Submitted:
13 February 2026
Posted:
14 February 2026
You are already at the latest version
Abstract
Candidozyma auris (formerly Candida auris) is an emerging multidrug-resistant pathogenic fungus with an increased ability to cause outbreaks in health care facilities leading to poor patient outcomes. Since its initial discovery in 2009, C.auris has spread rapidly across continents and is now classified by both the Centers for Disease Control and Prevention (CDC) and the World Health Organization (WHO) as a critical-priority pathogen. This review summarizes current knowledge on the origin, taxonomy, microbiology, and virulence mechanisms of C.auris, emphasizing its thermotolerance, osmotolerance, and biofilm-forming capacity on biotic and abiotic surfaces, as well as aspects related to its antifungal drug resistance and management. These features, together with its genomic plasticity, contribute to persistence, transmission, and drug resistance. Emerging evidence also supports a potential link between climate change and C. auris evolution, highlighting environmental adaptation as a driver of pathogenicity. Combating C. auris will require multidisciplinary efforts to mitigate its expanding global impact.
Keywords:
Candidozyma auris
; pathogenesis
; antifungal drug resistance
; biofilms
1. Introduction
Humanity has been afflicted historically by the emergence and spread of infectious diseases, which remain a leading cause of death globally (Piret and Boivin, 2021). Among these, fungal infections (FIs) represent an increasing threat to a variety of patients. The range of FIs can vary significantly in humans given the diverse routes of acquisition, epidemiology, tropisms, and virulence factors [1,2]. Invasive mycoses linked to these risk factors are increasingly associated with high global morbidity and mortality, with an estimated 1.7 million deaths annually [3]. Despite widespread awareness of their health implications, FIs continue to be poorly addressed. Persistent shortcomings in prevention, diagnosis, research funding, and treatment fuel the prevalence of these irrefutably important threats to our public health.
Among the causative agents of fungal infections, Candidozyma auris represents an increasingly significant emergent pathogen [4]. In 2019, the CDC ranked this nosocomial pathogen as an “urgent threat” due to its healthcare outbreak pervasiveness, ability to form biofilms on medical equipment (e.g., catheters, hospital beds, etc.), and multidrug resistance, conducive to refractory treatment [2,5,6,7]. Combined, these characteristics earned C. auris the accolade of “superbug”. More recently, the World Health Organization (WHO) identified fungi as emergent threats to human health, with C. auris being classified as a high-priority organism [8]. In this review, we summarize the origin, identification, biology, pathogenesis, mechanisms of antifungal resistance, and current and future directions that must be taken to successfully tackle the rapid dissemination of C. auris.
2. The Emergence Hypothesis
C. auris was first isolated in 2009 from the external ear canal of a 70-year-old female inpatient in a Japanese hospital. As it was isolated from otic discharge, it was given the species name auris, derived from the Latin word for ear [2,9,10,11]. According to the CDC, C. auris has been detected and isolated in over sixty different countries since its first appearance, spanning all continents except Antarctica [12]. Due to its rapid spread, the CDC had stopped tracking cases in the US until its recent resurgence. It is now considered endemic in healthcare across multiple continents [13], but where did it come from?
Casadevall and colleagues hypothesized that C. auris did not always possess thermotolerance and halotolerance [9]. A stress-adapted C. auris may have risen due to natural and/or anthropogenically induced global warming, making it the first fungal pathogen to emerge due to climate change [14]. Hostile climate conditions are believed to positively select for certain stress-associated genes, and overexpression of metabolic pathways regulated by Hsp90 and the Hog1 stress-activated protein kinase (SAPK) may contribute to virulence, drug resistance, and stress resistance. These same metabolic shifts are evolutionarily conserved in C. albicans. It is possible that global warming may continue exposing and spreading many fungi with pathogenic potential, including C. auris [9].
Given its resistance to hypersaline conditions, Casadevall et al (2021) also hypothesized that C. auris primitive environmental reservoir could have been the wetlands. These highly ionic environments exerted selective pressure which gave rise to unique adhesion factors. Among these, characterization of a surface colonization factor 1 (SCF1) was described, a cation rich adhesin conserved in C. auris and Candida haemulonii, but absent in other Candida species [15]. SCF1 mediates cation dependent surface interactions, enhancing biofilm formation in hypersaline ionized environments which supports the notion that this organism originates from wetlands. This adaptation also promotes adherence in high salinity and lipid-rich niches, thereby facilitating persistent colonization and survival on both human skin and abiotic surfaces [16]. C. auris has an abrogated ability to thrive under anaerobic and highly acidic conditions [17], which infers the notion that it first existed as a plant saprophyte rather than a commensal in the mammalian gut microbiome. This may also explain why C. auris preferentially colonizes cooler skin sites such as the nose, groin, axilla, ear, and other skin locations with cooler temperatures rather than the bloodstream. Another interesting hypothesis as to the acquisition of virulence is the possible interspecies mating between a pathogenic C. albicans and a non-pathogenic C. auris, which may have conferred virulence to the then-saprobe via plasmid DNA transfer. This is likely given the probability that wild forms of C. albicans could be found in the same wetlands [9].
Selection pressure may have also resulted in adaptation to basal temperatures observed in mammalian and avian species, which led Casadevall and colleagues to conceptualize a scheme to explain C. auris’s migratory patterns. Briefly, after acquiring the ability to grow at temperatures between 40 to 42 °C and resistance to hypersalinity in the wake of environmental metamorphosis, adapted C. auris strains may have integrated into avian intermediate hosts. Migration to rural areas with human populations would quickly ensue after C. auris use of birds as vehicular intermediaries. Moreover, the use of aviculture in rural areas may have increased fungal transmission between species, which would eventually lead to C. auris migration to urban areas, including hospital settings [9].
C. auris isolates have been exhaustively collected from various healthcare facilities across multiple continents [13], making it an exclusive human pathogen until recently. Isolation of wild C. auris samples from salt marshes in the Andaman Islands and from estuaries in Colombia suggested that wetland ecosystems may serve as reservoirs prior to human hosts [18,19]. Antifungal susceptibility testing found that 23 out of 24 C. auris samples had high MICs for fluconazole (FLU), and expected MICs for amphotericin B (AMB), while only one isolate had low MICs for both drugs. Remarkably, all isolates were obtained from salt marshes, but 95% of them still showed resistance. Similarly, an isolate from Colombian estuaries also exhibited low MIC profiles for FLU, AMB, voriconazole, and anidulafungin [19]. In both cases, the sensitive strains were isolated from areas with sparse human activity. WGS of the Andaman samples separated the susceptible strain from the resistant strains by 77 to 78 SNPs, which may explain the differences in susceptibility profiles [18]. Comparative and population genomics along with transcriptomic analyses should be performed to identify differences in drug target genes as a means of evolution. The potential environmental reservoirs of C. auris, including wastewater treatment plants, plastics, as well as natural environments, such as salt marshes, sand, seawater, estuaries, apples and dogs, are discussed widely within the literature [20,21,22].
3. Microbial Characteristics
3.1. Phenotypic
C. auris colonies have a white to grey appearance on Sabouraud agar plates. Yeast cells are ovoid in shape, appearing in groups, pairs or singles, and are typically between 2.0 and 5.0 µm in diameter [5,11]. Although the budding yeast morphology predominates in C. auris, a study found that pseudohyphae are formed upon HSP 90 depletion [23]. Pseudohyphal growth is characterized by a wide cell diameter compared to yeast cells, elongated cell morphology due to delayed or incomplete cell division (mother-daughter cell attachments remain), and a lack of cytoplasmic bridging and parallel divisions [24]. Growth as pseudohyphae can occur due to genotoxic stress or hypersaline conditions, and may also be triggered within the mammalian body environment [5,25,26]. C. auris grows well at 42 °C, and its carbon assimilation profile differs from other closely related Candida species [11]. Moreover, C. auris is unable to produce chlamydospores, which are described as thick-walled, hypha-specific cells produced by many fungi capable of filamentation [27,28]. When grown on fetal bovine serum, it fails to produce germ tubes compared to C. albicans, though it retains the ability to adhere to inert surfaces [27].
3.2. Genotypic
C. auris belongs to the order Saccharomycetales within the phylum Ascomycota. Molecular analyses, including sequencing of the 26S rDNA D1/D2 domain and DNA ITS regions place this species in the Clavispora clade of the Metschnikowiaceae family [29]. Phylogenetic relatives Candida duobushaemulonii, Candida pseudohaemulonii, and Candida haemulonii are also part of the same family [5,30]. At least 9 Candida spp. including Candida albicans, C. tropicalis, C. parapsilosis, and C. auris belong to the CTG clade species. In this clade CUG codes serine rather than the universal leucine, which contributes to proteomic diversity and stress adaptation [30]. This change is mediated by a mutant tRNA known as Ser tRNA(CAGSer). The mutation emerged from the continuous competition with its counterpart tRNA(CAGLeu) for CUG codons during the mRNA process. Eventually, this antagonism between tRNAs resulted in the loss of tRNA(CAGLeu) and survival of the analogous tRNA(CAGSer), begetting the CTG clade [31]. Initial epidemiological and whole-genome sequencing (WGS) studies helped identify four karyotypically distinct C. auris clades grouped by geographic regions where they were initially detected: South Asian (I), East Asian (II), South African (III), and South American (IV) [32]. WGS and multi-locus sequence typing determined that the emergence of C. auris was simultaneous and independent across these regions. Within each clade, isolates were clonal with minimal small-nucleotide polymorphism (SNPs) diversity [5,33,34]. The reasons for the concurrent emergence of genetically distinct clades remain a mystery. The majority of infections cluster within the four main clades. Beyond these, a sample belonging to a potential fifth clade was recovered from the ear of a 14-year-old female patient in an Iranian hospital. The patient had not been exposed to other isolates from previously identified clades. WGS confirmed that the strain indeed belonged to clade V, having a marked genetic divergence of >200,000 SNPs compared to the other clades [35]. Most recently, a sixth clade (VI), also known as Indomalayan, has been identified in Singapore and Bangladesh [36]. Bayesian molecular dating was used on a phylogeny of the four initial C. auris clades to determine the time to the most recent common ancestor.
Retrospectively, bloodstream isolates of C. auris date back as early as 1996 in South Korea, where, as a yeast misidentified by Vitek 2 YST and API 20C systems as C. haemulonii and Rhodotorula glutinis, caused the first three C. auris-related cases of hospital-acquired fungemia [37]. Additionally, ear isolates collected in South Korea between 2004-2006 also misidentified as C. haelumonii were later reclassified as C. auris [34]. It was estimated that the current C. auris clades have a common ancestor dating as far back as 360 years ago, and the primary outbreak-causing clade clusters I, III, and IV appeared as early as 36 to 38 years ago [5,38]. Recently, clade V has also been associated with nosocomial outbreaks (Ahmad and Alfouzan, 2021), and although clade II clusters can be sources of nosocomial spread, they are mainly associated with otomycoses [39]. Nonetheless, these factors all play an important function in clinical diagnostics.
3.2. Diagnostics and Detection
C. auris can often be misidentified using traditional biochemical methods, but the use of microbiology methods like plating clinical samples on chromogenic medium (CHROMAgar Candida, CHROMAgar Candida Plus) and blood cultures (BioFire’s BCID 2 and the GenMark Dx ePlex BCID-FP panels) is recommended to preserve fungal cell viability and rule out other Candida spp [40]. These methods are still undergoing refining, as they may still pose some challenges like cost, time-to-results, reliability, and protocol complexity, underscoring a need for affordable, rapid and reproducible high-value diagnostic tools, especially in regions where non-albicans Candida species (NCAC) are constantly reported [40,41].
Misdiagnoses are common with methods such as API 20C and VITEK2 YST ID, which can lead to inaccurate care plans for disease management [42]. Moreover, biochemical methods often require culturing yeast cells over several days, which can delay diagnosis and exacerbate disease progression [41]. Notably, the SENTRY Antifungal Surveillance Program reported no positive identification of C. auris between 1997 and 2016 from 135 clinical facilities in 39 countries [43]. Although C. auris belongs to the Candida/Clavispora clade like C. haemulonii, C. pseudohaemulonii, C. albicans and C. tropicalis, it has unique characteristics that clearly distinguish it genotypically and phenotypically from the others, prompting research and development initiatives of more sophisticated diagnostic tools, and optimization of currently available ones. The VITEK 2 YST ID card system is fast and dependable for the identification of yeasts and other yeast-like organisms. This is a fully automated system that employs fluorogenic methodologies and a series of carbohydrate fermentations, acid assimilations, and other biochemical reactions to determine yeast enzymatic activity. VITEK 2 can also yield results in 15h post-inoculation via fluorescence-based methods, compared to API 20C that requires 72h of incubation [44,45]. VITEK 2 does not, however, include ways to assess morphology, a characteristic that varies across species with and without dimorphism [44]. To avoid misidentification, further confirmation by microscopic morphological evaluation or agglutination tests may be necessary [44]. Although both systems are sufficiently reliable when discussing the identification of common pathogenic species, they are typically costly, time-consuming, intricate, and encounter setbacks when rare pathogens require detection [42].
Alternatively, the use of molecular methods such as rDNA and ITS sequencing, matrix assisted light desorption/ionization time-of-flight (MALDI TOF) mass spectrometry, and PCR/RT-PCR may bridge the diagnostic gap in surveillance studies [40,41]. In fact, DNA metagenomic and metabarcoding methods based on ITS regions have been used to identify potential anthropogenic and ecological niches of C. auris, yielding partial matches in available metagenomic and metabarcoding databases. Some results include the possible presence of C. auris in Floridian peanut fields, Kuwaiti air dust, South Korean activated sludge, and even animal reservoirs like Spanish dogs, suggesting a plethora of fungal inoculum intermediates via which C. auris could have been passaged to humans [6]. The ITS regions may provide more specificity with a lower margin of error than D1/D2 domain sequencing in certain instances, particularly when misidentification is highly probable. Although D1/D2 domains are the procedural approach and can be sequenced in almost all yeasts, reference ITS region sequences are available in public databases that can be used to compare sequences between pathogenic and non-pathogenic species for more reliable results [46]. In a study where ribosomal ITS1 and ITS2 regions were thoroughly sequenced in 373 clinically important yeasts, positive agreement rates of 96.8% and 99.7% were shown, respectively. ITS2 regions may be a more desirable molecular target when identifying pathogenic strains such as C. auris and could potentially standardized for clinical laboratory testing purposes [47]. With recent advances in bioinformatics and automation, culture-independent RT-PCR methods such as AurisID and FungiPlex Candida auris have yielded promising results and shown greater advantage than conventional methods [34]. Moreover, promising developments have been made with Raman spectroscopy and machine learning tools to identify C. auris, differentiate clades, and predict antifungal susceptibility profiles [48,49]. Together, diagnostic screening methods are critically important in supporting infection prevention and control teams.
The frequency of community transmission is difficult to estimate because C. auris can colonize and persist at multiple body sites in immunocompetent hosts without causing visible symptoms, thereby excluding them from high-IFI risk groups [50]. As C. auris spreads easily in healthcare settings, the CDC recommends screening for colonization in patients with high-risk healthcare exposures [51]. It is imperative to perform comprehensive screening studies by obtaining swabs from multiple body sites across immunocompetent and immunocompromised hosts at different settings to contribute to containment and mitigation efforts against C. auris [52]. It may also prove important to screen the near patient environment where dry biofilms are known to exist [53]. Many researchers have stressed the importance of a collective contribution between clinical, laboratory, public health, and government entities globally to reduce nosocomial spread [41]. By implementing commercial methods of screening and surveillance at the localized level, accurate surveying and reporting of positive symptomatic and asymptomatic cases can be assured. A recent study from Baltimore detected C. auris in 11.81% of wastewater influent samples by qPCR and reported a weak correlation with new cases [22].
4. Pathogenicity
Candida spp. is comprised of over 200 species that have been taxonomically tallied for decades, with only a handful of medically important fungi [54]. The CDC reported that 95% of invasive candidiasis cases are attributable to five species [55]. C. albicans remains responsible for the majority of candidiasis-related IFIs, both of mucocutaneous and systemic nature, in healthcare settings, as well as the third-to-fourth most reported infection across hospitals worldwide [56]. It is closely followed by Candida glabrata, C. parapsilosis, C. tropicalis, and C. kruseii [57]. Recently, however, trends have shifted to NCAC-related infections, including those caused by the novel C. auris. Comparative genomics identified several genes linked to drug resistance and virulence in this emerging species as compared to other Candida species, including expanded families of transporters and lipases, and this may contribute to differences that enhance C. auris virulence and pathogenicity. The following subsections highlight some of the main virulence factors contributing to the pathogenesis of this emergent pathogenic yeast.
4.1. Morphogenetic Switching
C. auris differs from C. albicans and other dimorphic fungi in that it largely exists as a yeast. In response to atypical or unfavorable growth conditions, certain C. auris isolates may form pseudohyphae-like and/or even hyphae-resembling structures [34]. After passage through a mammalian host, C. auris exhibited a unique phenotypic switch characterized by heritable and nonheritable changes between filamentous-form, yeast cells, and filamentous-competent cells. Interestingly, some orthologs of C. albicans needed for attachment and filamentation, like ALS4 and HGC1 were differentially expressed in the filamentous morphology of C. auris [26]. Notably, genomic amplification of ALS4 has been linked with enhanced cell-cell adherence in C. auris clinical isolates exhibiting aggregative phenotypes and robust biofilm formation [58]. EFG1 and WOR1, which are upregulated during C. albicans white-to-opaque transitions appear to be downregulated in C. auris filamentous cells. Moreover, C. auris filamentous cells differentially express orthologs of HGC1 and ALS4, involved in filamentation and adhesion respectively, whereas factors like NRG1 and CUP9 (negative regulators of filamentation) are overexpressed in C. auris yeast cells [26]. Genotoxic stress induced by treatment with hydroxyurea, methyl methanesulfonate, and 5-fluorocytosine, is able to trigger pseudohyphal morphogenesis in C. auris Δtup1 strains, a transcription factor belonging to the family of negative hyphal growth regulators in C. auris. Altogether, these findings indicate that, although C. auris shares many evolutionarily conserved alleles with other resistant Candida, it has diverged in some capacity due to stress and environmental selection pressure. Genome-wide association studies are practical tools that continue to provide insight into C. auris copy number variants, conserved and expanded gene families and how these may impact virulence, host-pathogen interactions, and persistence [25].
C. auris has also been found to include strains characterized by clumping, or aggregation, and non-aggregation phenotypes that seem to have a relationship with virulence. Using a Galleria mellonella model, they found that the virulence exhibited by a non-aggregating phenotype of C. auris was similar to that of C. albicans and C. tropicalis. The larvae were inoculated with several virulent Candida strains, incubated at 37 °C, and dissected 18 hours post-infection to reveal no C. auris hyphal or pseudohyphal formation compared to C. albicans. However, they did find that non-aggregate strains of C. auris resulted in similar larval killing rates to those of C. albicans when incubated at 37° C, while aggregate strains had less virulence [59]. It is important to note that aggregate strains were difficult to eradicate in vitro, and regardless of larval inoculation with single yeast cells obtained from aggregates, the hemolymph form larvae exhibited aggregation post-infection. This suggests that certain C. auris strains can maintain aggregation both in vivo and in vitro in physiologically unfavorable conditions but this does not necessarily correlate with increased virulence [59].
Despite the finding that aggregative strains are less virulent than non-aggregative strains, the former may still be associated with increased fungal burden and tissue colonization [60]. In a disseminated candidiasis mouse model, where virulence between C. haelumonii and C. auris was compared, immunosuppressed female BALB/c mice were challenged with both species and C. albicans. Results indicated that, despite administration of cyclophosphamide prophylactically and on infection day, all mice infected with C. haelumonii were asymptomatic and survived even 12 days post-infection. On the other hand, C. auris infected mice exhibited a 20% survival rate 5 days after inoculation but disseminated C. auris was not more virulent than C. albicans (10% survival rate by day 3 post-infection). Kidney fungal load showed a correlation between fungal burden and survival rate, which was species-specific. C. haelumonii cells were not detected in culture, while C. auris and C. albicans CFUs were 5.9 x 104 and 7.1 x 105 respectively. Lastly, C. auris aggregates were ubiquitous in kidney histopathology samples, which may denote aggregate morphology as an immune evasion mechanism [61].
4.2. Colony Phenotypic Switching
Besides resorting to morphological switching as an immunopathogenic and virulent tactic, C. auris can also undergo colony phenotypic changes [5]. In C. albicans the white-to-opaque switch plays distinct roles in its reproduction cycle, pathogenicity, and response to environmental stressors [62,63,64]. For instance, C. albicans was believed to be strictly asexual but following genome sequencing, its mating-type locus was further expounded when compared to the mating-type locus homologs of S. cerevisiae. This comparison led to the discovery of a/α heterozygosity present in the majority of C. albicans strains. Though homozygous strains (e.g., a/a and α/α) are able to mate, they do so at a much lower frequency than heterozygous strains. The mating-type locus also in part induces phenotypic switching from white to opaque, where opaque cells have a higher affinity for pheromone signaling and mating than white cells [62]. The white-to-opaque switching in C. albicans also plays a role in immune evasion, which contributes to its adaptability in changing environments, immunogenicity and antigenicity. Interestingly, the master regulator Wor1 plays a pivotal role in C. albicans colony phenotypic switching [63,64]. Several phase-specific genes are transcribed during phenotypic switching in C. albicans, and cell phenotype can dictate which genes are differentially expressed. For instance, white cells exclusively express WH11, whereas opaque cells express PEP1 which encodes secreted aspartyl proteases that catalyze protein degradation on host cells, potentially conferring them immunopathogenic roles.
Like C. albicans, colony phenotypic switching has also been documented in C. auris [5,34]. In a study aimed at improving classification of C. auris clinical isolates, C. auris was found to express pink, dark purple, and white colony phenotypes. Yeast cell size and shape remain invariable, and textures are smooth and glossy. In the same study, three strains across the four main C. auris clades were cultured in Salt Sabourad Dulcitol enriched broth and plated on CHROMagar Candida to investigate ability to undergo phenotypic switching, and to determine inter- and intraspecies differences in the latter. Pink colonies were predominant among all clades, but clades I, II and IV did show increased switching to dark purple colonies. Members of clades I, III and IV are commonly associated with nosocomial outbreaks, while clade II strains are mainly associated with superficial colonization and infection. Interestingly, clade III exhibited the least amount of phenotypic switching, while clade IV had the most. When tested for switching frequency, the clade IV strain that showed the greatest rate of phenotypic switching (B11245) appeared to switch between dark purple and pink more preferentially than between pink and white colonies. It is of interest that strain B11245 exhibited greater bias toward white colonies given its increased ability to proliferate while in this phenotype. Seemingly, this predilection, which induces propagation in C. auris B11245, may be homologous to the increased mating affinity opaque cells in C. albicans showcase upon switching [65]. On improved media such as CHROMagar Candida Plus, C. auris appears white with a surrounding blue halo. The medium also has improved sensitivity to multiple isolates, irrespective of clade or mutations, when compared against CHROMagar Candida [66]. This may suggest that phenotypic switching in C. auris is an inherited adaptive trait and could increase growth, survival, and adaptability.
Interestingly, colony phenotypic switching has been shown to correlate with ploidy plasticity in C. auris, particularly between pink and white colonies [67]. Cells within white colonies were smaller and had haploid genomes, whereas those in pink colonies tended to be larger in both size and genome (diploid). Transcriptomic analysis demonstrated differentially expressed genes between diploid and haploid types, with haploid cells having a more active TCA cycle and oxidative phosphorylation metabolism than diploid cells. In addition, ploidy plasticity appears to correlate with other virulence factors such as thermotolerance, antifungal resistance, disease, and cell membrane permeability [67]. Overall, such factors may increase fitness and allow C. auris to quickly adapt to different niches, ultimately increasing transmissibility.
Still, the community consensus remains that C. auris is haploid and sexual reproduction through nuclear fusion and meiotic mechanisms has not been recorded. In Munoz et al. study, evidence supported the presence of both mating type-like (MTL) loci within all 5 clades in a clade specific and homozygous manner [68]. Clades I, IV and V are strictly MTLa while clades II and III are MTLα. For sexual reproduction to occur, it is a requirement that strains with opposite idiomorphs come in contact. In C. auris, strains with differing mating loci have not been found within the same clade, but strains belonging to different clades and hence opposing mating loci have been found in different geographic locations [68]. With these findings and Casadevall’s hypothesis on interspecies horizontal gene transfer [9], sexual reproduction in C. auris may not be impossible. Such mechanisms would lead to increased genetic recombination and positive selection of diverse antifungal resistance genes.
4.3. Contextualizing The Damage Response Framework
The damage response framework originally conceived by Casadevall and Pirofski aimed at unifying host- and microorganism-centered views on pathogenesis. It proposes that pathogenicity cannot be attributed to either host or microorganism alone, but rather it results from interactions between them [69]. Moreover, host damage will dictate the degree of pathogenesis, and this damage could result from microbial metabolites, host responses, or both. The outcomes of these relationships are seldom simple to explain, as there is interplay between host and microbial factors that may exacerbate or attenuate benefit and/or damage for one or the other, both, or for neither. C. auris can be classified using this model, but because of its ability to colonize the skin irrespective of symptomology, it can be proposed that C. auris has the attributes of a commensal [70]. However, asymptomatic colonization by C. auris drives nosocomial outbreaks in hosts with weak immune systems, which can often manifest as life-threatening invasive infections. Alternatively, ear infections may occur in otherwise immunocompetent hosts, underscoring the importance of characterizing exogenous and endogenous factors that may fuel the commensal-to-pathogen transition in C. auris. The biology of C. auris can provide insight into this preferential colonization. Thermotolerance and osmotolerance are primary virulence factors that could be repurposed for adaptability on host skin. Moreover, C. auris can grow on biotic and abiotic surfaces, where it can persist for weeks, and has a predilection for colonizing cooler surfaces [52]. Therefore, C. auris could be a chronic colonizer of human skin, where it may micro-diversify and adapt to already present ecological consortia, establish mutualistic or commensal polymicrobial interactions with other fungi or bacteria on the skin, and trigger mild to modest immune responses that allow it to remain in homeostasis, much like Malassezia spp. The skin is also a source of lipids, lipid-producing enzymes, and carbohydrates that can ease the onerous task of nutrient scavenging in unfavorable environments [71]. It may be possible that in a comfortable niche constantly exposed to environmental factors such as the skin, C. auris could undergo transcriptional and epigenetic changes that enable optimal adherence, metabolic shifts, and survival. Fungi also grow well at varying pH levels, and given that the skin’s pH is generally acidic, C. auris may adapt and grow well under these conditions while other possible C. auris predators may be unwelcome [72].
Beyond the skin barrier, C. auris has the capacity to invade and become systemically important [73]. In an in vivo murine model of disseminated candidiasis, it was reported that C. auris is less virulent than C. albicans. After challenging immunocompetent C57BL/6J mice with the same colony-forming unit (CFUs) loads intravenously for both C. albicans and C. auris, survival rates were lower for C. albicans infection compared to C. auris. Likewise, they demonstrated decreased fungal burden in the kidneys of C. auris-infected mice after 7 days compared to organs infected with C. albicans. Importantly, results correlated with better immunomodulation and antifungal defense against C. auris in immunocompetent hosts. This was demonstrated by C. auris’s ability to induce a more potent immune response upon stimulation with patient-derived peripheral blood mononuclear cells (PBMCs). After 24 hours, PBMC exposure to C. auris resulted in broader transcriptional responses than those of C. albicans, where several unique C. auris differentially expressed genes correlated to enriched host responses, placing emphasis on cytokine production and certain interferon responses [74]. Furthermore, myeloperoxidase production in immunocompetent mice following C. auris and C. albicans infection was markedly robust, contradicting prior findings of impaired human neutrophil recruitment and neutrophil extracellular trap formation against C. auris relative to C. albicans [75]. Despite these observations, C. auris is more likely environmentally adapted, and should be considered an accidental pathogen. This is why it can persist, colonize skin, and survive within the healthcare environment so effectively.
4.4. Skin Colonization and Persistence
The persistence of C. auris on patients’ skin poses a major concern to all healthcare professionals. Unlike other Candida species that primarily inhabit mucosal membranes, C. auris shows a pronounced adhesion to and a predominant tendency for skin colonization [6,34]. Its prolific capacity for skin colonization and persistence accelerates its dissemination across the healthcare setting, as skin cells are constantly shed from infected patients, creating reservoirs for further transmission [72,76].
C. auris has been found to enrich populations of certain resident skin cells like type 3 innate lymphoid cells, ᵧδ cells, and IL17A, IL17F Th17 and Tc17 cells upon skin colonization. In fact, it preferentially lodges within deep skin tissue sections, where glands and hair follicles are found (Figure 1). This may be another adaptive mechanism for immune evasion, as host immune effector cells cannot readily reach those areas, concomitant with nutrient scavenging [77]. Colonized patients often exhibit no symptoms and can remain positive for long periods, with one surveillance study finding the median time for patients to become serially negative from an initial positive screen was 8.6 months (IQR 5.7–10.8 months) [76,78]. Further research found that C. auris remained within the follicles and sebaceous glands of murine skin months after skin surface screening was negative [79]. This explains how C. auris can proliferate on patient skin despite frequent chlorhexidine gluconate (CHG) washing, as the compound lacks permeation into the follicles and glands where the pathogen resides [76,80]. Recent studies have however shown that augmenting CHG with isopropyl alcohol or certain essential oils greatly improved decolonization in vitro and ex vivo [81,82].
Several factors give C. auris an advantage over other Candida species when colonizing the skin. The adaptation of several species-specific adhesion factors may play a prominent role in biofilm formation, skin colonization and virulence. The C. auris specific Surface Colonization Factor (SCF1) differs from the majority of fungal adhesins by utilizing cation-substrate interactions that enhances long term skin adherence and biofilm formation [83,84]. The identification of a C. auris specific IFF homolog (IFF4109), which is involved in skin surface adhesion and virulence, was also an important discovery [15]. In addition, the ALS homolog (ALS4112) has been shown to convey robust adherence to host ECM proteins and keratinocytes in vivo [85,86]. Further investigation using an in vivo murine model found that deletions of the adhesin genes SCF1 and ALS4112 significantly affected cell-cell adherence and biofilm formation depending on the aggregative growth phenotype [86]. This functional diversity of cell wall adhesins provides insight into how C. auris strains have adapted to effectively colonize and persist on the skin surface. A recent study found that Hog1 mitogen-activated protein kinase is essential for regulating cell wall organization and expression of cell adhesins, including IFF4109, ALS5 and a multitude of other uncharacterized adhesins involved in biofilm formation and skin adherence in vitro and ex vivo [87]. The nature of C. auris-host interactions is complex and understudied, with a multitude of skin specific adhesion factors yet to be characterized [84,87]. Furthermore, C. auris has specific immune evading properties and interactions with commensal skin microbiota that remain poorly understood [70]. Low neutrophil recruitment and a lack of IL-17A and IL-17F signaling by innate and adaptive lymphocytes have been associated with long-term C. auris skin colonization and persistence in immunocompromised individuals [75,79]. There is also evidence to support the dysbiosis of the skin microbiota involvement with C. auris colonization and persistence, with microbiome dysbiosis a common factor described among C. auris positive patients [88]. Microbiome dysbiosis is inevitable in patients who are given frequent CHG washes, possibly exacerbating C. auris persistence in immunocompromised patients [79]. Herein lies the value of in vivo and ex vivo studies, which help elucidate the mechanisms underlying skin persistence and inform the formulation of long-term decolonization strategies.
Current data highlights the widespread use of CHG in concentrations of 2-4% used in wipes and baths as a primary measure against C. auris colonization [79,81,89]. However, there is concern over its lack of permeation into the skin which is correlated to a lower reduction of fungal load and long-term persistence [82]. Using an ex vivo porcine model, it was shown that the efficacy of CHG/isopropanol is improved when combined with natural antiseptics like tea tree and lemongrass oil for decolonization [82]. Follow up studies are necessary to investigate the efficacy of improved CHG formulations in humans. Other natural antiseptics like manuka honey, eucalyptus oil and thymol have also shown promise in reducing C. auris fungal burden while synthetic compounds containing povidone iodine, octenidine and chlorine are most effective in vitro [90,91]. Additionally, research into liposomal technology for improved delivery of novel and repurposed antifungals may provide a future breakthrough into effective C. auris decolonization and treatment [92,93]. Despite these experimental breakthroughs there is currently no approved strategy for decolonization of infected patients, meaning that without active screening programs, colonized individuals may inadvertently spread across sensitive hospital areas. To address this problem, understanding of the factors regulating C. auris skin colonization and persistence in vivo is vital for developing effective decolonization strategies for infected patients.
4.5. Biofilm Lifestyle: Impact on Pathogenesis and Clinical Repercussions
Observations from ex vivo studies and animal models have demonstrated that C. auris can form biofilms in synthetic sweat media and persist in nutrient-poor, high salinity skin conditions [16,94]. Ongoing research suggests that C. auris employs alternative carbon metabolism enabling the use of sweat and sebum components as nutrient sources [71,95,96]. High-throughput profiling identified some unique genes from the C. auris clades enabling robust growth on alternative carbon sources and dipeptides as a nitrogen source [95]. These C. auris specific metabolic genes may explain its robust growth and persistence observed in the axilla and groin of patients [78]. It was also demonstrated that the presence of artificial sweat and sebum enhanced C. auris biofilm formation on porcine skin [16]. The findings showed that the fungal burden of C. auris was 10 times greater than C. albicans, indicating that C. auris can thrive as biofilms in high sweat body sites which correlates with clinical data [16,72,78].
C. auris is much like other Candida spp. by virtue of its ability to persist as a biofilm, much like other medically relevant fungal species. Biofilms are characterized by a dense consortia of adherent sessile cells that can form tightly woven, heterogeneous communities of multi-morphic cells embedded in an extracellular matrix (ECM). This enables them to adhere to biotic and abiotic surfaces [56,97]. As such, these highly organized microniches further complicate systemic, dermal and mucosal infections. In the case of yeasts like C. albicans, biofilm formation is a highly regulated process which takes place over 4 major steps: adherence, proliferation, maturation and dispersal [98]. Biofilm biogenesis relies on nutrient availability, sensory mechanisms, positive and negative transcriptional regulation, and divergent gene expression profiles that can induce metabolic and adaptive changes optimal for nutrient acquisition and survival [97]. Figure 2 illustrates the key biofilm development phases and accompanying transcriptional changes.
Albeit C. auris biofilms are substantially weaker than those of C. albicans, they have a persistent ability to adhere to both skin and abiotic surfaces [99]. Phenotypically, C. auris biofilms differ in that they do not contain true hyphae, but rather clumpy aggregates or single yeast cells with occasional pseudohyphae [5,27,28,34,100]. Biofilm spatial architecture largely depends on whether the strain is aggregate or non-aggregate. Vila et al. showcased fluorescent confocal images of C. auris isolates 0382 and 0387 belonging to the CDC AR-Bank. Isolate 0382 is classified as a high-biofilm former as opposed to isolate 0387, with the former exhibiting a dense and homogenous multilayer, while the latter formed a dispersed biofilm with scattered cell aggregates. Both presentations were synonymous to aggregate and non-aggregate C. auris phenotypes respectively [101]. Interestingly, both biofilm subtypes are associated with an increased secretion of exopolymeric materials into the biofilm matrix, an observation not supported from studies on a silicone elastomer [27]. Vila et al. also designed an oral infection mouse model in which C. auris did not colonize the tongue dorsum of infected mice, while C. albicans formed robust biofilms with hyphae penetrating and damaging epithelial tissue [101]. These observations are consistent with the findings that histatin 5, a cationic peptide found in saliva, has potent inhibitory activity even against FLU-resistant C. auris isolates, which may explain its inhibited growth in the oral flora [102].
Transcriptional differences exist between C. albicans and C. auris biofilms, which define their predilections for colonizing tissue in vivo and ex vivo and attaching to certain surfaces. Such heterogeneity may influence mechanisms of drug tolerance and sensitivity. Several virulence genes that encode glycophosphatidylinositol (GPI)-linked cell wall adhesins, integrins and agglutinin-like sequences become upregulated during biofilm formation and maturation in C. auris. Interestingly, seven of those upregulated GPI-anchored proteins (IFF4, PLB3, PGA52, PGA26, CSA1, HYR3, PLB3, and PGA7) are evolutionarily conserved between C. auris and its phylogenetic relatives C. haelumonii, C. pseudohaelumonii, and C. duobushaemuloni [30,34,68]. Transcriptomic analysis of in vitro grown C. auris biofilms revealed the presence of ALS1 and ALS5 adhesins, but not ALS3, a key adherence factor in C. albicans involved in host cell endocytosis [103]. Other putative virulence factors like lytic enzymes, which are critical for morphogenesis, adherence and invasion, such as hydrolases, hemolysin, secreted aspartic proteinases (SAPs) and phospholipases have also been documented in C. auris [26,28,30,68]. Different phenotypic switches induced by temperature in C. auris are also correlated to changes in SAP activity [100]. Together, these data indicate that the biofilm lifestyle is of critical importance to C. auris, and this has significant implications for its clinical management due to the tolerant phenotype associated with biofilms.
5. Antifungal Drug Resistance
C. auris has been called a “superbug” due to its synchronous geographical appearance, territorial distribution, tendency to cause hospital outbreaks, and quick multi-drug resistance evolution. All of which reinforces the global threat it represents. Its pan-resistance can be traced back to its phylogenetic relationships with other highly resistant NCAC species. Population genomics analyses found that four strains belonging to Clade I exhibited resistance against all antifungal classes used to treat systemic infections, underscoring the need to devise strategies that will mitigate resistant phenotypes [38].
Antifungal resistance in pathogenic fungi is a complex, multifactorial phenomenon that cannot be attributed to a single specific factor. Intrinsic resistance, stemming from an absence of drug pressure, is exemplified by the fact that 90% of global C. auris isolates exhibit intrinsic resistance to fluconazole [34]. This innate resistance commonly reflects specific physiological traits, including decreased drug binding due to target alteration, upregulation of efflux pumps or altered drug permeability rates [104]. C. auris tolerance and resistance mechanisms to the three main antifungal classes will be summarized:
5.1. Azoles
C. auris exhibits an intrinsic resistance to triazoles, likely due to its phylogenetic relationship to other intrinsically resistant species coupled with extensive azole use in fields and clinics [6,34]. Azoles target lanosterol-14-α-demethylase, an important enzyme in the ergosterol synthesis pathway encoded by ERG11. Mechanisms of triazole desensitization are exemplified in three categories: overexpression of the target enzyme, mutations in ERG11 that may alter the affinity of azoles for their target and increased efflux pump activity [5,104]. Fluconazole seems to be the main target for resistance and tolerance, given that it is the most frequently administered and readily available antifungal in an array of conditions. Prophylactic and empiric administration may lead to drug-induced selection pressure, which can open the floor to hypermutation events, loss of heterozygosity, and increased mutation rates conferring resistance or tolerance (e.g., point mutations, amino acid substitutions, insertions, etc.) [106,110]. For instance, Candida spp. have been found to possess hotspot mutations in the ERG11 alleles, namely amino acid substitutions in Y132F, K143R and F444L, all of which confer azole resistance. Because the C. auris genome has been sequenced, screening studies have found the same variants along ERG11 [6,104,105,106]. A study by Healey et al. found a 2-fold azole MIC increase upon insertion of Y132F or K143R substitutions in a Saccharomyces cerevisiae recombinant strain compared to a strain only expressing the wild-type C. auris ERG11 gene, suggesting decreased azole sensitivity in the presence of these allelic expressions [107].
Using a specialized Cas9-ribonucleoprotein transformation system, it was found that deleting the CDR1 homolog in C. auris, encoding an efflux pump, susceptibility to azoles largely increased in an azole-resistant strain up to 64 to 128-fold [108]. More recently it was shown that mutations in TAC1B, which encodes a zinc cluster transcription factor, conferred increased resistance to azoles via upregulation of CDR1 and MDR1 [109]. Multiple mechanisms of azole resistance related to physiological changes may also be present in C. auris, much like in other Candida spp. For instance, Hsp90 expresses the Heat Shock Protein 90 (HSP90), a chaperone protein that aids in stabilization of proteins during heat- and drug-induced stress, protein folding, and even heat-dependent morphogenetic changes in pathogenic C. albicans strains. In C. auris, blockage of Hsp90 using a doxycycline-repressible promoter resulted in abrogated azole tolerance, but strains possessing ERG11 mutations and a Hsp90 depletion remained unaffected. Depletion of the former seems to be responsible for filamentation in C. auris as well. The same group found that fluconazole resistance could also be mediated via CDR1 overexpression, independent of Hsp90 [23].
5.2. Polyenes
Resistance to polyenes, such as Amphotericin B (AMB) and nystatin, is often rare in almost all fungal species due to their fungicidal nature. Polyenes target the main component of the fungal cell membrane: ergosterol. The latter is not a byproduct of translation, thereby it is not considered a protein target [110]. Polyenes will form a complex with ergosterol present in the cell membrane leading to the creation of porous channels that will cause leakage of ions out of the cell, concentration gradient destabilization, and ultimately cell lysis [104,111]. If resistance does occur, it is often due to alterations in ergosterol biosynthesis in response to cell membrane stress that may lead to either upregulation of ERG genes or loss-of-function mutations that reduce membrane ergosterol levels. The exact mechanisms of resistance and tolerance to AMB in C. auris remain elusive, with some studies suggesting genetic expansion or intrinsic resistance, while others suggest metabolically transient pathways or transcriptome involvement. Worryingly, AMB resistance may also arise during treatment [6,104,105]. For instance, several studies have reported high AMB MICs across an array of C. auris isolates found in India, Russia, USA, and Colombia. The consensus is that C. auris AMB resistance ranges between 0-30%, though conservative studies suggest overestimation and a figure of 12% is accurate [112,113]. Since C. albicans notably exhibits amino acid substitutions in ERG2, ERG3, ERG5, ERG6 and ERG13 that can alter sterol composition in response to polyenes, it is hypothesized that C. auris may employ a similar mechanism, but further studies are required [105,107]. Interestingly, different genome wide studies found that C. auris intrinsically overexpresses ABC and MFS-type multidrug transporter gene orthologs, which code efflux pumps and contribute to virulence and resistance in C. albicans [106]. To further complicate matters, definitive clinical breakpoints have not been established by the Clinical Laboratory Standards Institute (CLSI) or the European Committee on Antimicrobial Susceptibility Testing (EUCAST) given the large genomic variability and hence susceptibility profiles among clades (Table 1). Although facsimile patterns were observed for fluconazole, no definitive parameters could be established for other antifungal classes [34,110].
5.3. Echinocandins
The mechanism of action (MOA) of echinocandins is focused on the uncompetitive inhibition of β-1,3-glucan synthase, the major enzyme involved in the synthesis of β-glucan, a major structural component of the fungal cell wall [114]. Glucan, along with chitin, intertwines with other major cell wall components like galactomannans and mannoproteins, conferring rigidity and protection to via cell wall biosynthesis while aiding in the preservation of cell membrane integrity [115]. Echinocandin resistance in C. auris is observed at lesser rates than those of azoles and polyenes, but there have been increasing reports where patients have become refractory to echinocandin therapy during treatment [106]. According to the CDC, less than 5% of C. auris USA isolates are echinocandin-resistant. Albeit rare, it is not uncommon to think that resistance rates may continue to rise as C. auris infections continue to be reported worldwide, especially in regions where individuals may have been previously treated with echinocandins [106,110]. In Candida spp., molecular methods have helped identify hotspots in FKS1 and FKS2 genes, termed HS1 and HS2, both of which encode the subunits of β-1,3-glucan synthase. In the case of C. auris, only FKS1 amino acid substitutions have been identified at S639F/Y/P positions, which also align with HS1 in C. albicans FKS1 [6,34,104]. S639F conferred echinocandin pan-resistance to 4 isolates in a study where 106 C. auris isolates were sequenced and tested for antifungal susceptibility [116]. Several other mutations in the FKS1 gene have been identified more recently associated with increased resistance to echinocandins [117]. These mutations can now be searched using a new benchmarking tool containing 100 characterized WGS strains [118].
Very few C. auris strains are intrinsically resistant, making echinocandiins first-line treatments in the event of a C. auris infection [5,34]. However, therapeutic failure is likely if echinocandin-resistant or pan-resistant C. auris isolates are encountered, much like in cases where C. albicans was exposed to echinocandins for prolonged periods during esophagitis [110]. Rising broad-spectrum resistance combined with fungal pathogens with intrinsic echinocandin resistance (e.g., C. neoformans) urges the need to expand the current antifungal armamentarium [119].
6. Future Therapeutic Directions
The first step in combating antifungal resistance and tolerance is to use methods to screen for and identify resistant strains and their associated determinants. Surveillance and epidemiological studies have been essential for characterizing microorganisms that are unresponsive to currently available therapies, and organizations such as CLSI and EUCAST have established standardized protocols for in vitro susceptibility testing, which are available directly from the CDC and EUCAST [120,121]. Methods include broth microdilution, disk diffusion, strip-based Etest, and azole agar screening. Though these reference methods are considered the gold standard in medical mycology for establishing clinical cut-offs, many breakpoints in understudied species are tentative or nonexistent, and these parameters need to be evaluated in all clinical isolates being tested where species-specific variability could posit reproducibility and sensitivity issues [5,34,42,110]. Additionally, breakpoints have only been established for a handful of commonly occurring dimorphic fungal pathogens (e.g, CLSI M27 A2 and M38 for yeasts and filamentous fungi) using only the main antifungal agents. Because of protocol variation, EUCAST and CLSI results are difficult to compare, and certain unculturable species cannot be tested [110]. Moreover, these standardized methods are limited to planktonic cell susceptibility, and many pathogenic fungi form biofilms [122]. More recently, novel automated methods such as the MALDI Biotyper antibiotic susceptibility test rapid assay (MBT ASTRA), based on MALDI-TOF mass spectrometry, and isothermal microcalorimetry (IMC) show promise as future susceptibility testing methods [123,124]. Nevertheless, MALDI-TOF MS requires highly skilled personnel for operation, and equipment costs may be too demanding for smaller facilities [41,42]. IMC may be a cost-effective alternative, as it has shown reliability when tested on biofilm and planktonic conditions in C. auris and compared against other available metabolic signature testing methods such as VITEK2 [124]. Limitations still linger, so further validation studies are required to determine the reliance of use in surveillance studies.
For C. auris, official clinical susceptibility cut-offs have not been established, but the CDC suggested tentative breakpoints based on common pathogenic Candida spp. and expert opinion for all major antifungals in drug-resistant strains [34]. The MIC breakpoints are summarized in Table 1. Given that >90% of C. auris strains are intrinsically resistant to fluconazole, which has the same MOA as other triazoles, MICs have not been established for second generation triazoles as susceptibility profiles vary across isolates [125].

Table 1.
C. auris tentative resistance breakpoints by CLSI*.
| Fluconazole | ≥32 mg/mL |
| Amphotericin B | ≥2 mg/mL |
| Caspofungin | ≥2 mg/mL |
| Micafungin | ≥4 mg/mL |
| Anidulafungin | ≥4 mg/mL |
Source: Centers for Disease Control and Prevention, National Center for Emerging and Zoonotic Infectious Diseases (NCEZID), Division of Foodborne, Waterborne, and Environmental Diseases (DFWED) [126]
Genes encoding fungal drug targets, and mutations conferring resistance can be identified using molecular methods such as sequencing, comparative genomics, RNAseq and RT-PCR. Although some of these techniques can be used in laboratory and clinical settings with minimal training due to their improvements in specificity, sensitivity and ease of use, DNA sequencing is still the preferred method to detect resistance, which limits its clinical use given the expenses associated with sequencing machinery [41,110]. Given that resistance and tolerance may have a polygenic and pleiotropic nature coupled with functional redundancy, developing more targeted methods together with the evolution of bioinformatics, techniques like spatial transcriptomics, in silico studies, nanopore sequencing and single cell RNAseq may be used to obtain a thorough outlook of drug-target interactions, tolerance and resistance pathways in the real space continuum, which could aid researchers in the drug discovery process.
High-throughput screening (HTS) of chemical libraries is an attractive method for the identifying compounds with antifungal activity and can be performed using in vivo and in vitro models coupled with robotics and software [127]. For instance, protocols have been developed for HTS against Candida biofilms using 96-well and 384-well microtiter plates [128,129]. The protocols are simple, reproducible, cost-effective, rapid and easy to implement across small and large laboratory settings. They are coupled with an XTT 2,3-Bis-(2-Methoxy-4-Nitro-5-Sulfophenyl)-2H-Tetrazolium-5-Carboxanilide reduction color-imetric assay, whereby water-soluble XTT is converted by actively respiring cells into a water-soluble, orange-colored formazan product [128,129,130]. Results can be read spectrophotometrically using a microtiter plate reader, whereby metabolically active sessile cell activity has a linear relationship to cell density and, subsequently, signals are detected from absorbance readings. HTS has also been used to identify compounds with repositioning potential. These compounds will generally have a high safety profile because they are already FDA-approved, as well as having well characterized mechanisms of action and ADMET properties. This makes drug repurposing an attractive solution to high attrition rates and time-consuming research and development facing the pharmaceutical industry [131].
Alternative strategies such as screening non-exhaustive natural repertoire of bioactive compounds, phenolic compounds, and even synthesizing silver nanoparticles with natural antioxidant and antivirulence properties have shown great promise and continue to be adopted globally in the war against multidrug resistance [7,104]. Combination therapy approaches have also proven successful, whereby classic antifungals have been administered in tandem for synergistic and/or additive effects. For example, the monoterpene carvacrol (CAR) was used in combination with AMB, fluconazole (FLU), nystatin (NYS) and caspofungin (CAS) in an in vitro screen against C. auris virulence factors using buccal epithelial cells. They found that CAR exhibited anti-adherence and anti-proteinase activities against C. auris at subclinical concentrations, without inducing pseudohyphal morphogenesis or penetration into epithelial cells. Furthermore, CAR-AMB and CAR-NYS combinations resulted in synergy across 28% of tested strains, of which 12% and 28% out of 25 total strains were resistant to both antifungals respectively. Additivity was also reported for 52% and 24% of strains in the CAR-FLU and CAR-CAS combinations, out of which 48% were FLU resistant and none were CAS resistant [132].
Prospective directions in antifungal therapy may include fungal vaccines and immunotherapy, which require an in-depth understanding of innate and adaptive responses in fungal immunity and careful safety considerations to avoid excessive reactogenicity and inflammatory dysregulation in immunocompromised hosts, sometimes referred to as cytokine storm [110]. Albeit no fungal vaccines have been licensed to date, several attempts have been carried out, with some such as the C. albicans-targeted NDV-3 making it to phase II clinical trials [133].
7. Conclusions and Outlook
C. auris has rapidly established itself as one of the most formidable fungal pathogens of the 21st century. Its remarkable adaptability, thermotolerance, osmotolerance, capacity to persist on biotic and abiotic surfaces and transmissibility have fueled its global emergence as a multidrug-resistant healthcare-associated threat. The convergence of virulence factors, antifungal resistance, and unique ecological versatility underscores the urgent need for continued surveillance, improved diagnostic tools, and coordinated infection control efforts. Understanding C. auris within the broader context of fungal pathogenesis highlights the intricate balance between microbial adaptation and host defense that determines disease outcomes.
Despite major advances in our understanding of C. auris biology and resistance mechanisms, significant knowledge gaps remain regarding its environmental reservoirs, transmission dynamics, and host immune interactions. The emergence of pan-resistant strains and the limited antifungal pipeline demand innovative approaches that combine drug repurposing, novel antifungal development, and immunotherapeutic strategies. Integrating high-throughput screening technologies with genomic and transcriptomic analyses will be instrumental in identifying new therapeutic targets and resistance determinants.
Ultimately, the story of C. auris serves as both a scientific and public health warning. Its rise illustrates how environmental change and global interconnectedness can converge to drive the emergence of new pathogens. Moving forward, a multidisciplinary approach that unites microbiology, immunology, ecology, and clinical medicine will be essential to mitigate the ongoing threat posed by C. auris and to strengthen preparedness against future fungal epidemics.
Funding
G.R. and R.K. are funded by NHS Assure Scotland (ENU129), and J.F.C. is funded by an IbioIC, Remora and Glasgow Caledonian University PhD funded studentship. Work in the J.L.R. laboratory is supported by NIH grant R33AI140823 from the National Institute of Allergy and Infectious Diseases. Additional support was provided by the Margaret Batts Tobin Foundation, San Antonio, TX.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results, and the content is solely the responsibility of the authors.
References
- Rodrigues, M.L.; Nosanchuk, J.D. Fungal Diseases as Neglected Pathogens: A Wake-up Call to Public Health Officials. PLOS Neglected Tropical Diseases 2020, 14, e0007964. [CrossRef]
- Clinical Alert to U.S. Healthcare Facilities - June 2016 | Candida Auris | Fungal Diseases | CDC Available online: https://archive.cdc.gov/www_cdc_gov/fungal/candida-auris/candida-auris-alert.html (accessed on 25 November 2025).
- Denning, D.W. Global Incidence and Mortality of Severe Fungal Disease. The Lancet Infectious Diseases 2024, 24, e428–e438. [CrossRef]
- Liu, F.; Hu, Z.-D.; Zhao, X.-M.; Zhao, W.-N.; Feng, Z.-X.; Yurkov, A.; Alwasel, S.; Boekhout, T.; Bensch, K.; Hui, F.-L.; et al. Phylogenomic Analysis of the Candida Auris- Candida Haemuli Clade and Related Taxa in the Metschnikowiaceae, and Proposal of Thirteen New Genera, Fifty-Five New Combinations and Nine New Species. Persoonia - Molecular Phylogeny and Evolution of Fungi 2024, 52, 22–43. [CrossRef]
- Du, H.; Bing, J.; Hu, T.; Ennis, C.L.; Nobile, C.J.; Huang, G. Candida Auris: Epidemiology, Biology, Antifungal Resistance, and Virulence. PLOS Pathogens 2020, 16, e1008921. [CrossRef]
- Chowdhary, A.; Jain, K.; Chauhan, N. Candida Auris Genetics and Emergence. Annual Review of Microbiology 2023, 77, 583–602. [CrossRef]
- Vazquez-Munoz, R.; Lopez, F.D.; Lopez-Ribot, J. Silver Nanoantibiotics Display Strong Antifungal Activity against the Emergent Multidrug-Resistant yeastCandida Aurisunder Both Planktonic and Biofilm Growing Conditions. 2020. [CrossRef]
- WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action Available online: https://www.who.int/publications/i/item/9789240060241 (accessed on 25 November 2025).
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the Emergence of Candida Auris: Climate Change, Azoles, Swamps, and Birds. mBio 2019, 10. [CrossRef]
- Egger, N.B.; Hoenigl, M.; Sprute, R.; Arastehfar, A.; Cornely, O.A.; Krause, R.; Lass-Flörl, C.; Prattes, J. The Rise of Candida Auris: From Unique Traits to Co-Infection Potential. Microbial Cell 2022, 9, 141–144.
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida Aurissp. Nov., a Novel Ascomycetous Yeast Isolated from the External Ear Canal of an Inpatient in a Japanese Hospital. Microbiology and Immunology 2009, 53, 41–44. [CrossRef]
- Jones, C.R.; Neill, C.; Borman, A.M.; Budd, E.L.; Cummins, M.; Fry, C.; Guy, R.L.; Jeffery, K.; Johnson, E.M.; Manuel, R.; et al. The Laboratory Investigation, Management, and Infection Prevention and Control of Candida Auris: A Narrative Review to Inform the 2024 National Guidance Update in England: This Article Is Part of the Candida Collection. Journal of Medical Microbiology 2024, 73. [CrossRef]
- Salmanton-García, J.; Nóbrega De Almeida, J.; Colombo, A.L. Candidozyma Auris (Formerly Candida Auris): Resistant, Long Lasting, and Everywhere. Clinical Microbiology and Infection 2026, S1198743X25006330. [CrossRef]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. Environmental Candida Auris and the Global Warming Emergence Hypothesis. mBio 2021, 12. [CrossRef]
- Santana, D.J.; Anku, J.A.E.; Zhao, G.; Zarnowski, R.; Johnson, C.J.; Hautau, H.; Visser, N.D.; Ibrahim, A.S.; Andes, D.; Nett, J.E.; et al. A Candida Auris–Specific Adhesin, Scf1, Governs Surface Association, Colonization, and Virulence. Science 2023, 381, 1461–1467. [CrossRef]
- Horton, M.V.; Johnson, C.J.; Kernien, J.F.; Patel, T.D.; Lam, B.C.; Cheong, J.Z.A.; Meudt, J.J.; Shanmuganayagam, D.; Kalan, L.R.; Nett, J.E. Candida Auris Forms High-Burden Biofilms in Skin Niche Conditions and on Porcine Skin. mSphere 2020, 5, e00910-19. [CrossRef]
- Day, A.M.; McNiff, M.M.; Da Silva Dantas, A.; Gow, N.A.R.; Quinn, J. Hog1 Regulates Stress Tolerance and Virulence in the Emerging Fungal Pathogen Candida Auris. mSphere 2018, 3, e00506-18. [CrossRef]
- Arora, P.; Singh, P.; Wang, Y.; Yadav, A.; Pawar, K.; Singh, A.; Padmavati, G.; Xu, J.; Chowdhary, A. Environmental Isolation of Candida Auris from the Coastal Wetlands of Andaman Islands, India. mBio 2021, 12. [CrossRef]
- Escandón, P. Novel Environmental Niches for Candida Auris: Isolation from a Coastal Habitat in Colombia. Journal of Fungi 2022, 8, 748. [CrossRef]
- Silva, I.; Miranda, I.M.; Costa-de-Oliveira, S. Potential Environmental Reservoirs of Candida Auris: A Systematic Review. Journal of Fungi 2024, 10, 336. [CrossRef]
- Akinbobola, A.; Kean, R.; Quilliam, R.S. Plastic Pollution as a Novel Reservoir for the Environmental Survival of the Drug Resistant Fungal Pathogen Candida Auris. Marine Pollution Bulletin 2024, 198, 115841. [CrossRef]
- Nwaubani, D.A.; Baral, R.; Solomon, T.; Idris, O.; Sherchan, S.P. Wastewater Surveillance of Candida Auris in Baltimore. International Journal of Hygiene and Environmental Health 2025, 263, 114486. [CrossRef]
- Kim, S.H.; Iyer, K.R.; Pardeshi, L.; Muñoz, J.F.; Robbins, N.; Cuomo, C.A.; Wong, K.H.; Cowen, L.E. Erratum for Kim et al., “Genetic Analysis of Candida Auris Implicates Hsp90 in Morphogenesis and Azole Tolerance and Cdr1 in Azole Resistance.” mBio 2019, 10. [CrossRef]
- Sudbery, P.; Gow, N.; Berman, J. The Distinct Morphogenic States of Candida Albicans. Trends in Microbiology 2004, 12, 317–324. [CrossRef]
- Bravo Ruiz, G.; Ross, Z.K.; Gow, N.A.R.; Lorenz, A. Pseudohyphal Growth of the Emerging Pathogen Candida Auris Is Triggered by Genotoxic Stress through the S Phase Checkpoint. mSphere 2020, 5. [CrossRef]
- Yue, H.; Bing, J.; Zheng, Q.; Zhang, Y.; Hu, T.; Du, H.; Wang, H.; Huang, G. Filamentation in Candida Auris , an Emerging Fungal Pathogen of Humans: Passage through the Mammalian Body Induces a Heritable Phenotypic Switch. Emerging Microbes & Infections 2018, 7, 1–13. [CrossRef]
- Larkin, E.; Hager, C.; Chandra, J.; Mukherjee, P.K.; Retuerto, M.; Salem, I.; Long, L.; Isham, N.; Kovanda, L.; Borroto-Esoda, K.; et al. The Emerging Pathogen Candida Auris: Growth Phenotype, Virulence Factors, Activity of Antifungals, and Effect of SCY-078, a Novel Glucan Synthesis Inhibitor, on Growth Morphology and Biofilm Formation. Antimicrob Agents Chemother 2017, 61, e02396-16. [CrossRef]
- Rossato, L.; Colombo, A.L. Candida Auris: What Have We Learned About Its Mechanisms of Pathogenicity? Frontiers in Microbiology 2018, 9. [CrossRef]
- Ramage, G.; Sherry, L.; Kean, R. Microbe Profile: Candidozyma Auris: An Emergent and Resilient Yeast and New Antifungal Strategies: This Article Is Part of the Microbe Profiles Collection. Microbiology 2026, 172. [CrossRef]
- Chybowska, A.D.; Childers, D.S.; Farrer, R.A. Nine Things Genomics Can Tell Us About Candida Auris. Frontiers in Genetics 2020, 11. [CrossRef]
- Santos, M.A.S.; Gomes, A.C.; Santos, M.C.; Carreto, L.C.; Moura, G.R. The Genetic Code of the Fungal CTG Clade. Comptes Rendus. Biologies 2011, 334, 607–611. [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida Auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin Infect Dis 2017, 64, 134–140. [CrossRef]
- Sharma, M.; Chakrabarti, A. On the Origin of Candida Auris: Ancestor, Environmental Stresses, and Antiseptics. mBio 2020, 11. [CrossRef]
- Ahmad, S.; Alfouzan, W. Candida Auris: Epidemiology, Diagnosis, Pathogenesis, Antifungal Susceptibility, and Infection Control Measures to Combat the Spread of Infections in Healthcare Facilities. Microorganisms 2021, 9, 807. [CrossRef]
- Chow, N.A.; de Groot, T.; Badali, H.; Abastabar, M.; Chiller, T.M.; Meis, J.F. Potential Fifth Clade ofCandida Auris,Iran, 2018. Emerging Infectious Diseases 2019, 25, 1780–1781. [CrossRef]
- Khan, T.; Faysal, N.I.; Hossain, M.M.; Mah-E-Muneer, S.; Haider, A.; Moon, S.B.; Sen, D.; Ahmed, D.; Parnell, L.A.; Jubair, M.; et al. Emergence of the Novel Sixth Candida Auris Clade VI in Bangladesh. Microbiology Spectrum 2024, 12. [CrossRef]
- Lee, W.G.; Shin, J.H.; Uh, Y.; Kang, M.G.; Kim, S.H.; Park, K.H.; Jang, H.-C. First Three Reported Cases of Nosocomial Fungemia Caused by Candida Auris. Journal of Clinical Microbiology 2011, 49, 3139–3142. [CrossRef]
- Chow, N.A.; Muñoz, J.F.; Gade, L.; Berkow, E.L.; Li, X.; Welsh, R.M.; Forsberg, K.; Lockhart, S.R.; Adam, R.; Alanio, A.; et al. Tracing the Evolutionary History and Global Expansion of Candida Auris Using Population Genomic Analyses. mBio 2020, 11. [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida Auris: A Rapidly Emerging Cause of Hospital-Acquired Multidrug-Resistant Fungal Infections Globally. PLoS Pathog 2017, 13, e1006290. [CrossRef]
- Lockhart, S.R.; Lyman, M.M.; Sexton, D.J. Tools for Detecting a “Superbug”: Updates on Candida Auris Testing. Journal of Clinical Microbiology 2022, 60. [CrossRef]
- Kordalewska, M.; Perlin, D.S. Molecular Diagnostics in the Times of Surveillance for Candida Auris. Journal of Fungi 2019, 5, 77. [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Kord, M.; Roudbary, M.; Zarrinfar, H.; Fang, W.; Hashemi, S.J.; Najafzadeh, M.J.; Khodavaisy, S.; Pan, W.; et al. Comparison of 21-Plex PCR and API 20C AUX, MALDI-TOF MS, and rDNA Sequencing for a Wide Range of Clinically Isolated Yeast Species: Improved Identification by Combining 21-Plex PCR and API 20C AUX as an Alternative Strategy for Developing Countries. Frontiers in Cellular and Infection Microbiology 2019, 9. [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Turnidge, J.D.; Castanheira, M.; Jones, R.N. Twenty Years of the SENTRY Antifungal Surveillance Program: Results for Candida Species From 1997–2016. Open Forum Infectious Diseases 2019, 6, S79–S94. [CrossRef]
- Graf, B.; Adam, T.; Zill, E.; Göbel, U.B. Evaluation of the VITEK 2 System for Rapid Identification of Yeasts and Yeast-Like Organisms. Journal of Clinical Microbiology 2000, 38, 1782–1785. [CrossRef]
- Ramani, R.; Gromadzki, S.; Pincus, D.H.; Salkin, I.F.; Chaturvedi, V. Efficacy of API 20C and ID 32C Systems for Identification of Common and Rare Clinical Yeast Isolates. Journal of Clinical Microbiology 1998, 36, 3396–3398. [CrossRef]
- Hinrikson, H.P.; Hurst, S.F.; Lott, T.J.; Warnock, D.W.; Morrison, C.J. Assessment of Ribosomal Large-Subunit D1-D2, Internal Transcribed Spacer 1, and Internal Transcribed Spacer 2 Regions as Targets for Molecular Identification of Medically Important Aspergillus Species. Journal of Clinical Microbiology 2005, 43, 2092–2103. [CrossRef]
- Leaw, S.N.; Chang, H.C.; Sun, H.F.; Barton, R.; Bouchara, J.-P.; Chang, T.C. Identification of Medically Important Yeast Species by Sequence Analysis of the Internal Transcribed Spacer Regions. Journal of Clinical Microbiology 2006, 44, 693–699. [CrossRef]
- Pezzotti, G.; Kobara, M.; Nakaya, T.; Imamura, H.; Fujii, T.; Miyamoto, N.; Adachi, T.; Yamamoto, T.; Kanamura, N.; Ohgitani, E.; et al. Raman Metabolomics of Candida Auris Clades: Profiling and Barcode Identification. IJMS 2022, 23, 11736. [CrossRef]
- Xue, J.; Yue, H.; Lu, W.; Li, Y.; Huang, G.; Fu, Y.V. Application of Raman Spectroscopy and Machine Learning for Candida Auris Identification and Characterization. Appl Environ Microbiol 2024, 90, e01025-24. [CrossRef]
- Bradley, S.F. Candida Auris Infection. JAMA 2019, 322, 1526. [CrossRef]
- Baker, A.D.; Gold, J.A.W.; Forsberg, K.; Jones, S.; Lyman, M.M. Progression from Candida Auris Colonization Screening to Clinical Case Status, United States, 2016–2023. Emerging Infectious Diseases 2025, 31. [CrossRef]
- Ramage, G.; Kean, R.; MacKay, W.G.; Vaccaro, A.; Lopez-Ribot, J.L.; Howe, M.H. Candidozyma Auris: Implications for Infection Prevention and Control in the Management of This Resilient Biofilm Former. In Fungal Biofilms; Springer Nature Switzerland, 2025; pp. 165–192 ISBN 978-3-031-93315-8.
- Ware, A.; Johnston, W.; Delaney, C.; Butcher, M.C.; Ramage, G.; Price, L.; Butcher, J.; Kean, R. Dry Surface Biofilm Formation by Candida Auris Facilitates Persistence and Tolerance to Sodium Hypochlorite. APMIS 2025, 133, e70022. [CrossRef]
- Spampinato, C.; Leonardi, D. CandidaInfections, Causes, Targets, and Resistance Mechanisms: Traditional and Alternative Antifungal Agents. BioMed Research International 2013, 2013, 1–13. [CrossRef]
- CDC Data and Statistics on Candidemia Available online: https://www.cdc.gov/candidiasis/data-research/facts-stats/index.html (accessed on 25 November 2025).
- Wall, G.; Montelongo-Jauregui, D.; Vidal Bonifacio, B.; Lopez-Ribot, J.L.; Uppuluri, P. Candida Albicans Biofilm Growth and Dispersal: Contributions to Pathogenesis. Current Opinion in Microbiology 2019, 52, 1–6. [CrossRef]
- CDC Data and Statistics on Candidemia Available online: https://www.cdc.gov/candidiasis/data-research/facts-stats/index.html (accessed on 25 November 2025).
- Bing, J.; Guan, Z.; Zheng, T.; Zhang, Z.; Fan, S.; Ennis, C.L.; Nobile, C.J.; Huang, G. Clinical Isolates of Candida Auris with Enhanced Adherence and Biofilm Formation Due to Genomic Amplification of ALS4. PLoS Pathog 2023, 19, e1011239. [CrossRef]
- Borman, A.M.; Szekely, A.; Johnson, E.M. Comparative Pathogenicity of United Kingdom Isolates of the Emerging Pathogen Candida Auris and Other Key Pathogenic Candida Species. mSphere 2016, 1, e00189-16. [CrossRef]
- Brown, J.L.; Delaney, C.; Short, B.; Butcher, M.C.; McKloud, E.; Williams, C.; Kean, R.; Ramage, G. Candida Auris Phenotypic Heterogeneity Determines Pathogenicity In Vitro. mSphere 2020, 5. [CrossRef]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Maor, Y.; Tarabia, J.; Schechner, V.; Adler, A.; et al. Multidrug-Resistant Candida Haemulonii and C. Auris , Tel Aviv, Israel. Emerging Infectious Diseases 2017, 23. [CrossRef]
- Nielsen, K.; Heitman, J. Sex and Virulence of Human Pathogenic Fungi. In Fungal Genomics; Elsevier, 2007; pp. 143–173.
- Kvaal, C.; Lachke, S.A.; Srikantha, T.; Daniels, K.; McCoy, J.; Soll, D.R. Misexpression of the Opaque-Phase-Specific Gene PEP1 ( SAP1 ) in the White Phase of Candida Albicans Confers Increased Virulence in a Mouse Model of Cutaneous Infection. Infection and Immunity 1999, 67, 6652–6662. [CrossRef]
- Kvaal, C.A.; Srikantha, T.; Soll, D.R. Misexpression of the White-Phase-Specific Gene WH11 in the Opaque Phase of Candida Albicans Affects Switching and Virulence. Infection and Immunity 1997, 65, 4468–4475. [CrossRef]
- Bentz, M.L.; Sexton, D.J.; Welsh, R.M.; Litvintseva, A.P. Phenotypic Switching in Newly Emerged Multidrug-Resistant Pathogen Candida Auris. Medical Mycology 2018, 57, 636–638. [CrossRef]
- Mulet Bayona, J.V.; Salvador García, C.; Tormo Palop, N.; Gimeno Cardona, C. Evaluation of a Novel Chromogenic Medium for Candida Spp. Identification and Comparison with CHROMagarTM Candida for the Detection of Candida Auris in Surveillance Samples. Diagnostic Microbiology and Infectious Disease 2020, 98, 115168. [CrossRef]
- Fan, S.; Li, C.; Bing, J.; Huang, G.; Du, H. Discovery of the Diploid Form of the Emerging Fungal Pathogen Candida Auris. ACS Infectious Diseases 2020, 6, 2641–2646. [CrossRef]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic Insights into Multidrug-Resistance, Mating and Virulence in Candida Auris and Related Emerging Species. Nature Communications 2018, 9. [CrossRef]
- Casadevall, A.; Pirofski, L. The Damage-Response Framework of Microbial Pathogenesis. Nature Reviews Microbiology 2003, 1, 17–24. [CrossRef]
- Tharp, B.; Zheng, R.; Bryak, G.; Litvintseva, A.P.; Hayden, M.K.; Chowdhary, A.; Thangamani, S. Role of Microbiota in the Skin Colonization of Candida Auris. mSphere 2023, 8, e00623-22. [CrossRef]
- Swaney, M.H.; Nelsen, A.; Sandstrom, S.; Kalan, L.R. Sweat and Sebum Preferences of the Human Skin Microbiota. Microbiol Spectr 2026, 11, e04180-22. [CrossRef]
- Sexton, D.J.; Bentz, M.L.; Welsh, R.M.; Derado, G.; Furin, W.; Rose, L.J.; Noble-Wang, J.; Pacilli, M.; McPherson, T.D.; Black, S.; et al. Positive Correlation Between Candida Auris Skin-Colonization Burden and Environmental Contamination at a Ventilator-Capable Skilled Nursing Facility in Chicago. Clin Infect Dis 2021, 73, 1142–1148. [CrossRef]
- Kim, H.Y.; Nguyen, T.A.; Kidd, S.; Chambers, J.; Alastruey-Izquierdo, A.; Shin, J.-H.; Dao, A.; Forastiero, A.; Wahyuningsih, R.; Chakrabarti, A.; et al. Candida Auris —a Systematic Review to Inform the World Health Organization Fungal Priority Pathogens List. Medical Mycology 2024, 62, myae042. [CrossRef]
- Bruno, M.; Kersten, S.; Bain, J.M.; Jaeger, M.; Rosati, D.; Kruppa, M.D.; Lowman, D.W.; Rice, P.J.; Graves, B.; Ma, Z.; et al. Transcriptional and Functional Insights into the Host Immune Response against the Emerging Fungal Pathogen Candida Auris. Nature Microbiology 2020, 5, 1516–1531. [CrossRef]
- Johnson, C.J.; Davis, J.M.; Huttenlocher, A.; Kernien, J.F.; Nett, J.E. H. mBio 2018, 9, e01403-18. [CrossRef]
- Biswal, M.; Rudramurthy, S.M.; Jain, N.; Shamanth, A.S.; Sharma, D.; Jain, K.; Yaddanapudi, L.N.; Chakrabarti, A. Controlling a Possible Outbreak of Candida Auris Infection: Lessons Learnt from Multiple Interventions. Journal of Hospital Infection 2017, 97, 363–370. [CrossRef]
- Proctor, D.M.; Drummond, R.A.; Lionakis, M.S.; Segre, J.A. One Population, Multiple Lifestyles: Commensalism and Pathogenesis in the Human Mycobiome. Cell Host & Microbe 2023, 31, 539–553. [CrossRef]
- Bergeron, G.; Bloch, D.; Murray, K.; Kratz, M.; Parton, H.; Ackelsberg, J.; Antwi, M.; Del Rosso, P.; Dorsinville, M.; Kubinson, H.; et al. Candida Auris Colonization After Discharge to a Community Setting: New York City, 2017–2019. Open Forum Infect Dis 2020, 8, ofaa620. [CrossRef]
- Huang, X.; Hurabielle, C.; Drummond, R.A.; Bouladoux, N.; Desai, J.V.; Sim, C.K.; Belkaid, Y.; Lionakis, M.S.; Segre, J.A. Murine Model of Colonization with Fungal Pathogen Candida Auris to Explore Skin Tropism, Host Risk Factors and Therapeutic Strategies. Cell Host & Microbe 2021, 29, 210-221.e6. [CrossRef]
- Eyre, D.W.; Sheppard, A.E.; Madder, H.; Moir, I.; Moroney, R.; Quan, T.P.; Griffiths, D.; George, S.; Butcher, L.; Morgan, M.; et al. A Candida Auris Outbreak and Its Control in an Intensive Care Setting. New England Journal of Medicine 2018, 379, 1322–1331. [CrossRef]
- Moore, G.; Schelenz, S.; Borman, A.M.; Johnson, E.M.; Brown, C.S. Yeasticidal Activity of Chemical Disinfectants and Antiseptics against Candida Auris. Journal of Hospital Infection 2017, 97, 371–375. [CrossRef]
- Johnson, C.J.; Eix, E.F.; Lam, B.C.; Wartman, K.M.; Meudt, J.J.; Shanmuganayagam, D.; Nett, J.E. Augmenting the Activity of Chlorhexidine for Decolonization of Candida Auris from Porcine Skin. J Fungi (Basel) 2021, 7, 804. [CrossRef]
- Santana, D.J.; Anku, J.A.E.; Zhao, G.; Zarnowski, R.; Johnson, C.J.; Hautau, H.; Visser, N.D.; Ibrahim, A.S.; Andes, D.; Nett, J.E.; et al. A Candida Auris-Specific Adhesin, SCF1, Governs Surface Association, Colonization, and Virulence. Science 2023, 381, 1461–1467. [CrossRef]
- Wang, T.W.; Sofras, D.; Montelongo-Jauregui, D.; Paiva, T.O.; Carolus, H.; Dufrêne, Y.F.; Alfaifi, A.A.; McCracken, C.; Bruno, V.M.; Van Dijck, P.; et al. Functional Redundancy in Candida Auris Cell Surface Adhesins Crucial for Cell-Cell Interaction and Aggregation. Nat Commun 2024, 15, 9212. [CrossRef]
- Zhao, G.; Lyu, J.; Veniaminova, N.A.; Zarnowski, R.; Mattos, E.; Johnson, C.J.; Quintanilla, D.; Hautau, H.; Hold, L.A.; Xu, B.; et al. Adhesin Als4112 Promotes Candida Auris Skin Colonization through Interactions with Keratinocytes and Extracellular Matrix Proteins. Nat Commun 2025, 16, 5673. [CrossRef]
- Wang, T.W.; Sofras, D.; Montelongo-Jauregui, D.; Paiva, T.O.; Carolus, H.; Dufrêne, Y.F.; Alfaifi, A.A.; McCracken, C.; Bruno, V.M.; Van Dijck, P.; et al. Functional Redundancy in Candida Auris Cell Surface Adhesins Crucial for Cell-Cell Interaction and Aggregation. Nat Commun 2024, 15, 9212. [CrossRef]
- Shivarathri, R.; Chauhan, M.; Datta, A.; Das, D.; Karuli, A.; Jenull, S.; Kuchler, K.; Thangamani, S.; Chowdhary, A.; Desai, J.V.; et al. The Candida Auris Hog1 MAP Kinase Is Essential for the Colonization of Murine Skin and Intradermal Persistence. bioRxiv 2024, 2024.03.18.585572. [CrossRef]
- Huang, X.; Welsh, R.M.; Deming, C.; Proctor, D.M.; Thomas, P.J.; Gussin, G.M.; Huang, S.S.; Kong, H.H.; Bentz, M.L.; Vallabhaneni, S.; et al. Skin Metagenomic Sequence Analysis of Early Candida Auris Outbreaks in U.S. Nursing Homes. mSphere 2026, 6, e00287-21. [CrossRef]
- Kean, R.; McKloud, E.; Townsend, E.M.; Sherry, L.; Delaney, C.; Jones, B.L.; Williams, C.; Ramage, G. The Comparative Efficacy of Antiseptics against Candida Auris Biofilms. International Journal of Antimicrobial Agents 2018, 52, 673–677. [CrossRef]
- Cerbu, D.; Seiser, S.; Phan-Canh, T.; Moser, D.; Freystätter, C.; Matiasek, J.; Kuchler, K.; Elbe-Bürger, A. Octenidine Effectively Reduces Candida Auris Colonisation on Human Skin. Sci Rep 2025, 15, 27034. [CrossRef]
- Wu, W.-G.; Luk, K.S.; Hung, M.-F.; Tsang, W.-Y.; Lee, K.-P.; Lam, B.H.-S.; Cheng, K.-L.; Cheung, W.-S.; Tang, H.-L.; To, W.-K. Antifungal Efficacy of Natural Antiseptic Products against Candida Auris. Med Mycol 2024, 62. [CrossRef]
- Baldim, I.; Paziani, M.H.; Grizante Barião, P.H.; Kress, M.R. von Z.; Oliveira, W.P. Nanostructured Lipid Carriers Loaded with Lippia Sidoides Essential Oil as a Strategy to Combat the Multidrug-Resistant Candida Auris. Pharmaceutics 2022, 14, 180. [CrossRef]
- Jaromin, A.; Zarnowski, R.; Markowski, A.; Zagórska, A.; Johnson, C.J.; Etezadi, H.; Kihara, S.; Mota-Santiago, P.; Nett, J.E.; Boyd, B.J.; et al. Liposomal Formulation of a New Antifungal Hybrid Compound Provides Protection against Candida Auris in the Ex Vivo Skin Colonization Model. Antimicrob Agents Chemother 2026, 68, e00955-23. [CrossRef]
- Eix, E.F.; Johnson, C.J.; Wartman, K.M.; Kernien, J.F.; Meudt, J.J.; Shanmuganayagam, D.; Gibson, A.L.F.; Nett, J.E. Ex Vivo Human and Porcine Skin Effectively Model Candida Auris Colonization, Differentiating Robust and Poor Fungal Colonizers. J Infect Dis 2022, 225, 1791–1795. [CrossRef]
- Brandt, P.; Mirhakkak, M.H.; Wagner, L.; Driesch, D.; Möslinger, A.; Fänder, P.; Schäuble, S.; Panagiotou, G.; Vylkova, S. High-Throughput Profiling of Candida Auris Isolates Reveals Clade-Specific Metabolic Differences. Microbiol Spectr 2026, 11, e00498-23. [CrossRef]
- Nicklas, J.P.; Deming, C.; Lee-Lin, S.; Conlan, S.; Shen, Z.; Blaustein, R.A.; Segre, J.A. Candida Auris Metabolism and Growth Preferences in Physiologically Relevant Skin-like Conditions. bioRxiv 2025, 2025.05.18.654780. [CrossRef]
- Fanning, S.; Mitchell, A.P. Fungal Biofilms. PLoS Pathogens 2012, 8, e1002585. [CrossRef]
- Ramage, G.; Kean, R.; Rautemaa-Richardson, R.; Williams, C.; Lopez-Ribot, J.L. Fungal Biofilms in Human Health and Disease. Nat Rev Microbiol 2025. [CrossRef]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-Forming Capability of Highly Virulent, Multidrug-Resistant Candida Auris. Emerg Infect Dis 2017, 23, 328–331. [CrossRef]
- Wang, X.; Bing, J.; Zheng, Q.; Zhang, F.; Liu, J.; Yue, H.; Tao, L.; Du, H.; Wang, Y.; Wang, H.; et al. The First Isolate of Candida Auris in China: Clinical and Biological Aspects. Emerging Microbes & Infections 2018, 7, 1–9. [CrossRef]
- Vila, T.; Montelongo-Jauregui, D.; Ahmed, H.; Puthran, T.; Sultan, A.S.; Jabra-Rizk, M.A. Comparative Evaluations of the Pathogenesis of Candida Auris Phenotypes and Candida Albicans Using Clinically Relevant Murine Models of Infections. mSphere 2020, 5, 10.1128/msphere.00760-20. [CrossRef]
- Pathirana, R.U.; Friedman, J.; Norris, H.L.; Salvatori, O.; McCall, A.D.; Kay, J.; Edgerton, M. Fluconazole-Resistant Candida Auris Is Susceptible to Salivary Histatin 5 Killing and to Intrinsic Host Defenses. Antimicrobial Agents and Chemotherapy 2018, 62. [CrossRef]
- Kean, R.; Delaney, C.; Sherry, L.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R.; Williams, C.; Ramage, G. Transcriptome Assembly and Profiling of Candida Auris Reveals Novel Insights into Biofilm-Mediated Resistance. mSphere 2018, 3, e00334-18. [CrossRef]
- Frías-De-León, M.G.; Hernández-Castro, R.; Vite-Garín, T.; Arenas, R.; Bonifaz, A.; Castañón-Olivares, L.; Acosta-Altamirano, G.; Martínez-Herrera, E. Antifungal Resistance in Candida Auris: Molecular Determinants. Antibiotics 2020, 9, 568. [CrossRef]
- Lockhart, S.R. Candida Auris and Multidrug Resistance: Defining the New Normal. Fungal Genetics and Biology 2019, 131, 103243. [CrossRef]
- Chaabane, F.; Graf, A.; Jequier, L.; Coste, A.T. Review on Antifungal Resistance Mechanisms in the Emerging Pathogen Candida Auris. Frontiers in Microbiology 2019, 10. [CrossRef]
- Healey, K.R.; Kordalewska, M.; Jiménez Ortigosa, C.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 Mutations Identified in Isolates of Candida Auris Directly Contribute to Reduced Azole Susceptibility. Antimicrobial Agents and Chemotherapy 2018, 62. [CrossRef]
- Rybak, J.M.; Doorley, L.A.; Nishimoto, A.T.; Barker, K.S.; Palmer, G.E.; Rogers, P.D. Abrogation of Triazole Resistance upon Deletion of CDR1 in a Clinical Isolate of Candida Auris. Antimicrobial Agents and Chemotherapy 2019, 63. [CrossRef]
- Barker, K.S.; Santana, D.J.; Zhang, Q.; Peters, T.L.; Rybak, J.M.; Morschhäuser, J.; Cuomo, C.A.; Rogers, P.D. Mutations in TAC1B Drive Increased CDR1 and MDR1 Expression and Azole Resistance in Candida Auris. Antimicrobial Agents and Chemotherapy 2025, 69. [CrossRef]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the Emerging Threat of Antifungal Resistance to Human Health. Nature Reviews Microbiology 2022, 20, 557–571. [CrossRef]
- Nett, J.E.; Andes, D.R. Antifungal Agents. Infectious Disease Clinics of North America 2016, 30, 51–83. [CrossRef]
- Chen, J.; Tian, S.; Han, X.; Chu, Y.; Wang, Q.; Zhou, B.; Shang, H. Is the Superbug Fungus Really so Scary? A Systematic Review and Meta-Analysis of Global Epidemiology and Mortality of Candida Auris. BMC Infect Dis 2020, 20, 827. [CrossRef]
- Siopi, M.; Peroukidou, I.; Beredaki, M.-I.; Spruijtenburg, B.; De Groot, T.; Meis, J.F.; Vrioni, G.; Tsakris, A.; Pournaras, S.; Meletiadis, J. Overestimation of Amphotericin B Resistance in Candida Auris with Sensititre YeastOne Antifungal Susceptibility Testing: A Need for Adjustment for Correct Interpretation. Microbiol Spectr 2023, 11, e04431-22. [CrossRef]
- Chen, S.C.A.; Sorrell, T.C. Antifungal Agents. Medical Journal of Australia 2007, 187, 404–409. [CrossRef]
- Gow, N.A.R.; Latge, J.-P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiology Spectrum 2017, 5. [CrossRef]
- Kordalewska, M.; Lee, A.; Park, S.; Berrio, I.; Chowdhary, A.; Zhao, Y.; Perlin, D.S. Understanding Echinocandin Resistance in the Emerging Pathogen Candida Auris. Antimicrobial Agents and Chemotherapy 2018, 62. [CrossRef]
- Ramos, L.S.; Barbosa, P.F.; Lorentino, C.M.A.; Lima, J.C.; Braga, A.L.; Lima, R.V.; Giovanini, L.; Casemiro, A.L.; Siqueira, N.L.M.; Costa, S.C.; et al. The Multidrug-Resistant Candida Auris, Candida Haemulonii Complex and Phylogenetic Related Species: Insights into Antifungal Resistance Mechanisms. Current Research in Microbial Sciences 2025, 8, 100354. [CrossRef]
- Misas, E.; Parnell, L.A.; Rajeev, M.; López, L.F.; Santos, A.R.D.; Mudge, Z.B.; Gade, L.; Forsberg, K.; Lyman, M.; Sexton, D.J.; et al. A Benchmark Dataset for Validating FKS1 Mutations in Candida Auris. Microbiol Spectr 2025, 13, e03147-24. [CrossRef]
- Cao, C.; Wang, Y.; Husain, S.; Soteropoulos, P.; Xue, C. A Mechanosensitive Channel Governs Lipid Flippase-Mediated Echinocandin Resistance in Cryptococcus Neoformans. mBio 2019, 10. [CrossRef]
- Arendrup, M.C.; Lockhart, S.R.; Wiederhold, N. Candida Auris MIC Testing by EUCAST and Clinical and Laboratory Standards Institute Broth Microdilution, and Gradient Diffusion Strips; to Be or Not to Be Amphotericin B Resistant? Clinical Microbiology and Infection 2025, 31, 108–112. [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Rhomberg, P.R.; Carvalhaes, C.G. Recent Increase in Candida Auris Frequency in the SENTRY Surveillance Program: Antifungal Activity and Genotypic Characterization. Antimicrob Agents Chemother 2024, 68, e00570-24. [CrossRef]
- Candida Auris: Methods and Protocols; Lorenz, A., Ed.; Methods in Molecular Biology; 1st ed. 2022.; Springer US: New York, NY, 2022; ISBN 978-1-0716-2417-3.
- Vatanshenassan, M.; Boekhout, T.; Lass-Flörl, C.; Lackner, M.; Schubert, S.; Kostrzewa, M.; Sparbier, K. Proof of Concept for MBT ASTRA, a Rapid Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry (MALDI-TOF MS)-Based Method To Detect Caspofungin Resistance in Candida Albicans and Candida Glabrata. Journal of Clinical Microbiology 2018, 56. [CrossRef]
- Di Luca, M.; Koliszak, A.; Karbysheva, S.; Chowdhary, A.; Meis, J.; Trampuz, A. Thermogenic Characterization and Antifungal Susceptibility of Candida Auris by Microcalorimetry. Journal of Fungi 2019, 5, 103. [CrossRef]
- CDC Antifungal Susceptibility Testing for C. auris Available online: https://www.cdc.gov/candida-auris/hcp/laboratories/antifungal-susceptibility-testing.html (accessed on 25 November 2025).
- CDC Antifungal Susceptibility Testing for C. auris Available online: https://www.cdc.gov/candida-auris/hcp/laboratories/antifungal-susceptibility-testing.html (accessed on 25 November 2025).
- Szymański, P.; Markowicz, M.; Mikiciuk-Olasik, E. Adaptation of High-Throughput Screening in Drug Discovery—Toxicological Screening Tests. International Journal of Molecular Sciences 2011, 13, 427–452. [CrossRef]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L.; Mowat, E.; Ramage, G.; Lopez-Ribot, J.L. A Simple and Reproducible 96-Well Plate-Based Method for the Formation of Fungal Biofilms and Its Application to Antifungal Susceptibility Testing. Nat Protoc 2008, 3, 1494–1500. [CrossRef]
- Ajetunmobi, O.H.; Wall, G.; Bonifacio, B.V.; Montelongo-Jauregui, D.; Lopez-Ribot, J.L. A 384-Well Microtiter Plate Model for Candida Biofilm Formation and Its Application to High-Throughput Screening. In Antifungal Drug Resistance; Springer US, 2023; pp. 53–64 ISBN 978-1-0716-3155-3.
- Singh, U.; Akhtar, S.; Mishra, A.; Sarkar, D. A Novel Screening Method Based on Menadione Mediated Rapid Reduction of Tetrazolium Salt for Testing of Anti-Mycobacterial Agents. Journal of Microbiological Methods 2011, 84, 202–207. [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nature Reviews Drug Discovery 2018, 18, 41–58. [CrossRef]
- Shaban, S.; Patel, M.; Ahmad, A. Improved Efficacy of Antifungal Drugs in Combination with Monoterpene Phenols against Candida Auris. Scientific Reports 2020, 10. [CrossRef]
- Lionakis, M.S.; Drummond, R.A.; Hohl, T.M. Immune Responses to Human Fungal Pathogens and Therapeutic Prospects. Nature Reviews Immunology 2023, 23, 433–452. [CrossRef]
Figure 1.
Host and cutaneous environmental factors promoting Candidozyma auris colonization, persistence and dissemination from the human skin. The epidermal microenvironment provides conditions favorable to C. auris, including moisture from sweat and lipid-rich sebum, which may support metabolic flexibility and utilization of alternative carbon sources. Continuous desquamation of corneocytes contributes to mechanical dissemination of C. auris onto surrounding environmental surfaces. Disruption of the resident cutaneous microbiota, including through chlorhexidine washing, may reduce microbial competition and promote C. auris persistence. C. auris can localize to protected niches such as hair follicles and associated sebaceous structures, potentially limiting exposure to topical antiseptics. Within the dermal/epidermal interface, interactions with immune cells are depicted, including neutrophils and lymphocytes. Evidence suggests altered neutrophil engagement and induction of interferon-γ (IFN-γ) associated responses, which may contribute to local inflammation and compromise of epithelial barrier integrity. Reduced or dysregulated IL-17A and IL-17F signaling is illustrated, consistent with impaired antifungal mucocutaneous defense pathways. Together, these factors create a permissive cutaneous niche that supports C. auris colonization, persistence, and transmission. Created in https://BioRender.com.
Figure 1.
Host and cutaneous environmental factors promoting Candidozyma auris colonization, persistence and dissemination from the human skin. The epidermal microenvironment provides conditions favorable to C. auris, including moisture from sweat and lipid-rich sebum, which may support metabolic flexibility and utilization of alternative carbon sources. Continuous desquamation of corneocytes contributes to mechanical dissemination of C. auris onto surrounding environmental surfaces. Disruption of the resident cutaneous microbiota, including through chlorhexidine washing, may reduce microbial competition and promote C. auris persistence. C. auris can localize to protected niches such as hair follicles and associated sebaceous structures, potentially limiting exposure to topical antiseptics. Within the dermal/epidermal interface, interactions with immune cells are depicted, including neutrophils and lymphocytes. Evidence suggests altered neutrophil engagement and induction of interferon-γ (IFN-γ) associated responses, which may contribute to local inflammation and compromise of epithelial barrier integrity. Reduced or dysregulated IL-17A and IL-17F signaling is illustrated, consistent with impaired antifungal mucocutaneous defense pathways. Together, these factors create a permissive cutaneous niche that supports C. auris colonization, persistence, and transmission. Created in https://BioRender.com.
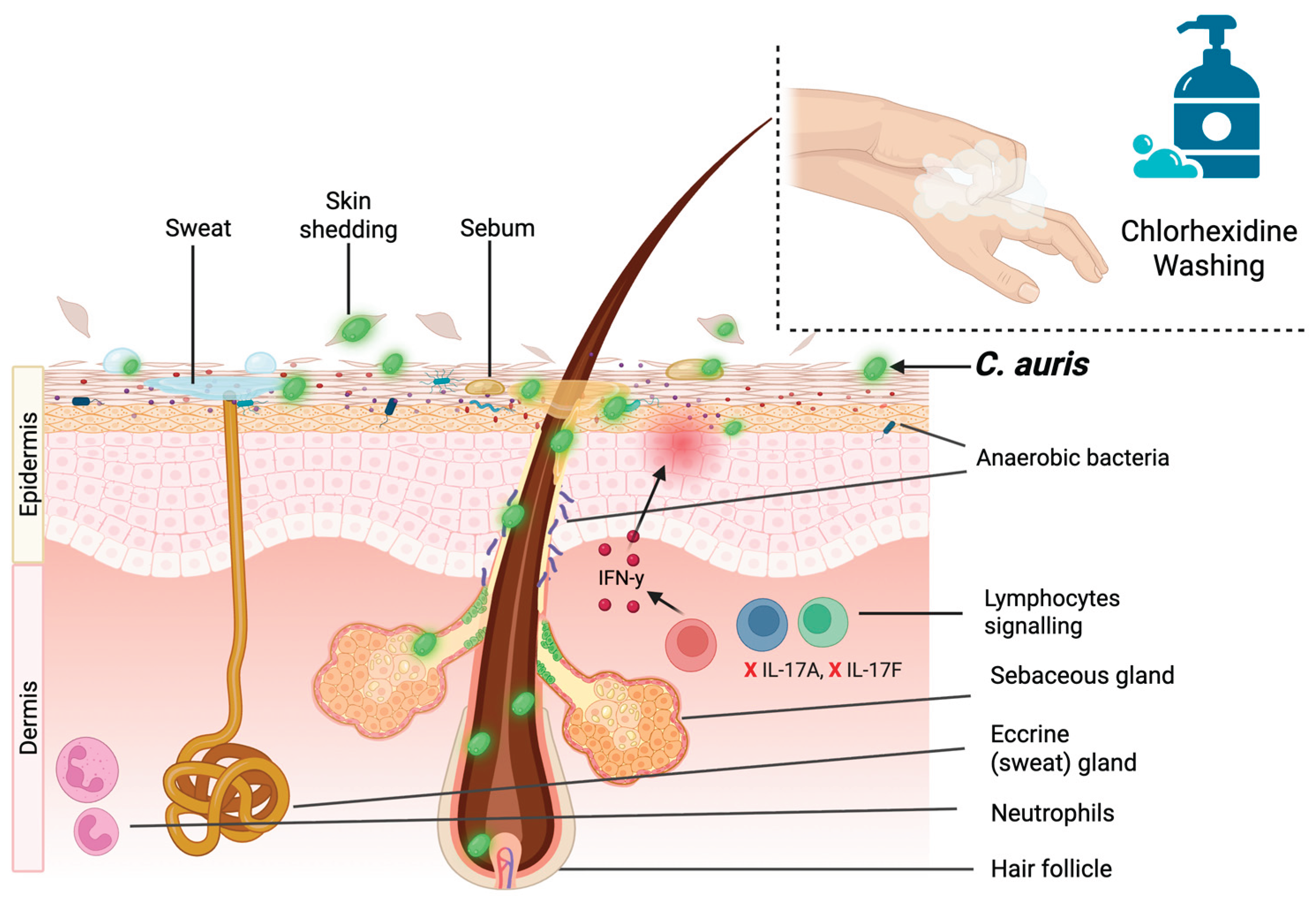
Figure 2.
Schematic representation of phase-enriched transcriptional programs underlying sequential stages of Candidozyma auris surface colonization and biofilm development. Schematic representation of key transcriptional patterns identified during each stage of Candidozyma auris surface colonization and biofilm development. Transcriptional Regulators / Signalling (Red), Adhesion Factors (Black), ECM / Cell Wall / Matrix Regulators (Blue), Stress Adaptation Factors (Green). During initial attachment (a), adhesins including IFF4/6/8, ALS1/4, CSA1, PGA26/52, and SCF1 facilitate surface adherence under regulation of EFG1 and BCR1, with cell wall organization supported by ECM33 and MP65. The response to drastic changes in heat (red arrows), osmotic pressure (rain cloud) and exposure to disinfectants/oxidising agents (spray bottle) is depicted in the stress adaptation phase (b). HOG1-mediated signalling and induction of stress response genes (HSP90, HSP104, SOD2, CAT1, PRX1) alongside metabolic regulators (TPS1/2) promote environmental tolerance. Persistence and early biofilm formation (c) involve regulatory activity of EFG1, ACE2, and TAC1, alongside cell wall and efflux-associated genes (CDR1, MDR1, FKS1, CHS1/2, GSC1) and sustained HSP90 expression. During biofilm maturation (d), ACE2, ZAP1, and BCR1 coordinate matrix development and intercellular adhesion, with contributions from FKS1, KRE6, EXG, CDR1, SNQ2, and RBT5. In dispersal (e), regulatory shifts involving NRG1, PES1, and ACE2 accompany expression of adhesins (IFF4, ALS5), cell wall remodelling (CHS1/2, KRE6, EXG), and stress tolerance genes, priming released cells for survival and reattachment. Created in https://BioRender.com.
Figure 2.
Schematic representation of phase-enriched transcriptional programs underlying sequential stages of Candidozyma auris surface colonization and biofilm development. Schematic representation of key transcriptional patterns identified during each stage of Candidozyma auris surface colonization and biofilm development. Transcriptional Regulators / Signalling (Red), Adhesion Factors (Black), ECM / Cell Wall / Matrix Regulators (Blue), Stress Adaptation Factors (Green). During initial attachment (a), adhesins including IFF4/6/8, ALS1/4, CSA1, PGA26/52, and SCF1 facilitate surface adherence under regulation of EFG1 and BCR1, with cell wall organization supported by ECM33 and MP65. The response to drastic changes in heat (red arrows), osmotic pressure (rain cloud) and exposure to disinfectants/oxidising agents (spray bottle) is depicted in the stress adaptation phase (b). HOG1-mediated signalling and induction of stress response genes (HSP90, HSP104, SOD2, CAT1, PRX1) alongside metabolic regulators (TPS1/2) promote environmental tolerance. Persistence and early biofilm formation (c) involve regulatory activity of EFG1, ACE2, and TAC1, alongside cell wall and efflux-associated genes (CDR1, MDR1, FKS1, CHS1/2, GSC1) and sustained HSP90 expression. During biofilm maturation (d), ACE2, ZAP1, and BCR1 coordinate matrix development and intercellular adhesion, with contributions from FKS1, KRE6, EXG, CDR1, SNQ2, and RBT5. In dispersal (e), regulatory shifts involving NRG1, PES1, and ACE2 accompany expression of adhesins (IFF4, ALS5), cell wall remodelling (CHS1/2, KRE6, EXG), and stress tolerance genes, priming released cells for survival and reattachment. Created in https://BioRender.com.
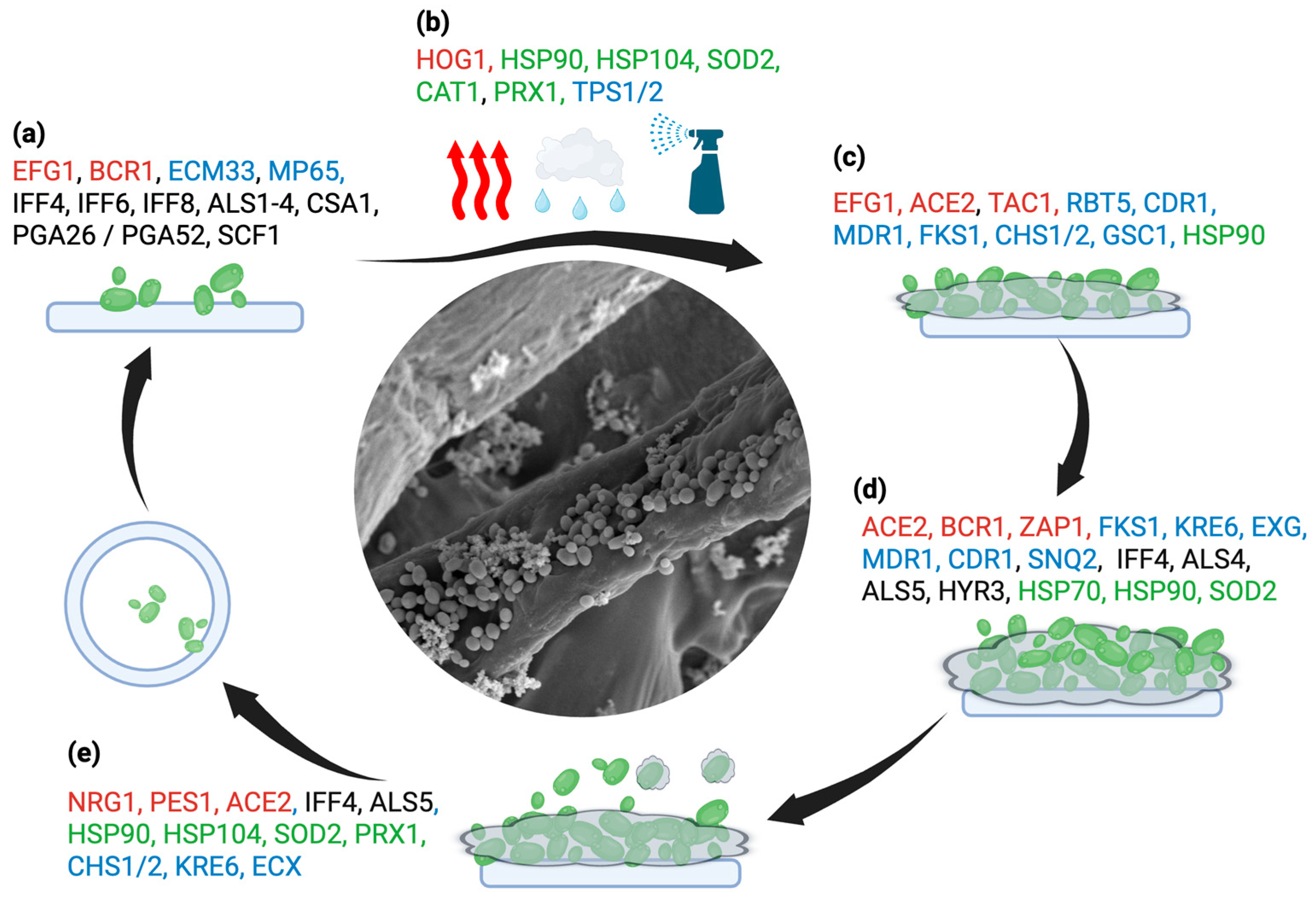
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



