
Submitted:
01 February 2026
Posted:
02 February 2026
You are already at the latest version
Abstract
Histamine is traditionally studied either as a peripheral mediator of inflammation and allergic responses or as a central neuromodulator of arousal and wakefulness. These domains, however, are rarely integrated within a unified conceptual framework. Here, we propose that histamine constitutes an evolutionarily conserved system for immune–brain coordination, signalling environmental volatility and mobilising physiological and cognitive resources in response to potential threat. Peripherally, rapid histamine release provides an early, non-specific alert to tissue disruption, while centrally, histaminergic projections reorganise brain state by increasing arousal, sensory gain, and learning under uncertainty. This coupling allows organisms to rapidly shift from energy conservation and stable behavioural policies toward vigilance, exploration, and adaptive updating when environmental conditions become unpredictable. We argue that neuropsychiatric vulnerability may emerge when this alarm system is chronically engaged or poorly calibrated, leading to persistent hyperarousal, sleep disruption, maladaptive learning, and fatigue. Framing histamine as an evolutionary alarm signal offers a coherent account of its roles across immune and neural systems and provides a conceptual basis for understanding immune-related psychiatric risk and for developing more targeted histaminergic interventions.
Keywords:
histamine
; neuroimmune communication
; arousal regulation
; environmental uncertainty
; evolutionary psychiatry
; brain–immune crosstalk
; neuromodulation
1. Introduction
The capacity to detect and respond to environmental challenge is a defining feature of biological systems. In multicellular organisms, this function is distributed across immune, neural, and endocrine systems that evolved not in isolation, but in continuous interaction. Immune signalling shapes motivation, cognition, and behaviour, while brain states regulate immune readiness and energy allocation. Increasing recognition of this bidirectional coupling has transformed our understanding of how inflammation influences mental health, shifting emphasis from discrete disease mechanisms toward dynamic immune–brain coordination [1,2,3].
Within this landscape, histamine occupies a distinctive yet under-theorised position [4]. It is one of the most rapid mediators of peripheral inflammatory responses [5] and one of the most diffusely projecting neuromodulators in the brain [6]. Peripherally, histamine release accompanies tissue injury, infection, and exposure to toxins, producing stereotyped physiological effects that prioritise defence and repair [5]. Centrally, histaminergic neurons modulate wakefulness, attention, sensory processing, and learning, exerting widespread influence over brain state despite their relatively small number [6]. These dual roles have traditionally been studied separately, resulting in parallel literatures that rarely converge.
This separation obscures a more fundamental question: why does the same molecule operate at both immune and neural levels, and what selective pressures shaped this arrangement? From an evolutionary perspective, the co-option of histamine into both systems suggests a shared functional logic rather than coincidental reuse. Histamine’s speed, diffuseness, and lack of specificity make it poorly suited for fine-grained discrimination, but ideal for signalling that prevailing conditions may no longer be safe or predictable [7]. In this sense, histamine appears optimised to function not as a marker of threat identity, but as a signal that existing physiological and behavioural policies require rapid reassessment.
Central histaminergic signalling is particularly well placed to support such reconfiguration [6]. By increasing arousal and sensory gain while modulating learning under uncertainty, histamine promotes rapid updating of internal models when environmental contingencies shift [8]. At the same time, peripheral histamine release mobilises immune and vascular responses that prepare the organism for potential injury or infection. Together, these effects suggest a coordinated system designed to bias the organism toward vigilance, exploration, and short-term survival in volatile contexts [9].
Here, we propose that histamine functions as an evolutionarily conserved alarm system that couples immune activation to brain-state reorganisation in response to environmental volatility [10]. We argue that this framework provides a unifying account of histamine’s peripheral and central actions and offers new insight into how immune processes shape cognition and behaviour. We further suggest that neuropsychiatric vulnerability may arise when this alarm system is chronically engaged or miscalibrated, leading to persistent hyperarousal, sleep disruption, and maladaptive learning. By reframing histamine within an evolutionary immune–brain coordination model, this Perspective aims to clarify its role in mental health and highlight new directions for translational research.
2. Histamine as a Peripheral Alarm Signal
Histamine is among the earliest mediators released during immune activation. Stored preformed in mast cells and basophils, it can be liberated within seconds in response to tissue injury, pathogen-associated signals, toxins, or mechanical stress. This speed distinguishes histamine from cytokines and chemokines that require de novo synthesis and positions it as an initial alarm rather than a downstream effector of immune specificity [11].
The peripheral actions of histamine are stereotyped and highly conserved. Vasodilation, increased vascular permeability, smooth muscle contraction, itch, and pain sensitivity serve to rapidly alter local tissue dynamics while broadcasting the presence of potential danger to the organism [12,13,14,15]. These effects are often described as pathological in modern clinical contexts, yet from an evolutionary standpoint they represent efficient mechanisms for amplifying weak or ambiguous signals of harm. Importantly, histamine does not encode information about the nature of the threat. Instead, it signals that tissue integrity or physiological stability may be compromised, triggering a state of heightened readiness.
This lack of specificity is a defining feature rather than a limitation [16]. In unpredictable environments, rapid detection of deviation from expected conditions is more valuable than accurate classification. Histamine thus functions as a low-threshold detector of volatility, prioritising speed and coverage over precision. By increasing blood flow, facilitating immune cell extravasation, and activating sensory afferents, histamine ensures that local perturbations are rapidly integrated into systemic responses [4].
Peripheral histamine signalling also interfaces directly with the nervous system. Sensory fibres expressing histamine receptors convey itch, pain, and discomfort, generating behavioural urgency and orienting attention toward the affected region. These signals are not simply nociceptive but motivational, biasing behaviour toward withdrawal, grooming, or avoidance [15]. In this way, histamine translates immune activation into action tendencies before higher-order evaluation is possible.
Crucially, peripheral histamine release rarely occurs in isolation. It often accompanies broader immune and neuroendocrine changes, acting as a trigger that lowers thresholds for subsequent responses [15]. Within the framework proposed here, histamine serves as the first component of an integrated alarm system, signalling that prevailing physiological assumptions may no longer hold and preparing both body and brain for rapid adaptation. In the next section, we turn to the central histaminergic system and examine how this peripheral alarm is mirrored by a coordinated reconfiguration of brain state, linking immune activation to arousal, learning, and behavioural flexibility.
3. Central Histamine and Brain-State Reconfiguration
Central histamine is produced by a small population of neurons located in the posterior hypothalamus, whose axons project broadly across cortical and subcortical regions. This anatomical organisation is characteristic of neuromodulatory systems that regulate global brain state rather than transmitting specific information. Despite their limited number, histaminergic neurons exert widespread influence over arousal, attention, sensory processing, and learning [6,8].
Histaminergic activity is tightly coupled to vigilance [17]. Firing rates are highest during wakefulness, decrease during non-rapid eye movement sleep, and are nearly silent during rapid eye movement sleep. Pharmacological blockade of histamine receptors produces sedation, while enhanced histaminergic signalling promotes alertness and resistance to sleep pressure [17]. These effects are often interpreted narrowly as regulation of the sleep–wake cycle. However, such an interpretation underestimates the functional scope of histamine, which extends beyond maintaining wakefulness to reshaping how information is processed during wake.
Central histamine increases neural gain across sensory and associative cortices, amplifying responsiveness to incoming stimuli [18]. This gain modulation enhances signal salience but also increases neural noise, a trade-off that becomes advantageous when environmental contingencies are unstable [19]. In such contexts, reliance on prior expectations may be maladaptive, and increased sensitivity to novel or unexpected input supports rapid updating of internal models [20]. Histamine thus biases cognition toward exploration and learning rather than exploitation of established strategies.
Histamine also interacts closely with other neuromodulatory systems involved in uncertainty and stress, including noradrenaline [21], acetylcholine [22], and corticotropin-releasing hormone [23]. Through these interactions, histamine contributes to a coordinated shift in brain state characterised by heightened arousal, attentional flexibility, and readiness for action. Importantly, this shift is not intrinsically valenced; histamine does not encode threat or reward but alters the conditions under which such information is evaluated.
From an evolutionary perspective, this pattern is consistent with a system designed to signal that the reliability of existing predictions has diminished. Central histamine does not specify what is wrong, but it alters the computational regime of the brain to prioritise rapid detection of change. In doing so, it mirrors the role of peripheral histamine, which signals disruption without defining its cause. Together, these properties suggest that central histamine functions as the neural counterpart of a peripheral immune alarm. By globally reconfiguring brain state, it ensures that immune-derived signals of potential danger are matched by appropriate cognitive and behavioural flexibility. This coordination provides a mechanistic bridge between inflammation, arousal, and learning, setting the stage for understanding how dysregulation of histaminergic signalling may contribute to psychiatric vulnerability. In the next section, we integrate peripheral and central actions of histamine into a unified evolutionary framework centred on environmental volatility and adaptive trade-offs.
4. Histamine, Environmental Volatility, and Adaptive Trade-Offs
Integrating peripheral and central actions, histamine can be understood as a system tuned to detect and respond to environmental volatility rather than to specific threats. Volatility, in this context, refers to situations in which the statistical structure of the environment changes rapidly or becomes unreliable, rendering previously adaptive physiological and behavioural strategies suboptimal. In such conditions, organisms benefit from mechanisms that prioritise rapid detection of change and flexible updating over efficiency and stability [24,25].
Histamine is well suited to this role [26,27]. Its release is rapid, its effects are diffuse, and its signalling lacks fine-grained specificity. These properties allow histamine to act as a global alert that prevailing assumptions may no longer hold [26,27]. Peripherally, this alert mobilises immune and vascular responses that prepare tissues for injury or infection. Centrally, it induces a shift in brain state characterised by increased arousal, enhanced sensory gain, and altered learning dynamics. Together, these changes bias the organism toward vigilance, exploration, and short-term survival.
This shift entails clear trade-offs. States promoted by histaminergic activation are metabolically costly and incompatible with processes optimised for predictable environments, such as sustained sleep, energy conservation, and reliance on habitual behaviour [28]. Increased neural gain enhances sensitivity to novel information but also increases susceptibility to distraction and noise. Similarly, immune readiness improves defence but raises the risk of collateral tissue effects [29]. From an evolutionary standpoint, these costs are acceptable when volatility is transient, but become maladaptive when the alarm state persists [30].
Framing histamine as a volatility signal helps reconcile its apparently paradoxical effects across domains. It explains why histamine promotes wakefulness without conferring positive affect, why it enhances learning under uncertainty while impairing performance in stable conditions, and why its peripheral actions are protective in acute contexts yet pathological when prolonged. Rather than serving a single function, histamine orchestrates a coordinated reallocation of resources in response to changing environmental demands [30].
This framework also highlights why histaminergic systems are vulnerable to dysregulation in modern contexts. Persistent low-grade inflammation, chronic stress, or repeated immune activation can engage the alarm system in the absence of genuine external volatility [30]. Under these conditions, histamine-mediated adaptations become decoupled from their original evolutionary function, leading to sustained hyperarousal and inefficient behavioural strategies. In the next section, we consider how chronic or miscalibrated activation of this alarm system may contribute to neuropsychiatric vulnerability, particularly in conditions characterised by disturbances of arousal, learning, and fatigue.
5. Histamine Dysregulation and Psychiatric Risk
If histamine functions as an adaptive alarm system under conditions of genuine environmental volatility, then psychiatric vulnerability may arise when this system is persistently engaged, insufficiently terminated, or poorly calibrated to actual threat. In such cases, mechanisms that evolved to support short-term survival may begin to undermine long-term cognitive, emotional, and physiological stability [31,32].
Chronic activation of histaminergic signalling is likely to bias the organism toward a sustained state of hyperarousal. Centrally, this may manifest as sleep disruption, heightened sensory sensitivity, attentional instability, and difficulty disengaging from threat-related information. While these features can be adaptive during acute danger, their persistence interferes with restorative processes such as sleep-dependent learning, synaptic homeostasis, and energy regulation. Over time, this imbalance may contribute to fatigue, cognitive inefficiency, and emotional dysregulation [28,32].
At the cognitive level, prolonged histamine-driven gain modulation may distort learning dynamics [28]. Increased sensitivity to novel or unexpected input can impair the consolidation of stable predictive models, leading to excessive updating in response to noise rather than meaningful change. This pattern resembles forms of maladaptive learning observed across several psychiatric conditions, in which individuals remain locked in states of heightened vigilance despite the absence of clear external threat [33]. Rather than reflecting simple anxiety or hyperreactivity, such states may represent a failure to appropriately downregulate an alarm system that was never designed for chronic engagement.
Peripheral immune processes may further amplify this vulnerability. Low-grade inflammation, recurrent infections, or immune dysregulation can repeatedly trigger histamine release, reinforcing central alarm states through both humoral and neural pathways. In this way, immune activity may shape psychiatric risk not only through cytokine-mediated effects on mood and motivation, but also by sustaining neuromodulatory conditions that favour hypervigilance and impaired recovery [34]. Histamine thus provides a plausible mechanistic bridge linking immune activation to disturbances of arousal, cognition, and affect.
This framework aligns with clinical features observed across a range of psychiatric and neuroimmune conditions, including anxiety-related disorders, obsessive–compulsive spectrum disorders, trauma-related syndromes, and post-infectious states. Common to these conditions are alterations in sleep, heightened threat sensitivity, cognitive rigidity, and fatigue, often in the context of ongoing immune or inflammatory activity [35]. Importantly, the present model does not imply that histamine is a primary cause of these disorders, but rather that dysregulated histaminergic signalling may act as a vulnerability factor that shapes symptom expression and persistence.
By reframing psychiatric symptoms as potential consequences of maladaptive alarm signalling, this perspective shifts emphasis away from static disease categories toward dynamic failures of immune–brain coordination. In the following section, we consider the translational implications of this view and its relevance for therapeutic targeting of histaminergic systems.
6. Translational and Therapeutic Implications
Viewing histamine as an evolutionary alarm system has important implications for how immune-related psychiatric symptoms are interpreted and treated [36]. Rather than conceptualising histaminergic activity as intrinsically pathological, this framework emphasises the context in which alarm signalling is engaged and the mechanisms responsible for its resolution. Symptoms such as insomnia, restlessness, sensory hypersensitivity, or cognitive hypervigilance may reflect the persistence of an adaptive response beyond its appropriate temporal window [37].
This perspective may help explain the mixed clinical effects of antihistaminergic interventions in psychiatric and neuroimmune conditions [37]. Non-specific suppression of histamine signalling can reduce arousal and improve sleep, yet may also impair attention, learning, or motivation when used indiscriminately. Such outcomes are consistent with the idea that histamine plays a regulatory role rather than serving as a simple driver of pathology. Therapeutic benefit may therefore depend on selectively retuning alarm thresholds or facilitating termination of histaminergic activation, rather than globally blocking the system.
Receptor-specific approaches may offer greater precision [38,39]. Different histamine receptor subtypes contribute to distinct aspects of immune and neural signalling, and their spatial distribution suggests that targeted modulation could attenuate maladaptive arousal without compromising necessary alertness or immune readiness. Importantly, the timing of intervention is likely to be critical. Interventions that dampen histaminergic activity during periods of unwarranted alarm may restore balance, whereas suppression during genuine environmental challenge could be counterproductive.
The proposed framework also highlights the potential value of histamine-related markers as indicators of immune–brain coupling. Peripheral measures of histamine release or mast cell activation, combined with central indices of arousal or sleep disruption, may help identify individuals in whom alarm systems are persistently engaged [40]. Such stratification could inform personalised treatment strategies and improve the interpretability of heterogeneous psychiatric presentations.
Finally, this perspective has particular relevance for conditions characterised by prolonged immune activation, such as post-infectious syndromes [41]. In these contexts, histaminergic signalling may remain elevated even after the initiating threat has resolved, sustaining symptoms of hyperarousal and fatigue. Understanding histamine as a mediator of immune–brain coordination rather than a disease-specific factor may therefore guide more nuanced approaches to treatment and recovery. In the final section, we summarise the key elements of this framework and outline directions for future research (see also Figure 1 for a summary).
Outstanding Questions for Future Research
Several fundamental questions remain unanswered and define priorities for future research. First, although histamine is rapidly mobilised during immune activation, the mechanisms that normally terminate histaminergic alarm signalling remain poorly understood. Identifying how histamine-driven arousal is actively resolved—at molecular, circuit, and endocrine levels—may be crucial for explaining why alarm states become persistent in some individuals, leading to chronic hyperarousal, sleep disruption, and fatigue.
A second open question concerns the informational content of central histaminergic signalling. While histamine is often linked to arousal or threat, it remains unclear whether it preferentially encodes environmental volatility itself rather than stimulus valence or motivational salience. Experimental paradigms that disentangle unpredictability from threat and reward, combined with computational models of learning under uncertainty, could directly test whether histamine modulates sensitivity to changing environmental statistics rather than specific outcomes.
Third, the pathways linking peripheral histamine release to central histaminergic activation remain insufficiently characterised. Peripheral immune activation may influence brain state through multiple routes, including humoral signalling, vagal afferents, mast cell–neuron interactions, or downstream cytokine cascades, operating over different timescales. Clarifying how fast and slow immune signals converge on central arousal systems will be essential for understanding post-infectious and inflammatory neuropsychiatric syndromes.
A further unresolved issue concerns the boundary between adaptive and maladaptive histaminergic engagement at the level of cognition and learning. While increased neural gain and exploratory learning may be advantageous during transient volatility, sustained histamine-driven gain modulation may impair the consolidation of stable predictive models, leading to excessive updating and cognitive rigidity. Determining how histamine shapes the exploration–exploitation balance across cognitive domains, and whether this balance can be recalibrated, represents a key empirical challenge.
There is also a need to identify reliable biomarkers of chronic histaminergic alarm states in humans. Integrating peripheral measures of histamine release or mast cell activity with central indices of arousal, sleep architecture, and large-scale network dynamics could allow detection of individuals in whom immune–brain alarm systems remain persistently engaged, providing a basis for stratified and mechanism-informed interventions.
Finally, an important translational question is whether histamine-targeted treatments can restore immune–brain coordination without indiscriminately suppressing adaptive alertness. Current antihistaminergic approaches often reduce arousal globally, with mixed cognitive effects. Future work should test whether receptor-specific, circuit-informed, or state-dependent modulation of histaminergic signalling can selectively dampen maladaptive alarm states while preserving the capacity for appropriate vigilance and learning under genuine uncertainty.
7. Conclusions
Histamine has traditionally been studied in fragmented domains, as a mediator of peripheral inflammation, a regulator of arousal and wakefulness, or a pharmacological target with undesirable side effects. In this Perspective, we propose an integrative framework in which histamine is understood as an evolutionarily conserved alarm system that coordinates immune activation with brain-state reconfiguration in response to environmental volatility.
By signalling disruption without encoding specificity, histamine enables rapid mobilisation of physiological and cognitive resources when prevailing assumptions about the environment may no longer be reliable. Peripherally, this alarm promotes immune readiness and tissue defence; centrally, it reshapes arousal, sensory gain, and learning dynamics to prioritise vigilance and adaptive updating. These functions are beneficial when engagement is transient, but become costly when the alarm state persists or is poorly calibrated.
Within this framework, psychiatric vulnerability emerges not from histamine activity per se, but from failures to appropriately engage and disengage this system. Chronic immune activation, sustained stress, or repeated inflammatory insults may prolong histaminergic signalling, contributing to hyperarousal, sleep disruption, maladaptive learning, and fatigue. Framing these features as consequences of dysregulated immune–brain coordination offers a coherent account of symptom overlap across neuropsychiatric and post-inflammatory conditions.
An evolutionary perspective on histamine shifts emphasis from static disease categories toward dynamic regulatory processes that govern adaptation to uncertainty. This view encourages more nuanced therapeutic strategies aimed at restoring balance within alarm systems rather than suppressing them indiscriminately, and highlights histamine as a key mediator at the interface of immunity, brain function, and mental health.
References
- Dantzer, R. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci 2008, 9(1), 46–56. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.H.; Raison, C.L. The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nat Rev Immunol 2016, 16(1), 22–34. [Google Scholar] [CrossRef] [PubMed]
- Hart, B.L., Biological basis of the behavior of sick animals. Neurosci Biobehav Rev 1988, 12(2), 123–37. [CrossRef] [PubMed]
- Schneider, E. Trends in histamine research: new functions during immune responses and hematopoiesis. Trends Immunol 2002, 23(5), 255–63. [Google Scholar] [CrossRef]
- Thurmond, R.L.; Gelfand, E.W.; Dunford, P.J. The role of histamine H1 and H4 receptors in allergic inflammation: the search for new antihistamines. Nat Rev Drug Discov 2008, 7(1), 41–53. [Google Scholar] [CrossRef]
- Haas, H.; Panula, P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci 2003, 4(2), 121–30. [Google Scholar] [CrossRef]
- Passani, M.B.; Blandina, P. Histamine receptors in the CNS as targets for therapeutic intervention. Trends Pharmacol Sci 2011, 32(4), 242–9. [Google Scholar] [CrossRef]
- Sansom, I.I. Histamine control of sleep, learning and memory. Drug Discov Today 2000, 5(3), 94–95. [Google Scholar] [CrossRef]
- Garnuszek, P. Marked survival prolongation of mice bearing a transplantable colon adenocarcinoma by treatment with radioactive platinum-[125I]histamine complex. Preliminary report. Nucl Med Rev Cent East Eur 2004, 7(2), 113–6. [Google Scholar]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef]
- Ribatti, D. The Discovery of Mast Cells: An Historical Note. Clin Anat 2025. [Google Scholar] [CrossRef]
- Wakim, K.G.; Peters, G.A. The effects of intravenously administered histamine on the peripheral circulation in man. J Lab Clin Med 1949, 34(3), 380–6. [Google Scholar] [PubMed]
- Zak-Nejmark, T. Binding of antagonists of H1 and H2 histamine receptors to peripheral blood lymphocytes of atopic and healthy subjects. Arch Immunol Ther Exp (Warsz) 1991, 39(5-6), 549–55. [Google Scholar] [PubMed]
- Baker, C.H.; Menninger, R.P. Histamine-induced peripheral volume and flow changes. Am J Physiol 1974, 226(3), 731–7. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.; Giesler, G.J. The multiple pathways for itch and their interactions with pain. Trends Neurosci 2010, 33(12), 550–8. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454(7203), 428–35. [Google Scholar] [CrossRef]
- Lieberman, P. Histamine, antihistamines, and the central nervous system. Allergy Asthma Proc 2009, 30(5), 482–6. [Google Scholar] [CrossRef]
- Brown, R.E.; Stevens, D.R.; Haas, H.L. The physiology of brain histamine. Prog Neurobiol 2001, 63(6), 637–72. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Cohen, J.D. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci 2005, 28, 403–50. [Google Scholar] [CrossRef]
- Yu, A.J.; Dayan, P. Uncertainty, neuromodulation, and attention. Neuron 2005, 46(4), 681–92. [Google Scholar] [CrossRef]
- Rothwell, N.J.; Stock, M.J.; Wyllie, M.G. Effects of histamine antagonists on noradrenaline-stimulated blood flow and oxygen consumption of brown adipose tissue in the rat. Pflugers Arch 1984, 402(3), 325–9. [Google Scholar] [CrossRef] [PubMed]
- Clapham, J.; Kilpatrick, G.J. Histamine H3 receptors modulate the release of [3H]-acetylcholine from slices of rat entorhinal cortex: evidence for the possible existence of H3 receptor subtypes. Br J Pharmacol 1992, 107(4), 919–23. [Google Scholar] [CrossRef] [PubMed]
- Kjaer, A. Histamine H1 and H2 receptor activation stimulates ACTH and beta-endorphin secretion by increasing corticotropin-releasing hormone in the hypophyseal portal blood. Neuroendocrinology 1992, 56(6), 851–5. [Google Scholar] [CrossRef] [PubMed]
- Behrens, T.E. Learning the value of information in an uncertain world. Nat Neurosci 2007, 10(9), 1214–21. [Google Scholar] [CrossRef]
- Mathys, C.D. Uncertainty in perception and the Hierarchical Gaussian Filter. Front Hum Neurosci 2014, 8, 825. [Google Scholar] [CrossRef]
- Green, J. Histamine and the Nervous System. Fed Proc 1964, 23, 1095–102. [Google Scholar]
- Green, M. Histamine in the central nervous system. Proc West Pharmacol Soc 1978, 21, 337–9. [Google Scholar]
- Tononi, G.; Cirelli, C. Sleep and the price of plasticity: from synaptic and cellular homeostasis to memory consolidation and integration. Neuron 2014, 81(1), 12–34. [Google Scholar] [CrossRef]
- Nathan, C. Points of control in inflammation. Nature 2002, 420(6917), 846–52. [Google Scholar] [CrossRef]
- Miller, A.H. Cytokine targets in the brain: impact on neurotransmitters and neurocircuits. Depress Anxiety 2013, 30(4), 297–306. [Google Scholar] [CrossRef]
- McEwen, B.S. Physiology and neurobiology of stress and adaptation: central role of the brain. Physiol Rev 2007, 87(3), 873–904. [Google Scholar] [CrossRef]
- Mehrhof, S.Z.; Fleming, H.; Nord, C.L. An interoceptive model of energy allostasis linking metabolic and mental health. Sci Adv 2025, 11(39), p. eady4356. [Google Scholar] [CrossRef] [PubMed]
- Browning, M. Anxious individuals have difficulty learning the causal statistics of aversive environments. Nat Neurosci 2015, 18(4), 590–6. [Google Scholar] [CrossRef] [PubMed]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: inflammation and the pathogenesis of depression. Trends Immunol 2006, 27(1), 24–31. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.G. Sleep disturbance as transdiagnostic: consideration of neurobiological mechanisms. Clin Psychol Rev 2011, 31(2), 225–35. [Google Scholar] [CrossRef]
- McEwen, B.S.; Akil, H. Revisiting the Stress Concept: Implications for Affective Disorders. J Neurosci 2020, 40(1), 12–21. [Google Scholar] [CrossRef]
- Kay, G.G. The effects of antihistamines on cognition and performance. J Allergy Clin Immunol 2000, 105 6 Pt 2, S622–7. [Google Scholar] [CrossRef]
- Haas, H.L.; Panula, P. Histamine receptors. Neuropharmacology 2016, 106, 1–2. [Google Scholar] [CrossRef]
- Akdis, C.A.; Simons, F.E. Histamine receptors are hot in immunopharmacology. Eur J Pharmacol 2006, 533(1-3), 69–76. [Google Scholar] [CrossRef]
- Kotrba, J.; Dudeck, A. Mast cells: The Janus of type 2 inflammation. Immunity 2024, 57(6), 1182–1184. [Google Scholar] [CrossRef]
- Tobi, M. Immune Signatures in Post-Acute Sequelae of COVID-19 (PASC) and Myalgia/Chronic Fatigue Syndrome (ME/CFS): Insights from the Fecal Microbiome and Serum Cytokine Profiles. Biomolecules 2025, 15(7). [Google Scholar] [CrossRef]
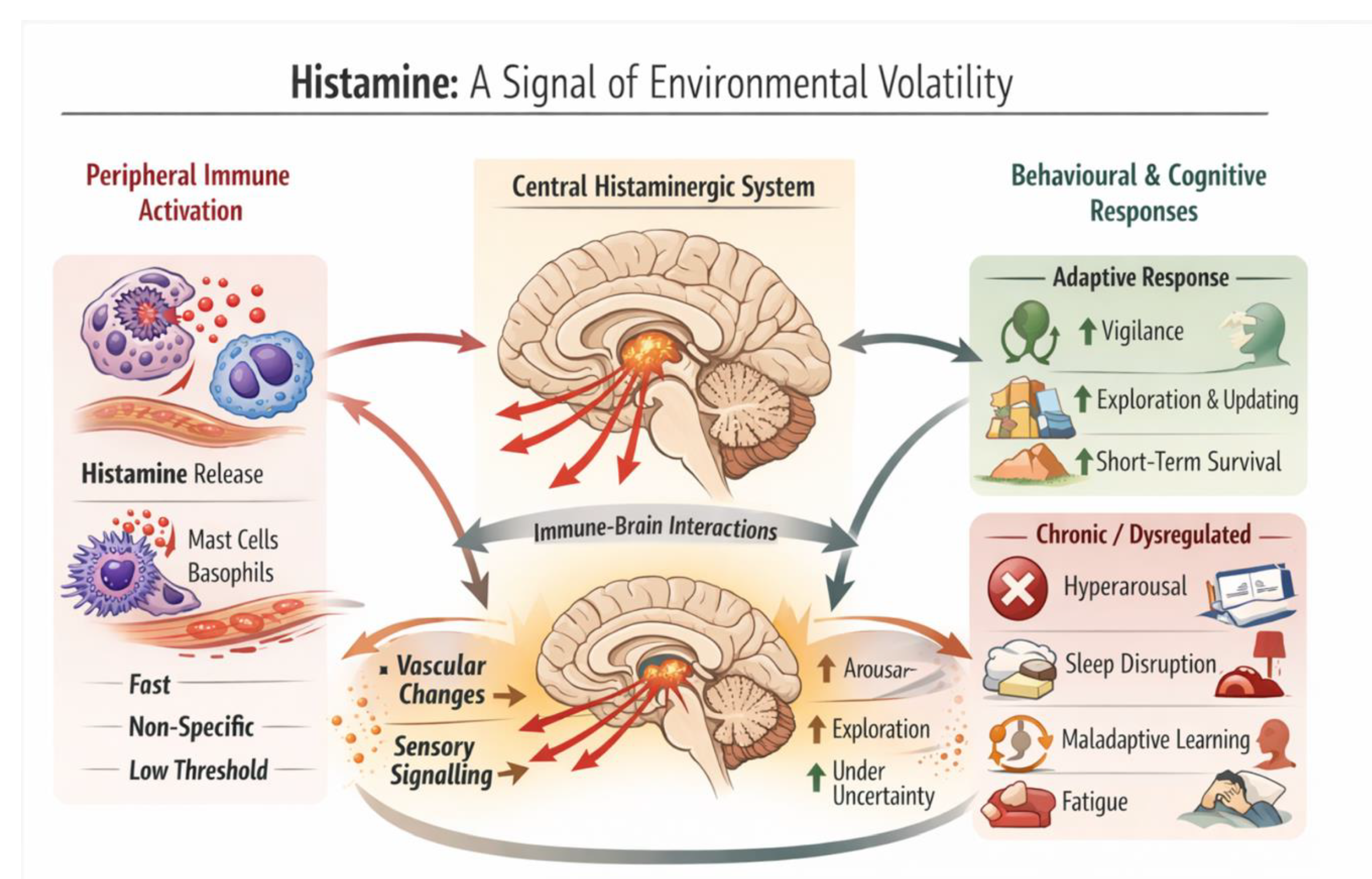
Figure 1.
Histamine as an evolutionary alarm system coordinating immune and brain responses to environmental volatility. Schematic illustration depicting histamine as a rapid, non-specific alarm signal linking peripheral immune activation to central brain-state reconfiguration. Peripheral histamine release from mast cells and basophils signals tissue disruption and mobilises immune and vascular responses. Central histaminergic neurons in the posterior hypothalamus project widely across the brain, increasing arousal, sensory gain, and learning under uncertainty. When transient, this coordinated response promotes vigilance, exploration, and short-term adaptive behaviour. When chronically engaged or dysregulated, persistent alarm signalling may contribute to hyperarousal, sleep disruption, maladaptive learning, fatigue, and increased psychiatric vulnerability. Figure created with the assistance of an AI-based image generation tool (OpenAI DALL·E), followed by conceptual specification, selection, and scientific curation by the author.
Figure 1.
Histamine as an evolutionary alarm system coordinating immune and brain responses to environmental volatility. Schematic illustration depicting histamine as a rapid, non-specific alarm signal linking peripheral immune activation to central brain-state reconfiguration. Peripheral histamine release from mast cells and basophils signals tissue disruption and mobilises immune and vascular responses. Central histaminergic neurons in the posterior hypothalamus project widely across the brain, increasing arousal, sensory gain, and learning under uncertainty. When transient, this coordinated response promotes vigilance, exploration, and short-term adaptive behaviour. When chronically engaged or dysregulated, persistent alarm signalling may contribute to hyperarousal, sleep disruption, maladaptive learning, fatigue, and increased psychiatric vulnerability. Figure created with the assistance of an AI-based image generation tool (OpenAI DALL·E), followed by conceptual specification, selection, and scientific curation by the author.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



