Submitted:
17 January 2026
Posted:
19 January 2026
Read the latest preprint version here
Preprints on COVID-19 and SARS-CoV-2
Abstract
COVID-19, caused by the new type of coronavirus SARS-CoV-2, has put an unprecedented impact on health, economy and social areas around the globe. It created an urgent global need for rapid diagnostics, effective therapeutics, and scalable vaccine manufacturing. The biomanufacturing industry played a central role in meeting this challenge by accelerating the development, production, and distribution of SARS‑CoV‑2 diagnostic assays and vaccines. This review provides an integrated overview of SARS‑CoV‑2 biology, clinical manifestations, transmission mechanisms, and major viral variants, followed by a detailed examination of diagnostic technologies. We further highlight the transformative impact of mRNA vaccine technologies, emphasizing advances in lipid nanoparticle formulation, large‑scale manufacturing, and regulatory‑aligned production strategies. The review also discusses the biomanufacturing sector’s rapid mobilization to overcome supply‑chain constraints, workforce shortages, and unprecedented global demand. Collectively, this work underscores how scientific innovation, industrial agility, and cross‑sector collaboration enabled the rapid deployment of diagnostics and vaccines that were essential to controlling the COVID‑19 pandemic.
Keywords:
COVID-19
; SARS-CoV-2
; mRNA vaccines
; nanomedicine
; biomanufacturing industry
1. Introduction
The coronavirus disease 19 (COVID-19) is a communicable acute pathogenic viral infectious disease caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which emerged in Wuhan, Hubei province, China in December 2019. In a span of small time it spread around the world causing considerable morbidity and mortality [1,2]. Until January 2026, around 780 million people have been infected with SARS-CoV-2 and over 7 million deaths have been reported worldwide (World Health Organization, 2025, https://covid19.who.int/, accessed on 01-05-2026). SARS-CoV-2 infection triggers the immune system which results in uncontrollable inflammatory response causing tissue damage [3] This disease causes severe atypical respiratory ailments which is its primary target but can affect other organs like GI tract and CNS causing severe complications [4]. FDA has approved several antiviral drugs for COVID-19 including remdesivir (Veklury) [5], molnupiravir (Lagevrio) [6] and nirmatrelvir/ritonavir (Paxlovid) [7], which have been authorized to reduce disease severity and hospitalization risk in patients with SARS-CoV-2 infection.
2. Structure
SARS-CoV-2 particles are round and surrounded by a membrane, measuring about 80–120 nm in diameter. They have many club-shaped S proteins on their surface, giving them a distinctive appearance similar to a solar corona, which is why they’re called coronaviruses (CoVs) [8].
SARS-COV-2 is a single stranded positive-sense RNA virus containing a large genome ranging from 29 kilobases and consists of 9,680 amino acids encoding 27 proteins including four structural proteins and 16 functional proteins such as RNA dependent RNA polymerase (RdRP) and helicase [9,10]. RNA is associated with nucleocapsid protein (N) and is surrounded by envelope containing membrane protein (M), spike protein (S) and envelope protein (E) [11]. S proteins are involved in the infection of host cell by binding to its receptor, angiotensin converting enzyme 2 (ACE2) mostly expressed in human lung alveolar epithelial cells [12]. For this reason, the S protein and ACE2 continue to be the primary target for various anti-coronavirus treatments, such as neutralizing monoclonal antibodies (mAbs), vaccines, and other inhibitors to block membrane-virus interaction [13]. Nucleocapsid protein (NP) allows the virus to hijack host cell machinery for its own replication and transcription. The M protein is crucial in coordinating the assembly and organization of the viral envelope by engaging with different structural proteins while E protein is involved in virus assembly, membrane permeability of the host cell and virus host interaction [14,15,16]. The N protein functions by interacting with the positive RNA strand of the viral genome, forming a helical ribonucleocapsid complex. It also engages with other structural membrane proteins during virion assembly [17].
The SARS-CoV-2 S protein is cleaved by host proteases into two parts: the receptor-binding fragment (S1) and the fusion fragment (S2) during biogenesis or virus assembly. The S1 subunit contains various domains, including the N-terminal domain (NTD), receptor-binding domain (RBD) with a receptor-binding motif (RBM), and C-terminal domains (CTDs). Several potential antibodies against SARS-CoV-2 have been identified which target the NTD, S2 or RBD [13].
3. Variants
Viruses exhibit instability because of mutations that occur from errors during their replication within host cells. [18]. While the majority of these mutations are biologically inconsequential, certain alterations can significantly modify viral characteristics, including transmissibility, pathogenicity, and immune evasion capacity [19]. Mutations in SARS-CoV-2 lead to the evolution of a variant which is different from other variants. These mutations have mostly been found in receptor binding domain (RBD) and the N-terminal domain (NTD) of S protein. Some of these mutations may increase the transmissibility of virus into host, evading host immune responses, causing altered symptoms and make available treatments and vaccinations less effective. Those variants are classified as Variants of Concern (VOC) by health organizations like WHO and CDC [20,21]. The classification of variants help in monitoring and responding to the pandemic. The main categories include:
3.1. Variants of Interest (VOI)
These are variants with specific genetic markers predicted to increase transmissibility, disease severity, or decrease vaccine/treatment efficacy, and disrupt diagnostic testing [22]. These VOIs likely require increased surveillance and further research [22]. As of late 2024, the CDC’s only recognized VOI is the Omicron sublineage JN.1 [23].
3.2. Variants of Concern (VOC)
The VOCs meet all the criteria for VOIs but are further classified as variants with evidence of increased transmissibility, severity, decreased vaccine/treatment efficacy, and diagnostic failures (1). These VOCs require reporting to public health agencies and increased research to detect public health risks [22].
3.3. Variants of High Consequence (VOHC)
Variants of High Consequence (VOHC) are those which possess all the characteristics of VOCs and for which there is clear evidence that prevention measures or medical countermeasures (including vaccines, treatments, or diagnostics) have significantly reduced effectiveness compared to previously circulating variants [24].
3.4. Major Variants of Coronavirus
3.4.1. Alpha (B.1.1.7)
First detected in the United Kingdom in late 2020, the Alpha variant carried mutations such as N501Y and P681H that and was characterized by increased transmissibility compared to the original virus. It was associated with higher risk of hospitalization and intensive care admission despite public health restrictions [25].
3.4.2. Beta (B.1.351)
This variant, first detected in South Africa, carried multiple mutations in the spike protein, notably K417N, E484K, and N501Y. It also showed increased transmissibility and a notable ability to evade the immune response generated by previous infection or vaccination. It was also linked to a higher risk of hospitalization [26].
3.4.3. Gamma (P.1)
First detected in Brazil, the Gamma variant shared some key mutations with the Beta variant besides D614G mutation in spike protein. It demonstrated increased transmissiblity and immune evasion capablities and was associated with a higher risk of hospitalization and intensive care admission [27].
3.4.4. Delta (B.1.617.2)
The Delta variant first identified in India. This variant has enhanced replication capacity compared to Alpha (B.1.1.7). It shows partial immune evasion, with reduced neutralization by vaccine-induced antibodies [28].
3.4.5. Omicron (B.1.1.529)
This variant, first detected in South Africa, carried 30 mutations in the spike protein, notably K417N, E484A, N501Y, D614G, P681H, among others. Besides, it carried mutations in RBD [20]. It uses cellular transmembrane serine protease 2 (TMPRSS2)-independent endosomal entry [29]. Omicron displayed marked genetic and phenotypic divergence from the original Wuhan strain, characterized by substantially increased transmissibility and a pronounced ability to evade both natural and vaccine-induced immunity, it rapidly displaced the Delta variant and achieved global dominance within weeks of its initial detection, triggering major waves of infection. Numerous studies documented a significant reduction in neutralizing activity of vaccine-elicited antibodies against these variants compared with the ancestral strain, contributing to increased breakthrough infections and driving the development of updated vaccine formulations [30].
4. Mechanism of Action
SARS-CoV-2 enters into body through the respiratory tract and infects epithelial cells of the trachea, bronchi, bronchioles and lungs [31]. Viral entry is initiated when the spike (S) glycoprotein binds to angiotensin converting enzyme 2 (ACE2), followed by proteolytic activation of the S protein by host proteases such as TMPRSS2 at the cell surface or cathepsins within endosomes [19,32]). This cleavage event triggers membrane fusion and release of the viral nucleocapsid into the cytoplasm. Once uncoated, the positive-sense single-stranded RNA genome functions directly as mRNA and is translated into the polyproteins pp1a and pp1ab. These polyproteins are cleaved by the viral proteases Mpro and PLpro to generate non-structural proteins that assemble into the replication–transcription complex (RTC) [33]. The RTC associates with double-membrane vesicles derived from the endoplasmic reticulum, where it mediates genomic replication and transcription of subgenomic RNAs encoding structural and accessory proteins [34].
The structural proteins S, M, and E are synthesized in the rough ER and transported to the ER–Golgi intermediate compartment (ERGIC). Meanwhile, the nucleocapsid (N) protein binds newly synthesized genomic RNA to form ribonucleoprotein complexes. Virion assembly occurs in the ERGIC, where budding into membranes enriched with S, M, and E proteins produces mature virions [35]. These virions are then transported in secretory vesicles through the Golgi apparatus and released via exocytosis, enabling infection of neighboring cells and dissemination throughout the respiratory tract [36].
5. Response of Biomanufacturing Industry to COVID-19 Pandemic
The biomanufacturing industry has played a transformative role in the global response to the COVID-19 pandemic, enabling the rapid development, scale-up, and distribution of essential medical countermeasures (Figure 1). This sector was instrumental in producing vast quantities of personal protective equipment (PPE), hand sanitizers, diagnostic assays, and most critically vaccines, all of which significantly reduced transmission and disease burden worldwide. According to global vaccination data sourced by the World Health Organization and compiled by Our World in Data, more than 13.64 billion COVID-19 vaccine doses have been administered globally, contributing to primary vaccination coverage in approximately 67% of the world’s population (World Health Organization. COVID-19 Vaccination Dashboard. WHO; 2023. Available at: https://data.who.int/dashboards/covid19/vaccines). WHO’s COVID-19 Vaccination Insights Report further documents the unprecedented scale of vaccine deployment, highlighting the coordinated efforts of manufacturers, governments, and international organizations to deliver billions of doses across all regions. These achievements were made possible through rapid biotechnological innovation, including mRNA platforms, viral-vector systems, and high-throughput diagnostic technologies, combined with cross-sector collaboration that accelerated regulatory pathways, expanded production capacity, and strengthened supply chains. Collectively, the biomanufacturing industry demonstrated an extraordinary ability to mobilize resources and expertise, ensuring that critical medical supplies reached global populations during a time of urgent need. We will expand these topics in the sections that follow.
5.1. SARS-CoV-2 Diagnostic Approaches
Ever since the COVID-19 pandemic, it has become immensely important to develop methods to accurately diagnose to effectively manage the disease. The biomanufacturing industry played a crucial role in the development of diagnostic tests for SARS-CoV-2. A diagnostic test indicates the presence of an infecting agent in the body which is causing disease. Since symptoms of COVID-19 are nonspecific, syndromic testing and CT scans may not be suitable for accurate diagnoses of the disease [37]. Molecular techniques that target genome or proteome of SARS-CoV-2 could be best methods for its detection. We discuss some methods of SARS-CoV-2 detection including commonly used polymerase chain reaction (PCR), and antigen rapid detection test [38,39].
5.1.1. Polymerase Chain Reaction (PCR)
Reverse-Transcription PCR or RT-qPCR [40] represents highly sensitive and specific methods for detecting viral genetic material. This testing approach involves conversion of viral RNA obtained from diverse specimens such as ocular secretions, saliva, sputum, bronchoalveolar lavage (BAL), blood, and fecal material, upper respiratory system samples such as oropharyngeal swabs (OPS) and nasopharyngeal swabs(NPSs) [41,42] complimentary DNA (cDNA) strands through targeted amplification of specific cDNA target sequences using enzyme reverse transcriptase. The cDNA is subsequently amplified in individual reaction tube by virus-specific primers and the PCR product is identified using various analytical techniques [43]. Notably, primers focusing on E gene, RNA dependent RNA Polymerase gene, ORF1 and N genes have been successfully developed and utilized [41,44]. The qPCR process mirrors standard PCR, cycling through denaturation, annealing, and extension, with annealing temperature carefully optimized for each primer set to ensure efficient amplification of SARS-CoV-2 targets. Across roughly 35–45 cycles, fluorescence increases as DNA doubles each round, producing a characteristic sigmoidal curve with baseline, exponential, and plateau phases. The cycle threshold (Ct), the point where fluorescence first rises above background, is used to determine whether a sample is positive and serves as the quantitative reference point in qPCR assays [42]. RT-qPCR detects amplified DNA in real time using either nonspecific dyes like SYBR Green or highly specific probe-based systems such as TaqMan. In TaqMan assays, fluorescence is released only when the probe is cleaved during amplification, making the method especially sensitive and specific. Automated instruments cycle the reaction up to ~40 times, enabling rapid detection, high throughput, and the option to multiplex multiple targets in a single reaction [42].
RT-qPCR test is considered to be gold standard method [45] as it is more accurate and reliable test for diagnosing COVID-19 disease. However, this method is not free of limitations as it requires expensive equipment and skilled technicians to perform tests in laboratory setting and hence is costly and has long wait time. RT-qPCR may not be the best option in emergency settings to detect SARS-CoV-2 [46]. The quality of specimen and the stage of disease my influence the testing accuracy of RT-PCR.
Accurate designing of primers that target conserved regions of viral genomes is essential for minimizing false-negative results and reducing the need for extensive standardization across different PCR protocols. Building on this principle, researchers applied evolutionary algorithms (EAs), an AI-based optimization approach, together with genomic data from the Global Initiative on Sharing Avian Influenza Data (GISAID) to identify sub-sequences suitable for developing RT-qPCR primers capable of detecting SARS-CoV-2 and its VOCs in human samples. The method focuses on uncovering 21-bp sequences unique to a specific viral strain, ranking these candidates according to their suitability as primers, and evaluating how effectively they discriminate the target virus from other circulating lineages. Importantly, primers generated through this strategy demonstrated improved sensitivity, identifying more positive cases than conventional primer sets [47].
5.1.2. Antigen Rapid Diagnostic Test
Antigen rapid diagnostic tests (Ag-RDTs) detect active SARS-CoV-2 infection by identifying viral proteins, typically nucleocapsid or spike antigens, in nasal swab samples using monoclonal antibodies embedded in a lateral-flow immunoassay. As the sample migrates along the test strip, viral antigens, if present, bind to antibodies conjugated to visible labels such as gold nanoparticles and form immune complexes that generate a visible signal within minutes. Although less sensitive than RT-qPCR, Ag-RDTs provide fast, low-cost, and easy-to-use detection, making them valuable tools for large-scale public health screening [48,49,50]. Their performance improves in individuals with higher viral loads and in symptomatic cases, where sensitivity increases substantially [51,52,53]. Because of their speed, simplicity, and scalability, antigen rapid tests have become widely used for timely identification of infectious individuals and for population-level surveillance during the COVID-19 pandemic.
Cochrane, an independent global organization that synthesizes high-quality systemic reviews following rigorous standardized methods to minimize bias. It has published and continuously updated systematic reviews assessing rapid tests for the detection of SARS-CoV-2 (www.cochrane.org). These reviews have played a pivotal role in synthesizing evidence on the accuracy and utility of Ag-RDTs outside of highly controlled laboratory settings. Cochrane reviews consistently highlight that Ag-RDT sensitivity varies significantly depending on factors such as the timing of testing relative to symptom onset, viral load, and the specific test brand [54,55]. In symptomatic individuals tested within the first week of illness, when viral loads are highest, Ag-RDTs perform substantially better, with sensitivity often exceeding 80% and, in some cases, approaching 90% compared with RT-PCR early symptomatic accuracy [54]. This high early-phase sensitivity supports their use for rapid identification of infectious individuals. In contrast, sensitivity declines in asymptomatic people or those tested later in infection, when viral loads tend to be lower [55]. A review reported that sensitivity in asymptomatic cases could fall below 60% [56].
Conversely, the specificity of Ag-RDTs has consistently been found to be very high across various real-world settings, typically above 98% [57]. This high specificity means that false positive results are rare, which is crucial for preventing unnecessary isolation and anxiety. The implications of these findings are profound for public health strategies. A review emphasized that while Ag-RDTs may not detect all infections, particularly those with low viral loads, their ease of use, rapid turnaround time, and high specificity make them effective tools for frequent, widespread testing, especially in settings where rapid results are critical [58]. Furthermore, the performance of Ag-RDTs can vary between different manufacturers and test brands, highlighting the importance of evaluating specific products [54].
In parallel, molecular lateral flow assays (LFA) have been developed to detect SARS-CoV-2 nucleic acids rather than proteins, including assays targeting the conserved NIID_2019-nCoV_N gene region. These tests pair nucleic-acid amplification with a simple paper-based readout, enabling rapid, equipment-free detection with good sensitivity and specificity. By targeting a highly conserved N-gene sequence, such molecular LFAs offer a portable and reliable alternative to laboratory-based RT-qPCR, complementing antigen tests in point-of-care and resource-limited settings [59].
5.1.3. Serology Test
SARS-CoV-2 infection triggers the production of IgM, IgG, and IgA antibodies in host, each appearing at different stages and providing valuable information about the timing and progression of infection [60]. Serology tests detect past or recent SARS-CoV-2 infection by identifying these antibodies. These assays expose a blood sample to SARS-CoV-2 proteins such as the spike or nucleocapsid, and antibody binding is then measured using enzyme-linked, chemiluminescent, or lateral-flow platforms. Antibody kinetics play a key role in interpretation: IgM typically rises during the first week of infection, IgA appears around days 4–10, and IgG develops in the second week and persists longer [61]. Because antibodies take time to develop, serology is not used to diagnose acute infection, but it is valuable for identifying prior exposure, assessing population-level immunity, evaluating vaccine responses, and supporting diagnosis in cases with negative RT-PCR or asymptomatic infection [62,63]. Overall, serological assays complement molecular tests by providing insight into the host immune response and the epidemiology of COVID-19.
5.1.4. Reverse Transcription Loop-Mediated Isothermal Amplification (RT-LAMP)
RT-LAMP is garnering a lot of attention for point-of-care (POC) applications. It has been used to detect diseases including the SARS-CoV-2 virus. Between quick antigen testing and RT-qPCR, RT-LAMP stands in the center. While the sensitivity is closer to that of RT-qPCR, the simplicity is comparable to that of fast antigen tests, making it appropriate for point-of-care application. [64,65]. This technique provides an efficient platform for identifying viral infections by amplifying the target nucleic acid in a single step using the PCR method at a fixed temperature (typically 60ºC). Three phases make up the LAMP reaction: elongation, cycling amplification, and initiation. This method uses four to six specially created primers to identify different nucleic acid sequences. Furthermore, strand-displacement polymerases reduce the reaction time by up to 30 min and do not require initial template denaturation. With RT-LAMP, amplified results can be seen in color, fluorescence, or under a UV lamp [66]
5.1.5. Next-Generation Sequencing
Next- generation sequencing (NGS) is a powerful technique that enables the simultaneous sequencing of thousands to billions of DNA fragments belonging to different organisms in a very short period of time with reduced cost compared to traditional sequencing techniques like Sanger’s sequencing [67]. Unlike PCR, which requires prior knowledge of target sequences, NGS provides an unbiased approach to pathogen identification, forming the molecular basis for diagnostics, vaccine development, and tracing pathogen spread [62].
During the COVID-19 pandemic, NGS became indispensable for epidemiological and molecular surveillance with sequencing data archived in several databases including Global Initiative on Sharing All Influenza Data (GISAID, https://www.gisaid.org/) [68], the National Center for Biotechnology Information (NCBI, https://www.ncbi.nlm.nih.gov/sars-cov-2/) [69], Genome Warehouse (https://bigd.big.ac.cn/gwh/), National Bioinformatics Center (CNCB)/National Genomics Data Center (NGDC) database (https://bigd.big.ac.cn/ncov/) [70] and Virus Pathogen Resource (https://www.viprbrc.org/) [71]. GISAID database has collected over 16 million SARS-CoV-2 sequences determined with NGS and the number continues to increase. This has enabled the researchers to identify SARS-CoV-2, take up studies aimed at developing diagnostics and vaccines, detect mutations unlike PCR, identify new variants and perform genomic surveillance [72,73,74]. NGS revealed the virus’s origin and intermediate host, refuted speculation of artificial origins, identified variants of concern such as Alpha, Delta, and Omicron, and provided functional insights through RNA sequencing of ACE2 expression, 16S rDNA sequencing of gut microbiota changes, and single-cell RNA sequencing of immune responses [73,75,76,77]. It also enabled environmental surveillance via metagenomic sequencing of air samples [76]. Platforms such as Illumina, Ion Torrent, Pacific Biosciences, and Oxford Nanopore Technology have independently developed specific library preparation protocols and are being used for SARS-CoV-2 sequencing [78,79,80,81] with Illumina being the most widely used due to its high throughput and accuracy [78]. Thus, NGS served as the backbone of COVID-19 genomic surveillance, providing real-time insights into viral evolution and guiding global public health responses.
The enormous amount of data generated by sequencing is analyzed by bioinformatics tools to unravel structure of virus, management of COVID-19 and discovery of vaccines [82]. Bioinformatics analysis workflow generally includes quality profiling, removing host genome data, de novo assembly of reads into contigs, taxonomic classification and finally verification of viral genome [83,84,85].
Next-generation sequencing (NGS) has become an essential component of epidemiological and molecular surveillance during the COVID-19 pandemic, offering a level of genomic resolution that far exceeds traditional diagnostic assays. Unlike targeted RT-PCR, which detects predefined genomic regions, NGS enables unbiased whole-genome characterization of SARS-CoV-2, allowing simultaneous identification of known variants, detection of novel mutations, and real-time assessment of viral evolution [86]. This comprehensive genomic view has been critical for tracking the emergence, geographic spread, and diversification of variants of concern, supporting rapid public-health responses and guiding updates to diagnostic assays and vaccine formulations [87].
Beyond variant identification, NGS provides the depth needed to monitor mutation patterns across the entire viral genome, including changes that may affect transmissibility, virulence, immune escape, or diagnostic performance. Mutations in primer- or probe-binding regions can compromise RT-PCR sensitivity, and NGS-based surveillance enables early detection of such changes, ensuring that diagnostic assays remain robust as the virus evolves [88]. Both amplicon-based and capture-based NGS approaches have proven effective for high-resolution surveillance, with studies demonstrating their ability to detect mixed infections, low-frequency variants, and emerging lineages such as Omicron [89].
NGS also plays a central role in phylodynamic modeling, where genomic data are integrated with epidemiological information to reconstruct transmission chains, estimate evolutionary rates, and identify outbreak clusters. These analyses have been instrumental in understanding superspreading events, assessing the impact of public-health interventions, and characterizing the introduction and establishment of new variants in different regions [90].
5.1.6. Nanotechnology Approaches in SARS-CoV-2 Diagnosis
Nanotechnology has emerged as a powerful tool in the rapid and early diagnosis of COVID-19 using nanostructured materials leading to improved therapeutic efficacy [91]. It has also been utilized to create infection-safe masks, self-sterilizing personal protective equipment (PPE) for medical professionals, and efficient disinfectants and surface coatings [92] (Table 1). Detecting viral RNA at early stages of disease when its concentration is low is challenging when using RT-PCR. Moreover, it is time consuming, needs lab space and equipment, needs trained personnel to perform test and is costly. Comparatively, nanotechnology offers sensitive and portable diagnostic platforms to detect ultralow levels of viral RNA, antigens or antibodies by combining electrical and photonic properties of nanomaterials with biologic molecules [93,94]. We will explore the role of nanotechnology in COVID-19 diagnosis in the following lines.
Nanomaterials and their derivative technologies have been used to create novel techniques for the detection of SARS-CoV-2. Because of their variable size, structure, and geometry, as well as their electrical conductivity, magnetic properties, surface reactivity for functionalization with biomolecules, and quantum or fluorescence properties, these nanomaterials are perfect for diagnostic support. Nanomaterial-based sensors, particularly those with porous properties, have been created with the purpose of directly detecting SARS-CoV-2 infectious virions [95]. Nano biosensors are analytical devices that combine transducer with biological sensing element to detect the specific biological molecules with high accuracy and speed, capture the biological response and translate it into electrical signals [96]. They have emerged as a pivotal tool in the rapid and sensitive detection of COVID-19 because of their high surface area-to-volume ratio and functional capabilities, offering high sensitivity and specificity in identifying SARS-CoV-2. These sensors use nanomaterials such as gold nanoparticles (AuNPs), carbon nanotubes, and quantum dots (QDs) to enhance the detection capabilities of traditional biosensors. The transducer converts biological interaction into electrical, optical or another measurable signal. For instance, nanobiosensors can detect viral RNA, antigens and antibodies with remarkable precision, often providing results in a matter of minutes [97]. Portable and reasonably priced point-of-care diagnostic devices have been developed as a result of the combination of nanotechnology with optical and electrochemical detection techniques [98]. These developments have greatly increased the COVID-19 diagnostics’ speed and precision, enabling prompt intervention and slowing the virus’s transmission. [99]. AuNPs have been employed in quantitative paper-based electrochemical sensor chips to detect the N gene and in colorimetric assays to identify target areas on the RdRp and N viral genes. They also make it possible to use lateral-flow assays based on colloidal AuNPs to identify certain IgM and IgG antibodies that recognize the S and N proteins of SARS-CoV-2. Using sialic acid-coated AuNPs for S protein detection or monoclonal antibodies (mAbs) coated on AuNPs, AuNPs-based lateral flow experiments identify S or N viral proteins [95].
Functional plasmonic photothermal biosensors have been utilized to detect viral genes, including ORF1ab, RdRp, and E, utilizing gold nanoislands (AuNIs). Iron oxide nanoparticles (IONPs) are used in real-time optomagnetic biosensors to identify the RdRp gene, whereas graphene oxide (GO) nanosheets combined with anti-S protein antibodies are used in field-effect transistor (FET)-based biosensing devices to identify the S antigen in clinical samples. Using MnFe3O4 nanospheres and QD nanobeads, fluorescence-linked immunoassays identify anti-SARS-CoV-2-specific IgG antibodies from serum. By combining a nanoparticle-based lateral flow biosensor with a multiplex reverse transcription LAMP (mRT-LAMP), dye streptavidin-coated polymer nanoparticles (SA-DNPs) can detect ORF1ab and N genes [95].
In low-resource contexts in particular, these tests are helpful for point-of-care (PoC) clinics and are typically inexpensive. Their significance lies in their ability to monitor, identify, and manage viral outbreaks and community transmissions. This aids healthcare systems in promptly diagnosing and treating critically ill individuals [95].
6. Development of SARS-CoV-2 mRNA Vaccines
During COVID-19 pandemic, the mRNA vaccine technology played a pivotal role in the controlling and management of the life-threatening disease. However, the successful development of mRNA vaccines is a result of three decades of groundwork research [100,101]. The mRNA was first described by Brenner and colleagues in 1961 as an unstable molecule that copies information from DNA [102] In1969 the mRNA was successfully transcribed in vitro [103]. However, further research in this area was hindered due to the challenge of rapid degradation of mRNA molecule, inefficient in vivo transportation, and immunogenicity. After years of research, Krieg and colleagues synthesized mRNA in vitro using SP6 RNA polymerase [104,105]. Malone et. al. demonstrated that cationic lipid N- [1-(2,3- dioleyloxy) propyl]-N, N, N-trimethylammonium chloride encapsulated mRNA could be used to transfect eukaryotic cells, for the expression of mRNA in vitro [106]. In 1990, an external mRNA called in-vitro transcription (IVT) was expressed in host animal for the first time. Wolff and colleagues transferred mRNA into mouse quadriceps muscle which was successfully expressed, thus laying the groundwork for using mRNA as a therapeutic tool [107]. In 1992, Jirikowski GF et. al. successfully reversed diabetes insipidus in rats by injecting vasopressin mRNA from normal mice into them [108]. One of the challenges of using mRNA for therapeutic purposes was activation of humoral and cellular immune system against it (Pardi et al., 2018a) ([109]. Frederic Martinon made the first mRNA vaccine in 1992 when they developed mRNA-nanoparticle delivery system encoding the influenza-virus nucleoprotein [101]. In 1995, cancer research got a boost when Conry and colleagues developed mRNA vaccine encoding the carcinoembryonic antigen (CEA) in mice [110]. Katalin Karik’o and Drew Weissman found that IVT mRNA could bind to and activate specific Toll Like receptors (TLR3, TLR7 and TLR8) [111]. However, incorporation of modified uridine into IVT mRNA drastically reduced its immune recognition and greatly improved its stability and expression [112,113,114]. COVID-19 vaccines produced by BioNTech and Moderna used N1-methyl-pseudouridine (1mΨ) incorporated mRNA. On January 10, 2020, GISAID (Global Initiative on Sharing Avian Influenza Data) published the first whole-genome sequence of SARS-CoV-2 which enabled the production of diagnostic tests and development of vaccines against COVID-19 by biomanufacturing industry worldwide (Figure 2) [115,116]. Same day, Wrapp D et. al substituted two prolines with serine at aa986 and 987 to produce prefusion-stabilized SARS-CoV-2S (S-2P) for structure analysis [117]. This was followed by development of COVID-19 vaccines by many companies including Pfizer/BioNTech, Moderna, Johnson & Johnson/ Jenssen (Table 2). Within two months, Moderna prepared mRNA-1273 vaccine for clinical trial. Moving in parallel, Pfizer/BioNTech prepared BNT162b2 and J&J/Jenssen synthesized Ad26.COV.S vaccine. By November 2020 production and clinical evaluation of mRNA/LNP expressing SARS-CoV-2 S-2P (mRNA-1273) [118], BNT162b2 [119] with around 95% efficacy [119,120,121] were announced. A single dose of Ad26.COV.S vaccine showed efficacy of around 66.9% [121]. Of late, COVID-19 vaccines have been effective against new variants of SARS-CoV-2 [122,123,124,125].
6.1. Vaccine Candidates
The distinguishing characteristics of a vaccine candidate includes long-lived and protective antibody response and reactogenicity [126]. An ideal vaccine has high efficacy and immunogenicity as well as being safe and devoid of infections [127]. During the present covid pandemic several candidates including attenuated, killed, subunit, recombinant and gene-based vaccines have been tested for potential vaccines against COVID-19 [128,129]. The major drawback of inactivated virus vaccines may be that they revert to virulence, while live attenuated virus vaccines may require multiple booster doses considering their minimal cell-mediated immune response [130,131]. Recombinant and protein-subunit vaccines have demonstrated solid and reliable efficacy, though results vary across platforms and study designs. Novavax’s NVX-CoV2373 remains the most extensively evaluated, showing about 90% efficacy against symptomatic infection and 100% protection against moderate-to-severe disease in Phase III trials [132]. Other recombinant vaccines, including ZF2001, SKYCovione (GBP510), Medigen MVC-COV1901, Abdala, and Soberana 02, have reported 60% to over 90% efficacy against symptomatic disease, with stronger protection against severe outcomes [129,133]. Many elicit robust neutralizing antibody responses, especially when paired with potent adjuvants, and several show improved performance when used as boosters [134,135,136]. However, these vaccines also face limitations including slower recombinant protein production, requirement of strong adjuvants to achieve optimal immunogenicity [129], and lower peak neutralizing antibody levels than those induced by mRNA platforms [133]. Despite these drawbacks, the overall evidence indicates that recombinant vaccines provide meaningful protection, particularly against severe disease, and remain valuable components of global vaccination programs [137](CDC, 2025). mRNA vaccines, wherein a genetic fragment encodes viral peptide induces robust immune response for longer periods of time [138]. The SARS-CoV-2 spike protein is important for host recognition and invasion. It is, therefore considered to be the main immunogen for vaccine design [139]. It has been shown that majority of antibodies produced against SARS-CoV-2 specifically bind to the S protein and efficiently block the binding of virus with ACE2 [140,141]. Researchers have been able to precisely construct the antigen while avoiding difficult protein purification processes and a risky viral culture process thanks to technologies for mRNA and vector vaccines [142]
6.2. Design and Modifications to mRNA
mRNA vaccine sequence is synthesized in vitro and consists of nucleosides bonded together by the phosphodiester bonds (Wadhwa et al., 2020). It is single stranded and acts as a template for protein synthesis [143]. mRNA is comprised of 5’cap, 5’and 3’untranslated regions (UTR), the coding region (or open reading frame) and poly(A) tail [144]. Since mRNA is highly unstable and immunogenic, nucleoside modifications have reportedly been used to combat this as described earlier [114]. For this reason, mRNA1273 and BNT162b2 used m1Ψ modified mRNA for their vaccine design [145,146]. At the 5’end of mRNA vaccines, 7-methylguanosinetriphosphate (m7GpppNp), a modified nucleotide structure, called 5’cap is added to protect the mRNA from exonuclease cleavage and allows it to transport to cytoplasm [147,148]. 5’and 3’UTRs are present upstream and downstream of coding region respectively. They are untranslated regions of mRNA and are thought to be involved in regulating mRNA expression, recognition of mRNA by ribosomes, and post-transcriptional modification of mRNA [149]. Studies have shown that high GC content in 5’ UTR has been associated with the formation of secondary structures and hence lower rate of protein initiation efficiency and therefore lower expression [150,151]. Conversely, high GC content within the 3’ UTR may help synthetic mRNAs to evade targeted degradation by ARE-binding proteins [152]. High GC content within the coding region has been associated with higher protein expression [153]. Coding region, also called as open reading frame (ORF) is comprised of triplets of nucleotides called codons that code for amino acids. It begins with starting codon (AUG) and culminates in stop codon (UAA, UAG, UGA) and its sequence dictates the sequence of amino acids in protein [148]. ORF sequences have been modified by several approaches to improve translational efficiency [148,154].
At the 3′ end of mRNA, a polymeric chain in the range of 50-250 adenylate residues is found. It is called as the poly(A) tail. Poly(A) tail contributes to the stability, export and translation of mRNA and protects it from exonucleases [147,155]. The translation efficiency and stability of mRNA vaccines is balanced by optimizing length of poly(A) tail [155,156].
6.3. Delivery of mRNA Vaccines
Delivering mRNA efficiently into host cells is a challenge because of the negative charge and hydrophilic nature of mRNA, degradation due to endonucleases and phagocytosis [157]. Thus, the need of delivery system arose to create effective vaccines [158]. Recent breakthroughs in nanoparticle (NP) research have made it possible to deliver mRNA vaccines to host cells while providing stability and protection against host nucleases [159,160]. Currently, mRNA vaccines are encapsulated in positively charged lipid-nanoparticles (LNP) to efficiently allow it to pass through cell membranes and escape RNase degradation ([161,162]. Both mRNA-1273 and BNT162b2 use LNP for encapsulation. Upon intramuscular administration, COVID-19 mRNA-LNPs bind to host cell surface and are internalized into antigen-presenting cells and mRNA delivered into cytosol where it expresses S protein antigen using host cell machinery [163]. S protein is presented at the cell surface in its prefusion confirmation leading to the production of antibodies and T-cell induction via MHC [164,165]. LNPs used in mRNA vaccines is composed of atleast four components including distereophosphatidylcholine (DSPC), a polyethylene-glycol (PEG)-lipid, cholesterol and an ionizable cationic lipid [166]. DSPC improves permeability, cholesterol increases membrane integrity and rigidity, PEG-lipids adjust particle size and zeta potential while ionizable lipid component increases transfection efficiency ([167].
6.4. Advantages, Limitations and Caveats
Historically, traditional vaccines utilized the innocuous carrier molecules of a weakened or inactivated pathogens or the entire organism to stimulate an immune response in the form of antibodies. Notable pioneers, including scientists like Plett and Jenner, employed cowpox or horsepox virus to develop vaccines against smallpox [168,169]. These vaccines have successfully been used to eradicate many infectious diseases [170]. However, there are number of limitations with this platform including failure to work against some viruses like HIV [170], incomplete viral inactivation, requirement of multiple doses to attain robust and enduring immunity, risk of infection, time consuming and requirement of large quantities of cell culture to grow virus and requirement of biosafety level 3 production facilities [170,171] In contrast, mRNA vaccines employ synthetic mRNA that encodes a viral spike protein to prompt cells to produce a viral protein, which allows for quick design and production. Moreover, LNPs protect the mRNA and ensure efficient delivery into cells, thereby enhancing both stability and efficacy. These distinctions underscore the innovative approach of mRNA vaccines in rapidly addressing emerging infectious diseases [172]. mRNA vaccines have several advantages including rapid clinical development potential, low-cost production at large-scale, least risk of integration into host genome, ability to target intracellular proteins, inducing effective humoral and cellular immune responses, leading to higher protection rates than conventional vaccines, as shown during the COVID-19 pandemic [173,174]. The effectiveness of inactivated vaccines is around 60%, adenovirus-vectored vaccines at 65%, and mRNA vaccines at 90%. These vaccines consistently provide protection against asymptomatic SARS-CoV-2 infection, symptomatic COVID-19, COVID-19-related hospitalization, severe or critical hospitalization, and death [175].
Although mRNA vaccines have demonstrated strong protective efficacy, several limitations have been reported. Myocarditis occurs more frequently in younger males, particularly after the second dose of the mRNA-1273 vaccine [176]. Cases of thrombocytopenia have also been documented, along with symptoms such as severe headache, dizziness, visual disturbances, fever, shortness of breath, arrhythmia, pericarditis, cardiogenic shock, and anaphylactic reactions [175,177]. mRNA vaccines also present important caveats. They require ultra-cold storage, which limits accessibility in low-resource settings. Their reactogenicity is generally higher than that of some other platforms, with common short-term effects including fatigue, and injection-site pain [178].
Despite these concerns, mRNA vaccines offer several advantages over conventional platforms. mRNA can encode a wide range of proteins, enabling rapid vaccine development through simple sequence modification [20,21]. Production and purification processes are largely uniform across different mRNA vaccines, facilitating standardization and scalability [20,22]. In vitro transcription further streamlines manufacturing [20,21,22]. Additionally, mRNA vaccines possess self-adjuvanting properties, inducing strong and durable adaptive immune responses through cytokines such as TNF-α and IFN-α [23], whereas protein-based vaccines require added adjuvants to achieve comparable immunogenicity [177].
Lipid nanoparticles (LNPs) play a central role in mRNA delivery, yet their components can contribute to toxicity. Ionizable lipids may activate immune pathways and promote inflammation, while PEGylated lipids can trigger immune reactions and alter biodistribution. Biodegradable alternatives such as trehalose glycolipids and polysarcosine are being explored to improve safety without compromising delivery efficiency [179]. Future LNP development aims to enhance organ- and cell-specific targeting, supported by advanced in-vitro platforms such as microfluidic systems. Strategies including cell-specific promoters, targeting ligands, optimized lipid compositions, and zwitterionic materials may further improve precision and biocompatibility, although active targeting increases manufacturing complexity [179,180].
Given their high effectiveness and rapid adaptability once a viral sequence is known, mRNA vaccines remain among the most promising vaccine technologies, particularly during pandemics [175]. A systemic review of 74 studies found that most adverse reactions were mild to moderate, with few serious events. The Pfizer-BioNTech vaccine had more reported side effects due to earlier and broader use. Serious adverse events were primarily allergic, neurological, and cardiovascular, including myocarditis and myocardial infarction. Local and systemic reactions were more common after the second dose and in younger individuals, but were generally short-lived. No significant association was found between mRNA vaccination and cardiac events other than myopericarditis. Pregnancy outcomes were similar between vaccinated and unvaccinated women, and allergic reactions were more frequent with Pfizer than Moderna. Neurological events were rare and causality remained uncertain. Continued safety monitoring, particularly for cardiac events, is recommended [181].
6.5. Innovations in Large-Scale mRNA Vaccines and Diagnostics
The biomanufacturing industry entered the COVID-19 pandemic facing an unprecedented mandate: to develop, validate, and mass-produce diagnostics and vaccines at a scale and speed without modern precedent. Analyses highlight how global demand for billions of doses and tests placed extraordinary strain on supply chains, raw-material availability, and production infrastructure, forcing rapid capacity expansion and real-time process redesign [182,183]. To overcome the cost, scalability, and distribution limitations of conventional mRNA vaccine manufacturing, next-generation platforms have emerged that integrate modular architectures, decentralized deployment, process intensification, and advanced digital automation to enable more agile and globally accessible production [184]. Among these innovations, a slug-flow–based scalable mRNA machine provides a modular, automated system capable of producing between 1,000 clinical doses and 10 million commercial doses within a single regulatory-aligned framework, offering a practical blueprint for rapid scale-up [185]. Broader adoption of decentralized facilities, continuous-flow processing, and intensified operations further helped manufacturers navigate raw-material shortages and accelerate global output, while underscoring persistent challenges in cost and quality management [184].
A major remaining barrier to widespread mRNA deployment is the need for technologies that can generate precisely defined LNP formulations at throughputs that scale seamlessly from discovery to commercial manufacturing while meeting stringent pharmaceutical quality standards [186]. A notable advance is a scalable microfluidic chip platform incorporating 1×, 10×, or 256× LNP-generation units, enabling production rates up to 17 L/h while maintaining consistent LNP size, composition, and in vivo potency. Fabricated from solvent-resistant silicon and glass, these chips are compatible with pharmaceutical workflows, fully resettable, and reusable. SARS-CoV-2 mRNA-LNP vaccines produced using this system elicited strong antibody responses in preclinical models, demonstrating its suitability for commercial-scale manufacturing [186]. Complementing these advances, another group developed an aqueous, scalable mRNA manufacturing process compatible with GMP-compliant production, along with the analytical methods required for product characterization, release testing, and stability assessment [187]. Optimization of plasmid preparation, in vitro transcription, purification, and GMP analytics further enabled the production of safe, potent, and globally deployable mRNA vaccines [187].
In parallel, diagnostic manufacturers addressed severe shortages of reagents, plastics, and enzymes through automation, high-throughput assembly, and regulatory flexibility [188]. While centralized RT-qPCR laboratories remained the gold standard for accuracy, their reliance on sample transport and batch processing created delays. Decentralized on-site and self-testing approaches—particularly antigen-based lateral flow assays—expanded access to rapid testing in clinics, workplaces, and homes. These point-of-care tools reduced exposure risks for healthcare workers and enabled frequent population-level screening, though with lower sensitivity than laboratory PCR [188].
Together, these innovations in automation, process engineering, supply-chain expansion, and regulatory alignment enabled the biomanufacturing sector to meet the extraordinary diagnostic and vaccine demands imposed by the COVID-19 pandemic.
6.5.1. Workforce and Expertise Gaps
During the COVID-19 vaccine and diagnostic scale-up, biomanufacturers faced severe workforce and expertise shortages, particularly in GMP operations, large-scale biologics production, and emerging modalities such as mRNA and viral-vector manufacturing. Analyses show that the surge in global demand collided with an already limited talent pipeline, creating bottlenecks in engineering, quality assurance, and bioprocess operations [182,183]. Many facilities struggled to recruit experienced staff and instead relied on accelerated cross-training, redeployment of personnel from non-critical programs, and emergency upskilling initiatives supported by governments and industry consortia (NIIMBL. 2022 Workforce Development Report). Regulatory agencies also introduced temporary flexibilities that allowed newly trained workers to operate under enhanced oversight, helping stabilize production during peak demand (FDA, 2020–2022; EMA, 2020). Although these measures mitigated immediate shortages, expert assessments emphasize that they were stop-gap solutions rather than long-term fixes, and that sustained investment in biomanufacturing education and workforce development remains essential for future pandemic preparedness (NIIMBL. 2022 Workforce Development Report [[183].
7. Discussion
The rapid global response to COVID-19 demonstrated how profoundly the biomanufacturing industry shapes modern public-health preparedness. While traditional vaccines have long been effective in controlling infectious diseases, the pandemic underscored the limitations of conventional platforms and accelerated the transition toward next-generation technologies. RNA-based vaccines emerged as transformative tools, with RNA platforms offering unparalleled speed in design, optimization, and production [189]. Decades of foundational research in mRNA biology, delivery systems, and lipid nanoparticle engineering enabled the swift development of the Pfizer-BioNTech and Moderna vaccines, which exceeded expectations in both efficacy and scalability [190]. Their success established mRNA technology as a versatile platform not only for pandemic response but also for future therapeutic applications.
The biomanufacturing industry played a central role in translating these scientific advances into deployable products at an unprecedented pace. To meet global demand, manufacturers rapidly expanded production capacity, implemented advanced bioprocessing strategies, and optimized upstream and downstream workflows. Strategic collaborations between pharmaceutical companies, contract manufacturing organizations, and regulatory agencies facilitated rapid technology transfer, harmonized quality standards, and accelerated time-to-market. These partnerships enabled efficient resource sharing, minimized production bottlenecks, and ensured consistent vaccine supply across diverse regions.
At the same time, the pandemic exposed critical vulnerabilities in global manufacturing systems, including shortages of raw materials, limited availability of specialized equipment, and workforce constraints. The cold-storage requirements for mRNA vaccines, which often demand ultra-low temperatures and specialized freezers, dry ice logistics, and strict handling protocols, added a further layer of complexity, increasing costs, limiting distribution in low-resource settings, and raising the risk of potency loss if the cold chain is interrupted. These challenges highlighted the need for more resilient supply chains, diversified manufacturing hubs, and sustained investment in biomanufacturing infrastructure and R&D. The lessons learned from COVID-19 emphasize that scientific innovation must be paired with robust, flexible, and globally coordinated manufacturing capabilities to ensure rapid and equitable access to diagnostics and vaccines during future health emergencies.
To address the unprecedented demand, the biomanufacturing industry has significantly scaled production, utilizing advanced facilities and bioprocessing techniques. Collaborations between pharmaceutical companies and contract manufacturing organizations have expedited vaccine availability through resource optimization and knowledge sharing.
8. Conclusion
The COVID-19 pandemic highlighted the essential role of the biomanufacturing industry in rapidly developing and scaling diagnostics and vaccines. Advances in molecular testing, next-generation sequencing, nanotechnology, and RNA-based platforms enabled an unprecedented global response, even as the crisis exposed vulnerabilities in supply chains, workforce capacity, and manufacturing infrastructure. Moving forward, the modularity of RNA therapeutics, combined with innovations such as self-amplifying RNA, next-generation microfluidic mixers, integrated production suites, and AI-enhanced platforms promises to transform biomanufacturing. These technologies support more flexible, decentralized, and scalable manufacturing models, aligning with global efforts to establish domestic RNA production capacity and improve preparedness for future pandemics. Emerging multiproduct facilities capable of producing diverse RNA therapeutics on demand will enhance resilience, expand access to rare-disease treatments, and accelerate progress toward personalized RNA medicines. The lessons and innovations from COVID-19 thus lay the foundation for a more agile and globally accessible biomanufacturing ecosystem.
Author Contributions
The conceptualization and design of this review article were initiated by Ishfaq Ahmed. He conducted extensive research, reviewed the relevant literature, and was responsible for writing the manuscript. Additionally, he prepared all the figures and tables included in this article. Quendrix Martinez, Shayne McRae and Ashwin Dharmalingam participated in the research process as part of their learning experience, following directions, and gaining valuable skills in research methodologies, although only a small portion of their writing was used in the manuscript.
Acknowledgments
I extend my heartfelt gratitude to all the undergraduate students who participated in my “Teaching research strategies to undergraduate students through COVID-19 studies” project, which commenced in 2020 and culminated in this comprehensive review article. While most of the writing and preparation of this article was carried out by the corresponding author, the involvement of co-authors in learning and applying research methodologies played a key role in the preparation of this manuscript (IA).
Conflicts of interest
Authors declare no conflict of interest.
References
- Zhou, P; Yang, X-L; Wang, X-G; Hu, B; Zhang, L; Zhang, W; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Lu, R; Zhao, X; Li, J; Niu, P; Yang, B; Wu, H; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. The Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, S; Herbert, JA; McNamara, PS; Hedrich, CM. COVID-19: Immunology and treatment options. Clinical Immunology 2020, 215, 108448. [Google Scholar] [CrossRef]
- Ghasemiyeh, P; Borhani-Haghighi, A; Karimzadeh, I; Mohammadi-Samani, S; Vazin, A; Safari, A; et al. Major Neurologic Adverse Drug Reactions, Potential Drug–Drug Interactions and Pharmacokinetic Aspects of Drugs Used in COVID-19 Patients with Stroke: A Narrative Review. Ther Clin Risk Manag 2020, 16, 595–605. [Google Scholar] [CrossRef]
- Beigel, JH; Tomashek, KM; Dodd, LE; Mehta, AK; Zingman, BS; Kalil, AC; et al. Remdesivir for the Treatment of Covid-19 - Final Report. N Engl J Med 2020, 383, 1813–1826. [Google Scholar] [CrossRef] [PubMed]
- Molnupiravir in unvaccinated patients with COVID-19. Drug Ther Bull 2022, 60, 35–35. [CrossRef]
- Vassilopoulos, A; Mylonakis, E. In patients with COVID-19 at risk for severe disease, nirmatrelvir + ritonavir reduced hospitalization or death. Ann Intern Med 2022, 175, JC63. [Google Scholar] [CrossRef]
- Yadav, R; Chaudhary, JK; Jain, N; Chaudhary, PK; Khanra, S; Dhamija, P; et al. Role of Structural and Non-Structural Proteins and Therapeutic Targets of SARS-CoV-2 for COVID-19. Cells 2021, 10, 821. [Google Scholar] [CrossRef]
- Yan, W; Zheng, Y; Zeng, X; He, B; Cheng, W. Structural biology of SARS-CoV-2: open the door for novel therapies. Signal Transduct Target Ther 2022, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Malone, B; Urakova, N; Snijder, EJ; Campbell, EA. Structures and functions of coronavirus replication–transcription complexes and their relevance for SARS-CoV-2 drug design. Nat Rev Mol Cell Biol 2022, 23, 21–39. [Google Scholar] [CrossRef]
- Brian, DA; Baric, RS. Coronavirus Genome Structure and Replication; 2005; pp. 1–30. [Google Scholar] [CrossRef]
- Jamison, DA; Anand Narayanan, S; Trovão, NS; Guarnieri, JW; Topper, MJ; Moraes-Vieira, PM; et al. A comprehensive SARS-CoV-2 and COVID-19 review, Part 1: Intracellular overdrive for SARS-CoV-2 infection. European Journal of Human Genetics 2022, 30, 889–898. [Google Scholar] [CrossRef]
- Yan, W; Zheng, Y; Zeng, X; He, B; Cheng, W. Structural biology of SARS-CoV-2: open the door for novel therapies. Signal Transduct Target Ther 2022, 7, 26. [Google Scholar] [CrossRef]
- Wu, W; Cheng, Y; Zhou, H; Sun, C; Zhang, S. The SARS-CoV-2 nucleocapsid protein: its role in the viral life cycle, structure and functions, and use as a potential target in the development of vaccines and diagnostics. Virol J 2023, 20, 6. [Google Scholar] [CrossRef]
- Zhang, Z; Nomura, N; Muramoto, Y; Ekimoto, T; Uemura, T; Liu, K; et al. Structure of SARS-CoV-2 membrane protein essential for virus assembly. Nat Commun 2022, 13, 4399. [Google Scholar] [CrossRef] [PubMed]
- Oronsky, B; Larson, C; Caroen, S; Hedjran, F; Sanchez, A; Prokopenko, E; et al. Nucleocapsid as a next-generation COVID-19 vaccine candidate. International Journal of Infectious Diseases 2022, 122, 529–530. [Google Scholar] [CrossRef]
- Papageorgiou, AC; Mohsin, I. The SARS-CoV-2 Spike Glycoprotein as a Drug and Vaccine Target: Structural Insights into Its Complexes with ACE2 and Antibodies. Cells 2020, 9, 2343. [Google Scholar] [CrossRef] [PubMed]
- Peeling, RW; Heymann, DL; Teo, Y-Y; Garcia, PJ. Diagnostics for COVID-19: moving from pandemic response to control. The Lancet 2022, 399, 757–768. [Google Scholar] [CrossRef]
- Hoffmann, M; Kleine-Weber, H; Schroeder, S; Krüger, N; Herrler, T; Erichsen, S; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Aleem, A; Akbar Samad, AB; Vaqar, S. Emerging Variants of SARS-CoV-2 and Novel Therapeutics Against Coronavirus (COVID-19). 2024. [Google Scholar]
- Scovino, AM; Dahab, EC; Vieira, GF; Freire-de-Lima, L; Freire-de-Lima, CG; Morrot, A. SARS-CoV-2’s Variants of Concern: A Brief Characterization. Front Immunol 2022, 13. [Google Scholar] [CrossRef]
- Ou, J; Lan, W; Wu, X; Zhao, T; Duan, B; Yang, P; et al. Tracking SARS-CoV-2 Omicron diverse spike gene mutations identifies multiple inter-variant recombination events. Signal Transduct Target Ther 2022, 7, 138. [Google Scholar] [CrossRef]
- Ma, KC; Castro, J; Lambrou, AS; Rose, EB; Cook, PW; Batra, D; et al. Genomic Surveillance for SARS-CoV-2 Variants: Circulation of Omicron XBB and JN.1 Lineages — United States, May 2023–September 2024. MMWR Morb Mortal Wkly Rep 2024, 73, 938–945. [Google Scholar] [CrossRef]
- Salehi-Vaziri, M; Fazlalipour, M; Seyed Khorrami, SM; Azadmanesh, K; Pouriayevali, MH; Jalali, T; et al. The ins and outs of SARS-CoV-2 variants of concern (VOCs). Arch Virol 2022, 167, 327–344. [Google Scholar] [CrossRef]
- Davies, NG; Abbott, S; Barnard, RC; Jarvis, CI; Kucharski, AJ; Munday, JD; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science (1979) 2021, 372. [Google Scholar] [CrossRef]
- Tegally, H; Wilkinson, E; Giovanetti, M; Iranzadeh, A; Fonseca, V; Giandhari, J; et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Expression of concern on: Comparative genomics and characterization of SARS-CoV-2 P.1 (Gamma) variant of concern from Amazonas, Brazil. Front Med (Lausanne) 2024, 11. [CrossRef]
- Mlcochova, P; Kemp, SA; Dhar, MS; Papa, G; Meng, B; Ferreira, IATM; et al. SARS-CoV-2 B.1.617.2 Delta variant replication and immune evasion. Nature 2021, 599, 114–119. [Google Scholar] [CrossRef]
- Willett, BJ; Grove, J; MacLean, OA; Wilkie, C; De Lorenzo, G; Furnon, W; et al. SARS-CoV-2 Omicron is an immune escape variant with an altered cell entry pathway. Nat Microbiol 2022, 7, 1161–1179. [Google Scholar] [CrossRef] [PubMed]
- Mambelli, F; de Araujo, ACVSC; Farias, JP; de Andrade, KQ; Ferreira, LCS; Minoprio, P; et al. An Update on Anti-COVID-19 Vaccines and the Challenges to Protect Against New SARS-CoV-2 Variants. Pathogens 2025, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Umakanthan, S; Chattu, VK; Ranade, A V; Das, D; Basavarajegowda, A; Bukelo, M. A rapid review of recent advances in diagnosis, treatment and vaccination for COVID-19. AIMS Public Health 2021, 8, 137–153. [Google Scholar] [CrossRef]
- Walls, AC; Park, Y-J; Tortorici, MA; Wall, A; McGuire, AT; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Hartenian, E; Nandakumar, D; Lari, A; Ly, M; Tucker, JM; Glaunsinger, BA. The molecular virology of coronaviruses. Journal of Biological Chemistry 2020, 295, 12910–12934. [Google Scholar] [CrossRef]
- Snijder, EJ; Limpens, RWAL; de Wilde, AH; de Jong, AWM; Zevenhoven-Dobbe, JC; Maier, HJ; et al. A unifying structural and functional model of the coronavirus replication organelle: Tracking down RNA synthesis. PLoS Biol 2020, 18, e3000715. [Google Scholar] [CrossRef]
- Klein, S; Cortese, M; Winter, SL; Wachsmuth-Melm, M; Neufeldt, CJ; Cerikan, B; et al. SARS-CoV-2 structure and replication characterized by in situ cryo-electron tomography. Nat Commun 2020, 11, 5885. [Google Scholar] [CrossRef]
- V’kovski, P; Kratzel, A; Steiner, S; Stalder, H; Thiel, V. Coronavirus biology and replication: implications for SARS-CoV-2. Nat Rev Microbiol 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Guan, W; Ni, Z; Hu, Y; Liang, W; Ou, C; He, J; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. New England Journal of Medicine 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Dutta, D; Naiyer, S; Mansuri, S; Soni, N; Singh, V; Bhat, KH; et al. COVID-19 Diagnosis: A Comprehensive Review of the RT-qPCR Method for Detection of SARS-CoV-2. Diagnostics 2022, 12, 1503. [Google Scholar] [CrossRef] [PubMed]
- Drain, PK. Rapid Diagnostic Testing for SARS-CoV-2. New England Journal of Medicine 2022, 386, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Lu, X; Wang, L; Sakthivel, SK; Whitaker, B; Murray, J; Kamili, S; et al. US CDC Real-Time Reverse Transcription PCR Panel for Detection of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg Infect Dis 2020, 26, 1654–1665. [Google Scholar] [CrossRef]
- Cheng, MP; Papenburg, J; Desjardins, M; Kanjilal, S; Quach, C; Libman, M; et al. Diagnostic Testing for Severe Acute Respiratory Syndrome–Related Coronavirus 2. Ann Intern Med 2020, 172, 726–734. [Google Scholar] [CrossRef]
- Dutta, D; Naiyer, S; Mansuri, S; Soni, N; Singh, V; Bhat, KH; et al. COVID-19 Diagnosis: A Comprehensive Review of the RT-qPCR Method for Detection of SARS-CoV-2. Diagnostics 2022, 12, 1503. [Google Scholar] [CrossRef]
- Chu, VT; Schwartz, NG; Donnelly, MAP; Chuey, MR; Soto, R; Yousaf, AR; et al. Comparison of Home Antigen Testing With RT-PCR and Viral Culture During the Course of SARS-CoV-2 Infection. JAMA Intern Med 2022, 182, 701. [Google Scholar] [CrossRef]
- Corman, VM; Landt, O; Kaiser, M; Molenkamp, R; Meijer, A; Chu, DK; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef]
- Liu, R; Han, H; Liu, F; Lv, Z; Wu, K; Liu, Y; et al. Positive rate of RT-PCR detection of SARS-CoV-2 infection in 4880 cases from one hospital in Wuhan, China, from Jan to Feb 2020. Clinica Chimica Acta 2020, 505, 172–175. [Google Scholar] [CrossRef]
- Shen, Z; Xiao, Y; Kang, L; Ma, W; Shi, L; Zhang, L; et al. Genomic Diversity of Severe Acute Respiratory Syndrome–Coronavirus 2 in Patients With Coronavirus Disease 2019. Clinical Infectious Diseases 2020, 71, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Perez-Romero, CA; Mendoza-Maldonado, L; Tonda, A; Coz, E; Tabeling, P; Vanhomwegen, J; et al. An Innovative AI-based primer design tool for precise and accurate detection of SARS-CoV-2 variants of concern. Sci Rep 2023, 13, 15782. [Google Scholar] [CrossRef]
- Peeling, RW; Olliaro, PL; Boeras, DI; Fongwen, N. Scaling up COVID-19 rapid antigen tests: promises and challenges. Lancet Infect Dis 2021, 21, e290–e295. [Google Scholar] [CrossRef] [PubMed]
- Brümmer, LE; Katzenschlager, S; Gaeddert, M; Erdmann, C; Schmitz, S; Bota, M; et al. Accuracy of novel antigen rapid diagnostics for SARS-CoV-2: A living systematic review and meta-analysis. PLoS Med 2021, 18, e1003735. [Google Scholar] [CrossRef]
- Schildgen, V; Demuth, S; Lüsebrink, J; Schildgen, O. Limits and Opportunities of SARS-CoV-2 Antigen Rapid Tests: An Experienced-Based Perspective. Pathogens 2021, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Lange, SJ; Kompaniyets, L; Freedman, DS; Kraus, EM; Porter, R; Blanck, HM; et al. Longitudinal Trends in Body Mass Index Before and During the COVID-19 Pandemic Among Persons Aged 2–19 Years — United States, 2018–2020. MMWR Morb Mortal Wkly Rep 2021, 70, 1278–1283. [Google Scholar] [CrossRef]
- Kohmer, N; Toptan, T; Pallas, C; Karaca, O; Pfeiffer, A; Westhaus, S; et al. The Comparative Clinical Performance of Four SARS-CoV-2 Rapid Antigen Tests and Their Correlation to Infectivity In Vitro. J Clin Med 2021, 10, 328. [Google Scholar] [CrossRef]
- Patriquin, G; LeBlanc, JJ; Williams, C; Hatchette, TF; Ross, J; Barrett, L; et al. Comparison between Nasal and Nasopharyngeal Swabs for SARS-CoV-2 Rapid Antigen Detection in an Asymptomatic Population, and Direct Confirmation by RT-PCR from the Residual Buffer. Microbiol Spectr 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Dinnes, J; Berhane, S; Walsh, J; Reidy, P; Doherty, A; Hillier, B; et al. Rapid, point-of-care antigen tests for diagnosis of SARS-CoV-2 infection. Cochrane Database of Systematic Reviews 2025, 2025. [Google Scholar] [CrossRef]
- Arshadi, M; Fardsanei, F; Deihim, B; Farshadzadeh, Z; Nikkhahi, F; Khalili, F; et al. Diagnostic Accuracy of Rapid Antigen Tests for COVID-19 Detection: A Systematic Review With Meta-analysis. Front Med (Lausanne) 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Baro, B; Rodo, P; Ouchi, D; Bordoy, AE; Saya Amaro, EN; Salsench, S V.; et al. Performance characteristics of five antigen-detecting rapid diagnostic test (Ag-RDT) for SARS-CoV-2 asymptomatic infection: a head-to-head benchmark comparison. Journal of Infection 2021, 82, 269–275. [Google Scholar] [CrossRef]
- Kim, AE; Bennett, JC; Luiten, K; O’Hanlon, JA; Wolf, CR; Magedson, A; et al. Comparative Diagnostic Utility of SARS-CoV-2 Rapid Antigen and Molecular Testing in a Community Setting. J Infect Dis 2024, 230, 363–373. [Google Scholar] [CrossRef]
- Wagenhäuser, I; Knies, K; Pscheidl, T; Eisenmann, M; Flemming, S; Petri, N; et al. SARS-CoV-2 antigen rapid detection tests: test performance during the COVID-19 pandemic and the impact of COVID-19 vaccination. EBioMedicine 2024, 109, 105394. [Google Scholar] [CrossRef]
- Derin, DÇ; Gültekin, E. Design of a lateral flow assay targeting the conserved NIID_2019-nCoV_N gene region for molecular viral diagnosis. Brazilian Journal of Medical and Biological Research 2025, 58. [Google Scholar] [CrossRef]
- Kumar, A; Tripathi, P; Kumar, P; Shekhar, R; Pathak, R. From Detection to Protection: Antibodies and Their Crucial Role in Diagnosing and Combatting SARS-CoV-2. Vaccines (Basel) 2024, 12, 459. [Google Scholar] [CrossRef]
- Kevadiya, BD; Machhi, J; Herskovitz, J; Oleynikov, MD; Blomberg, WR; Bajwa, N; et al. Diagnostics for SARS-CoV-2 infections. Nat Mater 2021, 20, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Udugama, B; Kadhiresan, P; Kozlowski, HN; Malekjahani, A; Osborne, M; Li, VYC; et al. Diagnosing COVID-19: The Disease and Tools for Detection. ACS Nano 2020, 14, 3822–3835. [Google Scholar] [CrossRef] [PubMed]
- Long, Q-X; Tang, X-J; Shi, Q-L; Li, Q; Deng, H-J; Yuan, J; et al. Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nat Med 2020, 26, 1200–1204. [Google Scholar] [CrossRef]
- Thapa, S; Singh, KR; Verma, R; Singh, J; Singh, RP. State-of-the-Art Smart and Intelligent Nanobiosensors for SARS-CoV-2 Diagnosis. Biosensors (Basel) 2022, 12, 637. [Google Scholar] [CrossRef]
- Choi, G; Moehling, TJ; Meagher, RJ. Advances in RT-LAMP for COVID-19 testing and diagnosis. Expert Rev Mol Diagn 2023, 23, 9–28. [Google Scholar] [CrossRef]
- Fernandes, RS; de Oliveira Silva, J; Gomes, KB; Azevedo, RB; Townsend, DM; de Paula Sabino, A; et al. Recent advances in point of care testing for COVID-19 detection. Biomedicine & Pharmacotherapy 2022, 153, 113538. [Google Scholar] [CrossRef]
- Behjati, S; Tarpey, PS. What is next generation sequencing? Arch Dis Child Educ Pract Ed 2013, 98, 236–238. [Google Scholar] [CrossRef]
- Shu, Y; McCauley, J. GISAID: Global initiative on sharing all influenza data – from vision to reality. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef]
- Hatcher, EL; Zhdanov, SA; Bao, Y; Blinkova, O; Nawrocki, EP; Ostapchuck, Y; et al. Virus Variation Resource – improved response to emergent viral outbreaks. Nucleic Acids Res 2017, 45, D482–90. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W-M; Song, S-H; Chen, M-L; Zou, D; Ma, L-N; Ma, Y-K; et al. The 2019 novel coronavirus resource. Yi Chuan 2020, 42, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y; Zmasek, C; Sun, G; Larsen, CN; Scheuermann, RH. Hepatitis C Virus Database and Bioinformatics Analysis Tools in the Virus Pathogen Resource (ViPR); 2019; pp. 47–69. [Google Scholar] [CrossRef]
- Zhu, N; Zhang, D; Wang, W; Li, X; Yang, B; Song, J; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. New England Journal of Medicine 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Zhou, H; Chen, X; Hu, T; Li, J; Song, H; Liu, Y; et al. A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Current Biology 2020, 30, 2196–2203.e3. [Google Scholar] [CrossRef] [PubMed]
- Chen, X; Kang, Y; Luo, J; Pang, K; Xu, X; Wu, J; et al. Next-Generation Sequencing Reveals the Progression of COVID-19. Front Cell Infect Microbiol 2021, 11. [Google Scholar] [CrossRef]
- Ramuta, MD; Newman, CM; Brakefield, SF; Stauss, MR; Wiseman, RW; Kita-Yarbro, A; et al. SARS-CoV-2 and other respiratory pathogens are detected in continuous air samples from congregate settings. Nat Commun 2022, 13, 4717. [Google Scholar] [CrossRef]
- Minor, NR; Ramuta, MD; Stauss, MR; Harwood, OE; Brakefield, SF; Alberts, A; et al. Metagenomic sequencing detects human respiratory and enteric viruses in air samples collected from congregate settings. Sci Rep 2023, 13, 21398. [Google Scholar] [CrossRef]
- Andersen, KG; Rambaut, A; Lipkin, WI; Holmes, EC; Garry, RF. The proximal origin of SARS-CoV-2. Nat Med 2020, 26, 450–452. [Google Scholar] [CrossRef]
- Yoo, HM; Kim, I-H; Kim, S. Nucleic Acid Testing of SARS-CoV-2. Int J Mol Sci 2021, 22, 6150. [Google Scholar] [CrossRef]
- Campos, GS; Sardi, SI; Falcao, MB; Belitardo, EMMA; Rocha, DJPG; Rolo, CA; et al. Ion torrent-based nasopharyngeal swab metatranscriptomics in COVID-19. J Virol Methods 2020, 282, 113888. [Google Scholar] [CrossRef]
- Paden, CR; Tao, Y; Queen, K; Zhang, J; Li, Y; Uehara, A; et al. Rapid, Sensitive, Full-Genome Sequencing of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg Infect Dis 2020, 26, 2401–2405. [Google Scholar] [CrossRef] [PubMed]
- Pillay, S; Giandhari, J; Tegally, H; Wilkinson, E; Chimukangara, B; Lessells, R; et al. Whole Genome Sequencing of SARS-CoV-2: Adapting Illumina Protocols for Quick and Accurate Outbreak Investigation during a Pandemic. Genes (Basel) 2020, 11, 949. [Google Scholar] [CrossRef]
- Mercatelli, D; Holding, AN; Giorgi, FM. Web tools to fight pandemics: the COVID-19 experience. Brief Bioinform 2021, 22, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Hu, T; Li, J; Zhou, H; Li, C; Holmes, EC; Shi, W. Bioinformatics resources for SARS-CoV-2 discovery and surveillance. Brief Bioinform 2021, 22, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I; Yusuf, K; Roy, BC; Stubbs, J; Anant, S; Attard, TM; et al. Dietary Interventions Ameliorate Infectious Colitis by Restoring the Microbiome and Promoting Stem Cell Proliferation in Mice. Int J Mol Sci 2021, 23, 339. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I; Roy, BC; Raach, R-MT; Owens, SM; Xia, L; Anant, S; et al. Enteric infection coupled with chronic Notch pathway inhibition alters colonic mucus composition leading to dysbiosis, barrier disruption and colitis. PLoS One 2018, 13, e0206701. [Google Scholar] [CrossRef] [PubMed]
- Charre, C; Ginevra, C; Sabatier, M; Regue, H; Destras, G; Brun, S; et al. Evaluation of NGS-based approaches for SARS-CoV-2 whole genome characterisation. Virus Evol 2020, 6. [Google Scholar] [CrossRef]
- Viana, R; Moyo, S; Amoako, DG; Tegally, H; Scheepers, C; Althaus, CL; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Wang, R; Hozumi, Y; Yin, C; Wei, G-W. Mutations on COVID-19 diagnostic targets. Genomics 2020, 112, 5204–5213. [Google Scholar] [CrossRef]
- Daviña-Núñez, C; Pérez, S; Cabrera-Alvargonzález, JJ; Rincón-Quintero, A; Treinta-Álvarez, A; Godoy-Diz, M; et al. Performance of amplicon and capture based next-generation sequencing approaches for the epidemiological surveillance of Omicron SARS-CoV-2 and other variants of concern. PLoS One 2024, 19, e0289188. [Google Scholar] [CrossRef]
- du Plessis, L; McCrone, JT; Zarebski, AE; Hill, V; Ruis, C; Gutierrez, B; et al. Establishment and lineage dynamics of the SARS-CoV-2 epidemic in the UK. Science 2021, 371, 708–712. [Google Scholar] [CrossRef]
- Haleem, A; Javaid, M; Singh, RP; Rab, S; Suman, R. Applications of nanotechnology in medical field: a brief review. Global Health Journal 2023, 7, 70–77. [Google Scholar] [CrossRef]
- Singh, P; Singh, D; Sa, P; Mohapatra, P; Khuntia, A; K Sahoo, S. Insights From Nanotechnology in COVID-19: Prevention, Detection, Therapy and Immunomodulation. Nanomedicine 2021, 16, 1219–1235. [Google Scholar] [CrossRef]
- Ayan, S; Aranci-Ciftci, K; Ciftci, F; Ustundag, CB. Nanotechnology and COVID-19: Prevention, diagnosis, vaccine, and treatment strategies. Front Mater 2023, 9. [Google Scholar] [CrossRef]
- Sharma, A; Kontodimas, K; Bosmann, M. Nanomedicine: A Diagnostic and Therapeutic Approach to COVID-19. Front Med (Lausanne) 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Fernández, A; Cabrera-Rodriguez, R; Ciuffreda, L; Perez-Yanes, S; Estevez-Herrera, J; González-Montelongo, R; et al. Nanomaterials to combat SARS-CoV-2: Strategies to prevent, diagnose and treat COVID-19. Front Bioeng Biotechnol 2022, 10. [Google Scholar] [CrossRef]
- Alhalaili, B; Popescu, IN; Kamoun, O; Alzubi, F; Alawadhia, S; Vidu, R. Nanobiosensors for the Detection of Novel Coronavirus 2019-nCoV and Other Pandemic/Epidemic Respiratory Viruses: A Review. Sensors 2020, 20, 6591. [Google Scholar] [CrossRef]
- Valerio, TL; Anastácio, R; da Silva, SS; de Oliveira, CC; Vidotti, M. An overview of electrochemical biosensors used for COVID-19 detection. Analytical Methods 2024, 16, 2164–2176. [Google Scholar] [CrossRef]
- Tekin, YS; Kul, SM; Sagdic, O; Rodthongkum, N; Geiss, B; Ozer, T. Optical biosensors for diagnosis of COVID-19: nanomaterial-enabled particle strategies for post pandemic era. Microchimica Acta 2024, 191, 320. [Google Scholar] [CrossRef]
- Harun-Ur-Rashid, M; Foyez, T; Jahan, I; Pal, K; Imran, A.B. Rapid diagnosis of COVID-19 via nano-biosensor-implemented biomedical utilization: a systematic review. RSC Adv 2022, 12, 9445–9465. [Google Scholar] [CrossRef]
- Pardi, N; Hogan, MJ; Porter, FW; Weissman, D. mRNA vaccines — a new era in vaccinology. Nat Rev Drug Discov 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Martinon, F; Krishnan, S; Lenzen, G; Magné, R; Gomard, E; Guillet, J; et al. Induction of virus-specific cytotoxic T lymphocytes in vivo by liposome-entrapped mRNA. Eur J Immunol 1993, 23, 1719–1722. [Google Scholar] [CrossRef]
- BRENNER, S; JACOB, F; MESELSON, M. An Unstable Intermediate Carrying Information from Genes to Ribosomes for Protein Synthesis. Nature 1961, 190, 576–581. [Google Scholar] [CrossRef]
- Lockard, RE; Lingrel, JB. The synthesis of mouse hemoglobin chains in a rabbit reticulocyte cell-free system programmed with mouse reticulocyte 9S RNA. Biochem Biophys Res Commun 1969, 37, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Krieg, PA; Melton, DA. 25] In vitro RNA synthesis with SP6 RNA polymerase; 1987; pp. 397–415. [Google Scholar] [CrossRef]
- Krieg, PA; Melton, DA. Functional messenger RNAs are produced by SP6 in vitro transcription of cloned cDNAs. Nucleic Acids Res 1984, 12, 7057–7070. [Google Scholar] [CrossRef]
- Malone, RW; Felgner, PL; Verma, IM. Cationic liposome-mediated RNA transfection. Proceedings of the National Academy of Sciences 1989, 86, 6077–6081. [Google Scholar] [CrossRef]
- Wolff, JA; Budker, V. The Mechanism of Naked DNA Uptake and Expression; 2005; pp. 1–20. [Google Scholar] [CrossRef]
- Jirikowski, GF; Sanna, PP; Maciejewski-Lenoir, D; Bloom, FE. Reversal of Diabetes Insipidus in Brattleboro Rats: Intrahypothalamic Injection of Vasopressin mRNA. Science (1979) 1992, 255, 996–998. [Google Scholar] [CrossRef]
- Hassett, KJ; Benenato, KE; Jacquinet, E; Lee, A; Woods, A; Yuzhakov, O; et al. Optimization of Lipid Nanoparticles for Intramuscular Administration of mRNA Vaccines. Mol Ther Nucleic Acids 2019, 15, 1–11. [Google Scholar] [CrossRef]
- Conry, RM; LoBuglio, AF; Loechel, F; Moore, SE; Sumerel, LA; Barlow, DL; et al. A carcinoembryonic antigen polynucleotide vaccine has in vivo antitumor activity. Gene Ther 1995, 2, 59–65. [Google Scholar]
- Karikó, K; Ni, H; Capodici, J; Lamphier, M; Weissman, D. mRNA Is an Endogenous Ligand for Toll-like Receptor 3. Journal of Biological Chemistry 2004, 279, 12542–12550. [Google Scholar] [CrossRef]
- Andries, O; Mc Cafferty, S; De Smedt, SC; Weiss, R; Sanders, NN; Kitada, T. N1-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. Journal of Controlled Release 2015, 217, 337–344. [Google Scholar] [CrossRef]
- Karikó, K; Muramatsu, H; Ludwig, J; Weissman, D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res 2011, 39, e142–e142. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K; Buckstein, M; Ni, H; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y-Z; Holmes, EC. A Genomic Perspective on the Origin and Emergence of SARS-CoV-2. Cell 2020, 181, 223–227. [Google Scholar] [CrossRef]
- Freed, NE; Vlková, M; Faisal, MB; Silander, OK. Rapid and inexpensive whole-genome sequencing of SARS-CoV-2 using 1200 bp tiled amplicons and Oxford Nanopore Rapid Barcoding. Biol Methods Protoc 2020, 5. [Google Scholar] [CrossRef]
- Wrapp, D; Wang, N; Corbett, KS; Goldsmith, JA; Hsieh, C-L; Abiona, O; et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science (1979) 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Corbett, KS; Edwards, D; Leist, SR; Abiona, OM; Boyoglu-Barnum, S; Gillespie, RA; et al. SARS-CoV-2 mRNA Vaccine Development Enabled by Prototype Pathogen Preparedness 2020. [CrossRef]
- Polack, FP; Thomas, SJ; Kitchin, N; Absalon, J; Gurtman, A; Lockhart, S; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. New England Journal of Medicine 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, LR; El Sahly, HM; Essink, B; Kotloff, K; Frey, S; Novak, R; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. New England Journal of Medicine 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Sadoff, J; Gray, G; Vandebosch, A; Cárdenas, V; Shukarev, G; Grinsztejn, B; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. New England Journal of Medicine 2021, 384, 2187–2201. [Google Scholar] [CrossRef]
- Yin, Z; Chen, J-L; Lu, Y; Wang, B; Godfrey, L; Mentzer, AJ; et al. Evaluation of T cell responses to naturally processed variant SARS-CoV-2 spike antigens in individuals following infection or vaccination. Cell Rep 2023, 42, 112470. [Google Scholar] [CrossRef]
- Tamandjou Tchuem, CR; Auvigne, V; Vaux, S; Montagnat, C; Paireau, J; Monnier Besnard, S; et al. Vaccine effectiveness and duration of protection of COVID-19 mRNA vaccines against Delta and Omicron BA.1 symptomatic and severe COVID-19 outcomes in adults aged 50 years and over in France. Vaccine 2023, 41, 2280–2288. [Google Scholar] [CrossRef] [PubMed]
- Powell, AA; Kirsebom, F; Stowe, J; Ramsay, ME; Lopez-Bernal, J; Andrews, N; et al. Protection against symptomatic infection with delta (B.1.617.2) and omicron (B.1.1.529) BA.1 and BA.2 SARS-CoV-2 variants after previous infection and vaccination in adolescents in England, August, 2021–March, 2022: a national, observational, test-negative, case-control study. Lancet Infect Dis 2023, 23, 435–444. [Google Scholar] [CrossRef]
- Abu-Raddad, LJ; Chemaitelly, H; Ayoub, HH; AlMukdad, S; Yassine, HM; Al-Khatib, HA; et al. Effect of mRNA Vaccine Boosters against SARS-CoV-2 Omicron Infection in Qatar. New England Journal of Medicine 2022, 386, 1804–1816. [Google Scholar] [CrossRef]
- Heinz, FX; Stiasny, K. Profiles of current COVID-19 vaccines. Wien Klin Wochenschr 2021, 133, 271–283. [Google Scholar] [CrossRef]
- Ada, GL. The ideal vaccine. World J Microbiol Biotechnol 1991, 7, 105–109. [Google Scholar] [CrossRef]
- Florindo, HF; Kleiner, R; Vaskovich-Koubi, D; Acúrcio, RC; Carreira, B; Yeini, E; et al. Immune-mediated approaches against COVID-19. Nat Nanotechnol 2020, 15, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Yadav, T; Srivastava, N; Mishra, G; Dhama, K; Kumar, S; Puri, B; et al. Recombinant vaccines for COVID-19. Hum Vaccin Immunother 2020, 16, 2905–2912. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Guardeño, JM; Regla-Nava, JA; Nieto-Torres, JL; DeDiego, ML; Castaño-Rodriguez, C; Fernandez-Delgado, R; et al. Identification of the Mechanisms Causing Reversion to Virulence in an Attenuated SARS-CoV for the Design of a Genetically Stable Vaccine. PLoS Pathog 2015, 11, e1005215. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y; Dai, T; Wei, Y; Zhang, L; Zheng, M; Zhou, F. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduct Target Ther 2020, 5, 237. [Google Scholar] [CrossRef]
- Hotez, PJ; Bottazzi, ME. Whole Inactivated Virus and Protein-Based COVID-19 Vaccines. Annu Rev Med 2022, 73, 55–64. [Google Scholar] [CrossRef]
- Wilson-Welder, JH; Torres, MP; Kipper, MJ; Mallapragada, SK; Wannemuehler, MJ; Narasimhan, B. Vaccine adjuvants: Current challenges and future approaches. J Pharm Sci 2009, 98, 1278–1316. [Google Scholar] [CrossRef]
- Heath, PT; Galiza, EP; Baxter, DN; Boffito, M; Browne, D; Burns, F; et al. Safety and Efficacy of NVX-CoV2373 Covid-19 Vaccine. New England Journal of Medicine 2021, 385, 1172–1183. [Google Scholar] [CrossRef]
- Keech, C; Albert, G; Cho, I; Robertson, A; Reed, P; Neal, S; et al. Phase 1–2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. New England Journal of Medicine 2020, 383, 2320–2332. [Google Scholar] [CrossRef]
- McConeghy, KW; Davidson, HE; Canaday, DH; Han, L; Hayes, K; Baier, RR; et al. Recombinant vs Egg-Based Quadrivalent Influenza Vaccination for Nursing Home Residents. JAMA Netw Open 2025, 8, e2452677. [Google Scholar] [CrossRef]
- Matsumura, T; Takano, T; Takahashi, Y. Immune responses related to the immunogenicity and reactogenicity of COVID-19 mRNA vaccines. Int Immunol 2023, 35, 213–220. [Google Scholar] [CrossRef]
- Duan, L; Zheng, Q; Zhang, H; Niu, Y; Lou, Y; Wang, H. The SARS-CoV-2 Spike Glycoprotein Biosynthesis, Structure, Function, and Antigenicity: Implications for the Design of Spike-Based Vaccine Immunogens. Front Immunol 2020, 11. [Google Scholar] [CrossRef]
- Li, K; Huang, B; Wu, M; Zhong, A; Li, L; Cai, Y; et al. Dynamic changes in anti-SARS-CoV-2 antibodies during SARS-CoV-2 infection and recovery from COVID-19. Nat Commun 2020, 11, 6044. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y; Yang, C; Xu, X; Xu, W; Liu, S. Structural and functional properties of SARS-CoV-2 spike protein: potential antivirus drug development for COVID-19. Acta Pharmacol Sin 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, R. mRNA and Adenoviral Vector Vaccine Platforms Utilized in COVID-19 Vaccines: Technologies, Ecosystem, and Future Directions. Vaccines (Basel) 2023, 11, 1737. [Google Scholar] [CrossRef]
- Pardi, N; Hogan, MJ; Porter, FW; Weissman, D. mRNA vaccines — a new era in vaccinology. Nat Rev Drug Discov 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Kim, SC; Sekhon, SS; Shin, W-R; Ahn, G; Cho, B-K; Ahn, J-Y; et al. Modifications of mRNA vaccine structural elements for improving mRNA stability and translation efficiency. Mol Cell Toxicol 2022, 18, 1–8. [Google Scholar] [CrossRef]
- Polack, FP; Thomas, SJ; Kitchin, N; Absalon, J; Gurtman, A; Lockhart, S; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. New England Journal of Medicine 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Walsh, EE; Pérez Marc, G; Zareba, AM; Falsey, AR; Jiang, Q; Patton, M; et al. Efficacy and Safety of a Bivalent RSV Prefusion F Vaccine in Older Adults. New England Journal of Medicine 2023, 388, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Mugridge, JS; Coller, J; Gross, JD. Structural and molecular mechanisms for the control of eukaryotic 5′–3′ mRNA decay. Nat Struct Mol Biol 2018, 25, 1077–1085. [Google Scholar] [CrossRef]
- Chaudhary, N; Weissman, D; Whitehead, KA. mRNA vaccines for infectious diseases: principles, delivery and clinical translation. Nat Rev Drug Discov 2021, 20, 817–838. [Google Scholar] [CrossRef]
- Chatterjee, S; Pal, JK. Role of 5′- and 3′-untranslated regions of mRNAs in human diseases. Biol Cell 2009, 101, 251–262. [Google Scholar] [CrossRef]
- Pelletier, J; Schmeing, TM; Sonenberg, N. The multifaceted eukaryotic cap structure. WIREs RNA 2021, 12. [Google Scholar] [CrossRef]
- Tuller, T; Zur, H. Multiple roles of the coding sequence 5′ end in gene expression regulation. Nucleic Acids Res 2015, 43, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Courel, M; Clément, Y; Bossevain, C; Foretek, D; Vidal Cruchez, O; Yi, Z; et al. GC content shapes mRNA storage and decay in human cells. Elife 2019, 8. [Google Scholar] [CrossRef]
- Kudla, G; Lipinski, L; Caffin, F; Helwak, A; Zylicz, M. High Guanine and Cytosine Content Increases mRNA Levels in Mammalian Cells. PLoS Biol 2006, 4, e180. [Google Scholar] [CrossRef]
- Silva, J; Fernandes, R; Romão, L. Translational Regulation by Upstream Open Reading Frames and Human Diseases; 2019; pp. 99–116. [Google Scholar] [CrossRef]
- Eckmann, CR; Rammelt, C; Wahle, E. Control of poly(A) tail length. WIREs RNA 2011, 2, 348–361. [Google Scholar] [CrossRef]
- Biziaev, N; Shuvalov, A; Salman, A; Egorova, T; Shuvalova, E; Alkalaeva, E. The impact of mRNA poly(A) tail length on eukaryotic translation stages. Nucleic Acids Res 2024, 52, 7792–7808. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N; Hogan, MJ; Porter, FW; Weissman, D. mRNA vaccines — a new era in vaccinology. Nat Rev Drug Discov 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Ramachandran, S; Satapathy, SR; Dutta, T. Delivery Strategies for mRNA Vaccines. Pharmaceut Med 2022, 36, 11–20. [Google Scholar] [CrossRef]
- Chen, S; Huang, X; Xue, Y; Álvarez-Benedicto, E; Shi, Y; Chen, W; et al. Nanotechnology-based mRNA vaccines. Nature Reviews Methods Primers 2023, 3, 63. [Google Scholar] [CrossRef]
- Kiaie, SH; Majidi Zolbanin, N; Ahmadi, A; Bagherifar, R; Valizadeh, H; Kashanchi, F; et al. Recent advances in mRNA-LNP therapeutics: immunological and pharmacological aspects. J Nanobiotechnology 2022, 20, 276. [Google Scholar] [CrossRef]
- Heshmati, N; Chakka, LRJ; Zhang, Y; Maniruzzaman, M. Fabrication of mRNA encapsulated lipid nanoparticles using state of the art SMART-MaGIC technology and transfection in vitro. Sci Rep 2024, 14, 22714. [Google Scholar] [CrossRef] [PubMed]
- Kiaie, SH; Majidi Zolbanin, N; Ahmadi, A; Bagherifar, R; Valizadeh, H; Kashanchi, F; et al. Recent advances in mRNA-LNP therapeutics: immunological and pharmacological aspects. J Nanobiotechnology 2022, 20, 276. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, PS; Rudra, A; Miao, L; Anderson, DG. Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Molecular Therapy 2019, 27, 710–728. [Google Scholar] [CrossRef] [PubMed]
- Jackson, LA; Anderson, EJ; Rouphael, NG; Roberts, PC; Makhene, M; Coler, RN; et al. An mRNA Vaccine against SARS-CoV-2 — Preliminary Report. New England Journal of Medicine 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Verbeke, R; Lentacker, I; De Smedt, SC; Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. Journal of Controlled Release 2021, 333, 511–520. [Google Scholar] [CrossRef]
- Hou, X; Zaks, T; Langer, R; Dong, Y. Lipid nanoparticles for mRNA delivery. Nat Rev Mater 2021, 6, 1078–1094. [Google Scholar] [CrossRef]
- Li, X; Qi, J; Wang, J; Hu, W; Zhou, W; Wang, Y; et al. Nanoparticle technology for mRNA: Delivery strategy, clinical application and developmental landscape. Theranostics 2024, 14, 738–760. [Google Scholar] [CrossRef]
- Plett, PC. [Peter Plett and other discoverers of cowpox vaccination before Edward Jenner]. Sudhoffs Arch 2006, 90, 219–232. [Google Scholar]
- Jenner, E. Dr. Jenner, on the Vaccine Inoculation. Med Phys J 1800, 3, 502–503. [Google Scholar]
- Maruggi, G; Zhang, C; Li, J; Ulmer, JB; Yu, D. mRNA as a Transformative Technology for Vaccine Development to Control Infectious Diseases. Molecular Therapy 2019, 27, 757–772. [Google Scholar] [CrossRef]
- Krammer, F. SARS-CoV-2 vaccines in development. Nature 2020, 586, 516–527. [Google Scholar] [CrossRef]
- Ruiz-Fresneda, MA; Ruiz-Pérez, R; Ruiz-Fresneda, C; Jiménez-Contreras, E. Differences in Global Scientific Production Between New mRNA and Conventional Vaccines Against COVID-19. Environmental Science and Pollution Research 2022, 29, 57054–57066. [Google Scholar] [CrossRef] [PubMed]
- Pollard, AJ; Bijker, EM. A guide to vaccinology: from basic principles to new developments. Nat Rev Immunol 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y-S; Kumari, M; Chen, G-H; Hong, M-H; Yuan, JP-Y; Tsai, J-L; et al. mRNA-based vaccines and therapeutics: an in-depth survey of current and upcoming clinical applications. J Biomed Sci 2023, 30, 84. [Google Scholar] [CrossRef]
- Liu, Y; Ye, Q. Safety and Efficacy of the Common Vaccines against COVID-19. Vaccines (Basel) 2022, 10, 513. [Google Scholar] [CrossRef] [PubMed]
- Patone, M; Mei, XW; Handunnetthi, L; Dixon, S; Zaccardi, F; Shankar-Hari, M; et al. Risk of Myocarditis After Sequential Doses of COVID-19 Vaccine and SARS-CoV-2 Infection by Age and Sex. Circulation 2022, 146, 743–754. [Google Scholar] [CrossRef]
- Xu, S; Yang, K; Li, R; Zhang, L. mRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. Int J Mol Sci 2020, 21, 6582. [Google Scholar] [CrossRef]
- Echaide, M; Chocarro de Erauso, L; Bocanegra, A; Blanco, E; Kochan, G; Escors, D. mRNA Vaccines against SARS-CoV-2: Advantages and Caveats. Int J Mol Sci 2023, 24, 5944. [Google Scholar] [CrossRef]
- Wang, J; Ding, Y; Chong, K; Cui, M; Cao, Z; Tang, C; et al. Recent Advances in Lipid Nanoparticles and Their Safety Concerns for mRNA Delivery. Vaccines (Basel) 2024, 12, 1148. [Google Scholar] [CrossRef]
- Webb, C; Ip, S; Bathula, N V.; Popova, P; Soriano, SK V.; Ly, HH; et al. Current Status and Future Perspectives on MRNA Drug Manufacturing. Mol Pharm 2022, 19, 1047–1058. [Google Scholar] [CrossRef]
- SeyedAlinaghi, S; Karimi, A; Pashaei, Z; Afzalian, A; Mirzapour, P; Ghorbanzadeh, K; et al. Safety and Adverse Events Related to COVID-19 mRNA Vaccines; a Systematic Review. Arch Acad Emerg Med 2022, 10, e41. [Google Scholar] [CrossRef]
- Feddema, JJ; Fernald, KDS; Schikan, HGCP; van de Burgwal, LHM. Upscaling vaccine manufacturing capacity - key bottlenecks and lessons learned. Vaccine 2023, 41, 4359–4368. [Google Scholar] [CrossRef]
- Iqbal, SM; Rosen, AM; Edwards, D; Bolio, A; Larson, HJ; Servin, M; et al. Opportunities and challenges to implementing mRNA-based vaccines and medicines: lessons from COVID-19. Front Public Health 2024, 12. [Google Scholar] [CrossRef]
- Seo, SH; Song, MK. Advancements and challenges in next-generation mRNA vaccine manufacturing systems. Clin Exp Vaccine Res 2025, 14, 299. [Google Scholar] [CrossRef] [PubMed]
- Hengelbrock, A; Schmidt, A; Helgers, H; Vetter, FL; Strube, J. Scalable mRNA Machine for Regulatory Approval of Variable Scale between 1000 Clinical Doses to 10 Million Manufacturing Scale Doses. Processes 2023, 11, 745. [Google Scholar] [CrossRef]
- Shepherd, SJ; Han, X; Mukalel, AJ; El-Mayta, R; Thatte, AS; Wu, J; et al. Throughput-scalable manufacturing of SARS-CoV-2 mRNA lipid nanoparticle vaccines. Proceedings of the National Academy of Sciences 2023, 120. [Google Scholar] [CrossRef] [PubMed]
- Whitley, J; Zwolinski, C; Denis, C; Maughan, M; Hayles, L; Clarke, D; et al. Development of mRNA manufacturing for vaccines and therapeutics: mRNA platform requirements and development of a scalable production process to support early phase clinical trials. Translational Research 2022, 242, 38–55. [Google Scholar] [CrossRef]
- Kumar, S; Basu, M; Chakraborty, D; Ghosh, P; Ghosh, MK. Unraveling COVID-19 Diagnostics: A Roadmap for Future Pandemic. Nature Cell and Science 2024, 000, 000–000. [Google Scholar] [CrossRef]
- Bouazzaoui, A; Abdellatif, AAH; Al-Allaf, FA; Bogari, NM; Al-Dehlawi, S; Qari, SH. Strategies for Vaccination: Conventional Vaccine Approaches Versus New-Generation Strategies in Combination with Adjuvants. Pharmaceutics 2021, 13, 140. [Google Scholar] [CrossRef]
- Gote, V; Bolla, PK; Kommineni, N; Butreddy, A; Nukala, PK; Palakurthi, SS; et al. A Comprehensive Review of mRNA Vaccines. Int J Mol Sci 2023, 24, 2700. [Google Scholar] [CrossRef]
- Bosch, I; de Puig, H; Hiley, M; Carré-Camps, M; Perdomo-Celis, F; Narváez, CF; et al. Rapid antigen tests for dengue virus serotypes and Zika virus in patient serum. Sci Transl Med 2017, 9. [Google Scholar] [CrossRef]
- Shaffaf, T; Ghafar-Zadeh, E. COVID-19 Diagnostic Strategies Part II: Protein-Based Technologies. Bioengineering 2021, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Aoki, MN; de Oliveira Coelho, B; Góes, LGB; Minoprio, P; Durigon, EL; Morello, LG; et al. Colorimetric RT-LAMP SARS-CoV-2 diagnostic sensitivity relies on color interpretation and viral load. Sci Rep 2021, 11, 9026. [Google Scholar] [CrossRef] [PubMed]
- Song, X; Coulter, FJ; Yang, M; Smith, JL; Tafesse, FG; Messer, WB; et al. A lyophilized colorimetric RT-LAMP test kit for rapid, low-cost, at-home molecular testing of SARS-CoV-2 and other pathogens. Sci Rep 2022, 12, 7043. [Google Scholar] [CrossRef] [PubMed]
- Atmar, RL; Ramani, S. Immunologic Detection and Characterization. Viral Infections of Humans; Springer US: New York, NY, 2022; pp. 1–30. [Google Scholar] [CrossRef]
- Najafabadi, ZY; Fanuel, S; Falak, R; Kaboli, S; Kardar, GA. The Trend of CRISPR-Based Technologies in COVID-19 Disease: Beyond Genome Editing. Mol Biotechnol 2023, 65, 146–161. [Google Scholar] [CrossRef]
- Li, X; Qi, J; Wang, J; Hu, W; Zhou, W; Wang, Y; et al. Nanoparticle technology for mRNA: Delivery strategy, clinical application and developmental landscape. Theranostics 2024, 14, 738–760. [Google Scholar] [CrossRef]
- Jones, I; Roy, P. Sputnik V COVID-19 vaccine candidate appears safe and effective. The Lancet 2021, 397, 642–643. [Google Scholar] [CrossRef] [PubMed]
- Pramod, S; Govindan, D; Ramasubramani, P; Kar, SS; Aggarwal, R; Manoharan, N; et al. Effectiveness of Covishield vaccine in preventing Covid-19 – A test-negative case-control study. Vaccine 2022, 40, 3294–3297. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Strategies Employed by the Biomanufacturing Industry to Combat COVID-19. The multi-faceted approach taken by the biomanufacturing industry to control the COVID-19 pandemic. Key strategies depicted include the development of diagnostic tests, the production and distribution of vaccines, and the manufacture of hand sanitizers and PPE such as masks. These efforts highlight the industry’s critical role in mitigating the spread of the virus and protecting public health.
Figure 1.
Strategies Employed by the Biomanufacturing Industry to Combat COVID-19. The multi-faceted approach taken by the biomanufacturing industry to control the COVID-19 pandemic. Key strategies depicted include the development of diagnostic tests, the production and distribution of vaccines, and the manufacture of hand sanitizers and PPE such as masks. These efforts highlight the industry’s critical role in mitigating the spread of the virus and protecting public health.
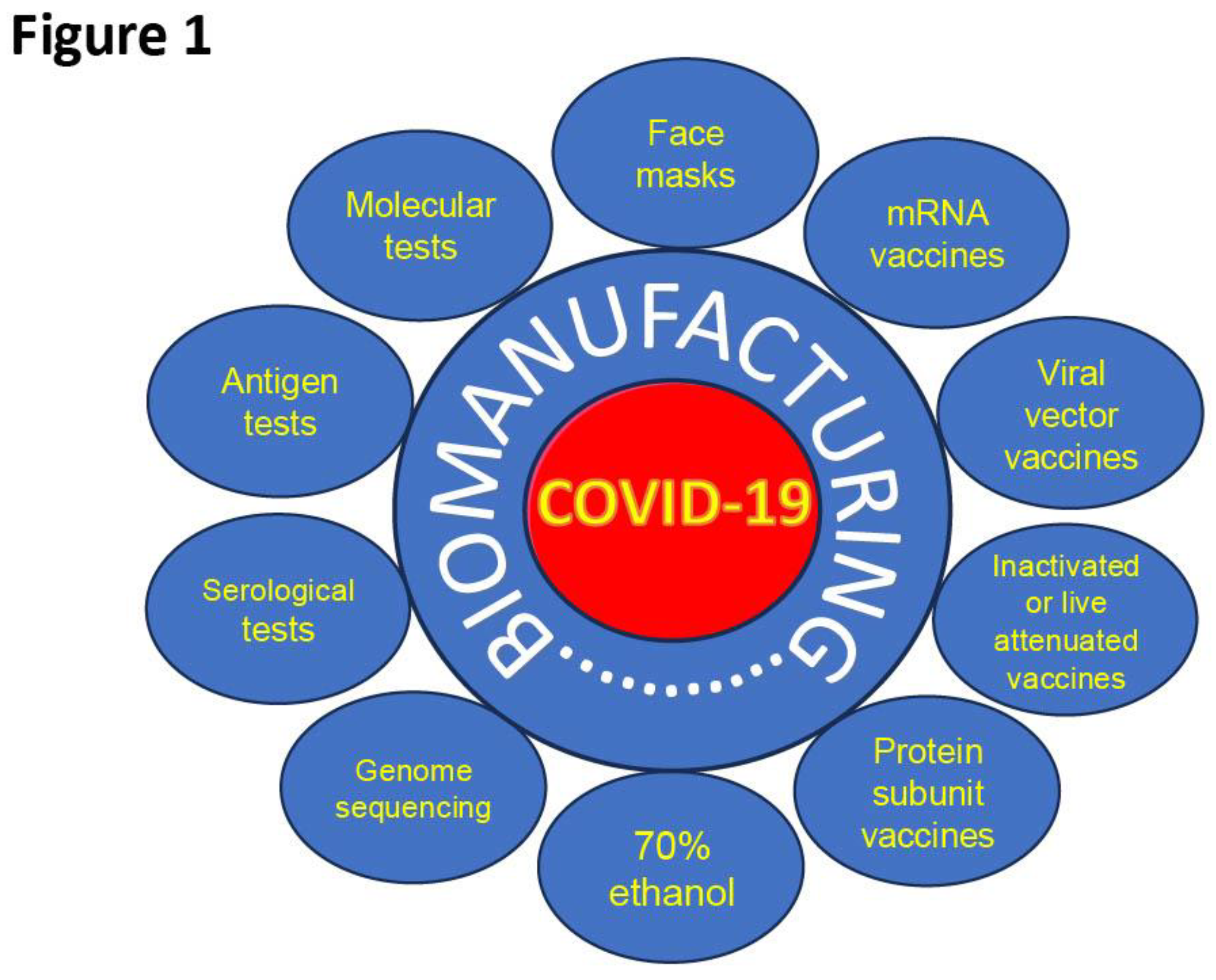
Figure 2.
Development strategies for mRNA-LNP COVID-19 vaccine. The key stages of COVID-19 vaccine development. A. Collection of nasopharyngeal samples from COVID-19 patients. B. Genome sequencing of SARS-CoV-2 obtained from patient samples. C. Design of mRNA sequences that encode the spike protein. D. Encapsulation of the synthetic mRNA in lipid nanoparticles (LNPs). E. Clinical trials and regulatory review and approval of the vaccine. F. Large-scale production and distribution of the mRNA-LNP vaccine.
Figure 2.
Development strategies for mRNA-LNP COVID-19 vaccine. The key stages of COVID-19 vaccine development. A. Collection of nasopharyngeal samples from COVID-19 patients. B. Genome sequencing of SARS-CoV-2 obtained from patient samples. C. Design of mRNA sequences that encode the spike protein. D. Encapsulation of the synthetic mRNA in lipid nanoparticles (LNPs). E. Clinical trials and regulatory review and approval of the vaccine. F. Large-scale production and distribution of the mRNA-LNP vaccine.
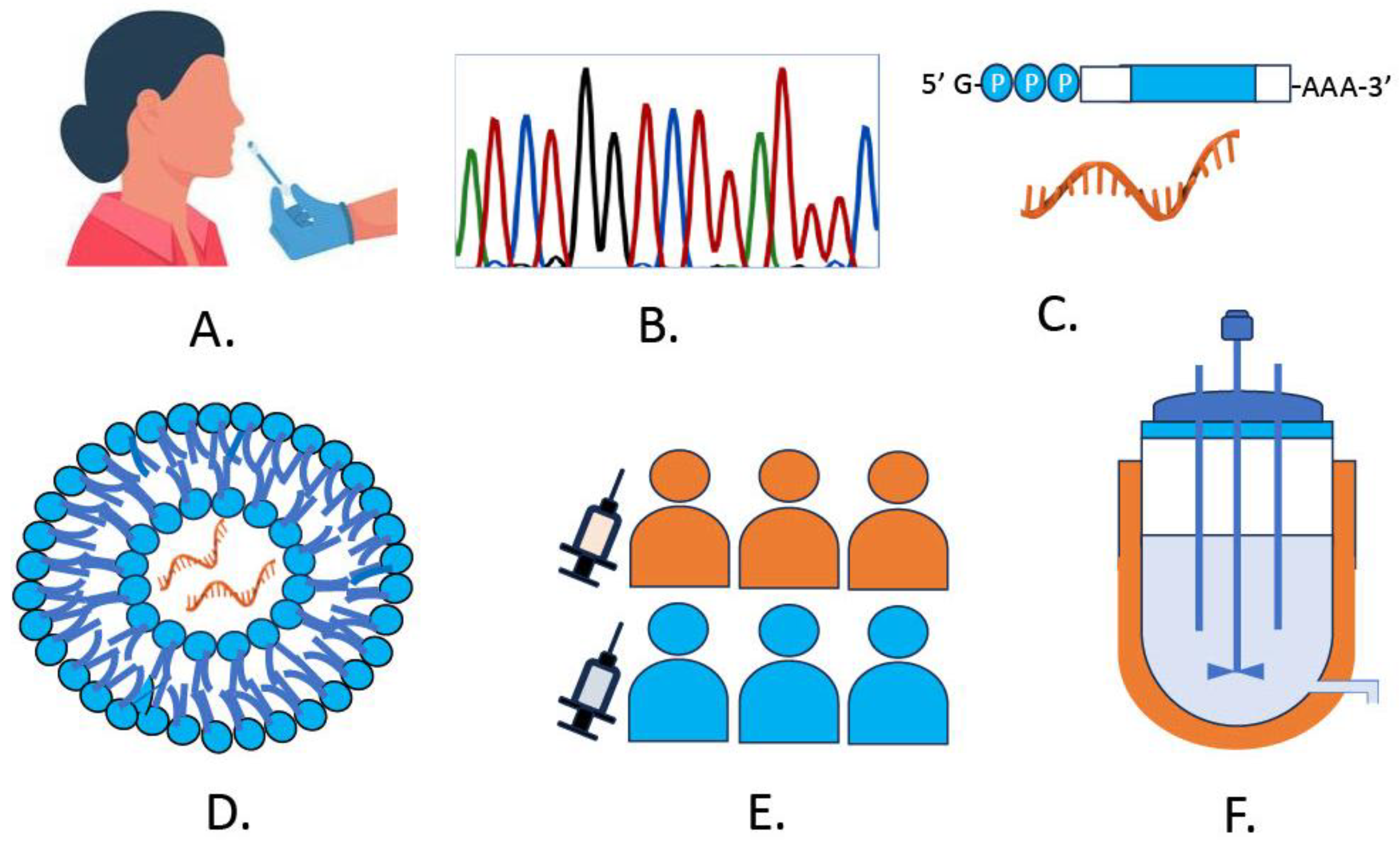

Table 1.
Nanotechnology based products for the prevention and detection of SARS-CoV-2. The application of nanotechnology in combating SARS-CoV-2 featuring innovative products developed for both prevention and detection of the virus.
Table 1.
Nanotechnology based products for the prevention and detection of SARS-CoV-2. The application of nanotechnology in combating SARS-CoV-2 featuring innovative products developed for both prevention and detection of the virus.
| Name of company | Country | Product | Function | References |
|---|---|---|---|---|
| GenScript USA Inc. | USA | Rapid antigen test | Gold-coated antibodies produce colorimetric signal on paper in presence of target serum | [191,192] |
| Abbot Diagnostics | USA | RT-LAMP | Assay amplifies target regions of the SARS-CoV-2 RdRp and N gene with the same fluorophore within a single well. | [193,194] |
| Bio-Rad Laboratories DRG Diagnostics GmbH Euroimmun US Inc. |
USA | ELISA | Viral antigens are immobilized on plate to catch antibodies from patient’s antibodies | [192,195] |
| Mammoth Biosciences/ Sherlock Biosciences | USA | CRISPR | Rapid detection of SARS-CoV-2 | [196] |
| Identify Sensors | USA | Check 4 | Rapid detection of SARS-CoV-2 |
www.statnano.com |
| NanoEnTek | South Korea | Frend COVID-19 Ag | Rapid detection of SARS-CoV-2 | |
| Zepto Life Technology | USA | COVID-19 Test | Rapid detection of SARS-CoV-2 | |
| Mologic Ltd | UK | COVIS-19 test | Rapid detection of SARS-CoV-2 | |
| Lucence Diagnostics Pte Ltd. | Singapore | SAFER Sample Kit | Rapid detection of SARS-CoV-2 | |
| Graphene Leaders Canada | Canada | GLCM Biosensor test | Rapid detection of SARS-CoV-2 | |
| Nanoshel | India | Hand Sanitizer | Anti-viral | |
| Curran Biotech | USA | Filter | Capture coating | |
| Medicevo Corporation | India | Face mask with graphene | Virus control and personal protection | |
| Flextrapower | USA | Graphene mask | Virus control and personal protection | |
| Integricote | USA | Respiratory mask | Virus control and personal protection | |
| Master Dynamic Limited | China | Diamond face mask | Virus removal | |
| Yamashin Filter Corp | Japan | Nanofiber mask | Virus removal | |
| MVX Prime Ltd. | UK | Nano mask | Anti-viral Anti-bacterial |
Table 2.
Recent vaccines developed for the immunity against COVID-19. Various vaccines recently developed to provide immunity against COVID-19. It includes key vaccines such as those based on mRNA technology, vector-based vaccines, and inactivated virus vaccines.
Table 2.
Recent vaccines developed for the immunity against COVID-19. Various vaccines recently developed to provide immunity against COVID-19. It includes key vaccines such as those based on mRNA technology, vector-based vaccines, and inactivated virus vaccines.
| Name of the company | Name of the vaccine | Adult dosage (www.who.int) |
Ingredients |
|---|---|---|---|
| Biontech/Pfizer (USA) |
BNT162b2 | 2 doses of 30µg each Intramuscularly |
Nucleoside-modified mRNA encoding the viral spike (S) glycoprotein of SARS-CoV-2 [145], DSPC, cholesterol, ALC-0315, ALC-0159 [197] |
| Moderna (USA) |
mRNA-1273 | 2 doses of 50µg each Intramuscularly |
Nucleoside-modified mRNA encoding the viral spike (S) glycoprotein , DSPC, cholesterol, SM102, DMG-PEG2000 [197] |
| Novavax (India) |
NVX-CoV2337 | 2 doses of 0.5 ml each Intramuscularly |
SARS-CoV-2 recombinant spike protein [135] |
| J&J/Janssen (USA) |
Ad26.CoV2.S | 1 dose of 0.5 ml each Intramuscularly |
Recombinant, replication-incompetent Ad26 vector, encoding a stabilized variant of the SARS-CoV-2 Spike (S) protein. [121] |
| Sinovac (China) |
Coronavac | 2 doses of 0.5 ml each Intramuscularly |
Aluminum hydroxide–adjuvanted, inactivated whole-virus vaccine [133] |
| Gamaleya (Russia) |
Sputnik V | 2 doses of 0.5 ml each Intramuscularly |
Recombinant, replication-incompetent Ad26 vector, encoding a stabilized variant of the SARS-CoV-2 Spike (S) protein. [198] |
| Bharat Biotech (India) |
Covaxin | 2 doses of 0.5 ml each Intramuscularly |
Alhydroxiquim-II–adjuvanted, inactivated whole-virus vaccine [133] |
| Astrazenica (UK) |
Covishield | 2 doses of 0.5 ml each Intramuscularly |
Recombinant, replication-incompetent ChAdOx1 vector, encoding a stabilized variant of the SARS-CoV-2 Spike (S) protein. [199] |
| Sinopharm (China) |
BBIBP-CorV | 2 doses of 0.5 ml each Intramuscularly |
Aluminum hydroxide–adjuvanted, inactivated whole-virus vaccine [133] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



