Submitted:
26 December 2025
Posted:
30 December 2025
You are already at the latest version
Abstract
UVB rays cause sunburn to lead to various skin damages, despite their necessity for vitamin D synthesis. The outermost layer of the epidermis is the cornified envelope (CE), characterized by a lipid-protein cross-linked complex structure. UVB radiation disrupted the CE structure, leading to skin barrier dysfunction and a decrease in epidermal moisture. Phycocyanin (PC) and its enzyme-digested product (EDPC) showed repair-enhancing action by their oral post-administration subsequent to UVB pre-radiation in hairless mice. The repair-promoting effect resulted from transcriptional events mediated by their administrations. DNA microarray analyses revealed a unique molecular mechanism underlying this effect. Several genes, including Lce, Sprr, Krt, Csta, and Degs, that encoded CE components of proteins, protease inhibitors, and/or enzymes, exhibited reasonable upregulation or downregulation in expression. Post-administration following UVB pre-radiation caused additionally/amelioratively up- or down-regulated expression of these genes.These findings indicate that PC, especially EDPC, had the repair-enhancing effect on UVB-induced skin barrier dysfunction when applied post-UVB exposure.
Keywords:
keratinocytes
; functional foods
; photo-aging
; anti-phototoxic compounds
1. Introduction
A burn is an injury to skin, caused by heat, electricity, chemicals, or radiation. The most common type of radiation burn is a sunburn resulting from exposure to ultraviolet (UV) rays. Solar UV radiation is divided into three categories based on wavelength, namely UVA (320-400 nm), UVB (280-320 nm) and UVC (100-280 nm) [1]. UVC rays are dispersed and reduced by the ozone layer and do not reach the ground. About 1%–10% of UVB rays, also known as sunburn rays, and 90%–99% of UVA rays reach the earth's surface. The skin epidermis functions as a barrier to protect the body from the external elements, particularly phototoxic UV rays [2]. Nonetheless, chronic exposure or intermittent overexposure of human skin to UVB rays leads to various skin dysfunctions, sunburn, skin photo-aging, and dermal pathologies. Sunscreen or anti-phototoxic foods/drinks are effective ways, apart from sun avoidance, to prevent phototoxic damage, such as sunburn, skin aging, collagen degradation-wrinkle formation, or pigmentation caused by UV radiation [3,4].
A group of blue-green algae of the genus Arthrospira (formerly Spirulina) are a rich source of pro-health metabolites): proteins, carbohydrates, and fats used in pharmaceutical and cosmetic formulations. Among these a photosynthetic protein phycocyanin (PC), which is blue, as its name suggests. It has been increasingly noticed that the consumption of Spirulina or PC extracts is associated with a variety of beneficial effects, such as wound healings, antimicrobial, antioxidant, anti-inflammatory, anti-melanogenic, immunomodulatory [3]. PC and its derivatives, like enzyme-digested product (EDPC), were known to have neuroprotective effects, including amelioration of Alzheimer’s disease [4,5]. PC and its derivatives have also been reported to protect against UVB-induced skin damage [6,7,8]. While similar protective effects have been observed in cosmetic applications on local skin [9], little information on these protective actions to skin via oral administration of PC and its derivatives has been accumulated except for a clinical report [10]. Additionally, there was no report concerning the skin effect of PC and its derivatives administered after UVB radiation.
In this study we aimed to examine the repair-enhancing effects of orally administered PC and EDPC subsequent to epidermal barrier failure induced by UVB pre-radiation in hairless mice. We also aimed to elucidate the mechanism underlying the PC and its derivatives-mediated ameliorating action using DNA microarray followed by genome informatics, which revealed previously unknown molecular events occurring in the mice's UVB-exposed skin.
2. Materials and Methods
2.1. Reagents
The following materials were used in this study: the RNAiso Plus, PrimeScript™ Reverse Transcriptase, Recombinant RNase Inhibitor, TBGreen. Premix Ex Taq™ (Tli RNaseH Plus), RR420 (Takara Bio Inc., Kyoto, Japan), RNeasy MinElute Cleanup Kit (QIAGEN, Tokyo, Japan), dNTP Mix and Oligo (dT)15 Primer (Promega, Tokyo, Japan), Low Input Quick Amp Labeling Kit, RNA6000 Nano Kit, Agilent Whole Human Genome DNA Microarray 4×44K v2, and Agilent Gene Expression Hybridization Kit (Agilent Technologies, Santa Clara, CA, USA) were also used in this study.
2.2. Animals
Twenty-four male HR-1 hairless mice (6 weeks old) were purchased from SLC, Inc. (Hamamatsu, Japan). The animal examinations were performed in accordance with the regulations set forth by the Institutional Animal Care and Use Committee of Intelligence and Technology Lab, Inc. (ITL), adhering to the standards for the Proper Conduct of Animal Experiments. The Animal Care Committee of ITL sanctioned these examinations, designating them with the code AE2023014 and granting approval on 26 September 2023.
2.3. Phycocyanin and Its Enzyme-Digested Products
Phycocyanin (PC), was purified from water extract of Spirulina, a dried product of Arthrospira platensis, which was obtained from DIC Co., Ltd. (Tokyo, Japan), by filtration followed by ultrafiltration. The enzyme-digested PC (EDPC) employed was a proteolytic product of PC generated by digestion with SD-NY10® from Amano Enzyme Co., Ltd. (Nagoya, Japan), in which no less than 58% were low-molecular-weight components with a molecular weight less than 6 kDa.
2.4. UVB Exposure
The UVB radiation device used was Dermaray®-200 (Termo, Tokyo, Japan). The UVB radiation at the mouse's back was measured using a UV illuminance meter (Dermaray UV Meter, Termo, Tokyo, Japan). The distance between the UV lamp and the animal’s back was adjusted before radiation to ensure a mean UVB radiance of 0.3 mW/cm². The UVB radiation was performed daily for four sequential days with each radiation duration of four minutes.
2.5. PC and EDPC Administration
HR-1 hairless mice were divided into 4 groups: control (Ctr), UVB radiation (UVB), UVB radiation followed by oral administration of PC (UVB+PC), and EDPC (UVB+EDPC). Group Ctr was given orally with PBS for 7 days without exposure to UVB rays. Group UVB was orally received with PBS for 7 days following a four-day pre-exposure to UVB rays. UVB+PC was orally administered for 7 days, with PC dosed at 1000 mg/kg after four-day pre-exposure to UVB radiation, and UVB+EDPC was administered with EDPC. During animal studies body weights of mice were 33 - 36 g.
2.6. Skin Moisture Measurement
The moisture content of the dorsal skin region was assessed thrice utilizing the Corneometer CM825 (Courage & Khazaka Electronic, Cologne, Germany), and the mean values were calculated and presented in arbitrary units (a.u.) ranging from 0 (no moisture) to 120 (maximum moisture).
2.7. Collagen Assay
A half of the sample from the dorsal skin region was subjected to a total collagen assay that was performed by means of the hydroxyproline method using HCl-hydrolyzed samples with a total collagen assay kit (Quick Zyme Biosciences, Leiden, The Netherlands) under the conditions mentioned in the kit manual.
2.8. RNA Extraction and Isolation
Total RNA was extracted from the remaining half of the dorsal skin region obtained from four groups—Ctr, UVB, UVB+PC, and UVB+EDPC—using the acid guanidine thiocyanate-phenol-chloroform method and silica membrane column-based purification [14]. Briefly, skin samples were homogenized in RNAiso Plus (Takara Bio Inc.), and the total RNA was isolated. The RNA was treated with DNase (QIAGEN, USA) in the aqueous phase and purified using the RNeasy MinElute Cleanup Kit (QIAGEN) according to the manufacturer’s instructions. The quantity and purity of RNA were evaluated at 230, 260, 280, and 320 nm using an Ultrospec 2000 spectrometer (GE Healthcare Bio-Sciences AB, Uppsala, Sweden). For DNA microarray analyses, RNA integrity number (RIN) was determined using an Agilent 2100 Bioanalyzer (Agilent Technologies Japan Ltd., Tokyo, Japan). Only high-quality RNA samples, which were defined by an A260/A230 of 1.5, A260/A280 of 1.8, and RIN 6.0, were used in the microarray analysis.
2.9. DNA Microarray Analyses
DNA microarray analyses were performed using the RNA samples from four groups-Ctr, UVB, UVB+PC,and UVB+EDPC. After cDNA synthesis, Cy3-labeled cRNA was synthesized and purified using the Low Input Quick Amp Labeling Kit (Agilent) according to the manufacturer’s instructions. Notably, reverse transcription was conducted using a T7 promoter-oligo(dT) primer. Absorbance was measured at 260, 280, 320, and 550 nm, and it was verified that the labeled cRNA had incorporated >6 pmol/mg of Cy3-CTP. Labeled cRNA was fragmented using the Gene Expression Hybridization Kit (Agilent) and applied to Whole Mouse Genome Array Ver2.0 slides (Agilent). After hybridization at 65 ℃ for 17 h, the slides were washed with Gene Expression Wash Buffers 1 and 2 (Agilent) according to the manufacturer’s instructions. The slides were scanned using a GenePix 4000B scanner (Molecular Devices Japan K.K., Tokyo, Japan). Scanned images were digitalized and normalized using GenePix Pro software (Molecular Devices Japan K.K., Tokyo, Japan).
2.10. Bioinformatics Analysis of Microarray Data
By comparing the control group vs. UVB, UVB+PC, UVB+EDPC groups, a list of genes that were >1.5-fold upregulated or <0.67-fold downregulated in the UVB, UVB+PC, and UVB+EDPC groups or the UVB+PC or UVB+EDPC group were generated. We referred to these genes as UVB-associated genes. The top 50 UVB-associated genes were annotated, and references were searched using NCBI databases, Google, and related information. The skin-related functions of annotated genes were deduced regarding the possible mechanisms of action of the PC or EDPC.
Time points of the examinations and samplings were shown in Figure 1.
3. Results
3.1.. Skin Moisture Level
The epidermal moisture levels of four groups-Ctr, UVB, UVB+PC, and UVB+EDPC-were assessed using the Corneometer CM825. Figure 2A shows the moisture levels of these groups at three time points: prior to UVB radiation, subsequent to four days of UVB radiation, and seven days post-administration of PC or EDPC following UV irradiation. UVB exposure caused a significant reduction in epidermal moisture levels (Figure 2B). Oral administration of PC and PCED for seven days resulted in the restoration of moisture loss induced by UVB.
These suggest that the administration of PC or PCED ameliorate UVB-induced epidermal moisture decrease. This repair-promoting effect on skin barrier dysfunction leading to dehydration is accompanied by transcriptional events and a change in the expression of functional genes, as mentioned below.
3.2. Skin Collagen Amount
Little difference in total collagen levels across skin samples from four groups-Ctr, UVB, UVB+PC, and UVB+EDPC-was found (data not shown), suggesting that UVB radiation did not influence skin collagen alterations. This has been also supported by comparative assessments of DNA microarray data. The expression levels of 34 collagen-related genes remained constant, ranging from 1.2 to 0.85-fold, among skin samples from four groups (data not shown). The expression levels of 44 collagen digesting protease (matrix metallopeptidase) genes were also constant (data not shown). From these, alteration in skin collagen amount by UVB radiation did not occur in this study.
3.3. DNA Microarray Analyses and Bioinformatics
We deduced the mechanism underlying the PC and/or EDPC-mediated repair-promoting effect of UV-induced skin barrier dysfunction in UVB-radiated mice. Therefore, we performed DNA microarray analyses using skin samples from four groups: Ctr, UVB, UVB+PC, and UVB+EDPC. We also clarified the difference in transcriptional events in the PC or EDPC groups.
3.3.1. Additionally Upregulated PC/EDPC-Related Genes
We compared the gene expression profiles of the two groups, Ctr and UVB, and identified xx genes that were upregulated by more than 1.5-fold in UVB (data not shown). In the subsequent comparison of the upregulated gene expression profiles between UVB and UVB+PC or UVB+EDPC, we aimed to identify genes associated with UV-mediated skin response, damage, inflammation, dehydration, and/or repair, which were additionally upregulated in UVB+PC or UVB+EDPC. As shown in Table 1, we identified 20 genes that were additionally upregulated by more than 1.5-fold in the UVB+PC and 27 genes in the UVB+EDPC. These genes were referred to as additionally upregulated PC/EDPC-related genes.
Table 1 lists the additionally upregulated PC/EDPC-related genes that are Lce gene family (Lce3b, Lce3c, Lce3d, Lce3e, Lce3f, and Lce6a; late cornified envelope proteins), Sprr gene family (Sprr1b, Sprr2a2, Sprr2d, and Sprr2h; small proline-rich proteins), Rptn (repetin; cornified envelope component), Degs2(delta 4-desaturase, sphingolipid 2), keratin gene family (Krt6b and Krt16; Keratins), Csta gene family (Stfa2, Stfa2l1, and Gm5416; A, cysteine-protease inhibitor), Klk gene subfamily (Klk9 and Klk13; kallikrein related peptidase, serine-protease), Serpin gene family (Serpina9 and Serpinb12; serpin family a member 9 and serpin family B member 12, respectively, serine-protease inhibitor), Spink7 (serine peptidase inhibitor Kazal type 7), Fetub (fetuin-B, protease inhibitor), Pitx1(paired-like homeodomain transcription factor 1), Aqp5 (aquaporin 5), Il1f6 (interleukin 36 alpha), and Ereg (epiregulin).
3.3.2. Amelioratively Downregulated EDPC-Related Genes.
As shown in Table 2, among upregulated genes in UVB, we identified one gene, Chi3l3 (chitinase-like protein 3, also known as the Ym1 gene), that was downregulated by less than 0.67-fold in UVB+EDPC but not in UVB+PC.The downregulation of Chi3l3 gene expression in UVB+EDPC resulted from EDPC administration to UVB-radiated mice (UVB). This downregulation is related to the ameliorative action of EDPC on UBV-induced skin barrier dysfunction. Thus, Chi3l3 was referred to as an amelioratively downregulated EDPC-related gene.
3.3.3. Amelioratively Upregulated PC/EDPC-Related Genes
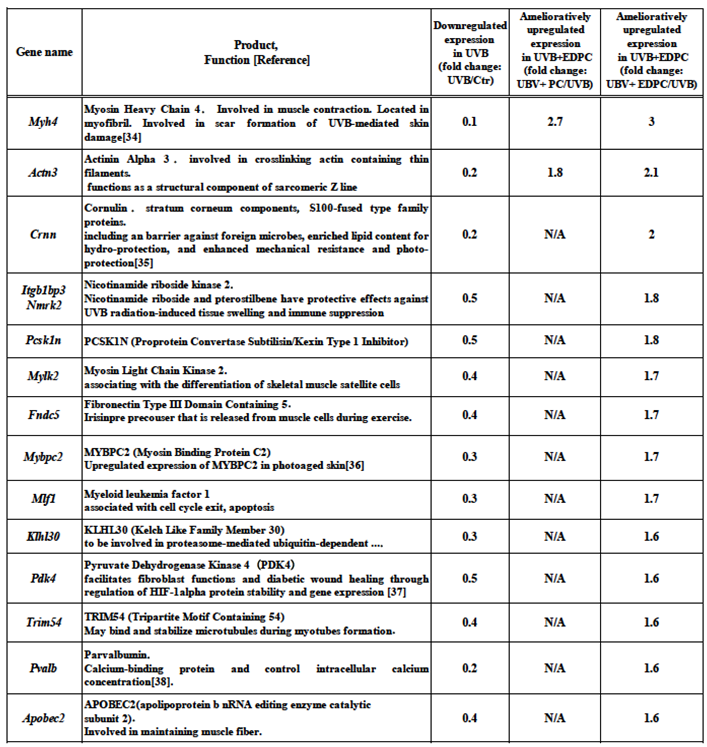
Table 3 shows fourteen genes whose expressions were upregulated by more than 1.5-fold in the UVB+EDPC among downregulated genes in UVB. This upregulation is associated with the ameliorative effect of EDPC on UVB-induced skin reactions, including damage. Thus, these genes were referred to as amelioratively upregulated PC/EDPC-related genes. Table 3 register the amelioratively upregulated PC-related genes that are just only two, Myh4, Actn3. On the other hand, the amelioratively upregulated EDPC-related genes were twelve genes. These genes were Myh4 (myosin heavy chain 4), Actn3(actinin alpha 3), Crnn (cornulin; S100-fused type family proteins), Itgb1bp3/Nmrk2 (nicotinamide riboside kinase 2), Pcsk1n (proprotein convertase subtilisin/kexin type 1 inhibitor), Mylk2 (myosin light chain kinase 2), Fndc5 (fibronectin type III domain containing 5), Mybpc2 (myosin binding protein c2), Mlf1 (myeloid leukemia factor 1), Klhl30 (kelch like family member 30), Pdk4 (pyruvate dehydrogenase kinase 4), Trim54 (tripartite motif containing 54), Pvalb (parvalbumin), and Apobec2 (apolipoprotein b nRNA editing enzyme catalytic subunit 2).
4. Discussion
4.1. UVB Rays
Sunlight (UVB) is essential to produce a fat soluble-vitamin, vitamin D, in the skin. Vitamin D participates in musculoskeletal health and reduces the risk of bone fractures []. A beneficial source of vitamin D is ultraviolet (UVB) radiation from the sun. However, a balance is needed between enough sun for adequate vitamin D and too much sun (sunburn), with increased risk of skin damage and cancer [10]. As less than 10% of UVB rays, also known as sunburn rays, reach the earth's surface, our skin epidermis was always exposed to the sunburn rays. The skin epidermis functions as a barrier to protect the body from the external elements, particularly phototoxic UV rays [11]. Nonetheless, chronic exposure or intermittent overexposure of human skin to UVB rays leads to various skin dysfunctions, such as sunburn, skin photo-aging, and dermal pathologies.
4.2. Epidermis: Its Structure and Function
As shown in Figure 3, the epidermis was composed of five layers, arranged from the inner to the outside: basal lamina, stratum basale, stratum spinosum, stratu granulosum, and stratum corneum. Stratum corneum is the outermost layer of the epidermis and mostly composed of corneocytes, which are terminally differentiated keratinocyte without nuclei and cytoplasmic organelles. The plasma membrane of the corneocyte periphery mostly was replaced to the cornified envelope (CE) that is a lipid-protein cross-linked structure. CE components, including the late cornified envelope (LCE) proteins, was reported to possess broad-spectrum antimicrobial activity [12]. Thus, CE and its components are known as multifunctional epidermal barrier not only to protects underlying tissue from UV rays, dehydration, infection, and mechanical stress, but also to possess lipidorganizing, antioxidant, and antimicrobial properties [13].
The epidermis has five layers: basal lamina, stratum basale, stratum spinosum, stratum granulosum, and stratum corneum. The cornified envelope (CE) is the outermost layer of the stratum corneum and is covered with a lipid-protein cross-linked structure composed of several proteins and ceramide.
4.3. Phycocyanin (PC), Enzyme-Digested Product (EDPC), and Skin Moisture
The consumption of Spirulina or phycocyanin (PC) is associated with a variety of beneficial effects, such as wound healing, antimicrobial, antioxidant, anti-inflammatory, anti-melanogenic, and immunomodulatory effects [3]. PC and its derivatives were known to protect against UVB-induced skin damage in cosmetic applications [6,7,8,9]. In a previous paper, we reported that orally administered PC and its enzyme-digested product (EDPC) had neuroprotective effects, including amelioration of Alzheimer’s disease [5]. To our knowledge, there is little information concerning a repair-promoting effect of orally administered PC and EDPC on UVB-induced skin barrier dysfunction.
By oral post-administration, both PC (UVB+PC) and EDPC (UVB+EDPC) alleviated skin moisture levels in hairless mice pre-radiated with UVB. EDPC (UVB+EDPC) had more of an alleviating effect than PC (UVB+PC) (Figure 2A,B). The potent action of EDPC compared to PC was also observed in the following results of gene expression analyses. About this difference it is thought that EDPC may be more easily absorbed in the gastrointestinal (GI) tract than PC and thus reach higher levels in target sites of the body, including the skin. This assumption was based on the fact that EDPC is a protease-digested product consisting of no less than 58% of low-molecular-weight components with a molecular weight less than 6 kDa.
4.4. Skin Collagen
Although decrease in skin collagen was reported in previous paper using UVB-exposed mice [14], we observed little difference among four groups, Ctr, UVB, UVB+PC and UVB+EDPC, indicating no effect of UVB radiation to digestion and/or synthesis of skin collagen. This was also supported by the DNA microarray data, little up-or down-regulated expression both of collagen- and its digestive proteas-related genes.
4.5. Change in Expression of Functional Genes, DNA Microarray Analyses, and Informatics
An efficient method for studying the expression levels of most genes in an organism’s functional genome, particularly the transcriptional product (mRNA), is through DNA microarray techniques, which provide high-throughput analysis of transcripts/mRNAs. In our previous studies on sublingual vaccines [15,16], we elucidated novel molecular events within the immune system and their related responses using a DNA microarray approach. In the present study, we further studied the mechanism underlying the repair-promoting action of orally administered PC and EDPC in the skin of hairless mice pre-radiated with UVB.
Epidermal keratinocytes are primarily responsible to transcriptional events and alterations in functional gene expression, which are mediate by UVB radiation and subsequent administration with PC or EDPC. DNA microarray analyses facilitate the observation of these molecular events in epidermal keratinocytes.
4.5.1. Additionally Upregulated PC/EDPC-Related Genes
UVB exposure caused skin barrier dysfunction, such as a decrease in skin moisture (Figure 1A,B). This dysfunction was partly recovered by spontaneous repair of CE with the relative moisture level from 57.5% to 65.7%. During this spontaneous repair, there was a marked upregulation or downregulation of gene expression primarily in epidermal keratinocytes (Table 1, Table 2 and Table 3). Oral post-administration of PC or EDPC following UVB radiation resulted in significant recovery of the relative moisture level and was accompanied by further upregulated expression of genes related to CE and its relatives. Thus, these genes, which have more upregulated expression than UBV radiation alone, are referred to as additionally upregulated PC/EDPC-related genes (Table 1). Interestingly, most of additionally upregulated PC/EDPC-related genes encoded proteins and/or components associated with CE and its relatives as mentioned follows.
Lce genes cluster contains six Lce genes: Lce3b, Lce3c, Lce3d, Lce3e, Lce3f, and Lce6a. These genes encode late cornified envelope component proteins, specifically LCE3B, LCE3C, LCE3D, LCE3E, LCE3F, and LCE6A, which may have functions related to the epidermal terminal. LCE3B and LCE3C have antimicrobial activity [8,13]. Sprr gene cluster contains four Sprr genes: Sprr1b, Sprr2a2, Sprr2d, and Sprr2h, encoding small proline-rich proteins, SPRR1B, SPRR2A2, SPRR2D, and SPRR2H, of the cornified envelope component [18]. As the two gene families of Lce and Sprr, located on human chromosome 1q21 and encoding structural and regulatory proteins, LCEs and SPPRs, are crucial to keratinocyte differentiation and stratum corneum functions, excess gene expression of Lce and/or Sprr is reported to be a risk for a pathologic state, psoriasis [10,13]. Krt gene cluster is two genes, Krt6b and Krt1, encode keratin 6B and keratin 16, respectively, which protect the skin's barrier and were produced when the epidermis is damaged [20]. Csta gene cluster is consists of three genes, Stfa2, Stfa2l1, Gm5416, encoding Stefin A2, Stefin A2 like1, and cystatin A family member 3 proteins, respectively. They are cysteine-type proteases inhibitors and associate with keratinocyte differentiation. These inhibitors are located in several cellular components, including CE, and regulate UV-induced keratinocyte apoptosis [21,22]. Degs2 gene encodes delta 4-desaturase, sphingolipid 2 that is involved in the biosynthesis of phytosphingolipids, ceramides, for CE barrier function [23]. Rptn gene’s product is repetin, which belongs to the S100 family, known as skin-specific protein with a role in calcium regulation within the skin [24]. Spink7 gene encodes serine peptidase inhibitor Kazal type 7 that regulates kallikrein and its related serine-proreases, and has a pivotal role in epidermal homeostasis [25]. Fetub gene’s product is Fetuin-B protein, which has the role of a cysteine protease inhibitor and is involved in the process with a focus on the regulation of the PI3K/AKT signaling pathway [26]. Pitx1 gene encodes Paired-Like Homeodomain Transcription Factor 1 protein, associating with wound healing in the skin by increasing cellular migration of keratinocytes and stimulating granulation tissue formation [27].
The following seven genes were observed only in additionally upregulated EDPC-rated genes but not in PC-related genes. Klk9 and Klk13 genes’ products are kallikrein-related peptidase 9 and kallikrein-related peptidase, respectively. These serine peptidases are involved in the different steps of skin wound healing and regulated by their inhibitor, Kazal type 7 [25,28]. Serpina9 and Serpinb12 genes encode each of serpin family A member 9 and serpin family B member 12, which relate to the regulation of epidermal barrier function and the pathogenic state in psoriasis [29]. Aqp5 gene product is aquaporin 5 protein, which participates in water-glycerol transportation proteins, proliferation of keratinocytes, and skin aging [30]. Il1f6 gene encodes the interleukin 36 alpha protein, which is associated with skin inflammation [31]. Ereg gene encodes epiregulin protein, a ligand of the EGF receptor contributing to inflammation, wound healing, and tissue repair [32].
These 27 genes relate to wound healing, regeneration, or cell proliferation of CE and/or keratinocytes. UVB-radiated skin damage was partly recovered by spontaneous repair of CE and/or keratinocytes with upregulated expression of these functional genes.
The oral post-administration of PC or EDPC after UVB radiation resulted in enhanced recovery of skin function and hydration, surpassing spontaneous repair, along with increased expression of PC/EDPC-related genes. Consequently, PC, particularly EDPC, appears to exert a repair-promoting effect on skin compromised by UVB exposure.
4.5.2. Amelioratively Downregulated EDPC-Related Gene
Chi3l3 gene encodes the chitinase-like protein 3 (also known as Ym1). Chi3l3 expression in skin wound reported to be upregulated during the early stage and downregulated in the late-stage coinciding with wound repair [33]. Thus, EDPC had an ameliorative effect on UVB-mediated skin damage by downregulating the expression of the Chi3L3 gene, which was upregulated due to UVB exposure.
4.5.3. Amelioratively Upregulated PC/EDPC-Related Gene
While only two PC-related genes, Myh4 and Actn3, were elevated, twelve EDPC-related genes exhibited ameliorative upregulation (Table 3). The downregulated expression of Myh4, Crnn, Mybpc2, Pdk4, and Pvalb genes was characteristic in their repair-enhancing activities among the EDPC-related genes. Myh4 gene encodes myosin heavy chain 4 that is located in myofibril and plays a role in scar formation resulting from UVB-induced skin damage [34]. Crnn gene’s product is cornulin, a component of the stratum corneum and a member of the S100-fused type family of proteins. Cornulin was known as a barrier against foreign microbes, enriched lipid content for hydro-protection, and enhanced mechanical resistance and photo-protection [35]. The coding product of the Mybpc2 gene is myosin light chain kinase 2which is involved in the healing of photoaged skin through its elevated expression of [36]. Pdk4 gene encodes pyruvate dehydrogenase kinase 4, which enhances fibroblast functions and wound healing by regulating HIF-1alpha protein stability and gene expression [37]. Pvalb gene’s product is parvalbumin, which is calcium-binding protein that regulates intracellular calcium level in various cell types including keratinocytes [38].
5. Conclusion
PC, especially EDPC, demonstrated the repair-enhancing effect on UVB-induced skin barrier dysfunction when administered orally following UVB exposure. Their repair-enhancing action was based on transcriptional events in keratinocytes via up- or down-regulated expression of multiple genes induced by PC or EDPC administration. Consequently, PC or EDPC appears to be promising functional food or beverage for the recovery of skin damage caused by sunburn.
Author Contributions
Conceptualization, Y. I., Y, K., and S. N.; Investigation, F. M. and S. N.; Methodology, F. M.; Interpretation of data and Project administration, Y. I. and Y, K.; Supervision, Y.I. and S. N.; Validation, Y. K. Writing – original draft, M.T. and S. N.
Funding
This study received no external funding.
Institutional Review Board Statement
This study was conducted according to the guidelines of Institutional Animal Care and Committee Guide of Intelligence and Technology Lab, Inc. (ITL) based on the Guidelines for Proper Conduct of Animal Experiments and approved by the Animal Care Committee of the ITL (approved number: AE2023011 date: 11 September 2023). This study was also approved by the ITL Biosafety Committee (approved number: BS2023011, date: 11 September 2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available from S.N. upon reasonable request.
Acknowledgments
We thank Kazuhiro Kawai, Masaki Matsuura, and Makiko Itoda (Intelligence and Technology Lab Inc.) for their invaluable technical assistances. We also appreciate Dr. Yuki Shiraishi (EP Mediate Co., Ltd.) for his help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- A.T. Slominski, et al., How UV light touches the brain and endocrine system through skin, and why, Endocrinology 159 (5) (2018) 1992–2007. [CrossRef]
- R.M. Slominski, J.Y. Chen, C. Raman, A.T. Slominski, Photo-neuro-immuno- endocrinology: how the ultraviolet radiation regulates the body, brain, and immune system, Proc. Natl. Acad. Sci. U. S. A. 121 (14) (2024) e2308374121. [CrossRef]
- S. Zhang, E. Duan, Fighting against skin aging: the way from bench to bedside, Cell Transplant. 27 (5) (2018) 729–738.
- McCarty, M.F.; DiNicolantonio, J.J. A Fundamental Role for Oxidants and Intracellular Calcium Signals in Alzheimer’s Pathogenesis-And How a Comprehensive Antioxidant Strategy May Aid Prevention of This Disorder. Int. J. Mol. Sci. 2021, 22, 2140. [CrossRef]
- Imai, Y.; Koseki, Y.; Hirano,M.; Nakamura, S. Nutrigenomic Studies on the Ameliorative Effect of Enzyme-Digested Phycocyanin in Alzheimer’s Disease Model Mice.Nutrients 2021, 13, 4431. [CrossRef]
- Jang YA, Kim BA. Protective Effect of Spirulina-Derived C-Phycocyanin against Ultraviolet B-Induced Damage in HaCaT Cells, Medicina (Kaunas). 2021 Mar 16;57(3):273. [CrossRef]
- Favas R, Morone J, Martins R, Vasconcelos V, Lopes G. Cyanobacteria Secondary Metabolites as Biotechnological Ingredients in Natural Anti-Aging Cosmetics: Potential to Overcome Hyperpigmentation, Loss of Skin Density and UV Radiation-Deleterious Effects, Mar Drugs. 2022 Mar 1;20(3):183. [CrossRef]
- Ramos SDP, Bürck M, Costa SFFD, Assis M, Braga ARC., Spirulina as a Key Ingredient in the Evolution of Eco-Friendly Cosmetics, BioTech (Basel). 2025 May 30;14(2):41. [CrossRef]
- Zhou Y, Bai R, Huang Y, Li W, Chen J, Cheng Z, Wu X, Diao Y.The anti-photoaging effect of C-phycocyanin on ultraviolet B-irradiated BALB/c-nu mouse skin, Front Bioeng Biotechnol, 2023 Aug 22:11:1229387. [CrossRef]
- lmai Y, Hirahashil T, Fukuda M. Phycocyanin Reduces Transepidermal water Loss and lmproves Skin Barrier Function in Healthy subjects-A Randomized, Double-blind, Placebo-controlled, Paralle-group Comparison Trial-, Jpn Pharmacol Ther 2019; 47:1833-40, UMINID:000035205.
- Knuschke P, Sun Exposure and Vitamin D,Curr Probl Dermatol, 2021:55:296-315. [CrossRef]
- Niehues H, Rikken G, Kersten FFJ, Eeftens JM, van Vlijmen-Willems IMJJ, Rodijk-Olthuis D, et al. CYSRT1: an antimicrobial epidermal protein that can interact with late cornified envelope proteins. J Invest Dermatol, 2023;143:1498e1508.e7.
- Nathalie Jonca1,2 and Michel Simon, The Cornified Envelope: A Versatile Contributor to the Epidermal Barrier, Journal of Investigative Dermatology (2023) 143, 1335e1337; [CrossRef]
- Tanaka M, Koyama Y, Nomura Y.,Effects of collagen peptide ingestion on UV-B-induced skin damage.,Biosci Biotechnol Biochem. 2009 Apr 23;73(4):930-2. [CrossRef]
- Yamamoto, T.; Mitsunaga, F.; Wasaki, K.; Kotani, A.; Tajima, K.; Tanji, M.; Nakamura, S. Mechanism Underlying the Immune Responses of Sublingual Vaccine for SARS-CoV-2 with RBD Antigen and Adjuvant, Poly(I:C) or AddaS03, in Non-human Primates. Arch. Microbiol. Immunol. 2023, 7, 150–164.
- Yamamoto, T.; HIrano, M.; Mitsunaga, F.;Wasaki, K.; Kotani, A.; Tajima, K.; Nakamura, S. Molecular Events in Immune Responses to Sublingua Influenza Vaccine with Hemagglutinin Antigen and Poly(I:C) Adjuvant in Nonhuman Primates, Cynomolgus Macaques. Vaccines 2024, 12, 643.
- Shen C, Gao J, Yin X, Sheng Y, Sun L, Cui Y, Zhang X, Association of the late cornified envelope-3 genes with psoriasis and psoriatic arthritis: a systematic review, J Genet Genomics. 2015 Feb 20;42(2):49-56. [CrossRef]
- Gibbs S, Fijneman R, Wiegant J, van Kessel AG, van De Putte P, Backendorf C.,Molecular characterization and evolution of the SPRR family of keratinocyte differentiation markers encoding small proline-rich proteins, Genomics. 1993 Jun;16(3):630-7. [CrossRef]
- Tian S, Chen S, Feng Y, Li Y., The Interactions of Small Proline-Rich Proteins with Late Cornified Envelope Proteins are Involved in the Pathogenesis of Psoriasis,Clin Cosmet Investig Dermatol. 2021 Sep 24:14:1355-1365. [CrossRef]
- Romashin DD, Tolstova TV, Varshaver AM, Kozhin PM, Rusanov AL, Luzgina NG., Keratins 6, 16, and 17 in Health and Disease: A Summary of Recent Findings., Curr Issues Mol Biol. 2024 Aug 6;46(8):8627-8641. [CrossRef]
- Kim SY, Bae CD., Calpain inhibitors reduce the cornified cell envelope formation by inhibiting proteolytic processing of transglutaminase 1, Exp Mol Med. 1998 Dec 31;30(4):257-62. [CrossRef]
- Doleckova I, Vidovic T, Jandova L, Gretzmeier C, Navarini AA, MacArthur MR, Goksel O, Nyström A, Ewald CY., Calpain Inhibition Protects against UVB-Induced Degradation of Dermal-Epidermal Junction-Associated Proteins, J Invest Dermatol., 2024 Sep;144(9):2103-2107.e2. [CrossRef]
- Ota A, Morita H, Naganuma T, Miyamoto M, Jojima K, Nojiri K, Matsuda J, Kihara A., Bifunctional DEGS2 has higher hydroxylase activity toward substrates with very-long-chain fatty acids in the production of phytosphingosine ceramides, J. Biol Chem., 2023 Apr;299(4):104603. [CrossRef]
- Krieg P, Schuppler M, Koesters R, Mincheva A, Lichter P, Marks F., Repetin (Rptn), a new member of the "fused gene" subgroup within the S100 gene family encoding a murine epidermal differentiation protein, GEnomics., 1997 Aug 1;43(3):339-48. [CrossRef]
- Liddle J, Beneton V, Benson M, Bingham R, Bouillot A, Boullay AB, et al., A Potent and Selective Kallikrein-5 Inhibitor Delivers High Pharmacological Activity in Skin from Patients with Netherton Syndrome, J Invest Dermatol. 2021 Sep;141(9):2272-2279. [CrossRef]
- Zhang W, Yao J, Chen C, Wang J, Zhou A., Fetuin-B Overexpression Promotes Inflammation in Diabetic Retinopathy Through Activating Microglia and the NF-κB Signaling Pathway, Curr Eye Res. 2024 Feb;49(2):168-179. [CrossRef]
- Overmiller AM, Uchiyama A, Hope ED, Nayak S, O'Neill CG, Hasneen K, Chen YW, Reprogramming of epidermal keratinocytes by PITX1 transforms the cutaneous cellular landscape and promotes wound healing., JCI Insight. 2024 Dec 20;9(24):e182844. [CrossRef]
- Nauroy P, Nyström A, Kallikreins: Essential epidermal messengers for regulation of the skin microenvironment during homeostasis, repair and disease, Matrix Biol Plus, 2019 Nov 21:6-7:100019. [CrossRef]
- Wang J, Li J, Zhou L, Hou H, Zhang K., Regulation of epidermal barrier function and pathogenesis of psoriasis by serine protease inhibitors, Front Immunol. 2024 Dec 16:15:1498067. [CrossRef]
- Zhou J, Dong Y, Liu J, Ren J, Wu J, Zhu N., AQP5 regulates the proliferation and differentiation of epidermal stem cells in skin aging, Braz J Med Biol Res 2020 Sep 18;53(11):e10009. [CrossRef]
- Calabrese L, Fiocco Z, Satoh TK, Peris K, French LE., Herapeutic potential of targeting interleukin-1 family cytokines in chronic inflammatory skin diseases, Br J Dermatol. 2022 Jun;186(6):925-941. [CrossRef]
- Odell ID, Steach H, Gauld SB, Reinke-Breen L, Karman J, Carr TL, Epiregulin is a dendritic cell-derived EGFR ligand that maintains skin and lung fibrosis, Sci Immunol. 2022 Dec 16;7(78):eabq6691. [CrossRef]
- Murase T, Shinba Y, Mitsuma M, Abe Y, Yamashita H, Ikematsu K., Wound age estimation based on chronological changes in chitinase 3-like protein 1 expression. Leg Med (Tokyo). 2022 Nov;59:102128. [CrossRef]
- Zhang Y, Inoue Y, Fardous J, Doi R, Ijima T, Fujibuchi T, et al, Prevention and Repair of Ultraviolet B-Induced Skin Damage in Hairless Mice via Transdermal Delivery of Growth Factors Immobilized in a Gel-in-Oil Nanoemulsion.,ACS Omega. 2023 Mar 3;8(10):9239-9249. [CrossRef]
- Makino T, Mizawa M, Takemoto K, Shimizu T., Ultraviolet B irradiation increases the expression of cornulin and retepin in human skin xenotransplants. Exp Dermatol. 2024 May;33(5):e15109. [CrossRef]
- Zhao J, Zhang X, Zhang D, Tang Q, Bi Y, Yuan L, Yang B, Li X, Li Z, Deng D, Cao W., Critical genes in human photoaged skin identified using weighted gene co-expression network analysis., Genomics. 2023 Sep;115(5):110682. [CrossRef]
- Ma Z, Mo R, Yang P, Ding Y, Zhang H, Dong Z, Chen Y, Tan Q., PDK4 facilitates fibroblast functions and diabetic wound healing through regulation of HIF-1α protein stability and gene expression., FASEB J. 2023 Oct;37(10):e23215. [CrossRef]
- Yáñez M, Gil-Longo J, Campos-Toimil M., Calcium binding proteins., Adv Exp Med Biol. 2012;740:461-82. [CrossRef]
- Schäfer M, Werner S., The cornified envelope: a first line of defense against reactive oxygen species., J Invest Dermatol. 2011 Jul;131(7):1409-11. [CrossRef]
Figure 1.
Examinations and samplings and their time points performed here.
Figure 2.
A: Skin moisture level (arbitrary unit) and B: relative moisture (%). Ctr group (open bar and circle), UVB group (red bar and circle), UVB+PC group (blue bar and circle), and UVB+ EDPC group (red bar and circle). ##p < 0.01, UVB compared with Ctr. $$ p < 0.01, UBV+ EDPC, compared with UBV.
Figure 2.
A: Skin moisture level (arbitrary unit) and B: relative moisture (%). Ctr group (open bar and circle), UVB group (red bar and circle), UVB+PC group (blue bar and circle), and UVB+ EDPC group (red bar and circle). ##p < 0.01, UVB compared with Ctr. $$ p < 0.01, UBV+ EDPC, compared with UBV.
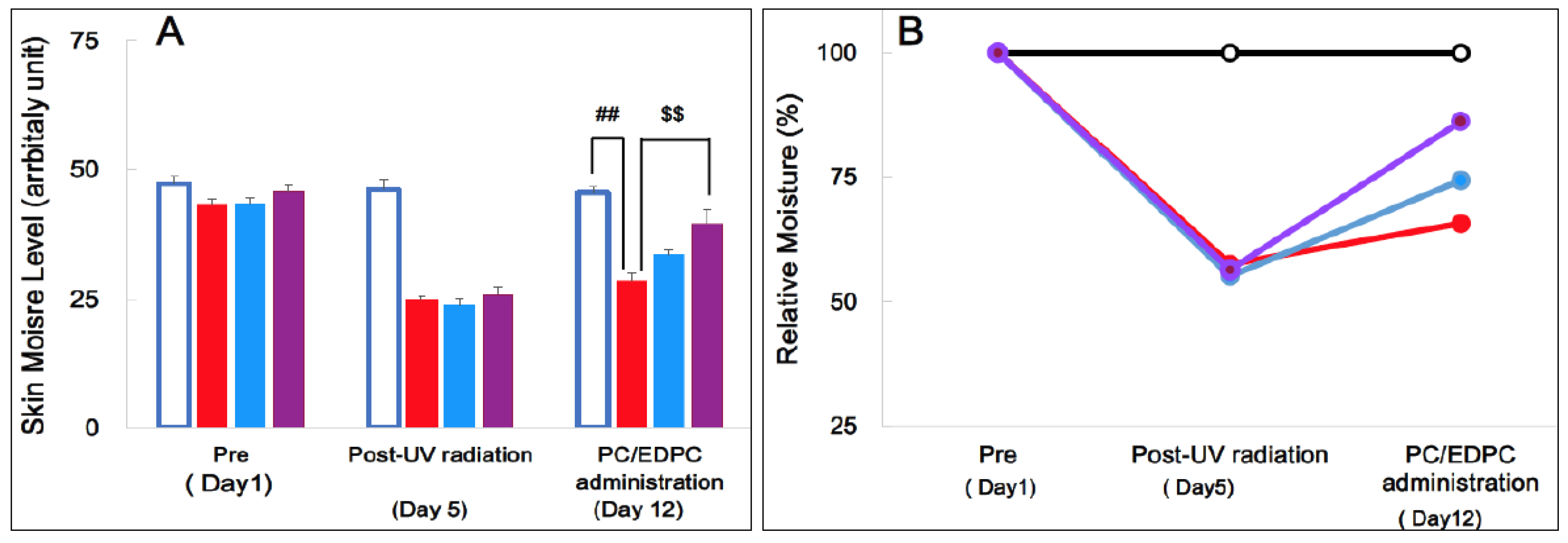
Figure 3.
Gross structure of epidermis and its components, modified from related references [13,39].
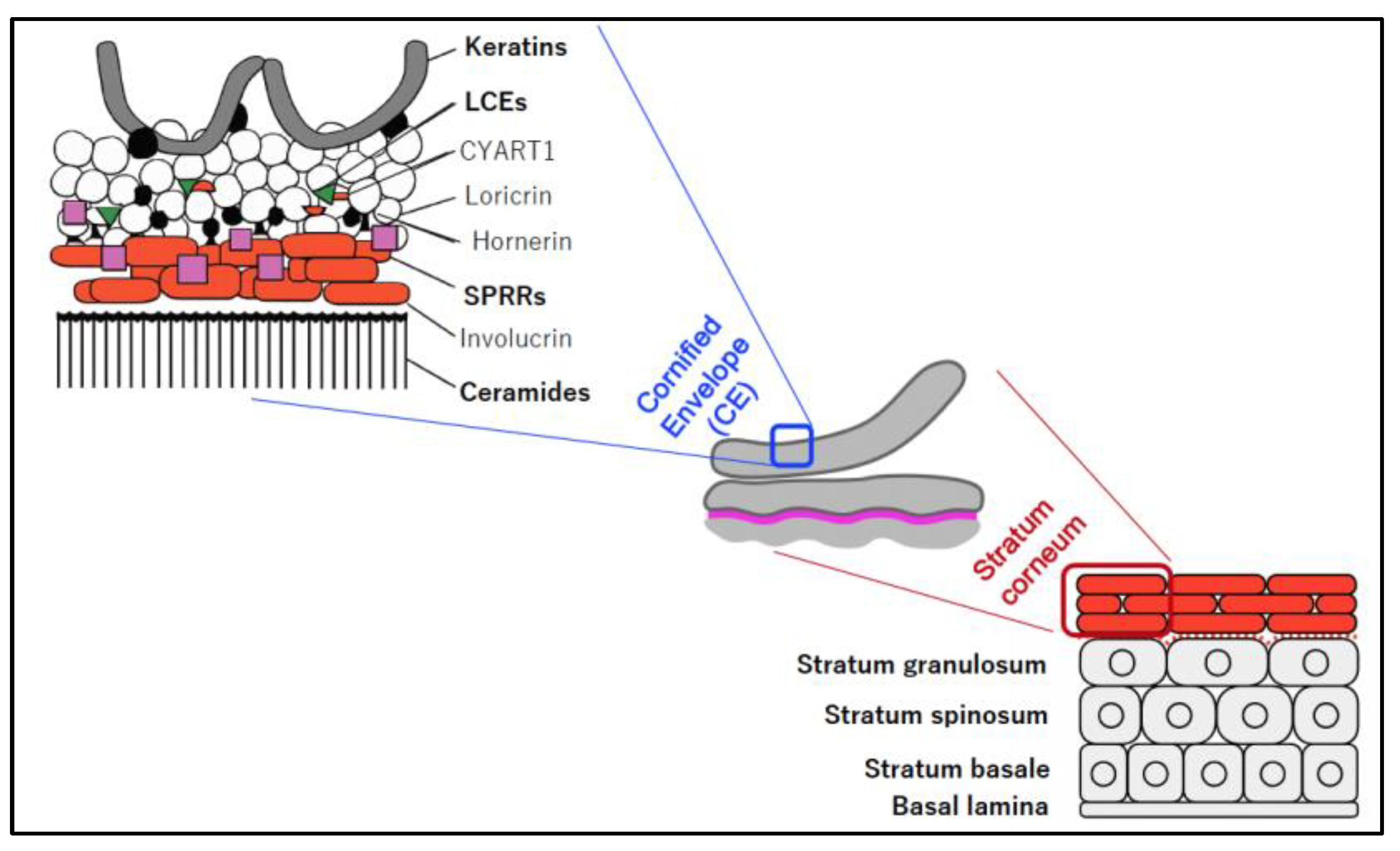

Table 1.
Additionally upregulated PC/EDPC-related genes. Genes exhibited additionally upregulated expression by 1.5-fold or more in the UVB+PC and/or UVB+EDPC groups, in comparison with that of the UVB group.
Table 1.
Additionally upregulated PC/EDPC-related genes. Genes exhibited additionally upregulated expression by 1.5-fold or more in the UVB+PC and/or UVB+EDPC groups, in comparison with that of the UVB group.
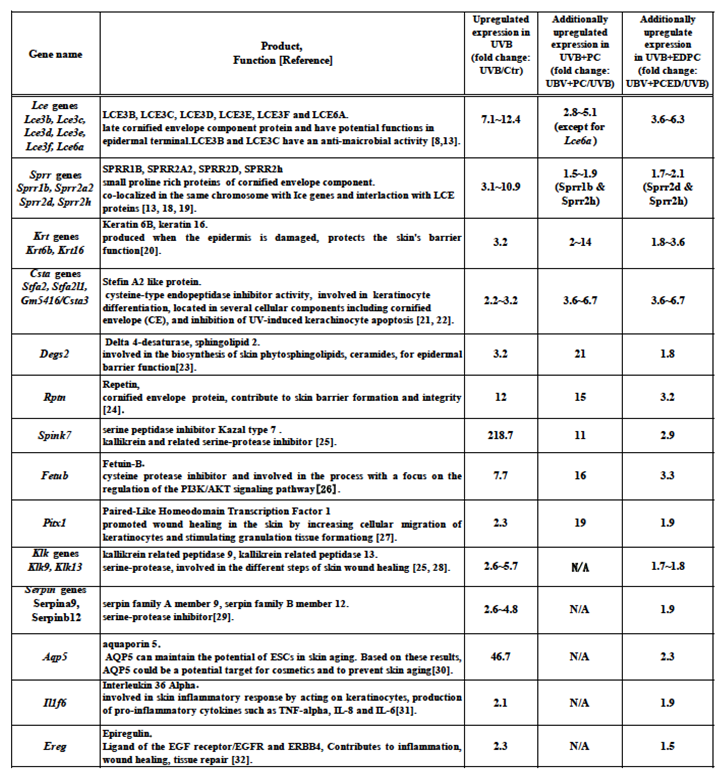 |
Table 2.
Amelioratively downregulated EDPC-related gene. Gene exhibited amelioratively downregulated expression by less than 0.67-fold in UVB+EDPC, in comparison with that of the UVB group.
Table 2.
Amelioratively downregulated EDPC-related gene. Gene exhibited amelioratively downregulated expression by less than 0.67-fold in UVB+EDPC, in comparison with that of the UVB group.
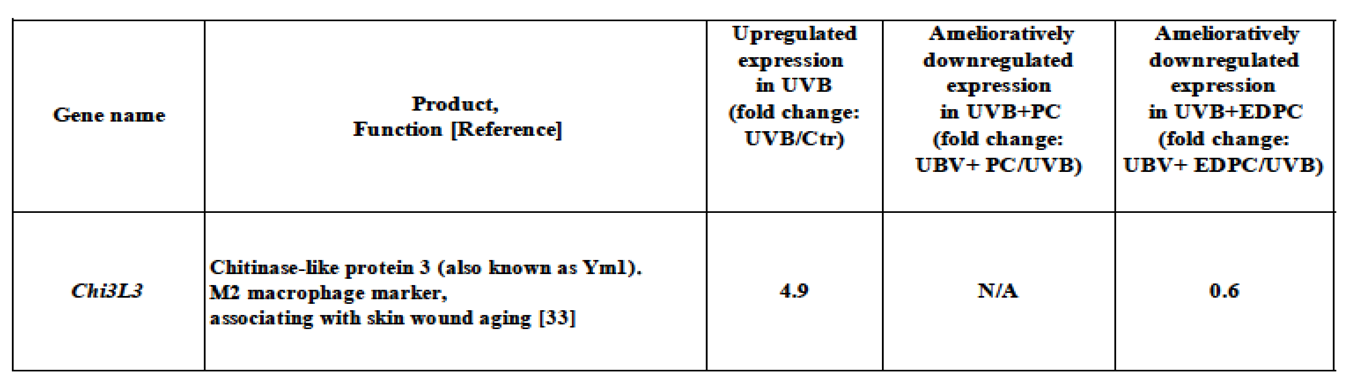 |
Table 3.
Amelioratively upregulated PC/EDPC-related genes. Genes exhibited amelioratively upregulated expression by more than1.5-fold or more in the UVB+PC and/or UVB+EDPC groups, in comparison with that of the UVB group.
Table 3.
Amelioratively upregulated PC/EDPC-related genes. Genes exhibited amelioratively upregulated expression by more than1.5-fold or more in the UVB+PC and/or UVB+EDPC groups, in comparison with that of the UVB group.
 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



