Submitted:
14 February 2026
Posted:
26 February 2026
You are already at the latest version
Abstract
Mycotoxin contamination of cattle feed commonly results in chronic, subclinical exposure with adverse effects on liver function and oxidative status. This controlled field trial evaluated a multi-component mycotoxin detoxifier in fattening mixed Limousin calves fed naturally contaminated diets under commercial production conditions. Twenty-four clinically healthy calves were randomly allocated to a control group or to a treatment group receiving a detoxifying additive containing adsorbent clays, phytogenic extracts (silymarin and curcumin) and yeast-derived components for 60 days. Feed, blood, fecal and liver samples were analyzed to assess biochemical, oxidative and histopathological re-sponses throughout the experimental period. The diet contained aflatoxin B1, fumonisins and zearalenone at concentrations exceeding guidance limits, indicating chronic mul-ti-mycotoxin exposure. No clinical signs of overt mycotoxicosis were observed. All bio-chemical parameters remained within physiological ranges; however, γ-glutamyl trans-ferase activity and protein carbonyl levels were significantly lower in treated calves com-pared with controls (p < 0.05). Other oxidative stress markers did not differ between groups, and histopathology revealed only mild, non-specific liver changes. In conclusion, supplementation with the tested mycotoxin detoxifier was safe and associated with im-proved hepatic enzyme regulation and reduced protein oxidative damage under field conditions.
Keywords:
mycotoxins
; fattening calves
; hepatic function
; oxidative stress
; γ-glutamyl transferase
; phytogenic feed additive
; mycotoxin detoxifier
1. Introduction
Mycotoxins are toxic secondary metabolites produced by filamentous fungi, mainly belonging to the genera Aspergillus, Fusarium, Penicillium, or Alternaria, and represent a major challenge for animal feed safety worldwide [1,2,3]. Their remarkable stability is primarily due to their chemical structure, while their prevalence is further increased by the inadequate implementation of good agricultural practices (GAPs) and, more recently, by climate change [4,5]. Taken together, these factors result in the extensive presence of mycotoxins in raw materials and finished animal feeds worldwide. Studies reported that at a global level, between 30 and 100% of food and feed samples are co-contaminated with mycotoxins [4,6,7]. In cattle production systems, contamination of cereal-based feeds with multiple mycotoxins is common, particularly under intensive feeding conditions and variable storage environments. In contrast to acute intoxications, chronic exposure to low or moderate concentrations of mycotoxins is far more prevalent and is increasingly recognized as a cause of subclinical metabolic disturbances rather than overt clinical disease [3,4].
Although ruminants possess a partial capacity to detoxify certain mycotoxins through microbial metabolism in the rumen, this capacity varies markedly among mycotoxin classes and may be insufficient during continuous dietary exposure [8,9]. Aflatoxin B1 (AFB1), fumonisins (FUMs) and zearalenone (ZEN) are among the most frequently detected mycotoxins in cattle feeds and have been associated with impaired liver function, oxidative stress, immune modulation and reduced productive efficiency, even at concentrations close to guidance or regulatory limits [4,10]. Importantly, such effects often develop in the absence of clear clinical signs, making subclinical mycotoxicosis difficult to detect under commercial conditions [3].
The liver plays a central role in mycotoxin biotransformation and is therefore a primary target organ during chronic exposure. Hepatic stress induced by mycotoxins has been linked to alterations in hepatobiliary enzymes, disruption of redox homeostasis and oxidative modification of cellular macromolecules [11]. Among routinely measured biochemical parameters, γ-glutamyl transferase has been proposed as a sensitive indicator of early hepatobiliary stress in cattle, particularly under subclinical exposure conditions where transaminase activities may remain within physiological ranges [3,12]. In parallel, oxidative stress biomarkers such as protein carbonyls and lipid peroxidation products provide valuable insight into the cellular impact of mycotoxins beyond classical clinical pathology [13,14].
Strategies to mitigate the adverse effects of mycotoxins in ruminant nutrition rely primarily on the inclusion of detoxifying feed additives. These products generally combine adsorbent materials, such as bentonite or sepiolite, with biologically active components including yeast derivatives and phytogenic compounds [15,16]. Among phytogenic extracts, curcumin and silymarin have attracted particular interest due to their hepatoprotective, antioxidant and anti-inflammatory properties [17,18,19,20,21]. In addition, bioactive constituents of silymarin have been reported to exhibit antibacterial activity [22,23]. Experimental studies have demonstrated that these compounds can stabilize hepatocyte membranes, enhance antioxidant defence systems and attenuate oxidative damage induced by mycotoxins [24,25,26]. More recently, multi-component formulations integrating adsorbents with phytogenic and yeast-derived constituents have been proposed as a comprehensive approach to mitigate the multifactorial effects of mycotoxin exposure [17,18,19].
Despite growing evidence supporting the efficacy of such detoxifiers in controlled experimental settings and in non-ruminant species, field data in fattening cattle remain limited. In particular, there is a lack of studies evaluating hepatic and oxidative responses under natural multi-mycotoxin exposure conditions representative of commercial feedlot systems [8,27]. The present study was therefore conducted as a controlled field trial to evaluate the effects of a multi-component mycotoxin detoxifier on hepatic function, oxidative stress status and general health indicators in fattening mixed Limousin calves exposed to naturally contaminated feed.
2. Materials and Methods
2.1. Ethical Approval and Compliance
The study was conducted in accordance with the Code of Practice for the Conduct of Clinical Trials for Veterinary Medicinal Products (European Medicines Agency, 2014) and the Guide for the Care and Use of Agricultural Animals in Research and Teaching (FASS, 2020). The experimental protocol was reviewed and approved by the Research Ethics & Conduct Committee, University of Thessaly, Greece (48/4-6-2024).
2.2. Study Design and Experimental Units
A controlled field trial was performed under commercial feedlot conditions. The experimental unit was the individual animal. Calves were allocated to two parallel groups, a control group and a treatment group receiving a multi-component mycotoxin detoxifier. The study duration was 60 consecutive days.
2.3. Animals, Housing, and Husbandry
The study included 24 clinically healthy mixed Limousin fattening calves, approximately six months of age, originating from the same commercial feedlot in Thessaly, Central Greece. All animals had a similar genetic background, management history, and nutritional status prior to enrolment. Inclusion criteria were absence of clinical disease during the two months preceding the study, normal appetite and rectal temperature, body weight within ±10% of the group mean, completed vaccination against clostridial diseases at 30 days of age, and antiparasitic treatment at approximately five months of age. Animals not meeting these criteria were excluded a priori.
Calves were housed in open group pens with concrete flooring and straw bedding, under natural ventilation and ambient environmental conditions typical of the region. Animals from both experimental groups were housed in the same open facility, physically separated by a barrier, and managed under identical environmental and husbandry conditions. Although animals were spatially separated by treatment, they shared the same housing environment, reflecting the inherent clustering typical of commercial feedlot studies.
Animals had ad libitum access to fresh drinking water and were fed a Total Mixed Ration (TMR) twice daily. No environmental enrichment beyond standard commercial practice was applied.
2.4. Sample Size Calculation
Sample size justification was based on published data in cows demonstrating biologically relevant changes in serum γ-glutamyl transferase activity following supplementation with a mycotoxin mitigation feed additive under field conditions [27,28]. Based on the between-group differences and variability reported in those studies, a sample size of approximately 10–12 animals per group was estimated to provide 80% power to detect meaningful changes in hepatic enzyme activity at a two-sided significance level of 0.05. Accordingly, the inclusion of 12 calves per group was considered adequate for the objectives of the present study, which focused on indicators of liver function and oxidative status under conditions of chronic, subclinical dietary mycotoxin exposure. The a priori sample size calculation was therefore intended to ensure adequate power to detect biologically relevant differences in selected hepatic and oxidative stress biomarkers under field conditions, rather than to provide confirmatory efficacy estimates or performance-related outcomes under commercial feeding conditions.
2.5. Randomization and Allocation Concealment
Animals were allocated to experimental groups using stratified randomization based on baseline body weight. Briefly, calves were first ranked according to body weight and assigned to weight strata to ensure comparable baseline distributions between groups. Within each stratum, animals were then allocated to control or treatment groups using simple random allocation.
Due to the nature of the dietary intervention and the commercial farm setting, caretakers were aware of group allocation. Clinical observations were performed on a weekly basis following predefined and standardized assessment criteria. Laboratory analyses of biochemical and oxidative stress parameters, as well as histopathological evaluations, were conducted by personnel blinded to treatment allocation.
2.6. Dietary Treatments
The control group (T1) received the basal Total Mixed Ration (TMR) without supplementation. The treatment group (T2) received the same TMR supplemented with 1.5 kg a commercial multi-component mycotoxin detoxifier per ton of TMR (BIŌNTE® QUIMITŌX® PLUS, BIŌNTE Nutrition S.L., Spain). The additive consisted of adsorbent clays (bentonite and sepiolite), phytogenic extracts of Silybum marianum (silymarin) and Curcuma longa (curcumin), and yeast-derived components. Supplementation was administered daily for 60 consecutive days prior to slaughter. A 7-day dietary adaptation period preceded the experimental phase, during which calves in the treatment group were exposed to the mycotoxin detoxifier to allow stabilization of feed intake and ruminal adaptation. No samples were collected during this period.
The TMR formulation was identical for both groups and prepared from the same batch of raw materials throughout the study to minimize variability in nutrient and mycotoxin content. Diets were formulated to meet the nutritional requirements of fattening calves (Table 1).
2.7. Clinical Monitoring and Animal Welfare
Animals were monitored throughout the experimental period by trained veterinarians. Weekly clinical examinations assessed general health status, behaviour, feed intake, fecal consistency, locomotion, and respiratory signs. Any adverse events or deviations from normal health status were recorded. Humane endpoints were predefined; however, none were reached, and no animals were withdrawn from the study.
Individual feed intake was not recorded during the trial due to the commercial group-feeding conditions. Therefore, feed intake was not analyzed as an outcome measure in the present study.
2.8. Sampling Procedures
2.8.1. Feed and Fecal Sampling and Mycotoxin Analysis
Composite TMR samples were collected every 10 days during the experimental period. Fecal samples were collected from all animals at the same intervals.
Feed samples were analyzed for aflatoxin B1 (AFB1), zearalenone (ZEN), deoxynivalenol (DON), fumonisin B1 (FUMB1), ochratoxin A (OTA), T-2 toxin (T-2), and HT-2 toxin (HT-2), using lateral flow strip kits supplied by ProGnosis Biotech (Larissa, Greece). Briefly, previously homogenized feed samples were ground until a fine powder was obtained. A total of 5 g of the obtained powder was weighed into a 50 mL falcon tube and 15 mL of the extraction solution (included in the kit) was added. The mixture was vortex shaken for 2 min. An aliquot (1 mL) of the supernatant was transferred to a 1.5 mL Eppendorf tube and centrifuged at 12,500 rpm for 2 min. A total of 100 µL of the supernatant was added to a dilution tube (included in the kit) and further mixed. Finally, 100 µL of the diluted extract was added to a microwell containing the lyophilized gold particles and homogenized using the micropipette until completely diluted. Once completely diluted, the strip was introduced. Then, 3 min after the introduction of the strip, it was retired from the microwell, and the strip’s cotton pad was peeled before its quantification using a 3PR-Mini instrument from ProGnosis. The quantification was performed using the S-Flow software (2.03.111) from the same company.
Regarding the methodology for the feces analysis, feces (1g) were extracted with 20 mL of a methanol/water (80:20) mixture using an ultraturrax for 15 min. After centrifugation, the supernatant was filtered and placed in an injection vial prior to its UHPLC-ESI-MS/TOF analysis. The chromatographic separation was performed with a Gemini C18 (50 x 2mm x 3 μm) column from Phenomenex. Detection and identification were performed using a quadrupole/time of flight mass spectrometer with a targeted approach.
For descriptive and statistical purposes, values below the LOD were not imputed and were treated as non-quantifiable measurements. It should be noted that feed and fecal mycotoxin levels are not directly comparable in quantitative terms, as fecal concentrations reflect combined effects of intake, ruminal metabolism, gastrointestinal binding, and excretion dynamics [9].
2.8.2. Blood Sampling and Biochemical Analyses
Blood samples were collected at the end of the 60-day feeding period via coccygeal vein puncture into EDTA-containing tubes. Plasma was separated by centrifugation and stored at −80°C until analysis.
Biochemical parameters including urea, creatinine, AST, ALT, γ-glutamyl transferase, total proteins, and albumin were measured spectrophotometrically using commercial diagnostic kits.
Oxidative stress markers included thiobarbituric-acid reactive substances (TBARS, expressed as malondialdehyde (MDA) equivalents and reflecting lipid peroxidation), protein carbonyls (CARBs, as an index of protein oxidation) and total antioxidant capacity (TAC, expressed as Trolox equivalents). Plasma levels of TBARS, CARBs, and TAC were assessed using commercial assay kits (Cayman Chemical, Ann Arbor, MI, USA) in accordance with the manufacturer’s protocols.
Baseline measurements were not obtained prior to the intervention; therefore, the reported results represent between-group differences at the study endpoint rather than within-animal changes over time.
2.8.3. Histopathology
Representative tissue samples were collected from the liver of all experimental animals at slaughter. All tissues were immediately fixed in 10% neutral buffered formalin for histological tissue processing. After fixation, the specimens were processed through graded alcohols and xylene and embedded in paraffin wax. Paraffin blocks were sectioned at 4–5 microns and slides were stained routinely with hematoxylin and eosin (H&E) for histopathological examination. All slides were examined under light microscopy by two pathologists blinded to groups.
2.9. Statistical Analysis
Statistical analyses were performed using IBM SPSS Statistics version 29.0. Data were assessed for normality using the Shapiro–Wilk test and visual inspection of distribution plots. Homogeneity of variances was evaluated with Levene’s test. Normally distributed variables were analyzed using independent-samples t-tests, while non-normally distributed variables were analyzed using the Mann–Whitney U test. Results are presented as mean ± standard deviation. Statistical significance was set at p < 0.05.
3. Results
3.1. Clinical Observations and General Health Status
All calves completed the 60-day experimental period and were included in the final analysis. No mortality or severe adverse events were recorded in either group. Animals in both the control (T1) and treatment (T2) groups remained in good general health throughout the study. Feed consumption behaviour at group level was normal, and no episodes of anorexia or depression were observed. Sporadic, mild clinical deviations such as transient diarrhea or mild lameness occurred in both groups with similar frequency and resolved spontaneously without therapeutic intervention. No clinical signs consistent with overt mycotoxicosis, ruminal acidosis, respiratory disease, or systemic illness were detected.
3.2. Mycotoxin Contamination of the Total Mixed Ration
Analysis of composite TMR samples collected at regular 10-day intervals throughout the experimental period revealed the presence of multiple mycotoxins (Table 2). AFB1, FUMs, and ZEN were consistently detected at concentrations exceeding selected technical or guidance values for cattle feed, whereas deoxynivalenol (DON), T-2 toxin (T-2), HT-2 toxin (HT-2) and ochratoxin A (OTA) remained below recommended thresholds. The values reported in Table 2 represent mean concentrations across all sampling points, reflecting sustained dietary exposure rather than isolated contamination events. The overall contamination pattern was comparable across sampling time points, supporting the presence of chronic multi-mycotoxin exposure under commercial feedlot conditions, without evidence of overt clinical mycotoxicosis.
For clarity, regulatory maximum levels (“EU limits”) refer to legally binding limits established by European Union legislation for complete feeds, whereas “guidance” or “technical” values correspond to non-binding reference thresholds proposed by EFSA or industry bodies to support risk management and the prevention of subclinical adverse effects [29,30,31,32]. Exceedance of guidance or technical values does not imply non-compliance with EU legislation but indicates increased toxicological relevance, particularly under conditions of chronic exposure.
Although FUM concentrations remained below the EU regulatory maximum level for complete feeds, they exceeded suggested guidance values, which are intended to indicate increased risk for performance impairment or subclinical effects rather than acute toxicosis.
3.3. Biochemical Parameters
Mean values and standard deviations of biochemical parameters measured at the end of the experimental period are presented in Table 3. No statistically significant differences were observed between groups for urea, creatinine, aspartate aminotransferase, alanine aminotransferase, total proteins, or albumin. All values were within physiological reference ranges for fattening calves.
γ-GT activity was significantly lower in the treatment group compared with the control group (p < 0.05). No other hepatic or renal biochemical markers differed significantly between groups.
3.3. Oxidative Stress Biomarkers
Oxidative stress indices are summarized in Table 4. CARBs levels were significantly lower in the treatment group compared with the control group (P < 0.05). No statistically significant differences were detected between groups for TBARS or TAC.
3.5. Fecal Mycotoxin Concentrations
Fecal mycotoxin analysis results are shown in Table 5. ZEN was detected in fecal samples from the control group (23.0±3.0), whereas it was not detected in samples from the treatment group. AFB1 was detected in the treatment group (19.0±3.0) but not in the control group. Deoxynivalenol was detected in the TMR at approximately 4,077 ppb. In the control group, low concentrations of DON were detected in fecal samples (90.6 ±11.1ppb), whereas in the group receiving the mycotoxin detoxifier, DON was not detected in feces. All other analyzed mycotoxins, including FUMB1, T-2/HT-2 toxins, and OTA, were below detection limits in both groups.
3.6. Histopathological Findings
Histopathological examination of liver samples did not reveal treatment-related differences between groups. Both groups exhibited only limited, non-specific lesions, including mild periportal infiltration by few lymphocytes and plasma cells, occasionally extended into the parenchyma. Occasional foci of vacuolar (likely glycogenic) degeneration were observed, along with sporadic presence of Ito cells containing lipofuscin pigment (Figure 1). The observed histological findings are considered non-specific. Lesions are minimal in severity, lack a characteristic pattern associated with a particular pathological process, and are commonly encountered as background or incidental changes in clinically healthy cattle. Mild periportal and parenchymal lymphoplasmacytic infiltrates may reflect physiological immune surveillance or mild, subclinical hepatic irritation, while the scattered vacuolar (likely glycogenic) degeneration and occasional lipofuscin-containing Ito cells represent common, non-pathognomonic age- or metabolism-related alterations. Collectively, these changes do not support a clear treatment-related hepatic effect.
4. Discussion
The present field trial evaluated the effects of a multi-component mycotoxin detoxifier on physiological and oxidative endpoints in fattening mixed Limousin calves exposed to naturally occurring, low-to-moderate levels of multiple mycotoxins under commercial feedlot conditions. The contamination profile of the TMR, characterized by the presence of AFB1, FUMs, and ZEN, is consistent with previous surveys of cattle feeds in Europe and reflects realistic field exposure rather than experimentally induced mycotoxicosis [4,8]. The absence of overt clinical signs throughout the experimental period indicates subclinical exposure, a scenario in which subtle metabolic and oxidative alterations are more likely to be detected than gross clinical disease [3].
Biochemical analyses showed that dietary supplementation with the detoxifier did not affect hepatic or renal function, as all measured parameters remained within physiological reference ranges. The significantly lower γ-GT activity observed in the treated group suggests a hepatoprotective effect under chronic mycotoxin exposure. γ-Glutamyl transferase is considered a sensitive indicator of hepatobiliary stress and has been reported to increase in cattle exposed to aflatoxins and other hepatotoxic mycotoxins, even without marked elevations in AST or ALT activities [3,27,28]. Accordingly, the lack of differences in transaminase activities in this study is consistent with the absence of overt hepatocellular damage and supports the interpretation that γ-GT may respond earlier to subclinical hepatic stress. In this study, ALT and AST activities remained within physiological ranges in both groups, indicating preservation of hepatocyte membrane integrity and absence of clinically relevant cytolysis. In contrast, the higher γ-GT activity observed in control calves indicates early or subclinical hepatobiliary stress and increased metabolic burden associated with mycotoxin biotransformation. The reduction of γ-GT activity in treated animals therefore reflects an alleviation of hepatic functional stress rather than liver injury, suggesting improved efficiency of hepatic detoxification processes. Maintenance of hepatic function is critical in ruminants exposed to dietary mycotoxins, as compromised liver capacity may increase systemic exposure to toxic metabolites and contribute to secondary effects on immune, reproductive, and gastrointestinal functions [33].
The phytogenic components of the tested anti-mycotoxin detoxifier, particularly silymarin and curcumin, have been shown to stabilize hepatocyte membranes, enhance antioxidant defense mechanisms, and limit oxidative modification of cellular macromolecules, thereby supporting hepatic resilience under toxic challenge [34,35]. These mechanisms are consistent with the reduced CARBs levels observed in treated calves, indicating attenuation of oxidative stress at the protein level.
The close association between hepatic function and oxidative status further supports this interpretation. The liver is both a major source and target of oxidative stress in ruminants, particularly during chronic dietary challenges such as mycotoxin exposure [36]. Although TBARS remained unchanged, the concurrent reduction in protein oxidation and γ-GT activity suggests an overall alleviation of subclinical hepatic and oxidative burden. Although no conclusions regarding productive performance can be drawn from this study, improved oxidative status has been associated with enhanced product quality and long-term resilience in livestock, highlighting the potential relevance of such physiological adaptations [37].
In addition, our study found a reduction in CARBs levels in treated calves, indicating decreased protein oxidative damage. Protein carbonyls are widely recognized as robust and sensitive biomarkers of oxidative stress, particularly under chronic, low-grade toxic challenge [36,38]. Mycotoxins such as AFB1 and FUMB1 are known to induce oxidative stress through excessive generation of reactive oxygen species during hepatic biotransformation, leading to oxidative modification of cellular proteins and disruption of redox homeostasis [39]. In this study, the absence of significant differences in TBARS and TAC suggests that lipid peroxidation and overall antioxidant reserves were not markedly affected at the observed exposure levels, whereas protein oxidation appeared more responsive to the intervention. Similar dissociations among oxidative stress indices have been reported in livestock exposed to moderate mycotoxin challenge [36,38].
The presence of AFB1 in the TMR at concentrations exceeding recommended guidance values confirms that animals were subjected to a nutritionally relevant mycotoxin challenge, reinforcing the practical relevance of this field study. Chronic AF exposure is well documented to promote oxidative stress via reactive oxygen species generation during hepatic metabolism, even in the absence of overt clinical disease [39].
The detoxifying agent evaluated in this study combines adsorptive, biological and phytogenic components with complementary modes of action [17]. Adsorbent clays such as bentonite and sepiolite can limit systemic absorption of polar mycotoxins, including AFB1, by gastrointestinal binding, while yeast-derived components may further support mycotoxin sequestration and intestinal barrier integrity. In parallel, the phytogenic compounds silymarin and curcumin exert antioxidant and hepatoprotective effects by stabilizing hepatocyte membranes and modulating cellular redox balance, as previously demonstrated in livestock species [19,40]. Collectively, these findings highlight the value of sensitive oxidative stress biomarkers for detecting early physiological responses to mycotoxin mitigation strategies under commercial feeding conditions, where exposure is often subclinical and clinical signs are absent.
The presence of AFB1 in the diets was demonstrated by the TMR analysis (37 ppb), thus indicating the availability of this mycotoxin for the calves. The detection of AFB1 in fecal samples from treated calves, in contrast to its absence in control animals, suggests that the mycotoxin was more efficiently excreted when the product was administered to the animal, thereby validating the efficacy of the tested product in mitigating AFB1. This observation is consistent with the known affinity of clay-based binders for AFB1 and supports, at a qualitative level, an adsorption-mediated mechanism of action for the product [41]. Fecal detection of AFB1 should not be interpreted as evidence of a complete mass balance, altered systemic bioavailability, or quantitative excretion kinetics, as fecal concentrations reflect only a partial and indirect snapshot of gastrointestinal binding and elimination processes within the complex toxicokinetic of mycotoxins in ruminants [42].
ZEN values found in TMR were low (342.3 ppb). However, it was detected at low concentrations in fecal samples from control animals but not in treated calves. This finding may reflect analytical variability at very low concentration levels, where measured values can fluctuate around the limit of detection, leading to occasional non-detection. Analysing a larger number of samples would likely reduce this variability and yield a more representative assessment. Given the low dietary exposure and the variability of fecal excretion near detection limits, this difference cannot be attributed to a treatment effect. In ruminants, ZEN detoxification involves additional mechanisms beyond adsorption, including microbial transformation and biological protection, which may not be adequately reflected by fecal concentrations alone [8,43]. Similarly, FUMB1 was not consistently detected in fecal samples despite their presence in the TMR. This could be due to extensive ruminal metabolism, alternative excretion pathways such as urinary excretion, or conversion to metabolites not captured by the analytical method used [9,42,44]. However, as the available toxicokinetics data reflecting the metabolism and excretion of these mycotoxins in calves is scarce, it cannot be assured that the analysis of fecal biomarkers could be of interest for FUMs. The potential significance of alternative excretion routes, such as the urinary system, in this context merits further investigation, as the observed absence of FUMs in the samples could be indicative of their presence in the urine, possibly as metabolites.
The low fecal DON concentration observed in the control group despite the relatively high dietary levels is consistent with the known metabolic fate of DON in ruminants. In these species, a large proportion of ingested DON is transformed by ruminal microflora, primarily through de-epoxidation to DOM-1, a substantially less toxic metabolite, resulting in limited excretion of the parent compound [3,8,45]. The absence of detectable DON in fecal samples of the treated group suggests a product-related effect within the digestive tract, potentially involving toxin binding, enhanced microbial transformation, or a combination of both mechanisms, resulting in concentrations below the analytical detection limit. Such findings are consistent with previous reports indicating reduced systemic availability or detectable residues of DON in animals receiving detoxifying feed additives, particularly under conditions of efficient ruminal metabolism [8,46,47]. Furthermore, enzymatic degradation of DON remains inefficient due to its stable epoxide ring and peculiar molecular structure, which limits complete biotransformation under practical feeding conditions [48]. In this context, the most effective strategy for DON mitigation in animal nutrition is not direct toxin binding, but rather the counteraction of its negative biological effects—particularly oxidative stress and hepatic dysfunction—by strengthening animal health [47,48]. This can be achieved through the administration of potent natural ingredients with antioxidant, hepatoprotective, and metabolism-stimulating properties that support the animal’s endogenous detoxification pathways, such as milk thistle (Silybum marianum) and turmeric (Curcuma longa) extracts [49]. Previous in vitro study with the tested anti-mycotoxin agent confirmed the cytotoxicity and the ability to increase ROS levels by DON in hepatic cells, while the tested anti – mycotoxin agent has a protective effect against DON-induced oxidative stress [50]. Moreover, more studies have shown that the tested anti-mycotoxin agent is effective in reducing the concentration of DON in the rumen fluid of dairy cows, significantly influencing the chemical profiles in the rumen, particularly enhancing the binding of mycotoxins, thereby supporting the role of phytogenic extracts in mitigating mycotoxin effects [17].
Overall, these findings underscore the complexity of interpreting fecal mycotoxin profiles in cattle under field conditions. Primary evidence for a biological effect of the detoxifier was derived from hepatic enzyme activity and oxidative stress biomarkers, which are more sensitive and physiologically relevant indicators of subclinical mycotoxin impact [11,51].
In our previous studies in non-ruminants, beneficial effects of supplementation of the same anti-mycotoxin detoxifier in the diets of weaners were noticed, including improved redox status, potential hepatoprotective properties and enhanced growth performance [18]. In addition, our recent study in ruminants exposed to natural mycotoxin contamination, indicated an improvement of oxidative balance and liver function after the administration of the same anti-mycotoxin detoxifier in the diet of dairy sheep [40]. Hepatoprotective and antioxidant properties of silymarin against mycotoxins such as AFB1, and OTA have been reported in poultry and rodents [52,53]. However, the administration of silymarin extract as oral drenches in periparturient dairy cows, has no adverse effect on the liver of lactating cows, and presents no objective evidence for a hepatoprotective effect [54].
In the current study, the histopathological examination of liver tissue revealed only mild, non-specific alterations in both experimental groups. Similar periportal lymphoplasmacytic infiltrates and vacuolar changes have been described as incidental findings in clinically healthy cattle and are not indicative of mycotoxin-induced hepatopathy [8]. The absence of treatment-related histological lesions supports the biochemical findings and confirms that supplementation with the detoxifier was not associated with hepatic toxicity. The absence of a semi-quantitative histopathological scoring system and formal assessment of inter-observer agreement limits the reproducibility and statistical comparison of liver lesions [55]. Consequently, histopathological findings should be interpreted descriptively and in conjunction with biochemical and oxidative stress parameters rather than as standalone comparative outcomes.
A limitation of this study is the lack of individual feed intake measurements, which could not be obtained under commercial group-feeding conditions. Consequently, potential effects of the additive on voluntary intake or feed efficiency could not be directly evaluated. Nevertheless, the primary objective of the trial was to assess physiological and oxidative responses under natural multi-mycotoxin exposure, and these endpoints are interpretable independently of individual intake data. Similar limitations are common in field trials conducted under commercial conditions and do not invalidate conclusions related to metabolic or hepatic health [8]. The absence of baseline and repeated measurements for biochemical and oxidative stress parameters limits the interpretation of the findings to endpoint differences between groups. Although stratified randomization was applied to minimize baseline imbalance, residual between-animal variability cannot be completely excluded. Future studies incorporating baseline and longitudinal sampling would allow a more robust assessment of temporal changes associated with dietary mycotoxin mitigation. Due to the limited sample size and the evaluation of multiple outcomes, statistically significant findings should be interpreted cautiously and considered exploratory.
In addition, the lack of blinding during routine clinical observations represents a potential source of observer bias, particularly for subjective or mild clinical endpoints. However, this limitation is inherent to field studies conducted under commercial conditions and does not affect laboratory-based or histopathological outcomes, which were assessed under blinded conditions. Although the a priori sample size calculation was informed by performance-related endpoints reported in previous cattle mycotoxicosis studies, the present findings suggest that γ-GT activity and protein oxidation markers may represent more sensitive indicators of biological response under subclinical exposure conditions. Future studies should therefore consider basing sample size calculations on hepatic or oxidative stress biomarkers to optimize the detection of hepatoprotective effects.
5. Conclusions
In conclusion, the supplementation of diet with the tested multi-component mycotoxin detoxifier in fattening calves exposed to naturally contaminated feed was safe and associated with improved hepatic enzyme regulation and reduced protein oxidative damage. These findings support previous evidence that combined adsorbent–phytogenic detoxifiers can mitigate the subclinical physiological impact of mycotoxins in ruminant production systems [17,18,19,27].
Author Contributions
Conceptualization, V.G.P, G.I.P, D.G., I.R., O.C., A.M.D., and K.D.; methodology, V.G.P, G.I.P, D.G., D.P., D.A., I.R., O.C., and C.E; software, D.G.; validation, V.G.P., D.P., and C.E.; formal analysis, D.G., D.P., C.M., and K.D.; investigation, K.D., D.A., G.I.P., I.R., O.C., and C.M.; resources, I.R., A.M.D. and O.C.; data curation, K.D., D.A., C.E. and C.M.; writing—original draft preparation, K.D., and D.G.; writing—review and editing, V.G.P., G.I.P., D.P., C.E., I.R., O.C., A.M.D. and D.G.; visualization, K.D., C.M.; supervision, D.G., and V.G.P.; project administration, D.G.; funding acquisition, I.R., A.M.D. and O.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by BIŌNTE NUTRITION S.L. (Calle México 33, Polígono Industrial Tecnoparc, 43204 Reus-Tarragona, Spain), through the Research Committee of the University of Thessaly (grant number 8040).
Institutional Review Board Statement
The animal study protocol was reviewed and approved by the Ethics and Research Deontology Committee (E.R.D.C.) of the University of Thessaly, Greece (protocol code: 48; date of approval: 04/06/2024).
Informed Consent Statement
Written informed consent was obtained from the owner of the client-owned animals prior to the initiation of the study.
Acknowledgments
We thank the farm owner for providing valuable data.
Conflicts of Interest
Authors Insaf Riahi and Óscar Castro were employed by BIŌNTE Nutrition S.L (Reus, Spain). The other authors declare no potential conflicts of interest related to the study.
Abbreviations
The following abbreviations are used in this manuscript:
| Abbreviation | Definition |
| γ-GT | Gamma-glutamyl transferase |
| AF | Aflatoxins |
| AFB1 | Aflatoxin B1 |
| ALT | Alanine aminotransferase |
| aNDFom | Amylase-treated neutral detergent fiber, expressed exclusive of residual ash |
| AST | Aspartate aminotransferase |
| BFH12 | Bovine fetal hepatocyte cell line |
| CARBs | Protein carbonyls |
| CP | Crude protein |
| CYP1A1 | Cytochrome P450 1A1 |
| CYP3A74 | Cytochrome P450 3A74 |
| DM | Dry matter |
| DON | Deoxynivalenol |
| E.R.D.C. | Ethics and Research Deontology Committee |
| EE | Ether extract |
| EU | European Union |
| FASS | Federation of Animal Science Societies |
| FUMs | Fumonisins |
| FUMB1 | Fumonisin B1 |
| H&E | Hematoxylin and eosin |
| HT-2 | HT-2 toxin |
| LC–MS/MS | Liquid chromatography–tandem mass spectrometry |
| LOD | Limit of detection |
| LOQ | Limit of quantification |
| MDA | Malondialdehyde |
| nd | Not detected |
| ppb | parts per billion |
| OTA | Ochratoxin A |
| SD | Standard deviation |
| SPSS | Statistical Package for the Social Sciences |
| T-2 | T-2 toxin |
| TAC | Total antioxidant capacity |
| TBARS | Thiobarbituric acid reactive substances |
| TMR | Total mixed ration |
| ZEN | Zearalenone |
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef]
- Magnoli, A.P.; Poloni, V.L.; Cavaglieri, L. Impact of mycotoxin contamination in the animal feed industry. Curr. Opin. Food Sci. 2019, 29, 99–108. [Google Scholar] [CrossRef]
- Zinedine, A.; El Akhdari, S. Food Safety and Climate Change: Case of Mycotoxins. In Handbook of Research on Global Environmental Changes and Human Health; IGI Global: Hershey, PA, USA, 2019; pp. 39–62. [Google Scholar]
- Rodrigues, I. A Review on the Effects of Mycotoxins in Dairy Ruminants. Anim. Prod. Sci. 2014, 54, 1155–1165. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-Mycotoxin Screening Reveals the Occurrence of 139 Different Secondary Metabolites in Feed and Feed Ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on mycotoxin issues in ruminants: Occurrence in forages, effects on health and animal performance and practical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef]
- Dänicke, S.; Brezina, U. Kinetics and metabolism of the Fusarium toxin deoxynivalenol in farm animals. Food Chem. Toxicol 2013, 60, 58–75. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Marroquin-Cardona, A.G.; Johnson, N.M.; Phillips, T.D.; Hayes, A.W. Mycotoxins in a changing global environment: A review. Food Chem. Toxicol. 2014, 69, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Iori, S.; Lahtela-Kakkonen, M.; D'Onofrio, C.; Maietti, F.; Mucignat, G.; Bardhi, A.; Barbarossa, A.; Zaghini, A.; Pauletto, M.; Dacasto, M.; Giantin, M. New insights into aflatoxin B1 mechanistic toxicology in cattle liver: an integrated approach using molecular docking and biological evaluation in CYP1A1 and CYP3A74 knockout BFH12 cell lines. Arch Toxicol. 2024, 98, 3097–3108. [Google Scholar] [CrossRef] [PubMed]
- Patsoukis, N.; Georgiou, C. D. Determination of the thiol redox state of organisms: new oxidative stress indicators. Anal. Bioanal. Chem. 2004, 378(7), 1783–1792. [Google Scholar] [CrossRef]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol Immunotoxicol. 2011, 33, 233–240. [Google Scholar] [CrossRef]
- Jouany, J.P. Methods for preventing, decontaminating and minimizing the toxicity of mycotoxins in feeds. Anim. Feed Sci. Technol. 2007, 137, 342–362. [Google Scholar] [CrossRef]
- Kolosova, A.; Stroka, J. Substances for reduction of the contamination of feed by mycotoxins: A review. World Mycotoxin J. 2011, 4, 225–256. [Google Scholar] [CrossRef]
- Fiorbelli, E.; Lapris, M.; Errico, M.; Della Badia, A.; Riahi, I.; Rocchetti, G.; Gallo, A. Mycotoxin Challenge in Dairy Cows: Assessment of the Efficacy of an Anti-Mycotoxin Agent by Adopting an In Vitro Rumen Simulation Method. Toxins. 2024, 16, 490. [Google Scholar] [CrossRef]
- Papatsiros, V.G.; Eliopoulos, C.; Voulgarakis, N.; Arapoglou, D.; Riahi, I.; Sadurní, M. Effects of a multi-component mycotoxin-detoxifying agent on oxidative stress, health and performance of sows. Toxins 2023, 15, 580. [Google Scholar] [CrossRef]
- Papatsiros, V.G.; Papakonstantinou, G.I.; Voulgarakis, N.; Eliopoulos, C.; Marouda, C.; Meletis, E. Effects of a curcumin/silymarin/yeast-based mycotoxin detoxifier on redox status and growth performance of weaned piglets. Toxins 2024, 16, 168. [Google Scholar] [CrossRef]
- Armanini, E.H.; Boiago, M.M.; de Oliveira, P.V.; Roscamp, E.; Strapazzon, J.V.; de Lima, A.G.; Copetti, P.M.; Morsch, V.M.; de Oliveira, F.C.; Wagner, R.; et al. Inclusion of a phytogenic bend in broiler diet as a performance enhancer and anti-aflatoxin agent: Impacts on health, performance, and meat quality. Res. Vet. Sci. 2021, 137, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; He, J.; Ahmad, H.; Shen, M.; Zhao, Y.; Gan, Z.; Zhang, L.; Zhong, X.; Wang, C.; Wang, T. Dietary Curcumin Supplementation Increases Antioxidant Capacity, Upregulates Nrf2 and Hmox1 Levels in the Liver of Piglet Model with Intrauterine Growth Retardation. Nutrients. 2019, 11, 2978. [Google Scholar] [CrossRef] [PubMed]
- Ledur, P.C.; Santurio, J.M. Cytoprotective Effects of Curcumin and Silymarin on PK-15 Cells Exposed to Ochratoxin A, Fumonisin B1 and Deoxynivalenol. Toxicon. 2020, 185, 97–103. [Google Scholar] [CrossRef]
- Tedesco, D.; Steidler, S.; Galletti, S.; Tameni, M.; Sonzogni, O.; Ravarotto, L. Efficacy of Silymarin-Phospholipid Complex in Reducing the Toxicity of Aflatoxin B1 in Broiler Chicks. Poult. Sci. 2004, 83, 1839–1843. [Google Scholar] [CrossRef]
- Tuong, D. T. C.; Moniruzzaman, M.; Smirnova, E.; Chin, S.; Sureshbabu, A.; Karthikeyan, A.; Min, T. Curcumin as a Potential Antioxidant in Stress Regulation of Terrestrial, Avian, and Aquatic Animals: A Review. Antioxidants 2023, 12, 1700. [Google Scholar] [CrossRef]
- Tedesco, D.E.A.; Guerrini, A. Use of milk thistle in farm and companion animals: A review. Planta Med. 2023, 89, 584–607. [Google Scholar] [CrossRef] [PubMed]
- Wellington, K.; Jarvis, B. Silymarin: A review of its clinical properties in the management of hepatic disorders. BioDrugs 2001, 15, 465–489. [Google Scholar] [CrossRef] [PubMed]
- Catellani, A.; Ghilardelli, F.; Trevisi, E.; Cecchinato, A.; Bisutti, V.; Fumagalli, F.; Gallo, A. Effects of supplementation of a mycotoxin mitigation feed additive in lactating dairy cows. Toxins 2023, 15, 546. [Google Scholar] [CrossRef] [PubMed]
- Helferich, W.G.; Baldwin, R.L.; Hinders, R.G. Effects of aflatoxin B1 on feed intake, growth, and liver function in dairy cattle. J. Dairy Sci. 1986, 69, 2237–2245. [Google Scholar]
- European Commission. European Commission Directive 2002/32/EC of The European Parliament and of The Council of 7 May 2002 on Undesirable Substances in Animal Feed. Off. J. Eur. Union 2019, L289, 32–36. [Google Scholar]
- European Commission. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol; zearalenone; ochratoxin A; T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, L229, 7–9. [Google Scholar]
- European Commission. Commission Recommendation of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products. Off. J. Eur. Union 2013, L91, 12–15. [Google Scholar]
- European Commission. Commission Regulation (EU) No 574/2011 of 16 June 2011 amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council as regards maximum levels for nitrite; melamine; Ambrosia spp. and car-ry-over of certain coccidiostats and histomonostats and consolidating Annexes I and II thereto. Off. J. Eur. Union 2011, L159, 7–24. [Google Scholar]
- Gatellier, P.; Mercier, Y.; Renerre, M. Effect of Diet Finishing Mode (Pasture or Mixed Diet) on Antioxidant Status of Charolais Bovine Meat. Meat Sci. 2004, 67, 385–394. [Google Scholar] [CrossRef]
- Bozakova, N.; Ivanov, V. Opportunities for sheep welfare improvement by silymarin additive–a review article. Bulg. J. Agric. Sci. 2022, 28, 502–509. [Google Scholar]
- Jaguezeski, A.M.; Gündel, S.S.; Favarin, F.R.; Gündel, A.; Souza, C.F.; Baldissera, M.D.; Cazarotto, C.C.; Volpato, A.; Fortuoso, B.F.; Ourique, A.F.; Da Silva, A.S. Low-dose curcumin-loaded Eudragit L100-nanocapsules in the diet of dairy sheep increases antioxidant levels and reduces lipid peroxidation in milk. J. Food Biochem. 2019, 43(8). [Google Scholar] [CrossRef]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front. Vet. Sci. 2015, 2. [Google Scholar] [CrossRef] [PubMed]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla Forage (Sulla coronariumL.) on the Oxidative Status and Milk Polyphenol Content in Goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef]
- Bacou, E.; Walk, C.; Rider, S.; Litta., G.; Perez-Calvo, E. Dietary Oxidative Distress: A Review of Nutritional Challenges as Models for Poultry, Swine and Fish. Antioxidants. 2021, 10(4), 525. [Google Scholar] [CrossRef]
- Bernabucci, U.; Colavecchia, L.; Danieli, P.P.; Basiricò, L.; Lacetera, N.; Nardone, A.; Ronchi, B. Aflatoxin B1 and fumonisin B1 affect the oxidative status of bovine peripheral blood mononuclear cells. Toxicol. In Vitro 2011, 25, 684–691. [Google Scholar] [CrossRef]
- Papakonstantinou, G. I.; Eliopoulos, C.; Meletis, E.; Riahi, I.; Stampinas, E. G.; Arapoglou, D.; Gougoulis, D.; Dimoveli, K.; Filippou, D.; Manouras, A.; Tsekouras, N.; Fotos, L.; Kostoulas, P.; Christodoulopoulos, G.; Papatsiros, V. G. Effects of a phytogenic mycotoxin detoxifier on oxidative status, health, and performance in dairy sheep. Toxins. 2025, 17, 425. [Google Scholar] [CrossRef] [PubMed]
- Vila-Donat, P.; Marín, S.; Sanchis, V.; Ramos, A.J. A review of the mycotoxin adsorbing agents, with an emphasis on multi-binding capacity. Food Chem. Toxicol. 2018, 114, 246–259. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Sung, H.G.; Lee, C.H. Comparative study on the aflatoxin B1 degradation ability of rumen fluid from Holstein steers and Korean native goats. J. Vet. Sci. 2009, 10, 29–34. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Faas, J.; Doupovec, B. Metabolism of zearalenone in the rumen of dairy cows with and without application of a zearalenone-degrading enzyme. Toxins 2021, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Bandyk, C.A. Review: Mycotoxins in ruminant livestock production. Appl. Anim. Sci. 2024, 40, 802–817. [Google Scholar] [CrossRef]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl Microbiol Biotechnol. 2011, 91(3), 491–504. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Risks to animal and human health related to deoxynivalenol. EFSA Journal. 2017, 15(9), 4718.
- Pestka, J.J. Deoxynivalenol: mechanisms of action, human exposure, and toxicological relevance. Arch Toxicol. 2010, 84, 663–79. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef]
- Surai, P.F. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef]
- Riahi, I.1; Sadurní, M.1; Codina, R.1; Ferrús, D.1; Escrivá, L.2; Meca, G. The in vitro efficacy of a detoxifying agent to mitigate don-induced oxidative stress in hepatic cells. Book of abstracts of 1st International Workshop of the Spanish network on mycotoxins, toxigenic fungi and their decontamination processes & MycoTWIN Workshops 5-6, Valencia, June 28 to 30, 2023; p. 44. [Google Scholar]
- Mavrommatis, A.; Giamouri, E.; Tavrizelou, S.; Zacharioudaki, M.; Danezis, G.; Simitzis, P. E.; Zoidis, E.; Tsiplakou, E.; Pappas, A. C.; Georgiou, C. A.; Feggeros, K. Impact of Mycotoxins on Animals' Oxidative Status. Antioxidants 2021, 10, 214. [Google Scholar] [CrossRef]
- Riahi, I.; Ramos, A.J.; Raj, J.; Jakovˇcevi´c, Z.; Farkaš, H.; Vasiljevi´c, M.; Pérez-Vendrell, A.M. Effect of a Mycotoxin Binder (MMDA) on the Growth Performance, Blood and Carcass Characteristics of Broilers Fed Ochratoxin A and T-2 Mycotoxin Contaminated Diets. Animals 2021, 11, 3205. [Google Scholar] [CrossRef]
- Damiano, S.; Longobardi, C.; Andretta, E.; Prisco, F.; Piegari, G.; Squillacioti, C.; Montagnaro, S.; Pagnini, F.; Badino, P.; Florio, S.; et al. Antioxidative Effects of Curcumin on the Hepatotoxicity Induced by Ochratoxin A in Rats. Antioxidants. 2021, 10, 125. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, D.; Domeneghini, C.; Sciannimanico, D.; Tameni, M.; Steidler, S.; Galletti, S. Silymarin, a possible hepatoprotector in dairy cows: biochemical and histological observations. J Vet Med A Physiol Pathol Clin Med. 2004, 51(2), 85–90. [Google Scholar] [CrossRef] [PubMed]
- Gibson-Corley, K. N.; Olivier, A. K.; Meyerholz, D. K. Principles for valid histopathologic scoring in research. Vet. Pathol 2013, 50(6), 1007–1015. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Liver, hematoxylin-eosin stain, magnification x 200, a: infiltration of the periportal area by lymphocytes and plasma cells (arrows)–(control group); Ito cells containing lipofuscin (arrowhead) (control group), b: focal infiltration of the hepatic parenchyma by scattered lymphocytes and plasma cells, c: micro-vacuolar degeneration of hepatocyte cytoplasm (arrowheads) (treated group).
Figure 1.
Liver, hematoxylin-eosin stain, magnification x 200, a: infiltration of the periportal area by lymphocytes and plasma cells (arrows)–(control group); Ito cells containing lipofuscin (arrowhead) (control group), b: focal infiltration of the hepatic parenchyma by scattered lymphocytes and plasma cells, c: micro-vacuolar degeneration of hepatocyte cytoplasm (arrowheads) (treated group).
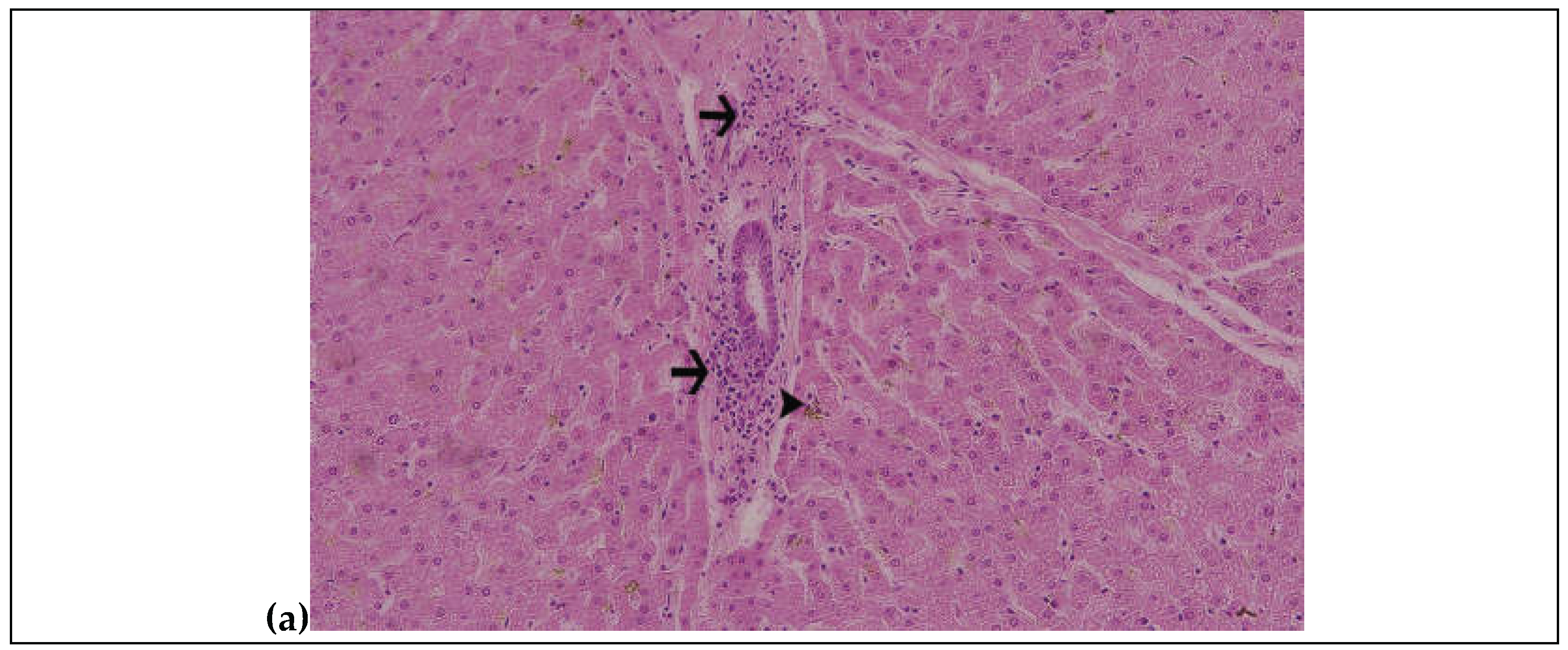
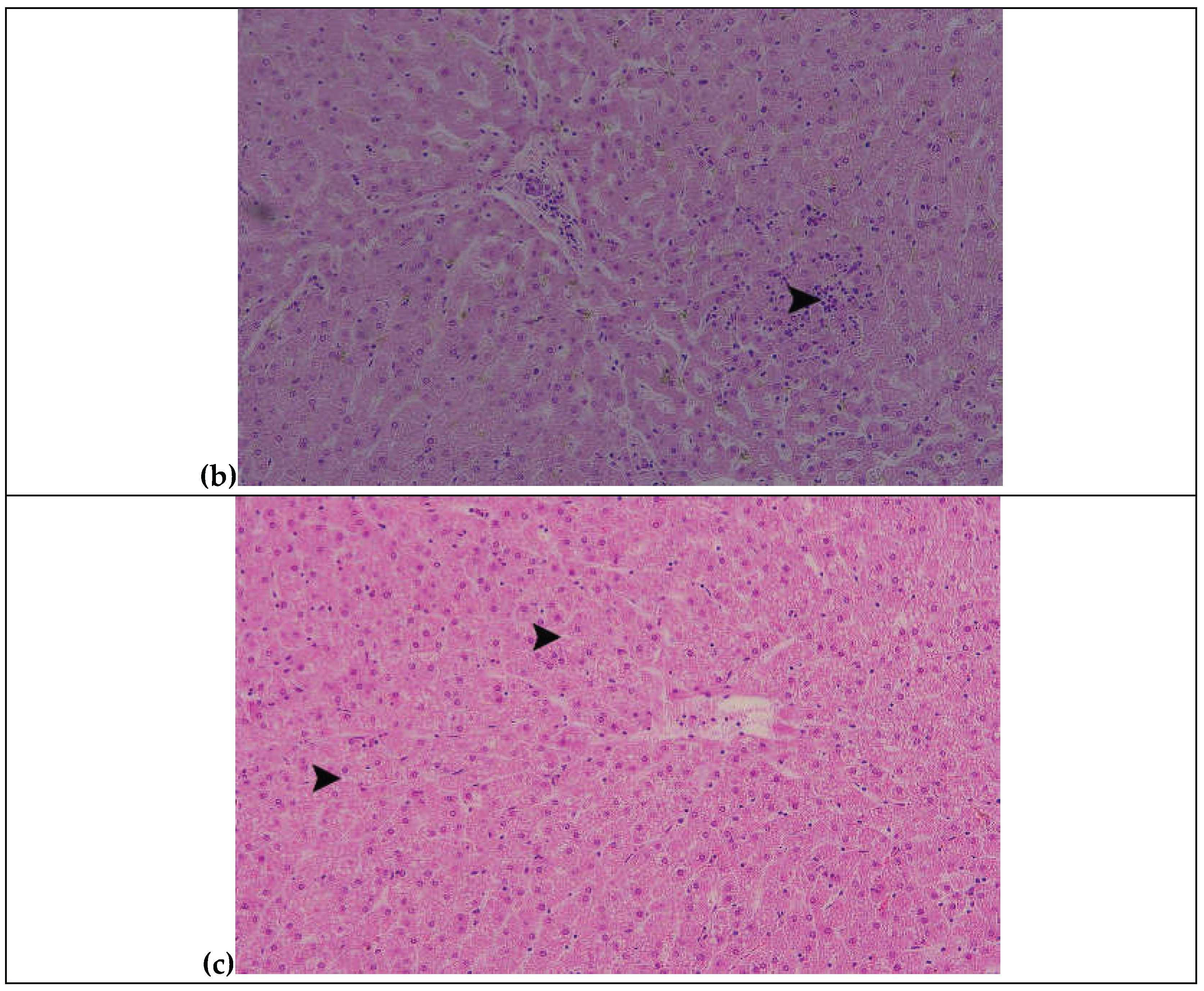

Table 1.
Chemical composition of ingredients and experimental diets (TMR; mean ± SD).
| Ingredients | Kg as fed | Chemicals | |||||
|---|---|---|---|---|---|---|---|
| DM | CP | EE | Starch | Ash | aNDFom | ||
| Oat Hay | 5.00 | 88.0 ± 1.0 | 8.0 ± 0.5 | 1.5 ± 0.2 | 2.0 ± 0.3 | 8.5 ± 0.5 | 50.0 ± 1.0 |
| Corn meal | 4.3 | 88.02 ± 0.28 | 10.73 ± 0.14 | 4.34 ± 0.15 | 69.47 ± 1.73 | 1.53 ± 0.11 | - |
| Soymeal (CP: 47%) | 0.550 | 88.83 ± 0.04 | 48.41 ± 0.71 | 3.70 ± 0.14 | - | 7.17 ± 0.48 | - |
| Barley | 3.75 | 88.0 ± 1.0 | 11.0 ± 0.5 | 2.0 ± 0.2 | 60.0 ± 1.5 | 2.5 ± 0.3 | 19.0 ± 1.0 |
| Wheat bran | 0.93 | 89.0 ± 1.0 | 15.5 ± 0.5 | 4.0 ± 0.2 | 18.0 ± 1.0 | 5.0 ± 0.3 | 42.0 ± 1.5 |
| Cotton seed (kg/DM) | 0.93 | 92.0 ± 1.0 | 23.0 ± 1.0 | 19.0 ± 0.5 | 0.5 ± 0.1 | 4.0 ± 0.2 | 30.0 ± 1.0 |
| Mineral and vitamin premix kg/DM* | 0.04 | 95.0 ± 1.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 90.0 ± 2.0 | 0.0 ± 0.0 |
| Salt | 0.04 | 95.0 ± 1.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 95.0 ± 2.0 | 0.0 ± 0.0 |
| CaCO3 | 0.06 | 95.0 ± 1.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 95.0 ± 2.0 | 0.0 ± 0.0 |
| *Chemical Analysis %: Sodium (Na): 7.70% Calcium (Ca): 22.13% Phosphorus (P): 5.15% Magnesium (Mg): 0.72% Additives per kg: Vitamins: Vitamin A (3a672a) 480,000 I.U. Vitamin D3 (3a671) 96,000 I.U. Vitamin E (3a700) 4,000 mg, Vitamin B1 (3a821) 200 mg, Niacin (3a314) 4,000 mg, Biotin (3a880) 200 mg, Choline (3a890) 2,000 mg Trace Elements: Manganese (Oxide-3b502) 2,080 mg, Iron (Oxide-E172) 2,000 mg, Zinc (Oxide-3b603) 4,800 mg, Iodine (Calcium Iodate-3b202) 80 mg, Selenium (Sodium Selenite-3b801) 20 mg DM: Dry Matter, CP: Crude Protein, EE: Ether Extract, aNDFom: amylase-treated Neutral Detergent Fiber, expressed exclusive of residual ash | |||||||
Table 2.
Mean mycotoxin concentrations (ppb) in the Total Mixed Ration (TMR) across the experimental period.
Table 2.
Mean mycotoxin concentrations (ppb) in the Total Mixed Ration (TMR) across the experimental period.
| Type of mycotoxin | TMR (ppb) |
EU regulatory maximum levels (p.p.b.) | Guidance values (p.p.b.) |
|---|---|---|---|
| AFB1 | 37 | 10 | 5 |
| FUMs | 8402 | 20,000 | 1,500 |
| DON | 4077 | 2,000 | 300 |
| ZEN | 342.3 | 500 | 100 |
| T-2 / HT-2 toxins | <40 | – | 50 |
| OTA | 1.75 | 100 | 50 |
| * European Commission [29,30,31,32] | |||
Table 3.
Mean values (±SD) of biochemical parameters in fattening mixed Limousin calves of the control (T1) and treatment (T2) groups at the end of the 60-day experimental period.
Table 3.
Mean values (±SD) of biochemical parameters in fattening mixed Limousin calves of the control (T1) and treatment (T2) groups at the end of the 60-day experimental period.
| Parameter | T1 (Control group) | T2 (Experimental group) |
|---|---|---|
| Urea (mg/dL) | 22.17 ± 2.79 | 18.17 ± 8.28 |
| Creatinine (mg/dL) | 1.91 ± 0.18 | 1.89 ± 0.17 |
| SGOT (AST) (U/L) | 111.5 ± 17.2 | 112.5 ± 15.33 |
| SGPT (ALT) (U/L) | 50.5 ± 6.02 | 53.17 ± 6.77 |
| γ-GT (U/L) | 20.0 ± 2.45* | 16.67 ± 2.70* |
| Total Proteins (g/dL) | 8.52 ± 0.60 | 8.62 ± 0.56 |
| Albumin (g/dL) | 4.42 ± 0.13 | 4.65 ± 0.15 |
| * A statistically significant decrease (P < 0.05) in γ-glutamyl transferase (γ-GT) activity was observed in the treatment group compared with the control group. | ||
Table 4.
Protein oxidation (CARBs), lipid peroxidation (TBARS) and total antioxidant capacity (TAC) in T1 and T2 calves. Values expressed as mean ± SD.
Table 4.
Protein oxidation (CARBs), lipid peroxidation (TBARS) and total antioxidant capacity (TAC) in T1 and T2 calves. Values expressed as mean ± SD.
| Biomarkers | T1 (control group) |
T2 (experimental group) |
P value |
|---|---|---|---|
| CARBs (nmol/mg protein) | 1.14 ± 0.2 | 0.87 ± 0.3 | <0.05 |
| TBARS (μM MDA) | 6.27 ± 0.2 | 5.45 ± 0.3 | 0.24 |
| TAC (mM Trolox Equivalent) | 0.83 ± 0.1 | 0.83 ± 0.01 | 0.11 |
Table 5.
Concentration of mycotoxins (ppb) detected in fecal samples from calves in the control (T1) and experimental (T2) groups. Values expressed as mean ± SD.
Table 5.
Concentration of mycotoxins (ppb) detected in fecal samples from calves in the control (T1) and experimental (T2) groups. Values expressed as mean ± SD.
| Mycotoxin | T1 (control group) |
T2 (experimental group) |
|---|---|---|
| AFB1 | nd | 19.0 ± 3.0 |
| FUMB1 | nd | nd |
| DON | 90.6 ± 11.1 | nd |
| ZEN | 23.0 ± 3.0 | nd |
| T2/HT-2 toxins | nd | nd |
| OTA | nd | nd |
| nd = not detected. | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



