Submitted:
01 February 2026
Posted:
03 February 2026
You are already at the latest version
Abstract
Histamine is a phylogenetically ancient neuromodulator with widespread projections from the tuberomammillary nucleus of the posterior hypothalamus. Rather than encoding specific perceptual, emotional, or motivational content, histamine regulates the context in which neural signals operate. Through a distinctive receptor architecture—most notably presynaptic H3 auto- and heteroreceptors—histamine interacts with all major neurochemical and neuropeptidergic systems, including dopamine, serotonin, noradrenaline, acetylcholine, glutamate, GABA, and hypothalamic peptides. In this review, we synthesise evidence across molecular, cellular, circuit, and systems neuroscience to argue that histamine functions as an interactional hub that coordinates the expression of other neuromodulatory signals. Histaminergic tone shapes monoaminergic gain, gates cholinergic cortical activation, regulates excitation–inhibition balance, and modulates neuropeptide-driven arousal, stress responsiveness, and social salience in a state-dependent manner. Rather than acting in parallel to these systems, histamine determines when their effects promote flexibility versus stability, vigilance versus recovery, and encoding versus consolidation. This interactional framework provides a unifying account of how arousal, cognition, sleep, inflammation, and behaviour are aligned, and suggests that neuropsychiatric symptoms may arise from disrupted cross-system coordination rather than isolated transmitter dysfunction.
Keywords:
histamine
; tuberomammillary nucleus
; neuromodulatory interactions
1. Introduction
The human brain relies on a small number of neuromodulatory systems to regulate global states such as arousal, attention, learning, and behavioural flexibility [1]. While monoamines such as dopamine, serotonin, and noradrenaline have dominated theoretical and clinical models of brain function, the histaminergic system has remained comparatively underappreciated outside its established role in sleep–wake regulation [2,3,4]. This neglect persists despite the fact that histamine possesses a set of anatomical, receptor-level, and physiological properties that position it uniquely as a global integrator of neurochemical signalling. By “global integrator,” we refer to a neuromodulatory system that regulates the expression, gain, and coordination of other neuromodulatory signals via presynaptic and state-dependent mechanisms, without encoding task-specific or motivational content.
Histaminergic neurons originate almost exclusively from the tuberomammillary nucleus of the posterior hypothalamus, yet project diffusely throughout the cortex, basal ganglia, limbic system, thalamus, and brainstem [3,5]. This compact-but-ubiquitous architecture mirrors that of other state-setting neuromodulators, but histamine differs in a critical respect: it exerts powerful presynaptic control over the release of multiple neurotransmitters via the histamine H3 receptor [4]. Acting as both an autoreceptor and a heteroreceptor, H3 receptors inhibit the release of histamine itself as well as dopamine [6], serotonin [7], noradrenaline [8], acetylcholine [9], glutamate [10], and GABA [11]. Through this mechanism, histamine is able to coordinate the activity of otherwise distinct neurochemical systems in a context-dependent manner [4].
Functionally, histamine does not encode specific perceptual, emotional, or motivational content [3,4]. Instead, it regulates the operating conditions under which neural computations take place. Histaminergic tone tracks wakefulness, metabolic state, circadian rhythms, stress, and immune activation, dynamically shaping cortical excitability, signal-to-noise ratio, and the balance between stability and flexibility in neural networks. These properties suggest that histamine plays a central role in determining how strongly internal signals are weighted, how persistently errors are experienced, and how readily beliefs and action policies are updated in response to new information [12,13].
Recent advances in molecular neuroscience, pharmacology, and systems-level modelling have begun to reveal the breadth of histamine’s interactions with other neuromodulatory systems, as well as its involvement in excitation–inhibition balance, neuropeptide signalling, and neuroimmune communication. At the same time, growing evidence implicates histaminergic dysfunction in neuropsychiatric conditions characterised by altered arousal, sleep disturbance, cognitive rigidity, fatigue, and impaired learning under uncertainty. These observations challenge the traditional view of histamine as a peripheral or secondary neuromodulator and instead motivate a reconceptualisation of histamine as a central coordinator of brain state [4,14].
In this article, we synthesise evidence across molecular, cellular, circuit, and translational levels to provide an integrated account of how histamine interacts with all major neurochemical systems in the brain. We argue that histamine functions as a systems-level regulator that aligns neuromodulatory, excitatory–inhibitory, neuropeptidergic to support adaptive cognition and behaviour. By framing histamine as a global integrator rather than a content-specific transmitter, we aim to clarify its role in health and disease and to highlight new opportunities for mechanism-based therapeutic interventions.
2. Anatomical and Receptor Architecture of the Histaminergic System
The histaminergic system is defined by a striking anatomical asymmetry: a small, spatially restricted population of neurons exerts influence over nearly the entire brain [5,12]. Histamine-producing neurons are located almost exclusively within the tuberomammillary nucleus (TMN) of the posterior hypothalamus. Despite their limited number, these neurons give rise to highly divergent axonal projections that innervate the cerebral cortex, hippocampus, amygdala, basal ganglia, thalamus, hypothalamus, and brainstem. This organisation closely resembles that of other neuromodulatory systems involved in global state regulation, yet the histamine system displays an especially diffuse and non-topographic projection pattern [3,12].
Histaminergic neurons fire tonically during wakefulness, reduce activity during non-rapid eye movement sleep, and are largely silent during rapid eye movement sleep. Their firing rate is modulated by circadian inputs, metabolic state, stress, and immune signals, positioning histamine as a key mediator between bodily state and brain-wide neural dynamics. Unlike fast synaptic transmitters, histamine is released in a manner consistent with volume transmission, enabling it to influence large populations of neurons simultaneously rather than conveying temporally precise signals [4,14].
At the molecular level, histamine acts through four G protein–coupled receptors (H1–H4), of which H1, H2, and H3 are most relevant in the central nervous system [15]. H1 and H2 receptors are predominantly postsynaptic and generally excitatory, increasing neuronal excitability through phospholipase C– and adenylyl cyclase–dependent pathways, respectively [4,16]. These receptors are widely expressed across cortical and subcortical regions and contribute to the arousing and activating effects of histamine [14].
In contrast, the H3 receptor has a unique and functionally critical role. H3 receptors are expressed primarily presynaptically, where they function as both autoreceptors on histaminergic terminals and heteroreceptors on non-histaminergic neurons [17]. Activation of H3 autoreceptors inhibits histamine synthesis and release, providing a powerful negative feedback mechanism [18]. As heteroreceptors, H3 receptors suppress the release of multiple neurotransmitters, including dopamine, serotonin, noradrenaline, acetylcholine, glutamate, and GABA. Through this single receptor class, histamine is able to exert coordinated control over diverse neurochemical systems [19].
This receptor architecture has two important implications. First, histamine can rapidly reshape the neurochemical environment without requiring direct postsynaptic action at every target site [18,20]. Second, the effects of histamine are inherently context-dependent: the impact of histaminergic signalling depends on which transmitter systems are active, which receptors are expressed locally, and the current state of the network [12]. As a result, histamine does not impose a fixed effect on brain function, but instead modulates how other systems express their influence [21].
Taken together, the anatomical compactness of the TMN, the diffuse projection pattern of histaminergic fibres, and the presynaptic dominance of the H3 receptor define histamine as a metamodulatory system [22]. Rather than encoding information directly, histamine sets the conditions under which other neurotransmitters operate, providing a structural foundation for its role as a global integrator of brain state.
Throughout this review, arrows and directional language refer to net functional influence at the systems level, rather than uniform changes in transmitter release or firing rates at all terminals.
3. Histamine–Monoamine Interactions: Coordinating Dopamine, Serotonin, and Noradrenaline
Monoaminergic systems such as dopamine, serotonin, and noradrenaline are central to contemporary models of cognition and psychopathology. These systems are typically ascribed distinct computational roles, including reward prediction, behavioural inhibition, and alerting. However, growing evidence indicates that their influence on behaviour depends critically on the global state of the brain [2]. Histamine plays a key role in establishing this state by modulating monoaminergic transmission at a presynaptic level.
3.1. Dopamine
Histamine exerts strong inhibitory control over dopaminergic signalling via H3 heteroreceptors located on dopaminergic terminals in both cortical and striatal regions [6]. Activation of these receptors reduces dopamine synthesis and release, whereas H3 antagonists or inverse agonists increase extracellular dopamine, particularly in the prefrontal cortex. Importantly, these effects do not resemble the direct dopaminergic stimulation produced by psychostimulants. Instead, histamine regulates the gain of dopaminergic signalling, shaping signal-to-noise ratio rather than absolute dopamine levels [6,23,24,25,26].
Functionally, this interaction influences action selection, habit formation, and the balance between goal-directed and habitual control. By constraining dopaminergic output under conditions of sustained arousal, histamine may prevent excessive reinforcement of transient or noisy signals. Conversely, reduced histaminergic tone may permit exaggerated dopaminergic influence, biasing behaviour toward rigid or overlearned action policies [23,24,25,26].
3.2. Serotonin
Serotonergic neurons are similarly subject to presynaptic regulation by histamine. H3 receptors expressed on serotonergic terminals inhibit serotonin release across widespread forebrain regions [27,28]. Through this mechanism, histamine modulates serotonergic contributions to behavioural inhibition, delay discounting and affective regulation [29,30,31].
Rather than acting as a primary determinant of mood or emotional valence, histamine shapes the context in which serotonergic signals operate. Under high histaminergic tone, serotonergic signalling may preferentially support flexible adjustment to changing contingencies. Under low histaminergic tone, the same serotonergic signals may promote perseveration or excessive caution. This interaction provides a potential bridge between arousal regulation and serotonergic models of compulsivity and anxiety [12,20].
3.3. Noradrenaline
The interaction between histamine and noradrenaline is central to the regulation of arousal and vigilance. Noradrenergic neurons of the locus coeruleus encode phasic responses to salient or unexpected events, while histamine provides a tonic arousal backdrop that increases the gain and functional impact of noradrenergic signalling at the cortical level [32]. Anatomical and functional studies demonstrate reciprocal connectivity between histaminergic and noradrenergic nuclei, supporting close coordination between these systems [8,33].
From a functional perspective, histamine stabilises wakefulness and attentional readiness, allowing noradrenergic signals to selectively highlight deviations from expectation rather than continuously driving alertness [3]. Disruption of this balance may lead to states of hypervigilance, in which phasic noradrenergic signalling is exaggerated against an unstable arousal background.
3.4. A Coordinated Monoaminergic Framework
Taken together, histamine does not compete with monoamines for control over behaviour. Instead, it regulates the conditions under which monoaminergic signals influence neural processing. By acting presynaptically through H3 heteroreceptors, histamine synchronises dopaminergic, serotonergic, and noradrenergic activity, ensuring that their effects remain proportionate to environmental demands and internal state.
This coordination suggests a hierarchical organisation in which histamine operates upstream of monoamines, shaping learning rates, uncertainty sensitivity, and behavioural flexibility across large-scale networks. Such a framework provides a natural explanation for why histaminergic perturbations can produce broad cognitive and behavioural effects without directly mimicking monoaminergic drugs.
4. Histamine and Acetylcholine: Cortical Activation, Attention, and Learning Readiness
Among histamine’s many interactions, its coupling with the cholinergic system is particularly important for understanding cortical activation and attention [9]. Acetylcholine is a primary driver of cortical desynchronisation, sensory gain, and experience-dependent plasticity, yet its effects are highly state-dependent [34,35,36]. Histamine plays a crucial role in establishing the arousal conditions under which cholinergic signalling can effectively support cognition.
Histaminergic neurons strongly excite basal forebrain cholinergic cells, predominantly via postsynaptic H1 receptor activation. This excitation promotes acetylcholine release throughout the cortex, facilitating wakefulness and attentional readiness. Experimental disruption of histaminergic signalling markedly attenuates cholinergic-driven cortical activation, highlighting the dependency of the cholinergic system on intact histaminergic tone [37,38,39].
Importantly, histamine does not directly encode attentional focus or task relevance. Instead, it sustains a cortical regime characterised by low synchrony and high responsiveness, within which acetylcholine can selectively enhance the processing of relevant sensory inputs. In this sense, histamine provides the permissive background state for cholinergic modulation rather than acting as an attentional signal itself [4,14].
Presynaptic H3 receptors further refine this interaction by modulating acetylcholine release at cortical terminals. Through H3-mediated inhibition, histamine can constrain excessive cholinergic activity, preventing overactivation and preserving signal fidelity. This dual mechanism—postsynaptic excitation of cholinergic neurons combined with presynaptic control of acetylcholine release—allows histamine to finely tune cortical activation across different behavioural contexts [40].
These interactions have clear implications for learning and memory. Acetylcholine is known to promote encoding over retrieval and to enhance synaptic plasticity in sensory and associative cortices. By regulating cholinergic tone, histamine influences when the brain is biased toward acquiring new information versus stabilising existing representations. Disruption of this balance may contribute to attentional instability, inefficient learning, or cognitive fatigue [40].
Clinically, the histamine–acetylcholine interaction provides a mechanistic explanation for the cognitive side effects of antihistaminergic medications, which often impair attention and vigilance [41], as well as for the procognitive effects observed with H3 receptor antagonists [42]. Rather than acting as classical stimulants, these agents appear to restore an optimal arousal–cholinergic coupling that supports sustained attention and learning efficiency.
5. Regulation of Excitation–Inhibition Balance: Histamine, Glutamate, and GABA
A defining feature of adaptive brain function is the capacity to balance excitation and inhibition across neural circuits. This balance determines not only overall network stability but also the flexibility with which neural representations can be updated in response to new information [43,44,45]. Histamine plays a central role in regulating this balance by acting simultaneously on excitatory and inhibitory neurotransmission through receptor-specific and location-dependent mechanisms [4]. The net effect of histamine on excitation–inhibition balance depends on circuit architecture, baseline activity, and receptor distribution.
Histamine modulates glutamatergic transmission at multiple levels [46,47]. Presynaptic H3 receptors expressed on glutamatergic terminals inhibit glutamate release, particularly within cortical and cortico-striatal pathways. This mechanism constrains excessive excitatory drive and limits the propagation of noisy or transient signals through large-scale networks. In parallel, postsynaptic H1 and H2 receptors increase the excitability of pyramidal neurons by enhancing intracellular calcium signalling and membrane depolarisation [4]. The coexistence of these opposing actions allows histamine to increase neuronal responsiveness while preventing runaway excitation [48].
GABAergic transmission is similarly regulated in a context-dependent manner [46]. H3 receptors are expressed on subsets of GABAergic terminals, where their activation suppresses inhibitory neurotransmitter release. The net effect of histamine on inhibition therefore depends on circuit architecture and baseline activity. In some microcircuits, histamine may transiently disinhibit pyramidal neurons, facilitating information flow; in others, it may stabilise inhibitory control by limiting excessive GABA release. This flexibility enables histamine to reshape local excitation–inhibition ratios without imposing a uniform effect across regions [48].
Rather than shifting excitation or inhibition globally, histamine functions as a gain controller [48]. By adjusting the depth and stability of attractor states within cortical and cortico-striatal networks, histamine influences whether neural activity patterns are resistant to perturbation or readily updated. High histaminergic tone favours responsiveness and adaptive updating, whereas reduced histaminergic signalling biases networks toward rigidity and perseveration. This framework aligns with computational models in which neuromodulators regulate learning rates and volatility sensitivity rather than encoding specific outcomes.
The implications of histamine’s role in excitation–inhibition balance extend beyond cognition to sleep and fatigue. During sustained wakefulness or inflammatory challenge, alterations in histaminergic signalling may destabilise local excitation–inhibition dynamics, contributing to cognitive variability, reduced processing efficiency, and the emergence of local sleep-like activity during wake. These effects provide a mechanistic bridge between arousal regulation, synaptic physiology, and behavioural performance [39,49].
In summary, histamine exerts a nuanced and dynamic influence on excitation–inhibition balance by coordinating glutamatergic and GABAergic transmission. Through this mechanism, it shapes the stability–flexibility trade-off that underlies learning, decision-making, and adaptive behaviour. This role positions histamine as a key regulator of network dynamics rather than a simple excitatory or inhibitory modulator.
6. Histamine, Neuropeptides, and the Regulation of Arousal, Motivation, and Social Salience
Histamine is deeply embedded within hypothalamic neuropeptide networks that regulate arousal, motivation, stress responsiveness, and social behaviour. Rather than acting in parallel to these systems, histamine operates as a state-setting intermediary, shaping the conditions under which neuropeptide signals exert their behavioural and cognitive effects. This positioning allows histamine to integrate metabolic, circadian, emotional, and immune information into coherent brain-wide operating modes [12].
6.1. Orexin/Hypocretin: Sustained Wakefulness and Motivational Drive
The strongest and best-characterised interaction between histamine and a neuropeptide system is with orexin (hypocretin). Orexin-producing neurons in the lateral hypothalamus provide dense excitatory input to histaminergic neurons in the tuberomammillary nucleus, robustly increasing histamine release during wakefulness. Conversely, many of the arousal-promoting effects of orexin depend on intact histaminergic signalling, indicating a partially hierarchical relationship between these systems [50].
Functionally, orexin initiates and stabilises wakefulness, while histamine maintains a globally activated cortical state that resists sleep pressure. This division of labour allows sustained alertness without continuous high-frequency firing of orexin neurons. Disruption of histamine–orexin coupling can therefore lead to fragmented wakefulness, fatigue, and impaired cognitive endurance, even in the absence of overt sleep deprivation [51,52].
6.2. Stress-Related Neuropeptides: CRH and Arousal Bias
Histamine also interacts closely with stress-related neuropeptides, particularly corticotropin-releasing hormone (CRH). CRH activates histaminergic neurons both directly and indirectly, while histamine amplifies the arousing and vigilance-enhancing effects of CRH at cortical and limbic levels [53]. This interaction biases the brain toward heightened environmental monitoring and threat sensitivity during stress.
Under acute conditions, this coupling is adaptive, promoting rapid detection of salient stimuli [54,55]. However, chronic activation—such as during prolonged stress or inflammation - may lock neural systems into a persistent high-arousal mode, reducing flexibility and impairing recovery [56]. In this context, histamine may contribute to the transition from adaptive vigilance to maladaptive hyperarousal.
6.3. Oxytocin and Social Neuropeptides: Contextual Modulation Rather than Valence
Although histamine does not directly encode social affiliation or bonding, it plays an important role in shaping the arousal context in which social neuropeptides such as oxytocin operate [57]. Histamine stimulates the release of oxytocin [57]. Oxytocin’s effects on social perception, trust, and emotional resonance are highly state-dependent [58]. Elevated histaminergic tone may bias oxytocinergic signalling toward heightened salience and alertness, whereas lower histamine levels may permit affiliative or calming effects to dominate [59].
This interaction provides a potential explanation for the heterogeneity of oxytocin effects observed across individuals and experimental paradigms. Rather than reflecting inconsistent oxytocin biology, variability may arise from differences in underlying histaminergic and arousal states that gate oxytocin’s downstream impact [60]. We propose this interaction as a hypothesis rather than a settled mechanism, requiring direct experimental testing.
6.4. Integrating Neuropeptide Signals into Global Brain State
Across neuropeptide systems, a common pattern emerges: histamine does not compete with peptides for behavioural control but determines how strongly their signals are expressed at the circuit level. By regulating cortical excitability, signal-to-noise ratio, and responsiveness to internal and external cues, histamine aligns neuropeptide-driven motivational and affective signals with the current physiological and environmental context.
Through this integrative role, histamine enables coherent transitions between behavioural states such as sleep and wakefulness, exploration and conservation, vigilance and social engagement. Disruption of this coordination may therefore contribute to conditions characterised by fatigue, emotional dysregulation, altered social processing, and impaired stress resilience.
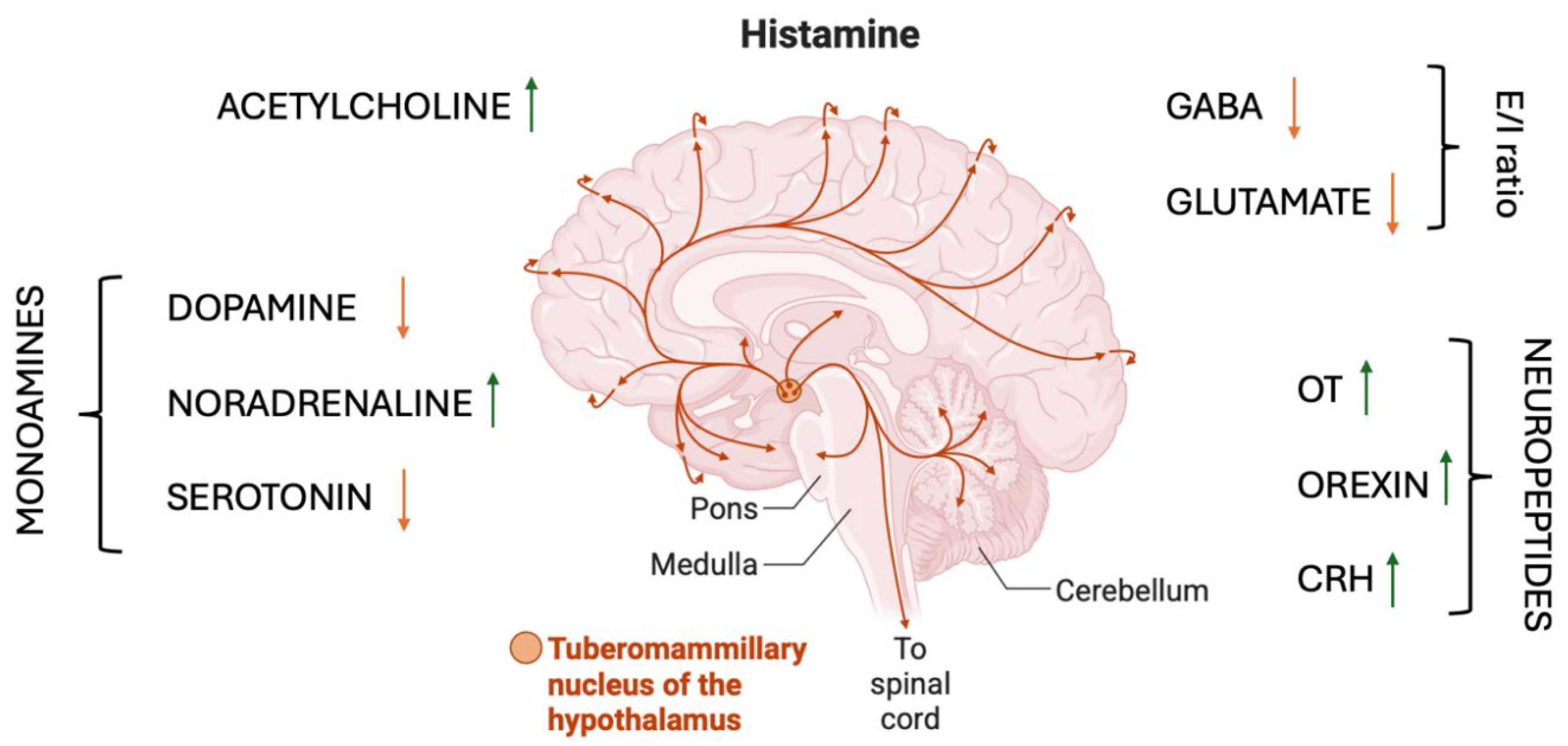
Figure 1.
Histamine as a global integrator of neurochemical systems. Histaminergic neurons originating in the tuberomammillary nucleus of the posterior hypothalamus project diffusely throughout the brain, positioning histamine as a system-level regulator of neural state rather than a content-specific transmitter. Through presynaptic H3 auto- and heteroreceptors, histamine exerts coordinated control over multiple neurotransmitter systems, including monoamines (dopamine, noradrenaline, serotonin), acetylcholine, glutamate, and GABA, while also interacting with key hypothalamic neuropeptides such as orexin, corticotropin-releasing hormone (CRH), and oxytocin (OT). Rather than uniformly exciting or inhibiting neural activity, histamine modulates cortical gain, excitation–inhibition balance, and the relative influence of other neuromodulators in a context-dependent manner. This interactional architecture allows histamine to align arousal, learning flexibility, vigilance, and recovery across behavioural and physiological states. Directional arrows indicate net systems-level influence rather than direct synaptic excitation or inhibition.
Figure 1.
Histamine as a global integrator of neurochemical systems. Histaminergic neurons originating in the tuberomammillary nucleus of the posterior hypothalamus project diffusely throughout the brain, positioning histamine as a system-level regulator of neural state rather than a content-specific transmitter. Through presynaptic H3 auto- and heteroreceptors, histamine exerts coordinated control over multiple neurotransmitter systems, including monoamines (dopamine, noradrenaline, serotonin), acetylcholine, glutamate, and GABA, while also interacting with key hypothalamic neuropeptides such as orexin, corticotropin-releasing hormone (CRH), and oxytocin (OT). Rather than uniformly exciting or inhibiting neural activity, histamine modulates cortical gain, excitation–inhibition balance, and the relative influence of other neuromodulators in a context-dependent manner. This interactional architecture allows histamine to align arousal, learning flexibility, vigilance, and recovery across behavioural and physiological states. Directional arrows indicate net systems-level influence rather than direct synaptic excitation or inhibition.
7. Five Avenues for Future Research
Future research on histamine should prioritise its role as an interactional hub rather than as an isolated neuromodulatory system. A central goal will be to identify the principles governing how histaminergic signalling coordinates monoaminergic systems such as dopamine, serotonin, and noradrenaline across different behavioural contexts. Rather than treating these interactions as static or additive, future studies should ask whether histamine synchronises or hierarchically constrains monoaminergic influences depending on arousal state, uncertainty, or cognitive demand, thereby shaping learning rates, flexibility, and behavioural persistence.
Equally important is a deeper characterisation of histamine–acetylcholine coupling as a mechanism for switching between cognitive modes. While acetylcholine supports attention and plasticity, its effects are highly contingent on global arousal. Future work should therefore examine how histaminergic tone determines whether cholinergic signalling enhances selective processing or instead contributes to distractibility and fatigue, particularly across cortical hierarchies and during sustained cognitive effort.
At the circuit level, a major challenge is to understand how histamine-mediated modulation of glutamatergic and GABAergic transmission interacts with slower neuromodulatory systems to stabilise excitation–inhibition balance. Rather than viewing histamine’s effects on excitation and inhibition in isolation, future studies should investigate whether histamine acts as a higher-order coordinator that aligns synaptic balance with broader neuromodulatory states, enabling networks to remain both responsive and stable across changing demands.
The interaction between histamine and hypothalamic neuropeptides also represents a key frontier. The proposal that histamine gates the expression of neuropeptide signals—such as orexin-driven arousal, CRH-mediated vigilance, or oxytocin-dependent social modulation—requires direct experimental testing. Variability in neuropeptide effects across individuals and paradigms may reflect differences in underlying histaminergic tone rather than intrinsic inconsistencies in peptide function, reframing long-standing debates in social and stress neuroscience.
Finally, future research should shift from transmitter-centric models of dysfunction toward frameworks that emphasise failures of cross-system coordination. Symptoms such as cognitive rigidity, hypervigilance, fatigue, or impaired recovery may arise not from abnormalities within a single system, but from misalignment between histamine and the systems it regulates. Understanding how histamine normally orchestrates interactions among monoamines, acetylcholine, fast synaptic transmission, and neuropeptides will be essential for developing integrative, mechanism-based models of brain state and vulnerability to neuropsychiatric disorders.
Conclusion
Histamine is more than a wake-promoting transmitter or peripheral immune mediator. Although other neuromodulatory systems also contribute to global state regulation, histamine is unique in its combination of compact anatomical origin, diffuse projections, and presynaptic heteroreceptor architecture. The evidence reviewed here supports a reconceptualisation of histamine as a systems-level integrator that coordinates multiple neurochemical and neuropeptidergic processes to regulate global brain state. Through its compact anatomical origin, diffuse projections, and distinctive receptor architecture - particularly presynaptic H3 auto- and heteroreceptors - histamine modulates the availability and impact of dopamine, serotonin, noradrenaline, acetylcholine, glutamate, and GABA in a context-dependent manner. While much mechanistic evidence derives from animal models, the conserved anatomy and receptor architecture of the histaminergic system support cautious translation to human brain function.
Rather than encoding specific behavioural content, histamine shapes the conditions under which neural computations unfold, influencing gain control, signal-to-noise ratio, and the balance between stability and flexibility in large-scale networks. Histamine is neither necessary nor sufficient for all state transitions, nor does it replace monoaminergic or neuropeptidergic signalling; rather, it constrains how these systems express their influence once engaged. This perspective provides a unifying framework for understanding how arousal, sleep, inflammation, and learning interact, and why their dysregulation produces cognitive rigidity, fatigue, and impaired adaptability across psychiatric and neurological disorders. This framework yields several testable predictions. Manipulations of histaminergic tone should preferentially alter learning rates, uncertainty sensitivity, and behavioural flexibility without proportionate changes in reward sensitivity or affective valence. Similarly, H3 receptor modulation should restore context-dependent cognitive flexibility and vigilance more robustly than baseline performance.
Recognising histamine as a global coordinator of brain state opens new avenues for mechanism-based interventions aimed at restoring adaptive neural dynamics rather than targeting individual neurotransmitter systems in isolation.
Acknowledgments
The author thanks colleagues and collaborators for numerous discussions that helped shape the ideas presented in this review, particularly those working at the interface of neuromodulation, systems neuroscience, and psychiatry. The author is grateful to members of the Department of Clinical Neurosciences and Mental Health at the Faculty of Medicine, University of Porto, and the Department of Neuroimaging at the Institute of Psychiatry, Psychology and Neuroscience, King’s College London, for providing an intellectually stimulating environment in which this work was developed. The author also acknowledges the broader neuroscience community whose experimental, theoretical, and clinical contributions underpin the synthesis presented here. No specific funding was received for the preparation of this review.
Declaration of Competing Interests: The author declares no competing interests.
References
- Marder, E. Neuromodulation of neuronal circuits: back to the future. Neuron 2012, 76, 1–11. [Google Scholar] [CrossRef]
- Robbins, T. W. Chemistry of the mind: neurochemical modulation of prefrontal cortical function. J Comp Neurol 2005, 493, 140–146. [Google Scholar] [CrossRef]
- Haas, H.; Panula, P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci 2003, 4, 121–130. [Google Scholar] [CrossRef]
- Brown, R. E.; Stevens, D. R.; Haas, H. L. The physiology of brain histamine. Prog Neurobiol 2001, 63, 637–672. [Google Scholar] [CrossRef]
- Wada, H.; Inagaki, N.; Itowi, N.; Yamatodani, A. Histaminergic neuron system in the brain: distribution and possible functions. Brain Res Bull 1991, 27, 367–370. [Google Scholar] [CrossRef]
- Ellenbroek, B. A. Histamine H(3) receptors, the complex interaction with dopamine and its implications for addiction. Br J Pharmacol 2013, 170, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Hersey, M.; et al. A tale of two transmitters: serotonin and histamine as in vivo biomarkers of chronic stress in mice. J Neuroinflammation 2022, 19, 167. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, N. J.; Stock, M. J.; Wyllie, M. G. Effects of histamine antagonists on noradrenaline-stimulated blood flow and oxygen consumption of brown adipose tissue in the rat. Pflugers Arch 1984, 402, 325–329. [Google Scholar] [CrossRef]
- Prast, H.; et al. Histaminergic neurons modulate acetylcholine release in the ventral striatum: role of H3 histamine receptors. Naunyn Schmiedebergs Arch Pharmacol 1999, 360, 558–564. [Google Scholar] [CrossRef]
- Karpati, A.; et al. Histamine elicits glutamate release from cultured astrocytes. J Pharmacol Sci 2018, 137, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; et al. Wakefulness Is Governed by GABA and Histamine Cotransmission. Neuron 2015, 87, 164–178. [Google Scholar] [CrossRef] [PubMed]
- Panula, P.; Nuutinen, S. The histaminergic network in the brain: basic organization and role in disease. Nat Rev Neurosci 2013, 14, 472–487. [Google Scholar] [CrossRef]
- Li, B.; et al. Histamine Increases Neuronal Excitability and Sensitivity of the Lateral Vestibular Nucleus and Promotes Motor Behaviors via HCN Channel Coupled to H2 Receptor. Front Cell Neurosci 2016, 10, 300. [Google Scholar] [CrossRef]
- Green, M. Histamine in the central nervous system. Proc West Pharmacol Soc 1978, 21, 337–339. [Google Scholar] [PubMed]
- Thangam, E. B.; et al. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front Immunol 2018, 9, 1873. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, H.; et al. Signaling Pathway of Histamine H(1) Receptor-Mediated Histamine H(1) Receptor Gene Upregulation Induced by Histamine in U-373 MG Cells. Curr Issues Mol Biol 2021, 43, 1243–1254. [Google Scholar] [CrossRef]
- Arrang, J. M.; Devaux, B.; Chodkiewicz, J. P.; Schwartz, J. C. H3-receptors control histamine release in human brain. J Neurochem 1988, 51, 105–108. [Google Scholar] [CrossRef]
- Arrang, J. M.; Garbarg, M.; Schwartz, J. C. Auto-inhibition of brain histamine release mediated by a novel class (H3) of histamine receptor. Nature 1983, 302, 832–837. [Google Scholar] [CrossRef]
- Lin, J. S.; Sergeeva, O. A.; Haas, H. L. Histamine H3 receptors and sleep-wake regulation. J Pharmacol Exp Ther 2011, 336, 17–23. [Google Scholar] [CrossRef]
- Schlicker, E.; Kathmann, M. Role of the Histamine H(3) Receptor in the Central Nervous System. Handb Exp Pharmacol 2017, 241, 277–299. [Google Scholar] [CrossRef]
- Bargmann, C. I. Beyond the connectome: how neuromodulators shape neural circuits. Bioessays 2012, 34, 458–465. [Google Scholar] [CrossRef]
- Llinas, R. R. Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective. Front Cell Neurosci 2014, 8, 320. [Google Scholar] [CrossRef]
- Alfaro-Rodriguez, A.; et al. Histamine H3 receptor activation prevents dopamine D1 receptor-mediated inhibition of dopamine release in the rat striatum: a microdialysis study. Neurosci Lett 2013, 552, 5–9. [Google Scholar] [CrossRef]
- Bolam, J. P.; Ellender, T. J. Histamine and the striatum. Neuropharmacology 2016, 106, 74–84. [Google Scholar] [CrossRef]
- Kononoff Vanhanen, J.; Nuutinen, S.; Tuominen, M.; Panula, P. Histamine H3 Receptor Regulates Sensorimotor Gating and Dopaminergic Signaling in the Striatum. J Pharmacol Exp Ther 2016, 357, 264–272. [Google Scholar] [CrossRef]
- Rapanelli, M.; et al. Histamine H3R receptor activation in the dorsal striatum triggers stereotypies in a mouse model of tic disorders. Transl Psychiatry 2017, 7, e1013. [Google Scholar] [CrossRef] [PubMed]
- Schlicker, E.; Betz, R.; Gothert, M. Histamine H3 receptor-mediated inhibition of serotonin release in the rat brain cortex. Naunyn Schmiedebergs Arch Pharmacol 1988, 337, 588–590. [Google Scholar] [CrossRef]
- Threlfell, S.; et al. Histamine H3 receptors inhibit serotonin release in substantia nigra pars reticulata. J Neurosci 2004, 24, 8704–8710. [Google Scholar] [CrossRef] [PubMed]
- Alguacil, L. F.; Perez-Garcia, C. Histamine H3 receptor: a potential drug target for the treatment of central nervous system disorders. Curr Drug Targets CNS Neurol Disord 2003, 2, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Dayan, P.; Huys, Q. J. Serotonin, inhibition, and negative mood. PLoS Comput Biol 2008, 4, e4. [Google Scholar] [CrossRef]
- Cools, R.; Roberts, A. C.; Robbins, T. W. Serotoninergic regulation of emotional and behavioural control processes. Trends Cogn Sci 2008, 12, 31–40. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Cohen, J. D. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci 2005, 28, 403–450. [Google Scholar] [CrossRef] [PubMed]
- Young, C. S.; Mason, R.; Hill, S. J. Studies on the mechanism of histamine-induced release of noradrenaline and 5-hydroxytryptamine from slices of rat cerebral cortex. Biochem Pharmacol 1988, 37, 2799–2805. [Google Scholar] [CrossRef] [PubMed]
- Hasselmo, M. E. The role of acetylcholine in learning and memory. Curr Opin Neurobiol 2006, 16, 710–715. [Google Scholar] [CrossRef]
- Goard, M.; Dan, Y. Basal forebrain activation enhances cortical coding of natural scenes. Nat Neurosci 2009, 12, 1444–1449. [Google Scholar] [CrossRef]
- Sarter, M.; Hasselmo, M. E.; Bruno, J. P.; Givens, B. Unraveling the attentional functions of cortical cholinergic inputs: interactions between signal-driven and cognitive modulation of signal detection. Brain Res Brain Res Rev 2005, 48, 98–111. [Google Scholar] [CrossRef]
- Cecchi, M.; Passani, M. B.; Bacciottini, L.; Mannaioni, P. F.; Blandina, P. Cortical acetylcholine release elicited by stimulation of histamine H1 receptors in the nucleus basalis magnocellularis: a dual-probe microdialysis study in the freely moving rat. Eur J Neurosci 2001, 13, 68–78. [Google Scholar] [CrossRef]
- Khateb, A.; Fort, P.; Pegna, A.; Jones, B. E.; Muhlethaler, M. Cholinergic nucleus basalis neurons are excited by histamine in vitro. Neuroscience 1995, 69, 495–506. [Google Scholar] [CrossRef]
- Thakkar, M. M. Histamine in the regulation of wakefulness. Sleep Med Rev 2011, 15, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Blandina, P.; et al. Inhibition of cortical acetylcholine release and cognitive performance by histamine H3 receptor activation in rats. Br J Pharmacol 1996, 119, 1656–1664. [Google Scholar] [CrossRef]
- Kay, G. G. The effects of antihistamines on cognition and performance. J Allergy Clin Immunol 2000, 105, S622–627. [Google Scholar] [CrossRef]
- Esbenshade, T. A.; et al. The histamine H3 receptor: an attractive target for the treatment of cognitive disorders. Br J Pharmacol 2008, 154, 1166–1181. [Google Scholar] [CrossRef]
- Yizhar, O.; et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature 2011, 477, 171–178. [Google Scholar] [CrossRef]
- Carcea, I.; Froemke, R. C. Cortical plasticity, excitatory-inhibitory balance, and sensory perception. Prog Brain Res 2013, 207, 65–90. [Google Scholar] [CrossRef]
- Froemke, R. C. Plasticity of cortical excitatory-inhibitory balance. Annu Rev Neurosci 2015, 38, 195–219. [Google Scholar] [CrossRef] [PubMed]
- Takei, H.; Yamamoto, K.; Bae, Y. C.; Shirakawa, T.; Kobayashi, M. Histamine H(3) Heteroreceptors Suppress Glutamatergic and GABAergic Synaptic Transmission in the Rat Insular Cortex. Front Neural Circuits 2017, 11, 85. [Google Scholar] [CrossRef]
- Chavez, H.; Vega, R.; Soto, E. Histamine (H3) receptors modulate the excitatory amino acid receptor response of the vestibular afferents. Brain Res 2005, 1064, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schlicker, E.; Malinowska, B.; Kathmann, M.; Gothert, M. Modulation of neurotransmitter release via histamine H3 heteroreceptors. Fundam Clin Pharmacol 1994, 8, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V. V.; Harris, K. D. Sleep and the single neuron: the role of global slow oscillations in individual cell rest. Nat Rev Neurosci 2013, 14, 443–451. [Google Scholar] [CrossRef]
- Huang, Z. L.; et al. Arousal effect of orexin A depends on activation of the histaminergic system. Proc Natl Acad Sci U S A 2001, 98, 9965–9970. [Google Scholar] [CrossRef]
- Burgess, C. R. Histamine and orexin in the control of arousal, locomotion, and motivation. J Neurosci 2010, 30, 2810–2811. [Google Scholar] [CrossRef]
- Saper, C. B.; Fuller, P. M. Wake-sleep circuitry: an overview. Curr Opin Neurobiol 2017, 44, 186–192. [Google Scholar] [CrossRef]
- Kjaer, A.; Larsen, P. J.; Knigge, U.; Jorgensen, H.; Warberg, J. Neuronal histamine and expression of corticotropin-releasing hormone, vasopressin and oxytocin in the hypothalamus: relative importance of H1 and H2 receptors. Eur J Endocrinol 1998, 139, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, A. E.; Lookingland, K. J.; Moore, K. E. Histaminergic neurons mediate restraint stress-induced activation of central 5-hydroxytryptaminergic neurons in the rat. Eur J Pharmacol 1994, 264, 163–167. [Google Scholar] [CrossRef]
- Provensi, G.; Passani, M. B.; Costa, A.; Izquierdo, I.; Blandina, P. Neuronal histamine and the memory of emotionally salient events. Br J Pharmacol 2020, 177, 557–569. [Google Scholar] [CrossRef]
- Arnsten, A. F. Stress signalling pathways that impair prefrontal cortex structure and function. Nat Rev Neurosci 2009, 10, 410–422. [Google Scholar] [CrossRef]
- Kjaer, A.; Knigge, U.; Warberg, J. Involvement of oxytocin in histamine- and stress-induced ACTH and prolactin secretion. Neuroendocrinology 1995, 61, 704–713. [Google Scholar] [CrossRef]
- Baettig, L.; et al. The awareness of the scared - context dependent influence of oxytocin on brain function. Brain Imaging Behav 2020, 14, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Shamay-Tsoory, S. G.; Abu-Akel, A. The Social Salience Hypothesis of Oxytocin. Biol Psychiatry 2016, 79, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Leong, I. L.; Wang, X.; Wu, H. Oxytocin's impact on the social brain: Individual differences and context shape a core amygdala-mediated mechanism. Neurosci Biobehav Rev 2026, 183, 106566. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



