: Implications and Consequences of Glycosylation in Alzheimer’s Disease and Other Tauopathies. Biomedicines 2025, 13, 2884. [Google Scholar] [CrossRef]
- Caradonna, E.; Nemni, R.; Bifone, A.; Gandolfo, P.; Costantino, L.; Giordano, L.; Mormone, E.; Macula, A.; Cuomo, M.; Difruscolo, R.; et al. The Brain–Gut Axis, an Important Player in Alzheimer and Parkinson Disease: A Narrative Review. J. Clin. Med. 2024, 13, 4130. [Google Scholar] [CrossRef]
- Moldovan, C.; Cochior, D.; Gorecki, G.; Rusu, E.; Ungureanu, F.-D. Clinical and Surgical Algorithm for Managing Iatrogenic Bile Duct Injuries during Laparoscopic Cholecystectomy: A Multicenter Study. Exp. Ther. Med. 2021, 22, 1385. [Google Scholar] [CrossRef]
- Tanase, C.; Cruceru, M.L.; Enciu, A.-M.; Popa, A.C.; Albulescu, R.; Neagu, M.; Constantinescu, S.N. Signal Transduction Molecule Patterns Indicating Potential Glioblastoma Therapy Approaches. OncoTargets Ther. 2013, 1737. [Google Scholar] [CrossRef]
- Mendonça, I.P.; Peixoto, C.A. The Double-Edged Sword: The Complex Function of Enteric Glial Cells in Neurodegenerative Diseases. J. Neurochem. 2025, 169, e70069. [Google Scholar] [CrossRef]
- Oliver, P.J.; Civitelli, L.; Hu, M.T. The Gut–Brain Axis in Early Parkinson’s Disease: From Prodrome to Prevention. J. Neurol. 2025, 272, 413. [Google Scholar] [CrossRef] [PubMed]
- Templeton, H.N.; Tobet, S.A.; Schwerdtfeger, L.A. Gut Neuropeptide Involvement in Parkinson’s Disease. Am. J. Physiol.-Gastrointest. Liver Physiol. 2025, 328, G716–G733. [Google Scholar] [CrossRef] [PubMed]
- Simionescu, O.; Tudorache, S.I. Autoimmune Pemphigus: Difficulties in Diagnosis and the Molecular Mechanisms Underlying the Disease. Front. Immunol. 2025, 16, 1481093. [Google Scholar] [CrossRef] [PubMed]
- Cord, D.; Rîmbu, M.C.; Iordache, M.P.; Albulescu, R.; Pop, S.; Tanase, C.; Popa, M.-L. Phytochemicals as Epigenetic Modulators in Chronic Diseases: Molecular Mechanisms. Molecules 2025, 30, 4317. [Google Scholar] [CrossRef]
- Adhikary, K.; Ganguly, K.; Sarkar, R.; Abubakar, Md.; Banerjee, P.; Karak, P. Phytonutrients and Their Neuroprotective Role in Brain Disorders. Front. Mol. Biosci. 2025, 12, 1607330. [Google Scholar] [CrossRef]
- Jaberi, K.R.; Alamdari-palangi, V.; Savardashtaki, A.; Vatankhah, P.; Jamialahmadi, T.; Tajbakhsh, A.; Sahebkar, A. Modulatory Effects of Phytochemicals on Gut–Brain Axis: Therapeutic Implication. Curr. Dev. Nutr. 2024, 8, 103785. [Google Scholar] [CrossRef] [PubMed]
- Lupu, M.; Gradisteanu Pircalabioru, G.; Chifiriuc, M.; Albulescu, R.; Tanase, C. Beneficial Effects of Food Supplements Based on Hydrolyzed Collagen for Skin Care (Review). Exp. Ther. Med. 2019. [Google Scholar] [CrossRef]
- Mocanu, A.-I.; Mocanu, H.; Moldovan, C.; Soare, I.; Niculet, E.; Tatu, A.L.; Vasile, C.I.; Diculencu, D.; Postolache, P.A.; Nechifor, A. Some Manifestations of Tuberculosis in Otorhinolaryngology – Case Series and a Short Review of Related Data from South-Eastern Europe. Infect. Drug Resist. 2022, Volume 15, 2753–2762. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Sun, L.; Liu, Y.; Chen, R. Safety and Efficacy of Fecal Microbiota Transplantation in the Treatment of Parkinson’s Disease: A Systematic Review of Clinical Trials. Front. Neurosci. 2025, 19, 1639911. [Google Scholar] [CrossRef]
- Cheng, Y.; Tan, G.; Zhu, Q.; Wang, C.; Ruan, G.; Ying, S.; Qie, J.; Hu, X.; Xiao, Z.; Xu, F.; et al. Efficacy of Fecal Microbiota Transplantation in Patients with Parkinson’s Disease: Clinical Trial Results from a Randomized, Placebo-Controlled Design. Gut Microbes 2023, 15, 2284247. [Google Scholar] [CrossRef] [PubMed]
- Duru, I.C.; Lecomte, A.; Shishido, T.K.; Laine, P.; Suppula, J.; Paulin, L.; Scheperjans, F.; Pereira, P.A.B.; Auvinen, P. Metagenome-Assembled Microbial Genomes from Parkinson’s Disease Fecal Samples. Sci. Rep. 2024, 14, 18906. [Google Scholar] [CrossRef]
- Jabbari Shiadeh, S.M.; Chan, W.K.; Rasmusson, S.; Hassan, N.; Joca, S.; Westberg, L.; Elfvin, A.; Mallard, C.; Ardalan, M. Bidirectional Crosstalk between the Gut Microbiota and Cellular Compartments of Brain: Implications for Neurodevelopmental and Neuropsychiatric Disorders. Transl. Psychiatry 2025, 15, 278. [Google Scholar] [CrossRef]
- Chai, Z.; Ouyang, Y.; Debebe, A.; Picker, M.; Lee, W.-J.; Fenton, S.; Becker-Dorison, A.; Augustin-Emmerichs, K.; Schwiertz, A.; Weber, S.N.; et al. Intestinal Biomarkers, Microbiota Composition, and Genetic Predisposition to Inflammatory Bowel Disease as Predictors of Parkinson’s Disease Manifestation. J. Park. Dis. 2025, 15, 766–779. [Google Scholar] [CrossRef]
- Chtioui, N.; Duval, C.; St-Pierre, D.H. The Impact of an Active Lifestyle on Markers of Intestinal Inflammation in Parkinson’s Disease: Preliminary Findings. Clin. Park. Relat. Disord. 2025, 12, 100301. [Google Scholar] [CrossRef]
- Fan, T.; Peng, J.; Liang, H.; Chen, W.; Wang, J.; Xu, R. Potential Common Pathogenesis of Several Neurodegenerative Diseases. Neural Regen. Res. 2026, 21, 972–988. [Google Scholar] [CrossRef]
- Terenzi, D.; Muth, A.-K.; Park, S.Q. Nutrition and Gut–Brain Pathways Impacting the Onset of Parkinson’s Disease. Nutrients 2022, 14, 2781. [Google Scholar] [CrossRef]
- Yan, Z.; Zhao, G. The Associations Among Gut Microbiota, Branched Chain Amino Acids, and Parkinson’s Disease: Mendelian Randomization Study. J. Park. Dis. 2024, 14, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Valle, J. Biofilm-Associated Proteins: From the Gut Biofilms to Neurodegeneration. Gut Microbes 2025, 17, 2461721. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, Q.; Su, R. Interplay of Human Gastrointestinal Microbiota Metabolites: Short-Chain Fatty Acids and Their Correlation with Parkinson’s Disease. Medicine (Baltimore) 2024, 103, e37960. [Google Scholar] [CrossRef] [PubMed]
- Buliman, A.; Calin, M.A.; Iordache, M.P. Targeting Anxiety with Light: Mechanistic and Clinical Insights into Photobiomodulation Therapy: A Mini Narrative Review. Balneo PRM Res. J. 2025, 16, 846–846. [Google Scholar] [CrossRef]
- Derkinderen, P.; Cossais, F.; Kulcsárová, K.; Škorvánek, M.; Sellier-Montaigne, L.; Coron, E.; Leclair-Visonneau, L.; Cerri, S.; Pellegrini, C.; Rolli-Derkinderen, M. How Leaky Is the Gut in Parkinson’s Disease? eBioMedicine 2025, 117, 105796. [Google Scholar] [CrossRef]
- Yue, M.; Chen, T.; Chen, W.; Wei, J.; Liao, B.; Zhang, J.; Li, F.; Hong, D.; Fang, X. The Engineered Probiotic Strain Lactococcus Lactis MG1363-pMG36e-GLP-1 Regulates Microglial Polarization and Gut Dysbiosis in a Transgenic Mouse Model of Parkinson’s Disease. Neural Regen. Res. 2026, 21, 1211–1221. [Google Scholar] [CrossRef]
- Cervantes-Arriaga, A.; Sarabia-Tapia, C.; Esquivel-Zapata, O.; López-Alamillo, S.; Reséndiz-Henriquez, E.; Corona, T.; Rodríguez-Violante, M. Pitfalls and Caveats in the Diagnostic Pathway of People with ParkinsonÓ?S Disease. Rev. Mex. Neurocienc. 2022, 23, 7436. [Google Scholar] [CrossRef]
- Faysal, Md.; Zehravi, M.; Sutradhar, B.; Al Amin, M.; Shanmugarajan, T.S.; Arjun, U.V.N.V.; Ethiraj, S.; Durairaj, A.; Dayalan, G.; Ahamad, S.K.; et al. The Microbiota-Gut-Brain Connection: A New Horizon in Neurological and Neuropsychiatric Disorders. CNS Neurosci. Ther. 2025, 31, e70593. [Google Scholar] [CrossRef]
- Ramos, H.; Araújo, A.M.; Ferreira, I.M.; Faria, M.A. The Neurotoxic Impact of Food Chemical Contaminants: A Growing Concern? Curr. Opin. Food Sci. 2026, 67, 101369. [Google Scholar] [CrossRef]
- Xie, Y.-A.; Kong, J.-D.; Li, S.; Wei, D.-F. What Is the Impact of Dopamine D2 Receptor in the Brain-Gut Axis? A Narrative Review of the Mechanism Based on Gut Microbiota in Modulating Emotion and Behavior. Alpha Psychiatry 2025, 26, 39226. [Google Scholar] [CrossRef]
- Feix, J.B.; Cheng, G.; Hardy, M.; Kalyanaraman, B. Microbial Metabolism of Levodopa as an Adjunct Therapeutic Target in Parkinson’s Disease. Antioxidants 2026, 15, 120. [Google Scholar] [CrossRef] [PubMed]
- Bi, M.; Liu, C.; Wang, Y.; Liu, S.-J. Therapeutic Prospect of New Probiotics in Neurodegenerative Diseases. Microorganisms 2023, 11, 1527. [Google Scholar] [CrossRef] [PubMed]
- Hey, G.; Nair, N.; Klann, E.; Gurrala, A.; Safarpour, D.; Mai, V.; Ramirez-Zamora, A.; Vedam-Mai, V. Therapies for Parkinson’s Disease and the Gut Microbiome: Evidence for Bidirectional Connection. Front. Aging Neurosci. 2023, 15, 1151850. [Google Scholar] [CrossRef]
- Patel, N.J.; Hajoori, M.; Desai, P. Evaluating Probiotic Properties of Gut Microflora for Gut Modulation as an Adjuvant Therapy for Parkinson’s Disease. J. Future Foods 2025, 5, 304–316. [Google Scholar] [CrossRef]
- Takahashi, R.; Yamakado, H.; Uemura, N.; Taguchi, T.; Ueda, J. The Gut–Brain Axis Based on α-Synuclein Propagation—Clinical, Neuropathological, and Experimental Evidence. Int. J. Mol. Sci. 2025, 26, 3994. [Google Scholar] [CrossRef]
- Domínguez Rojo, N.; Blanco Benítez, M.; Cava, R.; Fuentes, J.M.; Canales Cortés, S.; González Polo, R.A. Convergence of Neuroinflammation, Microbiota, and Parkinson’s Disease: Therapeutic Insights and Prospects. Int. J. Mol. Sci. 2024, 25, 11629. [Google Scholar] [CrossRef]
- Zheng, Y.; Bonfili, L.; Wei, T.; Eleuteri, A.M. Understanding the Gut–Brain Axis and Its Therapeutic Implications for Neurodegenerative Disorders. Nutrients 2023, 15, 4631. [Google Scholar] [CrossRef]
- Iyengar, A.R.S.; Dunkley, P.R.; Dickson, P.W. Immunity in Parkinson’s Disease - the Role of Adaptive and Auto-Immune Responses and Gut-Microbiome Axis. J. Neuroimmunol. 2025, 409, 578755. [Google Scholar] [CrossRef]
- Kearns, R. Gut–Brain Axis and Neuroinflammation: The Role of Gut Permeability and the Kynurenine Pathway in Neurological Disorders. Cell. Mol. Neurobiol. 2024, 44, 64. [Google Scholar] [CrossRef]
- Liang, Y.; Zhao, Y.; Fasano, A.; Su, C.-W. Gut Permeability and Microbiota in Parkinson’s Disease: Mechanistic Insights and Experimental Therapeutic Strategies. Int. J. Mol. Sci. 2025, 26, 9593. [Google Scholar] [CrossRef]
- Padhi, P.; Abdalla, A.; Schneider, B.; Backes, N.; Otto, A.A.; Scheibe, I.J.; Thomas, J.P.; Khadse, G.; Samidurai, M.; Jochmans, A.K.; et al. Bioengineered Gut Bacterium Synthesizing Levodopa Alleviates Motor Deficits in Models of Parkinson’s Disease. Cell Host Microbe 2025, 33, 1837–1854.e13. [Google Scholar] [CrossRef]
- Swain, D.; Panigrahi, L.L.; Pradhan, S.K.; Arakha, M.; Jadhao, K.R.; Rout, G.R. Elucidation of Dual Antimicrobial and Anti-Parkinsonian Activities through an In-Silico Approach of Ipomoea Mauritiana Jacq. in the Context of the Gut–Brain Axis. ACS Omega 2026, 11, 4021–4036. [Google Scholar] [CrossRef]
- Parrella, E.; Gennari, M.M.; Abate, G.; Pucci, M.; Schioppa, T.; Bosisio, D.; Tirelli, E.; Benarese, M.; Vegezzi, G.; Silletti, M.G.; et al. Early Alpha-Synuclein Accumulation, Oxidative Stress and Inflammation in the Proximal Colon of c-Rel-/- Mouse Model of Parkinson’s Disease. Neurobiol. Dis. 2025, 217, 107182. [Google Scholar] [CrossRef]
- Rust, C.; Tonge, D.; Van Den Heuvel, L.L.; Asmal, L.; Carr, J.; Pretorius, E.; Seedat, S.; Hemmings, S.M.J. Investigating the Blood Microbiome in Parkinson’s Disease, Schizophrenia, and Posttraumatic Stress Disorder. Brain Behav. 2025, 15, e70629. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; Borrego, J.J. Influence of Human Gut Microbiome on the Healthy and the Neurodegenerative Aging. Exp. Gerontol. 2024, 194, 112497. [Google Scholar] [CrossRef] [PubMed]
- Leta, V.; Zinzalias, P.; Batzu, L.; Mandal, G.; Staunton, J.; Jernstedt, F.; Rosqvist, K.; Timpka, J.; Van Vliet, T.; Trivedi, D.; et al. Effects of a Four-Strain Probiotic on Gut Microbiota, Inflammation, and Symptoms in Parkinson’s Disease: A Randomized Clinical Trial. Mov. Disord. 2025, 40, 2710–2721. [Google Scholar] [CrossRef] [PubMed]
- Panaitescu, P.-Ștefan; Răzniceanu, V.; Mocrei-Rebrean, Ștefania-M.; Neculicioiu, V.S.; Dragoș, H.-M.; Costache, C.; Filip, G.A. The Effect of Gut Microbiota-Targeted Interventions on Neuroinflammation and Motor Function in Parkinson’s Disease Animal Models—A Systematic Review. Curr. Issues Mol. Biol. 2024, 46, 3946–3974. [Google Scholar] [CrossRef]
- Han, M.N.; Di Natale, M.R.; Lei, E.; Furness, J.B.; Finkelstein, D.I.; Hao, M.M.; Diwakarla, S.; McQuade, R.M. Assessment of Gastrointestinal Function and Enteric Nervous System Changes over Time in the A53T Mouse Model of Parkinson’s Disease. Acta Neuropathol. Commun. 2025, 13, 58. [Google Scholar] [CrossRef]
- Morais, L.H.; Stiles, L.; Freeman, M.; Oguienko, A.D.; Hoang, J.D.; Ji, J.; Jones, J.; Quan, B.; Devine, J.; Bois, J.S.; et al. The Gut Microbiome Promotes Mitochondrial Respiration in the Brain of a Parkinson’s Disease Mouse Model. Npj Park. Dis. 2025, 11, 301. [Google Scholar] [CrossRef] [PubMed]
- Claudino Dos Santos, J.C.; Oliveira, L.F.; Noleto, F.M.; Gusmão, C.T.P.; Brito, G.A.D.C.; Viana, G.S.D.B. Gut-Microbiome-Brain Axis: The Crosstalk between the Vagus Nerve, Alpha-Synuclein and the Brain in Parkinson’s Disease. Neural Regen. Res. 2023, 18, 2611–2614. [Google Scholar] [CrossRef]
- Li, T.; Wu, J.; Zhou, S.; Li, M.; Zhao, L.; Wang, A.; Song, Y.; Huang, W.; Tan, L.; Qiao, C.; et al. Neuroprotective Effects of Time-Restricted Feeding Combined With Different Protein Sources in MPTP-Induced Parkinson’s Disease Mice Model and Its Modulatory Impact on Gut Microbiota Metabolism. Adv. Sci. 2026, e16502. [Google Scholar] [CrossRef]
- Carrossa, G.; Misenti, V.; Faggin, S.; Giron, M.C.; Antonini, A. The Small Intestinal Microbiota and the Gut–Brain Axis in Parkinson’s Disease: A Narrative Review. Biomedicines 2025, 13, 1769. [Google Scholar] [CrossRef] [PubMed]
- Hein, Z.M.; Arbain, M.F.F.; Kumar, S.; Mehat, M.Z.; Hamid, H.A.; Che Ramli, M.D.; Che Mohd Nassir, C.M.N. Intermittent Fasting as a Neuroprotective Strategy: Gut–Brain Axis Modulation and Metabolic Reprogramming in Neurodegenerative Disorders. Nutrients 2025, 17, 2266. [Google Scholar] [CrossRef]
- Dubey, S.; Ghosh, R.; Dubey, M.J.; Sengupta, S.; Das, S. From Allegory to Conceptualization, Hypothesis and Finally Evidences: Alzheimer’s Dementia, Parkinson’s Disease “Gut–Brain Axis” and Their Preclinical Phenotype. Egypt. J. Neurol. Psychiatry Neurosurg. 2024, 60, 89. [Google Scholar] [CrossRef]
- Mahbub, N.U.; Islam, M.M.; Hong, S.-T.; Chung, H.-J. Dysbiosis of the Gut Microbiota and Its Effect on α-Synuclein and Prion Protein Misfolding: Consequences for Neurodegeneration. Front. Cell. Infect. Microbiol. 2024, 14, 1348279. [Google Scholar] [CrossRef]
- Przewodowska, D.; Alster, P.; Madetko-Alster, N. Role of the Intestinal Microbiota in the Molecular Pathogenesis of Atypical Parkinsonian Syndromes. Int. J. Mol. Sci. 2025, 26, 3928. [Google Scholar] [CrossRef] [PubMed]
- Kroker Kimber, I.; Tremblay, M.-È. Food for Thought: Probiotic Modulation of Microglial Activity in Parkinson’s Disease. Front. Mol. Neurosci. 2025, 18, 1690507. [Google Scholar] [CrossRef]
- Pokora, B.; Pokora, K.; Binienda, A.; Fichna, J. The Ketogenic Diet in Parkinson’s Disease: A Potential Therapeutic Strategy. Pharmacol. Rep. 2025, 77, 1491–1513. [Google Scholar] [CrossRef] [PubMed]
- Theis, B.F.; Park, J.S.; Kim, J.S.A.; Zeydabadinejad, S.; Vijay-Kumar, M.; Yeoh, B.S.; Saha, P. Gut Feelings: How Microbes, Diet, and Host Immunity Shape Disease. Biomedicines 2025, 13, 1357. [Google Scholar] [CrossRef]
- Kwon, D.; Zhang, K.; Paul, K.C.; Folle, A.D.; Del Rosario, I.; Jacobs, J.P.; Keener, A.M.; Bronstein, J.M.; Ritz, B. Diet and the Gut Microbiome in Patients with Parkinson’s Disease. Npj Park. Dis. 2024, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, K.; Su, N.; Yuan, C.; Zhang, N.; Hu, X.; Fu, Y.; Zhao, F. Microbiota–Gut–Brain Axis in Health and Neurological Disease: Interactions between Gut Microbiota and the Nervous System. J. Cell. Mol. Med. 2024, 28, e70099. [Google Scholar] [CrossRef]
- Yuan, X.-Y.; Chen, Y.-S.; Liu, Z. Relationship among Parkinson’s Disease, Constipation, Microbes, and Microbiological Therapy. World J. Gastroenterol. 2024, 30, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Jo, S.; Lee, J.; Choi, M.; Kim, K.; Lee, S.; Kim, H.S.; Bae, J.-W.; Chung, S.J. Distinct Gut Microbiome Characteristics and Dynamics in Patients with Parkinson’s Disease Based on the Presence of Premotor Rapid-Eye Movement Sleep Behavior Disorders. Microbiome 2025, 13, 108. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Li, Y.; Yan, J.; Chang, R.; Xu, M.; Zhou, G.; Meng, J.; Liu, D.; Mao, Z.; Yang, Y. Gut Microbiota from Patients with Parkinson’s Disease Causes Motor Deficits in Honeybees. Front. Microbiol. 2024, 15, 1418857. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Zhao, Y.; Cheng, Y.; Huang, C.; Zhang, F. Helicobacter Pylori Infection and Parkinson’s Disease: Etiology, Pathogenesis and Levodopa Bioavailability. Immun. Ageing 2024, 21, 1. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, R.; Wen, G.; Xie, L.; Chen, T.; Liu, W. The Role of Gut Microbiota Tyrosine Decarboxylases in Levodopa Pharmacokinetics: Insights from a Levodopa Challenge Test. BMC Neurol. 2025, 25, 460. [Google Scholar] [CrossRef]
- Papić, E.; Rački, V.; Hero, M.; Zimani, A.N.; Čižek Sajko, M.; Rožmarić, G.; Starčević Čizmarević, N.; Ostojić, S.; Kapović, M.; Hauser, G.; et al. Microbial Diversity in Drug-Naïve Parkinson’s Disease Patients. PLOS One 2025, 20, e0328761. [Google Scholar] [CrossRef]
- Chambilo, B.; Dhiver, T.; Sahu, K.; Parashar, R. Gut Microbiota Modulation with Probiotics: Potential Therapeutic Avenues for Alzheimer’s and Parkinson’s Diseases. Neurol. Sci. Neurophysiol. 2025, 42, 75–83. [Google Scholar] [CrossRef]
- Du, Y.; Wang, L.; Cui, Y.; Xu, X.; Zhang, M.; Li, Y.; Gao, T.; Gao, D.; Sheng, Z.; Wang, S.; et al. Effect of Probiotics Supplementation on REM Sleep Behavior Disorder and Motor Symptoms in Parkinson’s Disease: A Pilot Study. CNS Neurosci. Ther. 2025, 31, e70541. [Google Scholar] [CrossRef]
- Papić, E.; Rački, V.; Hero, M.; Tomić, Z.; Starčević-Čižmarević, N.; Kovanda, A.; Kapović, M.; Hauser, G.; Peterlin, B.; Vuletić, V. The Effects of Microbiota Abundance on Symptom Severity in Parkinson’s Disease: A Systematic Review. Front. Aging Neurosci. 2022, 14, 1020172. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Pinto, M.F.; Candeias, E.; Melo-Marques, I.; Esteves, A.R.; Maranha, A.; Magalhães, J.D.; Carneiro, D.R.; Sant’Anna, M.; Pereira-Santos, A.R.; Abreu, A.E.; et al. Gut-First Parkinson’s Disease Is Encoded by Gut Dysbiome. Mol. Neurodegener. 2024, 19, 78. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Signalling Cascade Linking the Gut and Brain in PD. Created in BioRender. Iordache, M. (2026) https://BioRender.com/vs3gnrk. Abbreviations: BBB, brain blood barrier; IL-1β, interleukin 1β; IL-6, interleukin 6; SCFA, short chain fatty acids; TNF-α, tumor necrosis factor α.
Figure 1.
Signalling Cascade Linking the Gut and Brain in PD. Created in BioRender. Iordache, M. (2026) https://BioRender.com/vs3gnrk. Abbreviations: BBB, brain blood barrier; IL-1β, interleukin 1β; IL-6, interleukin 6; SCFA, short chain fatty acids; TNF-α, tumor necrosis factor α.
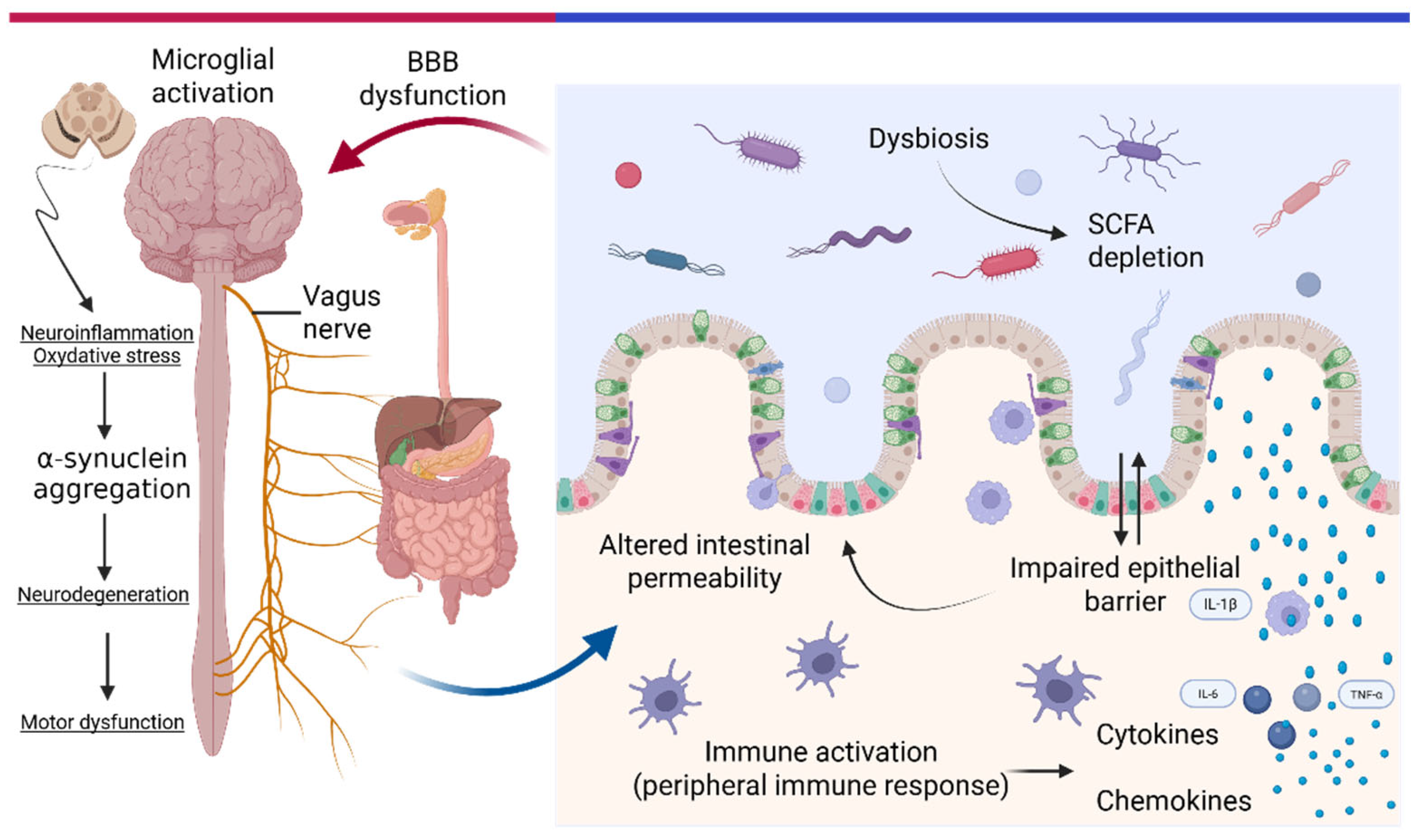

Table 1.
Reproducible Taxonomic Alterations in PD Across Meta-Analyses.
| Taxon |
Direction in PD |
Functional Role |
Mechanistic Implication |
||||
| Faecalibacterium prausnitzii | ↓ Decreased | Butyrate production | Reduced epithelial barrier integrity | [20] | |||
| Roseburia intestinalis | ↓ Decreased | SCFA production | Impaired anti-inflammatory signaling | [47,48] | |||
| Coprococcus spp. | ↓ Decreased | SCFA production; BCAA modulation | Altered metabolic homeostasis | [44,49,50] | |||
| Akkermansia muciniphila | ↑ Increased | Mucin degradation | Thinning of mucus layer | [42,51] | |||
| Bilophila spp. | ↑ Increased | Sulfur metabolism | Pro-inflammatory environment | [46,52] | |||
| Klebsiella variicola | ↑ Increased | LPS production | Immune activation | [13,53] | |||
| GABA-consuming species (Evtepia gabavorous) | ↑ Increased | GABA metabolism | Altered inhibitory neurotransmission | [16,54,55] |
Abbreviations: BCAA, branched-chain amino acid; GABA, gamma-aminobutyric acid; LPS, lipopolysaccharide; PD, Parkinson’s disease; SCFAs, short-chain fatty acids.
Table 2.
Functional Metabolic Pathways Altered in PD.
| Pathway | Direction in PD | Associated Consequence | |
| Riboflavin biosynthesis | ↓ Decreased | Reduced antioxidant capacity | [43,71,72] |
| Biotin biosynthesis | ↓ Decreased | Impaired fatty acid metabolism | [27,73] |
| CAZymes | ↓ Decreased | Reduced complex carbohydrate fermentation |
[74,75] |
| Secondary bile acid biosynthesis | ↓ Decreased | Altered gut–liver axis signaling |
[76,77] |
| Xenobiotic metabolism (solvent/pesticide) | ↑ Increased | Potential modulation of toxin exposure | [78,79,80,81] |
| Flagellar assembly genes | ↓ Decreased | Altered microbial motility | [53,68,82] |
Abbreviations: CAZymes, carbohydrate-active enzymes.
Table 3.
Microbial Metabolites Implicated in PD Pathogenesis.
| Metabolite | Direction in PD | Potential Impact | |
| Butyrate | ↓ Decreased | Reduced barrier integrity; microglial regulation |
[42] |
| Propionate | ↓ Decreased | Altered immune signaling | [105,106] |
| Polyamines | ↓ Decreased | Mucosal stability impairment | [53,107] |
| Isoleucine (BCAA) | ↓ Lower levels associated with higher PD risk | Dopaminergic metabolic modulation |
[108] |
| GABA | Dysregulated | Basal ganglia circuit imbalance | [9,47,59] |
| Lipopolysaccharide (LPS) | ↑ Increased systemic levels | TLR activation and neuroinflammation | [80,109] |
Abbreviations: BCAA, branched-chain amino acid; GABA, gamma-aminobutyric acid; LPS, lipopolysaccharide; PD, Parkinson’s disease; TLR, Toll-like receptor.
Table 4.
Clinical and Translational Microbiome-Targeted Interventions in PD.
| Intervention | Mechanism | Reported Effects | Limitations | |
| High-fiber / Mediterranean diet | Increases SCFA-producing taxa | Improved microbial diversity | Long-term adherence variability | [138] |
| Prebiotics | Stimulate SCFA production | Improved constipation | Limited large RCTs | [20,26] |
| Probiotics | Modulate inflammatory taxa | Improved GI symptoms | Strain heterogeneity | [50,103,139] |
| FMT | Restores microbial diversity | Motor and non-motor symptom improvement (preliminary) | Small sample sizes |
[140,141] |
| Bacterial decarboxylase inhibition | Reduces levodopa metabolism | Improved drug bioavailability (experimental) | Early translational stage | [102,121,142] |
Abbreviations: FMT, fecal microbiota transplantation; PD, Parkinson’s disease; RCT, randomized controlled trial; SCFAs, short-chain fatty acids.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



