Submitted:
09 February 2026
Posted:
10 February 2026
You are already at the latest version
Abstract
Candida albicans is the primary agent of acute vulvovaginal candidiasis(VVC) and its recurrent form(RVVC). Local innate immunity contributes to both defense and pathogenesis during vaginal Candida infection, where epithelial β-defensins(BD) constitute key components of the mucosal barrier. We previously reported that epithelial BD-1 expression is dynamically modulated during murine and human vaginitis, revealing strain-dependent and stimulus-specific regulation but leaving the host pathways involved unresolved. This study functionally defines the contribution of key immune pathways to epithelial antimicrobial peptide regulation. Using a murine model of VVC and the virulent C. albicans strain SC5314, we analyzed the temporal expression of BD-1 and extended our study to BD-3. In wild-type mice, both defensins displayed a biphasic pattern: early induction followed by attenuation as infection progressed. Genetic loss-of-function approaches revealed that NLRP3/IL-1β signaling is required for early BD-1 induction, whereas IL-17RA signaling preferentially supports sustained BD-3 expression. Together, these findings establish a causal and temporal link between host immune signaling and epithelial defensin regulation, and reveal a transient subversion of mucosal defenses by C. albicans. This work advances understanding of epithelial innate immunity, defining distinct temporal programs for BD-1 and BD-3, and identifying NLRP3/IL-1β and IL-17RA signaling as key pathways shaping mucosal defensin expression.
Keywords:
Candida albicans
; vulvovaginal candidiasis
; antimicrobial peptide
; β-Defensin
; NLRP3/IL-1β signaling
; IL-17RA signaling
1. Introduction
Candida albicans is a common commensal fungus of the human mucosal microbiota and the main etiological agent of vulvovaginal candidiasis (VVC). This acute inflammatory disease affects approximately 75% of women at least once during their reproductive lifetime and represents one of the most common causes of vaginal infection, following bacterial vaginosis. Between 6% and 10% of these women experience a distressing condition characterized by recurrent episodes of VVC (RVVC), defined as four or more symptomatic episodes per year [1,2,3]. Estimates indicate that RVVC affects around 138 million women worldwide annually, underscoring its significant public health impact [4]. Despite the high prevalence of Candida vaginitis, many aspects of the host–fungal interaction remain poorly understood [5].
Antifungal immune responses in the vaginal mucosa are distinct from those observed in other Candida infections. In this particular tract, the fungus coexists under a tightly regulated surveillance mechanism involving resident cells and basal levels of immune mediators [6]. During asymptomatic vaginal colonization, epithelial cells (ECs) sense its presence through surface-expressed pattern recognition receptors (PRRs) that recognize Candida as a harmless commensal [5,7]. The C. albicans yeast-to-hypha transition marks the shift to pathogenicity and triggers virulence programs that drive tissue invasion [7,8,9], epithelial activation, antimicrobial peptides (AMPs) expression, a strong interleukin-1 beta (IL-1β) driven inflammatory response, and polymorphonuclear leukocytes (PMNs) recruitment [10,11,12].
Among the cytokines involved in antifungal defense at mucosal sites, IL-1β and interleukin-17 (IL-17) play pivotal roles in orchestrating local immune responses [13,14]. IL-1β, produced by epithelial and innate immune cells upon fungal recognition, is a key mediator of inflammation and mucosal protection, and is considered a hallmark of VVC. In parallel, IL-17 signaling has been implicated in antifungal immunity, in part by promoting epithelial activation and PMNs recruitment. However, the contribution of IL-17 to VVC remains controversial, as several studies have reported low or undetectable levels of this cytokine in the vaginal environment, suggesting that its role may be limited or context-dependent [15,16].
AMPs are evolutionarily conserved effector molecules that display broad antimicrobial activity against pathogens. Among them, β-defensins (BDs) constitute one of the most important AMPs family [17,18]. These small (4–6 kDa) cationic peptides not only exert potent microbicidal effects but also act as immune modulators by recruiting PMNs, immature dendritic cells, and memory T cells to infection sites [18,19,20]. Human Beta Defensin 1 (hBD1) is constitutively expressed in the uninflamed mucosa of the female reproductive tract and is considered a major epithelial defense factor against invading microorganisms. In contrast, inducible defensins such as hBD3 are rapidly upregulated in response to inflammatory or infectious stimuli, contributing to the reinforcement of mucosal protection [21,22].
We recently demonstrated that C. albicans modulates epithelial BD-1 expression during vaginal infection, revealing strain-dependent and stimulus-specific regulation in both murine and human ECs models [23]. While these findings provided relevant descriptive and correlative data, the host immune pathways governing this regulation remained undefined. Here, using genetic loss-of-function approaches targeting NLRP3/IL-1β and IL-17RA signaling in a murine VVC model, we sought to define the contribution of these pathways to epithelial BD-1 and BD-3 expression in the vaginal mucosa.
2. Methods
2.1. Candida albicans
The Candida albicans SC5314 strain was used in all experiments. This strain is considered highly virulent based on its well-documented ability to express multiple virulence-associated traits, including elevated secreted aspartyl protease (SAP) and lipase activities, strong adherence to epithelial surfaces, and robust biofilm-forming capacity, as reported in previous studies [24]. These features are known to contribute to epithelial invasion and immune activation [16,25,26]. Yeast cells were cultured on Sabouraud Dextrose Agar (SDA; Britania, Los Patos, Argentina) at room temperature (RT).
For infection assays, the inoculum for the experiments was standardized by counting yeast cells in phosphate-buffered saline (PBS) using a Neubauer hemocytometer (Boeco, Germany). The number of viable yeasts was determined in triplicate by plating on SDA and incubating for 48 h at RT.
2.2. Mice
C57BL/6J wild-type (WT) mice were purchased from the Faculty of Veterinary Science, National University of La Plata, Argentina. Nlrp3(−/−) and Il17ra (−/−) mice were used to genetically define the contribution of inflammasome and IL-17–dependent pathways to epithelial antimicrobial peptide regulation during vaginal infection.
Knockout mice with targeted disruption of the IL17-RA receptor (Il17ra(-/-)) on the C57BL/6 background were provided by Amgen Inc. (MTA#7939181). Knockout mice with targeted disruption of the NLRP3 receptor (nlrp3/Jtm1Bhk) on the C57BL/6 background were obtained from The Jackson Laboratory (Bar Harbor, ME, USA).
All animals were maintained under a standard light cycle (12 h light/dark) with free access to food and water. All experimental procedures were approved and conducted in accordance with the guidelines of the Animal Experimentation Ethics Committee of the Faculty of Chemical Sciences, National University of Córdoba (Approval number: RD-2022-2789-E-UNC-DEC#FCQ), and strictly followed the recommendations of the Guide to the Care and Use of Experimental Animals published by the Canadian Council on Animal Care (OLAW Assurance number A5802-01). Every effort was made to minimize animal suffering during the experiments.
2.3. In Vivo VVC Model
Female WT, Il17ra(-/-) and Nlrp3(-/-) mice (8–10 weeks old) were treated subcutaneously with 0.2 mg of β-estradiol 17-valerate (Merck-Millipore, Darmstadt, Germany) dissolved in 100 μl of sesame oil (Merck-Millipore) on days −6 and −3, before vaginal infection on day 0. On days 2 and 4 post infection (pi), mice received additional estradiol injections to maintain the pseudoestrus state [27]. Estrogenized mice were then inoculated intravaginally with 20 μl of a C. albicans SC5314 suspension (infected group; 5 × 106 viable yeasts/PBS) or PBS (uninfected group) [26].
For each experimental condition, WT mice were included as internal controls and processed in parallel with Il17ra(-/-) or Nlrp3(-/-) animals. WT and knockout mice were infected, sampled, and analyzed within the same experimental runs to ensure direct comparability.
2.4. CVL Collection
Mice were anesthetized with isoflurane, and cervicovaginal lavages (CVL) were collected by washing the vaginal tract with 70 μl of sterile PBS [27,28]. The collected CVL were immediately processed for quantification of CFUs or PMNs. Another portion of each CVL was centrifuged for 10 min at 4 °C, and the resulting cell-free supernatant was stored at −80 °C for subsequent cytokine analysis.
2.5. Fungal Burden (CFU/ml)
CVL samples were serially diluted (1:10, 1:100) in sterile PBS, and 100 μl aliquots of each dilution were plated in duplicate onto Petri dishes containing SDA. Plates were incubated at RT for 48 h, and the number of yeast colonies was recorded as colony-forming units per milliliter (CFU/ml) [26,29]. For clarity of data presentation, WT control groups corresponding to each knockout experiment are shown separately in the figures, as they were processed in parallel under identical experimental conditions.
2.6. PMNs Quantification
Cells from CVL samples were adjusted to a concentration of 5 × 104 cells per 100 μl by resuspension in PBS, based on total cell counts determined using a Neubauer hemocytometer. Subsequently, samples were subjected to cytocentrifugation at 500 rpm for 5 min onto glass slides using a Shandon Elliot cytospin system. The cells were then stained with May-Grünwald Giemsa (MGG; Biopak, Buenos Aires, Argentina) and examined under a NIKON ECLIPSE microscope (Nikon, Tokyo, Japan). PMNs were identified based on morphological criteria. For each sample, PMNs were manually counted in ten non-adjacent fields at a 40× objective. PMNs counts were averaged across fields for each sample, and results were expressed as mean PMNs counts per group ± standard error of the mean (SEM) [26].
2.7. Cytokines Quantification
Cytokine concentrations in CVL samples were quantified using commercially available IL-1β and IL-17A ELISA detection kits (R&D Systems, McKinley Place, MN, USA), as previously described [26,30]. Absorbance was measured using a BIO-RAD microplate reader, and cytokine concentrations were interpolated from standard curves. Data were expressed as pg/ml.
2.8. Immunofluorescence for the Evaluation of mBD1 and mBD3 Expression
To detect BDs by immunofluorescence, 10 μl of CVL from each experimental group was placed on glass slides and air-dried at RT. Samples were fixed with pure methanol (Sintorgan) for 5 min at RT and washed with PBS. Blocking was performed with 10% bovine serum albumin (BSA) and 0.3% Triton X-100 in PBS for 2 h at RT. Slides were then incubated overnight (ON) at 4°C with a rabbit IgG primary antibody against mouse BD-1 (Santa Cruz Biotechnology) or mouse BD-3 (Santa Cruz Biotechnology) (1:50). After three PBS washes, samples were incubated for 2 h at RT with a rabbit anti-IgG secondary antibody conjugated to Alexa Fluor 488 (Invitrogen) at a 1:500 dilution.
Slides were mounted with FluorSave™ Reagent (Merck Millipore). Images were acquired using a LEICA DMi8 inverted fluorescence microscope (Leica Microsystems, Wetzlar, Germany) equipped with a 20× objective. At least ten images were captured per cytospin preparation, each containing a minimum of ten cells per field. mBD1 and mBD3 expression was evaluated in the cytoplasm of vaginal ECs, identified in bright-field images based on their characteristic morphology. Selection masks were generated to individually delineate each cell and were superimposed onto the corresponding fluorescence images, allowing specific quantification of the cytoplasmic signal. This approach ensured accurate and cell-specific assessment of BDs expression. Semi-quantitative analysis of mBD1 and mBD3 expression was performed using ImageJ software. Fluorescence intensity (FI) was expressed as the mean per-cell intensity normalized to the control group treated on day 2 of the kinetic study, according to the following equation:
Normalized fluorescence = FI of infected mouse ECs
Mean FI of day 2 treated control ECs
2.9. Statistical Analysis
Experiments were performed in triplicate unless otherwise indicated. Data were expressed as the mean ± standard error of the mean (SEM). Differences between groups were analyzed using parametric statistical tests, including Student’s t-test (for two-group comparisons) or one-way ANOVA, followed by appropriate post hoc tests (Dunnett’s, Bonferroni’s, or Welch’s), depending on the assumptions of normality and homogeneity of variance. Statistical significance was defined as p< 0.05 (*), p< 0.01 (**), p< 0.001 (***), and p< 0.0001 (****). All analyses were performed using GraphPad Prism 10 software (GraphPad, San Diego, CA, USA).
3. Results
3.1. Establishment of Infection and Local Inflammatory Profile During Experimental Vulvovaginal Candidiasis in C57BL/6 (WT) Mice
To characterize the modulation and kinetic expression of mBD1 and mBD3 during VVC, we first explored the key local parameters associated with the vaginal mucosal response to C. albicans SC5314 infection in C57BL/6 (WT) mice. C. albicans SC5314 was chosen based on its well-documented virulence and its ability to elicit robust local immune responses in experimental models of vaginal candidiasis [16,25]. Estrogen-treated WT animals were intravaginally infected with 5 ×106 viable yeast on day 0 (Infected group), while a control group received PBS (uninfected group) (Figure 1A). CVL were collected on days 2, 4, and 8 post infection (pi) for subsequent analysis. First, to assess the progression of C. albicans infection, local fungal burden was quantified by CFUs on each day of the study. WT infected mice displayed robust colonization that persisted throughout the evaluated period, with similar values on days 2 and 4, followed by a significant decrease in fungal burden by day 8 (Figure 1B). MGG-stained slides of cells present in CVL from both groups of animals revealed abundant ECs, consistent with shedding of superficial epithelial layers and estrogen-induced epithelial turnover (Figure 1C). During VVC, PMNs recruited to the vaginal lumen contribute to tissue damage and the local inflammation [12]. In this model, we observed a peak recruitment of PMNs in WT infected mice at day 2, followed by a marked decrease at days 4 and 8, showing significant differences compared with the uninfected group (Figure 1D).
In line with the aim of the present study, we evaluated the intravaginal concentrations of IL-1β, a key marker associated with Candida vaginitis [10,11], and IL-17A, a cytokine linked to antifungal immunity whose role in VVC remains controversial [15,16]. We observed that the highest levels of IL-1β in CVL from infected mice were detected at days 2 and 4 (Figure 1E) with a significant decrease by the end of the study (day 8). These increases were significant compared with the uninfected WT group at all evaluated time points (Figure 1E). Regarding IL-17A, our findings showed low local production of this cytokine in both infected and uninfected WT animals, with IL-17A concentrations remaining below 15 pg/ml, which corresponds to the detection limit of the assay.
3.2. C. albicans SC5314 Infection Differentially Modulates mBD1 Expression in the Vaginal Tract
At mucosal surfaces, ECs function as an active barrier, and BD-1 is constitutively expressed in uninflamed mucosa as part of the epithelial surveillance system [31]. Our previous findings using low-virulence and RVVC-associated C. albicans isolates revealed strain-dependent modulation of epithelial BD-1 expression [23]. Here, we extended this analysis to infection with the more virulent strain SC5314 to define the temporal dynamics of BD-1 expression during VVC.
mBD1 protein expression was assessed in ECs recovered from CVL of infected and uninfected WT mice using a specific anti-mBD1 antibody and IF assay (Figure 2A). We observed distinct kinetic profiles in each experimental group associated with exposure to treatments (Figure 2B). At day 2 pi, infected animals exhibited a significant increase in mBD1 expression compared with the uninfected group, showing an approximately 380% rise, indicating an early and strong response to C. albicans presence (Figure 2C). At day 4 pi, mBD1 expression levels in infected animals were similar to those at day 2 but showed a significant decrease compared with the uninfected group, highlighting the marked response of the vaginal tract to estrogenic treatment and the ability of the fungus to inhibit peptide expression. Analysis of percentage variation indicated a 37% reduction in the infected group (Figure 2C). By the end of the study (day 8), while estrogen treatment continued to stimulate mBD1 expression, infection progression resulted in a 75% reduction in peptide levels. These results illustrate the mucosal tract’s ability to upregulate peptide expression early during infection (day 2), as well as the capacity of C. albicans to inhibit mBD1 as the infection advances.
These results deepen our earlier observations by demonstrating that mBD1 modulation reflects a coordinated and temporally regulated epithelial response during vaginal infection.
3.3. C. albicans SC5314 Infection Differentially Modulates mBD3 Expression in the Vaginal Tract
BD-3 is an inducible peptide that is rapidly produced in response to inflammatory or infectious stimuli and displays strong candidacidal activity, exceeding that of the constitutively expressed BD-1 [32]. However, its role during vulvovaginal candidiasis remains poorly characterized. We therefore analyzed the temporal expression of mBD-3 in the vaginal tract during experimental infection.
mBD3 expression was studied in ECs recovered from CVL of infected and uninfected WT mice using a specific anti-mBD3 antibody and IF assay (Figure 3A). At the early time point of the kinetic study (day 2), mBD3 exhibited a peak of expression associated with the presence of C. albicans, with a significant difference compared with the uninfected group (Figure 3B). The percentage variation analysis indicated an increase of 159 % in mBD3 expression relative to the control group (Figure 3C). At day 4 pi, mBD3 expression levels in infected mice showed a significant decrease compared with day 2 pi and with the uninfected group. The percentage variation between groups at day 4 revealed a 68% reduction in peptide expression in infected animals compared with uninfected controls. Interestingly, estrogen treatment also stimulates mBD3 expression in this tract, reaching its highest levels at this time point (Figure 3B). By day 8 of the study, estrogen treatment continued to stimulate mBD3 expression in the uninfected group, whereas infection progression led to an approximately 75% reduction in mBD3 expression (Figure 3C).
Together, these findings indicate that mBD3 exhibits a biphasic expression pattern during C. albicans infection, with early induction followed by sustained downregulation, similar to that observed for mBD1.
3.4. NLRP3 Inflammasome Pathway Activation Drives mBD1 but not mBD3 Induction during C. albicans Vaginal Infection
The canonical IL-1β pathway, activated through fungal pathogen-associated molecular patterns (PAMPs) and NLRP3 inflammasome signaling, plays a central role in cytokine secretion [33]. We previously reported that recombinant human IL-1β induces hBD1 expression in human ECs from the female genital tract, supporting a role for IL-1β in BD-1 regulation [23].
To genetically define the contribution of NLRP3/IL-1β –dependent signaling to epithelial AMPs regulation during VVC, we developed the experimental model described above in NLRP3-deficient mice (Nlrp3(⁻/⁻)) and compared their responses with those obtained from WT animals. For this purpose, estrogen-treated Nlrp3(⁻/⁻) animals were iv infected with 5 ×106 viable C. albicans SC5314 yeasts on day 0 (Infected Nlrp3(⁻/⁻) group), while a control group received PBS inoculation (uninfected Nlrp3(⁻/⁻) group). CVL were collected on days 2, 4, and 8 pi to assess the progression of fungal infection by CFU counts. Nlrp3(⁻/⁻) mice exhibited robust vaginal colonization by C. albicans, which persisted throughout the evaluation period (Figure 4A, left panel). Fungal burdens in infected Nlrp3(⁻/⁻) mice were comparable to those observed in WT animals at days 2 and 4, followed by a significant reduction at day 8 (Figure 4A, right panel).
Based on these findings, day 2 pi was selected to investigate the contribution of the NLRP3/IL-1β signaling to mBD1 and mBD3 regulation. At this time point, both peptides exhibited peak expression in infected WT mice, while fungal burden remained comparable between WT and Nlrp3(⁻/⁻) groups, allowing comparison independent of colonization differences. In addition, estrogen-mediated stimulation of defensin expression becomes more prominent at later time points, allowing infection-driven regulation to be examined independently of hormonal effects. Accordingly, CVL from both genotypes was collected on day 2 of the experimental schema. Figure 4B shows a representative image of mBD1 expression in ECs recovered from infected and uninfected mice, and Figure 4C displays the corresponding fold change in peptide expression. As observed, C. albicans infection induced a significant increase in mBD1 expression in WT animals compared with uninfected animals. In contrast, Nlrp3(⁻/⁻) mice showed comparable mBD1 expression levels in infected and uninfected groups. Moreover, when comparing infected animals of both genotypes, Nlrp3(⁻/⁻) mice exhibited significantly lower mBD1 expression than WT mice.
For mBD3, similar to what was observed in infected WT animals (Figure 4D and 4E), Nlrp3(⁻/⁻) mice showed a significant increase in mBD3 levels in response to C. albicans infection compared with the uninfected Nlrp3(⁻/⁻) group. Interestingly, comparison between infected groups of both genotypes revealed that mBD3 levels in the Nlrp3(⁻/⁻) group were comparable to those observed in WT mice (Figure 4E).
Collectively, these findings indicated NLRP3/IL-1β signaling is required for optimal mBD1 induction during early infection, whereas mBD3 regulation occurs independently of this pathway.
3.5. IL-17RA Signaling Differentially Regulates mBD1 and mBD3 Expression during C. albicans Vaginal Infection
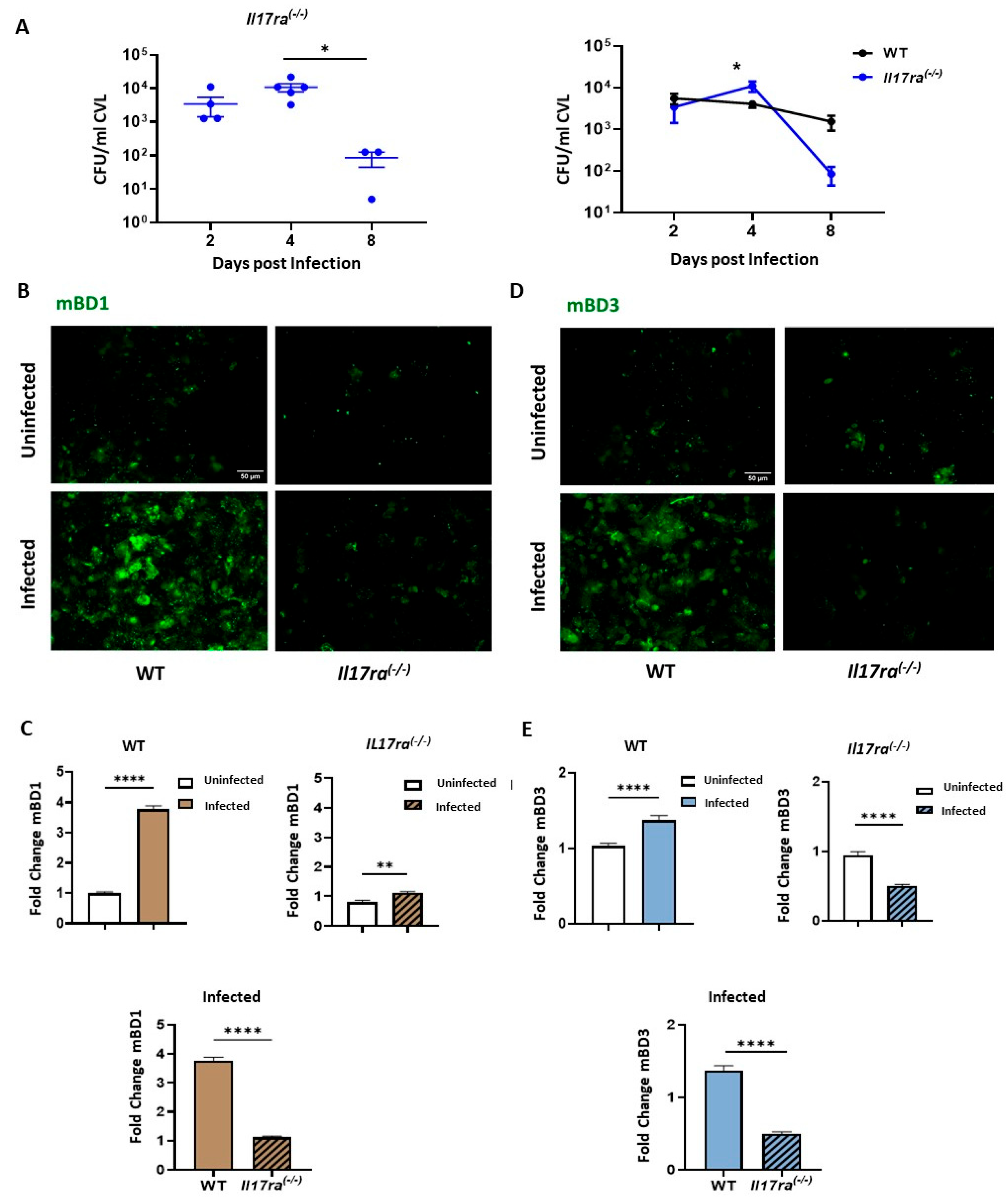
While IL-17 plays a well-established role in intestinal candidiasis [14], its contribution to the immunopathogenesis of VVC remains unclear [15,16]. To evaluate the role of IL-17RA signaling in vaginal infection and BD regulation, the VVC model described above was established in IL-17RA-deficient mice (Il17ra(⁻/⁻)). Estrogen-treated Il17ra(⁻/⁻) animals were iv infected with 5×106 viable C. albicans SC5314 yeasts on day 0 (Infected Il17ra(⁻/⁻) group), while a control group received PBS inoculation (uninfected Il17ra(⁻/⁻) group). The progression of infection was evaluated by CFU in the CVL collected on days 2, 4, and 8 pi (Figure 5A, left panel). No significant differences in fungal burden were observed between infected Il17ra(⁻/⁻) mice at days 2 and 4 pi; but at day 8, these animals showed a decrease in fungal burden compared to previous days. When compared with infected WT mice, no significant differences in CFU were observed between both strains at day 2. A transient increase in vaginal fungal burden was observed in Il17ra⁻/⁻ mice at day 4 pi; however, this difference was no longer evident by day 8. (Figure 5A, left panel). At the end of the study (day 8), infected Il17ra(⁻/⁻) mice showed a reduction in fungal burden compared to the WT group.
Since fungal burden at day 2 remained similar between WT and Il17ra(⁻/⁻) mice, and both mBD1 and mBD3 expression at this time point are independent of estrogenic stimulation, we selected this day to evaluate the contribution of IL-17 signaling to BDs regulation in the vaginal tract after C. albicans infection (Figure 5B and 5D). As previously demonstrated in WT mice, fungal infection induced a significant increase in mBD1 expression compared to uninfected controls. In Il17ra(⁻/⁻) mice, C. albicans infection also resulted in a significant increase in mBD1 expression relative to uninfected Il17ra(⁻/⁻) animals. However, comparison between infected groups of both genotypes revealed that mBD1 levels in Il17ra(⁻/⁻) mice were significantly lower than those observed in WT animals.
For mBD3, meanwhile, C. albicans infection induced an increase in peptide expression at day 2 pi compared with uninfected WT control, Il17ra(⁻/⁻) mice failed to upregulate mBD3 expression in response to C. albicans infection, showing significantly lower levels compared with infected WT mice (Figure 5D and 5E). Finally, when comparing both genotypes, infected Il17ra(⁻/⁻) mice exhibited significantly lower mBD3 levels than infected WT animals.
Collectively, these findings indicate that IL-17RA signaling contributes to mBD3 induction during C. albicans vaginal infection, whereas mBD1 expression is only partially dependent on this pathway.
4. Discussion
The onset of clinical symptoms in patients with VVC is characterized by the presence of Candida in the vaginal exudate, a local inflammatory reaction driven by PMNs recruitment into the lumen, and high concentrations of proinflammatory cytokines, with IL-1β being considered pathognomonic of this infection [7,34]. The VVC model in C57BL/6 mice infected with C. albicans SC5314 used in this study reproduces these key infection parameters. The fungus was isolated from the vaginal cavity throughout the course of infection. A high fungal burden was present at days 2 and 4 pi, accompanied by a strong recruitment of PMNs into the vaginal lumen (day 2) and elevated IL-1β levels. Despite their abundance, PMNs fail to control C. albicans growth, and their recruitment does not correlate with fungal clearance [35,36]. Evidence from murine and human studies indicates that neutrophil depletion does not modify fungal burden and that components of the vaginal environment, such as heparan sulfate, can neutralize PMNs fungicidal activity [7,37]. Thus, although neutrophils are classically regarded as key effector cells, their antimicrobial function appears limited in the vaginal milieu [3,5], underscoring the relevance of exploring additional microbicidal mechanisms, such as the contribution of epithelial-derived AMPs.
Studies in the human female genital tract have demonstrated that hBDs are key components of the mucosal defense system, acting as broad-spectrum AMPs that contribute to the control of viral, bacterial, parasitic, and fungal pathogens [31,38]. In addition to their antimicrobial role, their expression is modulated by hormones. Estrogen-dependent upregulation of BD-1 through ERα and ERβ signaling pathways has also been demonstrated [39,40]. In line with this concept, our results showed a peak in the expression of both the constitutive mBD1 and the inducible mBD3 at days 4 and 8 in estrogen-treated but uninfected WT mice. These results provide novel data on the interplay between hormonal signaling and epithelial antimicrobial responses in the murine vaginal mucosa.
Both mBD1 and mBD3 exhibit direct candidacidal activity, although with different efficacies. In vitro studies have shown that mBD3 is the more potent fungicidal peptide, retaining activity under physiological salt conditions and acting against both yeast and hyphal forms, whereas mBD1 is weaker and more salt-sensitive [41,42]. Beyond their antimicrobial properties, these peptides exert distinct immunoregulatory functions. mBD1 contributes to mucosal homeostasis by maintaining epithelial integrity and controlling commensal balance, thereby limiting excessive inflammation [31,43]. In contrast, mBD3 displays a broader immunomodulatory profile, enhancing cytokine and chemokine production and promoting the recruitment of PMNs, dendritic cells, and T cells. Moreover, its interaction with PRR signaling pathways suggests a dual role in amplifying antifungal responses while modulating immune activity under certain inflammatory or chronic conditions [44].
An important finding from our time-course studies of vaginal infection reveals a dynamic epithelial response, as the vaginal epithelium senses C. albicans and mounts a robust upregulation of both mBD1 and mBD3 by day 2 pi, reflecting an early barrier-driven attempt to contain fungal growth. However, by days 4 and 8, the fungus markedly suppresses the expression of both the constitutive mBD1 and the inducible mBD3. This biphasic pattern reflects our previous clinical observations in women with VVC, where ECs from acute cases showed high hBD1/hBD3 expression, but patients with RVVC exhibited strong transcriptional and protein downregulation. Consistently, in vitro experiments using SAP and LIP inhibitors demonstrated that these fungal virulence factors contribute to this regulatory switch [23]. In this context, it is important to note that BD expression in this study was assessed at the protein level in cells recovered from the vaginal lumen, a sampling approach used in our study in both acute and recurrent VVC patients due to its minimally invasive nature and ease of collection, while reliably reflecting the local mucosal immune microenvironment. This strategy provides biologically relevant information on the availability of AMPs at the mucosal interface, where host–fungus interactions occur. Previous studies have shown that BD transcript levels do not necessarily correlate with protein abundance in VVC, supporting the relevance of protein-based analyses [23]. Moreover, the evaluation of defensin expression in vaginal lavage cells allows the identification of responding cellular subsets, an aspect that cannot be captured by bulk transcriptomic approaches.
Similar to our results, an inhibitory pattern of BDs expression has been reported in other genital tract infections. In women infected with Chlamydia trachomatis or Neisseria gonorrhoeae, levels of hBD1, hBD2, and hBD3 in CVL were significantly lower than in uninfected controls [45,46]. In a larger cohort of women with bacterial vaginosis, BD expression was also reduced, particularly for hBD2 [45]. This inhibitory effect on BD production is not restricted to the genital tract. In gastric and intestinal epithelia, Helicobacter pylori, Vibrio cholerae, and enterotoxigenic Escherichia coli have been shown to downregulate hBD1 through virulence factors such as CagA or bacterial toxins [47,48,49]. Given the critical antimicrobial and immunomodulatory functions of epithelial BDs, their suppression can represent an efficient and conserved mechanism of immune evasion at mucosal surfaces. Our findings in patients and experimental models clearly demonstrate that C. albicans exploits this strategy to establish and persist in vaginal tracts, weakening local defenses and promoting conditions that favor the permanence of infection.
Having established the epithelial response pattern in WT animals infected with C. albicans SC5314, we next wanted to functionally define the contribution of key immune pathways to epithelial AMPs regulation. mBD1 and mBD3 expression were assessed in mice deficient in NLRP3 and IL-17RA using the VVC model. The canonical NLRP3/IL-1β pathway has been well documented in animal models [8,50] and in patients with NLRP3 polymorphisms [51]. Pietrella et al. [52] reported that components of the NLRP3 inflammasome pathway were overexpressed in vaginal samples from patients with VVC, demonstrating that inflammasome activation occurs during Candida vaginitis. Similarly, NLRP3 induction was observed in vaginal tissue following murine C. albicans infection [33,53]. Nevertheless, our results in infected Nlrp3(⁻/⁻) mice revealed that the absence of NLRP3 did not impair the control of vaginal fungal burden, as these mice showed colonization levels comparable to WT animals at early time points and even lower CFU counts at the end of the study. Thus, NLRP3 signaling is dispensable for the control of vaginal fungal burden under the experimental conditions tested. Consistently, inhibition of NLRP3 with glyburide in infected WT animals mimicked the results observed in Nlrp3(⁻/⁻) mice [33], further supporting that NLRP3 signaling is dispensable for fungal clearance in this mucosal site.
To evaluate the contribution of the NLRP3 pathway to the functional regulation of BDs, we next examined the expression of mBD1 and mBD3. Day 2 pi was selected for these analyses, as both peptides showed a clear increase in response to C. albicans at this time point. Notably, this represents an early phase of infection when estrogen-driven stimulation is not detected and before the fungus exerts its inhibitory effects on defensin expression. Although NLRP3 was nonessential for fungal control, its absence markedly affected the epithelial response. The significant reduction of mBD1 expression in infected Nlrp3(⁻/⁻) mice compared with WT animals indicates that mBD1 induction is strongly dependent on NLRP3/IL-1β signaling. This result supports a model in which infection-driven inflammasome activation and IL-1β release act as upstream cues for the transcriptional upregulation of constitutive epithelial defensin. In contrast, mBD3 expression was preserved in Nlrp3(⁻/⁻) mice, suggesting that its induction follows an inflammasome-independent pathway. Given that mBD3 is an inducible peptide typically associated with inflammatory stimuli, these findings highlight distinct regulatory networks governing mBD1 and mBD3 expression during the early response to C. albicans challenge. Altogether, our data reveal that while NLRP3/IL-1β signaling is essential for optimal mBD1 expression, alternative mechanisms are sufficient to trigger mBD3 upregulation in the vaginal tract. In agreement, previous evidence positions IL-1β as a major regulator of BD-1. Pahl et al. [54] reported in a Candida esophagitis infection model that blockade of IL-1β receptor-dependent signaling strongly reduces the induction of hBDs. They also describe a signaling loop between IL-1β and hBDs regulation. Several in vitro results have confirmed this report [55,56]. We likewise demonstrated that recombinant hIL-1β markedly upregulates hBD1 expression in hECs from the female genital tract, identifying IL-1β as a key regulator of this peptide [23]. Loss of NLRP3 selectively impaired early BD-1 induction, providing genetic evidence that inflammasome-dependent signaling governs the initial epithelial antimicrobial response suggested by our prior in vitro observations. Together, these data support IL-1β–mediated signaling as a central link between fungal sensing and epithelial antimicrobial responses in the vaginal tract.
Consistent with previous reports, the absence of IL-17RA signaling did not impair the control of C. albicans in the vaginal mucosa. Il17ra(⁻/⁻) mice showed fungal kinetics similar to WT animals early in infection and a faster decline at later stages. These observations align with studies demonstrating that IL-17 –mediated responses, while essential for antifungal protection at oral and intestinal sites [13,14], play a limited or dispensable role in VVC [15,16]. The tolerogenic nature of the vaginal mucosa, together with estrogen-mediated modulation, could explain the limited contribution of IL-17 to fungal clearance [57,58]. Despite this, IL-17RA signaling contributes to the regulation of BD. Our results identify mBD3 as the defensin most strongly associated with IL-17RA-dependent signaling during vaginal C. albicans infection. The marked reduction of mBD3 expression in Il17ra(⁻/⁻) mice, compared with the partial effect observed for mBD1, suggests that IL-17–mediated pathways play a dominant and selective role in driving BD-3 transcription. Notably, this regulatory role was observed even though IL-17 remained undetectable in CVL samples from infected mice, suggesting that cytokine production is stringently controlled within the vaginal mucosa and may occur at levels below detection in vaginal fluids. Such a scenario could reflect transient or spatially restricted IL-17 expression, limited recruitment of IL-17–producing cells to the vaginal tissue, or modulation by local immune regulatory mechanisms [26,59,60]. Collectively, these observations emphasize the finely tuned and compartmentalized nature of IL-17 responses in the mucosal vaginal tract.
In our model, the absence of IL-17RA signaling strongly impaired mBD3 expression, supporting a central role for IL-17-mediated pathways in its induction during vaginal C. albicans infection. Similar observations have been reported in other mucosal sites where IL-17 enhances BDs expression. Verma et al. [61] demonstrated that Th17 cytokines can activate epithelial and immune cells to release alarmins and AMPs, including BDs. Likewise, IL-17 stimulation increased mBD3 expression in murine colonic epithelial cells [62], and in nasal tissue, IL-17A promoted mBD3 upregulation in response to Staphylococcus aureus colonization [63]. Also, it was reported that IL-17 potently induces BD through epithelial activation of NF-κB, MAPK, and C/EBP pathways, which are essential for the transcription of AMPs genes such as Defb2 and Defb3 [64,65]. The partial induction of mBD1 in Il17ra⁻/⁻ mice suggests that additional regulatory pathways, including NLRP3/IL-1β axis, contribute to its expression, as also demonstrated in this work. Complementary to this, prior studies have demonstrated that IFN-α activated during acute viral challenge rapidly upregulates hBD1 in human immune cells [66], indicating a direct link between IFN-I signaling and BD-1 transcriptional induction. Interestingly, other authors and our group have reported that IFN-I is relevant in mucosal defense in Candida vaginitis [25,26,67]. Overall, the data presented here provide a framework for future mechanistic studies of defensin regulation during VVC.
In summary, this study reveals a dynamic interplay between host epithelial defenses and C. albicans during vaginal infection, defining distinct regulatory programs for constitutive and inducible BDs throughout the course of VVC. Our findings demonstrate that cytokine-driven pathways differentially control mBD1 and mBD3 in the vaginal mucosa, uncovering nonredundant mechanisms of epithelial antimicrobial regulation. Specifically, we propose a biphasic model in which NLRP3/IL-1β signaling predominates early mBD1 induction, whereas IL-17RA-dependent pathways are preferentially associated with mBD3 regulation during vaginal infection. By integrating genetic deficiency models with temporal analyses, this work provides strong evidence supporting a regulatory role for NLRP3/IL-1β and IL-17RA pathways in epithelial AMPs expression during vaginal candidiasis and provides new insight into the temporal coordination of mucosal innate immunity, laying a conceptual foundation for the development of targeted or combinatorial therapeutic strategies against VVC.
Author Contributions
Conceptualization, S.C.A. and C.E.S.; methodology, S.C.A., E.R., M.S.M. P.A.I. and C.M.R.; investigation, F.O.R., J.P.C., P.I and C.E.S; formal analysis, S.C.A., E.R., C.M.R., P.I. and C.E.S; data curation, S.C.A., E.R. and C.M.R.; software, S.C.A., C.M.R. and P.I.; visualization, S.C.A. and E.R.; validation, S.C.A., E.R. and C.M.R; supervision, F.O.R., J.P.C. and C.E.S.; project administration, P.A.I. and C.E.S.; funding acquisition, F.O.R., P.I., J.P.C. and C.E.S.; writing—original draft preparation, S.C.A. and C.E.S.; writing—review and editing, S.C.A., E.R., C.M.R., P.A.I., P.I., F.O.R., J.P.C. and C.E.S.; resources, P.I., F.O.R., J.P.C. and C.E.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from Agencia Nacional de Promoción Científica y Tecnológica- FONCyT (PICT-2019–2012) and -FONCyT (PICT 2021 00333) (Argentina); SECyT-UNC (30720150100934CB) and SECyT-UNC (33620230100340CB); CONICET (PIP 11220220100579CO) (Argentina). AltaMed Health Service Corporation (USA).
Institutional Review Board Statement
All experimental procedures were approved and conducted in accordance with the guidelines of the Animal Experimentation Ethics Committee of the Faculty of Chemical Sciences, National University of Córdoba (Approval number: RD-2022-2789-E-UNC-DEC#FCQ), and strictly followed the recommendations of the Guide to the Care and Use of Experimental Animals published by the Canadian Council on Animal Care (OLAW Assurance number A5802-01).
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding authors.
Acknowledgments
We acknowledge the staff from Microscopy Core, Cell Culture, Histotechnology, and Animal facility from the Department of Clinical Biochemistry and CIBICI-CONICET for their excellent technical assistance.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Sobel JD. Vulvovaginal candidosis. The Lancet 2007;369:1961–71. [CrossRef]
- Achkar JM, Fries BC. Candida Infections of the Genitourinary Tract. Clinical Microbiology Reviews 2010;23:253–73. [CrossRef]
- Valentine M, Wilson D, Gresnigt MS, Hube B. Vaginal Candida albicans infections: host–pathogen–microbiome interactions. FEMS Microbiology Reviews 2025;49. [CrossRef]
- Denning DW, Kneale M, Sobel JD, Rautemaa-Richardson R. Global burden of recurrent vulvovaginal candidiasis: a systematic review. The Lancet Infectious Diseases 2018;18:e339–47. [CrossRef]
- MacAlpine J, Lionakis MS. Host-microbe interaction paradigms in acute and recurrent vulvovaginal candidiasis. Cell Host & Microbe 2024;32:1654–67. [CrossRef]
- Rosati D, Bruno M, Jaeger M, ten Oever J, Netea MG. Recurrent Vulvovaginal Candidiasis: An Immunological Perspective. Microorganisms 2020;8:144. [CrossRef]
- Peters BM, Palmer GE, Nash AK, Lilly EA, Fidel PL, Noverr MC. Fungal Morphogenetic Pathways Are Required for the Hallmark Inflammatory Response during Candida albicans Vaginitis. Infection and Immunity 2014;82:532–43. [CrossRef]
- Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence 2013;4:119–28. [CrossRef]
- Roselletti E, Monari C, Sabbatini S, Perito S, Vecchiarelli A, Sobel JD, et al. A Role for Yeast/Pseudohyphal Cells of Candida albicans in the Correlated Expression of NLRP3 Inflammasome Inducers in Women With Acute Vulvovaginal Candidiasis. Frontiers in Microbiology 2019;10. [CrossRef]
- Yano J, Sobel JD, Nyirjesy P, Sobel R, Williams VL, Yu Q, et al. Current patient perspectives of vulvovaginal candidiasis: incidence, symptoms, management and post-treatment outcomes. BMC Women’s Health 2019;19:48. [CrossRef]
- Ardizzoni A, Wheeler RT, Pericolini E. It Takes Two to Tango: How a Dysregulation of the Innate Immunity, Coupled With Candida Virulence, Triggers VVC Onset. Frontiers in Microbiology 2021;12. [CrossRef]
- Cheng KO, Montaño DE, Zelante T, Dietschmann A, Gresnigt MS. Inflammatory cytokine signalling in vulvovaginal candidiasis: a hot mess driving immunopathology. Oxford Open Immunology 2024;5. [CrossRef]
- Conti HR, Shen F, Nayyar N, Stocum E, Sun JN, Lindemann MJ, et al. Th17 cells and IL-17 receptor signaling are essential for mucosal host defense against oral candidiasis. Journal of Experimental Medicine 2009;206:299–311. [CrossRef]
- Dixon BREA, Radin JN, Piazuelo MB, Contreras DC, Algood HMS. IL-17a and IL-22 Induce Expression of Antimicrobials in Gastrointestinal Epithelial Cells and May Contribute to Epithelial Cell Defense against Helicobacter pylori. PLOS ONE 2016;11:e0148514. [CrossRef]
- Pietrella D, Rachini A, Pines M, Pandey N, Mosci P, Bistoni F, et al. Th17 Cells and IL-17 in Protective Immunity to Vaginal Candidiasis. PLoS ONE 2011;6:e22770. [CrossRef]
- Peters BM, Coleman BM, Willems HME, Barker KS, Aggor FEY, Cipolla E, et al. The Interleukin (IL) 17R/IL-22R Signaling Axis Is Dispensable for Vulvovaginal Candidiasis Regardless of Estrogen Status. The Journal of Infectious Diseases 2020;221:1554–63. [CrossRef]
- Schutte BC, Mitros JP, Bartlett JA, Walters JD, Jia HP, Welsh MJ, et al. Discovery of five conserved β-defensin gene clusters using a computational search strategy. Proceedings of the National Academy of Sciences 2002;99:2129–33. [CrossRef]
- Shelley JR, Davidson DJ, Dorin JR. The Dichotomous Responses Driven by β-Defensins. Frontiers in Immunology 2020;11. [CrossRef]
- Yang D, Chertov O, Bykovskaia SN, Chen Q, Buffo MJ, Shogan J, et al. β-Defensins: Linking Innate and Adaptive Immunity Through Dendritic and T Cell CCR6. Science 1999;286:525–8. [CrossRef]
- Shimbo M, Ito N, Kadonosono K. [Investigation of beta-D-glucan values in the vitreous]. Nippon Ganka Gakkai Zasshi 2002;106:579–82.
- Funderburg N, Lederman MM, Feng Z, Drage MG, Jadlowsky J, Harding C V., et al. Human β-defensin-3 activates professional antigen-presenting cells via Toll-like receptors 1 and 2. Proceedings of the National Academy of Sciences 2007;104:18631–5. [CrossRef]
- Fesahat F, Firouzabadi AM, Zare-Zardini H, Imani M. Roles of Different β-Defensins in the Human Reproductive System: A Review Study. American Journal of Men’s Health 2023;17. [CrossRef]
- Miró MS, Caeiro JP, Rodriguez E, Vargas L, Vigezzi C, Icely PA, et al. Candida albicans Modulates Murine and Human Beta Defensin-1 during Vaginitis. Journal of Fungi 2021;8:20. [CrossRef]
- Vigezzi C, Icely PA, Dudiuk C, Rodríguez E, Miró MS, Castillo GDV, et al. Frequency, virulence factors and antifungal susceptibility of Candida parapsilosis species complex isolated from patients with candidemia in the central region of Argentina. Journal de Mycologie Médicale 2019:100907. [CrossRef]
- Pekmezovic M, Hovhannisyan H, Gresnigt MS, Iracane E, Oliveira-Pacheco J, Siscar-Lewin S, et al. Candida pathogens induce protective mitochondria-associated type I interferon signalling and a damage-driven response in vaginal epithelial cells. Nature Microbiology 2021;6:643–57. [CrossRef]
- Rodriguez E, Savid-Frontera C, Angiolini SC, Hernáez-Sánchez ML, Miró MS, Viano ME, et al. Type-I interferons in Vulvovaginal Candidiasis: Mechanism of epithelial early defense and immune regulation against Candida albicans. Mucosal Immunology 2025;18:1124–38. [CrossRef]
- Miró MS, Rodríguez E, Vigezzi C, Icely PA, Gonzaga de Freitas Araújo M, Riera FO, et al. Candidiasis vulvovaginal: una antigua enfermedad con nuevos desafíos. Revista Iberoamericana de Micología 2017;34:65–71. [CrossRef]
- Yano J, Palmer GE, Eberle KE, Peters BM, Vogl T, McKenzie AN, et al. Vaginal Epithelial Cell-Derived S100 Alarmins Induced by Candida albicans via Pattern Recognition Receptor Interactions Are Sufficient but Not Necessary for the Acute Neutrophil Response during Experimental Vaginal Candidiasis. Infection and Immunity 2014;82:783–92. [CrossRef]
- de Freitas Araújo MG, Pacífico M, Vilegas W, Dos Santos LC, Icely PA, Miró MS, et al. Evaluation of Syngonanthus nitens (Bong.) Ruhl. extract as antifungal and in treatment of vulvovaginal candidiasis. Medical Mycology 2013;51:673–82. [CrossRef]
- Renna MS, Correa SG, Porporatto C, Figueredo CM, Aoki MP, Paraje MG, et al. Hepatocellular apoptosis during Candida albicans colonization: involvement of TNF- and infiltrating Fas-L positive lymphocytes. International Immunology 2006;18:1719–28. [CrossRef]
- Yarbrough VL, Winkle S, Herbst-Kralovetz MM. Antimicrobial peptides in the female reproductive tract: a critical component of the mucosal immune barrier with physiological and clinical implications. Human Reproduction Update 2015;21:353–77. [CrossRef]
- Krishnakumari V, Rangaraj N, Nagaraj R. Antifungal Activities of Human Beta-Defensins HBD-1 to HBD-3 and Their C-Terminal Analogs Phd1 to Phd3. Antimicrobial Agents and Chemotherapy 2009;53:256–60. [CrossRef]
- Bruno VM, Shetty AC, Yano J, Fidel PL, Noverr MC, Peters BM. Transcriptomic Analysis of Vulvovaginal Candidiasis Identifies a Role for the NLRP3 Inflammasome. MBio 2015;6. [CrossRef]
- Sobel JD. Recurrent vulvovaginal candidiasis. American Journal of Obstetrics and Gynecology 2016;214:15–21. [CrossRef]
- Fidel PL, Barousse M, Espinosa T, Ficarra M, Sturtevant J, Martin DH, et al. An Intravaginal Live Candida Challenge in Humans Leads to New Hypotheses for the Immunopathogenesis of Vulvovaginal Candidiasis. Infection and Immunity 2004;72:2939–46. [CrossRef]
- Yano J, Peters BM, Noverr MC, Fidel PL. Novel Mechanism behind the Immunopathogenesis of Vulvovaginal Candidiasis: “Neutrophil Anergy.” Infection and Immunity 2018;86. [CrossRef]
- Yano J, Noverr MC, Fidel PL. Vaginal Heparan Sulfate Linked to Neutrophil Dysfunction in the Acute Inflammatory Response Associated with Experimental Vulvovaginal Candidiasis. MBio 2017;8. [CrossRef]
- Fusco A, Savio V, Donniacuo M, Perfetto B, Donnarumma G. Antimicrobial Peptides Human Beta-Defensin-2 and -3 Protect the Gut During Candida albicans Infections Enhancing the Intestinal Barrier Integrity: In Vitro Study. Frontiers in Cellular and Infection Microbiology 2021;11. [CrossRef]
- Hickey DK, Fahey J V, Wira CR. Mouse estrous cycle regulation of vaginal versus uterine cytokines, chemokines, α-/β-defensins and TLRs. Innate Immunity 2013;19:121–31. [CrossRef]
- Prado-Montes de Oca E. Human β-defensin 1: A restless warrior against allergies, infections and cancer. The International Journal of Biochemistry & Cell Biology 2010;42:800–4. [CrossRef]
- Burd RS, Furrer JL, Sullivan J, Smith AL. Murine ??-Defensin-3 Is an Inducible Peptide with Limited Tissue Expression and Broad-Spectrum Antimicrobial Activity. Shock 2002;18:461–4. [CrossRef]
- Tomalka J, Azodi E, Narra HP, Patel K, O’Neill S, Cardwell C, et al. β-Defensin 1 Plays a Role in Acute Mucosal Defense against Candida albicans. The Journal of Immunology 2015;194:1788–95. [CrossRef]
- Palrasu M, Marudamuthu A, Kakar K, Hamida H, Thada S, Gupta R, et al. AhR-Dependent Induction of β-Defensin 1 in Colonic Epithelial Cells Regulates Cross-Talk between Gut Microbiota and Immune Response Leading to Attenuation of Colitis. Advanced Science 2025;12. [CrossRef]
- Semple F, Dorin JR. β-Defensins: Multifunctional Modulators of Infection, Inflammation and More? Journal of Innate Immunity 2012;4:337–48. [CrossRef]
- Noda-Nicolau NM, Bastos LB, Bolpetti AN, Pinto GVS, Marcolino LD, Marconi C, et al. Cervicovaginal Levels of Human β-Defensin 1, 2, 3, and 4 of Reproductive-Aged Women With Chlamydia trachomatis Infection. Journal of Lower Genital Tract Disease 2017;21:189–92. [CrossRef]
- Zhai Y-J, Feng Y, Ma X, Ma F. Defensins: defenders of human reproductive health. Human Reproduction Update 2023;29:126–54. [CrossRef]
- Chakraborty K, Ghosh S, Koley H, Mukhopadhyay AK, Ramamurthy T, Saha DR, et al. Bacterial exotoxins downregulate cathelicidin (hCAP-18/LL-37) and human β-defensin 1 (HBD-1) expression in the intestinal epithelial cells. Cellular Microbiology 2008;10:2520–37. [CrossRef]
- Bauer B, Pang E, Holland C, Kessler M, Bartfeld S, Meyer TF. The Helicobacter pylori Virulence Effector CagA Abrogates Human β-Defensin 3 Expression via Inactivation of EGFR Signaling. Cell Host & Microbe 2012;11:576–86. [CrossRef]
- Patel SR, Smith K, Letley DP, Cook KW, Memon AA, Ingram RJM, et al. H elicobacter pylori downregulates expression of human β-defensin 1 in the gastric mucosa in a type IV secretion-dependent fashion. Cellular Microbiology 2013;15:2080–92. [CrossRef]
- Hise AG, Tomalka J, Ganesan S, Patel K, Hall BA, Brown GD, et al. An Essential Role for the NLRP3 Inflammasome in Host Defense against the Human Fungal Pathogen Candida albicans. Cell Host & Microbe 2009;5:487–97. [CrossRef]
- Jaeger M, Carvalho A, Cunha C, Plantinga TS, van de Veerdonk F, Puccetti M, et al. Association of a variable number tandem repeat in the NLRP3 gene in women with susceptibility to RVVC. European Journal of Clinical Microbiology & Infectious Diseases 2016;35:797–801. [CrossRef]
- Pietrella D, Pandey N, Gabrielli E, Pericolini E, Perito S, Kasper L, et al. Secreted aspartic proteases of Candida albicans activate the NLRP3 inflammasome. European Journal of Immunology 2013;43:679–92. [CrossRef]
- Pericolini E, Gabrielli E, Amacker M, Kasper L, Roselletti E, Luciano E, et al. Secretory Aspartyl Proteinases Cause Vaginitis and Can Mediate Vaginitis Caused by Candida albicans in Mice. MBio 2015;6. [CrossRef]
- Pahl R, Brunke G, Steubesand N, Schubert S, Böttner M, Wedel T, et al. IL-1β and ADAM17 are central regulators of β-defensin expression in Candida esophagitis. American Journal of Physiology-Gastrointestinal and Liver Physiology 2011;300:G547–53. [CrossRef]
- Zaga-Clavellina V, Martha RV-M, Flores-Espinosa P. In Vitro Secretion Profile of Pro-Inflammatory Cytokines IL-1β, TNF-α, IL-6, and of Human Beta-Defensins (HBD)-1, HBD-2, and HBD-3 from Human Chorioamniotic Membranes After Selective Stimulation with Gardnerella vaginalis. American Journal of Reproductive Immunology 2012;67:34–43. [CrossRef]
- Mao F-Y, Lv Y-P, Hao C-J, Teng Y-S, Liu Y-G, Cheng P, et al. Helicobacter pylori–Induced Rev-erbα Fosters Gastric Bacteria Colonization by Impairing Host Innate and Adaptive Defense. Cellular and Molecular Gastroenterology and Hepatology 2021;12:395–425. [CrossRef]
- Fidel PL, Cutright J, Steele C. Effects of Reproductive Hormones on Experimental Vaginal Candidiasis. Infection and Immunity 2000;68:651–7. [CrossRef]
- Yano J, Kolls JK, Happel KI, Wormley F, Wozniak KL, Fidel PL. The Acute Neutrophil Response Mediated by S100 Alarmins during Vaginal Candida Infections Is Independent of the Th17-Pathway. PLoS ONE 2012;7:e46311. [CrossRef]
- Zelante T, Iannitti RG, Cunha C, De Luca A, Giovannini G, Pieraccini G, et al. Tryptophan Catabolites from Microbiota Engage Aryl Hydrocarbon Receptor and Balance Mucosal Reactivity via Interleukin-22. Immunity 2013;39:372–85. [CrossRef]
- Lee AJ, Ashkar AA. The Dual Nature of Type I and Type II Interferons. Frontiers in Immunology 2018;9. [CrossRef]
- Verma A, Wuthrich M, Deepe G, Klein B. Adaptive Immunity to Fungi. Cold Spring Harbor Perspectives in Medicine 2015;5:a019612–a019612. [CrossRef]
- Rahman A, Fahlgren A, Sundstedt C, Hammarström S, Danielsson Å, Hammarström M-L. Chronic colitis induces expression of β-defensins in murine intestinal epithelial cells. Clinical and Experimental Immunology 2010;163:123–30. [CrossRef]
- Archer NK, Adappa ND, Palmer JN, Cohen NA, Harro JM, Lee SK, et al. Interleukin-17A (IL-17A) and IL-17F Are Critical for Antimicrobial Peptide Production and Clearance of Staphylococcus aureus Nasal Colonization. Infection and Immunity 2016;84:3575–83. [CrossRef]
- Kao C-Y, Chen Y, Thai P, Wachi S, Huang F, Kim C, et al. IL-17 Markedly Up-Regulates β-Defensin-2 Expression in Human Airway Epithelium via JAK and NF-κB Signaling Pathways. The Journal of Immunology 2004;173:3482–91. [CrossRef]
- Liang SC, Tan X-Y, Luxenberg DP, Karim R, Dunussi-Joannopoulos K, Collins M, et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. The Journal of Experimental Medicine 2006;203:2271–9. [CrossRef]
- Corleis B, Lisanti AC, Körner C, Schiff AE, Rosenberg ES, Allen TM, et al. Early type I Interferon response induces upregulation of human β-defensin 1 during acute HIV-1 infection. PLOS ONE 2017;12:e0173161. [CrossRef]
- Smeekens SP, Ng A, Kumar V, Johnson MD, Plantinga TS, van Diemen C, et al. Functional genomics identifies type I interferon pathway as central for host defense against Candida albicans. Nature Communications 2013;4:1342. [CrossRef]
Figure 1.
Establishment of Infection and Local Inflammatory Profile During Experimental Vaginal Candidiasis in C57BL/6 (WT) Mice. (A) Schematic representation of the murine VVC model in WT mice. (B) Individual (top) and mean (bottom) intravaginal fungal load, expressed as C. albicans CFU per mL of CVL, in Infected WT mice at days 2, 4, and 8 post infection. Values represent the mean ± SEM (n ≥ 14). Data were analyzed using the Mann-Whitney test at each specific time point. *p<0.05. (C) Representative images of the cell populations present in CVL cytospin from Uninfected or Infected mice at day 2 post infection, stained with May-Grünwald Giemsa (x200 magnification). Red arrows indicate the ECs, and yellow arrows indicate the PMNs. (D) PMNs counts from CVL cytospin preparations of Uninfected (gray line) and Infected (black line) WT mice at days 2, 4, and 8 post infection, stained with May-Grunwald Giemsa (x40 magnification). Values represent the mean ± SEM (n ≥ 10). (E) IL-1β levels in CVL samples from Uninfected (gray line) and Infected (black line) mice at days 2, 4, and 8 post infection, determined by ELISA. Values represent the mean ± SEM (n ≥ 6). (D-E) Data were analyzed using two-way ANOVA. Black asterisks indicate significant differences within the Infected group relative to day 2. Gray asterisks indicate significant differences between the Uninfected and Infected groups at the same time point. *p<0.05. **p<0.01. #p=0.06 (Uninfected and Infected, day 2). WT: wild-type; SC: subcutaneous; IV: intravaginal; CFU: Colony Forming Units; CVL: Cervicovaginal lavage; PMNs: Polymorphonuclear neutrophils; ECs: Epithelial cells. SEM = standard error of the mean; ANOVA = analysis of variance.
Figure 1.
Establishment of Infection and Local Inflammatory Profile During Experimental Vaginal Candidiasis in C57BL/6 (WT) Mice. (A) Schematic representation of the murine VVC model in WT mice. (B) Individual (top) and mean (bottom) intravaginal fungal load, expressed as C. albicans CFU per mL of CVL, in Infected WT mice at days 2, 4, and 8 post infection. Values represent the mean ± SEM (n ≥ 14). Data were analyzed using the Mann-Whitney test at each specific time point. *p<0.05. (C) Representative images of the cell populations present in CVL cytospin from Uninfected or Infected mice at day 2 post infection, stained with May-Grünwald Giemsa (x200 magnification). Red arrows indicate the ECs, and yellow arrows indicate the PMNs. (D) PMNs counts from CVL cytospin preparations of Uninfected (gray line) and Infected (black line) WT mice at days 2, 4, and 8 post infection, stained with May-Grunwald Giemsa (x40 magnification). Values represent the mean ± SEM (n ≥ 10). (E) IL-1β levels in CVL samples from Uninfected (gray line) and Infected (black line) mice at days 2, 4, and 8 post infection, determined by ELISA. Values represent the mean ± SEM (n ≥ 6). (D-E) Data were analyzed using two-way ANOVA. Black asterisks indicate significant differences within the Infected group relative to day 2. Gray asterisks indicate significant differences between the Uninfected and Infected groups at the same time point. *p<0.05. **p<0.01. #p=0.06 (Uninfected and Infected, day 2). WT: wild-type; SC: subcutaneous; IV: intravaginal; CFU: Colony Forming Units; CVL: Cervicovaginal lavage; PMNs: Polymorphonuclear neutrophils; ECs: Epithelial cells. SEM = standard error of the mean; ANOVA = analysis of variance.
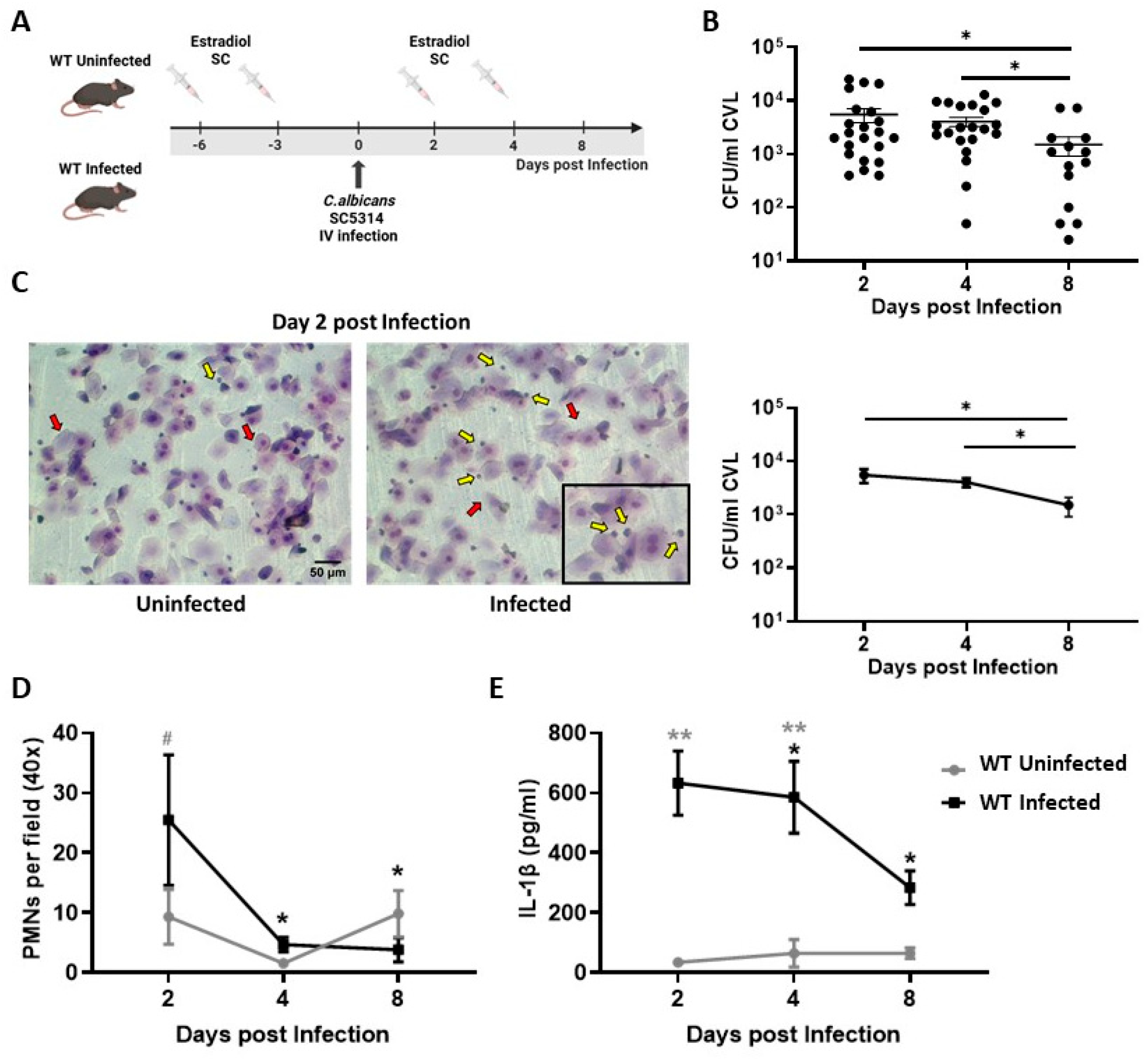
Figure 2.
Differential expression of Beta Defensin-1 (mBD1) in ECs from cervicovaginal lavage cells during VVC. (A) Representative images of CVL cytospin preparations from Uninfected and Infected WT mice at days 2, 4, and 8 post infection. mBD1 protein expression was detected by indirect immunofluorescence (IF) using a rabbit anti-mouse BD-1 primary antibody and an Alexa Fluor 488-conjugated anti-rabbit IgG secondary antibody. Images were captured with a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD1 expression appears in green. (B) Semi-quantification of mBD1 fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change. Analysis was performed using ImageJ software (n≥4 mice per time point). Values represent the mean ± SEM. Data were analyzed using one-way ANOVA. ++++p<0.0001 (Infected vs Uninfected, days 2, 4, and 8). ****p<0.0001 (Infected, days 2 and 4 vs. day 8). #### p<0.0001 (Uninfected, days 4 and 8 vs. day 2). (C) Fluorescence intensity in the Infected group expressed as a percentage change in mBD1 relative to the Uninfected group at days 2, 4, and 8 post infection. CVL: Cervicovaginal lavage. WT: wild-type. SEM = standard error of the mean; ANOVA = analysis of variance.
Figure 2.
Differential expression of Beta Defensin-1 (mBD1) in ECs from cervicovaginal lavage cells during VVC. (A) Representative images of CVL cytospin preparations from Uninfected and Infected WT mice at days 2, 4, and 8 post infection. mBD1 protein expression was detected by indirect immunofluorescence (IF) using a rabbit anti-mouse BD-1 primary antibody and an Alexa Fluor 488-conjugated anti-rabbit IgG secondary antibody. Images were captured with a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD1 expression appears in green. (B) Semi-quantification of mBD1 fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change. Analysis was performed using ImageJ software (n≥4 mice per time point). Values represent the mean ± SEM. Data were analyzed using one-way ANOVA. ++++p<0.0001 (Infected vs Uninfected, days 2, 4, and 8). ****p<0.0001 (Infected, days 2 and 4 vs. day 8). #### p<0.0001 (Uninfected, days 4 and 8 vs. day 2). (C) Fluorescence intensity in the Infected group expressed as a percentage change in mBD1 relative to the Uninfected group at days 2, 4, and 8 post infection. CVL: Cervicovaginal lavage. WT: wild-type. SEM = standard error of the mean; ANOVA = analysis of variance.
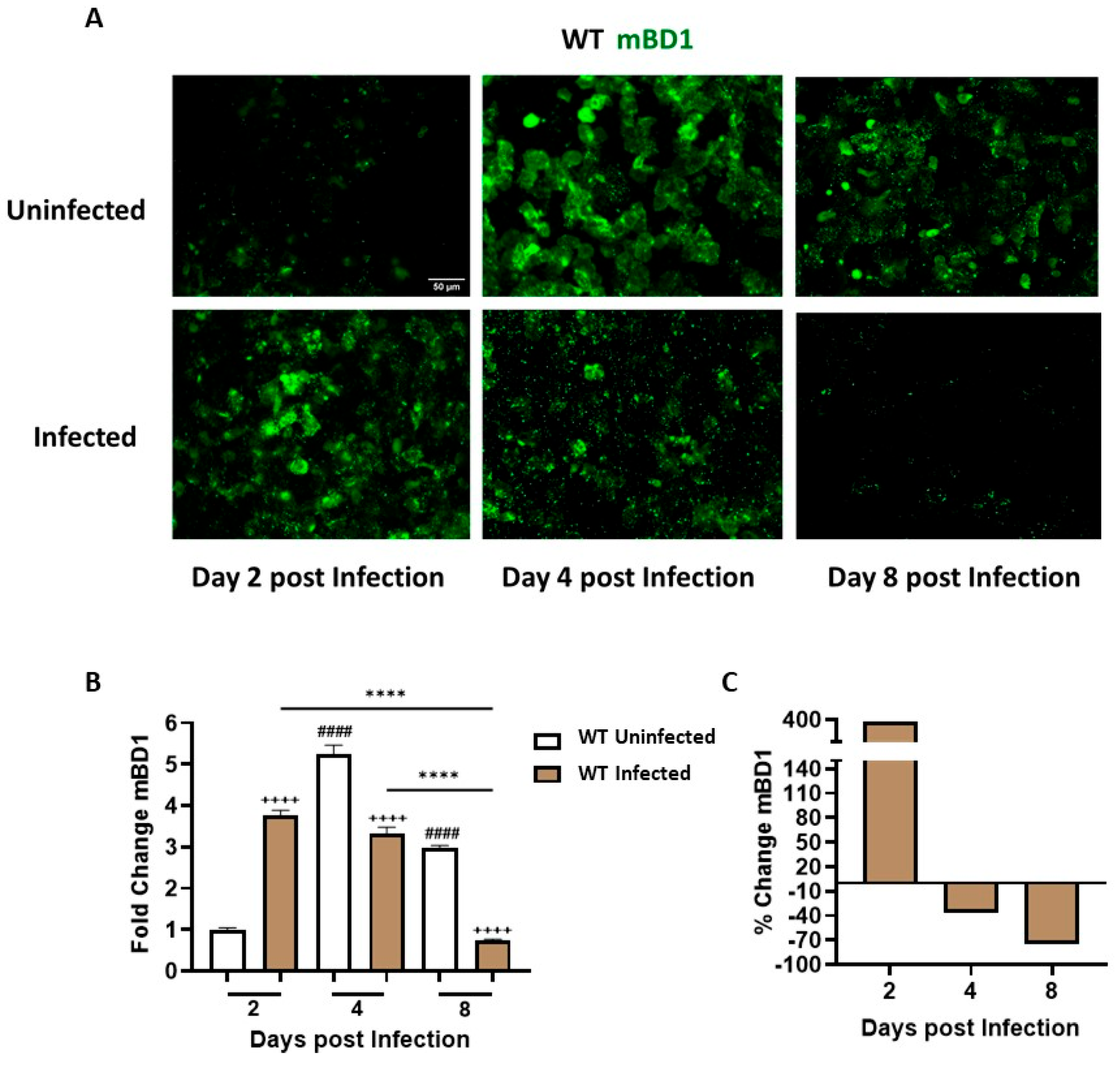
Figure 3.
Differential expression of Beta Defensin-3 (mBD3) in ECs from cervicovaginal lavage cells during VVC. (A) Representative images of CVL preparations from Uninfected and Infected WT mice at days 2, 4, and 8 post infection. mBD3 protein expression in ECs was detected by indirect immunofluorescence (IF) using a rabbit anti-mouse BD-3 primary antibody and an Alexa Fluor 488-conjugated anti-rabbit IgG secondary antibody. Images were obtained with a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD3 expression appears in green. (B) Semi-quantification of mBD3 fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change. Analysis was performed with ImageJ software (n≥4 mice per time point). Values represent the mean ± SEM. Data were analyzed using one-way ANOVA. ++++p<0.0001 (Infected vs Uninfected days 2, 4, and 8). **p<0.01 (Infected day 4 vs day 8). ***p<0.001 (Infected day 2 vs day 4). ****p<0.0001 (Infected day 2 vs day 8). #### p<0.0001 (Uninfected days 4 and 8 vs day 2). (C) Fluorescence intensity of the Infected group expressed as a percentage change in mBD3 relative to the Uninfected group at days 2, 4, and 8 post infection. CVL: cervicovaginal lavage. WT: wild-type. SEM = standard error of the mean; ANOVA = analysis of variance.
Figure 3.
Differential expression of Beta Defensin-3 (mBD3) in ECs from cervicovaginal lavage cells during VVC. (A) Representative images of CVL preparations from Uninfected and Infected WT mice at days 2, 4, and 8 post infection. mBD3 protein expression in ECs was detected by indirect immunofluorescence (IF) using a rabbit anti-mouse BD-3 primary antibody and an Alexa Fluor 488-conjugated anti-rabbit IgG secondary antibody. Images were obtained with a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD3 expression appears in green. (B) Semi-quantification of mBD3 fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change. Analysis was performed with ImageJ software (n≥4 mice per time point). Values represent the mean ± SEM. Data were analyzed using one-way ANOVA. ++++p<0.0001 (Infected vs Uninfected days 2, 4, and 8). **p<0.01 (Infected day 4 vs day 8). ***p<0.001 (Infected day 2 vs day 4). ****p<0.0001 (Infected day 2 vs day 8). #### p<0.0001 (Uninfected days 4 and 8 vs day 2). (C) Fluorescence intensity of the Infected group expressed as a percentage change in mBD3 relative to the Uninfected group at days 2, 4, and 8 post infection. CVL: cervicovaginal lavage. WT: wild-type. SEM = standard error of the mean; ANOVA = analysis of variance.
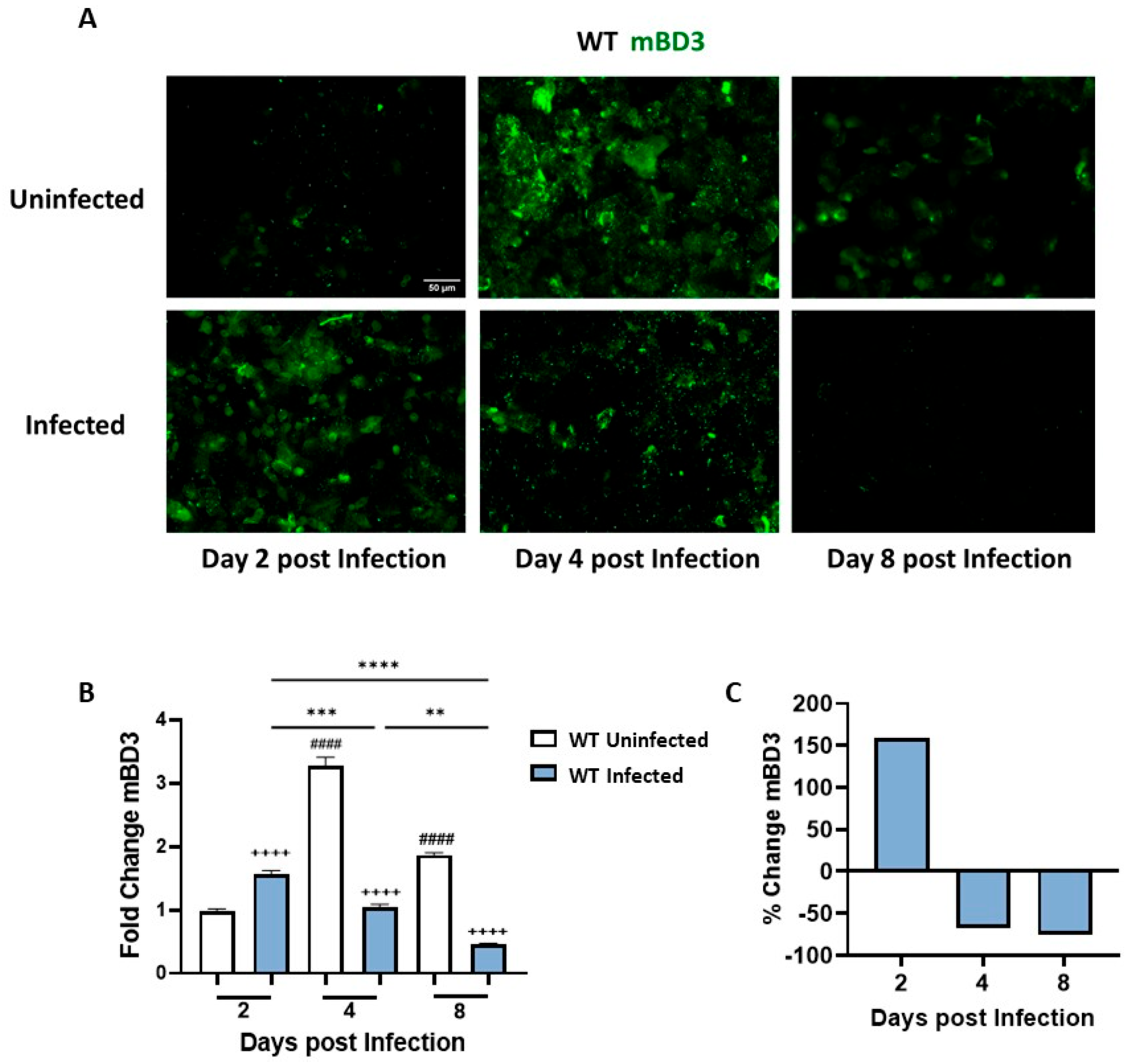
Figure 4.
Regulation of Beta Defensin-1 (mBD1) and Beta Defensin-3 (mBD3) in Nlrp3(-/-) mice during VVC. (A) Individual intravaginal fungal load counts, expressed as C. albicans CFUs/mL of CVL, in Infected Nlrp3(-/-) mice at days 2, 4, and 8 post infection (left). Comparative analysis of CFU/mL between WT (black line) and Nlrp3(-/-) (red line) mice throughout infection kinetics (right). Values represent the mean ± SEM (n ≥ 3). Data were analyzed using the Mann-Whitney test at each specific time point and two-way ANOVA for comparisons between WT and Nlrp3(-/-) animals. (B-D) Representative images of CVL cytospin preparations from WT and Nlrp3(-/-) mice, Uninfected and Infected at day 2 post-infection. ECs were stained by IF to detect mBD1 (panel B) or mBD3 (panel D) expression. Images were captured using a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD1 and mBD3 expression is shown in green (top panels). (C-E) The lower panels show semi-quantification of mBD1 (panel C) and mBD3 (panel E) fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change in WT and Nlrp3(-/-) mice, comparing infected and uninfected groups, using ImageJ software (n=5 mice). Values represent the mean ± SEM. Data were analyzed using Student's t-test. ****p<0.0001. WT: wild-type; CFU: Colony Forming Units; CVL: Cervicovaginal lavage; ECs: Epithelial cells. SEM = standard error of the mean; ANOVA = analysis of variance.
Figure 4.
Regulation of Beta Defensin-1 (mBD1) and Beta Defensin-3 (mBD3) in Nlrp3(-/-) mice during VVC. (A) Individual intravaginal fungal load counts, expressed as C. albicans CFUs/mL of CVL, in Infected Nlrp3(-/-) mice at days 2, 4, and 8 post infection (left). Comparative analysis of CFU/mL between WT (black line) and Nlrp3(-/-) (red line) mice throughout infection kinetics (right). Values represent the mean ± SEM (n ≥ 3). Data were analyzed using the Mann-Whitney test at each specific time point and two-way ANOVA for comparisons between WT and Nlrp3(-/-) animals. (B-D) Representative images of CVL cytospin preparations from WT and Nlrp3(-/-) mice, Uninfected and Infected at day 2 post-infection. ECs were stained by IF to detect mBD1 (panel B) or mBD3 (panel D) expression. Images were captured using a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD1 and mBD3 expression is shown in green (top panels). (C-E) The lower panels show semi-quantification of mBD1 (panel C) and mBD3 (panel E) fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change in WT and Nlrp3(-/-) mice, comparing infected and uninfected groups, using ImageJ software (n=5 mice). Values represent the mean ± SEM. Data were analyzed using Student's t-test. ****p<0.0001. WT: wild-type; CFU: Colony Forming Units; CVL: Cervicovaginal lavage; ECs: Epithelial cells. SEM = standard error of the mean; ANOVA = analysis of variance.
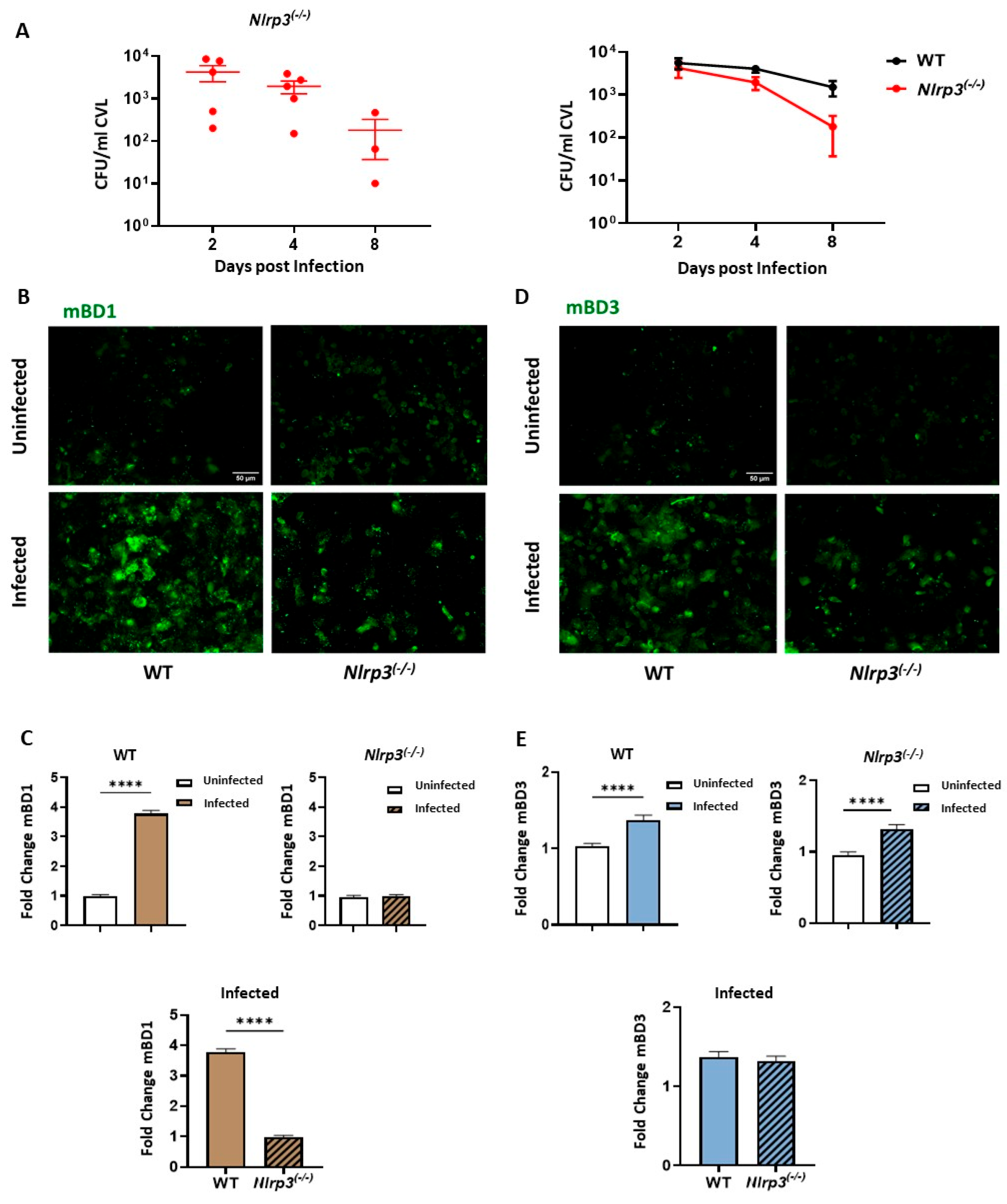
Figure 5.
Regulation of Beta Defensin-1 (mBD1) and Beta Defensin-3 (mBD3) in Il17ra(-/-) mice during VVC. (A) Individual intravaginal fungal load counts, expressed as C. albicans CFU per mL CVL, in Infected Il17ra(-/-) mice at days 2, 4, and 8 post infection (left). Comparative analysis of CFU/mL between WT (black line) and Il17ra(-/-) (blue line) mice during infection kinetics (right). Values represent the mean ± SEM (n ≥ 3). Data were analyzed using the Mann-Whitney test at each specific time point and two-way ANOVA for comparisons between WT and Il17ra(-/-) animals. *p<0.05. (B-D) Representative images of CVL cytospin preparations from WT and Il17ra(-/-) mice, Uninfected and Infected at day 2 post infection. ECs were stained by IF to detect mBD1 (panel B) or mBD3 (panel D) expression. Images were captured with a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD1 and mBD3 expression is shown in green (top panels). (C-E) The lower panels show the semi-quantification of mBD1 (panel C) and mBD3 (panel E) fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change in WT and Il17ra(-/-) mice, comparing infected and uninfected groups, using ImageJ software (n=4 mice). Values represent the mean ± SEM. Data were analyzed using Student's t-test. ****p<0.0001. WT: wild-type; CFU: Colony Forming Units; CVL: Cervicovaginal lavage; ECs: Epithelial cells. SEM = standard error of the mean; ANOVA = analysis of variance.
Figure 5.
Regulation of Beta Defensin-1 (mBD1) and Beta Defensin-3 (mBD3) in Il17ra(-/-) mice during VVC. (A) Individual intravaginal fungal load counts, expressed as C. albicans CFU per mL CVL, in Infected Il17ra(-/-) mice at days 2, 4, and 8 post infection (left). Comparative analysis of CFU/mL between WT (black line) and Il17ra(-/-) (blue line) mice during infection kinetics (right). Values represent the mean ± SEM (n ≥ 3). Data were analyzed using the Mann-Whitney test at each specific time point and two-way ANOVA for comparisons between WT and Il17ra(-/-) animals. *p<0.05. (B-D) Representative images of CVL cytospin preparations from WT and Il17ra(-/-) mice, Uninfected and Infected at day 2 post infection. ECs were stained by IF to detect mBD1 (panel B) or mBD3 (panel D) expression. Images were captured with a LEICA DMi8 inverted fluorescence microscope (x200 magnification). mBD1 and mBD3 expression is shown in green (top panels). (C-E) The lower panels show the semi-quantification of mBD1 (panel C) and mBD3 (panel E) fluorescence intensity, normalized to the Uninfected group at day 2 post infection and expressed as fold change in WT and Il17ra(-/-) mice, comparing infected and uninfected groups, using ImageJ software (n=4 mice). Values represent the mean ± SEM. Data were analyzed using Student's t-test. ****p<0.0001. WT: wild-type; CFU: Colony Forming Units; CVL: Cervicovaginal lavage; ECs: Epithelial cells. SEM = standard error of the mean; ANOVA = analysis of variance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



