Submitted:
08 February 2026
Posted:
10 February 2026
You are already at the latest version
Abstract
Background: Traditional two-dimensional (2D) neurobiological models fail to capture the complex spatial, mechanical, and biochemical microenvironment of the human brain. This limitation has contributed to a high failure rate in translating neuroprotective drugs from bench to bedside. Main body: This review explores the technological evolution toward three-dimensional (3D) systems, including neural organoids, scaffold-based hydrogels, and microfluidic Organ-on-Chip (OoC) platforms. This review discusses how these systems facilitate realistic synaptic connectivity and neurovascular unit modeling. By mimicking the native extracellular matrix (ECM) and introducing dynamic fluid flow, these models provide a more accurate representation of human CNS pathophysiology. Specific examination is provided regarding the role of mechanotransduction and the blood-brain barrier (BBB) in disease progression. Furthermore, the integration of patient-derived induced pluripotent stem cells (iPSCs) for high-fidelity disease modeling and personalized drug screening are highlighted, which allows for the observation of disease phenotypes—such as amyloid-beta aggregation and phosphorylated tau—that are often absent in 2D or animal models. Conclusion: 3D neurobiological models represent a transformative shift toward precision medicine. They offer more predictive platforms for treating neurodegenerative and neurodevelopmental disorders, ultimately aiming to reduce the economic burden of drug failure and improve patient-specific therapeutic outcomes.
Keywords:
3D cell culture
; organoids
; organ-on-chip
; neurobiology
; precision medicine
; iPSCs
; neurovascular unit
Background
The central nervous system (CNS) is characterized by an intricate hierarchy of cellular and extracellular components. For decades, the gold standard for studying this system in vitro has been two-dimensional (2D) culture. However, 2D systems force neurons into an unnatural planar geometry, leading to altered gene expression, abnormal protein distribution, and a lack of the complex 3D connectivity found in vivo [1,2]. In a 2D environment, cells are exposed to a uniform concentration of nutrients and oxygen, which does not reflect the nutrient gradients or the specialized niches found in the human brain.
With the rising global burden of neurodegenerative diseases such as Alzheimer’s (AD) and Parkinson’s (PD), there is an urgent need for more predictive human-relevant models. The paradigm shift toward 3D cultures—ranging from self-assembled organoids to engineered Organ-on-Chip (OoC) platforms—aims to solve the “translational failure” where 90% of CNS drugs fail in clinical trials despite success in animal and 2D models [3,4]. While animal models provide systemic complexity, they often fail to replicate human-specific cortical expansion and metabolic pathways, particularly in the context of neuroinflammation and glial responses [5,6].
Aim of this review: This article provides an in-depth analysis of the transition from 2D to 3D neurobiological technologies. The objective of this review is to elucidate the architectural benefits of 3D systems, the physiological advantages of microfluidic integration for the neurovascular unit, and the clinical implications of using patient-specific “avatars” for precision medicine.
1. Architectural Superiority of 3D Systems
The human brain is not merely a collection of neurons; it is a highly organized structure where the extracellular matrix (ECM) plays a dynamic role in signaling. In 2D cultures, neurons are typically grown on glass or plastic surfaces coated with a thin layer of adhesive proteins. This setup imposes a forced polarity that does not exist in the brain [7]. As summarized in Table 1 and Table 3D environments offer a critical improvement in mechanical realism, transitioning from the gigapascal rigidity of plastic to the kilopascal softness of native brain tissue.
3D models allow cells to interact with their surroundings in all directions, promoting natural neuronal morphology, including more complex dendrite branching and the formation of functional synapses [8]. Furthermore, the 3D environment enables the study of mechanotransduction—the process by which cells convert mechanical stimuli into biochemical signals. In the brain, the stiffness of the tissue (expressed as Young’s modulus) is approximately 0.5–1.0 kPa. Traditional 2D culture plastics are known to trigger pathological activation in glial cells, potentially confounding experimental results in neuroinflammation studies [9].
1.1. Scaffold-Free: Brain Organoids
Brain organoids, or “mini-brains,” represent a significant breakthrough as self-organized 3D tissues derived from human iPSCs that recapitulate embryonic developmental processes [10]. Unlike 2D monolayers, organoids exhibit distinct regionalization, containing structures reminiscent of the cerebral cortex and hippocampus. As illustrated in Figure 1, the self-assembly process allows for the development of distinct layers that mimic the human cortical architecture, a feature entirely absent in planar cultures.
These models have been instrumental in studying neurodevelopmental disorders. For example, cerebral organoids allowed researchers to observe the premature differentiation of neural progenitors in microcephaly [11]. However, the lack of a vascular system remains a challenge, as the inner core often becomes necrotic due to limited oxygen diffusion once the organoid reaches a size typically greater than 2 mm [12].
1.2. Scaffold-Based: Hydrogels and 3D Bioprinting
To address the structural limitations of organoids, researchers utilize scaffold-based 3D models. Natural hydrogels like collagen, fibrin, and hyaluronic acid mimic the native ECM of the brain [13]. Synthetic hydrogels, such as polyethylene glycol (PEG), offer the advantage of being “tunable,” meaning researchers can precisely control the stiffness and biochemical composition of the environment to study specific disease states [14].
3D bioprinting takes this a step further by using “bio-inks” to deposit cells and ECM components in a layer-by-layer fashion. This allows for the creation of heterogeneous tissues where neurons, astrocytes, and microglia are placed in specific spatial arrangements [15]. This spatial control is vital for modeling the “neurogenic niche,” where the proximity of specific cell types dictates stem cell fate and neuronal maturation.
2. Organ-on-Chip (OoC) and the Neurovascular Unit
The neurovascular unit (NVU) is a complex interface consisting of endothelial cells, pericytes, astrocytes, and neurons. It is responsible for maintaining the blood-brain barrier (BBB) and regulating cerebral blood flow.
2.1. The Blood-Brain Barrier (BBB) on a Chip
The BBB is a highly selective semipermeable border that prevents solutes in the circulating blood from non-selectively crossing into the CNS. Traditional 2D “Transwell” models of the BBB are static and lack the fluid shear stress that is essential for the maturation of endothelial cells.
Organ-on-Chip (OoC) platforms integrate microfluidic channels that simulate blood flow [16]. Studies have demonstrated that endothelial cells exposed to shear stress in these chips show significantly higher expression of tight-junction proteins like Claudin-5 and Occludin, leading to trans-endothelial electrical resistance (TEER) values that approach in vivo levels [17,18]. These models are now being used to test the permeability of new neurotherapeutic drugs, providing a much more accurate prediction of whether a drug will actually reach the brain in a human patient [19].
2.2. Microfluidic Modeling of Axonal Transport
Microfluidics also allows for the physical separation of neuronal cell bodies from their axons. Using microgrooves, researchers can isolate axons in a separate compartment. This is particularly useful for studying neurodegenerative diseases where the primary pathology begins at the synapse or along the axon, such as Amyotrophic Lateral Sclerosis (ALS) or glaucoma [20,21]. By applying toxic proteins (like amyloid-beta oligomers) solely to the axonal compartment, researchers can track the retrograde transport of damage signals to the nucleus, a process that is obscured in 2D cultures where the entire cell is exposed simultaneously [22].
3. Precision Medicine and Patient-Specific Modeling
Precision medicine aims to tailor medical treatment to the individual characteristics of each patient. The integration of 3D cell culture with patient-derived iPSCs has made the concept of “clinical trials in a dish” a reality.
3.1. High-Fidelity Disease Modeling (AD, PD, ALS)
Many neurodegenerative diseases are characterized by protein misfolding. In Alzheimer’s Disease (AD), 2D models often fail to show plaque formation because the secreted amyloid-beta (Aβ) is washed away into the large volume of culture media. In a 3D hydrogel or organoid, restricted diffusion leads to the accumulation and aggregation of these proteins, closely mimicking the pathology seen in patient brains [23,24].
In Parkinson’s Disease (PD), 3D midbrain organoids have revealed that patient-specific genetic mutations result in mitochondrial dysfunctions that are only fully apparent when the cells are grown in a 3D architecture supporting metabolic maturity [25,26]. The wide array of these applications across various neurological conditions is detailed in Table 2, highlighting the specific molecular insights gained from 3D architectures.
3.2. Personalized Drug Screening and Toxicity
One of the most beneficial outcomes of 3D neurobiology is the ability to perform personalized drug screening. For example, in patients with glioblastoma, 3D “tumoroids” can be generated from the patient’s own biopsy. These models can be treated with various combinations of chemotherapeutics to identify the most effective treatment for that specific individual’s tumor [27,28].
In neurobiology, this approach is being expanded to identify potential neurotoxic side effects of drugs. Since every individual has a unique genetic makeup, a drug that is safe for the general population might be toxic to a specific patient. High-throughput 3D platforms allow for the testing of these drugs across diverse genetic backgrounds before they are administered to patients, significantly increasing safety and efficacy [29,30].
Conclusions
The transition from 2D to 3D cell culture in neurobiology represents a paradigm shift from simplistic reductionism to physiological realism.
Beneficial Outcomes and Future Directions: The evidence presented in this review suggests that 3D organoids and Organ-on-Chip technologies provide the necessary structural and physiological depth to overcome the translational gap in drug development. Specifically:
- Reduced Clinical Failure: By providing more predictive data during the preclinical phase, 3D models can filter out ineffective or toxic compounds before they reach expensive human trials.
- Patient-Centric Care: The use of patient-derived iPSCs in 3D systems allows for the development of “patient avatars,” enabling clinicians to test treatments in vitro before treating the patient.
- Ethical Advancement: These models offer a robust alternative to animal testing, particularly for studying human-specific neurological conditions that do not have accurate animal counterparts.
Future efforts must focus on the integration of multiple “organs” on a single chip (e.g., gut-brain axis) and the development of vascularized organoids to maintain long-term viability. As these technologies become more scalable and standardized, they will undoubtedly become the cornerstone of neurobiological research and precision medicine.
Authors’ Contributions
Farnam Gholipour Maralan performed the conceptualization, extensive literature review, data synthesis, drafting of technical sections, and final manuscript preparation.
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
The author declares no competing interests.
Funding
No funding was received for this study.
Acknowledgments
Not applicable.
References
- Jensen, C; Teng, Y. Is it time to start transitioning from 2D to 3D cell culture? Front Mol Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Abbott, A. Biology’s new dimension. Nature 2003, 424(6951), 870–2. [Google Scholar] [CrossRef]
- Cummings, JL; Morstorf, T; Zhong, K. Alzheimer’s disease drug-development pipeline: few candidates, frequent failures. Alzheimers Dement. 2014, 10(4), 432–8. [Google Scholar] [CrossRef]
- Gribkoff, VK; Kaczmarek, LK. The need for new models in antiepileptic drug discovery. Neuropharmacology 2017, 120, 1–2. [Google Scholar]
- Lancaster, MA; Knoblich, JA. Organogenesis in a dish: modeling development and disease using pluripotent stem cells. Science 2014, 345(6194), 1247125. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y; Inoue, H; Wu, JC; Yamanaka, S. Induced pluripotent stem cell technology: a decade of progress. Nat Rev Drug Discov. 2017, 16(2), 115–30. [Google Scholar] [CrossRef] [PubMed]
- Baker, BM; Chen, CS. Deconstructing the third dimension: how 3D culture microenvironments alter cellular cues. J Cell Sci. 2012, 125(13), 3015–24. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, AM; DeLaporte, L; Vunjak-Novakovic, G; et al. Nerve in a dish: materials for 3D in vitro neurons. Soft Matter 2015, 11(37), 7254–67. [Google Scholar]
- Tang-Schomer, MD; White, JD; Tien, LW; et al. Bioengineered functional 3D brain-like tissue. PNAS 2014, 111(38), 13811–6. [Google Scholar] [CrossRef]
- Lancaster, MA; Renner, M; Martin, CA; et al. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501(7467), 373–9. [Google Scholar] [CrossRef]
- Cugola, FR; Fernandes, IR; Russo, FB; et al. The Brazilian Zika virus strain causes birth defects in experimental models. Nature 2016, 534(7606), 267–71. [Google Scholar] [CrossRef] [PubMed]
- Quadrato, G; Nguyen, T; Macosko, EZ; et al. Cell diversity and network dynamics in photosensitive human brain organoids. Nature 2017, 545(7652), 48–53. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R; Stevens, L; Thompson, BC; et al. 3D printing of novel gellan gum-based scaffolds for neural tissue engineering. Biomaterials 2015, 67, 264–73. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q; Tomaskovic-Crook, E; Wallace, GG; et al. 3D bioprinting of human neural progenitors and their activity in 3D-printed structures. Adv Healthc Mater. 2016, 5(12), 1429–38. [Google Scholar] [CrossRef]
- Murphy, SV; Atala, A. 3D bioprinting of tissues and organs. Nat Biotechnol. 2014, 32(8), 773–85. [Google Scholar] [CrossRef]
- Bhatia, SN; Ingber, DE. Microfluidic organs-on-chips. Nat Biotechnol. 2014, 32(8), 760–72. [Google Scholar] [CrossRef]
- Booth, R; Kim, H. Characterization of a microfluidic blood-brain barrier model. Lab Chip 2012, 12(10), 1784–92. [Google Scholar] [CrossRef]
- Herland, A; van der Meer, AD; FitzGerald, EA; et al. Distinct contributions of astrocytes and pericytes to neuroinflammation in a 3D microfluidic human blood-brain barrier model. PLoS One 2016, 11(3), e0150360. [Google Scholar] [CrossRef]
- Vatine, GD; Barrile, R; Kiernan, MJ; et al. Human iPSC-derived blood-brain barrier chips enable disease modeling and personalized medicine applications. Cell Stem Cell. 2019, 24(6), 995–1005. [Google Scholar] [CrossRef]
- Osaki, T; Sivathanu, V; Kamm, RD. Engineered 3D vascular and neural tissue models. Sci Adv. 2018, 4(10), eaat5847. [Google Scholar] [CrossRef]
- Taylor, AM; Blurton-Jones, M; Rhee, SW; et al. A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat Methods 2005, 2(8), 599–605. [Google Scholar] [CrossRef]
- Park, J; Wetzel, I; Marriott, I; et al. A 3D human triculture system modeling neurodegeneration and neuroinflammation in Alzheimer’s disease. Lab Chip 2014, 14(24), 4740–50. [Google Scholar] [CrossRef]
- Choi, SH; Kim, YH; Hebisch, M; et al. A three-dimensional human neural cell culture model of Alzheimer’s disease. Nature 2014, 515(7526), 274–8. [Google Scholar] [CrossRef] [PubMed]
- Lin, YT; Seo, J; Gao, F; et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer’s disease phenotypes in human iPSC-derived brain cell types. Neuron 2018, 98(6), 1144–54. [Google Scholar] [CrossRef]
- Jo, J; Xiao, Y; Sun, AX; et al. Midbrain-like organoids from human pluripotent stem cells contain dopaminergic neurons. Cell Stem Cell. 2016, 19(2), 248–57. [Google Scholar] [CrossRef]
- Di Domenico, A; Carola, G; Calatayud, C; et al. Patient-specific iPSC-derived astrocytes contribute to non-cell autonomous neurodegeneration in Parkinson’s disease. Stem Cell Reports 2019, 12(2), 213–29. [Google Scholar] [CrossRef]
- Hubert, CG; Rivera, M; Spangler, LC; et al. A three-dimensional organoid culture system derived from human glioblastomas. Cancer Res. 2016, 76(8), 2468–77. [Google Scholar] [CrossRef]
- Linkous, A; Balamatsli, D; Snuderl, M; et al. Modeling patient-derived glioblastoma with cerebral organoids. Cell Rep. 2019, 26(12), 3203–11. [Google Scholar] [CrossRef] [PubMed]
- Pașca, AM; Sloan, SA; Clarke, LE; et al. Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat Methods 2015, 12(7), 671–8. [Google Scholar] [CrossRef]
- Truong, DD; Fan, Y; Jureczek, J; et al. Engineering a neurovascular unit on a chip. Biomaterials 2019, 198, 63–77. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparative analysis of neurodevelopmental models. (A) 2D adherent systems provide high-throughput monitoring of neural growth. (B) 3D brain organoids recapitulate complex spatial patterning and cellular diversity of the human brain.
Figure 1.
Comparative analysis of neurodevelopmental models. (A) 2D adherent systems provide high-throughput monitoring of neural growth. (B) 3D brain organoids recapitulate complex spatial patterning and cellular diversity of the human brain.
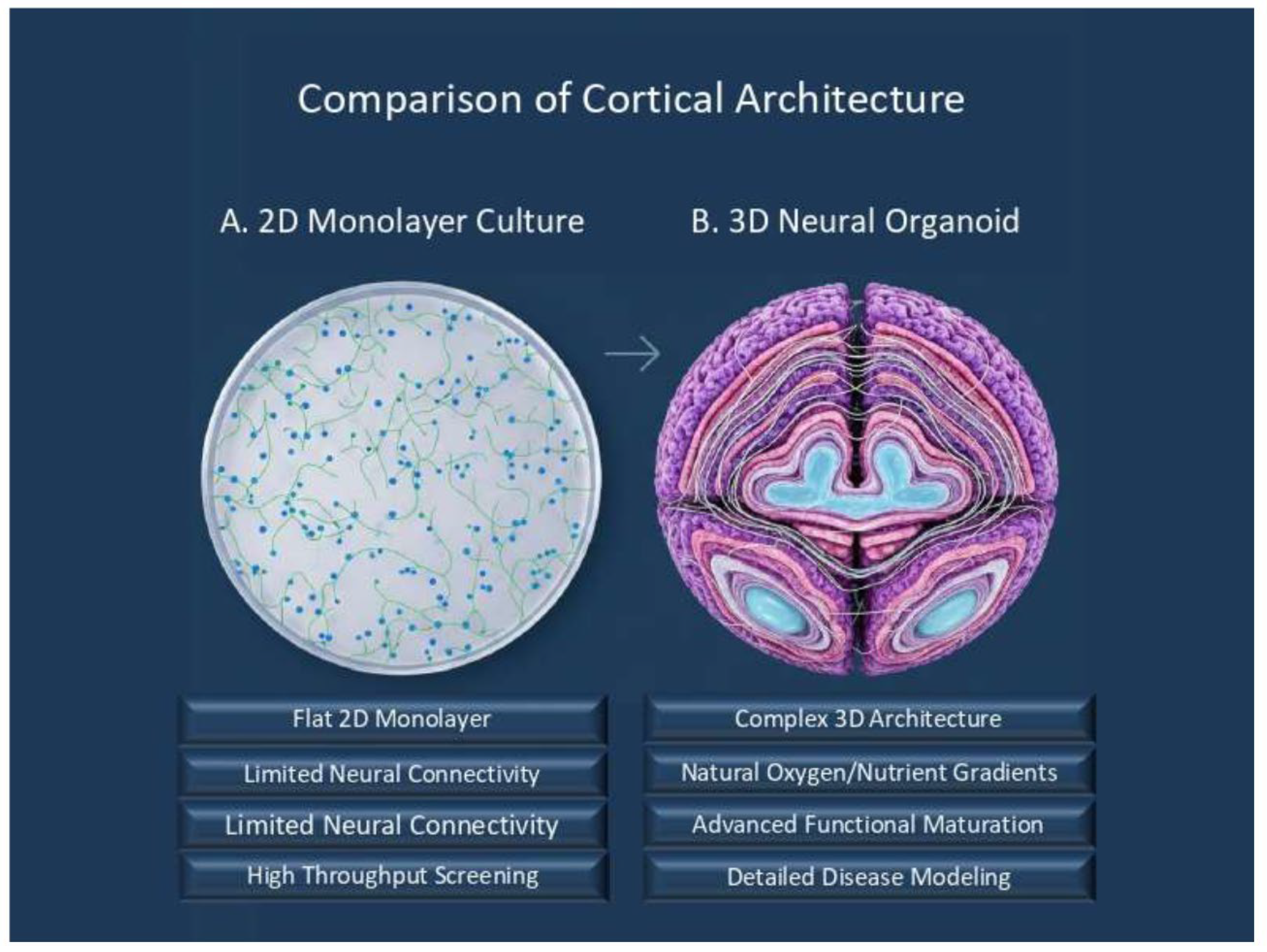

Table 1.
Comparison of 2D vs. 3D Neurobiological Models This table compares the fundamental physical and predictive characteristics of traditional and modern culture systems.
Table 1.
Comparison of 2D vs. 3D Neurobiological Models This table compares the fundamental physical and predictive characteristics of traditional and modern culture systems.
| Feature | 2D Culture | 3D Organoids | Organ-on-Chip (3D) |
|---|---|---|---|
| Mechanical Environment | Non-physiological (Rigid) | Physiological (Soft) | Physiological (Soft + Flow) |
| Cell Morphology | Flattened, unnatural | Natural, 3D branching | Controlled, polarized |
| Nutrient Diffusion | Uniform | Gradient-based | Controlled/Dynamic |
| Predictive Power | Low (30-40%) [3,4] | High (60-80%) [10,23] | Very High (>80%) [19,30] |
| Translational Use | Basic signaling | Disease mechanisms | Drug screening/PK-PD |
Table 2.
Applications of 3D Models in Neurological Diseases A summary of how 3D platforms have successfully modeled specific molecular hallmarks of CNS disorders.
Table 2.
Applications of 3D Models in Neurological Diseases A summary of how 3D platforms have successfully modeled specific molecular hallmarks of CNS disorders.
| Disease | 3D Model Type | Key Molecular Finding | Ref |
|---|---|---|---|
| Alzheimer’s | iPSC 3D Hydrogel | Aβ plaque and Tau tangle formation | [23] |
| Parkinson’s | Midbrain Organoids | Selective dopaminergic loss | [25] |
| ALS | Motor Neuron Chip | Defective neuromuscular junction (NMJ) | [20] |
| Zika Virus | Cerebral Organoids | Viral-induced apoptosis of NPC cells | [11] |
| Glioblastoma | Patient-derived Spheroids | Personalized drug resistance profiling | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



