
Submitted:
27 January 2026
Posted:
28 January 2026
You are already at the latest version
Abstract
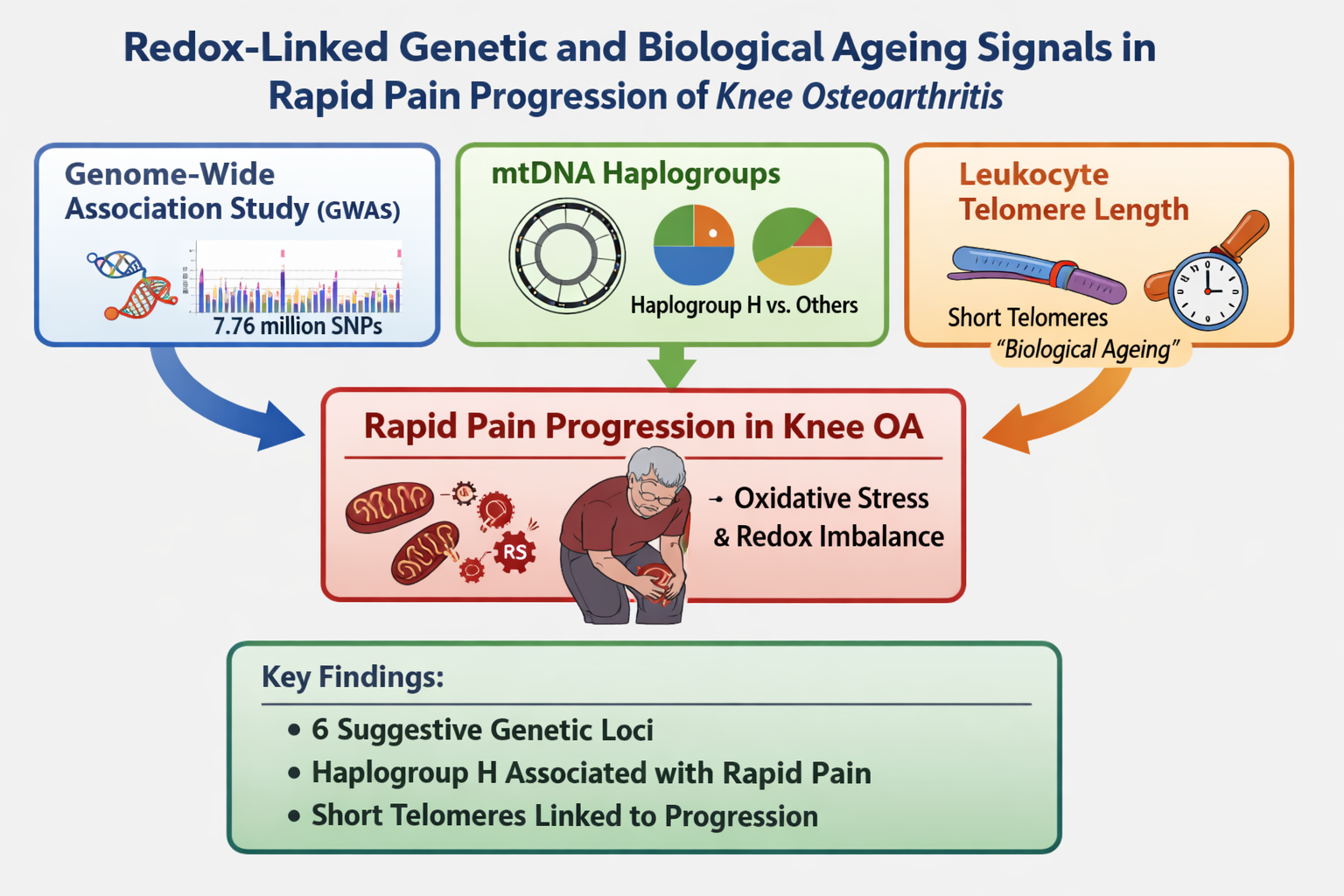
Rapid pain progression in knee osteoarthritis (OA) is heterogeneous and may reflect redox-related mechanisms. We performed an exploratory analysis in Osteoarthritis Initiative (OAI) participants combining nuclear genome-wide association, mitochondrial DNA (mtDNA) haplogroups, and leukocyte telomere length. Rapid pain progression was defined using rescaled Western Ontario and McMaster Universities Osteoarthritis Index (WOMAC) pain (0–100) within 24-month windows. An additive genome-wide association study (GWAS) in 2,946 participants tested 7,762,204 imputed variants, adjusting for age, sex, body mass index (BMI) and three principal components. Haplogroups were analyzed in 3,357 participants, and telomere length (telomere-to-single-copy gene, T/S, ratio) in 301 participants. No variant reached genome-wide significance (p < 5×10−8), but six loci were suggestive (p < 5×10−6) with minimal inflation (λ = 0.995). mtDNA haplogroup H was nominally associated with rapid pain progression (odds ratio, OR = 1.179, p = 0.023). Rapid pain progressors had shorter baseline telomeres (0.825 ± 0.268 vs 0.985 ± 0.375; p < 0.001), and telomere length was inversely associated with progression (OR per 1-unit T/S = 0.260, p = 0.007). These findings are consistent with a redox-linked framework for rapid pain worsening and provide candidates for replication and mechanistic studies.
Keywords:
knee osteoarthritis
; rapid pain progression
; oxidative stress
; redox imbalance
; mitochondrial DNA haplogroups
; leukocyte telomere length
; genome-wide association study
; pain rajectories
1. Introduction
Osteoarthritis (OA) is a chronic and heterogeneous disease and one of the leading causes of pain and disability worldwide. In 2020, an estimated 595 million people were affected (≈7.6% of the global population), representing a marked increase since 1990 [1]. In Spain, the EPISER2016 study estimated a high prevalence of symptomatic OA, including knee OA [2]. With population ageing and the persistence of modifiable risk factors, OA prevalence is projected to increase substantially over coming decades, with major health-economic consequences due to clinical care, pharmacological treatments, and joint replacement surgery [1,3].
Although structural changes are central to OA pathophysiology, pain is the symptom that most strongly drives disability, impaired quality of life, and health-care use. The course of pain in OA is heterogeneous; while some patients maintain stable trajectories or even improve, others experience rapid and sustained progression, suggesting the existence of distinct clinical subphenotypes or patterns [4,5,6,7]. Recent work has standardised operational definitions of rapid progression using 24-month windows, improving comparability across cohorts and supporting the development of predictive models [8,9].
Genetic studies focusing on pain rather than structural damage in knee OA remain limited, but they already suggest that pain has a partly distinct genetic architecture [10,11]. Candidate-gene associations have implicated variants in TRPV1, PCSK6 and COMT in painful versus painless knee OA, supporting nociceptive and monoaminergic contributions that are not fully explained by radiographic severity [12,13,14]. This pain-structure discordance has also been observed in the Spanish PROCOAC cohort, including patients with KL ≤2 who still reported significant pain [15]. More recently, variation in TACR1 and COL11A2 has been linked to knee OA pain and experimental pain sensitivity, consistent with a role for pain sensitisation pathways [16,17]. In addition, functional variation in SCN9A provides a clear proof of concept that germline changes in nociceptive ion channels can shape pain susceptibility [18,19]. At the polygenic level, a GWAS of knee pain in UK Biobank identified loci near GDF5 and COL27A1, indicating that common variants can influence both joint damage and the propensity to report pain [20].
Mitochondrial biology offers a plausible mechanistic bridge between joint pathology and pain amplification. Mitochondria regulate cellular bioenergetics and are major sources of reactive oxygen species (ROS), which act as redox signalling mediators in inflammation and stress responses. Mitochondrial DNA (mtDNA) haplogroups have emerged as modifiers of OA phenotypes; several cohort studies and meta-analyses have shown that specific mtDNA haplogroups influence the prevalence, incidence and progression of OA, including data from the Osteoarthritis Initiative (OAI) and other populations [21,22,23,24]. These effects are consistent with experimental evidence showing that mitochondrial dysfunction, mtDNA damage and oxidative stress contribute to chondrocyte apoptosis, matrix degradation and joint inflammation [25,26,27]. Importantly, mitochondrial energetics and redox imbalance can also modulate neuroimmune pathways and nociceptor sensitisation, raising the possibility that mitochondrial DNA (mtDNA) background influences pain trajectories in knee OA [28,29].
Markers of biological ageing provide an additional layer of vulnerability relevant to redox biology. Leukocyte telomere length reflects cumulative exposure to oxidative stress and systemic low-grade inflammation, processes implicated in OA and in peripheral/central sensitisation of pain pathways [30,31,32].
Combining inherited genetic variation with ageing-related biomarkers may help define a biologically meaningful subphenotype of rapid pain worsening that is plausibly linked to redox imbalance and oxidative stress, processes that can modulate both joint tissue homeostasis and nociceptor sensitisation. To address this hypothesis, we performed a hypothesis-generating exploratory analysis in knee OA participants from the OAI, combining three complementary layers of information: (i) an exploratory single-cohort GWAS to screen for nuclear variants associated with rapid pain progression, (ii) an association analysis of mitochondrial DNA haplogroups as proxies of mitochondrial bioenergetics and redox balance, and (iii) in a smaller subsample, an evaluation of leukocyte telomere length as a marker of biological ageing and cumulative oxidative/inflammatory burden. Together, these analyses were designed to identify candidate genetic and ageing-related signals consistent with an oxidative stress–related contribution to rapid pain progression, providing a framework for future replication and mechanistic studies.
2. Materials and Methods
2.1. Cohort Description
For this study, we used data from the OAI, a prospective, multicentre cohort designed to investigate the onset and progression of knee OA, with publicly accessible longitudinal clinical data, imaging and biospecimens (https://nda.nih.gov/oai) [33]. Participants were followed annually for up to ten years using standardised protocols to quantify changes in knee health over time, capturing the onset and progression of symptoms and functional limitations, the development and worsening of structural abnormalities, and longitudinal variation in imaging and biomarker-based indicators of knee OA, with ancillary assessments in other joints when available.
For the descriptive characterisation of the study cohort, we included key clinical variables available in the OAI dataset. Specifically, sex, age and body mass index (BMI) were recorded at baseline for OAI participants.
All participating clinical centres complied with the Declaration of Helsinki and obtained local ethics approval. All participants provided written informed consent.
2.2. Rapid Pain Progression Criteria
We operationalised rapid pain progression using scaled WOMAC pain scores (0–100) and standardised 24-month windows, following previously published approaches [8,9]. Participants were classified as rapid pain progressors if, in any valid 24-month window, they met at least one of the following criteria:
- Increase of at least 10 points over 24 months and substantial pain at the end of the window (WOMAC pain ≥40); OR
- Increase of at least 20 points over 24 months with end pain ≥35; OR
- Sustained substantial pain (WOMAC pain ≥40 at both the beginning and end of the 24-month window).
A 24-month window was considered valid if WOMAC pain scores were available at all annual visits within the window. Participants were classified as non-rapid pain progressors only if they had at least one valid observation period of 48 months or longer in which they did not meet rapid pain progression criteria in any embedded 24-month interval. This conservative control definition was used to reduce misclassification of participants with intermittent rapid worsening or missing follow-up assessments.
2.3. Genetic Analyses
Genetic information for this study was obtained from two different sources: nuclear genome-wide data and mitochondrial data.
2.3.1. Nuclear Genome-Wide Data
We used nuclear genome-wide genotype data generated as part of the Genetic Components of Knee Osteoarthritis (GeCKO) study [34] in OAI participants. These data had been previously deposited in dbGaP (accession phs000955.v1.p1) and were used to extract single nucleotide polymorphisms (SNPs) across the genome for association analyses.
Genotyping was performed in 4,219 OAI participants using the Illumina HumanOmni2.5 BeadChip. Quality control (QC) was conducted with PLINK 2.0 [35] and excluded single nucleotide polymorphisms (SNPs) with low call rate, duplicates, monomorphic variants, minor allele frequency (MAF) < 0.05 or significant deviation from Hardy–Weinberg equilibrium (p < 1×10−6). At the sample level, individuals with >3% missing genotypes were removed; at the variant level, SNPs with >2% missingness were filtered out. We further excluded duplicate samples, heterozygosity outliers (>±4 SD from the mean) and ancestry outliers (>±8 SD from the European cluster in the 1000 Genomes reference principal component analysis (PCA) space). Post-QC VCF files were imputed on the Michigan Imputation Server (Minimac4), retaining variants with imputation quality (Rsq) >0.8, yielding approximately 8 million SNPs.
2.3.2. Mitochondrial Data
Mitochondrial genetic variation was captured through European mitochondrial DNA (mtDNA) haplogroups, which had been previously assigned by our group to OAI participants following a single base extension (SBE) assay-based protocol described elsewhere [21].
2.4. Telomere Length Assay
A subset of (N=301) OAI participants was used to examine the influence of baseline telomere length on the risk of rapid pain progression. The telomere length of peripheral blood leukocytes (PBLs) from this subset of patients had been previously characterised [31] using a validated quantitative PCR-based assay, as described elsewhere [36]. Briefly, this method quantifies the average ratio of telomere repeat copy number to a single-copy reference gene (36B4) in each sample (T/S ratio) and provides an indirect estimate of mean telomere length.
2.5. Statistical Analyses
Data were analyzed using IBM SPSS Statistics, version 29 (IBM Corp., Armonk, NY, USA) and R software, version 4.4.1 (R Foundation for Statistical Computing, Vienna, Austria).
The genome-wide association study (GWAS) was performed using PLINK 2.0 for the post-imputation processing, and rvtests [37] for the association analyses under an additive genetic model, adjusting for age, sex, body mass index (BMI) and the first three principal components of ancestry derived from PCA on a pruned set of hard-called SNPs using stricter missingness and relatedness thresholds. The final dataset comprised 2,946 QC-passed OAI participants of European ancestry and 7,762,204 SNPs, with rapid pain progression (yes/no) as the outcome. Genome-wide significance was defined as p<5×10−8. To summarise the results and prioritise independent association signals, we applied linkage disequilibrium (LD)-based clumping in PLINK 2.0 (r2 > 0.2 within the study sample), retaining the lead SNP with the lowest p-value in each linkage disequilibrium cluster. Lead variants with p<5×10−6 were considered suggestive and were reported as hypothesis-generating candidates.
Haplogroup and allele frequencies between rapid pain progressors and non-rapid pain progressors were compared using chi-square tests from contingency tables. Odds ratios (ORs) and 95% confidence intervals (CIs) were calculated to assess the odds of carrying each allele in rapid pain progressors compared with non-progressors. For the haplogroup analysis, we followed a previously described approach, comparing each haplogroup with all remaining haplogroups pooled into a single reference category. In addition, a regression model adjusting for the confounder variables of sex, age and BMI was constructed to assess independent associations between mtDNA haplogroups and rapid pain progression.
PBL telomere length (T/S ratio) was first compared between rapid and non-rapid pain progressors using the non-parametric Mann-Whitney U test. We then performed a logistic regression, adjusting for sex, age and BMI, to evaluate PBL telomere length as a potential risk factor associated with rapid pain progression.
2.6. Declaration of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this work the author(s) used ChatGPT in order to adjust the different R scripts. After using this tool, the author(s) reviewed and edited the content as needed and take(s) full responsibility for the content of the publication.
3. Results
3.1. Study Population
The complete dataset included a total of 3,395 OAI participants of European ancestry, of which 1,906 (56.1%) were women and 1,489 (43.9%) were men. Overall, 1,260 (37.1%) participants met the criteria for rapid pain progression and 2,135 (62.9%) were classified as non-rapid pain progressors. A descriptive analysis of this population showed that rapid pain progressors were more likely to be female (p<0.001) and to have higher BMI (p<0.001) than non-rapid pain progressors (Table 1).
3.2. Nuclear Genome-Wide Analysis
We conducted a GWAS to identify nuclear genetic variants associated with rapid pain progression. As described in the Methods section, the final GWAS dataset included 2,946 QC-passed OAI participants of European ancestry and 7,762,204 imputed SNPs, with rapid pain progression (yes/no) as the outcome. Overall, the GWAS showed no evidence of substantial test statistic inflation (genomic inflation factor λ = 0.995), and the quantile-quantile (QQ) plot suggested adequate control of population stratification and relatedness (Supplementary Figure S1). The Manhattan plot is shown in Figure 1.
No SNP reached the conventional threshold for genome-wide significance (p < 5×10−8). However, we identified 6 independent loci that met the predefined suggestive significance threshold (p < 5×10−6) and were therefore considered candidate signals for further evaluation (Table 2). The strongest association signal was observed for rs73631790 on chromosome 19q13.43 (p = 7.195×10−7), located in the proximal 5′ flanking region of DUXA, a double homeobox transcription factor gene. Another prominent signal, rs2862774 on chromosome 2q14.1 (p = 1.03×10−6), maps intronically within IL36B, which encodes the interleukin-36β cytokine. Among the remaining loci, rs9912678 on 17p11.2 (p = 3.37×10−6) lies in the upstream flanking region of RAI1; rs57987665 on 14q21.3 (p = 3.62×10−6) falls in an intergenic region with the closest annotated transcript being the long non-coding RNA LOC105378178; rs34699810 on 3p26.1 (p = 4.11×10−6) is intergenic and nearest to the antisense lncRNA LMCD1-AS1; and rs62147861 on chromosome 2 (p = 4.06×10−6) is located in proximity to the uncharacterised non-coding RNA LOC124907827. Effect sizes for these variants were modest to moderate (Table 2), as expected for complex traits.
Given the limited sample size and the absence of genome-wide significance findings, these association signals should be interpreted as exploratory and hypothesis-generating. Replication in independent cohorts and functional follow-up will be required to confirm their role in rapid pain progression.
3.3. Mitochondrial DNA Analysis
Haplogroup data were available for 3,357 OAI participants, of which 1,239 (36.91%) were rapid pain progressors and 2,118 (63.09%) were non-rapid pain progressors. The frequency distribution of major European mtDNA haplogroups ranged from 9% for haplogroup J to 40.9% for the most common haplogroup H (Table 3). In unadjusted analyses, only mtDNA haplogroup H showed a nominally significant over-representation in the rapid pain progressors group (43.4%) compared with non-rapid progressors (39.4%) (OR = 1.179; 95%CI = 1.023-1.359; p = 0.023). In addition, the pooled “other” haplogroup category (individual haplogroup frequency <5%) was nominally associated with a lower risk of rapid pain progression (OR = 0.792; 95%CI = 0.650-0.966; p = 0.021); however, given its heterogeneous composition, this finding was considered exploratory and was not interpreted further. No other haplogroup showed a statistically significant difference in frequency (Table 3).
In multivariable logistic regression, carriers of haplogroup H remained more likely to be rapid pain progressors (OR = 1.172; 95%CI = 1.012-1.357; p = 0.034), and rapid pain progression was also independently associated with female sex (OR = 1.494; 95%CI = 1.289-1.732; p < 0.001), age (OR = 1.009; 95%CI = 1.001-1.017; p = 0.031) and higher BMI (OR = 1.113; 95%CI = 1.095-1.131; p < 0.001) (Table 4).
3.4. Telomere Length Analysis
In the subset of 301 OAI participants with available PBL telomere measurements, 103 (34.22%) were classified as rapid pain progressors and 198 (65.78%) as non-rapid pain progressors. Baseline PBL telomere length (T/S ratio) was 0.825 ± 0.268 in rapid pain progressors and 0.985 ± 0.375 in non-rapid pain progressors (p < 0.001, Mann–Whitney U test). In multivariable logistic regression adjusting for sex, age and BMI, PBL telomere length was inversely associated with rapid pain progression (OR per 1-unit increase in T/S = 0.260; 95%CI = 0.098-0.693; p = 0.007) (Table 5).
4. Discussion
This hypothesis-generating study aimed to investigate redox-linked genetic and biological ageing determinants of rapid pain progression in knee OA using participants from the OAI. Using a standardised 24-month window definition, we combined a discovery GWAS with mtDNA haplogroup profiling, and additionally evaluated baseline PBL telomere length in a subset of participants. As an exploratory GWAS without external replication or meta-analysis, these findings should be interpreted cautiously: no variants reached the conventional genome-wide significance, but six suggestive loci were identified as hypothesis-generating candidates. In parallel, mtDNA haplogroup H was nominally over-represented among rapid pain progressors, and shorter baseline telomere length was associated with increased odds of rapid pain progression after adjustment for sex, age and BMI.
To our knowledge, none of the six lead SNPs identified in this study has been previously reported as associated with OA or pain phenotypes, nor do they overlap with the established set of genome-wide significant loci for structural OA described in recent large-scale meta-analyses [11,38,39,40]. Notably, our GWAS focused on a strictly defined longitudinal phenotype of rapid pain worsening, whereas most prior OA pain genetics has examined cross-sectional symptomatic status or broader pain traits; together with limited power in a single-cohort design, this may partly explain the lack of overlap with previously reported loci.
At the gene level, IL36B encodes interleukin-36β, a member of the IL-1 cytokine family that stimulates the production of IL-6, IL-8 and matrix metalloproteinases by synovial fibroblasts and articular chondrocytes, and IL-36 family cytokines are increasingly recognised as mediators of joint inflammation [41,42,43]. Moreover, spinal IL-36γ/IL-36R signalling has been shown to participate in the maintenance of chronic inflammatory pain in preclinical models, supporting a plausible link between this locus and pathways of inflammatory nociception [43,44]. Notably, IL-36 signalling has also been shown to modulate glutathione homeostasis and reactive oxygen species (ROS) resolution in experimental systems, providing a potential mechanistic bridge between inflammatory activation and redox imbalance [45].
RAI1, the nearest gene to the 17p11.2 signal, is the dosage-sensitive transcription factor responsible for Smith–Magenis syndrome, a neurodevelopmental disorder characterised by altered temperature and pain sensitivity, and RAI1 dosage modulates pain sensitivity in mouse models [46,47]; in addition, RAI1 has recently been prioritised as a putative effector gene at an all-OA locus in integrative analyses that combine OA GWAS with osteoclast regulatory data [48]. Beyond nociception-related phenotypes, patient-derived and cellular models of RAI1 haploinsufficiency have reported both mitochondrial and autophagy alterations accompanied by increased oxidative stress/ROS signatures, further supporting a redox-linked biological plausibility for this locus [49].
By contrast, DUXA is a paralogue of DUX4, an embryonic transcription factor causally involved in facioscapulohumeral muscular dystrophy, and although members of the DUX family are important regulators of early myogenic programmes, no direct role of DUXA in OA or pain has yet been reported [50,51]. However, DUX4 has been tightly connected to oxidative stress biology in muscle, including evidence that oxidative stress can upregulate DUX4 expression and that DUX4-related phenotypes involve ROS/DNA damage pathways that can be mitigated by antioxidant strategies, providing an indirect redox-relevant context for this gene family [52].
The remaining loci map to poorly characterised long non-coding RNAs: LMCD1-AS1, which has been implicated as an oncogenic lncRNA in osteosarcoma, cervical and thyroid cancers, and the uncharacterised ncRNAs LOC105378178 and LOC124907827, for which no specific function in musculoskeletal disease or nociceptive pathways is currently known [53,54].
Taken together, our findings did not identify definitive genome-wide significant variants for rapid pain progression, but they highlighted a small set of biologically plausible candidate loci, particularly at IL36B and RAI1, with potential links to inflammatory–redox mechanisms that merit further investigation. In line with the single-cohort design and the use of a suggestive significance threshold, these association signals should be regarded as exploratory and primarily hypothesis-generating, and replication in independent cohorts and functional studies will be required to confirm whether these loci contribute to the genetic architecture of rapid pain worsening in knee OA.
Our finding that mtDNA haplogroup H was nominally over-represented among rapid pain progressors is consistent with a growing body of evidence indicating that mitochondrial genetic background can modulate OA phenotypes. In European-descent populations, mtDNA haplogroups have been linked to knee OA prevalence and severity, as well as to disease incidence and progression across cohorts including the OAI, supporting the concept that mtDNA lineages may influence joint vulnerability beyond traditional risk factors [21,23,55,56]. Mechanistically, mtDNA haplogroups differ in oxidative phosphorylation performance and redox balance; haplogroup H has been associated with higher oxygen consumption and oxidative damage compared with haplogroups such as J, which has been discussed as a more uncoupled/low-ROS background in OA-related contexts [22,57]. Given the established links between mitochondrial dysfunction/oxidative stress and neuroimmune signalling in pain sensitisation, mtDNA haplogroups could plausibly contribute not only to structural trajectories but also to pain trajectories in knee OA [28,58]. Nevertheless, because the observed association is nominal and the “Other” haplogroup category aggregates heterogeneous lineages, these results should be interpreted cautiously and require replication in independent datasets with comparable longitudinal pain phenotyping.
In the telomere substudy, rapid pain progressors exhibited shorter baseline leukocyte telomere length, supporting the notion that biological ageing and cumulative systemic stress may modulate symptomatic worsening in knee OA. Prior work in the OAI has linked shorter leukocyte telomere length to radiographic knee OA and greater structural burden, and longitudinal analyses have further suggested that accelerated telomere attrition is associated with incident knee OA during follow-up [31,59]. Importantly, telomere length has also been related to the biological burden of chronic knee OA pain: individuals with higher chronic pain severity, and/or high perceived stress in the context of chronic knee OA pain, show shorter telomeres, consistent with an accelerated ageing signal in more severe symptomatic presentations [30,60]. Conceptually, telomere length is an integrative marker of biological aging, influenced by oxidative and inflammatory stressors that are relevant to OA pathophysiology and pain sensitisation [32]. While our results align with this framework, they derive from a subset of patients and, therefore, warrant cautious interpretation and replication, ideally in cohorts with repeated telomere measures and harmonized longitudinal pain phenotypes.
This study has several limitations. First, the genome-wide component was conducted in a single discovery cohort without external replication or meta-analysis, and no associations reached conventional genome-wide significance; therefore, the identified loci should be regarded as exploratory and hypothesis-generating. Second, although we used a strict and clinically meaningful definition of rapid pain progression, misclassification remains possible given the inherent variability of self-reported pain and the influence of time-varying factors such as analgesic use, intra-articular treatments and comorbidities. Third, the telomere analysis was restricted to a relatively small subset of participants, which limits power and may introduce selection bias, and the “other” mtDNA haplogroup category aggregates heterogeneous lineages, reducing biological interpretability. As contingency measures, we report effect sizes with confidence intervals, emphasise internal consistency across complementary genetic layers (nuclear GWAS, mtDNA haplogroups and telomere length), and interpret all signals cautiously within an explicitly exploratory framework. Future work will prioritise replication in independent cohorts with harmonised longitudinal pain phenotyping and, where possible, repeated telomere measurements and functional follow-up to strengthen causal inference and clarify underlying mechanisms.
5. Conclusions
This hypothesis-generating analysis (Figure 2) suggests that rapid pain progression in knee OA may be associated with biological signals consistent with a redox-linked biology framework, spanning nuclear immune–neuroimmune genetic variation (including a suggestive IL36B signal), mitochondrial genetic background, and systemic biological ageing indexed by leukocyte telomere length. Together, these findings provide hypothesis-generating leads for replication and mechanistic studies; however, as oxidative stress was not directly measured and the genome-wide component lacked external replication, independent validation, functional follow-up and integrative multi-omics studies will be required to confirm these associations and to identify actionable redox and mitochondrial targets for preventing rapid pain worsening.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figure S1. Quantile–quantile (QQ) plot of GWAS p-values for rapid pain progression in knee osteoarthritis. Observed −log10(P) values are plotted against those expected under the null hypothesis for 7,762,204 imputed SNPs tested in OAI participants of European ancestry under an additive model adjusted for age, sex, BMI and the first three ancestry principal components. The close agreement with the null line indicates adequate control of population stratification and relatedness, with minimal test statistic inflation (genomic inflation factor λ = 0.995); deviation is confined to the extreme tail, consistent with a small number of suggestive association signals.
Author Contributions
FJB and IRP contributed equally in the design and coordination of the study; both conceived the study, participated in its design and helped to draft the final version of the manuscript; NO coordinated the access to the biological samples and the clinical information used in this study; LMS and DMC contributed equally in the statistical analysis of data; JVG and AMT helped in the analysis of the GWAS data; JLFG, AM and FO coordinated and performed the telomere assays; SR performed the genetic assignments of mtDNA haplogroups.
Funding
This study has been funded by Instituto de Salud Carlos III (ISCIII) through the projects RD21/0002/0009, RD24/0007/0026, PMP22/00101, PMPTA22/00115, PI22/01155 and PI23/00913 and co-funded by the European Union. This work was also funded by grants IN607A 2021/07 and IN607D 2021/13 from Axencia Galega de Innovación-Xunta de Galicia. IRP is supported by Contrato Miguel Servet-II Fondo de Investigación Sanitaria (CPII17/00026) SERGAS-stabilized. JVG is supported by grant IN606A 2022/048 from Xunta de Galicia, Spain. This work was also funded by Pfizer & Eli Lilly and Company through the 3rd Global Awards for Advancing Chronic Pain Research, ADVANCE (grant ID#64122119).
Institutional Review Board Statement
This study has received the favorable opinion of the ethics committee of XUNTA de Galicia, registration number 2024/074.
Informed Consent Statement
To the best of our knowledge, patients of the OAI cohort are informed about the goal of their enrollment in these prospective studies. The entity responsible of this cohort will be the first to receive a copy of this manuscript once it is published. In addition, all the clinical centers involved in the recruitment of the patients have made provisions to ensure the safety, confidentiality and ethical treatment of study participants according to the Declaration of Helsinki. In this sense, all the participants signed an informed consent.
Data availability statement
Publicly available datasets were analyzed in this study. OAI data can be accessed via https://nda.nih.gov/oai. Derived data are available from the corresponding author upon reasonable request.
Acknowledgments
The Osteoarthritis Initiative (OAI) is a public-private partnership comprised of five contracts (N01-AR-2-2258; N01-AR-2-2259; N01-AR-2-2260; N01-AR-2-2261; N01-AR-2-2262) funded by the National Institutes of Health, a branch of the Department of Health and Human Services, and conducted by the OAI Study Investigators. Private funding partners include Pfizer, Inc.; Novartis Pharmaceuticals Corporation; Merck Research Laboratories; and GlaxoSmithKline. Private sector funding for the OAI is managed by the Foundation for the National Institutes of Health. This manuscript was prepared using an OAI public use data set and does not necessarily reflect the opinions or views of the OAI investigators, the NIH, or the private funding partners.
Conflicts of interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BMI | Body mass index |
| CI | Confidence interval |
| COL11A2 | Collagen type XI alpha 2 chain |
| COL27A1 | Collagen type XXVII alpha 1 chain |
| COMT | Catechol-O-methyltransferase |
| dbGaP | Database of Genotypes and Phenotypes |
| DNA | Deoxyribonucleic acid |
| DUX4 | Double homeobox 4 |
| DUXA | Double homeobox A |
| GDF5 | Growth differentiation factor 5 |
| GeCKO | Genetic Components of Knee Osteoarthritis |
| GWAS | Genome-wide association study |
| HWE | Hardy–Weinberg equilibrium |
| IBM | International Business Machines |
| IL | Interleukin |
| IL-1 | Interleukin-1 |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| IL-36 | Interleukin-36 |
| IL-36γ | Interleukin-36 gamma |
| IL-36R | Interleukin-36 receptor |
| IL36B | Interleukin 36 beta |
| LD | Linkage disequilibrium |
| LMCD1-AS1 | LMCD1 antisense RNA 1 |
| lncRNA | Long non-coding RNA |
| LOC105378178 | Uncharacterised non-coding RNA locus |
| LOC124907827 | Uncharacterised non-coding RNA locus |
| MAF | Minor allele frequency |
| Minimac4 | Minimac4 imputation software |
| mtDNA | Mitochondrial DNA |
| OA | Osteoarthritis |
| OAI | Osteoarthritis Initiative |
| OR | Odds ratio |
| PBL | Peripheral blood leukocytes |
| PCA | Principal component analysis |
| PC | Principal component |
| PCR | Polymerase chain reaction |
| QC | Quality control |
| Quantile–quantile | |
| R | R statistical software/environment |
| RAI1 | Retinoic acid induced 1 |
| ROS | Reactive oxygen species |
| r2 | Squared correlation coefficient |
| RNA | Ribonucleic acid |
| Rsq | Imputation quality metric |
| SBE | Single base extension |
| SCN9A | Sodium voltage-gated channel alpha subunit 9 |
| SD | Standard deviation |
| SNP | Single nucleotide polymorphism |
| SPSS | Statistical Package for the Social Sciences |
| T/S | Telomere-to-single-copy gene ratio |
| TACR1 | Tachykinin receptor 1 |
| TRPV1 | Transient receptor potential vanilloid 1 |
| UK | United Kingdom |
| USA | United States of America |
| VCF | Variant call format |
| WOMAC | Western Ontario and McMaster Universities Osteoarthritis Index |
References
- Collaborators GO. Global, regional, and national burden of osteoarthritis, 1990-2020 and projections to 2050: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Rheumatol 2023, 5(9), e508–e522. [Google Scholar] [CrossRef] [PubMed]
- Blanco, FJ; Silva-Díaz, M; Quevedo Vila, V; Seoane-Mato, D; Pérez Ruiz, F; Juan-Mas, A; et al. Prevalence of symptomatic osteoarthritis in Spain: EPISER2016 study. Reumatol Clin (Engl Ed) 2021, 17(8), 461–470. [Google Scholar] [CrossRef] [PubMed]
- Leifer, VP; Katz, JN; Losina, E. The burden of OA-health services and economics. Osteoarthritis Cartilage 2022, 30(1), 10–16. [Google Scholar] [CrossRef]
- Collins, JE; Katz, JN; Dervan, EE; Losina, E. Trajectories and risk profiles of pain in persons with radiographic, symptomatic knee osteoarthritis: data from the osteoarthritis initiative. Osteoarthritis Cartilage 2014, 22(5), 622–630. [Google Scholar] [CrossRef] [PubMed]
- Lee, AC; Harvey, WF; Han, X; Price, LL; Driban, JB; Bannuru, RR; et al. Pain and functional trajectories in symptomatic knee osteoarthritis over up to 12 weeks of exercise exposure. Osteoarthritis Cartilage 2018, 26(4), 501–512. [Google Scholar] [CrossRef]
- Radojčić, MR; Arden, NK; Yang, X; Strauss, VY; Birrell, F; Cooper, C; et al. Pain trajectory defines knee osteoarthritis subgroups: a prospective observational study. Pain 2020, 161(12), 2841–2851. [Google Scholar] [CrossRef] [PubMed]
- Johnson, AJ; Vasilopoulos, T; Booker, SQ; Cardoso, J; Terry, EL; Powell-Roach, K; et al. Knee pain trajectories over 18 months in non-Hispanic Black and non-Hispanic White adults with or at risk for knee osteoarthritis. BMC Musculoskelet Disord 2021, 22(1), 415. [Google Scholar] [CrossRef]
- Widera, P; Welsing, PMJ; Ladel, C; Loughlin, J; Lafeber, FPFJ; Petit Dop, F; et al. Multi-classifier prediction of knee osteoarthritis progression from incomplete imbalanced longitudinal data. Sci Rep. 2020, 10(1), 8427. [Google Scholar] [CrossRef]
- Castagno, S; Birch, M; van der Schaar, M; McCaskie, A. Predicting rapid progression in knee osteoarthritis: a novel and interpretable automated machine learning approach, with specific focus on young patients and early disease. Ann Rheum Dis. 2025, 84(1), 124–135. [Google Scholar] [CrossRef]
- Thakur, M; Dawes, JM; McMahon, SB. Genomics of pain in osteoarthritis. Osteoarthritis Cartilage 2013, 21(9), 1374–1382. [Google Scholar] [CrossRef]
- Aubourg, G; Rice, SJ; Bruce-Wootton, P; Loughlin, J. Genetics of osteoarthritis. Osteoarthritis Cartilage 2022, 30(5), 636–649. [Google Scholar] [CrossRef]
- Valdes, AM; De Wilde, G; Doherty, SA; Lories, RJ; Vaughn, FL; Laslett, LL; et al. The Ile585Val TRPV1 variant is involved in risk of painful knee osteoarthritis. Ann Rheum Dis. 2011, 70(9), 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Malfait, AM; Seymour, AB; Gao, F; Tortorella, MD; Le Graverand-Gastineau, MP; Wood, LS; et al. A role for PACE4 in osteoarthritis pain: evidence from human genetic association and null mutant phenotype. Ann Rheum Dis. 2012, 71(6), 1042–1048. [Google Scholar] [CrossRef]
- Neogi, T; Soni, A; Doherty, SA; Laslett, LL; Maciewicz, RA; Hart, DJ; et al. Contribution of the COMT Val158Met variant to symptomatic knee osteoarthritis. Ann Rheum Dis. 2014, 73(1), 315–317. [Google Scholar] [CrossRef] [PubMed]
- Oreiro-Villar, N; Raga, AC; Rego-Pérez, I; Pértega, S; Silva-Diaz, M; Freire, M; et al. PROCOAC (PROspective COhort of A Coruña) description: Spanish prospective cohort to study osteoarthritis. Reumatol Clin (Engl Ed) 2022, 18(2), 100–104. [Google Scholar] [CrossRef]
- Warner, SC; Walsh, DA; Laslett, LL; Maciewicz, RA; Soni, A; Hart, DJ; et al. Pain in knee osteoarthritis is associated with variation in the neurokinin 1/substance P receptor (TACR1) gene. Eur J Pain 2017, 21(7), 1277–1284. [Google Scholar] [CrossRef]
- Ho, KWD; Wallace, MR; Sibille, KT; Bartley, EJ; Cruz-Almeida, Y; Glover, TL; et al. Single Nucleotide Polymorphism in the COL11A2 Gene Associated with Heat Pain Sensitivity in Knee Osteoarthritis. Mol Pain. 2017, 13, 1744806917724259. [Google Scholar]
- Reimann, F; Cox, JJ; Belfer, I; Diatchenko, L; Zaykin, DV; McHale, DP; et al. Pain perception is altered by a nucleotide polymorphism in SCN9A. Proc Natl Acad Sci U S A 2010, 107(11), 5148–5153. [Google Scholar] [CrossRef]
- Klein, CJ; Wu, Y; Kilfoyle, DH; Sandroni, P; Davis, MD; Gavrilova, RH; et al. Infrequent SCN9A mutations in congenital insensitivity to pain and erythromelalgia. J Neurol Neurosurg Psychiatry 2013, 84(4), 386–391. [Google Scholar] [CrossRef]
- Meng, W; Adams, MJ; Palmer, CNA; Shi, J; Auton, A; Ryan, KA; et al. Genome-wide association study of knee pain identifies associations with GDF5 and COL27A1 in UK Biobank. Commun Biol. 2019, 2, 321. [Google Scholar] [CrossRef] [PubMed]
- Rego-Perez, I; Fernandez-Moreno, M; Fernandez-Lopez, C; Arenas, J; Blanco, FJ. Mitochondrial DNA haplogroups: role in the prevalence and severity of knee osteoarthritis. Arthritis Rheum 2008, 58(8), 2387–2396. [Google Scholar] [CrossRef] [PubMed]
- Soto-Hermida, A; Fernandez-Moreno, M; Oreiro, N; Fernandez-Lopez, C; Pertega, S; Cortes-Pereira, E; et al. Mitochondrial DNA (mtDNA) haplogroups influence the progression of knee osteoarthritis. Data from the Osteoarthritis Initiative (OAI). PloS one 2014, 9(11), e112735–e112735. [Google Scholar] [CrossRef]
- Fernandez-Moreno, M; Soto-Hermida, A; Vazquez-Mosquera, ME; Cortes-Pereira, E; Pertega, S; Relano, S; et al. A replication study and meta-analysis of mitochondrial DNA variants in the radiographic progression of knee osteoarthritis. Rheumatology 2017, 56(2), 263–270. [Google Scholar] [CrossRef] [PubMed]
- Durán-Sotuela, A; Fernandez-Moreno, M; Suárez-Ulloa, V; Vázquez-García, J; Relaño, S; Hermida-Gómez, T; et al. A meta-analysis and a functional study support the influence of mtDNA variant m.16519C on the risk of rapid progression of knee osteoarthritis. Ann Rheum Dis 2023. [Google Scholar] [CrossRef] [PubMed]
- Gavriilidis, C; Miwa, S; von Zglinicki, T; Taylor, RW; Young, DA. Mitochondrial dysfunction in osteoarthritis is associated with down-regulation of superoxide dismutase 2. Arthritis Rheum 2013, 65(2), 378–387. [Google Scholar] [CrossRef]
- Wang, Y; Zhao, X; Lotz, M; Terkeltaub, R; Liu-Bryan, R. Mitochondrial Biogenesis Is Impaired in Osteoarthritis Chondrocytes but Reversible via Peroxisome Proliferator-Activated Receptor γ Coactivator 1α. Arthritis Rheumatol 2015, 67(8), 2141–2153. [Google Scholar] [CrossRef]
- Liu, D; Cai, ZJ; Yang, YT; Lu, WH; Pan, LY; Xiao, WF; et al. Mitochondrial quality control in cartilage damage and osteoarthritis: new insights and potential therapeutic targets. Osteoarthritis Cartilage 2022, 30(3), 395–405. [Google Scholar] [CrossRef]
- Carrasco, C; Naziroǧlu, M; Rodríguez, AB; Pariente, JA. Neuropathic Pain: Delving into the Oxidative Origin and the Possible Implication of Transient Receptor Potential Channels. Front Physiol. 2018, 9, 95. [Google Scholar] [CrossRef]
- Silva Santos Ribeiro, P; Willemen, HLDM; Eijkelkamp, N. Mitochondria and sensory processing in inflammatory and neuropathic pain. Front Pain Res (Lausanne) 2022, 3, 1013577. [Google Scholar] [CrossRef]
- Sibille, KT; Langaee, T; Burkley, B; Gong, Y; Glover, TL; King, C; et al. Chronic pain, perceived stress, and cellular aging: an exploratory study. Mol Pain 2012, 8, 12. [Google Scholar] [CrossRef]
- Mosquera, A; Rego-Pérez, I; Blanco, FJ; Fernández, JL. Leukocyte Telomere Length in Patients with Radiographic Knee Osteoarthritis. Environ Mol Mutagen 2019, 60(3), 298–301. [Google Scholar] [CrossRef]
- Vaiserman, A; Krasnienkov, D. Telomere Length as a Marker of Biological Age: State-of-the-Art, Open Issues, and Future Perspectives. Front Genet. 2020, 11, 630186. [Google Scholar] [CrossRef]
- Wesseling, J; Dekker, J; van den Berg, WB; Bierma-Zeinstra, SM; Boers, M; Cats, HA; et al. CHECK (Cohort Hip and Cohort Knee): similarities and differences with the Osteoarthritis Initiative. Ann Rheum Dis. 2009, 68(9), 1413–1419. [Google Scholar] [CrossRef] [PubMed]
- Yerges-Armstrong, LM; Yau, MS; Liu, Y; Krishnan, S; Renner, JB; Eaton, CB; et al. Association analysis of BMD-associated SNPs with knee osteoarthritis. J Bone Miner Res. 2014, 29(6), 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Chang, CC; Chow, CC; Tellier, LC; Vattikuti, S; Purcell, SM; Lee, JJ. Second-generation PLINK: rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Tamayo, M; Mosquera, A; Ignacio Rego, J; Luis Fernandez-Sueiro, J; Blanco, FJ; Luis Fernandez, J. Differing patterns of peripheral blood leukocyte telomere length in rheumatologic diseases. Mutation Research-Fundamental and Molecular Mechanisms of Mutagenesis 2010, 683(1-2), 68–73. [Google Scholar] [CrossRef]
- Zhan, X; Hu, Y; Li, B; Abecasis, GR; Liu, DJ. RVTESTS: an efficient and comprehensive tool for rare variant association analysis using sequence data. Bioinformatics 2016, 32(9), 1423–1426. [Google Scholar] [CrossRef]
- Tachmazidou, I; Hatzikotoulas, K; Southam, L; Esparza-Gordillo, J; Haberland, V; Zheng, J; et al. Identification of new therapeutic targets for osteoarthritis through genome-wide analyses of UK Biobank data. Nat Genet. 2019, 51(2), 230–236. [Google Scholar] [CrossRef]
- Boer, CG; Hatzikotoulas, K; Southam, L; Stefánsdóttir, L; Zhang, Y; Coutinho de Almeida, R; et al. Deciphering osteoarthritis genetics across 826,690 individuals from 9 populations. Cell 2021, 184(18), 4784–4818.e17. [Google Scholar] [CrossRef] [PubMed]
- Hatzikotoulas, K; Southam, L; Stefansdottir, L; Boer, CG; McDonald, ML; Pett, JP; et al. Translational genomics of osteoarthritis in 1,962,069 individuals. Nature 2025, 641(8065), 1217–1224. [Google Scholar] [CrossRef]
- Magne, D; Palmer, G; Barton, JL; Mézin, F; Talabot-Ayer, D; Bas, S; et al. The new IL-1 family member IL-1F8 stimulates production of inflammatory mediators by synovial fibroblasts and articular chondrocytes. Arthritis Res Ther. 2006, 8(3), R80. [Google Scholar] [CrossRef]
- Queen, D; Ediriweera, C; Liu, L. Function and Regulation of IL-36 Signaling in Inflammatory Diseases and Cancer Development. Front Cell Dev Biol. 2019, 7, 317. [Google Scholar] [CrossRef]
- Chen, WJ; Yu, X; Yuan, XR; Chen, BJ; Cai, N; Zeng, S; et al. The Role of IL-36 in the Pathophysiological Processes of Autoimmune Diseases. Front Pharmacol. 2021, 12, 727956. [Google Scholar] [CrossRef] [PubMed]
- Li, Q; Liu, S; Li, L; Ji, X; Wang, M; Zhou, J. Spinal IL-36γ/IL-36R participates in the maintenance of chronic inflammatory pain through astroglial JNK pathway. Glia 2019, 67(3), 438–451. [Google Scholar] [CrossRef]
- Wang, P; Yang, W; Guo, H; Dong, HP; Guo, YY; Gan, H; et al. IL-36γ and IL-36Ra Reciprocally Regulate NSCLC Progression by Modulating GSH Homeostasis and Oxidative Stress-Induced Cell Death. Adv Sci (Weinh) 2021, 8(19), e2101501. [Google Scholar] [CrossRef] [PubMed]
- Girirajan, S; Patel, N; Slager, RE; Tokarz, ME; Bucan, M; Wiley, JL; et al. How much is too much? Phenotypic consequences of Rai1 overexpression in mice. Eur J Hum Genet. 2008, 16(8), 941–954. [Google Scholar] [CrossRef]
- Elsea, SH; Girirajan, S. Smith-Magenis syndrome. Eur J Hum Genet 2008, 16(4), 412–421. [Google Scholar] [CrossRef] [PubMed]
- Mullin, BH; Zhu, K; Brown, SJ; Mullin, S; Dudbridge, F; Pavlos, NJ; et al. Leveraging osteoclast genetic regulatory data to identify genes with a role in osteoarthritis. Genetics 2023, 225(2), iyad150. [Google Scholar] [CrossRef]
- Turco, EM; Giovenale, AMG; Sireno, L; Mazzoni, M; Cammareri, A; Marchioretti, C; et al. Retinoic acid-induced 1 gene haploinsufficiency alters lipid metabolism and causes autophagy defects in Smith-Magenis syndrome. Cell Death Dis. 2022, 13(11), 981. [Google Scholar] [CrossRef]
- Leidenroth, A; Hewitt, JE. A family history of DUX4: phylogenetic analysis of DUXA, B, C and Duxbl reveals the ancestral DUX gene. BMC Evol Biol. 2010, 10, 364. [Google Scholar] [CrossRef]
- Tihaya, MS; Mul, K; Balog, J; de Greef, JC; Tapscott, SJ; Tawil, R; et al. Facioscapulohumeral muscular dystrophy: the road to targeted therapies. Nat Rev Neurol. 2023, 19(2), 91–108. [Google Scholar] [CrossRef]
- Karpukhina, A; Galkin, I; Ma, Y; Dib, C; Zinovkin, R; Pletjushkina, O; et al. Analysis of genes regulated by DUX4 via oxidative stress reveals potential therapeutic targets for treatment of facioscapulohumeral dystrophy. Redox Biol. 2021, 43, 102008. [Google Scholar] [CrossRef] [PubMed]
- Shao, J; Xu, Y; Li, H; Chen, L; Wang, W; Shen, D; et al. LMCD1 antisense RNA 1 (LMCD1-AS1) potentiates thyroid cancer cell growth and stemness via a positive feedback loop of LMCD1-AS1/miR-1287-5p/GLI2. Ann Transl Med. 2020, 8(22), 1508. [Google Scholar] [CrossRef] [PubMed]
- Liang, M; Li, Y; Chen, C. LMCD1-AS1 Facilitates Cell Proliferation and EMT by Sponging miR-873-3p in Cervical Cancer. Crit Rev Eukaryot Gene Expr. 2023, 33(2), 13–25. [Google Scholar] [CrossRef]
- Fernandez-Moreno, M; Soto-Hermida, A; Vazquez-Mosquera, ME; Cortes-Pereira, E; Relano, S; Hermida-Gomez, T; et al. Mitochondrial DNA haplogroups influence the risk of incident knee osteoarthritis in OAI and CHECK cohorts. A meta-analysis and functional study. Ann Rheum Dis. 2017, 76(6), 1114–1122. [Google Scholar] [CrossRef]
- Zhao, Z; Li, Y; Wang, M; Jin, Y; Liao, W; Fang, J. Mitochondrial DNA haplogroups participate in osteoarthritis: current evidence based on a meta-analysis. Clin Rheumatol. 2020, 39(4), 1027–1037. [Google Scholar] [CrossRef]
- Tan, S; Sun, Y; Li, S; Wu, H; Ding, Y. The impact of mitochondrial dysfunction on osteoarthritis cartilage: current insights and emerging mitochondria-targeted therapies. Bone Res. 2025, 13(1), 77. [Google Scholar] [CrossRef]
- Willemen, HLDM; Santos Ribeiro, PS; Broeks, M; Meijer, N; Versteeg, S; Tiggeler, A; et al. Inflammation-induced mitochondrial and metabolic disturbances in sensory neurons control the switch from acute to chronic pain. Cell Rep Med. 2023, 4(11), 101265. [Google Scholar] [CrossRef]
- Guillén, R; Otero, F; Mosquera, A; Vázquez-Mosquera, M; Rego-Pérez, I; Blanco, FJ; et al. Association of accelerated dynamics of telomere sequence loss in peripheral blood leukocytes with incident knee osteoarthritis in Osteoarthritis Initiative cohort. Sci Rep. 2021, 11(1), 15914. [Google Scholar] [CrossRef]
- Sibille, KT; Chen, H; Bartley, EJ; Riley, J; Glover, TL; King, CD; et al. Accelerated aging in adults with knee osteoarthritis pain: consideration for frequency, intensity, time, and total pain sites. Pain Rep. 2017, 2(3), e591. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Manhattan plot of the genome-wide association study (GWAS) for rapid pain progression in knee osteoarthritis. Association results for 7,762,204 imputed SNPs tested in OAI participants of European ancestry under an additive genetic model, adjusting for age, sex, BMI and the first three ancestry principal components. Each point represents a SNP (y-axis: −log10(P)) plotted by chromosomal position (x-axis; alternating colors by chromosome). The red dashed line indicates the conventional genome-wide significance threshold (P = 5 × 10−8), and the blue dashed line indicates the suggestive threshold (P = 5 × 10−6). No variant reached genome-wide significance; six loci exceeded the suggestive threshold and were carried forward as hypothesis-generating candidates (Table 2).
Figure 1.
Manhattan plot of the genome-wide association study (GWAS) for rapid pain progression in knee osteoarthritis. Association results for 7,762,204 imputed SNPs tested in OAI participants of European ancestry under an additive genetic model, adjusting for age, sex, BMI and the first three ancestry principal components. Each point represents a SNP (y-axis: −log10(P)) plotted by chromosomal position (x-axis; alternating colors by chromosome). The red dashed line indicates the conventional genome-wide significance threshold (P = 5 × 10−8), and the blue dashed line indicates the suggestive threshold (P = 5 × 10−6). No variant reached genome-wide significance; six loci exceeded the suggestive threshold and were carried forward as hypothesis-generating candidates (Table 2).
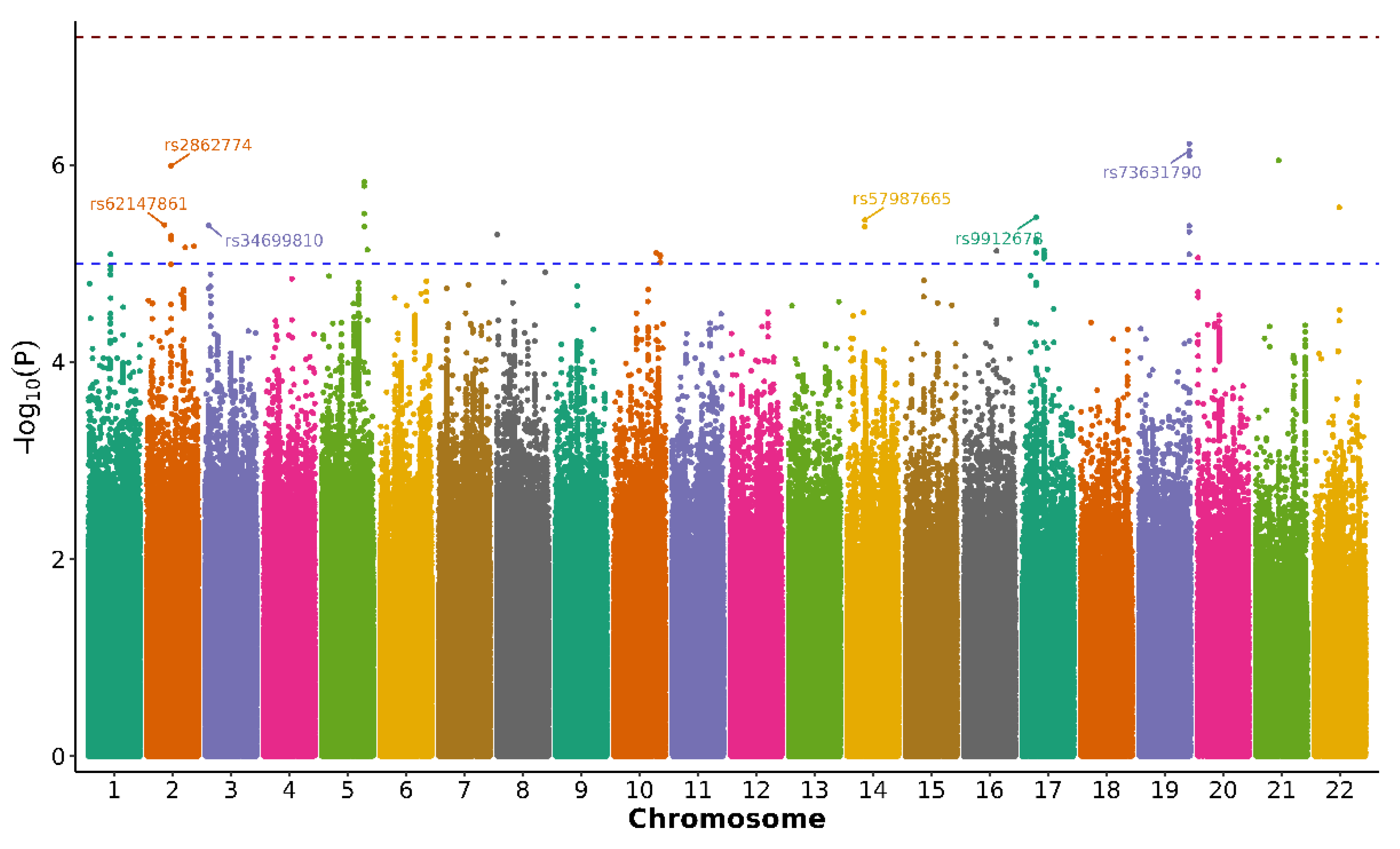
Figure 2.
Hypothesized pathways linking oxidative stress to rapid pain progression in knee osteoarthritis in a hypothesis-generating analysis. Conceptual framework combining the three biological layers evaluated in this study—nuclear genetic variation (exploratory GWAS signals), mtDNA haplogroups, and leukocyte telomere length as a marker of biological ageing—converging on oxidative stress and mitochondrial dysfunction (increased ROS), inflammation, and cellular ageing. These processes are proposed to promote pain sensitisation and joint damage, ultimately contributing to rapid pain progression in knee OA. Image generated using artificial intelligence.
Figure 2.
Hypothesized pathways linking oxidative stress to rapid pain progression in knee osteoarthritis in a hypothesis-generating analysis. Conceptual framework combining the three biological layers evaluated in this study—nuclear genetic variation (exploratory GWAS signals), mtDNA haplogroups, and leukocyte telomere length as a marker of biological ageing—converging on oxidative stress and mitochondrial dysfunction (increased ROS), inflammation, and cellular ageing. These processes are proposed to promote pain sensitisation and joint damage, ultimately contributing to rapid pain progression in knee OA. Image generated using artificial intelligence.
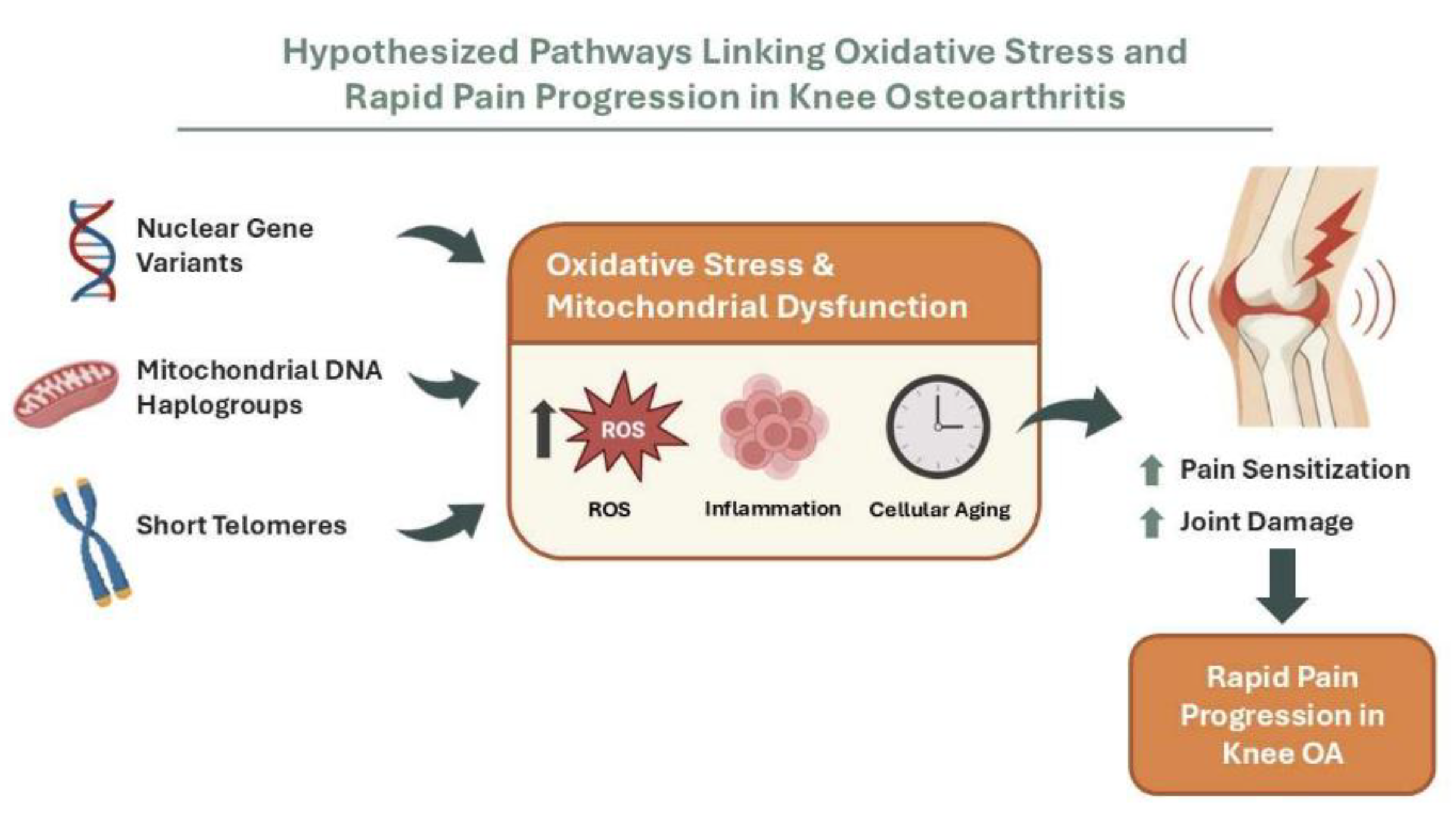

Table 1.
Descriptive analysis of the OAI participants based on their pain progression status.
| Rapid pain progressors (N=1,260) |
Non-rapid pain progressors (N=2,135) |
p-value | |
|---|---|---|---|
| Age at baseline (years) | 61.82±9.05 | 61.33±9.32 | 0.116* |
| Sex: | <0.001# | ||
| Male | 494 (39.2) | 995 (46.6) | |
| Female | 766 (60.8) | 1140 (53.4) | |
| BMI (Kg/m2) | 29.43±4.90 | 27.25±4.31 | <0.001* |
Values are mean±standard deviation or number of patients with percentage in parentheses; (*) Mann-Whitney U test; (#) chi-square test; BMI: body mass index; significant p-values are in bold.
Table 2.
Suggestive associations with rapid pain progression in the discovery GWAS*.
| rsID | Chr. | Ref. | Alt. | Nearest gene | OR (95% CI) | p-value |
|---|---|---|---|---|---|---|
| rs73631790 | 19 | G | A | DUXA | 0.605 (0.496 – 0.738) | 7.195 x 10-7 |
| rs9912678 | 17 | A | T | RAI1 | 1.787 (1.400 – 2.284) | 3.374 x 10-6 |
| rs57987665 | 14 | A | G | LOC105378178 | 1.332 (1.180 – 1.503) | 3.618 x 10-6 |
| rs34699810 | 3 | A | G | LMCD1-AS1 | 1.680 (1.347 – 2.095) | 4.107 x 10-6 |
| rs2862774 | 2 | A | C | IL36B | 0.687 (0.591 – 0.798) | 1.027 x 10-6 |
| rs62147861 | 2 | C | A | LOC124907827 | 1.530 (1.277 – 1.833) | 4.059 x 10-6 |
(*) Adjusted for age, sex, body mass index and first 3 principal components; Chr.: chromosome; Ref.: reference; Alt.: alternative allele; OR: odds ratio; CI: confidence interval.
Table 3.
Frequencies and odds ratios (ORs) of mitochondrial DNA (mtDNA) haplogroups in rapid pain progressors.
Table 3.
Frequencies and odds ratios (ORs) of mitochondrial DNA (mtDNA) haplogroups in rapid pain progressors.
| Haplogroups | Rapid pain progressors | Non-rapid pain progressors | Total | 95% CI | |||
|---|---|---|---|---|---|---|---|
| (N=1,239) | (N=2,118) | (N=3,357) | OR | Lower CI | Upper CI | p-value | |
| H | 538 (43.4) | 835 (39.4) | 1373 (40.9) | 1.179 | 1.023 | 1.359 | 0.023# |
| Uk | 287 (23.2) | 526 (24.8) | 813 (24.2) | 0.912 | 0.774 | 1.076 | 0.275 |
| T | 125 (10.1) | 219 (10.3) | 344 (10.2) | 0.973 | 0.772 | 1.227 | 0.817 |
| J | 119 (9.6) | 184 (8.7) | 303 (9.0) | 1.117 | 0.877 | 1.423 | 0.371 |
| Others* | 170 (13.7) | 354 (16.7) | 524 (15.6) | 0.792 | 0.650 | 0.966 | 0.021# |
Values are number of patients with percentage in parentheses: (*) the group “others” includes mtDNA lineages with a population frequency below 5%; (#) Chi-square test; statistical significance declared at P ≤ 0.05 (in bold); OR: odds ratio; CI: confidence interval.
Table 4.
Multivariable logistic regression model to predict the influence of mtDNA haplogroup H on the risk of rapid pain progression in participants belonging to the OAI.
Table 4.
Multivariable logistic regression model to predict the influence of mtDNA haplogroup H on the risk of rapid pain progression in participants belonging to the OAI.
| Variable | B | Adjusted OR | 95% CI | p-value |
|---|---|---|---|---|
| Sex (female) | 0.402 | 1.494 | 1.289 – 1.732 | <0.001 |
| Age | 0.009 | 1.009 | 1.001 – 1.017 | 0.031 |
| BMI | 0.107 | 1.113 | 1.095 – 1.131 | <0.001 |
| Haplogroup H | 0.159 | 1.172 | 1.012 – 1.357 | 0.034 |
B: regression coefficient; OR: odds ratio; BMI: body mass index; OAI: Osteoarthritis Initiative; CI: confidence interval; statistical significance declared at p<0.05 (in bold).
Table 5.
Multivariable logistic regression model to predict the influence of PBL telomere length on the risk of rapid pain progression in a subset of 301 participants belonging to the OAI.
Table 5.
Multivariable logistic regression model to predict the influence of PBL telomere length on the risk of rapid pain progression in a subset of 301 participants belonging to the OAI.
| Variable | B | Adjusted OR | 95% CI | p-value |
|---|---|---|---|---|
| Sex (female) | 0.140 | 1.150 | 0.688 – 1.923 | 0.593 |
| Age | 0.012 | 1.012 | 0.982 – 1.042 | 0.449 |
| BMI | 0.132 | 1.141 | 1.076 – 1.211 | <0.001 |
| T/S ratio* | -1.347 | 0.260 | 0.098 – 0.693 | 0.007 |
B: regression coefficient; OR: odds ratio; PBL: peripheral blood leukocyte; BMI: body mass index; OAI: Osteoarthritis Initiative; CI: confidence interval; (*) average ratio of telomere repeat copy number to a single-copy reference gene (36B4) in each sample; statistical significance declared at p<0.05 (in bold).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



