
Submitted:
24 December 2025
Posted:
26 December 2025
You are already at the latest version
Abstract
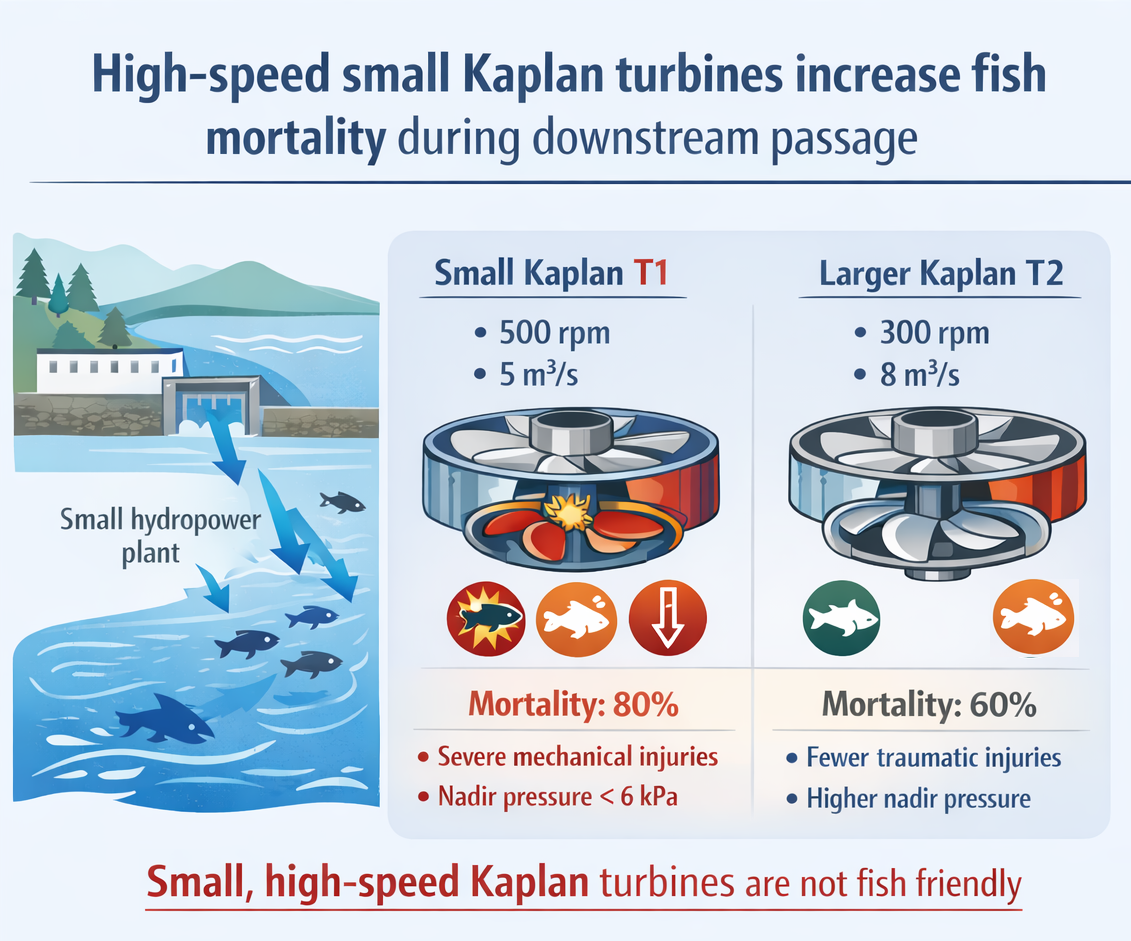
Fish passage through turbines is one of the main environmental impacts of hydropower. Turbine type is a key factor influencing fish survival, and widespread Kaplan turbines are generally considered less dangerous than other turbine types. Nevertheless, while large Kaplan turbines have been extensively studied, there is limited empirical evidence about the biological impact of small, high-speed Kaplan turbines on fish survival. In this study, we conducted controlled in situ fish experiments at a small and low head hydropower plant (1 MW; head 8 m) using balloon tags and pressure sensors to quantify real mortality in two horizontal Kaplan turbines operating at full capacity: one small turbine (1.2 m Ø, 500 rpm and 5 m3/s) and one larger unit (1.55 m Ø, 300 rpm and 8 m3/s). Fish (9.5–19 cm) were released into the intake flow and monitored post-passage. Results showed higher mortality in the small turbine, with ~80% in 24 hours, many exhibiting severe mechanical injuries such as complete sectioning of the head or spinal cord, with significantly higher mortality in greater fish. In contrast, the larger turbine showed a ~60% mortality rate, and fewer traumatic injuries. Our findings highlight the underestimated impact of small, high-rpm Kaplan turbines on fish survival and underscore the need for adaptive turbine operation or structural modifications to minimize ecological damage during critical migration periods.
Keywords:
Kaplan turbine
; fish mortality
; balloon tag
; downstream migration
; smolts
; salmonids
; hydropower impact
1. Introduction
Free and safe downstream movements are necessary for fish conservation [1,2]. Downstream migration is a vital process for diadromous fish species, and it is also a key factor for fitness in potamodromous species that only move within freshwater systems [3]. For diadromous fishes, the on-time descent to estuarine or marine habitats marks a key life stage, often tied to reproduction or maturation, while potamodromous species require longitudinal connectivity, whether by seasonal habitat use or gene flow. Despite its ecological relevance, downstream migration has historically received less attention in impact assessments compared to upstream movements This bias persists in regulatory frameworks and mitigation efforts, which often prioritize fishways for upstream movements [4,5]. Consequently, the hazards associated with diversion channel entrainment, turbine passage, and lack of effective downstream routes and solutions remain underestimated.
Although occasionally upstream fishways can be used for downward passage [1,5], fish moving downstream and encountering a dam face two main pathways: falling over the spillway or being drawn into diversion devices (canals, pipes, and/or turbines). Dams and weirs are rarely equipped with specific devices for downstream bypassing obstacles such as protective screens (physical, mechanical, and/or behavioral) that avoid the entry into facilities (diversion canals or turbines), or guiding structures to safety bypasses (e.g., sluices) [6]. Spillway routes can be relatively safe when the drop is moderate and tailwater depth is sufficient to avoid traumatic impact [7]. However, when the diversion takes the most discharge and there are no protective screens, many fish may not detect the spillway, and they are funneled into intake structures. This can lead to entering irrigation, water supply, or hydropower canals/pipes, causing migratory delays and substantially increasing the risk of mechanical and physiological injuries [8,9,10].
In the case of hydroelectric power plants (HPP), passage through turbines usually becomes the only or most likely alternative for fish moving downstream. It exposes individuals to several stressors: sudden pressure changes (rapid decompression, even cavitation), fluid shear and turbulence, caused by high turbulent flow, and mechanical strike from mainly mobile turbine runner blades or stay vanes, wicket gates, and screens [11]. Rapid decompression can exceed physiological tolerances, leading to barotrauma, especially in physoclistous species (fish with swim bladders not connected to the digestive tract and thus slowly dissolve swim bladder gases into the blood) [11,12]. Shear forces disrupt fish equilibrium and cause internal damage [13,14,15], and direct blade strike remains one of the most severe causes of mortality, particularly at high rotational speeds [15]. These stressors may act independently or synergistically, increasing the cumulative mortality risk. Average worldwide fish mortality from hydroelectric turbine passage was estimated at 22.3% [16]. However, fish mortality rates vary widely depending on turbine (type, diameter, runner speed, shape, and number of blades), operating conditions (flow and head), and fish characteristics (size, species-specific traits). Studies show that large-bodied fish and physoclistous species are particularly susceptible to turbine-induced injuries [4,7]. While larger and lower-head operating Kaplan turbines are known to be less dangerous than common Francis turbines [17].
Kaplan turbines represent one of the most widely implemented turbine types in hydropower plants worldwide since the early 20th century [18]. These propeller-type turbines feature both adjustable runner blades and wicket gates, making them double-regulated machines that operate at a constant rotational speed (Figure 1). This design enables high efficiency, particularly in low-head hydropower applications (2-20 m) and lower discharge (1-100 m3/s) [19]. Kaplan turbines can be installed in various configurations, including both vertical and horizontal orientations, depending on site-specific requirements. For lower head drop (2-10 m) and lower discharge (1-10 m3/s), these turbines are configured with small diameters (<2 m) to work at high rotational speeds (more than 300 rpm) to reach optimal energy production (0,1-2 MW) [20,21]. Small and high-speed Kaplan turbines are also used as a complement to larger turbines’ installations to work with base/ecological flows or in low river discharge conditions. Thus, their combined use is common in the lower reach of rivers and/or streams with high seasonal flow variations. For instance, in the case of small HPP (<10 MW) in Spain, Kaplan turbines are estimated at 37-42% of the total turbines, and of these, high-speed turbines account for around 30-40% [22,23].
To assess fish passage mortality through turbines, various predictive tools have been developed. E.g. deterministic blade strike models [25,26,27] which estimate only fish collision and its derived mortality; empirical models, such as those by [28], which rely on direct observed injury patterns related to turbine, flow and fish features; and more complex numerical models such as BioPA (Biological Performance Assessment Tool) which uses computational fluid dynamics (CFD) models to estimate the likelihood that a fish will be exposed to a stressor at a specific magnitude and applies the biological response models to estimate the probability of injury or mortality [15]. However, previous models were mainly tested and calibrated using data from large Kaplan or Francis turbines and may not adequately reflect the hydraulic conditions of small, high-rpm units.
Kaplan turbines have generally been associated with moderate mortality rates when operated at low rotational speeds (under 300 rpm) [29], but injury rates can rise sharply under higher velocities or smaller diameters [7]. Nevertheless, despite the widespread use of small and high-rpm Kaplan turbines, no studies have directly verified their biological impact [4,7,30]. Consequently, many operators and environmental authorities assume low ecological risk for smaller Kaplan turbines based on models calibrated for slower, larger turbines. This may lead to underestimation of injury and mortality rates, neglecting key differences in design and hydraulics.
Therefore, this study hypothesizes that fish mortality in small and high-rpm Kaplan turbines is underestimated. Thus, we aim to empirically test the mortality range caused by such Kaplan turbines on fish during downstream migration as a first approximation to their potential damage. The findings will help refine environmental risk assessments and promote more accurate ecological evaluations of small hydropower installations.
2. Materials and Methods
2.1. Ethical Statement
All fish experiments were performed following European Union ethical guidelines (Directive 2010/63/UE), Spanish Act RD 53/2013 and RD 118/2021, with the approval of the competent authorities (Regional Government of Navarra and University of Valladolid). All field work was carried out under the permission of the regional fisheries service and water management authorities. Experiments and procedures were adopted to minimize fish stress and avoid unnecessary damage. Animal welfare principles of replacement, reduction, and refinement [31] were strictly applied in experimental design.
2.2. Study Site
Experiments were carried out in the most downstream HPP of the Bidasoa River, located at the village of Lesaka (Navarre, Spain; Figure 2). This infrastructure has a run-of-the-river configuration, with a net head of 8 m (ranging from 7.8 to 8.2 m depending on river discharge) and an installed capacity of 1 MW. It counts with two different sizes of horizontal Kaplan turbines: a smaller unit (T1) of 1.20 m diameter to operate under low flow conditions (4 blades, 500 rpm, 5 m3/s and 0.34 MW) and a larger unit (T2) of 1.55 m diameter (4 blades, 300 rpm, 8 m3/s and 0.66 MW), which serves as main working unit. Flow stems from a 2 m height dam and a diversion canal of 560 m. There is a fishway for upstream migration but no downstream migration facility to avoid fish entering the intake canal. The mean annual discharge in the river reach is 24.3 m3/s and the mean annual water temperature is 13.2 ºC [32]. According to physical and chemical analysis, water quality is “very good” based on the Spanish Act RD 817/2015.
The HPP is 12 km upstream of the sea and its dam is the first barrier for ascending fish. Atlantic salmon (Salmo salar Linnaeus, 1758) is the most emblematic species in the Bidasoa basin. There, salmon is in the southern limit of its natural distribution range [33,34], with a population nearing extinction, threatened mainly by HPP and global warming [35]. It is estimated that more than 27% of salmon smolts pass through the turbines of this HPP during their downstream migration to the sea [36]. This proportion could be higher in climate scenarios characterized by an increasing frequency of low flow conditions.
Fish assemblage in this area also include other diadromous species, such as sea brown trout (Salmo trutta Linnaeus, 1758); European eel (Anguilla Anguilla Linnaeus, 1758), sea lamprey (Petromyzon marinus Linnaeus, 1758), and Allis shad (Alosa alosa Linnaeus, 1758); and potamodromous species as riverine brown trout (Salmo trutta), Ebro nase (Parachondrostoma miegii Steindachner, 1866), Pyrenean gudgeon (Gobio lozanoi Doadrio & Madeira, 2004), Pyrenean minnow (Phoxinus bigerri Kottelat, 2007), and stone loach (Barbatula barbatula Linnaeus, 1758) [37].
2.3. Experimental Design
The experiment consisted of passing a group of Atlantic salmon smolts (n ≈ 20) through each of the two turbines, operating at 100% of their nominal discharge. As the objective was to estimate a mortality range, releasing a sufficient number of tagged smolts per trial (nt=20) and expecting at least a 50% recapture rate [38] (nr≈10) allowed a statistically reliable prior estimate of turbine-passage mortality range, while complying with reduction and refinement principles by minimizing the total number of fish required for robust inference [39,40].
Because the turbines operate at a constant rotational speed, the most favorable scenario for fish passage occurs when running at 100% capacity. At full discharge, the wicket gates and the gaps between runner blades are maximally open, reducing the probability of blade strike, whereas rapid decompression and shear-related stressors remain relatively stable [25]. For animal welfare reasons, if mortality exceeded 50% under this most favorable condition, the experiment was discontinued.
Experiments were carried out in April 2025. All smolts came from Mugaire fish farm (fork length range: 95–190 mm) (Table 1), avoiding using wild stocking. Fish were acclimated for 2 hours to local river conditions and subsequently anaesthetized (50 mg/L of eugenol diluted in ethyl alcohol), sized (fork length ±1 mm) and tagged. Immediately thereafter, they were released into the intake turbine flow chamber, recovered after turbine passage by a kayak and hand net, and finally monitored 24 hours post-passage in a perforated box placed inside the river.
Balloon tags were used as the method for recapturing fish after their passage through the turbines, as this technique is considered one of the least harmful options (e.g., compared with net capture) [41]. Two balloons were attached to the fish by anchoring small hooks at the mouth, one in the upper jaw and the other in the lower jaw (Figure 2). Inside each balloon, two reaction capsules were inserted containing 1 g of oxalic acid dihydrate and sodium bicarbonate in a 1.4:2 ratio. These capsules were activated at the moment of release by injecting 5 ml of water at approximately 40 °C. The balloons were sealed with a custom-made plug (liquid silicone used for mold casting), through which the activation water was injected. The connection between the balloon and the plug was secured with an O-ring, and the connection between the balloon and the hook was made using braided fishing line [42,43].
Fish were introduced into the turbines through an opening located immediately downstream of the gates of the intake chamber. A PVC pipe, fixed to the gate structure, extended to the suction zone of the turbines (Figure 2). Using a manual plunger, each fish was gently pushed until it was drawn into the water column. Observers were stationed both in the intake chamber—to verify whether fish floated away without being suctioned—and at the turbine outflow, to provide guidance to the kayaker.
To complement the information collected while reducing the number of individuals used, one barotrauma detection probe per turbine was injected to obtain a detailed pressure profile during passage through the facilities (Figure 2).
The barotrauma detection probe is based on the Sentinel board architecture [44], powered by an ESP32 microcontroller. To capture the rapid hydraulic transients characteristic of turbine passage, the unit features a TE Connectivity MS5837-30BA pressure sensor, configured to log high-resolution data at 50 Hz.
Beyond pressure monitoring, the device can be used to characterize physical trauma (shear, strikes, and turbulence) using an Adafruit BNO055 9-DOF Absolute Orientation Sensor [45], which integrates a triaxial accelerometer, gyroscope, and magnetometer. Data is synchronized via an onboard RTC (Real-Time Clock) and written to a high-speed data storage unit. The electronics are powered by a lithium battery with a dedicated charge management system and status LED. For structural integrity, the 3D-printed housing is fully encapsulated in 3M Scotchcast™ Resin 40, a flexible polyurethane potting compound that ensures IP68 waterproofing and exceptional shock absorption against mechanical impact.
2.4. Data Processing
2.4.1. Mortality
Mortality (M) was calculated using the following expression (Equation (1)) [38]. First, the experimental mortality (Me) was determined as the ratio between the number of fish found dead within 24 hours after passage through the turbines and the total number of recovered fish. All fish that died, both immediately after passage and within 24 hours, were necropsied to determine the cause of death. Live fish were also carefully examined for potential injuries. Those with a high likelihood of survival were returned to the river, while individuals with poor survival prospects were euthanized using a high dose of MS-222.
Fish handling, tagging, transport, and recapture may significantly affect post-passage mortality. For this reason, turbine-passage studies commonly include a control group to distinguish handling-related mortality from mortality caused specifically by turbine passage. In this context, the mortality in the control group (Mc) is defined as the proportion of tagged control fish released in the turbine outlet that died after 24 hours, relative to the total number of recovered fish. To adhere as closely as possible to the 3Rs principle, a dedicated control batch was not directly used in this study. Instead, a targeted literature review was conducted to identify an appropriate range of control-group mortality values consistent with the characteristics of our experimental design.
Reported control mortality using balloon-tag studies ranges from <5% in alosines [40] and Chinook salmon smolts [39], to <10% for pike and North American percids [41], and up to 14.6-22.4% for rainbow trout smolts [38]. Based on this, control mortality (Mc) values of 5% and 20% were selected to represent conservative and precautionary scenarios, respectively.
The confidence interval (CI 95%) was obtained through the standard error (SE), calculated using the following expression (Equation (2)) [38], where Se = 1-Me; Sc = 1-Mc; Ne is the number of deployed fish in the experiment, and Nc is a hypothetical control sample size (considered equal to the number of deployed fish in the theoretical control Ne = Nc =20).
Accordingly, the resulting CI should be interpreted as a sensitivity interval conditional on the assumed control mortality, rather than as an inference based on an observed control group.
2.4.2. Pressure Profile
In addition to being represented as the temporal variation in pressure values, the information recorded by the pressure tags is intended to provide a predictor of potential turbine-induced barotrauma. For this purpose, the Ratio of Pressure Change RPC [12,15] was applied. This ratio is calculated as the fish’s acclimation pressure before turbine entry divided by the nadir pressure, defined as the minimum absolute pressure experienced by fish during turbine passage. Lower nadir pressures combined with higher acclimation pressures increase the risk of barotrauma [46], and RPC values equal to or below 2.5 (i.e., a nadir pressure at least 40% of the acclimation pressure) have been suggested to be “fish-friendly” [47].
2.5. Data Analysis
To compare fish fork length between groups (T1, T2), a two-sample Student’s t-test was applied after verifying that sample size and distributional assumptions supported the use of a parametric test. The normality of the sample was evaluated based on skewness and kurtosis (±2 considered as acceptable).
Differences in mortality proportions between turbines, both immediately and 24 hours after turbine passage, were evaluated using Fisher’s exact test. To evaluate whether fish length influenced mortality outcomes, Mann–Whitney U test was used to compare median fork lengths between surviving and dead individuals, as sample sizes were small and normality could not be assumed within outcome groups. In addition, differences in mean mortality between turbines were assessed using Welch’s t-test, which does not assume equal variances and is therefore appropriate for groups with heterogeneous variance or sample size. Significance was evaluated at α = 0.05.
3. Results
3.1. Mortality
a) Experimental mortality (Me)
Following turbine passage, 11/20 fish from T1 and 9/20 from T2 were recovered downstream. The experimental mortality 24 hours after turbine passage was approximately 82% for T1 and 67% for T2. In the case of T1, slightly more than half of the fish died immediately after passing through the turbines, whereas for T2 the majority died within the following 24 hours. Overall, decapitation was the most frequently observed injury, although other types of damage were also detected, including internal trauma (stomach, spinal column) and embolism in the ocular system (Table 2).
Based on these results, and in accordance with animal welfare and ethical principles, it was decided not to conduct additional experimental replicates.
Fish length influenced mortality outcomes following turbine passage. As shown in Figure 3, for turbine T1, individuals classified as dead were significantly larger than those that survived, with survival restricted to smaller fish (<110 mm fork length).
b) Mortality (M)
According to the results for experimental mortality (Me) and the selected reference values for control mortality (Mc), 95% confidence intervals for final mortality (M) were estimated (Table 3). Final mortality exceeded 50% in both turbines and, although differences were not statistically significant due to high variability and overlapping confidence intervals, values were higher for turbine T1.
3.2. Pressure Profile
Recovery of sensors released (two per turbine) with readable data proved to be highly challenging, likely because the devices remained trapped or stationary within the draft tube due to intense turbulence and buoyancy effects (Figure 3). As a result, usable pressure records were obtained only for turbine T1.
Pressure records from turbine T1 (Figure 4) indicated an acclimation pressure of 100.5 kPa, corresponding to fish swimming near the water surface prior to turbine entry. The maximum recorded pressure reached 160.1 kPa, while the mean nadir pressure (i.e., minimum pressure experienced) was 49.9 kPa. Notably, an extreme minimum nadir pressure as low as 5.92 kPa was also recorded.
Based on the mean pressure data, the resulting ratio of pressure change (RPC) was approximately 2.04 (100.5/49.9). However, fish passing close to the blade tips may experience substantially lower nadir pressures, such as 5.92 kPa, resulting in an RPC of approximately 17.
4. Discussion
Small Kaplan turbines with high rotational speeds are widely deployed in river systems experiencing seasonal flow variation and commonly serve as support for larger generation units. These turbines are typically installed in low-head, mainstem reaches, where their productivity is greatest. In regions such as Spain, small and high speed Kaplan units comprise an estimated ~15% of all installed turbines [22,23], highlighting their widespread application in medium and large river networks. Despite their growing prevalence, environmental assessments for these systems frequently rely on data and risk models derived from larger, slower-turning turbines, often presuming a limited biological impact on fish downstream passage based on extrapolation rather than site-specific evidence.
The limited sample size, imposed by animal welfare constraints, resulted in relatively wide confidence intervals and prevented the detection of statistically significant differences in mortality between the two turbines. Nevertheless, survival appeared to be restricted to fish smaller than 110 mm in turbine T1, whereas larger individuals were able to survive passage only through turbine T2. Given the substantial physical differences between the two turbines, clearer contrasts in mortality would have been expected, with higher mortality rates anticipated in T1 compared to T2 [15,48]. Even so, our results provide empirical evidence that small, high-rotational-speed Kaplan turbines pose a severe threat to downstream migrating Atlantic salmon smolts.
Contrary to the general assumption that Kaplan turbines are relatively “fish-friendly” compared to Francis turbines—an assumption mostly based on studies of large, slow-rotating units [17,18]—we also observed high mortality rates on them. The small turbine (T1, 500 rpm) resulted in a mean final mortality of nearly 80%, while the larger unit (T2, 300 rpm) showed a mortality greater than 50%, both under full-flow operating conditions, the most favorable hydraulic scenario for fish survival [25]. These values far exceed the global mean turbine-passage mortality (≈22%; [16]) and the typical <10–15% expected for smolts in larger Kaplan turbines in small HPP [29,38] and large-scale HPP Kaplan installations [4,39]. However, our findings align more closely with recent assessments of small hydropower plants, which suggest that mortality can rise sharply in facilities with narrower runners and higher rotational speeds [49], highlighting a major gap in current environmental impact assumptions for small hydropower.
The predominance of severe mechanical injuries—complete sectioning of the head or trunk—indicates that blade strike was the dominant mortality mechanism, mainly in T1. This is consistent with deterministic models predicting steep increases in strike probability with decreasing runner diameter and increasing rotational speed [25,26,27], particularly when blade count is high and gaps are narrow.
Beyond mechanical trauma, barotrauma also contributed to mortality. Our mean nadir pressure recorded in turbine T1 (≈49 kPa) was in accordance with reported values from Kaplan turbines with similar head [50]. Although it produced an RPC of ≈2.0—typically interpreted as a “low-risk” of mortality associated with rapid decompression in published thresholds [12,47]—several fish exhibited injuries consistent with decompression (ocular hemorrhage, intestinal rupture). Importantly, the minimum recorded nadir pressure (5.92 kPa) is physically near water vapor pressure, implying extreme localized pressure drops likely occurring near blade leading edges or tip vortices, where the lowest pressure zones are predicted [50,51]. In such regions, RPC can exceed 3.0, a level associated with >20–40% barotrauma-related injury in salmonids [15,50] and may even reach values around 17.0, which have been associated with immediate barotrauma-induced mortality [15]. Our finding that at least 4 out of 14 fish with a known cause of death showed barotrauma-compatible lesions supports this interpretation. Nevertheless, barotrauma injuries could be higher, because they are hindered by previous blade strikes and fish sectioning mortality. Nadir pressures in turbine T2 are expected to be higher than those observed in T1, owing to its larger runner diameter and lower rotational speed, as previously reported by [48].
In the case of shear forces, they are known to occur within wicket-gate wakes, tip vortices, and inter-blade acceleration zones; however, their biological effects could not be determined in this study. Shear-related injuries are typically difficult to isolate from strike and decompression trauma, and published thresholds are derived almost exclusively from laboratory flumes rather than from Kaplan runners [15,25]. Consequently, the relative importance of shear in our turbines—both small, high-rpm units—remains uncertain and cannot be classified as either minor or substantial based on available evidence.
Fish forced-entry experiments generally amplify mortality caused by overall handling [52]. Moreover, strike and rapid decompression survival is influenced by volitional behavior, as fish can use physical or physiological responses to avoid or reduce those stressors. Thus, mortality may have been overestimated by handling stress inherent to sedation, tagging, forced turbine passage, and recapture—factors known to contribute 5–20% mortality in balloon-tagged fish [38,39,40]. Nevertheless, even adopting a conservative Mc range (5–20%) does not alter the conclusion: both turbines, particularly the smaller unit T1, caused exceptionally high mortality far beyond what could be explained by handling alone.
The hydropower plant studied lies only 12 km from the sea, in a river reach where >27% of smolts from the whole river basin enter the diversion canal [36], with this proportion likely to increase in dry years. Once inside the long (560 m) canal, fish have limited ability to return, causing migration delay in the turbine’ intake area, which itself increases predation risk and reduces seaward arrival success [8,9,10]. Importantly, the small turbine (T1)—the most lethal—was also positioned in the main bulk-flow region of the intake chamber, making it the most likely path for migrating smolts. Moreover, immediate predation on surviving fish after turbine passage is expected to be important due to injury-related impairment, disorientation, and reduced swimming performance, all of which markedly increase vulnerability to piscivorous fauna [6,18]. Because small Kaplan turbines often operate during the peak smolt migration season, their ecological influence extends beyond immediate mortality, potentially affecting entire cohorts.
The mainstem of the Bidasoa River basin consists of six small HPP (three equipped with low high speed Kaplan turbines and the other three with Francis turbines) along a 41.6 km river reach. Hydraulic/mechanical guidance structures, behavioral barriers, or alternative bypass routes once inside the canals were absent at all HPP, further increasing turbine entrainment probability [6,36]. Therefore, the accumulative impact on salmon populations already near extinction at the edge of their biogeographical range [33,34,35] can be considered unacceptable from a conservation and management perspective.
Furthermore, our study was centered on salmon smolts, but if impacts on adult salmon and riverine sea trout kelts (fish that return to sea after spawning), as well as on eels and lampreys, are considered, the expected mortality may exceed 50%. These findings underscore the broader ecological implications of small HPP for fish assemblages in this basin.
Taken together, our empirical evidence supports the conclusion that small, high-rpm Kaplan turbines pose a disproportionately high risk to downstream fish migrants. Given their widespread use in low-head rivers across Europe and elsewhere, this issue deserves urgent attention. The immplementation of effective downstream bypasses, physical or exclusion screens, behavioral guidance systems, and operational adjustments (e.g., shutting down small turbines during peak migrations) may significantly reduce mortality, as recommended in international best-practice guidelines [4,50].
Ultimately, the current regulatory assumption that “Kaplan turbines are safe” cannot be generalized across scales and rotational speeds. Our study shows that small, high-rpm Kaplan turbines represent a distinct hydraulic environment requiring specific assessment, monitoring, and mitigation. In this context, for future studies, we have identified several aspects that should be improved and/or emphasized. We found that, when recovering balloon tags, using at least two individually identifiable balloons is essential in this type of facility due to the high likelihood of impact. It would also be advisable to place them at both the anterior and posterior ends of the fish to reliably distinguish fragments (the upper jaw proved to be a more secure anchoring site than the lower jaw). The fish or sensor-injection system also warrants attention, as in cases where suction is not sufficiently strong, it is necessary to position the inlet as close to the turbine as possible. We also consider it crucial to perform replicated trials under different turbine operating regimes in order to confirm that the greatest damage occurs when these systems operate at full capacity. Finally, future studies should also incorporate the tracking of actively swimming fish, not only passive (sedated) individuals, to obtain a clearer understanding of processesoccuring in the vicinity of these facilities [53].
5. Conclusions and Management Applications
This study shows that small, high-speed Kaplan turbines pose a far greater threat to downstream-migrating salmon smolts than generally assumed. Mortality exceeded 80% in the 1.20 m, 500 rpm turbine and approximately 60% in the 1.55 m, 300 rpm unit—values far above typical mortality reported for larger Kaplan turbines. Injuries were dominated by severe mechanical trauma, confirming blade strike as the primary mechanism, while internal lesions consistent with barotrauma were also observed, supported by measured nadir pressures and expected localized pressure extremes in small runners.
These findings challenge the widespread regulatory assumption that Kaplan turbines are relatively “fish-friendly,” showing instead that turbine diameter and rotational speed are critical predictors of mortality. Small, high-rpm units constitute a distinct hydraulic environment that is not well represented in existing empirical or mechanistic models, many of which underestimate strike probability and fail to capture extreme nadir pressure.
From a management perspective, the results emphasize the need to:
(i) re-evaluate impact assessments, explicitly incorporating turbine size and rpm;
(ii) implement guidance or exclusion systems in diversion canals to prevent turbine entrainment;
(iii) adopt operational measures, such as shutting down turbines during peak migration periods;
(iv) require improved modelling or field measurements for licensing, particularly in rivers supporting endangered salmon populations.
Given the high proportion of smolts entrained into the intake and the proximity of the HPP to the sea, turbine-related mortality at the observed levels may significantly reduce cohort survival. Mitigation actions focusing on reducing turbine passage are therefore essential for effective conservation and recovery of salmon in this basin.
Author Contributions
Conceptualization, FJSR and FJBC; methodology, FJBC, JFFP and JVC; investigation, FJSR and FJBC; data adquisition, JVC, JMA, FJBC and FJSR; data curation, FJBC and AGV; writing—original draft preparation, FJSR; writing—review and editing, JFFP, JVC, AGV, JMA and FJBC; project administration, FJSR;. All authors have read and agreed to the published version of the manuscript.
Funding
This work has received funding from the project LIFE21- NAT- ES- LIFE KANTAURIBAI (Ref. 101074197) supported by the LIFE programme of the European Commission and the “III Agreetment between Confederación Hidrográfica del Duero-ITAGRA-Universidad de Valladolid). Juan Francisco Fuentes- Pérez’s contribution was funded by the project RYC2022-0 36557- Ministry of Science, Innovation and Universities, the Spanish Research Agency, through the State Plan for Scientific, Technical and Innovation Research 2021–2023 (MCIU/AEI/10.13039/501100011033), and co- financed by the European Social Fund Plus (FSE+).
Data Availability Statement
Data supporting reported results can be found along the manuscript.
Acknowledgments
The authors would like to thank the Fish Management Service of the Government of Navarre and OREKAN for their support. José Manuel Checa Díaz-Maroto, consulting engineer in HGM S.A., helped us in the overview of Kaplan turbines in Spain. During the preparation of this manuscript, the authors used Gemini 3 pro and ChatGPT (OpenAI) to assist with language editing, figure formatting, and the rephrasing of text. All scientific content, data analysis, and interpretation were performed by the authors. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Celestino, L.F.; Sanz-Ronda, F.J.; Miranda, L.E.; Makrakis, M.C.; Dias, J.H.P.; Makrakis, S. Bidirectional connectivity via fish ladders in a large Neotropical river. River Res. Appl. 2019, 35, 236–246. [Google Scholar] [CrossRef]
- Fernandes Celestino, L.; Sanz-Ronda, F.J.; Miranda, L.E.; Cavicchioli Makrakis, M.; Dias, J.H.P.; Makrakis, S. Bidirectional connectivity via fish ladders in a large Neotropical river: Response to a comment. River Res. Appl. 2020, 36, 1377–1381. [Google Scholar] [CrossRef]
- Lucas, M.C.; Baras, E.; Thom, T.J.; Duncan, A.; Slavík, O. Migration of freshwater fishes; Wiley Online Library: Oxford, UK, 2001. [Google Scholar]
- Pracheil, B.M.; DeRolph, C.R.; Schramm, M.P.; Bevelhimer, M.S. A fish-eye view of riverine hydropower systems: the current understanding of the biological response to turbine passage. Rev. Fish Biol. Fish. 2016 262 2016, 26, 153–167. [Google Scholar] [CrossRef]
- Sanz-Ronda, F.J.; Fuentes-Pérez, J.F.; García-Vega, A.; Bravo-Córdoba, F.J. Fishways as Downstream Routes in Small Hydropower Plants: Experiences with a Potamodromous Cyprinid. Water 2021, 13, 1041. [Google Scholar] [CrossRef]
- Schwevers, U.; Adam, B. Fish protection technologies and fish ways for downstream migration; Springer, 2020; Vol. 279. [Google Scholar]
- Mueller, M.; Knott, J.; Pander, J.; Geist, J. Experimental comparison of fish mortality and injuries at innovative and conventional small hydropower plants. J. Appl. Ecol 2022, 59, 2360–2372. [Google Scholar] [CrossRef]
- Aarestrup, K.; Jepsen, N.; Rasmussen, G.; Okland, F. Movements of two strains of radio tagged Altlantic salmon, Salmo salar L., smolts through a reservoir. Fish. Manag. Ecol. 1999, 6, 97–107. [Google Scholar] [CrossRef]
- Jepsen, N.; Aarestrup, K.; Økland, F.; Rasmussen, G. Survival of radiotagged Atlantic salmon (Salmo salar L.)–and trout (Salmo trutta L.) smolts passing a reservoir during seaward migration. Hydrobiologia 1998, 371, 347–353. [Google Scholar] [CrossRef]
- Karppinen, P.; Hynninen, M.; Vehanen, T.; Vähä, J. Variations in migration behaviour and mortality of Atlantic salmon smolts in four different hydroelectric facilities. Fish. Manag. Ecol. 2021, 28, 253–267. [Google Scholar] [CrossRef]
- Cada, G.F.; Coutant, C.C.; Whitney, R.R. Development of biological criteria for the design of advanced hydropower turbines; EERE Publication and Product Library: Washington, DC (United States), 1997. [Google Scholar]
- Brown, R.S.; Pflugrath, B.D.; Colotelo, A.H.; Brauner, C.J.; Carlson, T.J.; Deng, Z.D.; Seaburg, A.G. Pathways of barotrauma in juvenile salmonids exposed to simulated hydroturbine passage: Boyle’s law vs. Henry’s law. Fish. Res. 2012, 121, 43–50. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, F.; Shi, X.; Adu-Poku, K.A.; Zhang, J.; Yuan, S. A systematic investigation on the damage characteristics of fish in axial flow pumps. Processes 2022, 10, 2228. [Google Scholar] [CrossRef]
- Neitzel, D.A.; Dauble, D.D.; Čada, G.F.; Richmond, M.C.; Guensch, G.R.; Mueller, R.P.; Abernethy, C.S.; Amidan, B. Survival estimates for juvenile fish subjected to a laboratory-generated shear environment. Trans. Am. Fish. Soc. 2004, 133, 447–454. [Google Scholar] [CrossRef]
- Pflugrath, B.D.; Harnish, R.A.; Rhode, B.; Engbrecht, K.; Beirao, B.; Mueller, R.P.; McCann, E.L.; Stephenson, J.R.; Colotelo, A.H. The susceptibility of Juvenile American shad to rapid decompression and fluid shear exposure associated with simulated hydroturbine passage. Water 2020, 12, 586. [Google Scholar] [CrossRef]
- Radinger, J.; van Treeck, R.; Wolter, C. Evident but context-dependent mortality of fish passing hydroelectric turbines. Conserv. Biol. 2022, 36, e13870. [Google Scholar] [CrossRef] [PubMed]
- Algera, D.A.; Rytwinski, T.; Taylor, J.J.; Bennett, J.R.; Smokorowski, K.E.; Harrison, P.M.; Clarke, K.D.; Enders, E.C.; Power, M.; Bevelhimer, M.S. What are the relative risks of mortality and injury for fish during downstream passage at hydroelectric dams in temperate regions? A systematic review. Environ. Evid. 2020, 9, 3. [Google Scholar] [CrossRef]
- Čada, G.F. The development of advanced hydroelectric turbines to improve fish passage survival. Fisheries 2001, 26, 14–23. [Google Scholar] [CrossRef]
- Dincer, I.; Ishaq, H. Renewable Hydrogen Production; Elsevier, 2021. [Google Scholar]
- Ball, I.; Hendricks, D.; Jawaid, T.S.; Rutz, D.; Steller, J. Small Hydropower Technologies—European State-of-the-Art Innovations; WIP Renewable Energies: Munich, Germany, 2020. [Google Scholar]
- Halder, P.; Doppalapudi, A.T.; Azad, A.K.; Khan, M.M.K. Chapter 7 - Efficient hydroenergy conversion technologies, challenges, and policy implication. In; Azad, A.K.B.T.-A. in C.E.T., Ed.; Academic Press, 2021; pp. 295–318. ISBN 978-0-12-821221-9. [Google Scholar]
- CHD Consultoría y asistencia para la inspección y vigilancia del cumplimiento del condicionado de las concesiones de aprovechamientos hidroeléctricos en la Confederación Hidrográfica del Duero. Clave: 02.803.267/0811. In forme técnico; Confederación Hidrográfica del Duero.: Valladolid, Spain, 2011.
- Checa-Díaz, J.M. Consulting Engineer in HGM. Personal Comunication, 2025. [Google Scholar]
- Sale, M.J.; Cada, G.F.; Carlson, T.J.; Dauble, D.D.; Hunt, R.T.; Sommers, G.L. DOE Hydropower Program Annual Report for FY 2002 (DOE ID-11107); U.S. Department of Energy Office of Energy Efficiency and Renewable Energy Wind and Hydropower Technologies, 2003. [Google Scholar]
- Deng, Z.; Carlson, T.J.; Ploskey, G.R.; Richmond, M.C.; Dauble, D.D. Evaluation of blade-strike models for estimating the biological performance of Kaplan turbines. Ecol. Modell. 2007, 208, 165–176. [Google Scholar] [CrossRef]
- Von Raben, K. Regarding the problem of mutilations of fishes by hydraulic turbines. Die Wasserwirtschaft 1957, 4, 97–100. [Google Scholar]
- Montén, E. Fish and turbines: fish injuries during passage through power station turbines; Norstedts Tryckeri: Stockholm, Sweden, 1985. [Google Scholar]
- Bosc, S.; Larinier, M. Définition d’une statégie de réouverture de la Garonne et de l’Ariège à la dévalaison des salmonidés grands migrateurs: simulation des mortalités induites par les aménagements hydroélectriques lors de la migration de dévalaison; Irstea: Toulouse,France, 2000. [Google Scholar]
- Larinier, M.; Dartiguelongue, J. La circulation des poissons migrateurs: le transit à travers les turbines des installations hydroélectriques. Bull. Français la Pêche la Piscic. 1989, 1–87. [Google Scholar] [CrossRef]
- Rutschmann, P.; Kampa, E.; Wolter, C.; Albayrak, I.; David, L.; Stoltz, U.; Schletterer, M. Novel Developments for Sustainable Hydropower; Springer Nature, 2022. [Google Scholar]
- Russell, W.M.S.; Burch, R.L. The principles of humane experimental technique.; Universities Federation For Animal Welfare (UFAW): Wheathampstead, UK, 1959. [Google Scholar]
- MAPAMA Anuario de aforos 2019-2020. Estación 1106 río Bidasoa en Endarlatza; Ministerio de Agricultura y Pesca Alimentación y Medio Ambiente: Madrid, Spain, 2023.
- Horreo, J.L.; Machado-Schiaffino, G.; Griffiths, A.M.; Bright, D.; Stevens, J.R.; Garcia-Vazquez, E. Atlantic salmon at risk: apparent rapid declines in effective population size in southern European populations. Trans. Am. Fish. Soc. 2011, 140, 605–610. [Google Scholar] [CrossRef]
- Nicola, G.G.; Elvira, B.; Jonsson, B.; Ayllón, D.; Almodóvar, A. Local and global climatic drivers of Atlantic salmon decline in southern Europe. Fish. Res. 2018, 198, 78–85. [Google Scholar] [CrossRef]
- García-Vega, A.; Fuentes-Pérez, J.F.; Bravo-Córdoba, F.J.; Elso, J.; Ardaiz-Ganuza, J.; Sanz-Ronda, F.J. Assessing Trends and Challenges: Insights From 30 Years of Monitoring and Management of Threatened Southern Atlantic Salmon Populations. Aquat. Conserv. Mar. Freshw. Ecosyst. 2025, 35, e70052. [Google Scholar] [CrossRef]
- Sanz-Ronda, F.J.; Manzano, S.; Bravo-Córdoba, F.J.; Fuentes-Pérez, J.F.; García-Vega, A.; Valbuena-Castro, J. Evaluación de la mortalidad de la fauna piscícola asociada al descenso por turbinas en centrales hidroeléctricas del río Bidasoa. Resultados preliminares. Technical report, Project LIFE21-NAT-ES-LIFE KANTAU-RIBAI (101074197). 2025. [Google Scholar]
- Government of Navarre Registro ictiológico de Navarra (1978-2015). Base de datos inédita. 2016.
- Ballesteros, F.; Vázquez, V.M. Evaluación de las mortalidad de peces tras su paso por turbinas hidroeléctricas en ríos del norte de España. Ecología 2001, 275–284. [Google Scholar]
- Skalski, J.R.; Townsend, R.; Lady, J.; Giorgi, A.E.; Stevenson, J.R.; McDonald, R.D. Estimating route-specific passage and survival probabilities at a hydroelectric project from smolt radiotelemetry studies. Can. J. Fish. Aquat. Sci. 2002, 59, 1385–1393. [Google Scholar] [CrossRef]
- Heisey, P.G.; Mathur, D.; D’Allesandro, L. A new technique for assessing fish passage survival at hydro power stations. In Proceedings of the Proceedings of the workshop on fish passage at hydroelectric developments, 1993; pp. 32–38. [Google Scholar]
- Tuononen, E.I.; Cooke, S.J.; Timusk, E.R.; Smokorowski, K.E. Extent of injury and mortality arising from entrainment of fish through a Very Low Head hydropower turbine in central Ontario, Canada. Hydrobiologia 2022, 849, 407–420. [Google Scholar] [CrossRef]
- Salalila, A.; Martinez, J.; Tate, A.; Acevedo, N.; Salalila, M.; Deng, Z.D. Balloon Tag Manufacturing Technique for Sensor Fish and Live Fish Recovery. J. Vis. Exp. 2023, e65632. [Google Scholar] [CrossRef] [PubMed]
- Mathur, D.; Heisey, P.G.; Euston, E.T.; Skalski, J.R.; Hays, S. Turbine passage survival estimation for chinook salmon smolts (Oncorhynchus tshawytscha) at a large dam on the Columbia River. Can. J. Fish. Aquat. Sci. 1996, 53, 542–549. [Google Scholar] [CrossRef]
- MyCircuits The Sentinel board. Available online: https://github.com/MyCircuitsTV/Sentinel (accessed on 17 Sep 2023).
- Adafruit Industries Adafruit BNO055 Absolute Orientation Sensor. Adafruit Industries. Available online: https://learn.adafruit.com/adafruit-bno055-absolute-orientation-sensor (accessed on 25 Feb 2020).
- Becker, J.M.; Abernathy, C.S.; Dauble, D.D. Identifying the effects on fish of changes in water pressure during turbine passage; EERE Publication and Product Library: Washington, DC (United States), 2003. [Google Scholar]
- Fraser, R.; Deschênes, C.; O’Neil, C.; Leclerc, M. VLH: development of a new turbine for very low head sites. Proceeding 15th Waterpower 2007, 10, 23–26. [Google Scholar]
- Martinez, J.J.; Deng, Z.D.; Titzler, P.S.; Duncan, J.P.; Lu, J.; Mueller, R.P.; Tian, C.; Trumbo, B.A.; Ahmann, M.L.; Renholds, J.F. Hydraulic and biological characterization of a large Kaplan turbine. Renew. energy 2019, 131, 240–249. [Google Scholar] [CrossRef]
- Mueller, M.; Pander, J.; Geist, J. Evaluation of external fish injury caused by hydropower plants based on a novel field-based protocol. Fish. Manag. Ecol. 2017, 24, 240–255. [Google Scholar] [CrossRef]
- Boys, C.A.; Pflugrath, B.D.; Mueller, M.; Pander, J.; Deng, Z.D.; Geist, J. Physical and hydraulic forces experienced by fish passing through three different low-head hydropower turbines. Mar. Freshw. Res. 2018, 69, 1934–1944. [Google Scholar] [CrossRef]
- Cook, T.C.; Hecker, G.E.; Faulkner, H.B.; Jansen, W. Development of a more fish-tolerant turbine runner, advanced hydropower turbine project; Worcester Polytechnic Inst.: Holden, MA (United States); Alden Research Lab …, 1997. [Google Scholar]
- EPRI Report No. TR-101231, Project 2694-01.; EPRI Fish entrainment and turbine mortality review and guidelines. Prepared by Stone & Webster Environmental Services. Electric Power Research Institute: California, USA, 1992.
- Pflugrath, B.D.; Mueller, R.P.; Deters, K.A.; Watson, S.M.; Schneider, A.D.; Deng, Z.D. Maximizing Safe Passage for Large Fish: Evaluating Survival of Rainbow Trout Through a Novel Hydropower Turbine. Environ. Sustain. Indic. 2025, 100801. [Google Scholar] [CrossRef]
Figure 1.
Schematic of a Kaplan turbine illustrating potential fish injury zones modified from [24].
Figure 1.
Schematic of a Kaplan turbine illustrating potential fish injury zones modified from [24].
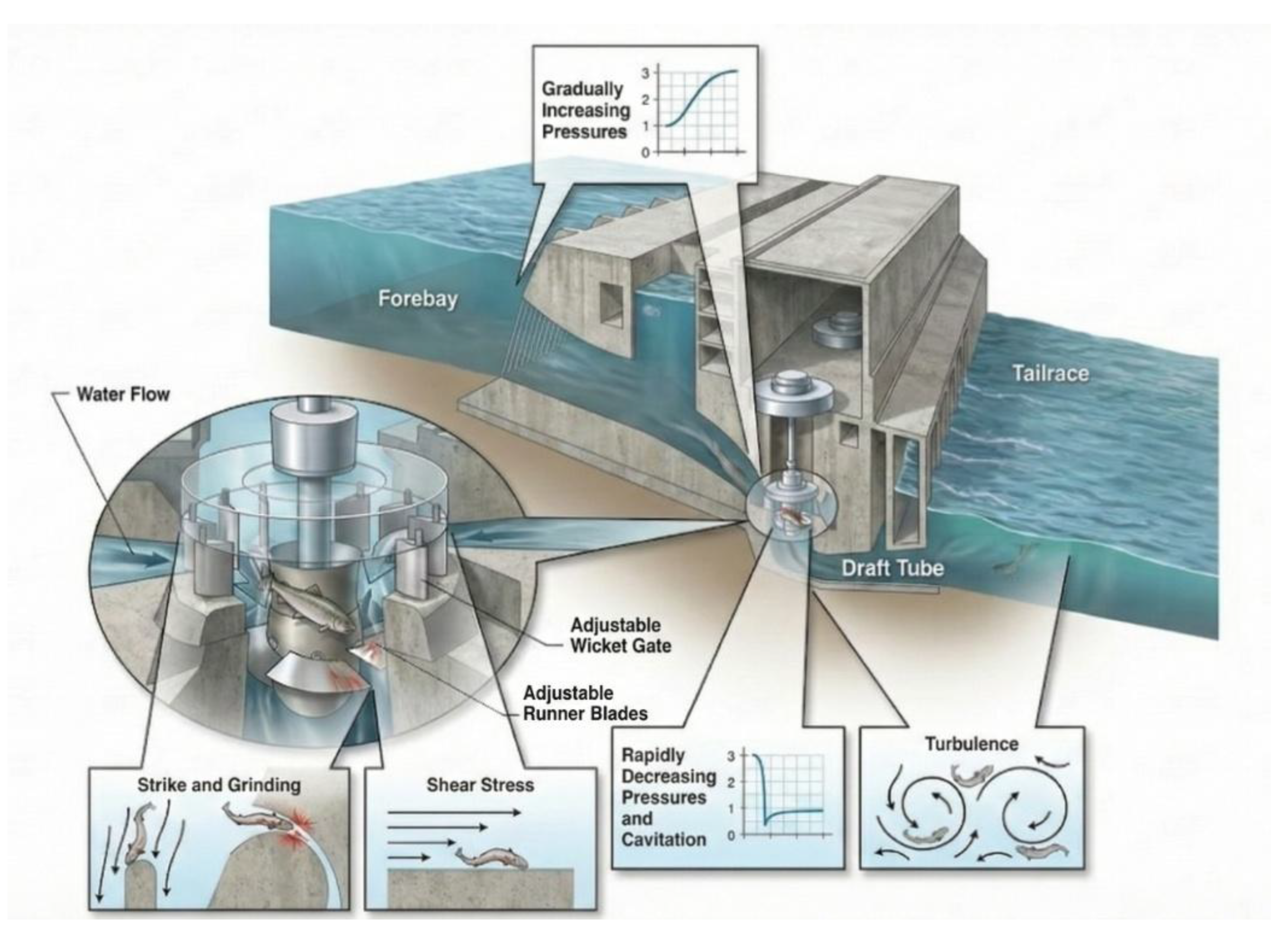
Figure 2.
Tagging and injection with balloon tags. Left: injection pipe with pressure tag ready for release. Right: salmon smolt tagged with two balloon tags.
Figure 2.
Tagging and injection with balloon tags. Left: injection pipe with pressure tag ready for release. Right: salmon smolt tagged with two balloon tags.

Figure 3.
Box-plot and length comparison based on whether they were dead or alive after 24 h of passing through the turbines (T1: small turbine; T2: large turbine). Different letters above the box plots indicate statistically significant differences between groups based on pairwise comparisons (Mann–Whitney U test, p = 0.045).
Figure 3.
Box-plot and length comparison based on whether they were dead or alive after 24 h of passing through the turbines (T1: small turbine; T2: large turbine). Different letters above the box plots indicate statistically significant differences between groups based on pairwise comparisons (Mann–Whitney U test, p = 0.045).
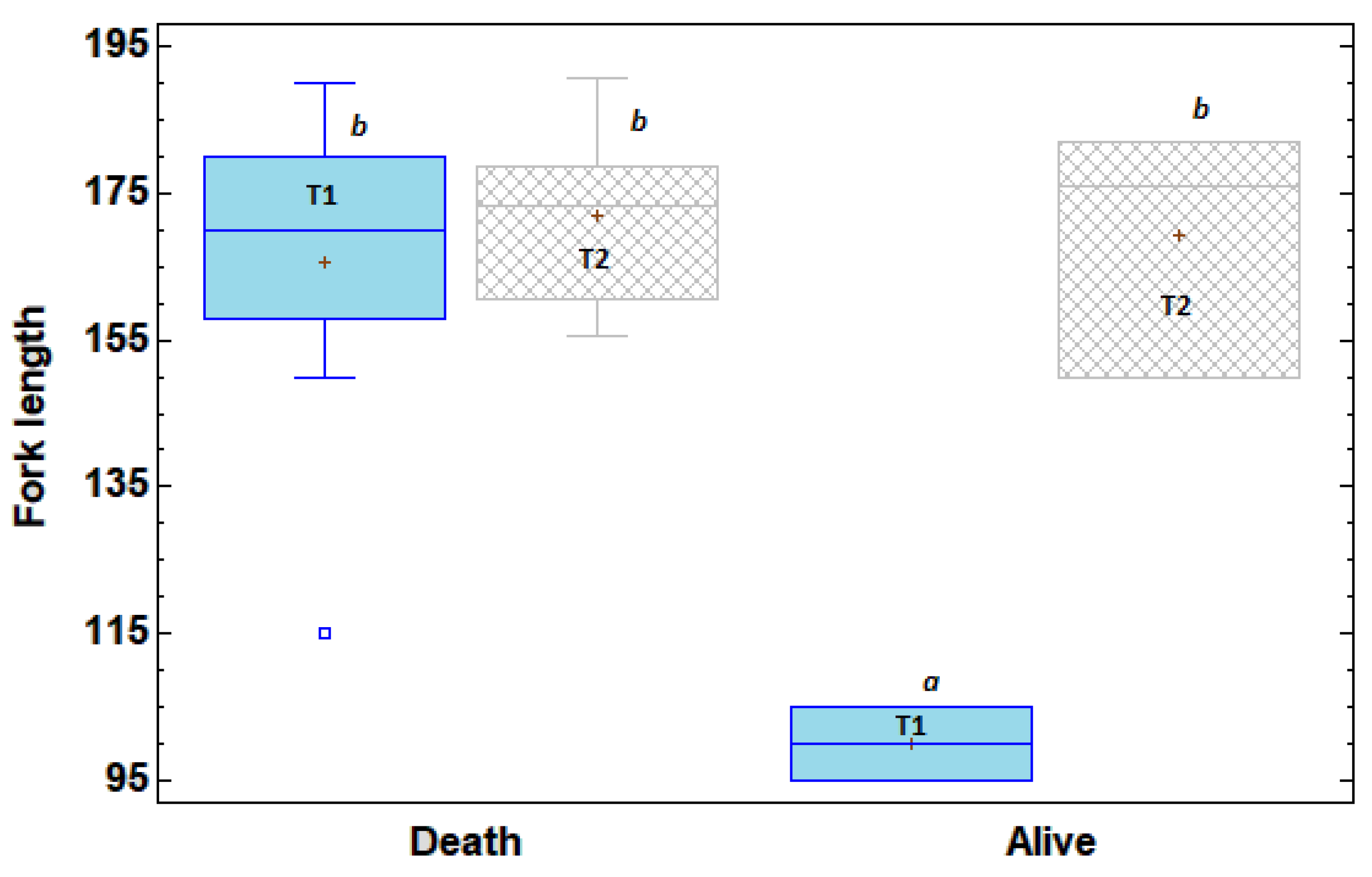
Figure 4.
Pressure profile recorded during the path passage through the Kaplan turbine exhibiting the lowest nadir pressure. Note that the time axis includes a break to exclude a static period during which the sensor remained stationary in the draft tube, thereby highlighting pressure changes during turbine passage.
Figure 4.
Pressure profile recorded during the path passage through the Kaplan turbine exhibiting the lowest nadir pressure. Note that the time axis includes a break to exclude a static period during which the sensor remained stationary in the draft tube, thereby highlighting pressure changes during turbine passage.
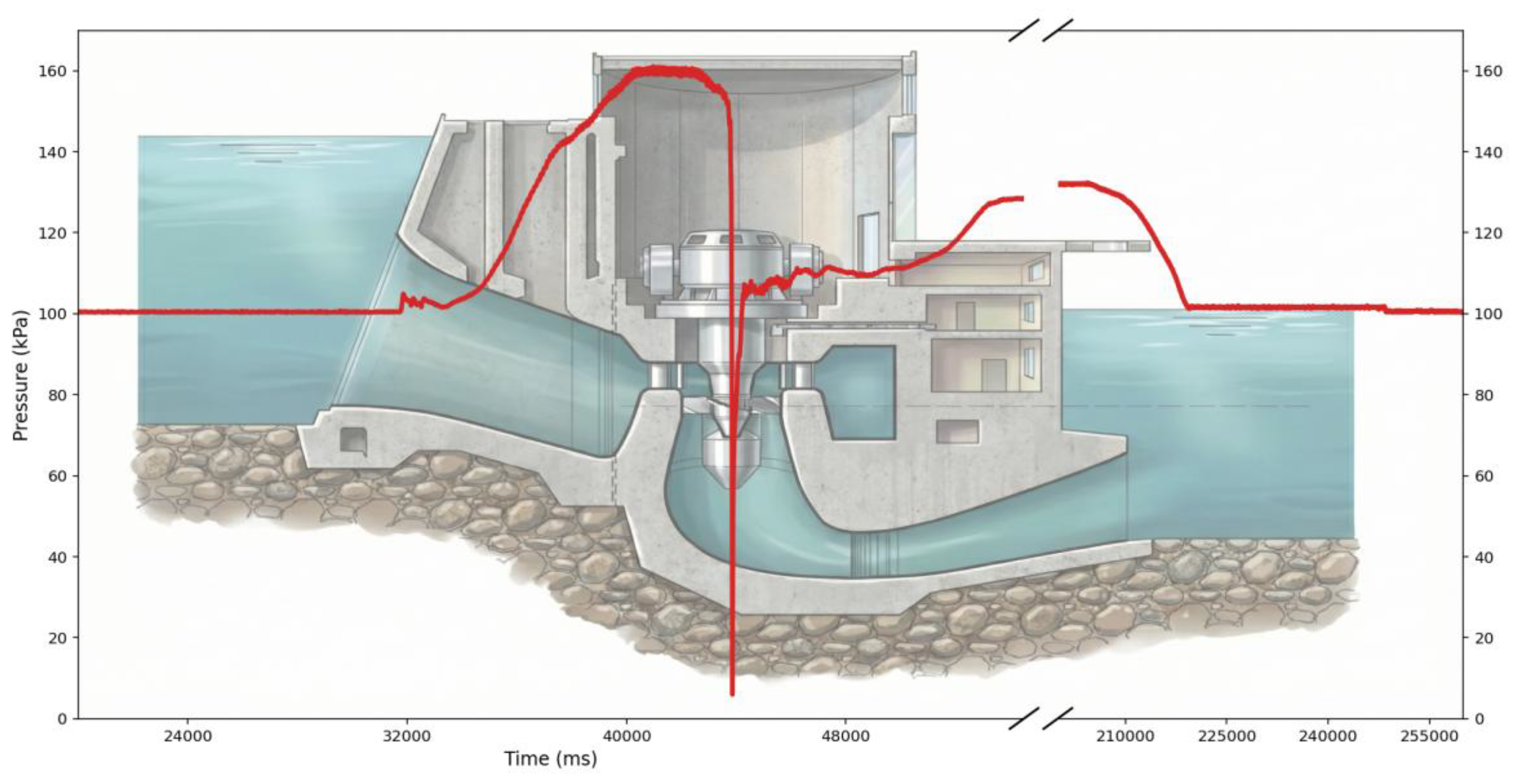

Table 1.
Fish sample (Salmo salar smolts). SD: Standard Deviation.
| Turbine | N | Fork length (mm)* | |
| Mean ± SD | Range | ||
| T1 (Ø: 1.20 m) | 2 | 153 ± 14 | 95 - 190 |
| T2 (Ø: 1.50 m) | 20 | 161 ± 7 | 130 - 190 |
* No statistically significant differences were found between groups (data normally distributed; mean comparison test, p = 0.253).
Table 2.
Estimated mortality (Me), with details of the smolts recovered after passing through the turbines, with details of their condition at the time and after 24 hours, as well as the cause of death and the type of injury.
Table 2.
Estimated mortality (Me), with details of the smolts recovered after passing through the turbines, with details of their condition at the time and after 24 hours, as well as the cause of death and the type of injury.
| Turbine | Fork length | After turbine |
After 24 hours |
Cause of death | Injury type | Me just after turbine passage | Me after 24 hours |
| T1 | 15 | Dead | Turbine passage | Decapitation | 45.5%* (5/11) |
81.8%* (9/11) | |
| 15.8 | Dead | Turbine passage | Decapitation | ||||
| 17.8 | Dead | Turbine passage | Decapitation | ||||
| 16.4 | Dead | Turbine passage | Decapitation | ||||
| 17 | Dead | Turbine passage | Decapitation | ||||
| 19 | Alive | Dead | Turbine passage | Stomach rupture | |||
| 11.5 | Alive | Dead | Turbine passage | Eye embolism | |||
| 18 | Alive | Dead | Undetermined | ||||
| 18.5 | Alive | Dead | Undetermined | ||||
| 9.5 | Alive | Alive | |||||
| 10.5 | Alive | Alive | |||||
| T2 | 17.5 | Dead | Turbine passage | Decapitation | 11.1%* (1/9) |
66.7%* (6/9) | |
| 15.5 | Alive | Dead | Turbine passage | Eye embolism | |||
| 17.8 | Alive | Dead | Turbine passage | Stomach rupture | |||
| 16 | Alive | Dead | Turbine passage | Spinal cord brake | |||
| 19 | Alive | Dead | Undetermined | ||||
| 17 | Alive | Dead | Undetermined | ||||
| 15 | Alive | Alive | |||||
| 18.2 | Alive | Alive | |||||
| 17.6 | Alive | Alive |
* No statistically significant differences were found between turbines either immediately or 24 hours after turbine passage (Fisher’s exact test; p > 0.150).
Table 3.
Mortality (M), Standard Error (SE) and Confidence Interval (IC), based on the two selected Control Mortality values (Mc). Calculated according to Equation (1) and Equation (2).
Table 3.
Mortality (M), Standard Error (SE) and Confidence Interval (IC), based on the two selected Control Mortality values (Mc). Calculated according to Equation (1) and Equation (2).
|
MORTALITY (M ± SE) |
Mc = 5% | Mc = 20% | ||||
| M (%) | SE | IC (95%) | M (%) | SE | IC (95%) | |
| T1* | 80.9 | 10.8 | 56.8 – 100 | 77.3 | 11.1 | 52.6 – 100 |
| T2* | 64.9 | 9.9 | 42.1 – 87.7 | 58.3 | 10.8 | 33.4 – 83.2 |
*No statistically significant differences were detected in final mortality between turbines (Welch’s t-test: If Mc = 5%: t = 1.092; p= 0.290. If Mc= 20%: t = 1.228; p = 0.235).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



