Submitted:
16 February 2026
Posted:
18 February 2026
You are already at the latest version
Abstract
Diabetic kidney disease (DKD) is one of the most serious complications of diabetes and the leading cause of end-stage renal disease worldwide. Recently, renin-angiotensin system inhibitors, non-steroidal mineralocorticoid receptor antagonists, sodium-glucose cotransporter 2 inhibitors, and glucagon-like peptide-1 receptor agonists have been proposed as the four pillars for treating DKD. To understand the molecular mechanisms by which these drugs improve DKD, we described the histological and molecular changes due to diabetes. Based on our understanding of the molecular changes in DKD, we present evidence on the efficacy of these drugs in improving DKD and discuss why such drugs improve the prognosis of DKD. In addition to diabetes and hypertension, insulin resistance, dyslipidemia and hyperuricemia are risk factors for DKD. Metformin, fibrates, and febuxostat have been reported to improve DKD, however, caution is required when administering these drugs to patients with renal impairment due to concerns about the onset of lactic acidosis, rhabdomyolysis, and deterioration of renal function, respectively. Imeglimin, pemafibrate, and dotinurad have similar chemical structures or effects to metformin, fibrates, and febuxostat, respectively, but are safer in patients with renal impairment. Furthermore, they have specific mechanisms to improve DKD and may offer new options for its treatment.
Keywords:
diabetic kidney disease
; dotinurad
; glucagon-like peptide-1 receptor agonists
; imeglimin
; nonsteroidal mineralocorticoid receptor antagonists
; pemafibrate
; sodium-glucose cotransporter-2 inhibitors
1. Introduction
Diabetic kidney disease (DKD) is defined by albuminuria and progressive reduction in estimated glomerular filtration rate (eGFR) in the setting of a long duration of diabetes and is typically associated with retinopathy [1]. The diagnosis of DKD is most conclusively made by findings of mesangial expansion and nodular glomerulosclerosis on kidney biopsy, though kidney biopsy is rarely necessary. DKD is one of the most serious complications of diabetes and the leading cause of end-stage renal disease (ESRD) worldwide [2]. DKD is widespread, causing serious health problems and imposing a significant economic burden on human societies worldwide [3].
Risk factors of DKD include hyperglycemia, hypertension, dyslipidemia, and obesity/insulin resistance [4]. Hyperglycemia is considered one of the most prominent and independent risk factors for DKD, as it worsens renal function by altering the antioxidant system, leading to an increased formation of advanced glycation end products (AGEs) [5]. Hyperglycemia also activates the polyol pathway, enhancing the development of DKD [5]. Randomized controlled trials (RCTs), such as the ACCORD trial and VADT trial, have demonstrated the beneficial effects of intensive glucose control on the delayed onset and prevention of albuminuria progression in patients with type 2 diabetes [6,7].
Hypertension is also a pivotal risk factor for DKD. A meta-analysis including 27 studies showed an increased risk of DKD with hypertension by 67% compared to without hypertension [8]. In the prospective cohort study of 158,365 Chinese men and women, compared with those with normal blood pressure, prehypertension, and stage 1 and stage 2 hypertension increased the development of ESRD by 30%, 47% and 106%, respectively [9]. Systolic blood pressure was a stronger predictor of ESRD than diastolic blood pressure or pulse pressure.
Dyslipidemia plays an important role in the development and progression of DKD.
A recent meta-analysis comprising 165,230 studies found that low high-density lipoprotein cholesterol (HDL-C) levels (< 40 mg/dL) were associated with a 70% increase in DKD incidence [10]. Another recent meta-analysis demonstrated that low-density lipoprotein-cholesterol (LDL-C) was significantly higher in DKD patients who developed ESRD [11].
Obesity/insulin resistance is a critical risk factor for type 2 diabetes, hypertension, and dyslipidemia; therefore, obesity is closely related to the development of DKD. In the meta-analysis, abdominal obesity was significantly associated with an increased odds of DKD [12]. A recent meta-analysis showed that high body mass index (BMI) was a risk factor for adverse kidney events such as the onset of DKD, the doubling of serum creatinine, ESRD, or death in patients [13].
In diabetes, the renin-angiotensin system (RAS) is activated, especially in the kidneys and heart, driven by hyperglycemia and insulin resistance, leading to increased angiotensin II, which causes inflammation, oxidative stress, fibrosis, and further insulin resistance, thereby driving diabetic complications such as nephropathy and cardiovascular disease [14]. RAS is also implicated in the pathogenesis of primary hypertension [15]. Obesity activates the RAS through multiple pathways, including increased sympathetic nervous system activity, adipose tissue inflammation, insulin resistance, and direct effects of adipokines such as leptin, leading to higher blood pressure and sodium retention [16]. Dyslipidemia and RAS activation have a harmful "crosstalk": oxidized LDL increases RAS components, while activated RAS promotes further LDL oxidation, inflammation, and vascular damage, accelerating atherosclerosis and metabolic syndrome [17]. Activated RAS is significantly associated with the development of DKD, which may explain why RAS inhibitors (RASis) and non-steroidal mineralocorticoid receptor antagonists (nsMRAs) have improved renal outcomes in DKD.
Recently, RASis, nsMRAs, sodium-glucose cotransporter 2 inhibitors (SGLT2is), and glucagon-like peptide-1 receptor agonists (GLP-1RAs) have been proposed as the four pillars for treating DKD [18]. These drugs have demonstrated a significant prevention of DKD progression via different mechanisms in clinical trials. To understand how and where these drugs act in the kidney, we first consider how diabetes and related metabolic disorders alter kidney histology and hemodynamics. Second, we consider how these drugs act on kidneys that have been pathologically altered by diabetes and related metabolic disorders.
We will also introduce dotinurad, imeglimin, and pemafibrate, which are all developed in Japan and potentially effective for treating DKD.
2. The Histological and Molecular Characteristics of DKD
The histological and molecular characteristics of DKD are illustrated in Figure 1. Histological findings of DKD include proliferation of mesangial cells that support the glomerular structure, thickening of the glomerular basement membrane (GBM) which acts as a selective filter for the blood, preventing the passage of proteins and cells while allowing the passage of water and waste products, podocyte damage, glomerular endothelial dysfunction and arteriolosclerosis, interstitial fibrosis, and proximal and distal tubular damage [19].
Glomerular hyperfiltration contributes to the development of DKD. Hyperglycemia induces the dilatation of afferent arterioles through the release of vasoactive mediators, including insulin-like growth factor 1 (IGF-1), glucagon, nitric oxide (NO), vascular endothelial growth factor (VEGF), and prostaglandins [20]. Due to a high filtered load of glucose, reabsorption of glucose and sodium is increased because of the upregulation of SGLT2 in the proximal tubules. Reduced delivery of sodium to the macula densa of distal tubules causes dilatation of the afferent arteriole because of enhanced tubuloglomerular feedback (TGF) [21]. At the same time, constriction of the efferent arteriole occurs due to a high local level of angiotensin II, which causes glomerular hypertension [22].
Erythropoietin levels increase after initiation of SGLT2is treatment in diabetic patients. In patients with diabetes, the proximal tubules are overloaded with excessive glucose reabsorption, and the increased oxygen demand leads to tubulointerstitial hypoxia [23]. Consequently, erythropoietin production is impaired because of the dysfunction of fibroblasts. SGLT2is reduce the workload of the proximal tubules and improve tubulointerstitial hypoxia, allowing fibroblasts to resume normal erythropoietin production.
A study investigating the effect of arteriosclerosis on new-onset renal damage in a Chinese community population with diabetes demonstrated that atherosclerosis is a risk factor for new-onset renal damage, particularly new-onset proteinuria, in patients with diabetes [24]. A cross-sectional study revealed that vascular inflammation, atherosclerosis, and altered lipid metabolism were associated with the development of DKD [25].
The causes of mesangial cell proliferation are still largely unknown; however, hyperglycemia, hypertension, and angiotensin II are considered the primary factors contributing to mesangial cell proliferation [26]. Podocytes play a crucial role in maintaining the integrity of the glomerular filtration barrier by synthesizing components of the filtration barrier, forming slit membranes between their foot processes, and interacting with endothelial cells [27]. Hyperglycemia induces oxidative stress, inflammation, and mitochondrial damage, thereby leading to podocyte injury [28]. Hypertension, dyslipidemia, and vascular endothelial cell damage also contribute to podocyte injury and subsequent loss of glomerular filtration barrier integrity [29]. Podocytes are rich in mitochondria and heavily dependent on them for energy to maintain normal functions. Emerging evidence suggests that mitochondrial dysfunction is a key driver of podocyte injury in DKD [24]. Impairment of mitochondrial function results in an energy crisis, oxidative stress, inflammation, and cell death [29].
Renal interstitial fibrosis, caused by hyperglycemia and inflammation, is characterized by the accumulation of extracellular matrix (ECM) and by the transdifferentiation of renal tubular epithelial cells and fibroblasts into myofibroblasts, ultimately leading to excessive ECM production. Transforming growth factor beta (TGF-β) promotes this cellular transformation and ECM deposition, leading to renal fibrosis [30]. Hyperglycemia, dyslipidemia, oxidative stress, hypoxia, activation of RAS, endoplasmic reticulum (ER) stress, inflammation, epithelial–mesenchymal transition, and programmed cell death have been shown to induce renal tubular injury [31].
In the kidney with DKD, the infiltrating macrophages were mainly M1 macrophages [32]. In DKD, crosstalk between the Notch pathway and nuclear factor-κB (NF-κB) signaling in macrophages contributes to macrophage polarization. Hyperpolarized macrophages secrete great amounts of inflammatory cytokines, exacerbating the inflammatory response, ECM secretion, fibrosis, and necroptosis in glomerular mesangial cells and renal tubular epithelial cells [32].
Diabetes was associated with a 3.5-fold increase in collecting duct renin production via the angiotensin II type 1 receptor (AT1R) [33]. When DKD occurs, it stimulates the RAS, one of the most significant indicators of developing renal impairment. Intrarenal RAS elements are upregulated in patients with DKD, whereas systemic RAS elements are downregulated. The intrarenal RAS plays a key role in the development of DKD [34]. Increased urinary renin could contribute to RAS activation, a feature of DKD. Renin can activate the RAS locally within the kidney, which is relevant to sodium retention, hypertension, glomerular hemodynamics, inflammation, and the progression of DKD. Elevated urinary renin levels were found in patients with DKD [35]. The AT1R in the proximal tubule plays a crucial role in regulating blood pressure and fluid balance by controlling sodium (Na+) reabsorption, primarily through the Na+/H+ exchanger 3 (NHE3). Angiotensin II activates this receptor to increase Na+ uptake [36].
In diabetes, AT1Rs in the glomerulus and renal tubules are overactivated by hyperglycemia, inducing DKD by increasing oxidative stress, inflammation, and ECM deposition, which leads to poor filtration and protein leakage [37]. Angiotensin II binds to AT1Rs on mesangial cells, podocytes, and tubules. AT1R activation induces mesangial cell proliferation and stimulates them to produce excess ECM, impairing filtration. Hyperglycemia and angiotensin II lead to the overproduction of reactive oxygen species (ROS) and inflammatory signals, ultimately damaging podocytes [37].
In diabetes, angiotensin II binding to AT1R on endothelial cells promotes vascular damage by increasing ROS, reducing nitric oxide (NO) production, inducing inflammation, and causing cellular senescence and endothelial-to-mesenchymal transition [38]. AT1Rs are found on the vascular smooth muscle cells of both the afferent and efferent arterioles. Angiotensin II, acting primarily through AT1R, constricts both the afferent and efferent renal arterioles, a crucial mechanism for regulating GFR and systemic blood pressure [39]. Glomerular pressure is maintained by the balance in tone between the afferent and efferent arterioles. Glomerular hyperfiltration is an early manifestation of DKD and most likely results from dysregulation of afferent and efferent arteriolar tone [40]. Angiotensin II can cause relative vasoconstriction of the efferent arteriole, increase glomerular pressure, and lead to hyperfiltration [41].
MR in the kidney, primarily activated by aldosterone, regulates salt/water balance in the distal tubules, which is crucial for maintaining blood pressure. MR activation contributes to the propagation of kidney injury, inflammation, and fibrosis, as well as the progression of chronic kidney disease (CKD) [42]. MR can be activated by nonligand activation with the regulatory protein Rac family small guanosine triphosphatase 1 (Rac1), oxidative stress, elevated glucose, or high salt levels [43]. Rac1 mediates MR activation, and constitutively active Rac1 has been shown to induce nuclear translocation and activation of MR in podocytes [44]. Mice with elevated Rac1 activity developed podocyte injury and severe albuminuria, which were prevented by MR antagonism [44,45]. MR overactivation in podocytes may induce podocyte injury.
MR is expressed in mesangial cells, where its activation by aldosterone promotes inflammation, oxidative stress, fibrosis, and excessive cell proliferation through pathways such as the extracellular signal-regulated kinase pathway, contributing to kidney injury in DKD [46].
Autophagy is an essential process for podocyte maintenance and is disrupted in DKD [47]. Spironolactone improved DKD by promoting podocyte autophagy [48]. Aldosterone mediates podocyte injury through the NLRP3 inflammasome, a member of the NOD-like receptor family, and pyrin domain-containing 3 (NLRP3) [49]. Furthermore, aldosterone induces podocyte injury by reducing podocyte-associated molecules and activating the Wnt/β-catenin signaling pathway [50].
Endothelial dysfunction is reported in patients with CKD, which may be mediated by the endothelial MR [51]. Aldosterone increases the expression of cell adhesion molecules and monocyte chemoattractant protein-1 (MCP-1) in endothelial cells [52]. Aldosterone also promotes oxidative injury [53,54] and reduces endothelial nitric oxide synthase (eNOS) activity [55].
Aldosterone activates MR in the distal tubule and collecting duct to increase sodium reabsorption via the epithelial sodium channel (ENaC), thereby increasing blood pressure [56]. It has been shown that ENaC activation increases salt sensitivity in patients with diabetes [57,58]. Increased sodium retention enhances intraglomerular pressure that could lead to podocyte injury, mesangial expansion, and kidney fibrosis, histological characteristics of DKD. Thus, regulating ENaC to stimulate sodium excretion would be an important therapeutic target for preventing the progression of DKD.
In myeloid cells, aldosterone stimulates the expression of tumor necrosis factor-α (TNF-α), interleukin-12, and MCP-1 [59]. Infiltration of M1 proinflammatory macrophages promotes maladaptive repair and perpetuation of injury, whereas an early switch from M1 to M2 macrophages promotes effective repair [60]. Mice lacking MR in the myeloid lineage exhibit increased expression of M2 polarization markers in infiltrating macrophages [61].
Increased MR activation may lead to fibroblast proliferation and the production of profibrotic molecules, inducing renal fibrosis [62]. Aldosterone stimulates ECM synthesis [63], but platelet-derived growth factor-induced ECM production has been shown to be reduced in fibroblasts cultured with spironolactone [64], indicating a significant contribution of MR activation to renal fibrosis.
The endothelial cell MR is a central regulator of vascular dysfunction [65]. The endothelial cell MR disrupts vasomotor control via ENaC activation, eNOS dysregulation, and glycocalyx injury, amplifies inflammation and oxidative stress and mitochondrial ROS, and drives fibrosis and remodeling via TGF-β [65]. The endothelial cell MR contributes to diabetes- and obesity-related vascular disease.
3. RASis for DKD
RASis such as angiotensin-converting enzyme inhibitors (ACEis) and AT1R blockers (ARBs) reduce systemic blood pressure by vasodilation. RASis-induced renal vasodilation results in an increase in renal blood flow, leading to improvement of renal ischemia and hypoxia. ARBs reduce urinary albumin by lowering intraglomerular pressure, thereby protecting the glomerular endothelium and podocytes [66]. In addition to blocking angiotensin II-induced renal tissue injury, ARBs can decrease intrarenal angiotensin II levels by reducing proximal tubular angiotensinogen and renin production in the collecting duct, thereby reducing intrarenal RAS activation. As shown in Figure 1, ATR1 activation has been reported to have adverse effects across various kidney regions; therefore, ARBs are thought to improve DKD pathology in multiple ways.
In the RENAAL Study, ARB (losartan) reduced the incidence of doubling of serum creatinine concentration by 25% (P = 0.006) and ESRD by 28% (P = 0.002) compared with placebo in patients with DKD [67]. The level of proteinuria declined by 35% with losartan (P < 0.001 vs. placebo).
Another ARB, irbesartan reduced the risk of a doubling of the serum creatinine concentration by 33% (P = 0.003 vs. placebo) and by 37% (P < 0.001 vs. amlodipine) in patients with DKD [68]. Treatment with irbesartan reduced the relative risk of ESRD by 23% compared with placebo and amlodipine (P = 0.07 vs. placebo and amlodipine). Serum creatinine increased more slowly in the irbesartan group than in the placebo and amlodipine group.
4. nsMRA for DKD
Although a meta-analysis showed a 31% reduction in urinary protein or albumin excretion after treatment with a steroidal MRA in patients with CKD, data on hard clinical outcomes are lacking [69]. Finerenone, a nonsteroidal, selective MRA, had more potent anti-fibrotic effects than steroidal MRAs in preclinical models [70]. Finerenone has been shown to reduce the urinary albumin-to-creatinine ratio (UACR) in patients with CKD treated with an RASis [71].
Finerenone reduced the risk of the primary composite outcome, including renal failure, a sustained decrease of at least 40% in eGFR from baseline, or death from renal causes, in patients with CKD and type 2 diabetes treated with RASis by 18% in the FIDELIO-DKD study [72]. In this study, the early reduction in albuminuria, the early separation of the Kaplan-Meier curves for the secondary outcome (death from cardiovascular causes, nonfatal myocardial infarction, nonfatal stroke, or hospitalization for heart failure), and the modest reduction in blood pressure suggested that some of the benefits of finerenone may be mediated by its natriuretic effect.
In the RCT to determine whether the direct renin inhibitor aliskiren would reduce cardiovascular and renal events in patients with type 2 diabetes and CKD treated with RASis, systolic and diastolic blood pressures were lower with aliskiren, and the mean reduction in UACR was greater in the aliskiren group [73]. Although aliskiren demonstrated significant hemodynamic effects in this trial, it did not reduce the risk of the secondary outcome, suggesting that the addition of aliskiren with RASis in patients with type 2 diabetes who are at high risk for cardiovascular and renal events was not supported by this study. Another RCT showed that combination therapy with an ACEi and an ARB was associated with an increased risk of adverse events among DKD patients [74]. These results, along with the FIDELIO-DKD study, highlight the significance of selective MR blockade beyond strong RAS inhibition for the secondary outcome.
Aldosterone breakthrough is a well-known phenomenon that occurs in patients with long-term use of RASis [75]. The blockade of MR is effective in managing patients with resistant hypertension, defined as uncontrollable blood pressure despite the concurrent use of three antihypertensive drugs [75]. Aldosterone breakthrough puts hypertensive patients at a higher risk of cardiovascular disease and worsens future outcomes.
As shown in Figure 1, MR activation has been reported to contribute to deleterious effects in various parts of the kidney through mechanisms distinct from ATR1 activation. Therefore, nsMRAs are thought to ameliorate DKD pathology through multiple mechanisms, not just RAS inhibition. The delayed separation of the Kaplan–Meier curves for the primary outcome and for persistent benefit over the trial duration provides evidence supporting the hypothesis that finerenone may slow CKD progression by influencing tissue remodeling.
5. SGLT2is for DKD
In the EMPA-REG OUTCOME trial, SGLT2i, empagliflozin, slowed the progression of kidney disease and reduced the rate of clinically relevant renal events compared with placebo when added to standard care in patients with type 2 diabetes [76]. In this study, during long-term administration, the eGFR remained stable in the empagliflozin groups and declined steadily in the placebo group. In the CANVAS Program, another SGLT2i, canagliflozin showed a possible benefit with respect to the progression of albuminuria (hazard ratio [HR], 0.73; 95% confidence interval [CI], 0.67 to 0.79) and the composite outcome of a sustained 40% reduction in eGFR, the need for renal-replacement therapy, or death from renal causes (HR, 0.60; 95% CI, 0.47 to 0.77) [77]. In patients with DKD, the risk of renal failure and cardiovascular events was lower in the canagliflozin group than in the placebo group at a median follow-up of 2.62 years, in the CREDENCE trial [78].
The DAPA-CKD trial showed that the risk of a composite endpoint, including a sustained decline in eGFR of at least 50%, ESRD, or death from renal or cardiovascular causes, was significantly lower with dapagliflozin than with placebo in patients with CKD, regardless of diabetes status [79]. In the EMPA-KIDNEY trial, empagliflozin therapy led to a lower risk of progression of kidney disease or death from cardiovascular causes than placebo, in a wide range of patients with CKD regardless of the presence or absence of diabetes [80].
The mechanisms for possible renal protective effects of SGLT2is were shown in Figure 2. The main mechanism of the renal protective effect of SGLT2is is the sodium-related physiological effects of SGLT2is, proposed by Heerspink H.J.L. et al. [21]. An increased SGLT2 mRNA expression increases the renal NaCl reabsorption in the proximal tubule, leading to a marked reduction in distal NaCl delivery to the macula densa [81]. The decline in macula densa NaCl delivery is sensed by the juxtaglomerular apparatus as a reduction in circulating plasma volume, a phenomenon called TGF, leading to maladaptive glomerular afferent arterial vasodilatation and increased intraglomerular pressure [82]. SGLT2is increase distal renal NaCl delivery, inducing an increased afferent tone, thereby reducing the intraglomerular pressure and glomerular hyperfiltration [83], which may be the renal protective effect.
SGLT2is reduce the overload of the proximal tubules and improve tubulointerstitial hypoxia, which leads to the recovery of erythropoietin production by fibroblasts [23]. Increased hematocrit during SGLT2i therapy indicates the recovery of tubulointerstitial function in diabetic kidneys [23]. In fact, serum erythropoietin increased from baseline in the dapagliflozin group through week 4 [84]. Elevated erythropoietin may contribute to the renal protective effect of SGLT2is [85]. The treatment with recombinant human erythropoietin showed renal protective effects beyond hematopoiesis in streptozotocin-induced diabetic rats [86]. Erythropoietin has been reported to protect podocytes from AGEs-induced damage [87]. Erythropoietin ameliorated podocyte injury in advanced diabetic nephropathy in db/db mice [88].
Hyperuricemia may be associated with a higher risk of DKD progression in individuals with type 2 diabetes [89]. Hyperuricemia results from increased purine production coupled with reduced renal or intestinal excretion of uric acid (UA) [90]. Renal excretion of UA is the major regulator of serum UA concentration [91,92]. In humans, the reabsorption of UA into the blood plays a crucial role in regulating serum UA levels. Renal UA reabsorption is mainly mediated by urate transporter 1 (URAT1) and glucose transporter 9 (GLUT9) expressed in the proximal tubule [93,94,95]. ATP-binding cassette, subfamily G, 2 (ABCG2) has been identified as a high-capacity UA exporter that mediates renal and/or intestinal UA excretion [96]. The meta-analysis using 59 RCTs, all SGLT2is significantly decreased serum UA levels compared with the placebo [97]. Another meta-analysis showed that SGLT2is significantly reduced the risk of gout in individuals with type 2 diabetes and heart failure [98]. SGLT2is increase urinary glucose and sodium by blocking SGLT2 in the proximal tubule, and then, glucosuria and natriuresis augment tubular flow, diluting luminal UA and downregulating URAT1, which reduces UA reabsorption [99,100,101,102]. SGLT2is increases urinary glucose, and increased urinary glucose may compete with urinary UA for apical GLUT9 in the proximal tubules, thereby reducing UA reabsorption [103]. SGLT2is have been reported to upregulate ABCG2, thus promoting urinary UA excretion, in diabetic mice [104]. The serum UA-lowering or increased urinary UA excretion effects of SGLT2is may contribute to its renal protective effect.
Exposure of mesangial cells to high glucose levels significantly increased SGLT2 expression [105]. Canagliflozin inhibited high-glucose-induced activation of the protein kinase C (PKC)-NAD(P)H oxidase pathway and increases in ROS production [105]. Inhibition of mesangial SGLT2 may reduce PKC activation and ROS production in DKD, contributing to the renoprotective effects of SGLT2 inhibitors.
Podocyte damage disrupts the normal structure of GBM, leading to proteinuria, the most common clinical sign of DKD [106]. Recently, SGLT2 has been reported to be expressed in podocytes, and SGLT2is have been shown to improve podocyte damage in kidney diseases [107,108]. Empagliflozin showed structural and functional benefits on podocytes of BTBR ob/ob mice [109]. These results indicate that SGLT2is’s protective effect against DKD may be mediated by its direct effects on podocytes.
Mitochondria-associated ER membranes (MAMs) are dynamic contact sites between ER and mitochondria that play an important role in the exchange of metabolic substances and messengers between the two organelles, as well as in modulating calcium and lipid homeostasis, apoptosis, inflammation, ER stress, mitochondrial dynamics, and bioenergetics [110]. MAMs dysfunction may be a crucial pathogenesis of podocyte damage in DKD [111,112]. SGLT2is have been previously reported to protect mitochondrial and ER functions [113,114]. Inhibition of SGLT2 attenuates MAMs imbalance in diabetic podocytes by activating the AMP-activated protein kinase (AMPK) pathway [115].
Endothelial dysfunction is associated with the development of DKD, and advanced diabetic glomerulopathy often exhibits thrombotic microangiopathy, including glomerular capillary microaneurysms and mesangiolysis, which are typical manifestations of endothelial dysfunction in the glomerulus [116]. Diabetic mice with severe endothelial dysfunction owing to eNOS deficiency develop progressive nephropathy, indicating a significant role of endothelial dysfunction in DKD. SGLT2is have been reported to improve vascular endothelial dysfunction, including improving eNOS [117].
6. GLP-1RAs for DKD
In the LEADER Trial, the renal outcome occurred in fewer participants in the liraglutide group than in the placebo group (HR, 0.78; 95% CI, 0.67 to 0.92; P=0.003). The new onset of persistent macroalbuminuria occurred in fewer participants in the liraglutide group than in the placebo group (HR, 0.74; 95% CI, 0.60 to 0.91; P=0.004) [118].
In the FLOW Trial, the risk of a primary-outcome event was 24% lower in the semaglutide group than in the placebo group (HR, 0.76; 95% CI, 0.66 to 0.88; P = 0.0003) [119]. The mean annual eGFR slope was less steep in the semaglutide group (P<0.001).
The mechanisms of possible renal protective effects of GLP-1RAs were shown in Figure 3. Podocyte apoptosis was detected in early DKD [18]. The phosphatidylinositol 3'-kinase-Akt (PI3K-Akt) signaling pathway plays an important role in podocyte apoptosis [120,121]. GLP-1RAs inhibit podocyte apoptosis by upregulating the PI3K/Akt pathway by promoting white fat browning [122]. GLP-1RAs reversed diabetes- and high-glucose-induced decreases in podocyte GLP-1R expression, suggesting that GLP-1RAs may directly affect podocytes via GLP-1R [122]. GLP-1RA may directly and indirectly act on podocytes.
Ferroptosis is a regulated mechanism of cellular demise distinguished by iron-dependent oxidative stress and lipid peroxidation [123]. Oxidative stress can damage tubular and vascular endothelial cells and induce the secretion of profibrotic factors, ultimately leading to fibroblast activation and fibroblast/macrophage recruitment [124]. Targeting ferroptosis could be a significant approach in the fight against fibrosis. GLP-1RAs attenuated renal fibrosis injury in db/db mice by inhibiting ferroptosis [125].
The natriuretic effect of GLP-1RAs has been proposed to underlie the GLP-1RAs–induced reduction in blood pressure [126]. GLP-1RAs increase natriuresis by reducing NHE3 activity in the proximal tubule [127]. Elevated salt sensitivity by ENaC overexpression may be a risk factor for DKD. GLP-1RAs could ameliorate salt sensitivity by decreasing ENaC expression and help prevent kidney injury in DKD [128]. The increased natriuresis induced by inhibition of NHE3 and ENaC by GLP-1RAs may increase Na+ delivery to macula densa, reducing TGF and resulting in alleviation of glomerular hypertension, which may also contribute to renoprotection by GLP-1RAs.
Lipotoxicity, the excessive accumulation of lipids in non-adipose tissues due to abnormal lipid metabolism, is associated with the progression of DKD by inducing ER stress, mitochondrial dysfunction, impaired autophagy, inflammation, and oxidative stress [129]. GLP-1RAs may attenuate lipotoxicity by improving insulin resistance in adipose tissue, enhancing fatty acid (FA) oxidation, reducing FA synthesis, and improving mitochondrial function [130]. Mesangial cells are susceptible to lipotoxicity, and lipotoxicity-induced apoptosis is involved in DKD [131]. Palmitate-mediated lipotoxicity increased apoptosis and decreased GLP-1R expression in a rat mesangial cell line [132]. Metformin attenuated lipotoxicity-induced mesangial cell apoptosis and restored GLP-1R expression [132], suggesting that GLP-1R activation contributes to an improvement in mesangial cells. To our knowledge, no studies have examined the direct effects of GLP-1RAs on mesangial cells, so future research is awaited.
GLP-1RAs improve endothelial dysfunction induced by diabetes [133], helping to prevent the progression of DKD. Improvement in endothelial dysfunction decreases systemic production of inflammatory cytokines and ROS, which may also be reno-protective. GLP-1RAs have been associated with increased mitochondrial biogenesis and improved mitochondrial function [134], thereby reducing systemic ROS production, contributing to renoprotection.
7. Other Possible Drugs Beneficial for DKD
7.1. Dotinurad
Hyperuricemia may be associated with a higher risk of DKD progression [89]. DKD combined with hyperuricemia constitutes a vicious cycle, where diabetic microvascular complications affect the kidneys and interact with persistent hyperuricemia. Very recently, a meta-analysis evaluated febuxostat, an xanthine oxidase (XO) inhibitor, for treating DKD with hyperuricemia showed that febuxostat significantly reduced serum UA, UACR, and serum creatinine levels, and improved eGFR [135].
CKD patients are at increased risk of developing cardiovascular disease due to CKD-specific risk factors [136]. The accumulation of uremic toxins in the circulation and in tissues is associated with the progression of CKD and its co-morbidities, such as cardiovascular disease [136]. ABCG2 is a major transporter of the uremic toxin indoxyl sulfate (IS) [137]. ABCG2 regulates the pathophysiological excretion of IS and strongly influences CKD prognosis. Febuxostat has been reported to inhibit human ABCG2 at a clinical dose [138]. IS accumulates in the body in CKD. An ABCG2 inhibitor, such as febuxostat, causes a marked accumulation of renal IS in rats by suppressing its excretion [139]. Hypouricemic agents that do not affect ABCG2 are effective therapeutic options for the treatment of hyperuricemia in patients with CKD.
Figure 4.
The inhibitory effects of febuxostat and dotinurad on molecules associated with uric acid and uremic toxin indoxyl sulfate metabolism. Black arrows indicate the flow of substance, and blue arrows indicate inhibitory effects of febuxostat and dotinurad on molecules associated with uric acid and indoxyl sulfate metabolism. ATP-binding cassette, subfamily G, 2 (ABCG2); IS, indoxyl sulfate; UA, uric acid; URAT1, urate transporter 1; XO, xanthine oxidase.
Figure 4.
The inhibitory effects of febuxostat and dotinurad on molecules associated with uric acid and uremic toxin indoxyl sulfate metabolism. Black arrows indicate the flow of substance, and blue arrows indicate inhibitory effects of febuxostat and dotinurad on molecules associated with uric acid and indoxyl sulfate metabolism. ATP-binding cassette, subfamily G, 2 (ABCG2); IS, indoxyl sulfate; UA, uric acid; URAT1, urate transporter 1; XO, xanthine oxidase.
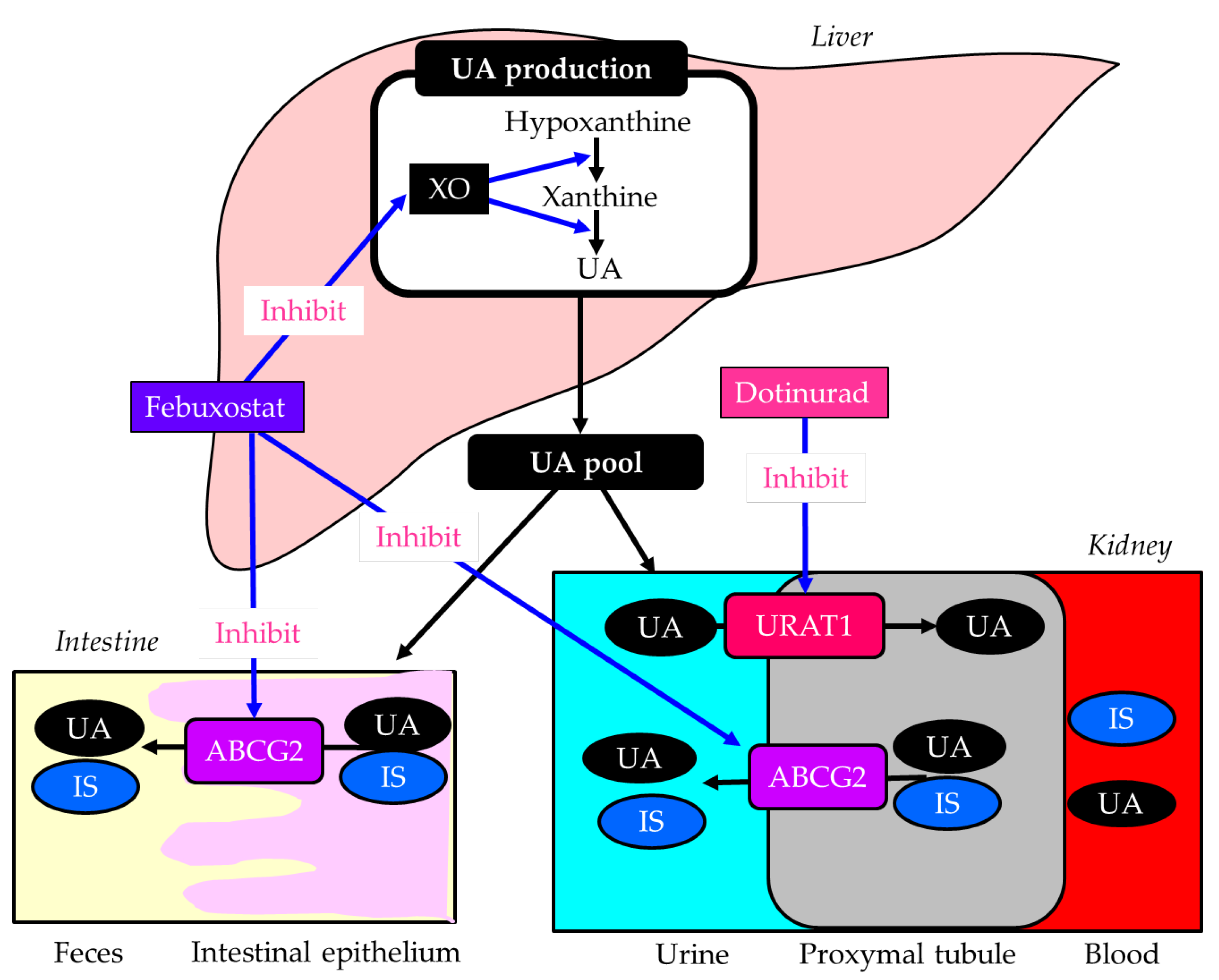
Dotinurad is a new therapeutic medicine for gout and hyperuricemia discovered by FUJI YAKUHIN, a Japanese pharmaceutical company. Dotinurad reduces serum UA levels by selectively inhibiting URAT1 [140]. Dotinurad is a potent selective URAT1 inhibitor with minimal effect on other urate transporters, such as ABCG2 [140], indicating that dotinurad reduces serum UA by not affecting IS clearance. Such characteristics of dotinurad are beneficial for DKD. In fact, dotinurad improved UACR and eGFR in patients with DKD/CKD in our study [141]. Another study showed that the proportion of patients with improved eGFR was significantly higher in patients with eGFR < 30 than in patients with 30 ≤ eGFR, and dotinurad significantly improved eGFR in advanced CKD patients with eGFR < 30 [142]. These results suggest that dotinurad may improve renal function in patients with severe renal dysfunction. A recent comparative study of the effects of dotinurad and febuxostat on renal function in CKD patients with hyperuricemia showed that dotinurad and febuxostat decreased serum UA, and eGFR improved only with dotinurad, but no change was observed with febuxostat, thus highlighting the renal protective effect of dotinurad beyond the reduction in serum UA levels [143]. The difference in the effects of dotinurad and febuxostat on eGFR may be due to different effects of these drugs on ABCG2 and the resulting effects on IS clearance. A very recent study evaluated switching from febuxostat to dotinurad and found that urinary IS increased significantly at weeks 4 and 12, and that IS clearance was significantly improved by dotinurad at week 24 [144], supporting our hypothesis.
Dotinurad can be a potential therapeutic option for DKD complicated with hyperuricemia.
7.2. Imeglimin
Imeglimin was discovered by the French company Poxel and developed in Japan by Sumitomo Pharma. It was approved in Japan in 2021, making it the first in the world. Imeglimin has a chemical structure similar to metformin, but unlike metformin, it is thought to exert its blood glucose-lowering effect by enhancing insulin secretion [145]. Imeglimin is a novel oral hypoglycemic agent with a unique mechanism of action that targets mitochondrial bioenergetics.
Figure 5.
Differential effects of metformin and imeglimin on mitochondria, with resulting differential renal protective effects and risk of lactic acidosis. Black arrows indicate effects, and red upward arrows indicate an increase in activity. AMPK, AMP-activated protein kinase; GSIS, glucose-stimulated insulin secretion; mGPDH, mitochondrial glycerol-3-phosphate dehydrogenase; NAMPT, nicotinamide phosphoribosyltransferase.
Figure 5.
Differential effects of metformin and imeglimin on mitochondria, with resulting differential renal protective effects and risk of lactic acidosis. Black arrows indicate effects, and red upward arrows indicate an increase in activity. AMPK, AMP-activated protein kinase; GSIS, glucose-stimulated insulin secretion; mGPDH, mitochondrial glycerol-3-phosphate dehydrogenase; NAMPT, nicotinamide phosphoribosyltransferase.
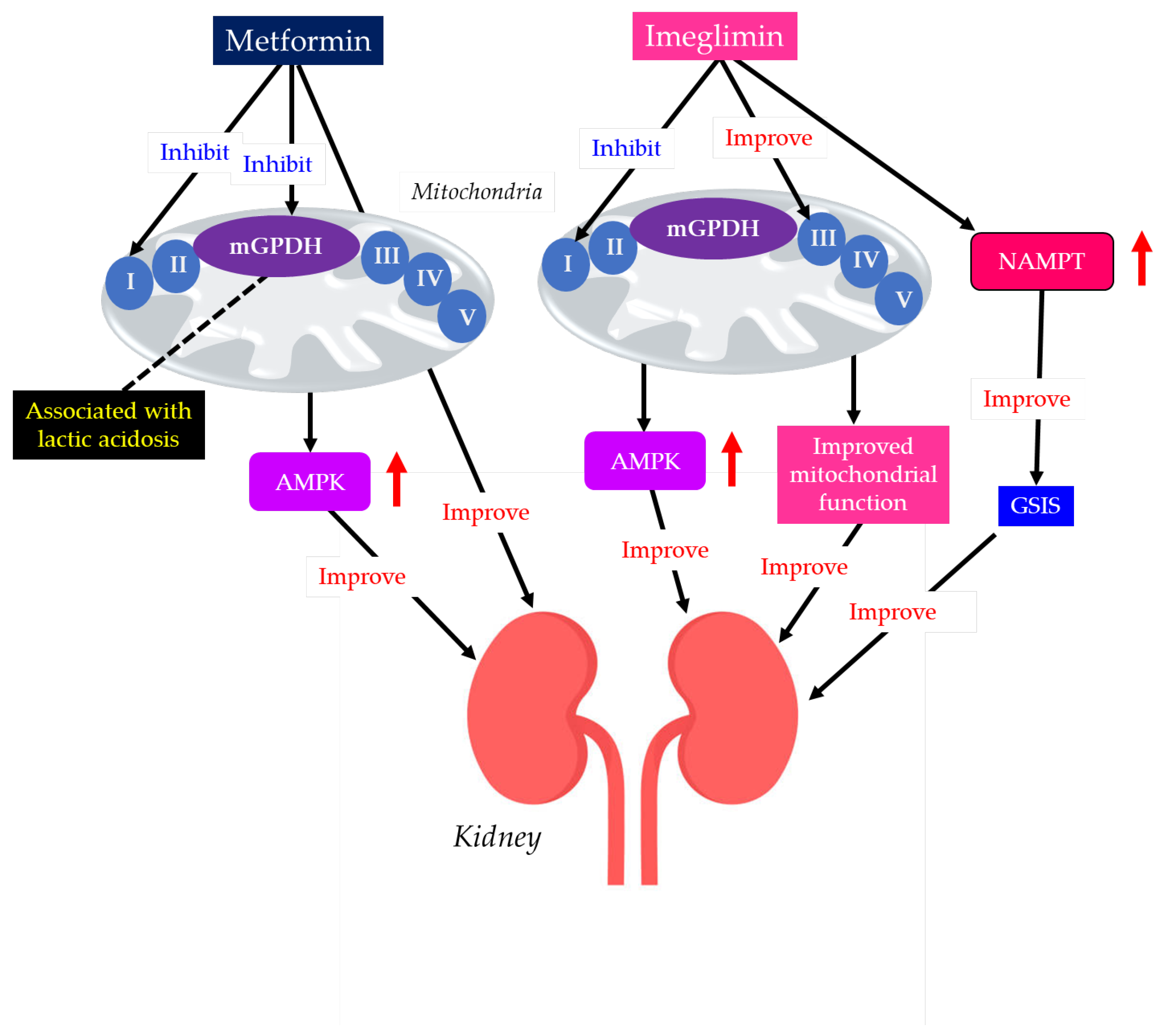
Metformin acts primarily by mildly and reversibly inhibiting mitochondrial complex I in the electron transport chain, especially in hepatocytes. This inhibition reduces ATP production, elevates the AMP/ATP ratio, and activates AMPK, thereby improving insulin resistance [146]. An improvement in insulin resistance reduces hepatic gluconeogenesis and increases glucose uptake in muscles [146].
Recently, a retrospective observational multicenter cohort study including 316 693 patients with type 2 diabetes demonstrated that the metformin group showed a lower incidence of doubling of serum creatinine (HR, 0.71; 95% CI, 0.65-0.77), eGFR ≤ 15 mL/min/1.73 m2 (HR 0.61; 95% CI, 0.53-0.71), and ESRD (HR 0.55; 95% CI, 0.47-0.66) [147]. The multivariable Cox survival model showed that metformin users had significantly better renal outcomes, with a notably lower risk of sustained eGFR of < 60 mL/min/1.73 m2 (HR, 0.71; 95% CI, 0.56-0.90) and new CKD onset (HR, 0.78; 95% CI, 0.65-0.94) [148].
Metformin protects the kidneys mainly via AMPK signaling and AMPK-independent pathways [149]. Metformin exerts multiple mechanisms of renal protection, including induction of autophagy, antioxidant properties, alleviation of ER stress, anti-inflammatory effects, attenuation of lipotoxicity, and antifibrotic effects [149]. Dysregulation of TGF-β signaling is implicated in renal fibrosis. Angiotensin, glucose, and oxidative stress induce TGF-β overexpression in tubular epithelial cells, macrophages, and renal interstitial fibroblasts. Metformin attenuated TGF-β expression in renal tissues from a rat model of renal fibrosis [150].
Imeglimin redirects substrate flux towards complex II, inhibits complex I, restores complex III activity in mitochondria, and promotes FA oxidation [145]. Imeglimin also decreases ROS production and improves mitochondrial and ER function. Imeglimin enhances glucose-stimulated insulin secretion (GSIS) by β-cells and inhibits β-cell apoptosis by maintaining mitochondrial and ER function and structure [145].
Pancreatic islet β-cell dysfunction, characterized by defective GSIS, is a predominant component of the pathophysiology of diabetes. Imeglimin acutely and directly amplifies GSIS in pancreatic islets by increasing the cellular nicotinamide adenine dinucleotide (NAD+) through the induction of nicotinamide phosphoribosyltransferase (NAMPT), along with the augmentation of glucose-induced ATP levels [151], which is the crucial difference between imeglimin and metformin.
Some individuals, such as patients with renal impairment, may not be prescribed metformin due to the risk of lactic acidosis [152]. Due to the predominant renal excretion, DKD patients treated with metformin are likely to develop lactic acidosis. A high dose of metformin or imeglimin was administered to rats with renal insufficiency, but only metformin developed fatal lactic acidosis [153]. Only metformin showed an inhibitory effect on the mitochondrial glycerol-3-phosphate dehydrogenase (mGPDH), resulting in a decrease in entry of glycerol into gluconeogenic flux, disrupting the glycerophosphate shuttle and inducing accumulation of cytosolic NADH, which is closely associated with the development of lactic acidosis [154,155], which is also the crucial difference between imeglimin and metformin.
The 90-day imeglimin treatment reduced albuminuria in Zucker rats [156]. This imeglimin treatment significantly reduced renal interstitial fibrosis, glomerular injury, and renal interstitial inflammation. In a retrospective study investigating the safety of imeglimin in patients with an eGFR <45 mL/min per 1.73 m2, no deterioration in renal or hepatic function was observed [157]. Furthermore, significant improvements in proteinuria and acidemia were observed. Another study also found that imeglimin reduced UACR in patients with type 2 diabetes [158].
Imeglimin activates AMPK in the same way as metformin [159], which is thought to explain its renal protective effect. Improvement of mitochondrial function and reduction of ER stress by imeglimin are thought to result in reduced vascular endothelial dysfunction and improved pathology in DKD [145]. The interaction of insulin with the kidneys is a dynamic, multifocal process, as it acts at multiple sites throughout the nephron [160]. Insulin acts on a range of tissues, from the glomerulus to the renal tubule, by modulating functions such as glomerular filtration, gluconeogenesis, natriuresis, glucose uptake, ion transport regulation, and apoptosis prevention [160]. Therefore, enhancement of GSIS by imgelimin may also contribute to renal protection [151].
7.3. Pemafibrate
Hypertriglyceridemia is a common feature of dyslipidemia in DKD and significantly contributes to its progression, acting as an independent predictor of albuminuria and renal function decline, driving renal damage through lipotoxicity, inflammation, and oxidative stress [161,162]. A large population cohort study showed that HDL-C <40 mg/dL in men and <50 mg/dL in women was associated with a 27% higher risk of low eGFR and a 28% higher risk of an eGFR reduction >30%, with a 24% higher risk of developing albuminuria and a 44% higher risk of developing one abnormality [163]. Low HDL reduces anti-inflammatory and antioxidant protection, contributing to renal structural damage [164].
Figure 6.
Effects of fenofibrate and pemefibrate on lipid, uric acid, and indoxyl sulfate metabolism. Black horizontal arrows indicate the flow of substance and effects, and red upward and downward arrows indicate an increase and a decrease of activity or substances, respectively. The red dotted arrows indicate activating effects, and the blue dotted arrows indicate inhibitory effects. ATP-binding cassette, subfamily G, 2 (ABCG2); HDL, high-density lipoprotein; IS, indoxyl sulfate; LPL, lipoprotein lipase; PPARα, peroxisome proliferator-activated receptor alpha; TG, triglyceride; UA, uric acid; URAT1, urate transporter 1; VLDL, very-low-density lipoprotein; XO, xanthine oxidase.
Figure 6.
Effects of fenofibrate and pemefibrate on lipid, uric acid, and indoxyl sulfate metabolism. Black horizontal arrows indicate the flow of substance and effects, and red upward and downward arrows indicate an increase and a decrease of activity or substances, respectively. The red dotted arrows indicate activating effects, and the blue dotted arrows indicate inhibitory effects. ATP-binding cassette, subfamily G, 2 (ABCG2); HDL, high-density lipoprotein; IS, indoxyl sulfate; LPL, lipoprotein lipase; PPARα, peroxisome proliferator-activated receptor alpha; TG, triglyceride; UA, uric acid; URAT1, urate transporter 1; VLDL, very-low-density lipoprotein; XO, xanthine oxidase.
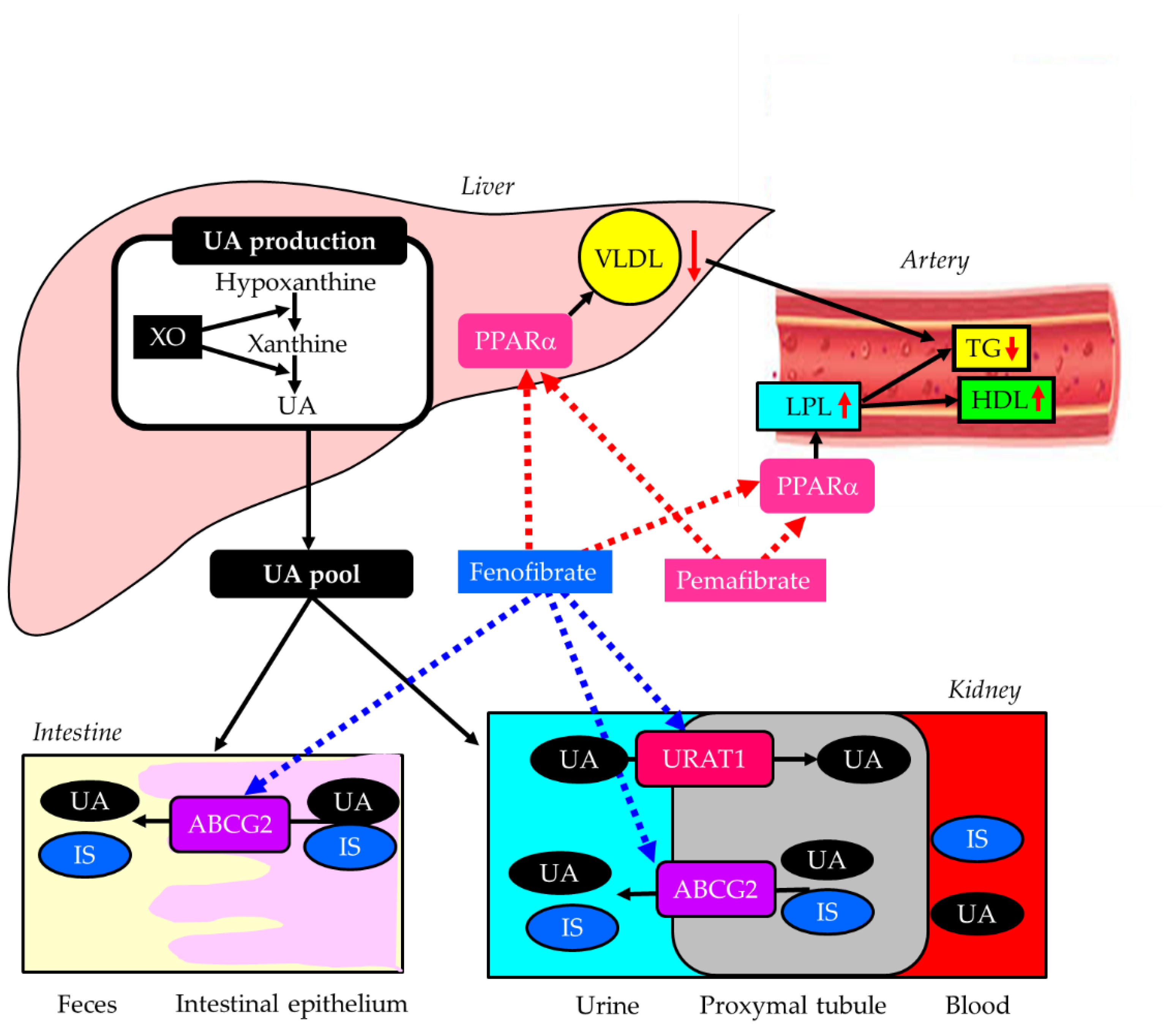
Fibrates are effective, first-line medications for managing hypertriglyceridemia and low HDL-C [165]. They act as peroxisome proliferator-activated receptor alpha (PPARα) agonists, increasing lipoprotein lipase (LPL) activity and reducing hepatic very-low density lipoprotein (VLDL) production [165]. Elevated LPL activity by fibrates also increases serum HDL-C [167]. Fibrates are particularly beneficial for combined high triglyceride (TG) and low HDL-C; therefore, managing such lipid abnormality with fibrates may be a promising option for renal protection. Fenofibrate offers notable reno-protective effects, particularly in DKD. It reduces albuminuria and slows the progression of kidney damage by suppressing inflammatory markers such as TGF-β, reducing oxidative stress, and improving endothelial function [166]. The FIELD study showed that the progression to albuminuria was significantly reduced by fenofibrate compared with placebo [167]. However, fibrates should be prescribed with caution in DKD patients, as impaired urinary excretion of fibrates can lead to serious adverse effects, such as rhabdomyolysis. The review of 76 published cases reported that gemfibrozil was the most frequent agent associated with rhabdomyolysis, followed by bezafibrate, fenofibrate, ciprofibrate, and clofibrate [168]. Twenty-three cases were associated with fibrate monotherapy. Sixteen cases had chronic renal failure before fibrate therapy [168]. Another concern when administering fenofibrate to patients with DKD is its inhibitory effect on ABCG2, a major transporter of IS [137]. Fenofibrate has been shown to inhibit human ABCG2 by over 80% [138]. The accumulation of uremic toxins, such as IS, in the kidney is associated with the progression of DKD [136].
Pemafibrate is a drug for the treatment of dyslipidemia developed by Japanese Kowa Co., Ltd. Pemafibrate is a novel, selective PPARα modulator with higher potency and selectivity than conventional fibrates [169]. Pemafibrate showed higher activity and more selectivity for PPARα than fenofibrate in vitro [169]. Additionally, since pemafibrate is excreted into bile, it can be used in patients with renal impairment, unlike conventional fibrates. Pemafibrate significantly attenuated histological tubular injury in FA overload nephropathy and decreased renal FA content and oxidative stress in mice, suggesting it is a promising drug for DKD [170]. Another animal study showed that decreases in GFR and increases in serum creatinine were markedly greater after conventional fibrate (fenofibrate, bezafibrate) use than after pemafibrate use, in a unilateral ureteral obstruction (UUO)-induced renal fibrosis model (UUO mice) and an adenine-induced CKD model (CKD mice) [171]. Pemafibrate suppressed the increase in gene expressions of collagen-I, fibronectin, and interleukin-1b in the kidneys of UUO mice. In CKD mice, pemafibrate suppressed the elevation in plasma creatinine and blood urea nitrogen levels, as well as renal fibrosis. Moreover, pemafibrate inhibited the upregulation of MCP-1, interleukin-1β, TNF-α, and interleukin-6 in the kidneys of CKD mice.
Fenofibrate was reported to decrease serum UA levels by increasing urinary excretion, most likely through inhibition of URAT1 [172]. We evaluated the effects of switching from fenofibrate to pemafibrate, and found that eGFR significantly increased at 3, 6, and 12 months despite an increase in serum UA levels in patients with type 2 diabetes [173]. Another study also found that the switching from fenofibrate to pemafibrate was significantly associated with an elevation in eGFR in diabetic patients with CKD [174].
8. Conclusion
Because overactivation of AT1R and MR, which are widely distributed in the kidney, is strongly associated with DKD development via various mechanisms, RASis and nsMRAs are considered strong tools for treating DKD. SGLT2is improve glomerular hypertension and UA metabolism by inhibiting the reabsorption of glucose and sodium, and suppress renal fibrosis by improving renal interstitial hypoxia. SGLT2is also improve podocytes and mesangial cells, both directly and by reducing oxidative stress and inflammation. The improvement of endothelial dysfunction by SGLT2is may also contribute significantly to the renal protection. GLP-1RAs directly and indirectly improve podocytes and mesangial cells, and alleviate endothelial dysfunction by improving metabolic abnormalities due to diabetes. GLP-1RAs increase natriuresis by inhibiting NHE3 and ENaC, thereby reducing intraglomerular pressure, contributing to renal protection.
Dotinurad, a UA-lowering drug, does not inhibit ABCG2, which is involved in the excretion of uremic toxins, unlike febuxostat, and therefore can be used safely in DKD patients. A small number of studies have shown that it also improves DKD. Imeglimin is structurally similar to metformin but has a different effect on mitochondria and therefore a different renal protective effect. Because imeglimin does not inhibit mGPDH, unlike metformin, it is thought that the risk of developing lactic acidosis is low even in DKD patients with renal impairment. Because pemafibrate is excreted in the bile, unlike conventional fibrates, it may be possible to use it in DKD patients with impaired renal function without concern for the development of rhabdomyolysis. Pemafibrate does not inhibit ABCG2, unlike fenofibrate, and therefore can be used safely in DKD patients.
We believe that the use of four dishes (RASis, nsMRAs, SGLT2is, GLP-1RAs) with DIP (dotinurad, imeglimin, pemafibrate) for DKD patients may contribute to improving the prognosis of DKD (Figure 7).
Author Contributions
H.Y., M.H., H.A. and H. K. conceived the review; H. Y. wrote the paper; H. K. edited the paper and provided critical guidance. All authors read and approved the final version of this paper.
Funding
This review research received no external funding.
Institutional Review Board Statement
Not applicableInformed Consent Statement: Not applicable. Data Availability Statement: No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest in relation to the present review paper.
Abbreviations
| ABCG2 | ATP-binding cassette, subfamily G, 2 |
| ACEis | angiotensin-converting enzyme inhibitors |
| AGEs | advanced glycation end products |
| AMPK | AMP-activated protein kinase |
| ARBs | AT1R blockers |
| AT1R | angiotensin II type 1 receptor |
| BMI | body mass index |
| CI | confidence interval |
| CKD | chronic kidney disease |
| DKD | diabetic kidney disease |
| ECM | extracellular matrix |
| eGFR | estimated glomerular filtration rate |
| ENaC | epithelial sodium channel |
| eNOS | endothelial nitric oxide synthase |
| ER | endoplasmic reticulum |
| ESRD | end-stage renal disease |
| FA | fatty acid |
| GBM | glomerular basement membrane |
| GLP-1RAs | glucagon-like peptide-1 receptor agonists |
| GLUT9 | glucose transporter 9 |
| GSIS | glucose-stimulated insulin secretion |
| HDL-C | high-density lipoprotein-cholesterol |
| HR | hazard ratio |
| IGF-1 | insulin-like growth factor 1 |
| IS | indoxyl sulfate |
| LDL-C | low-density lipoprotein-cholesterol |
| LPL | lipoprotein lipase |
| MAMs | mitochondria-associated endoplasmic reticulum membranes |
| MCP-1 | monocyte chemoattractant protein-1 |
| mGPDH | mitochondrial glycerol-3-phosphate dehydrogenase |
| MR | mineralocorticoid receptor |
| NAD+ | nicotinamide adenine dinucleotide |
| NAMPT | nicotinamide phosphoribosyltransferase |
| NF-κB | nuclear factor-κB |
| NHE3 | Na+/H+ exchanger 3 |
| NO | nitric oxide |
| NLRP3 | NOD-like receptor family, and pyrin domain-containing 3 |
| nsMRAs | non-steroidal mineralocorticoid receptor antagonists |
| PI3K-Akt | phosphatidylinositol 3'-kinase-Akt |
| PKC | protein kinase C |
| PPARα | peroxisome proliferator-activated receptor alpha |
| Rac1 | Rac family small guanosine triphosphatase 1 |
| RAS | renin-angiotensin system |
| RASis | RAS inhibitors |
| RCTs | randomized controlled trials |
| ROS | reactive oxygen species |
| SGLT2 | sodium-glucose cotransporter 2 |
| SGLT2is | sodium-glucose cotransporter 2 inhibitors |
| TG | triglyceride |
| TGF | tubuloglomerular feedback |
| TGF-β | transforming growth factor beta |
| TNF-α | tumor necrosis factor-alpha |
| UA | uric acid |
| URAT1 | urate transporter 1 |
| VEGF | vascular endothelial growth factor |
| VLDL | very-low-density lipoprotein |
| XO | xanthine oxidase |
References
- American Diabetes Association. Standards of care in diabetes - 2025. Diabetes. Care 2025, 48 (suppl 1), S1–344. [Google Scholar] [CrossRef] [PubMed]
- Sagoo, M.K.; Gnudi, L. Diabetic Nephropathy: An Overview. Methods. Mol. Biol. 2020, 2067, 3–7. [Google Scholar]
- Kidney Disease: Improving Global Outcomes (KDIGO) Diabetes Work Group. KDIGO 2022 Clinical Practice Guideline for Diabetes Management in Chronic Kidney Disease. Kidney. Int. 2022, 102(5S), S1–S127. [CrossRef] [PubMed]
- Hussain, S.; Jamali, M.C.; Habib, A.; Hussain, M.S.; Akhtar, M.; Najmi, A.K. Diabetic kidney disease: An overview of prevalence, risk factors, and biomarkers. Clinical. Epidemiology. And. Global. Health. 2021, 9, 2–6. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, C. Oxidative Stress: A Culprit in the Progression of Diabetic Kidney Disease. Antioxidants (Basel). 2024, 13, 455. [Google Scholar] [CrossRef]
- Ismail-Beigi, F.; Craven, T.; Banerji, M.A.; Basile, J.; Calles, J.; Cohen, R.M.; Cuddihy, R.; Cushman, W.C.; Genuth, S.; Grimm, R.H., Jr.; et al. Effect of intensive treatment of hyperglycaemia on microvascular outcomes in type 2 diabetes: an analysis of the ACCORD randomised trial. Lancet 2010, 376, 419–30. [Google Scholar] [CrossRef]
- Duckworth, W.; Abraira, C.; Moritz, T.; Reda, D.; Emanuele, N.; Reaven, P.D.; Zieve, F.J.; Marks, J.; Davis, S.N.; Hayward, R.; et al. Glucose control and vascular complications in veterans with type 2 diabetes. N. Engl. J. Med. 2009, 360, 129–139. [Google Scholar] [CrossRef]
- Wagnew, F.; Eshetie, S.; Kibret, G.D.; Zegeye, A.; Dessie, G.; Mulugeta, H.; Alemu, A. Diabetic nephropathy and hypertension in diabetes patients of sub-Saharan countries: a systematic review and meta-analysis. BMC. Res. Notes 2018, 11, 565. [Google Scholar] [CrossRef]
- Reynolds, K.; Gu, D.; Muntner, P.; Kusek, J.W.; Chen, J.; Wu, X.; Duan, X.; Chen, C.S.; Klag, M.J.; Whelton, P.K.; et al. A population-based, prospective study of blood pressure and risk for end-stage renal disease in China. J. Am. Soc. Nephrol. 2007, 18, 1928–1935. [Google Scholar] [CrossRef]
- Wudu, M.A.; Birhanu, T.A.; Tegegne, K.D.; Wondifraw, E.B. Incidence and predictors of diabetic kidney disease among type 2 diabetes mellitus adult patients in Ethiopia: a systematic review and meta-analysis. BMC. Endocr. Disord. 2025, 25, 190. [Google Scholar] [CrossRef]
- Tian, D.; Chen, Q.; Zeng, L.; Hao, Y. The impact of blood lipids and statins on renal function and mortality in patients with diabetic nephropathy: A meta-analysis. Acta. Pharm. 2025, 75, 1–22. [Google Scholar] [CrossRef]
- Zhao, Q.; Yi, X.; Wang, Z. Meta-Analysis of the Relationship between Abdominal Obesity and Diabetic Kidney Disease in Type 2 Diabetic Patients. Obes. Facts 2021, 14, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.F.; Chen, Y.; Yao, T.H.; Wu, Y.Z.; Dai, H.Z. Impact of body mass index on adverse kidney events in diabetes mellitus patients: A systematic-review and meta-analysis. World. J. Clin. Cases. 2024, 12, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.M.; Brown, N.J. The renin-angiotensin-aldosterone system and glucose homeostasis. Trends. Pharmacol. Sci. 2011, 32, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Manrique, C.; Lastra, G.; Gardner, M.; Sowers, J.R. The renin angiotensin aldosterone system in hypertension: roles of insulin resistance and oxidative stress. Med. Clin. North. Am. 2009, 93, 569–582. [Google Scholar] [CrossRef]
- Schütten, M.T.; Houben, A.J.; de Leeuw, P.W.; Stehouwer, C.D. The Link Between Adipose Tissue Renin-Angiotensin-Aldosterone System Signaling and Obesity-Associated Hypertension. Physiology (Bethesda). 2017, 32, 197–209. [Google Scholar] [CrossRef]
- Chen, J.; Li, D.; Schaefer, R.; Mehta, J.L. Cross-talk between dyslipidemia and renin-angiotensin system and the role of LOX-1 and MAPK in atherogenesis studies with the combined use of rosuvastatin and candesartan. Atherosclerosis. 2006, 184, 295–301. [Google Scholar] [CrossRef]
- Han, S.; Kim, S.A. New Era in Diabetic Kidney Disease Treatment: The Four Pillars and Strategies to Build Beyond. Electrolyte. Blood. Press. 2024, 22, 21–28. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Adeva-Contreras, L.; Fernández-Fernández, C.; Carneiro-Freire, N.; Domínguez-Montero, A. Histological Manifestations of Diabetic Kidney Disease and its Relationship with Insulin Resistance. Curr. Diabetes. Rev. 2023, 19, e280322202705. [Google Scholar] [CrossRef]
- Tonneijck, L.; Muskiet, M.H.; Smits, M.M.; van Bommel, E.J.; Heerspink, H.J.; van Raalte, D.H.; Joles, J.A. Glomerular Hyperfiltration in Diabetes: Mechanisms, Clinical Significance, and Treatment. J. Am. Soc. Nephrol. 2017, 28, 1023–1039. [Google Scholar] [CrossRef]
- Heerspink, H.J.; Perkins, B.A.; Fitchett, D.H.; Husain, M.; Cherney, D.Z. Sodium Glucose Cotransporter 2 Inhibitors in the Treatment of Diabetes Mellitus: Cardiovascular and Kidney Effects, Potential Mechanisms, and Clinical Applications. Circulation. 2016, 134, 752–772. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chang, Y.H.; Yang, S.Y.; Wu, K.D.; Chu, T.S. Update of pathophysiology and management of diabetic kidney disease. Journal. Of the Formosan. Medical. Association 2018, 117, 662–675. [Google Scholar] [CrossRef] [PubMed]
- Sano, M.; Takei, M.; Shiraishi, Y.; Suzuki, Y. Increased Hematocrit During Sodium-Glucose Cotransporter 2 Inhibitor Therapy Indicates Recovery of Tubulointerstitial Function in Diabetic Kidneys. J. Clin. Med. Res. 2016, 8, 844–847. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Liu, R.; Wu, S.; Chen, S.; Zhang, L.; Qiu, X.; Liu, K. The effect of arteriosclerosis on new-onset renal damage in diabetic patients. Endocr. J. 2023, 70, 173–183. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, P.; Liu, Y.; Chen, X.; He, Y.; Feng, J. Vascular inflammation, atherosclerosis, and lipid metabolism and the occurrence of non-high albuminuria diabetic kidney disease: A cross-sectional study. Diab. Vasc. Dis. Res. 2021, 18, 1479164121992524. [Google Scholar] [CrossRef]
- Thomas, H.Y.; Ford Versypt, A.N. Pathophysiology of mesangial expansion in diabetic nephropathy: mesangial structure, glomerular biomechanics, and biochemical signaling and regulation. J. Biol. Eng. 2022, 16, 19. [Google Scholar] [CrossRef]
- Cunanan, J.; Zhang, D.; Peired, A.J.; Barua, M. Podocytes in health and glomerular disease. Front. Cell. Dev. Biol. 2025, 13, 1564847. [Google Scholar] [CrossRef]
- Demirci, H. Glomerular alteration in diabetic kidney disease: from morphology to metabolism. Kidney. Res. Clin. Pract. Online ahead of print. 4 Jul 2025. [CrossRef]
- Liu, S.; Yuan, Y.; Xue, Y.; Xing, C.; Zhang, B. Podocyte Injury in Diabetic Kidney Disease: A Focus on Mitochondrial Dysfunction. Front. Cell. Dev. Biol. 2022, 10, 832887. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, D.; Kang, X.; Zhou, R.; Sun, Y.; Lian, F.; Tong, X. Signaling Pathways Involved in Diabetic Renal Fibrosis. Front. Cell. Dev. Biol. 2021, 9, 696542. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, M.; Cheng, C.K.; Li, Q. Tubular injury in diabetic kidney disease: molecular mechanisms and potential therapeutic perspectives. Front. Endocrinol (Lausanne) 2023, 14, 1238927. [Google Scholar] [CrossRef]
- Ma, T.; Li, X.; Zhu, Y.; Yu, S.; Liu, T.; Zhang, X.; Chen, D.; Du, S.; Chen, T.; Chen, S.; et al. Excessive Activation of Notch Signaling in Macrophages Promote Kidney Inflammation, Fibrosis, and Necroptosis. Front. Immunol. 2022, 13, 835879. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.J.; Toma, I.; Sipos, A.; Meer, E.J.; Vargas, S.L.; Peti-Peterdi, J. The collecting duct is the major source of prorenin in diabetes. Hypertension 2008, 51, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Karimi, F.; Maleki, M.; Movahedpour, A.; Alizadeh, M.; Kharazinejad, E.; Sabaghan, M. Overview of the renin-angiotensin system in diabetic nephropathy. Journal. of the Renin-Angiotensin. Aldosterone. System 2024, 25, 1–15. [Google Scholar] [CrossRef]
- Tang, J.; Wysocki, J.; Ye, M.; Vallés, P.G.; Rein, J.; Shirazi, M.; Bader, M.; Gomez, R.A.; Sequeira-Lopez, M.S.; Afkarian, M.; et al. Urinary Renin in Patients and Mice With Diabetic Kidney Disease. Hypertension. 2019, 74, 83–94. [Google Scholar] [CrossRef]
- Leite, A.P.O.; Li, X.C.; Nwia, S.M.; Hassan, R.; Zhuo, J.L. Angiotensin II and AT1a Receptors in the Proximal Tubules of the Kidney: New Roles in Blood Pressure Control and Hypertension. Int. J. Mol. Sci. 2022, 23, 2402. [Google Scholar] [CrossRef]
- Burns, K.D. Angiotensin II and its receptors in the diabetic kidney. Am. J. Kidney. Dis. 2000, 36, 449–467. [Google Scholar] [CrossRef]
- Wong, W.T.; Tian, X.Y.; Xu, A.; Ng, C.F.; Lee, H.K.; Chen, Z.Y.; Au, C.L.; Yao, X.; Huang, Y. Angiotensin II type 1 receptor-dependent oxidative stress mediates endothelial dysfunction in type 2 diabetic mice. Antioxid. Redox. Signal. 2010, 13, 757–768. [Google Scholar] [CrossRef]
- Vavrinec, P.; Henning, R.H.; Goris, M.; Vavrincova-Yaghi, D.; Buikema, H.; van Dokkum, R.P. Vascular smooth muscle function of renal glomerular and interlobar arteries predicts renal damage in rats. Am. J. Physiol. Renal. Physiol. 2012, 303, F1187–F195. [Google Scholar] [CrossRef]
- Ricciardi, C.A.; Gnudi, L. Kidney disease in diabetes: From mechanisms to clinical presentation and treatment strategies. Metabolism 2021, 124, 154890. [Google Scholar] [CrossRef]
- Patinha, D.; Fasching, A.; Pinho, D.; Albino-Teixeira, A.; Morato, M.; Palm, F. Angiotensin II contributes to glomerular hyperfiltration in diabetic rats independently of adenosine type I receptors. Am. J. Physiol. Renal. Physiol. 2013, 304, F614–622. [Google Scholar] [CrossRef]
- Nakamura, T.; Girerd, S.; Jaisser, F.; Barrera-Chimal, J. Nonepithelial mineralocorticoid receptor activation as a determinant of kidney disease. Kidney. Int. Suppl (2011) 2022, 12, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Epstein, M.; Kovesdy, C.P.; Clase, C.M.; Sood, M.M.; Pecoits-Filho, R. Aldosterone, Mineralocorticoid Receptor Activation, and CKD: A Review of Evolving Treatment Paradigms. Am. J. Kidney. Dis. 2022, 80, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Nagase, M.; Yoshida, S.; Kawarazaki, W.; Kurihara, H.; Tanaka, H.; Miyoshi, J.; Takai, Y.; Fujita, T. Modification of mineralocorticoid receptor function by Rac1 GTPase: implication in proteinuric kidney disease. Nat. Med. 2008, 14, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Ishizawa, K.; Uchida, S. Mineralocorticoid receptor as a therapeutic target in chronic kidney disease and hypertension. Hypertens. Res. 2017, 40, 221–225. [Google Scholar] [CrossRef]
- Rafiq, K.; Hitomi, H.; Nakano, D.; Nishiyama, A. Pathophysiological roles of aldosterone and mineralocorticoid receptor in the kidney. J. Pharmacol. Sci. 2011, 115, 1–7. [Google Scholar] [CrossRef]
- Tang, C.; Livingston, M.J.; Liu, Z.; Dong, Z. Autophagy in kidney homeostasis and disease. Nat. Rev. Nephrol. 2020, 16, 489–508. [Google Scholar] [CrossRef]
- Dong, D.; Fan, T.T.; Ji, Y.S.; Yu, J.Y.; Wu, S.; Zhang, L. Spironolactone alleviates diabetic nephropathy through promoting autophagy in podocytes. Int. Urol. Nephrol. 2019, 51, 755–764. [Google Scholar] [CrossRef]
- Bai, M.; Chen, Y.; Zhao, M.; Zhang, Y.; He, J.C.; Huang, S.; Jia, Z.; Zhang, A. NLRP3 inflammasome activation contributes to aldosterone-induced podocyte injury. Am. J. Physiol. Renal. Physiol. 2017, 312, F556–F564. [Google Scholar] [CrossRef]
- Zhu, J.J.; Chen, Y.P.; Yang, M.; Liu, B.L.; Dong, J.; Dong, H.R.; Rui, H.L.; Cheng, H. Aldosterone is involved in the pathogenesis of obesity-related glomerulopathy through activation of Wnt/β-catenin signaling in podocytes. Mol. Med. Rep. 2018, 17, 4589–4598. [Google Scholar] [CrossRef]
- Davel, A.P.; Anwar, I.J.; Jaffe, I.Z. The endothelial mineralocorticoid receptor: mediator of the switch from vascular health to disease. Curr. Opin. Nephrol. Hypertens. 2017, 26, 97–104. [Google Scholar] [CrossRef]
- Caprio, M.; Newfell, B.G.; la Sala, A.; Baur, W.; Fabbri, A.; Rosano, G.; Mendelsohn, M.E.; Jaffe, I.Z. Functional mineralocorticoid receptors in human vascular endothelial cells regulate intercellular adhesion molecule-1 expression and promote leukocyte adhesion. Circ. Res. 2008, 102, 1359–1367. [Google Scholar] [CrossRef]
- Taye, A.; Morawietz, H. Spironolactone inhibits NADPH oxidase-induced oxidative stress and enhances eNOS in human endothelial cells. Iran. J. Pharm. Res. 2011, 10, 329–337. [Google Scholar]
- Iwashima, F.; Yoshimoto, T.; Minami, I.; Sakurada, M.; Hirono, Y.; Hirata, Y. Aldosterone induces superoxide generation via Rac1 activation in endothelial cells. Endocrinology. 2008, 149, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Nagata, D.; Takahashi, M.; Sawai, K.; Tagami, T.; Usui, T.; Shimatsu, A.; Hirata, Y.; Naruse, M. Molecular mechanism of the inhibitory effect of aldosterone on endothelial NO synthase activity. Hypertension 2006, 48, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.M.; Fogo, A.B. The role of mineralocorticoid receptor activation in kidney inflammation and fibrosis. Kidney. Int. Suppl. (2011) 2022, 12, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Pitzer, A.; Kleyman, T.R.; Kirabo, A. Kidney Tubular IL-1β ENaCtivation in Diabetes and Salt-Sensitive Hypertension. Circ. Res. 2022, 131, 74–76. [Google Scholar] [CrossRef]
- Pavlov, T.S.; Palygin, O.; Isaeva, E.; Levchenko, V.; Khedr, S.; Blass, G.; Ilatovskaya, D.V.; Cowley, A.W., Jr.; Staruschenko, A. NOX4-dependent regulation of ENaC in hypertension and diabetic kidney disease. FASEB. J. 2020, 34, 13396–13408. [Google Scholar] [CrossRef]
- Usher, M.G.; Duan, S.Z.; Ivaschenko, C.Y.; Frieler, R.A.; Berger, S.; Schütz, G.; Lumeng, C.N.; Mortensen, R.M. Myeloid mineralocorticoid receptor controls macrophage polarization and cardiovascular hypertrophy and remodeling in mice. J. Clin. Invest. 2010, 120, 3350–3364. [Google Scholar] [CrossRef]
- Vinuesa, E.; Hotter, G.; Jung, M.; Herrero-Fresneda, I.; Torras, J.; Sola, A. Macrophage involvement in the kidney repair phase after ischaemia/reperfusion injury. J. Pathol. 2008, 214, 104–113. [Google Scholar] [CrossRef]
- Barrera-Chimal, J.; Estrela, G.R.; Lechner, S.M.; Giraud, S.; El Moghrabi, S.; Kaaki, S.; Kolkhof, P.; Hauet, T.; Jaisser, F. The myeloid mineralocorticoid receptor controls inflammatory and fibrotic responses after renal injury via macrophage interleukin-4 receptor signaling. Kidney. Int. 2018, 93, 1344–1355. [Google Scholar] [CrossRef]
- Barrera-Chimal, J.; Rocha, L.; Amador-Martínez, I.; Pérez-Villalva, R.; González, R.; Cortés-González, C.; Uribe, N.; Ramírez, V.; Berman, N.; Gamba, G.; et al. Delayed spironolactone administration prevents the transition from acute kidney injury to chronic kidney disease through improving renal inflammation. Nephrol. Dial. Transplant. 2019, 34, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chen, Z.; Park, C.; Centrella, M.; McCarthy, T.; Chen, L.; Al-Omari, A.; Moeckel, G.W. Aldosterone stimulates fibronectin synthesis in renal fibroblasts through mineralocorticoid receptor-dependent and independent mechanisms. Gene 2013, 531, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Koszegi, S.; Molnar, A.; Lenart, L.; Hodrea, J.; Balogh, D.B.; Lakat, T.; Szkibinszkij, E.; Hosszu, A.; Sparding, N.; Genovese, F.; et al. RAAS inhibitors directly reduce diabetes-induced renal fibrosis via growth factor inhibition. J. Physiol. 2019, 597, 193–209. [Google Scholar] [CrossRef] [PubMed]
- An, W.; Jaffe, I.Z. Emerging Roles for Endothelial Mineralocorticoid Receptor in Disease Pathogenesis: Potential New Indications for MR Antagonism. Current. Opinion. in Physiology 2025, 100893. [Google Scholar]
- Kobori, H.; Mori, H.; Masaki, T.; Nishiyama, A. Angiotensin II blockade and renal protection. Curr. Pharm. Des. 2013, 19, 3033–3042. [Google Scholar] [CrossRef]
- Brenner, B.M.; Cooper, M.E.; de Zeeuw, D.; Keane, W.F.; Mitch, W.E.; Parving, H.H.; Remuzzi, G.; Snapinn, S.M.; Zhang, Z.; Shahinfar, S.; et al. RENAAL Study Investigators. Effects of losartan on renal and cardiovascular outcomes in patients with type 2 diabetes and nephropathy. N. Engl. J. Med. 2001, 345, 861–869. [Google Scholar] [CrossRef]
- Lewis, E.J.; Hunsicker, L.G.; Clarke, W.R.; Berl, T.; Pohl, M.A.; Lewis, J.B.; Ritz, E.; Atkins, R.C.; Rohde, R.; Raz, I.; et al. Collaborative Study Group. Renoprotective effect of the angiotensin-receptor antagonist irbesartan in patients with nephropathy due to type 2 diabetes. N. Engl. J. Med. 2001, 345, 851–860. [Google Scholar] [CrossRef]
- Currie, G.; Taylor, A.H.; Fujita, T.; Ohtsu, H.; Lindhardt, M.; Rossing, P.; Boesby, L.; Edwards, N.C.; Ferro, C.J.; Townend, J.N.; et al. Effect of mineralocorticoid receptor antagonists on proteinuria and progression of chronic kidney disease: a systematic review and meta-analysis. BMC. Nephrol. 2016, 17, 127. [Google Scholar] [CrossRef]
- Grune, J.; Beyhoff, N.; Smeir, E.; Chudek, R.; Blumrich, A.; Ban, Z.; Brix, S.; Betz, I.R.; Schupp, M.; Foryst-Ludwig, A.; et al. Selective Mineralocorticoid Receptor Cofactor Modulation as Molecular Basis for Finerenone's Antifibrotic Activity. Hypertension 2018, 71, 599–608. [Google Scholar] [CrossRef]
- Bakris, G.L.; Agarwal, R.; Chan, J.C.; Cooper, M.E.; Gansevoort, R.T.; Haller, H.; Remuzzi, G.; Rossing, P.; Schmieder, R.E.; Nowack, C.; et al. Effect of Finerenone on Albuminuria in Patients With Diabetic Nephropathy: A Randomized Clinical Trial. JAMA. 2015, 314, 884–894. [Google Scholar] [CrossRef]
- Bakris, G.L.; Agarwal, R.; Anker, S.D.; Pitt, B.; Ruilope, L.M.; Rossing, P.; Kolkhof, P.; Nowack, C.; Schloemer, P.; Joseph, A.; et al. Effect of Finerenone on Chronic Kidney Disease Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2020, 383, 2219–2229. [Google Scholar] [CrossRef]
- Parving, H.H.; Brenner, B.M.; McMurray, J.J.; de Zeeuw, D.; Haffner, S.M.; Solomon, S.D.; Chaturvedi, N.; Persson, F.; Desai, A.S.; Nicolaides, M.; et al. Cardiorenal end points in a trial of aliskiren for type 2 diabetes. N. Engl. J. Med. 2012, 367, 2204–2213. [Google Scholar] [CrossRef] [PubMed]
- Fried, L.F.; Emanuele, N.; Zhang, J.H.; Brophy, M.; Conner, T.A.; Duckworth, W.; Leehey, D.J.; McCullough, P.A.; O'Connor, T.; Palevsky, P.M.; et al. Combined angiotensin inhibition for the treatment of diabetic nephropathy. N. Engl. J. Med. 2013, 369, 1892–1903. [Google Scholar] [CrossRef] [PubMed]
- Mogi, M. Aldosterone breakthrough from a pharmacological perspective. Hypertens. Res. 2022, 45, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Wanner, C.; Inzucchi, S.E.; Lachin, J.M.; Fitchett, D.; von Eynatten, M.; Mattheus, M.; Johansen, O.E.; Woerle, H.J.; Broedl, U.C.; Zinman, B.; et al. Empagliflozin and Progression of Kidney Disease in Type 2 Diabetes. N. Engl. J. Med. 2016, 375, 323–334. [Google Scholar] [CrossRef]
- Neal, B.; Perkovic, V.; Mahaffey, KW.; de Zeeuw, D.; Fulcher, G.; Erondu, N.; Shaw, W.; Law, G.; Desai, M.; Matthews, D.R.; et al. Canagliflozin and Cardiovascular and Renal Events in Type 2 Diabetes. N. Engl. J. Med. 2017, 377, 644–657. [Google Scholar] [CrossRef]
- Perkovic, V.; Jardine, M.J.; Neal, B.; Bompoint, S.; Heerspink, H.J.L.; Charytan, D.M.; Edwards, R.; Agarwal, R.; Bakris, G.; Bull, S.; et al. Canagliflozin and Renal Outcomes in Type 2 Diabetes and Nephropathy. N. Engl. J. Med. 2019, 380, 2295–2306. [Google Scholar] [CrossRef]
- Heerspink, H.J.L.; Stefánsson, B.V.; Correa-Rotter, R.; Chertow, G.M.; Greene, T.; Hou, F.F.; Mann, J.F.E.; McMurray, J.J.V.; Lindberg, M.; Rossing, P.; et al. Dapagliflozin in Patients with Chronic Kidney Disease. N. Engl. J. Med. 2020, 383, 1436–1446. [Google Scholar] [CrossRef]
- Herrington, W.G.; Staplin, N.; Wanner, C.; Green, J.B.; Hauske, S.J.; Emberson, J.R.; Preiss, D.; Judge, P.; Mayne, K.J.; Ng, S.Y.A.; et al. Empagliflozin in Patients with Chronic Kidney Disease. N. Engl. J. Med. 2023, 388, 117–127. [Google Scholar]
- Vallon, V.; Mühlbauer, B.; Osswald, H. Adenosine and kidney function. Physiol. Rev. 2006, 86, 901–940. [Google Scholar] [CrossRef]
- Vallon, V.; Richter, K.; Blantz, R.C.; Thomson, S.; Osswald, H. Glomerular hyperfiltration in experimental diabetes mellitus: Potential role of tubular reabsorption. J. Am. Soc. Nephrol. 1999, 10, 2569–2576. [Google Scholar] [CrossRef] [PubMed]
- Thomson, S.C.; Rieg, T.; Miracle, C.; Mansoury, H.; Whaley, J.; Vallon, V.; Singh, P. Acute and chronic effects of SGLT2 blockade on glomerular and tubular function in the early diabetic rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R75–R83. [Google Scholar] [CrossRef] [PubMed]
- Lambers Heerspink, H.J.; de Zeeuw, D.; Wie, L.; Leslie, B.; List, J. Dapagliflozin a glucose-regulating drug with diuretic properties in subjects with type 2 diabetes. Diabetes. Obes. Metab. 2013, 15, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Katsuyama, H. A Possible Mechanism for Renoprotective Effect of Sodium-Glucose Cotransporter 2 Inhibitor: Elevation of Erythropoietin Production. J. Clin. Med. Res. 2017, 9, 178–179. [Google Scholar] [CrossRef]
- Toba, H.; Sawai, N.; Morishita, M.; Murata, S.; Yoshida, M.; Nakashima, K.; Morita, Y.; Kobara, M.; Nakata, T. Chronic treatment with recombinant human erythropoietin exerts renoprotective effects beyond hematopoiesis in streptozotocin-induced diabetic rat. Eur. J. Pharmacol. 2009, 612, 106–114. [Google Scholar] [CrossRef]
- Ruester, C.; Franke, S.; Bondeva, T.; Wolf, G. Erythropoietin protects podocytes from damage by advanced glycation end-products. Nephron. Exp. Nephrol. 2011, 117, e21–30. [Google Scholar] [CrossRef]
- Loeffler, I.; Ruster, C.; Franke, S.; Liebisch, M.; Wolf, G. Erythropoietin ameliorates podocyte injury in advanced diabetic nephropathy in the db/db mouse. Am. J. Physiol. Renal. Physiol. 2013, 305, F911–918. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, X.; Sun, J.; Qian, Q.; Yu, J.; An, X. Hyperuricemia Predicts the Progression of Type 2 Diabetic Kidney Disease in Chinese Patients. Diabetes. Ther. 2023, 14, 581–591. [Google Scholar] [CrossRef]
- Yanai, H.; Adachi, H.; Hakoshima, M.; Katsuyama, H. Molecular Biological and Clinical Understanding of the Pathophysiology and Treatments of Hyperuricemia and Its Association with Metabolic Syndrome, Cardiovascular Diseases and Chronic Kidney Disease. Int. J. Mol. Sci. 2021, 22, 9221. [Google Scholar] [CrossRef]
- Gibson, T.; Waterworth, R.; Hatfield, P.; Robinson, G.; Bremner, K. Hyperuricaemia, gout and kidney function in New Zealand Maori men. Br. J. Rheumatol. 1984, 23, 276–282. [Google Scholar] [CrossRef]
- Simmonds, H.A.; McBride, M.B.; Hatfield, P.J.; Graham, R.; McCaskey, J.; Jackson, M. Polynesian women are also at risk for hyperuricaemia and gout because of a genetic defect in renal urate handling. Br. J. Rheumatol. 1994, 33, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, A.; Kimura, H.; Chairoungdua, A.; Shigeta, Y.; Jutabha, P.; Cha, S.H.; Hosoyamada, M.; Takeda, M.; Sekine, T.; Igarashi, T.; et al. Molecular identification of a renal urate anion exchanger that regulates blood urate levels. Nature 2002, 417, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sanna, S.; Maschio, A.; Busonero, F.; Usala, G.; Mulas, A.; Lai, S.; Dei, M.; Orrù, M.; Albai, G. The GLUT9 gene is associated with serum uric acid levels in Sardinia and Chianti cohorts. PLoS. Genet. 2007, 3, e194. [Google Scholar] [CrossRef] [PubMed]
- Vitart, V.; Rudan, I.; Hayward, C.; Gray, N.K.; Floyd, J.; Palmer, C.N.; Knott, S.A.; Kolcic, I.; Polasek, O.; Graessler, J.; et al. SLC2A9 is a newly identified urate transporter influencing serum urate concentration, urate excretion and gout. Nat. Genet. 2008, 40, 437–442. [Google Scholar] [CrossRef]
- Matsuo, H.; Takada, T.; Ichida, K.; Nakamura, T.; Nakayama, A.; Ikebuchi, Y.; Ito, K.; Kusanagi, Y.; Chiba, T.; Tadokoro, S. Common defects of ABCG2, a high-capacity urate exporter, cause gout: A function-based genetic analysis in a Japanese population. Sci. Transl. Med. 2009, 1, 5ra11. [Google Scholar] [CrossRef]
- Akbari, A.; Rafiee, M.; Sathyapalan, T.; Sahebkar, A. Impacts of Sodium/Glucose Cotransporter-2 Inhibitors on Circulating Uric Acid Concentrations: A Systematic Review and Meta-Analysis. J. Diabetes. Res. 2022, 2022, 7520632. [Google Scholar] [CrossRef]
- Banerjee, M.; Pal, R.; Maisnam, I.; Chowdhury, S.; Mukhopadhyay, S. Serum uric acid lowering and effects of sodium-glucose cotransporter-2 inhibitors on gout: A meta-analysis and meta-regression of randomized controlled trials. Diabetes. Obes. Metab. 2023, 25, 2697–2703. [Google Scholar] [CrossRef]
- Zapf, A.M.; Woodward, O.M. SGLT2 Inhibitors and uric acid homeostasis. Gout. Urate. Cryst. Depos. Dis. 2024, 2, 157–172. [Google Scholar] [CrossRef]
- Sanchez-Lozada, L.G.; Lanaspa, M.A.; Rodriguez-Iturbe, B.; Brown, J.M.; Madero, M.; Johnson, R.J. Sodium-glucose cotransporter 2 inhibitors and uric acid. Nephron 2025, 149, 488–492. [Google Scholar] [CrossRef]
- Billing, A.M.; Kim, Y.C.; Gullaksen, S.; Schrage, B.; Raabe, J.; Hutzfeldt, A.; Demir, F.; Kovalenko, E.; Lassé, M.; Dugourd, A.; et al. Metabolic communication by SGLT2 inhibition. Circulation. 2024, 149, 860–884. [Google Scholar] [CrossRef]
- Suijk, D.L.S.; van Baar, M.J.B.; van Bommel, E.J.M.; Iqbal, Z.; Krebber, M.M.; Vallon, V.; Touw, D.; Hoorn, E.J.; Nieuwdorp, M.; Kramer, M.M.H.; et al. SGLT2 inhibition and uric acid excretion in patients with type 2 diabetes and normal kidney function. Clin. J. Am. Soc. Nephrol. 2022, 17, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J. Uric acid and the cardio-renal effects of SGLT2 inhibitors. Diabetes. Obes. Metab. 2019, 21, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.H.; Chang, Y.P.; Li, T.; Han, F.; Li, C.J.; Li, X.Y.; Xue, M.; Cheng, Y.; Meng, Z.Y.; Han, Z.; Sun, B.; Chen, L.M. Empagliflozin Attenuates Hyperuricemia by Upregulation of ABCG2 via AMPK/AKT/CREB Signaling Pathway in Type 2 Diabetic Mice. Int. J. Biol. Sci. 2020, 16, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Maki, T.; Maeno, S.; Maeda, Y.; Yamato, M.; Sonoda, N.; Ogawa, Y.; Wakisaka, M.; Inoguchi, T. Amelioration of diabetic nephropathy by SGLT2 inhibitors independent of its glucose-lowering effect: A possible role of SGLT2 in mesangial cells. Sci. Rep. 2019, 9, 4703. [Google Scholar] [CrossRef]
- Tagawa, A.; Yasuda, M.; Kume, S.; Yamahara, K.; Nakazawa, J.; Chin-Kanasaki, M.; Araki, H.; Araki, S.; Koya, D.; Asanuma, K.; et al. Impaired Podocyte Autophagy Exacerbates Proteinuria in Diabetic Nephropathy. Diabetes 2016, 65, 755–767. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Li, S.S.; He, Y.X.; Yan, L.J.; Lv, F.; Liang, Q.M.; Gan, Y.H.; Han, L.P.; Xu, H.D.; Li, Y.C.; et al. SGLT2 inhibitors alleviated podocyte damage in lupus nephritis by decreasing inflammation and enhancing autophagy. Ann. Rheum. Dis. 2023, 82, 1328–1340. [Google Scholar] [CrossRef]
- Ge, M.; Molina, J.; Kim, J.J.; Mallela, S.K.; Ahmad, A.; Varona Santos, J.; Al-Ali, H.; Mitrofanova, A.; Sharma, K.; Fontanesi, F.; et al. Empagliflozin reduces podocyte lipotoxicity in experimental Alport syndrome. Elife. 2023, 12, e83353. [Google Scholar] [CrossRef]
- Hudkins, K.L.; Li, X.; Holland, A.L.; Swaminathan, S.; Alpers, C.E. Regression of diabetic nephropathy by treatment with empagliflozin in BTBR ob/ob mice. Nephrol. Dial. Transpl. 2022, 37, 847–859. [Google Scholar] [CrossRef]
- Rowland, A.A.; Voeltz, G.K. Endoplasmic reticulum-mitochondria contacts: function of the junction. Nat. Rev. Mol. Cell. Biol. 2012, 13, 607–625. [Google Scholar] [CrossRef]
- Li, X.; Yang, Q.; Liu, S.; Song, S.; Wang, C. Mitochondria-associated endoplasmic reticulum membranes promote mitochondrial fission through AKAP1-Drp1 pathway in podocytes under high glucose conditions. Exp. Cell. Res. 2023, 424, 113512. [Google Scholar] [CrossRef]
- Wei, X.; Wei, X.; Lu, Z.; Li, L.; Hu, Y.; Sun, F.; Jiang, Y.; Ma, H.; Zheng, H.; Yang, G.; et al. Activation of TRPV1 channel antagonizes diabetic nephropathy through inhibiting endoplasmic reticulum-mitochondria contact in podocytes. Metabolism. 2020, 105, 154182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, S.; Zhu, P.; Hu, S.; Chen, Y.; Ren, J. Empagliflozin rescues diabetic myocardial microvascular injury via AMPK-mediated inhibition of mitochondrial fission. Redox. Biol. 2018, 15, 335–346. [Google Scholar] [CrossRef]
- Ren, F.F.; Xie, Z.Y.; Jiang, Y.N.; Guan, X.; Chen, Q.Y.; Lai, T.F.; Li, L. Dapagliflozin attenuates pressure overload-induced myocardial remodeling in mice via activating SIRT1 and inhibiting endoplasmic reticulum stress. Acta. Pharmacol. Sin. 2022, 43, 1721–1732. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Q.; Jiang, X.; Song, S.; Zou, W.; Yang, Q.; Liu, S.; Chen, S.; Wang, C. Inhibition of SGLT2 protects podocytes in diabetic kidney disease by rebalancing mitochondria-associated endoplasmic reticulum membranes. Cell. Commun. Signal. 2024, 22, 534. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Tanabe, K.; Croker, B.P.; Johnson, R.J.; Grant, M.B.; Kosugi, T.; Li, Q. Endothelial dysfunction as a potential contributor in diabetic nephropathy. Nat. Rev. Nephrol. 2011, 7, 36–44. [Google Scholar] [CrossRef]
- Yanai, H.; Adachi, H.; Hakoshima, M.; Katsuyama, H. Significance of Endothelial Dysfunction Amelioration for Sodium-Glucose Cotransporter 2 Inhibitor-Induced Improvements in Heart Failure and Chronic Kidney Disease in Diabetic Patients. Metabolites. 2023, 13, 736. [Google Scholar] [CrossRef]
- Mann, J.F.E.; Ørsted, D.D.; Brown-Frandsen, K.; Marso, S.P.; Poulter, N.R.; Rasmussen, S.; Tornøe, K.; Zinman, B.; Buse, J.B. LEADER Steering Committee and Investigators. Liraglutide and Renal Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2017, 377, 839–848. [Google Scholar] [CrossRef]
- Perkovic, V.; Tuttle, K.R.; Rossing, P.; Mahaffey, K.W.; Mann, J.F.E.; Bakris, G.; Baeres, F.M.M.; Idorn, T.; Bosch-Traberg, H.; Lausvig, N.L.; et al. Effects of Semaglutide on Chronic Kidney Disease in Patients with Type 2 Diabetes. N. Engl. J. Med. 2024, 391, 109–121. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Qiu, X.J.; Dong, S.S.; Zhou, L.N.; Zhu, Y.; Wang, M.D.; Jin, L.W. MicroRNA-770-5p is involved in the development of diabetic nephropathy through regulating podocyte apoptosis by targeting TP53 regulated inhibitor of apoptosis 1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1248–1256. [Google Scholar]
- Ren, Q.; Yu, S.Y. CD2-associated protein participates in podocyte apoptosis via PI3K/Akt signaling pathway. J. Recept. Signal. Transduct. Res. 2016, 36, 288–291. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Y.; Long, D.; Wu, Y.; Liu, F. GLP-1 receptor agonist, liraglutide, protects podocytes from apoptosis in diabetic nephropathy by promoting white fat browning. Biochem. Biophys. Res. Commun. 2023, 664, 142–151. [Google Scholar] [CrossRef]
- Jiang, X.; Stockwell, B. R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell. Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Daenen, K.; Andries, A.; Mekahli, D.; Van Schepdael, A.; Jouret, F.; Bammens, B. Oxidative stress in chronic kidney disease. Pediatr. Nephrol. 2019, 34, 975–991. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Song, J.X.; Zhang, Z.; An, J.R.; Gou, Y.J.; Tan, M.; Zhao, Y. Exploring Liraglutide's mechanism in reducing renal fibrosis: the Fsp1-CoQ10-NAD(P)H pathway. Sci. Rep. 2025, 15, 1754. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Park, S.Y.; Lee, D.Y.; Kim, N.H.; Seo, J.A. GLP-1 receptor agonists in diabetic kidney disease: current evidence and future directions. Kidney. Res. Clin. Pract. 2022, 41, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Carraro-Lacroix, L.R.; Malnic, G.; Girardi, A.C. Regulation of Na+/H+ exchanger NHE3 by glucagon-like peptide 1 receptor agonist exendin-4 in renal proximal tubule cells. Am. J. Physiol. Renal. Physiol. 2009, 297, F1647–1655. [Google Scholar] [CrossRef]
- Kodama, G.; Taguchi, K.; Ito, S.; Nishino, Y.; Mori, K.; Taguchi, S.; Mitsuishi, Y.; Yamashita, Y.; Yamagishi, S.I.; Fukami, K. Glucagon like peptide-1 modulates urinary sodium excretion in diabetic kidney disease via ENaC activation. Sci. Rep. 2025, 15, 11486. [Google Scholar] [CrossRef]
- Anumas, S.; Inagi, R. Mitigating Lipotoxicity: A Potential Mechanism to Delay Chronic Kidney Disease Progression Using Current Pharmacological Therapies. Nephrology (Carlton) 2025, 30, e70098. [Google Scholar] [CrossRef]
- Yanai, H.; Adachi, H.; Hakoshima, M.; Iida, S.; Katsuyama, H. Metabolic-Dysfunction-Associated Steatotic Liver Disease-Its Pathophysiology, Association with Atherosclerosis and Cardiovascular Disease, and Treatments. Int. J. Mol. Sci. 2023, 24, 15473. [Google Scholar] [CrossRef]
- Park, M.J.; Han, H.J.; Kim, D.I. Lipotoxicity-Induced PRMT1 Exacerbates Mesangial Cell Apoptosis via Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2017, 18, 1421. [Google Scholar] [CrossRef]
- Kim, D.I.; Park, M.J.; Heo, Y.R.; Park, S.H. Metformin ameliorates lipotoxicity-induced mesangial cell apoptosis partly via upregulation of glucagon like peptide-1 receptor (GLP-1R). Arch. Biochem. Biophys. 2015, 584, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Adachi, H.; Hakoshima, M.; Katsuyama, H. Glucagon-Like Peptide 1 Receptor Agonists Versus Sodium-Glucose Cotransporter 2 Inhibitors for Atherosclerotic Cardiovascular Disease in Patients With Type 2 Diabetes. Cardiol. Res. 2023, 14, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.Y.; Oh, T.J.; Cho, Y.M. Glucagon-like peptide-1 increases mitochondrial biogenesis and function in INS-1 rat insulinoma cells. Endocrinol. Metab. 2015, 30, 216–220. [Google Scholar] [CrossRef]
- Lin, M.; Zhang, H.; Wu, H.; Zhao, D.; Nan, Z.; Fu, Y. Evaluation of febuxostat in treating diabetic kidney disease with hyperuricemia: a systematic review and meta-analysis of randomized controlled trials. Front. Med (Lausanne) 2025, 12, 1657274. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.J.; Sidor, N.A.; Tonial, N.C.; Che, A.; Urquhart, B.L. Uremic Toxins in the Progression of Chronic Kidney Disease and Cardiovascular Disease: Mechanisms and Therapeutic Targets. Toxins (Basel) 2021, 13, 142. [Google Scholar] [CrossRef]
- Takada, T.; Yamamoto, T.; Matsuo, H.; Tan, J.K.; Ooyama, K.; Sakiyama, M.; Miyata, H.; Yamanashi, Y.; Toyoda, Y.; Higashino, T.; et al. Identification of ABCG2 as an Exporter of Uremic Toxin Indoxyl Sulfate in Mice and as a Crucial Factor Influencing CKD Progression. Sci. Rep. 2018, 8, 11147. [Google Scholar] [CrossRef]
- Miyata, H.; Takada, T.; Toyoda, Y.; Matsuo, H.; Ichida, K.; Suzuki, H. Identification of Febuxostat as a New Strong ABCG2 Inhibitor: Potential Applications and Risks in Clinical Situations. Front. Pharmacol. 2016, 7, 518. [Google Scholar] [CrossRef]
- Taniguchi, T.; Omura, K.; Motoki, K.; Sakai, M.; Chikamatsu, N.; Ashizawa, N.; Takada, T.; Iwanaga, T. Hypouricemic agents reduce indoxyl sulfate excretion by inhibiting the renal transporters OAT1/3 and ABCG2. Sci. Rep. 2021, 11, 7232. [Google Scholar] [CrossRef]
- Taniguchi, T.; Ashizawa, N.; Matsumoto, K.; Saito, R.; Motoki, K.; Sakai, M.; Chikamatsu, N.; Hagihara, C.; Hashiba, M.; Iwanaga, T. Pharmacological Evaluation of Dotinurad, a Selective Urate Reabsorption Inhibitor. J. Pharmacol. Exp. Ther. 2019, 371, 162–170. [Google Scholar] [CrossRef]
- Yanai, H.; Katsuyama, H.; Hakoshima, M.; Adachi, H. Urate Transporter 1 Can Be a Therapeutic Target Molecule for Chronic Kidney Disease and Diabetic Kidney Disease: A Retrospective Longitudinal Study. Biomedicines 2023, 11, 567. [Google Scholar] [CrossRef]
- Kurihara, O.; Yamada, T.; Kato, K.; Miyauchi, Y. Efficacy of dotinurad in patients with severe renal dysfunction. Clin. Exp. Nephrol. 2024, 28, 208–216. [Google Scholar] [CrossRef]
- Takata, T.; Taniguchi, S.; Mae, Y.; Kageyama, K.; Fujino, Y.; Iyama, T.; Hikita, K.; Sugihara, T.; Isomoto, H. Comparative assessment of the effects of dotinurad and febuxostat on the renal function in chronic kidney disease patients with hyperuricemia. Sci. Rep. 2025, 15, 8990. [Google Scholar] [CrossRef]
- Osonoi, T.; Shirabe, S.; Saito, M.; Hosoya, M.; Watahiki, N.; Douguchi, S.; Ofuchi, K.; Katoh, M. Switching from febuxostat to dotinurad, a novel selective urate reabsorption inhibitor, may substantially reduce serum urate levels, through URAT1 inhibition, potentially without adversely affecting ABCG2 function: findings from the SWITCH SURI study. Front. Endocrinol (Lausanne) 2025, 16, 1655502. [Google Scholar] [CrossRef]
- Yanai, H.; Adachi, H.; Hakoshima, M.; Katsuyama, H. Glucose-Lowering Effects of Imeglimin and Its Possible Beneficial Effects on Diabetic Complications. Biology (Basel) 2023, 12, 726. [Google Scholar] [CrossRef] [PubMed]
- Drewe, J.; Foretz, M.; Krähenbühl, S. Metformin-mechanisms of its glycemia-reducing effect. Pharmacol. Rev. 2025, 78, 100106. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Lin, S.H.; Hung, S.Y.; Chiou, Y.Y.; Hsu, W.C.; Chang, C.M.; Liou, H.H.; Chang, M.Y.; Ho, L.C.; Wu, C.F.; et al. Renal Protective Effect of Metformin in Type 2 Diabetes Patients. J. Clin. Endocrinol. Metab. 2025, 110, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Lin, S.H.; Wang, H.H.; Hsu, W.C.; Hung, S.Y.; Chiou, Y.Y.; Liou, H.H.; Chang, M.Y.; Ho, L.C.; Wu, C.F.; et al. Role of Metformin in Preventing New-Onset Chronic Kidney Disease in Patients with Type 2 Diabetes Mellitus. Pharmaceuticals (Basel). 2025, 18, 95. [Google Scholar] [CrossRef]
- Pan, Q.; Lu, X.; Zhao, C.; Liao, S.; Chen, X.; Guo, F.; Yang, C.; Liu, H.F. Metformin: the updated protective property in kidney disease. Aging (Albany NY) 2020, 12, 8742–8759. [Google Scholar] [CrossRef]
- Yi, H.; Huang, C.; Shi, Y.; Cao, Q.; Zhao, Y.; Zhang, L.; Chen, J.; Pollock, C.A.; Chen, X.M. Metformin attenuates folic-acid induced renal fibrosis in mice. J. Cell. Physiol. 2018, 233, 7045–7054. [Google Scholar] [CrossRef]
- Hallakou-Bozec, S.; Kergoat, M.; Fouqueray, P.; Bolze, S.; Moller, D.E. Imeglimin amplifies glucose-stimulated insulin release from diabetic islets via a distinct mechanism of action. PLoS. ONE 2021, 16, e0241651. [Google Scholar] [CrossRef]
- DeFronzo, R.; Fleming, G.A.; Chen, K.; Bicsak, T.A. Metformin-associated lactic acidosis: Current perspectives on causes and risk. Metabolism 2016, 65, 20–29. [Google Scholar] [CrossRef]
- Theurey, P.; Vial, G.; Fontaine, E.; Monternier, P.A.; Fouqueray, P.; Bolze, S.; Moller, D.E.; Hallakou-Bozec, S. Reduced lactic acidosis risk with Imeglimin: Comparison with Metformin. Physiol. Rep. 2022, 10, e15151. [Google Scholar] [CrossRef]
- Madiraju, A.K.; Erion, D.M.; Rahimi, Y.; Zhang, X.M.; Braddock, D.T.; Albright, R.A.; Prigaro, B.J. Metformin suppresses gluconeogenesis by inhibiting mitochondrial glycerophosphate dehydrogenase. Nature 2014, 510, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of hepatic glucose metabolism in health and disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef] [PubMed]
- Lachaux, M.; Soulié, M.; Hamzaoui, M.; Bailly, A.; Nicol, L.; Rémy-Jouet, I.; Renet, S.; Vendeville, C.; Gluais-Dagorn, P.; Hallakou-Bozec, S.; et al. Short-and long-term administration of imeglimin counters cardiorenal dysfunction in a rat model of metabolic syndrome. Endocrinol. Diabetes. Metab. 2020, 3, e00128. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, S.; Matsumoto, K.; Tanno, Y.; Yokoo, T. Imeglimin in Patients with an Estimated Glomerular Filtration Rate Less than 45 mL/min per 1.73 m(2): A Case Series. JMA. J. 2025, 8, 1301–1306. [Google Scholar]
- Uto, A.; Ishinoda, Y.; Asaga, T.; Tanahashi, Y.; Kobayashi, A.; Meshino, H.; Okazaki, M.; Tomita, K.; Kasuga, A.; Oshima, N. Imeglimin for Type 2 Diabetes Mellitus: Its Efficacy and Insight Into the Potential Benefit for Renal and Liver Function. Cureus. 2024, 16, e66322. [Google Scholar] [CrossRef]
- Hozumi, K.; Sugawara, K.; Ishihara, T.; Ishihara, N.; Ogawa, W. Effects of imeglimin on mitochondrial function, AMPK activity, and gene expression in hepatocytes. Sci. Rep. 2023, 13, 746. [Google Scholar] [CrossRef]
- Daza-Arnedo, R.; Rico-Fontalvo, J.; Aroca-Martínez, G.; Rodríguez-Yanez, T.; Martínez-Ávila, M.C.; Almanza-Hurtado, A.; Cardona-Blanco, M.; Henao-Velásquez, C.; Fernández-Franco, J.; Unigarro-Palacios, M.; et al. Insulin and the kidneys: a contemporary view on the molecular basis. Front. Nephrol. 2023, 3, 1133352. [Google Scholar] [CrossRef]
- Migdalis, I.N.; Ioannidis, I.M.; Papanas, N.; Raptis, A.E.; Sotiropoulos, A.E.; Dimitriadis, G.D. Hypertriglyceridemia and Other Risk Factors of Chronic Kidney Disease in Type 2 Diabetes: A Hospital-Based Clinic Population in Greece. J. Clin. Med. 2022, 11, 3224. [Google Scholar] [CrossRef]
- Li, D.; Liu, Z.; Jiang, H. Hypertriglyceridemia in chronic kidney disease: pathophysiological mechanisms, cardiovascular risk, and emerging therapeutics. Lipids. Health. Dis. Online ahead of print. 20 Jan 2026. [CrossRef]
- Russo, G.T.; De Cosmo, S.; Viazzi, F.; Pacilli, A.; Ceriello, A.; Genovese, S.; Guida, P.; Giorda, C.; Cucinotta, D.; Pontremoli, R.; et al. Plasma Triglycerides and HDL-C Levels Predict the Development of Diabetic Kidney Disease in Subjects With Type 2 Diabetes: The AMD Annals Initiative. Diabetes. Care 2016, 39, 2278–2287. [Google Scholar] [CrossRef] [PubMed]
- Tu, Q.M.; Jin, H.M.; Yang, X.H. Lipid abnormality in diabetic kidney disease and potential treatment advancements. Front. Endocrinol (Lausanne) 2025, 16, 1503711. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Adachi, H.; Hakoshima, M.; Katsuyama, H. Molecular Biological and Clinical Understanding of the Statin Residual Cardiovascular Disease Risk and Peroxisome Proliferator-Activated Receptor Alpha Agonists and Ezetimibe for Its Treatment. Int. J. Mol. Sci. 2022, 23, 3418. [Google Scholar] [CrossRef]
- Al-Rasheed, N.M.; Al-Rasheed, N.M.; Attia, H.A.; Al-Amin, M.A.; Al-Ajmi, H.N.; Hasan, I.H.; Mohamad, R.A.; Sinjilawi, N.A. Renoprotective Effects of Fenofibrate via Modulation of LKB1/AMPK mRNA Expression and Endothelial Dysfunction in a Rat Model of Diabetic Nephropathy. Pharmacology 2015, 95, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Keech, A.; Simes, R.J.; Barter, P.; Best, J.; Scott, R.; Taskinen, M.R.; Forder, P.; Pillai, A.; Davis, T.; Glasziou, P.; et al. Effects of long-term fenofibrate therapy on cardiovascular events in 9795 people with type 2 diabetes mellitus (the FIELD study): randomised controlled trial. Lancet 2005, 366(9500), 1849–61. [Google Scholar] [CrossRef]
- Wu, J.; Song, Y.; Li, H.; Chen, J. Rhabdomyolysis associated with fibrate therapy: review of 76 published cases and a new case report. Eur. J. Clin. Pharmacol. 2009, 65, 1169–1174. [Google Scholar] [CrossRef]
- Fruchart, J.C.; Santos, R.D.; Aguilar-Salinas, C.; Aikawa, M.; Al Rasadi, K.; Amarenco, P.; Barter, P.J.; Ceska, R.; Corsini, A.; Després, J.P.; et al. The selective peroxisome proliferator-activated receptor alpha modulator (SPPARMα) paradigm: conceptual framework and therapeutic potential: A consensus statement from the International Atherosclerosis Society (IAS) and the Residual Risk Reduction Initiative (R3i) Foundation. Cardiovasc. Diabetol. 2019, 18, 71. [Google Scholar]
- Aomura, D.; Harada, M.; Yamada, Y.; Nakajima, T.; Hashimoto, K.; Tanaka, N.; Kamijo, Y. Pemafibrate Protects against Fatty Acid-Induced Nephropathy by Maintaining Renal Fatty Acid Metabolism. Metabolites 2021, 11, 372. [Google Scholar] [CrossRef]
- Horinouchi, Y.; Murashima, Y.; Yamada, Y.; Yoshioka, S.; Fukushima, K.; Kure, T.; Sasaki, N.; Imanishi, M.; Fujino, H.; Tsuchiya, K.; et al. Pemafibrate inhibited renal dysfunction and fibrosis in a mouse model of adenine-induced chronic kidney disease. Life. Sci. 2023, 321, 121590. [Google Scholar] [CrossRef]
- Uetake, D.; Ohno, I.; Ichida, K.; Yamaguchi, Y.; Saikawa, H.; Endou, H.; Hosoya, T. Effect of fenofibrate on uric acid metabolism and urate transporter 1. Intern. Med. 2010, 49, 89–94. [Google Scholar] [CrossRef]
- Yanai, H.; Katsuyama, H.; Hakoshima, M.A. Significant Increase of Estimated Glomerular Filtration Rate After Switching From Fenofibrate to Pemafibrate in Type 2 Diabetic Patients. Cardiol. Res. 2021, 12, 358–362. [Google Scholar] [CrossRef]
- Izumihara, R.; Nomoto, H.; Kito, K.; Yamauchi, Y.; Omori, K.; Shibayama, Y.; Yanagiya, S.; Miya, A.; Kameda, H.; Cho, K.Y.; et al. Switching from Conventional Fibrates to Pemafibrate Has Beneficial Effects on the Renal Function of Diabetic Subjects with Chronic Kidney Disease. Diabetes. Metab. J. 2024, 48, 473–481. [Google Scholar] [CrossRef]
Figure 1.
The histological and molecular characteristics of DKD. Blue arrows indicate effects, black horizontal arrows indicate the flow of substance, and red upward and downward arrows indicate an increase and a decrease of activity or substances, respectively. AT1R, angiotensin II type 1 receptor; ECM, extracellular matrix; ENaC, epithelial sodium channel; Epo, erythropoietin; G, glucose; GBM, glomerular basement membrane; MR, mineralocorticoid receptor; NHE3, Na+/H+ exchanger 3; ROS, reactive oxygen species; SGLT2, sodium-glucose cotransporter 2; TGF, tubuloglomerular feedback.
Figure 1.
The histological and molecular characteristics of DKD. Blue arrows indicate effects, black horizontal arrows indicate the flow of substance, and red upward and downward arrows indicate an increase and a decrease of activity or substances, respectively. AT1R, angiotensin II type 1 receptor; ECM, extracellular matrix; ENaC, epithelial sodium channel; Epo, erythropoietin; G, glucose; GBM, glomerular basement membrane; MR, mineralocorticoid receptor; NHE3, Na+/H+ exchanger 3; ROS, reactive oxygen species; SGLT2, sodium-glucose cotransporter 2; TGF, tubuloglomerular feedback.
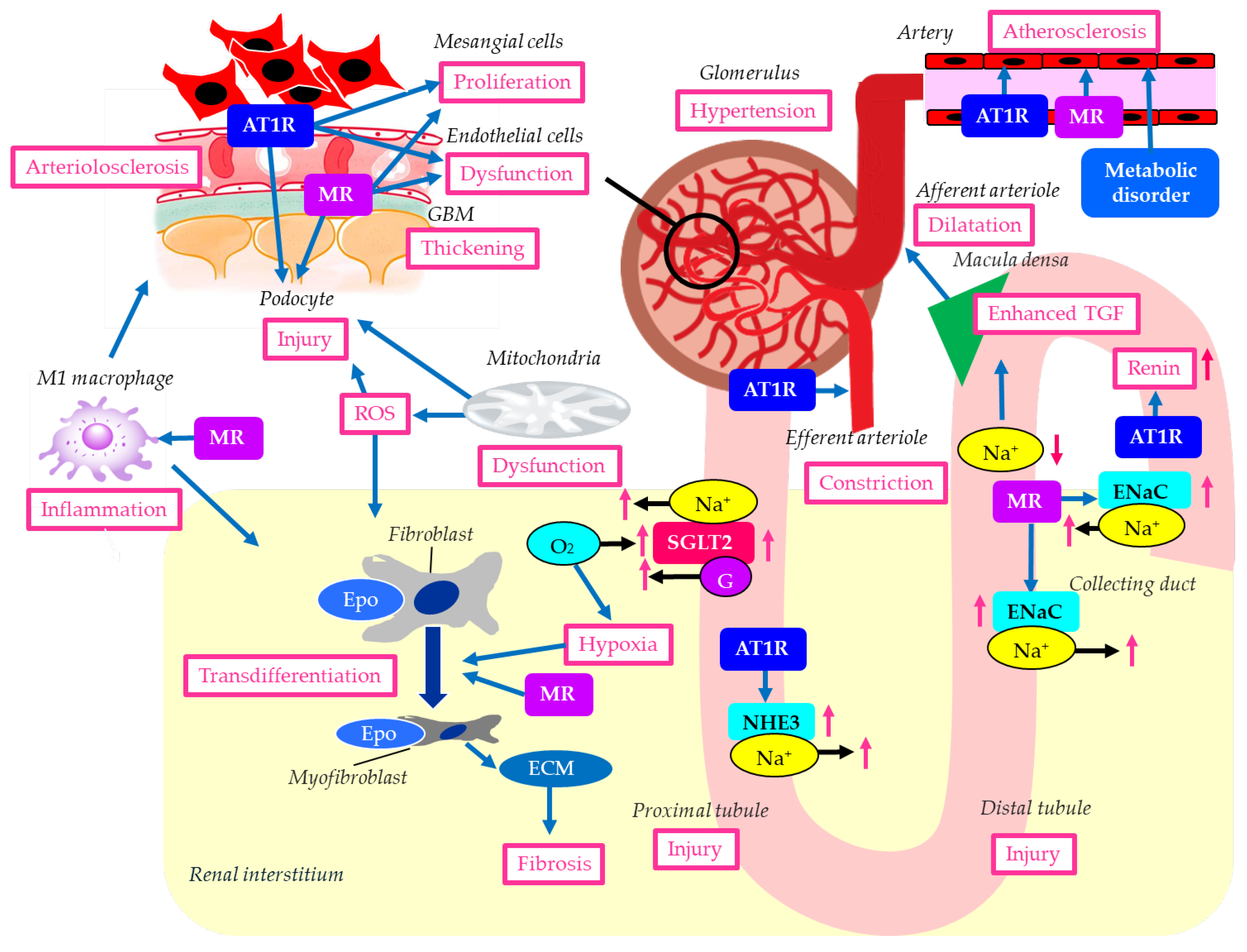
Figure 2.
The mechanisms of possible renal protective effects of SGLT2is. Blue arrows indicate effects, black horizontal arrows indicate the flow of substance, and red upward and downward arrows indicate an increase and a decrease of activity or substances, respectively. ATP-binding cassette, subfamily G, 2 (ABCG2); ECM, extracellular matrix; Epo, erythropoietin; ER, endoplasmic reticulum; G, glucose; GLUT9, glucose transporter 9; ROS, reactive oxygen species; SGLT2, sodium-glucose cotransporter 2; SGLT2is, sodium-glucose cotransporter 2 inhibitors; TGF, tubuloglomerular feedback; UA, uric acid; URAT1, urate transporter 1.
Figure 2.
The mechanisms of possible renal protective effects of SGLT2is. Blue arrows indicate effects, black horizontal arrows indicate the flow of substance, and red upward and downward arrows indicate an increase and a decrease of activity or substances, respectively. ATP-binding cassette, subfamily G, 2 (ABCG2); ECM, extracellular matrix; Epo, erythropoietin; ER, endoplasmic reticulum; G, glucose; GLUT9, glucose transporter 9; ROS, reactive oxygen species; SGLT2, sodium-glucose cotransporter 2; SGLT2is, sodium-glucose cotransporter 2 inhibitors; TGF, tubuloglomerular feedback; UA, uric acid; URAT1, urate transporter 1.
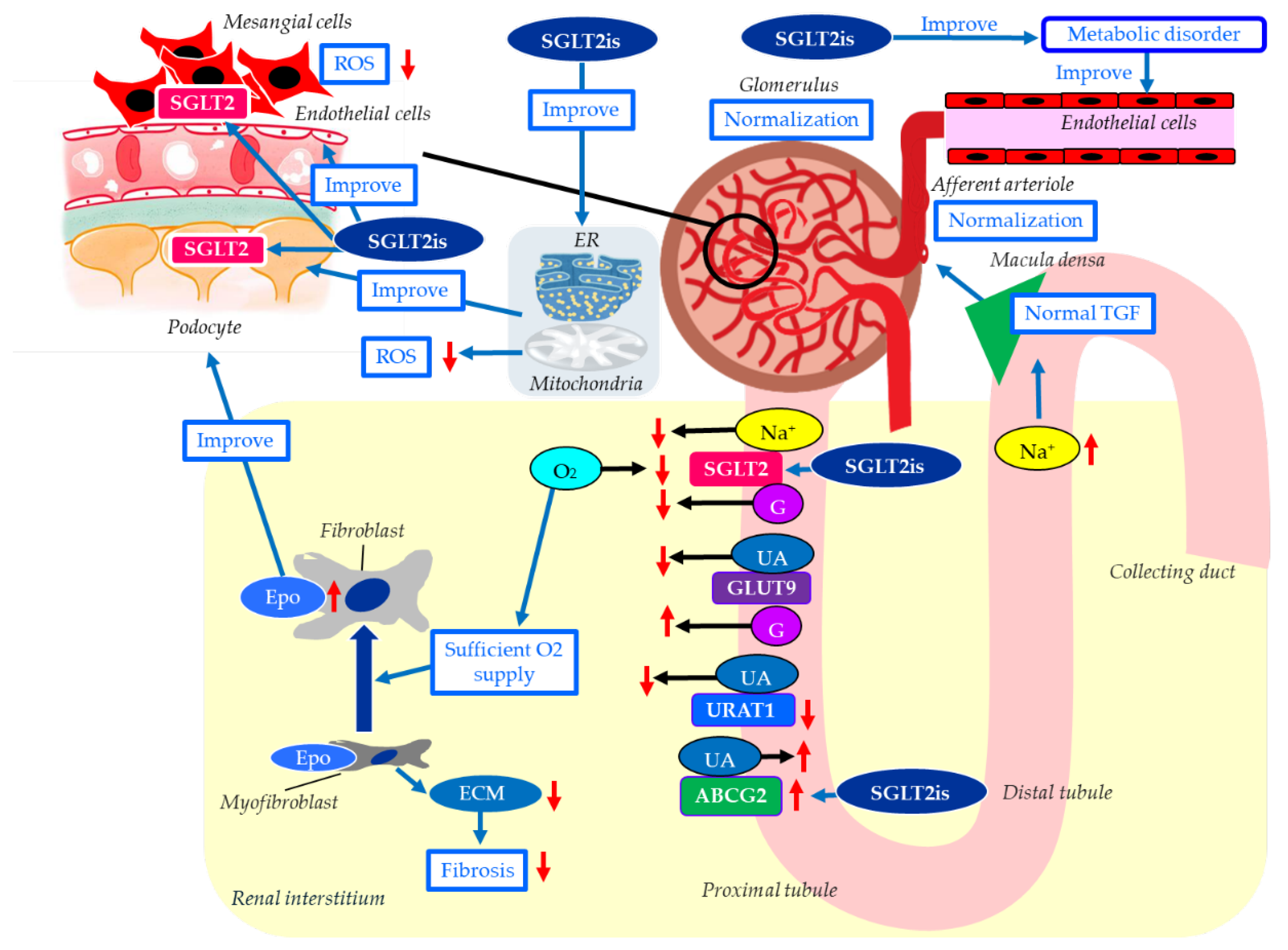
Figure 3.
The mechanisms of possible renal protective effects of GLP-1RAs. Blue arrows indicate effects, black horizontal arrows indicate the flow of substance, and red upward and downward arrows indicate an increase and a decrease of activity and substances, respectively. ENaC, epithelial sodium channel; GLP-1R, glucagon-like peptide-1 receptor; GLP-1RAs; glucagon-like peptide-1 receptor agonists; NHE3, Na+/H+ exchanger 3; PI3K-Akt, phosphatidylinositol 3'-kinase-Akt; ROS, reactive oxygen species; TGF, tubuloglomerular feedback.
Figure 3.
The mechanisms of possible renal protective effects of GLP-1RAs. Blue arrows indicate effects, black horizontal arrows indicate the flow of substance, and red upward and downward arrows indicate an increase and a decrease of activity and substances, respectively. ENaC, epithelial sodium channel; GLP-1R, glucagon-like peptide-1 receptor; GLP-1RAs; glucagon-like peptide-1 receptor agonists; NHE3, Na+/H+ exchanger 3; PI3K-Akt, phosphatidylinositol 3'-kinase-Akt; ROS, reactive oxygen species; TGF, tubuloglomerular feedback.
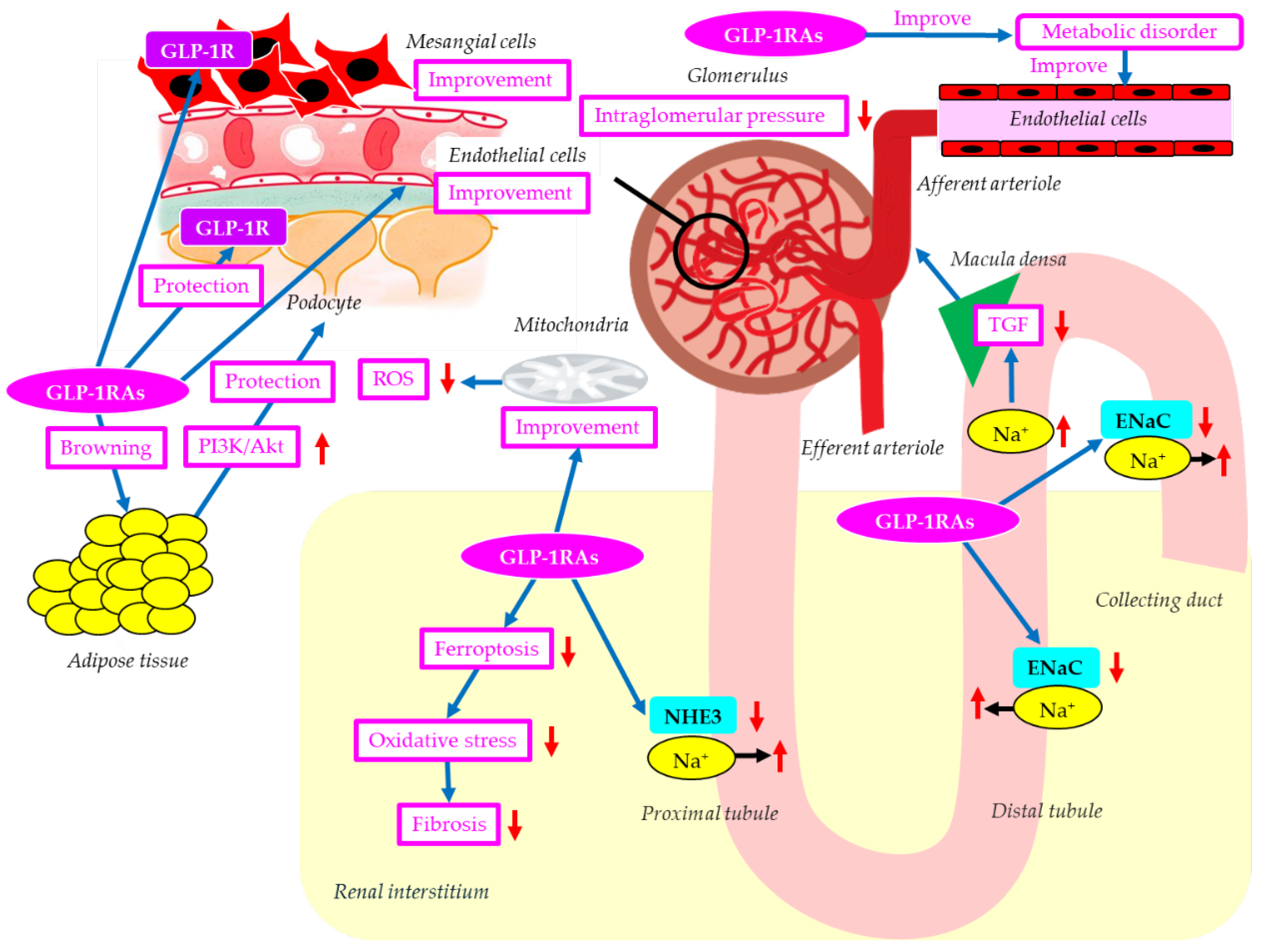
Figure 7.
DKD patients had better take four dishes with DIP. GLP-1RAs, glucagon-like peptide-1 receptor agonists; nsMRAs, non-steroidal mineralocorticoid receptor antagonists; RASis, renin-angiotensin system inhibitors; SGLT2is, sodium-glucose cotransporter 2 inhibitors.
Figure 7.
DKD patients had better take four dishes with DIP. GLP-1RAs, glucagon-like peptide-1 receptor agonists; nsMRAs, non-steroidal mineralocorticoid receptor antagonists; RASis, renin-angiotensin system inhibitors; SGLT2is, sodium-glucose cotransporter 2 inhibitors.
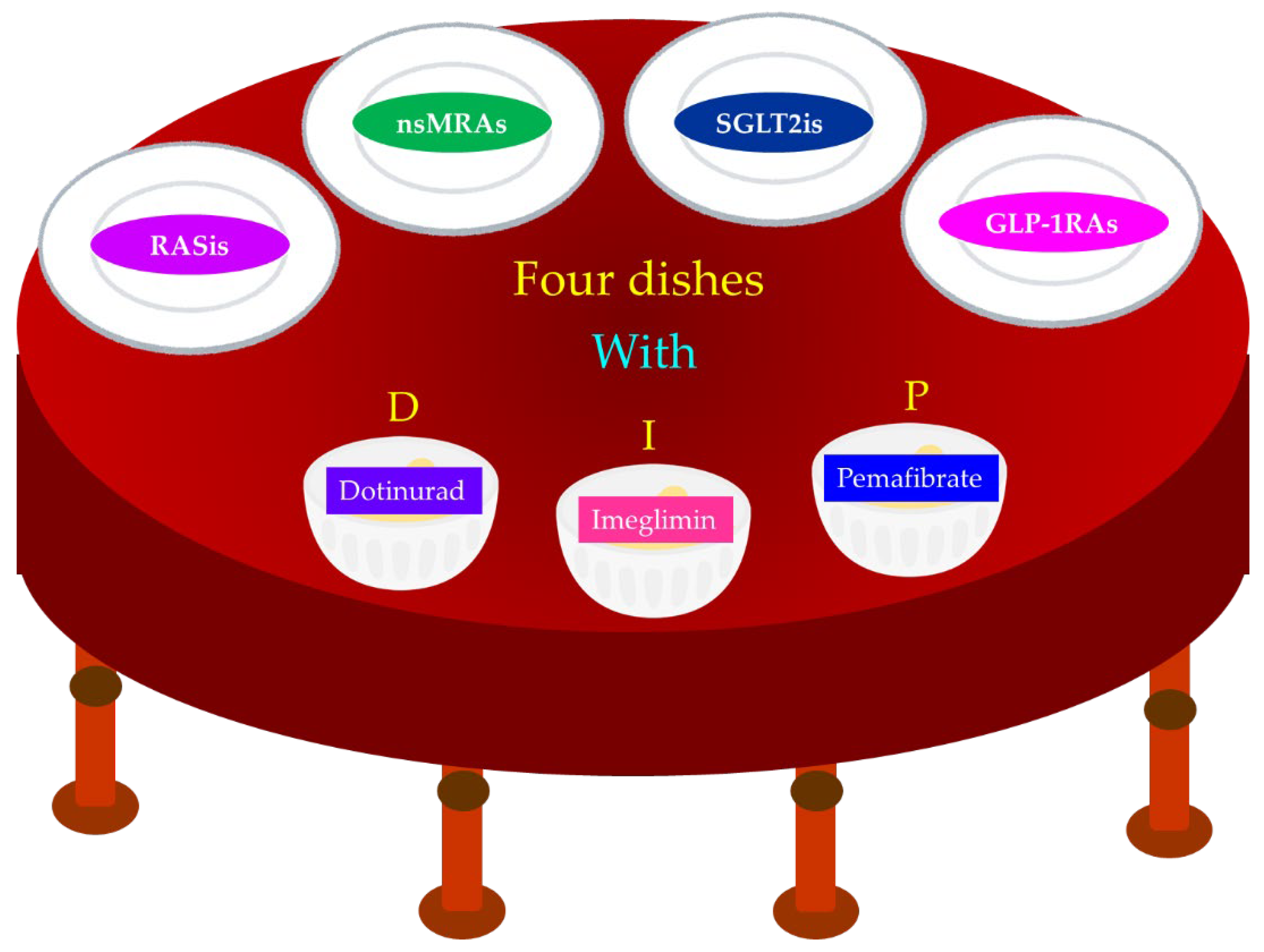
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



