Submitted:
21 December 2025
Posted:
22 December 2025
You are already at the latest version
Abstract
Background: As diets shift towards more plant-based patterns, nutrients mainly supplied by animal-sourced foods are under increased scrutiny, including sulphur-containing amino acids (SCAAs) such as taurine (TAU), methionine (MET) and cysteine (CYS), which support neuroprotection, antioxidant defences and cellular signalling and may therefore be relevant for cognitive health. Method: This systematic review examined the relationship between SCAA supplementation and cognitive function in randomised controlled trials (RCTs). PubMed, EMBASE and PsycINFO were searched in July–August 2024 and updated in September 2025 (PROSPERO: CRD42024574453) for RCTs assessing SCAA supplementation or depletion and cognitive or psychological outcomes. Data were narratively synthesised with explicit consideration of contextual factors (e.g., co-ingested ingredients, exercise state, testing conditions). Results: Eight RCTs (N = 244) were included; all investigated TAU, with no MET or CYS trials. All used acute single-dose interventions, mostly in crossover designs, assessing core cognitive domains and mood. Across trials, acute TAU doses (typically 1–3 g; up to ~50 mg/kg) produced at best small, isolated cognitive benefits, with most outcomes unchanged. Combinations of TAU with caffeine more consistently improved performance but did not clarify TAU’s independent contribution, and effects on mood and wellbeing were generally small, inconsistent and only apparent under specific conditions (e.g., co-ingestion with caffeine, exercise or sleep loss. No trial characterised habitual diet or baseline SCAA status or recruited participants with low animal-source food intake, so cognitive vulnerability under reduced SCAA intake remains untested. Conclusion: Acute TAU trials provide limited evidence of short-term cognitive or mood benefits and do not clarify whether lower SCAA intake in plant-based diets affects cognition or mood. Longer-term, well-powered studies that characterise habitual diet and SCAA status, include MET and CYS, and assess cognition and mood in populations with lower animal-source food intake are needed to determine whether SCAAs should form part of strategies to support brain health as diets shift towards plant-based eating.
Keywords:
taurine
; sulphur-containing amino acids
; cognitive function
; mood
; plant-based diets
; sustainable diets
1. Introduction
Climate change is a significant global challenge facing society. Broadly defined as the alteration in temperature and weather patterns, climate change occurs as a result of human activity either directly or indirectly [1]. If not addressed, climate change will have significant consequences for global temperatures, weather and flooding [2], with further negative impacts on health [3,4], food security [5] and water supplies. Greenhouse gas emissions are the key driver of climate change with approximately a third of all greenhouse gas emissions coming from the food system e.g., Methane produced by cattle digestion, Nitrous Oxide used to fertilise crops etc. [6]. Consequently, shifting populations to more sustainable diets is high on the global sustainability agenda [7]. The EAT-Lancet commission have recommended a healthy and sustainable reference diet, largely consisting of plant-based foods, small amounts of dairy and very little meat [8,9]. However, shifting dietary patterns to reduce or remove meat consumption may have unintended consequences for cognition, as a result of compromising intake of nutrients with key roles in supporting neuroprotection and cognition [10,11,12,13]. Additionally, multiple studies have indicated that following and adapting to plant-based diets result in significantly lower plasma sulphur-containing amino acid levels [14,15,16].
Sulphur containing amino acids (SCAAs) are of particular interest. SCAAs, namely Methionine, Cysteine and Taurine (TAU), are amino acids crucial for maintenance of cellular function and health [17]. Methionine and Cysteine are incorporated into proteins and contribute to the synthesis of Taurine [18]. While Taurine is not incorporated into proteins, it has physiological significance due to anti-inflammatory properties [19] and its role in regulating blood pressure [20] and insulin sensitivity [21]. Specifically, TAU has garnered increased attention due to its potential neuroprotective properties and influence on cognitive function [22,23,24]. In animal models, supplementing TAU led to improvements in memory [14,25]. Additionally in mice, mood disorders such as depression were associated TAU level in the extra cellular fluid of the medial frontal cortex, where TAU concentration was lower in mice exhibiting depressive symptoms [26,27]. A recent study also demonstrated that TAU may be neuroprotective, with lower blood plasma levels of TAU being associated with higher dementia risk level [28]. As such, evidence suggests that TAU, and potentially SCAAs more broadly have importance for cognitive health. While CYS and TAU can be synthesised endogenously [29], the highest concentrations of SCAAs in humans are obtained through animal-based dietary sources [18] including meat, dairy products and fish. Therefore, it is important to consider that shifting dietary patterns towards sustainable, plant-based diets may inadvertently reduce SCAAs intake with potential consequences for cognitive health.
To date, the literature examining the cognitive effects of SCAAs has predominantly focused on acute supplementation, often investigating taurine within the context of energy drinks and at supradietary doses far exceeding typical daily intake (estimated at 40–400 mg) [57]. However, rather than solely representing a limitation, these supradietary levels may be necessary to elicit acute neurocognitive effects, for instance by achieving the concentrations required to activate specific inhibitory receptors (e.g., GABA-A) that are less sensitive to lower physiological levels [49]. Crucially, a distinction must be drawn between the effects of supplementation—aimed at optimizing performance in replete individuals—and the issue of dietary depletion, where the goal is restoring homeostatic levels to prevent decline. While the transition to plant-based diets raises specific concerns regarding the latter, current research largely assesses the former, creating a disconnect between the available evidence and the emerging public health concern.
Therefore, this review was designed to address two primary research questions: 1) Is there evidence regarding the cognitive effects of SCAA restoration in the context of dietary depletion? and 2) Does SCAA supplementation have measurable acute effects on cognitive function and mood? To answer these questions, this review focuses on Randomised Controlled Trials (RCTs) that measure cognitive outcomes (e.g., memory, attention, executive function), and mood (e.g., anxiety, depression, and emotional well-being). Additionally, it aims to identify which dosages, if any, yield the most substantial benefits. Ultimately, this review seeks to clarify the current evidence for SCAAs as cognitive modulators, highlighting critical knowledge gaps that must be addressed to understand the cognitive implications of the global shift toward sustainable diets.
2. Materials and Methods
This systematic review was conducted following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [30] to ensure methodological transparency and reproducibility. The review was developed using the Population, Intervention, Comparison, Outcomes, Context (PICOC) framework [31]. This framework guided the exploration of SCAA efficacy (Intervention) versus placebo control (Comparison) on cognition and mood (Outcomes) in healthy adults (Population), within the contexts of both dietary reduction and varying supplementation states (Context). The review protocol was pre-registered with PROSPERO (ID: CRD42024574453).
2.1. Search Strategy
An initial comprehensive search was carried out across PubMed, EMBASE, and PsycINFO to identify RCTs published in English that investigate the effects of SCAAs on cognition and mood. A manual search of reference lists from relevant articles was conducted to identify additional studies. This search was conducted between July and August 2024 and included all studies up to September 2024. A further update search was conducted on 1st September 2025. Key words for the database searches were identified from existing literature and the search strategy used variations of the key words in conjunction with Boolean operators (AND/OR) to account for differences in spelling. The key words included: sulphur-containing amino acids, sulphuric amino acid, methionine, cysteine, TAU, cognition, memory, attention, executive function, mental performance, neurocognitive, cognitive score, brain function, information processing, reaction time, mood, depression, anxiety, energy.
2.2. Eligibility Criteria
The PICO framework guided the development of the inclusion and exclusion criteria (Table 1.) with additional considerations applied relating to type of publication and publication language.
2.3. Screening and Data Extraction
Two independent reviewers (JM and RT) performed the initial screening of titles and abstracts to identify potentially relevant studies. Full-text articles were then retrieved and assessed against the eligibility criteria by both reviewers. A third reviewer (HY) acted as an arbitrator in cases of disagreement. To ensure fidelity, reviewers met to discuss any disagreements and conflicts were resolved through discussion until full agreement was reached.
The review team developed a data extraction template to ensure comprehensive and systematic extraction of study characteristics. The extracted data included publication details, participant characteristics, study design, intervention details, cognitive and mood outcomes, results and conclusions. Where necessary, the authors of studies were contacted to provide missing or unclear data.
2.4. Risk of Bias Assessment
The risk of bias for each study was assessed independently by reviewers JM and RT using the Cochrane Risk of Bias 2 Tool [32]. Any disagreements were resolved through discussion. The following sources of bias were assessed: randomisation process, deviations from intended interventions, missing outcome data, measurement of the outcome and the selection of the reported result which all contribute to the source for overall bias. Studies were classified as either low risk of bias, some concerns of bias or high risk of bias. Across all studies, there were no classifications for high risk of bias with most sources classed as low risk of bias. Four of the seven studies were marked as some concerns of bias in the selection of the reported result. Therefore, the sources’ reported results were carefully examined to make interpretations for the present study.

Table 2.
Cochrane Risk of Bias 2 assessment for randomisation trials.
| Study | Randomisation Process | Deviations from intended interventions | Missing outcome data | Measurement of the outcome | Selection of the reported result | Overall Bias |
|---|---|---|---|---|---|---|
| Alford et al. [33] |  |
|
|
|
|
|
| García et al. [34] | |
|
|
|
 |
|
| Giles et al. [35] | |
|
|
|
|
|
| Lassiter et al. [36] | |
|
|
|
|
|
| Liu and Rong [37] | |
|
|
|
|
|
| Ozan et al. [38] | |
|
|
|
|
|
| Peacock et al. [39] | |
|
|
|
|
|
| Seidl et al. [40] | |
|
|
|
|
|
Note. ‘+’ = low risk of bias, ‘-‘= high risk of bias, and ‘!’ = some concerns of bias.
2.5. Data Synthesis
A narrative synthesis was performed to summarise the characteristics and findings of the included studies. A meta-analysis was not suitable due to the complex differences in population outcomes, dosages and combinations with differing active ingredients. This narrative synthesis included tabulated data that described each study’s design, intervention details, cognitive and mood measures, and main findings and conclusions. The narrative synthesis identified key trends, inconsistencies, and gaps in the literature regarding the effects of sulphur-containing amino acids on cognition and mood.
3. Results
3.1. Search Results
The PRISMA flowchart (Figure 1.) outlines the study screening and selection process. The initial search returned 2285 studies, of which 155 were identified as randomised controlled trials and moved forward to the abstract and title screening stage. Access was requested for 2 studies which were granted and further screened. After reading the abstracts of each text, 12 studies met the eligibility criteria for full text review. Upon reading of full text, five texts were excluded due to no standardised cognitive tests used (n=3), not double-blinded (n=1) and too many active ingredients (n=1). A total of 7 studies were identified for data extraction and included in this review. The updated search in September 2025 showed an extra study eligible for review, out of a total of 9 additional results.
3.2. Included Studies
While all included studies utilised a validated cognitive test, mood was not assessed in every trial. Across all eligible RCTs, taurine (TAU) was the sole SCAA investigated; no studies utilising methionine or cysteine met the inclusion criteria. Crucially, regarding the review's first objective, no studies were found that assessed cognitive outcomes in the context of dietary depletion or SCAA restoration. Consequently, all eight included studies investigated the acute effects of supplementation in healthy populations.
3.3. Taurine and Cognitive Function
All eight studies were RCTs assessing cognition through standardised tests. N=244 (n range: 10-80), 109 female, 135 male aged 18-45. Cognitive domains assessed were attention (selective, sustained, and divided attention), executive function (cognitive control, and decision-making), memory (working memory & immediate recall), perceptual processing, psychomotor performance, reaction time, and motor control. The findings in relation to the effects of TAU on each of these domains are outlined below.
3.3.1. Attention
Attention was a cognitive domain explored across two studies. Giles et al. [35] used the Attention Network Test to investigate the effects of TAU on alerting, orienting, and executive control components of attention. TAU administered alone showed no significant effect on attention. However, when combined with glucose, there was a significant improvement in attentional control, particularly in tasks requiring orienting attention. Seidl et al. [40] utilised the D2 test of attention to measure selective and sustained attention in a sample of graduate students. Findings demonstrated a combination of caffeine (CAF) (80mg), TAU (1g), and glucuronolactone (600mg) significantly improved sustained attention scores, although the independent effects of TAU alone were not explored. From the limited available data, TAU alone does not appear to significantly improve attention but may work synergistically with other active compounds to enhance attention, particularly during complex, attention-demanding tasks.
3.3.2. Executive Function
Three studies explored the influence of TAU on executive function e.g., cognitive control, inhibition, and decision-making. Ozan et al. [38] explored the effect of TAU on cognitive control via the Stoop test in elite athletes, finding that 3g of TAU alone significantly improved accuracy and reaction times in incongruent Stroop trials, compared to placebo, indicating a selective benefit of TAU supplementation. Interestingly, those who received the combination of TAU and CAF showed improved performance across all Stroop trials, indicating a general effect of enhanced cognitive control and faster, accurate responses. Liu and Rong [37] challenges these claims with contrasting findings which demonstrated CAF to be the driver of improved performance during the Stroop task. Interestingly, they found CAF to significantly improve reaction time for incongruent trials compared to the combination of TAU and CAF, but not for congruent trials. The authors found no evidence to suggest that TAU, on its own, influences cognitive control, even in incongruent trials. Additionally, Lassiter et al. [36] explored the impact of TAU-containing beverages on executive function. The intervention (160 mg CAF, 2 g TAU, 400 mg Panax ginseng) did not produce specific effects on performance; improvements in Go/No Go tests and Stroop reaction times were observed, but as these occurred in both placebo and active groups, they were attributed to exercise rather than the intervention. Overall, the evidence concerning the effects of TAU-based interventions for executive function are mixed; while TAU appeared to selectively enhance inhibitory control on incongruent Stroop trials in some findings (faster reaction times with fewer errors [38]), this finding was not supported by other studies using the same task [37]. Where consistent enhancements were observed, this was largely attributable to combination interventions where CAF was present.
3.3.3. Working Memory & Immediate Recall
Working memory, particularly the ability to temporarily hold and process information, is crucial for complex cognitive tasks. This was examined in two studies. Giles et al. [35] used the N-back Task to assess working memory performance in habitual CAF users. TAU only reduced response time on the verbal N-back in the 1-Back condition in the absence of CAF. Response time was also lower on the object 3-Back condition compared to placebo. Ultimately, this suggests that TAU significantly improves participants’ working memory for both low, and high cognitive load. More precisely, in a verbal N-back, this reflects working memory maintenance/updating efficiency rather than episodic ‘recall’, whereby faster responses without a loss of accuracy indicate more efficient processing. Furthermore, there was no antagonistic effect of TAU and caffeine recorded for working memory. A second study [34] explored effects of three different commercially available energy drinks containing active ingredients on working memory, operationalised using the N-back task. In contrast to findings of Giles, no influence of TAU was observed. Results demonstrated a significant improvement in working memory for those who were in condition A (CAF and glucose, no TAU) compared to those in the control condition. When TAU was combined with CAF and glucose, this significant effect diminished, suggesting an antagonistic effect from TAU.
With regards to immediate memory recall performance, Alford et al. [33] found that short term memory performance improved after supplying their participants with a commercially available energy drink. Findings demonstrated that participants who received the combination of TAU (1g), CAF (80mg), glucose (5.25g) & glucuronolactone (600mg) in a 250ml beverage demonstrated significantly improved immediate recall memory, across three different controlled studies, compared to those who received either a placebo “energy drink”, sparkling water, water, and no drink. However, as TAU was only administered in combination with other active compounds, it is difficult to draw conclusions on its effects on immediate memory recall. In summary, while TAU shows potential for improving performance in memory tasks, study methodologies and findings are heterogeneous, where the effects of TAU appear context-dependent, varying across task complexity and ingredient interactions.
3.3.4. Reaction Time
Several studies examined the effect of TAU on reaction time using a range of cognitive paradigms including the Stroop test, Visual Oddball Task, Stimulus Degradation Task, Reaction Time Task and Choice Reaction Time. When administered in isolation, the effects of TAU were inconsistent. Two studies found that supplementation of TAU alone did not enhance reaction time across any of the reaction time paradigms [37,39]. In one study [35], TAU significantly improved reaction times in the N-back task, indicating superior performance vs placebo. Findings suggest task specific effects where benefits may be more pronounced where cognitive control is required.
Performance during reaction time tasks often improved following a combination of CAF and TAU. Using the Stroop Test, Ozan et al. [38], examined reaction times in elite athletes under anaerobic and aerobic stress. The combination of CAF and TAU significantly improved both reaction times and accuracy, demonstrating their synergistic effect even under physically taxing conditions. Similarly, Alford et al. [33] studied the effects of CAF and TAU on reaction time using the 5-Choice Reaction Time Task. Participants’ who received a combination of CAF and TAU demonstrated a significant reduction in reaction times. In contrast, findings from three studies suggest CAF to be the active ingredient which elicit benefits on reaction time. Liu and Rong [37] found that reaction time reduced during the Stroop task for those who received a combination of TAU with CAF, although CAF alone appeared to have the clearest benefit for reaction times. Peacock et al. [39] assessed reaction times using both the Visual Oddball Task and the Stimulus Degradation Task. Findings demonstrated a limited effect of TAU but rather, CAF alone significantly improved performance under degraded visual conditions. Furthermore, Giles et al. [35] also found that CAF alone significantly improved reaction time across all tasks, further supporting CAF’s robust facilitatory effect.
Lassiter et al. [36] further illustrate how task context and additional components can influence outcomes. In a sample of health trained cyclists, participants completed the Choice Reaction Time Task after consumption of a carbonated energy drink, no differences were found between placebo and the active drink containing CAF, TAU and panax ginseng. The authors reported that faster reaction times were observed after exercise in both active and placebo groups. Overall, findings show that the effects of TAU upon reaction time are context and task dependent, with evidence suggesting further synergistic effects of administration of TAU in combination with other active ingredients.
3.4.. Taurine and Mood
Four studies assessed mood in their RCT’s. N=174 (n range: 10-80) 83 female, 91 male aged 18-35. Only one study investigated the effects of TAU in isolation of other active ingredients [35]. All other studies used combinations of active components. Aspects of mood assessed across studies included alertness, fatigue, well-being, and anxiety.
Giles et al. [35] used the Profile of Mood States (POMS) and a Caffeine Withdrawal Questionnaire to evaluate mood states in habitual CAF consumers. There was a significant effect of TAU on fatigue, but lowered feelings of vigour in absence of CAF, and was associated with heightened withdrawal symptoms of CAF. In contrast, CAF improved vigour, reduced fatigue and alleviated withdrawal symptoms such as headaches. The combination of TAU and CAF generally mirrored the results from CAF, however one interaction showed increased vigour over time. Overall, findings indicate that TAU’s influence on mood was inconsistent and context-dependent, in contrast to the more robust and predictable effects of CAF and its combination with TAU.
Alford et al. [33] measured subjective alertness using a 100mm VAS scale after participants consumed Red Bull. Following supplementation, subjective ratings of alertness significantly increased compared to placebo, while fatigue ratings declined. This suggests that the combination of CAF and TAU contributes to increased energy and alertness levels.
Seidl et al. [40] used the Basler-Befindlichkeitsbogen Questionnaire to evaluate the effects of CAF, TAU, and glucuronolactone on well-being. Participants in the placebo group showed significant declines in well-being, vitality, and social extroversion over the session, whereas those in the active intervention group did not. These findings suggest that the combination may help maintain mood and well-being, particularly during periods of fatigue such as late at night. One study specifically examined the effects of energy drink consumption on anxiety using the State-Trait Anxiety Inventory (STAI).
García et al. [34] conducted a study on medical students to determine the acute effects of energy drinks on mood, including state anxiety, using the STAI. The study divided participants into groups consuming different energy drinks (labelled as A, B, and C) and a control group that consumed carbonated water. Drink A contained 149.5mg CAF and 23g of glucose. Drink B contained 147.2mg CAF, 49.6g glucose and 1.84g of TAU. Drink C contained 155mg CAF, 52.8g of glucose and 1.95g of TAU. The results indicated that Drink C significantly reduced STAI scores post-consumption, suggesting an that Drink C was responsible for significantly reducing anxiety. This reduction implies that some active ingredients may have calming effects, potentially counteracting the stimulating effects of CAF through the interaction of other ingredients. However, the comparative analysis across all groups showed no significant differences in the percent change in STAI scores, indicating that while Drink C had a notable impact on reducing state anxiety, the effect was not uniformly observed with the other Drink conditions. Overall, evidence suggests that TAU’s mood effects are less robust than those of CAF, and its influence may depend on interactions with other compounds.
4. Discussion
This systematic review was designed to address two distinct objectives: 1) to evaluate the cognitive effects of SCAA restoration in the context of dietary depletion, and 2) to determine the acute effects of supplementation on cognition and mood. Critically, regarding the first objective, no eligible trials were identified, leaving the specific risks of SCAA depletion in plant-based transitions currently untested. Consequently, the findings discussed here pertain exclusively to the second objective—acute supplementation. Within this literature, taurine (TAU) was the only SCAA represented, meaning the effects of methionine and cysteine remain undefined. Overall, the findings present a complex and inconsistent picture of TAU supplementation on cognition and mood. While few studies explored TAU in isolation, where this was implemented, TAU elicited domain-specific effects rather than global improvement. Interestingly, enhancements in cognitive performance and mood were often more pronounced when TAU was administered in combination with additional active ingredients, such as caffeine (CAF) or glucose. However, the findings surrounding combination interventions were also inconsistent suggesting that enhancements associated with TAU may be context dependent, with effects influenced by task demands and ingredient synergy.
In the domain of attention, TAU alone did not enhance attentional performance. However, when combined with glucose [35], or with CAF and glucuronolactone [40], enhancements in attentional control, orienting, sustained attention, and selective attention were observed. Findings suggest that TAU’s effects may be contingent upon synergistic interactions between ingredients. Evidence for TAU’s effects on executive function was also largely heterogeneous. One study reported enhanced inhibitory control following TAU supplementation [38], while other studies did not replicate these effects [36,37]. Moreover, although CAF and TAU in combination appeared to enhance cognitive control [38], other findings contrasted this [36,37]. Such discrepancies likely reflect methodological variation, including differences in sample characteristics, cognitive paradigms, and the composition of active interventions.
Findings relating to working memory were similarly heterogeneous and context dependent. Only one study found TAU to enhance working memory capabilities when cognitive demands are high [35], whereas García et al. [34] did not support this finding and reported a possible antagonistic interaction when combined with CAF and glucose where performance diminished. Reaction time effects were the most inconsistent: in some paradigm's TAU alone produced no significant effects [37,39], while in others it enhanced reaction speed under cognitively demanding conditions [35]. Combinations with CAF produced clearer benefits, particularly during exercise or stress, but in several studies, CAF alone accounted for the improvements. Collectively, the literature indicates that TAU supplementation does not consistently enhance cognition in isolation, with observed effects more plausibly explained by interactions with co-administered bioactive compounds or by task-specific contextual factors.
Mood-related outcomes were largely assessed in combination studies, limiting firm conclusions about TAU’s independent contribution. Across trials, TAU with CAF and glucose was associated with increased alertness [33], and reduced state anxiety [34]. When TAU was combined with CAF and glucuronolactone, cognitive performance and mood was significantly improved [40]. However, in the only study to assess TAU alone [35], supplementation influenced ratings of fatigue and lowered ratings of vigour. Divergent outcomes likely reflect variations in study design, intervention duration, and mood assessment sensitivity. Although TAU-containing formulations may modulate mood, evidence does not support a consistent independent effect. Therefore, further controlled trials isolating TAU’s contribution are warranted.
The inconsistent findings may also be partially explained by the neurobiological pathways through which TAU influences psychological outcomes. By modulating inhibitory neurotransmitter systems, such as GABAergic and Glycinergic pathways [41], TAU may influence processes such as arousal, attention and inhibitory control, particularly under conditions of high cognitive load and stress. Importantly, TAU acts on these systems in a dose dependent manner. Lower concentrations of TAU preferentially activate glycinergic pathways, whereas higher concentrations activate GABAA receptors [42]. Context dependent factors such as variation in dose, receptor activation and task demand may explain heterogeneous effects.
On the other hand, the lack of consistent findings may also be attributed to the influence of TAU on cognition occurring through slower-acting, longer-term mechanisms rather than rapid, transient changes associated with acute supplementation. For example, the human body is relatively inefficient in disposing of an accumulation of unstable molecules known as free radicals [43] which, when abundant, can lead to oxidative stress. Oxidative stress has been associated with disorders of the central nervous system [44]. Owing to its powerful antioxidant properties, TAU has been found to attenuate the effects of oxidative stress on cognitive health and even neurodegenerative disorders [45]. Given the established links between TAU and cognitive health, there is substantial scope for future work to examine the long-term effects of TAU supplementation on cognition. This line of research may be especially relevant for certain population groups, such as older adults, who tend to have lower circulating TAU levels [29] and consume less TAU through diet which will be an important area of research for the future.
When the cognitive findings are considered in relation to the conditions under which they were generated, it becomes clear that any apparent effects of TAU are highly context dependent and should be interpreted with caution. Several trials were conducted in athletic populations during, or immediately after, intense exercise bouts. In realist terms, exercise functions as a powerful contextual modifier: it alters cerebral blood flow, arousal, fatigue, and motivation, and triggers hormonal and catecholaminergic responses (adrenaline, noradrenaline, dopamine, norepinephrine) that are themselves capable of changing cognitive performance [46,47,48,49]. These co-occurring changes create distinct context–mechanism–outcome configurations in which TAU is only one element of a much broader physiological state. The type, intensity, and timing of exercise relative to testing further shape these configurations, producing substantial heterogeneity both within and between studies and limiting the extent to which “the same” cognitive domains are truly comparable across trials. Under such conditions, it is difficult to disentangle any independent contribution of TAU from the consequences of the exercise context itself, and any observed benefits are better viewed as arising from specific combinations of context, mechanisms, and tasks, rather than as generalisable drug-like effects of TAU.
Beyond the internal validity of the acute supplementation trials, these findings have to be interpreted against a broader nutritional and sustainability backdrop. Many national and international dietary guidelines now encourage shifts towards more plant-based, environmentally sustainable eating patterns, which typically reduce intakes of animal-sourced foods that are the primary contributors of TAU and other SCAAs [50,51]. Recent expert group guidelines on nutrition and brain health trials also emphasise the importance of characterising background diet and baseline nutrient status when evaluating cognitive effects of specific compounds, and the present literature provides a clear illustration of the limitations that arise when this is not done [52,53]. However, none of the trials in this review assessed habitual diet, baseline SCAA status, or explicitly recruited individuals with low animal-source food intake (Table 3). Although sulphur amino acid restriction paradigms have been explored in both animals and humans, these studies have primarily targeted metabolic health and ageing outcomes, with cognitive endpoints assessed only in selected rodent models under methionine restriction and not in human trials, and none has specifically examined cognitive vulnerability during real-world plant-based dietary transitions [54,55,56,57,58]. Notably, in rodents, methionine or sulphur amino acid restriction consistently reduces adiposity, improves insulin sensitivity and lipid profiles, enhances antioxidant defences, and extends lifespan, effects that appear to be mediated by hormonal and transcriptional adaptations such as increased FGF21 signalling [54,55,56]. Emerging human trials in adults with overweight and obesity indicate that whole-food sulphur amino acid restriction is feasible and can produce modest reductions in fat mass alongside characteristic changes in circulating sulphur metabolites and improvements in selected cardiometabolic risk markers[57,58]. However, these studies have not included cognitive outcomes, and none has modelled plant-based dietary transitions in otherwise healthy populations. As a result, the central question motivating this review—whether lower SCAA intake during plant-based transitions confers cognitive vulnerability that can be mitigated by dietary restoration or supplementation—remains unanswered. Future work will need to move beyond short-term, single-dose paradigms in assumed replete samples and instead focus on longer-term interventions in populations with lower SCAA intakes, incorporating careful assessment of habitual diet, biomarkers, and cognitive outcomes. Such studies are essential if we are to understand how to support cognitive health while progressing towards more sustainable dietary patterns.
This systematic review has several methodological strengths. We implemented a rigorous and transparent protocol, preregistered on PROSPERO, and conducted a comprehensive search across multiple major databases in line with PRISMA guidelines. To maximise internal validity, we restricted inclusion to randomised controlled trials and excluded non-randomised or observational designs. Study screening and selection were undertaken independently by two reviewers, minimising selection bias, and all included trials were subjected to duplicate risk-of-bias assessment, further strengthening the reliability of our conclusions.
Despite these methodological advantages, the conclusions should be interpreted considering several limitations. Firstly, there was substantial heterogeneity among the included studies. While the reviewed evidence and proposed mechanisms suggest the potential for TAU to influence psychological function, it is difficult to draw definitive conclusions regarding TAU’s effects on psychological outcomes from the available data. Moreover, most studies implemented single-dose or short-term interventions using TAU doses exceeding typical dietary intake (estimated at 40–400 mg [59]). This raises concerns about ecological validity and highlights the need for longitudinal studies examining TAU supplementation under more naturalistic conditions. In particular, future work should embed TAU interventions within habitual dietary patterns, including plant-based and plant-forward diets where TAU exposure from animal-sourced foods is lower, to clarify whether any benefits observed at pharmacological doses translate to real-world eating patterns. Longitudinal investigation would better determine the potential cumulative effects of TAU on cognition and mood and more closely replicate human consumption of TAU through diet. In addition, findings may have limited generalisability as important subgroups, such as older adults, were underrepresented. Given that endogenous TAU concentrations available in the body and brain change across the lifespan [29], further research is warranted in diverse demographic groups, such as older adults, who may be particularly susceptible to deficits in TAU and benefit from supplementation. Finally, to maintain a focus on high-quality evidence from randomised controlled trials, we restricted inclusion to peer-reviewed publications and excluded grey literature.
Overall, while the SCAA taurine (TAU) has a plausible biological basis for influencing cognitive function and mood, heterogeneity in study designs, populations, and outcomes means that current evidence does not support firm conclusions about its acute effects. As societies grapple with a shift towards more sustainable, plant-rich diets that may lower exposure to TAU and other sulphur-containing amino acids, clarifying their role in maintaining cognitive health becomes increasingly important. Well-powered, longer-term trials that embed TAU within real-world dietary patterns and explicitly consider background SCAA status are now needed to determine whether preserving adequate SCAA intake should form part of strategies for maintaining brain health within sustainable diets.
Author Contributions
Conceptualization, J.A.M., A.R.G. and H.A.Y.; methodology, J.A.M., A.L.C. and H.A.Y.; software, J.A.M.; validation, J.A.M., A.L.C., R.M.J.T. and H.A.Y.; formal analysis, J.A.M.; investigation, J.A.M., R.M.J.T. and A.L.C.; resources, H.A.Y.; data curation, J.A.M. and R.M.J.T.; writing—original draft preparation, J.A.M.; writing—review and editing, J.A.M., A.L.C., R.M.J.T., A.R.G. and H.A.Y.; visualization, J.A.M.; supervision, A.L.C. and H.A.Y.; project administration, H.A.Y.; funding acquisition, H.A.Y. All authors have read and agreed to the published version of the manuscript.
Funding
Jack Moores and Hayley A. Young receive research funding from Viridian Nutrition for work on taurine. Viridian Nutrition had no role in the conception, design, analysis, or writing of this review. The remaining authors declare no conflicts of interest.
Data Availability Statement
Pre registration and data extraction are available Prospero: CRD42024574453
Acknowledgments
We would like to thank Swansea University for supporting this research.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| SCAA | Sulphur-containing amino acid |
References
- Evseeva, O.; Evseeva, S.; Dudarenko, T. The impact of human activity on the global warming. E3S Web of Conferences, 2021; EDP Sciences. [Google Scholar]
- Hansen, J.E. Global warming has accelerated: are the united nations and the public well-informed? Environment Science and Policy for Sustainable Development 2025, 67(1), 6–44. [Google Scholar] [CrossRef]
- Haines, A. Climate change and human health: impacts, vulnerability and public health. Public health 2006, 120(7), 585–596. [Google Scholar] [CrossRef] [PubMed]
- Patz, J.A. Impact of regional climate change on human health. Nature 2005, 438(7066), 310–317. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.; Von Braun, J. Climate change impacts on global food security. Science 2013, 341(6145), 508–513. [Google Scholar] [CrossRef]
- Crippa, M. Food systems are responsible for a third of global anthropogenic GHG emissions. Nature food 2021, 2(3), 198–209. [Google Scholar] [CrossRef]
- WHO, W.H.O. Sustainable healthy diets: Guiding principles; Food & Agriculture Org, 2019. [Google Scholar]
- Li, Y. Reducing climate change impacts from the global food system through diet shifts. Nature Climate Change 2024, 14(9), 943–953. [Google Scholar] [CrossRef]
- Willett, W. Food in the Anthropocene: the EAT–Lancet Commission on healthy diets from sustainable food systems. The lancet 2019, 393(10170), 447–492. [Google Scholar] [CrossRef]
- Markus, C.R. Effect of different tryptophan sources on amino acids availability to the brain and mood in healthy volunteers. Psychopharmacology 2008, 201, 107–114. [Google Scholar] [CrossRef]
- McAfee, A.J. Red meat consumption: An overview of the risks and benefits. Meat science 2010, 84(1), 1–13. [Google Scholar] [CrossRef]
- Sato, H. Neurodegenerative processes accelerated by protein malnutrition and decelerated by essential amino acids in a tauopathy mouse model. Science advances 2021, 7(43), p. eabd5046. [Google Scholar] [CrossRef]
- Venero, J.L. Changes in neurotransmitter levels associated with the deficiency of some essential amino acids in the diet. British journal of nutrition 1992, 68(2), 409–420. [Google Scholar] [CrossRef] [PubMed]
- Aamer, H. Exploring Taurine's Potential in Alzheimer’s Treatment: A Comprehensive Review. Cureus 2024, 16(5). [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A. Amino acid changes during transition to a vegan diet supplemented with fish in healthy humans . European Journal of Nutrition 2017, 56(5), 1953–1962. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.A. Plasma concentrations and intakes of amino acids in male meat-eaters, fish-eaters, vegetarians and vegans: a cross-sectional analysis in the EPIC-Oxford cohort . European journal of clinical nutrition 2016, 70(3), 306–312. [Google Scholar] [CrossRef]
- Colovic, M.B. Sulphur-containing amino acids: protective role against free radicals and heavy metals . Current medicinal chemistry 2018, 25(3), 324–335. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: an overview . The Journal of nutrition 2006, 136(6), 1636S–1640S. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino acids 2014, 46(1), 7–20. [Google Scholar] [CrossRef]
- Waldron, M. The effects of oral taurine on resting blood pressure in humans: A meta-analysis. Current hypertension reports 2018, 20, 1–8. [Google Scholar] [CrossRef]
- Bae, M.; Ahmed, K.; Yim, J.-E. Beneficial effects of taurine on metabolic parameters in animals and humans. Journal of Obesity & Metabolic Syndrome 2022, 31(2), p. 134. [Google Scholar] [CrossRef]
- Lee, N.-Y.; Kang, Y.-S. Taurine protects glutamate neurotoxicity in motor neuron cells. In in Taurine 10; Springer, 2017. [Google Scholar]
- Leon, R. Protective function of taurine in glutamate -induced apoptosis in cultured neurons. Journal of neuroscience research 2009, 87(5), 1185–1194. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: metabolism, functions, and nutrition . Amino acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-L. Taurine improves the spatial learning and memory ability impaired by sub-chronic manganese exposure. Journal of biomedical science 2014, 21(1), p. 51. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y. Taurine alleviates chronic social defeat stress-induced depression by protecting cortical neurons from dendritic spine loss . Cellular and Molecular Neurobiology 2023, 43(2), 827–840. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yuan, M.; Zhang, W. The major biogenic amine metabolites in mood disorders . Frontiers in Psychiatry 2024, 15, 1460631. [Google Scholar] [CrossRef]
- Chouraki, V. Association of amine biomarkers with incident dementia and Alzheimer's disease in the Framingham Study . Alzheimer's & Dementia 2017, 13(12), 1327–1336. [Google Scholar] [CrossRef]
- Singh, P. Taurine deficiency as a driver of aging . Science 2023, 380(6649), p. eabn9257. [Google Scholar] [CrossRef]
- Page, M.J. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews . bmj 2021, 372. [Google Scholar]
- Richardson, W.S. The well-built clinical question: a key to evidence-based decisions . ACP journal club 1995, 123(3), A12–3. [Google Scholar] [CrossRef]
- Sterne, J.A. RoB 2: a revised tool for assessing risk of bias in randomised trials . bmj 2019, 366. [Google Scholar] [CrossRef]
- Alford, C.; Cox, H.; Wescott, R. The effects of red bull energy drink on human performance and mood . Amino acids 2001, 21, 139–150. [Google Scholar] [CrossRef]
- García, A. Acute effects of energy drinks in medical students . European journal of nutrition 2017, 56, 2081–2091. [Google Scholar] [CrossRef] [PubMed]
- Giles, G.E. Differential cognitive effects of energy drink ingredients: caffeine, taurine, and glucose . Pharmacology Biochemistry and Behavior 2012, 102(4), 569–577. [Google Scholar] [CrossRef]
- Lassiter, D.G. Effect of an energy drink on physical and cognitive performance in trained cyclists . Journal of Caffeine Research 2012, 2(4), 167–175. [Google Scholar] [CrossRef]
- Liu, J.; Rong, W. Effects of taurine combined with caffeine on repetitive sprint exercise performance and cognition in a hypoxic environment . Scientific Reports 2025, 15(1), 5386. [Google Scholar] [CrossRef] [PubMed]
- Ozan, M. Does single or combined caffeine and taurine supplementation improve athletic and cognitive performance without affecting fatigue level in elite boxers? A double-blind, placebo-controlled study . Nutrients 2022, 14(20), 4399. [Google Scholar] [CrossRef]
- Peacock, A.; Martin, F.H.; Carr, A. Energy drink ingredients. Contribution of caffeine and taurine to performance outcomes . Appetite 2013, 64, 1–4. [Google Scholar] [CrossRef]
- Seidl, R. A taurine and caffeine-containing drink stimulates cognitive performance and well-being . Amino acids 2000, 19, 635–642. [Google Scholar] [CrossRef]
- Song, N.-y. Interaction between taurine and GABAA/glycine receptors in neurons of the rat anteroventral cochlear nucleus . Brain research 2012, 1472, 1–10. [Google Scholar] [CrossRef]
- Jia, F. Taurine is a potent activator of extrasynaptic GABAA receptors in the thalamus . Journal of Neuroscience 2008, 28(1), 106–115. [Google Scholar] [CrossRef]
- Ripps, H.; Shen, W. taurine: a “very essential” amino acid. Molecular vision 2012, 18, 2673. [Google Scholar]
- Di Monte, D.A.; Chan, P.; Sandy, M.S. Glutathione in Parkinson's disease: a link between oxidative stress and mitochondrial damage? Annals of Neurology Official Journal of the American Neurological Association and the Child Neurology Society 1992, 32(S1), S111–S115. [Google Scholar] [CrossRef] [PubMed]
- Menzie, J. Taurine and central nervous system disorders . Amino acids 2014, 46(1), 31–46. [Google Scholar] [CrossRef] [PubMed]
- Box, A.G. Individual differences influence exercise behavior: how personality, motivation, and behavioral regulation vary among exercise mode preferences . Heliyon 2019, 5(4). [Google Scholar] [CrossRef] [PubMed]
- Ide, K.; Secher, N.H. Cerebral blood flow and metabolism during exercise. Progress in neurobiology 2000, 61(4), 397–414. [Google Scholar] [CrossRef]
- Lambourne, K.; Tomporowski, P. The effect of exercise-induced arousal on cognitive task performance: a meta-regression analysis . In Brain research; 2010; Volume 1341, pp. 12–24. [Google Scholar]
- McMorris, T.; Tomporowski, P.; Audiffren, M. Exercise and cognitive function; John Wiley & Sons, 2009. [Google Scholar]
- Young, H.A.; Freegard, G.; Benton, D. Mediterranean diet, interoception and mental health: Is it time to look beyond the ‘Gut-brain axis’? Physiology & Behavior 2022, 257, 113964. [Google Scholar] [CrossRef]
- Romijn, A.R. Perspective: advancing dietary guidance for cognitive health—focus on solutions to harmonize test selection, implementation, and evaluation . Advances in Nutrition 2023, 14(3), 366–378. [Google Scholar] [CrossRef]
- Young, H.A. Multi-nutrient interventions and cognitive ageing: are we barking up the right tree? Nutrition Research Reviews 2023, 36(2), 471–483. [Google Scholar] [CrossRef]
- Taylor, R.M.J. Unveiling Dietary Complexity: A Scoping Review and Reporting Guidance for Network Analysis in Dietary Pattern Research . Nutrients 2025, 17(20), 3261. [Google Scholar] [CrossRef]
- Perrone, C.E. Metabolic adaptations to methionine restriction that benefit health and lifespan in rodents . Exp Gerontol 2013, 48(7), 654–60. [Google Scholar] [CrossRef]
- Dong, Z.; Sinha, R.; Richie, J.P., Jr. Disease prevention and delayed aging by dietary sulfur amino acid restriction: translational implications . Annals of the New York Academy of Sciences 2018, 1418(1), 44–55. [Google Scholar] [CrossRef]
- Lail, H. Effects of Dietary Methionine Restriction on Cognition in Mice . Nutrients 2023, 15(23), 4950. [Google Scholar] [CrossRef]
- Stolt, E. Sulfur amino acid restriction, energy metabolism and obesity: a study protocol of an 8-week randomized controlled dietary intervention with whole foods and amino acid supplements . J Transl Med 2021, 19(1), p. 153. [Google Scholar] [CrossRef]
- Olsen, T. Dietary sulfur amino acid restriction in humans with overweight and obesity: Evidence of an altered plasma and urine sulfurome, and a novel metabolic signature that correlates with loss of fat mass and adipose tissue gene expression . Redox Biol 2024, 73, 103192. [Google Scholar] [CrossRef]
- Santulli, G. Functional role of taurine in aging and cardiovascular health: an updated overview . Nutrients 2023, 15(19), 4236. [Google Scholar] [CrossRef]
Figure 1.
PRISMA flow chart of the screening process.
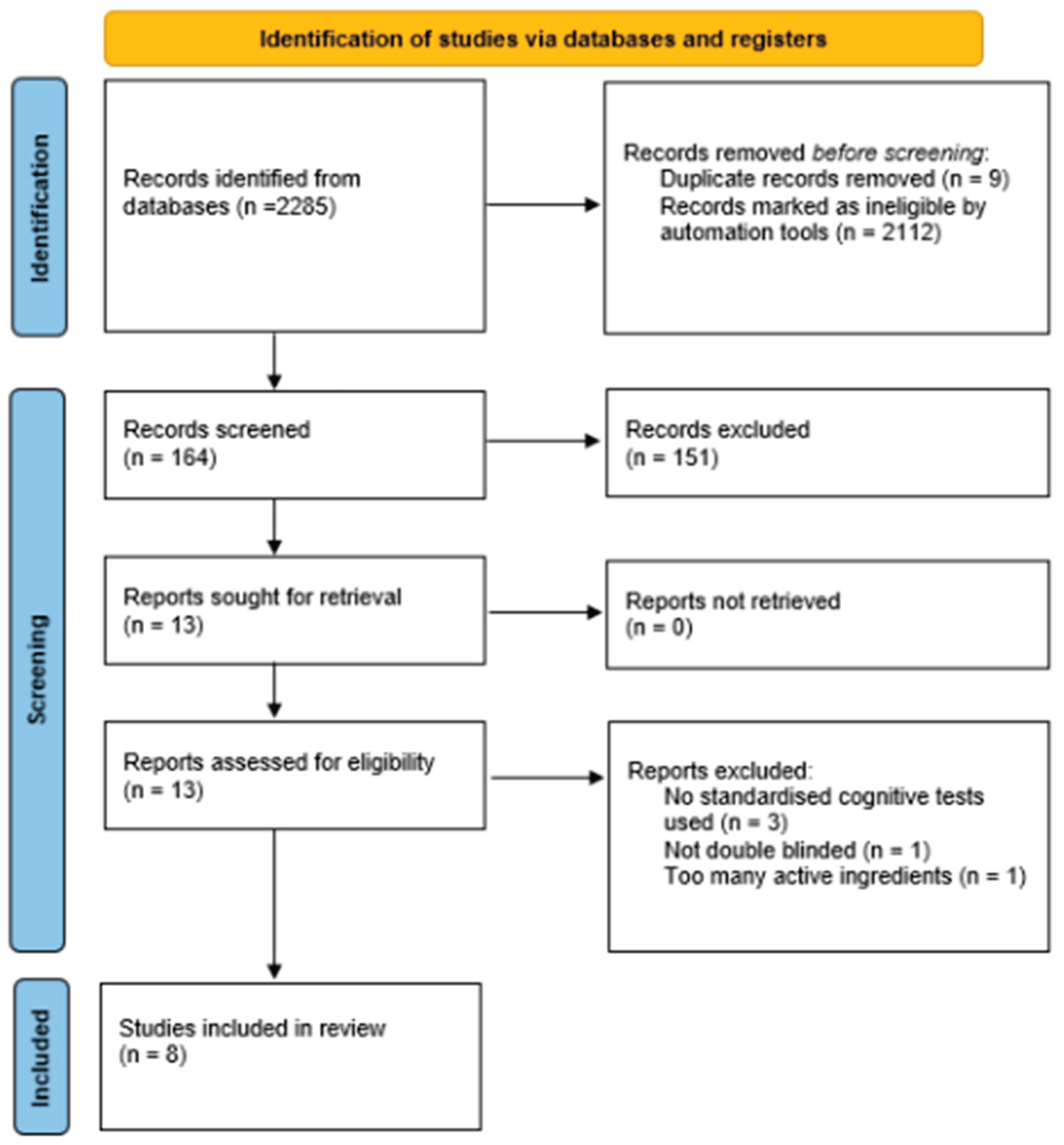
Table 1.
Inclusion and exclusion criteria applied during study selection.
| Inclusion | Exclusion | |
|---|---|---|
| Population | Studies involving healthy adults aged 18 years and over were eligible. No restrictions were placed on gender, ethnicity, or geographical location. | Animal studies, as well as those involving children or participants with neurodegenerative diseases or metabolic disorders, were excluded to maintain relevance to the primary population of interest. |
| Intervention | Supplementation of one or more SCAA (e.g., taurine, methionine, or cysteine), administered alone or in combination with up to four ingredients (e.g., caffeine, glucose). | Studies using over four total ingredients in the intervention, or those including medications as comparators, were excluded to avoid confounding effects. |
| Comparison | Must include a placebo control group. | Does not include a placebo control group. |
| Outcomes | Studies must include outcomes related to cognition (e.g., memory, attention, reaction time, executive function) and/or mood (e.g., anxiety, depression, emotional well-being), measured through validated scales or cognitive tests. | Studies which did not focus on facets of cognitive function and / or mood as outcomes. |
| Context | Studies conducted in contexts of either: 1. Dietary Depletion/Restoration: (e.g., low habitual SCAA intake, plant-based diet transitions). 2. Acute Supplementation/Enhancement: (e.g., supradietary dosing in replete individuals). |
Contexts involving clinical malnutrition, disease-related deficiency, or recovery from surgery/trauma. |
| Study type | Peer reviewed, double-blind, placebo-controlled RCTs. | Non-peer reviewed, non-randomised trials, cohort or case-control studies, observational designs, and any studies lacking a placebo control were excluded. |
| Publication Language | English language. | Published not using English Language. |
Table 3.
Characteristics of included studies, including population, aims, design, intervention (with dosages), outcome measures (with cognitive tests), mood measures, main findings, and authors’ conclusions.
Table 3.
Characteristics of included studies, including population, aims, design, intervention (with dosages), outcome measures (with cognitive tests), mood measures, main findings, and authors’ conclusions.
| Reference | Population | Aims | Design | Intervention & dosage | Cognitive tests | Mood Measures | Main Findings | Habitual diet / SCAA status assessed? | Authors Conclusions |
|---|---|---|---|---|---|---|---|---|---|
| Alford et al. [33] | 3 studies* Study 1: N=10 (5f, 5m) aged 18-30 Study 2: N=14 (7f, 7m) aged 18-35 Study3: N= 12 (5f, 7m) aged 20-21 Total N=36 (17f, 19m) aged 18-35 healthy, moderate CAF users |
To investigate the effects of Red Bull Energy Drink on physical endurance (aerobic and anaerobic), psychomotor performance (reaction time, concentration, memory), subjective alertness, and mood. | Double-blind, repeated-measures, randomised crossover across three separate studies. Each study completed within a 4-week period with a one week break for each participant between their two testing sessions. Participants received Red Bull, water (still or carbonated), or a PLA (Still water replaced carbonated water in the 3rd study). |
All studies used a single dose combo of Red Bull (250ml) which included: TAU 1g, CAF 80mg, Glucose 5.25g, Glucuronolactone 600mg Study 1 PLA = carbonated water Study 2 PLA = carbonated water OR no drink control Study 3 PLA = Still water OR "Dummy Energy drink" (flavoured carbonated water) |
5-Choice reaction time ‘Concentration Task’ Immediate Recall Memory Task |
VAS scale - 100mm | Red Bull significantly improved choice reaction time, concentration, and immediate recall compared to control drinks and PLA. Participants reported increased alertness after consuming Red Bull compared to PLA and control drinks. |
No – only caffeine-use status (moderate CAF users) recorded; no assessment of habitual diet or SCAA intake | Red Bull Energy Drink improves both mental and physical performance, including reaction time, memory, concentration, and endurance. These effects are attributed to the combined ingredients, CAF, TAU, and glucose. The drink also increases subjective alertness without significant cardiovascular side effects at rest. Authors commend TAU for “other” positive effects on mood |
| García et al. [34] | N=80 healthy medical students (50m, 30f) mean age 21.45 All participants had consumed energy drinks in their lifetime |
To determine the acute effects of different energy drinks on cardiovascular parameters, stress levels, and working memory in medical students. | Double-blinded, randomised placebo-controlled trial. 4 groups: Control group (carbonated water) Groups A, B & C were commercially available energy drinks Tests were conducted before and after consumption of intervention. |
Single dose Energy drink intervention: All drinks 460ml A: CAF = 149.5mg, Glucose=23g, TAU=0g B: CAF=147.2mg, Glucose=49.6, TAU=1.84g C: CAF= 155mg, Glucose=52.8g, TAU= 1.95g Control: carbonated water |
N-back Task | State-Trait Anxiety Inventory (STAI) | Group A showed an increase in working memory performance (no TAU) compared to the control, but no significant differences between groups were found. The STAI test showed a decrease in anxiety in group C. |
No – prior energy drink exposure noted; no systematic assessment of habitual diet, TAU, or SCAA intake | The results highlight the variability of energy drink effects on physiological and cognitive functions, likely due to differing compositions of the drinks. Authors suggest anxiety reduction could be due to ingredient composition. |
| Giles et al. [35] | N=48 Habitual CAF consumers, 18M, 30F Good health, CAF consumers (200 mg/day+), non-smokers, No use of prescription medication except for oral contraceptives. |
To evaluate the individual and combined effects of CAF, TAU, and glucose on cognitive performance and mood in habitual CAF consumers who were CAF-deprived for 24 hours. | Double blind mixed design Within subject, 4 conditions, 3-day washout: Within-participants factors: CAF and TAU treatment. Between-participants factor: glucose treatment or PLA. |
Single dose, combined and PLA separated by a 3-day washout: PLA= 0 CAF+ 0 TAU TAU= 0 CAF+ 2000mg TAU CAF= 200mg CAF+ 0mg TAU CAF*TAU= 200mg + 2000mg Between group factor: Half participants administered 50g Glucose (250mlGLU+sparkling water) or PLA (250ml sparkling water+250ml PLA) |
Attention (alerting, orienting, Executive control), Reaction Time, Working Memory & Psychomotor Performance Attention Network Test (ANT) N-back Task Reaction Time Task (RTT) |
Mood states, CAF withdrawal symptoms Profile of Mood States (POMS) Withdrawal Questionnaire (WQ) |
TAU increased choice reaction time accuracy and improved reaction times in particular working memory tasks (verbal and object N-back). TAU+GLU increased orienting attention Glucose improved object working memory in combination with CAF CAF improved executive control, working memory, and reduced reaction times. It also increased tension, vigour, and reduced fatigue and withdrawal symptoms. |
No – caffeine consumption used as inclusion criterion; no assessment of habitual diet or SCAA intake | TAU had inconsistent effects on mood and cognitive performance. CAF was the main driver of cognitive performance improvements, particularly in attention, working memory, and psychomotor performance. Glucose had limited effects on cognitive performance, and its interaction with CAF and TAU requires further research. |
| Lassiter et al. [36] | N= 15 healthy, trained cyclists (7f, 8 m), aged 20-45 years. | To evaluate the effect of an ED containing CAF, carbohydrates, TAU, and Panax ginseng on cycling time-trial performance and cognitive performance at rest, during exercise, and after exercise. |
Double-blind, placebo-controlled, randomised, crossover repeated measures. Each participant completed two experimental trials separated by 6-21 days Participants consumed either the ED or a PLA following a 12-hour fast and CAF abstention followed by a 35km cycling time trial course after intervention. |
Single dose Energy drink intervention: 480 mL containing 54 g carbohydrate, 160 mg CAF, 2 g TAU, 400 mg Panax ginseng PLA = 480mL 0 kcal, CAF-free, no herbal or amino acids. |
Choice Reaction Time Task Go/no-go task (executive function) Stroop Test Tapping task - taps per second psychomotor control test |
N/a | Improved performance was observed on the executive function task and reduced movement times after the race in both the choice reaction and executive function tasks, this was a time effect, PLA also improved post-race. Stroop test reaction times improved post-race but showed no significant treatment effects. Energy drink intervention increased taps per second in the tapping task both pre- and post-exercise compared to PLA. |
No – trained athlete status reported; no habitual diet or SCAA intake assessed | The energy drink enhanced both aerobic performance and certain aspects of cognitive function (tapping speed, executive function) during and after exercise. |
| Liu and Rong [37] | N=16 healthy male university footballers (mean 23.7 y) |
Assess acute effects of TAU, CAF, and TAU+CAF on cognition (Stroop) and exercise performance under hypoxia | Double-blind randomised placebo-controlled crossover RCT 4 groups : PLA, CAF, TAU, TAU*CAF Tests were 60 min after ingestion, 3-day washout period |
Single dose and combination, 3-day washout CAF = 5mg/kg TAU = 50mg/kg TAU*CAF= 50+5mg/kg PLA= maltodextrin 5mg/kg Stroop was administered after a physical warm up (BL), after an exhaustion test (MID), and after intense sprinting (END). |
Stroop task | N/a | CAF improved reaction time vs PLA for congruent and incongruent Stroop trials, no change in accuracy. TAU alone showed no significant results For incongruent and congruent trials, CAF had significantly faster RT than both TAU and PLA. For incongruent trials, CAF improved RT vs TAU*CAF. |
No – no habitual diet or SCAA intake reported | CAF is the primary driver of for cognitive enhancement in Stroop task performance. TAU alone or with CAF is ineffective did not enhance performance during the Stroop task |
|
Ozan et al. [38] |
N=20 male, elite boxers (>10 years’ experience) 18-24 years old (M=22.14±1.42) |
To evaluate the effects of CAF, TAU, and their combination CAF*TAU compared to PLA on athletic performance and exercise-induced fatigue cognitive performance levels. | Double-blind randomised crossover. Four conditions: CAF, TAU, CAF*TAU, PLA All conditions met by each participant in a 72-hour period. |
Single dose, combination, and PLA within 72 hours window. TAU=3g CAF= 6mg/kg CAF*TAU= 6mg/kg + 3g PLA = 300mg of Maltodextrin |
Reaction times and accuracy Stroop test |
N/a | CAF*TAU improved cognitive reaction times and accuracy compared to PLA. TAU significantly improved incongruent Stroop trial accuracy and incongruent trial reaction times vs PLA |
No – no habitual diet or SCAA intake reported | Co-ingesting CAF and TAU improves anaerobic performance, balance, agility, and cognitive function in elite male boxers more effectively than either supplement alone or PLA. |
| Peacock et al. [39] | N=19 right-handed females 19-22 years old (M=20.8) |
To investigate the independent and combined effects of CAF and TAU on behavioural performance, specifically reaction time. | Double-blind, placebo-controlled, crossover design. Four counterbalanced conditions: PLA, TAU, CAF, CAF*TAU participant’s sessions were separated by a 2-7-day washout period |
Single dose, combined and PLA separated by 2-7-day washout TAU=1g CAF=80mg PLA=matched to active counterparts' weight with cornflour |
Reaction times (Visual Oddball Task) Stimulus Degradation Task: Measures reaction time to identify digits at three levels of visual degradation (intact, low degradation, high degradation). |
N/a | Non-significant effects of TAU for reaction times in either task. No significant effects of CAF on visual oddball task. CAF significantly improved reaction times in the stimulus degradation task compared to PLA. CAF*TAU did not enhance reaction times compared to CAF. TAU may attenuate CAF's beneficial effects on reaction time. |
No – no habitual diet or SCAA intake reported | Treatments are task dependent. TAU did not have significant independent effects on reaction time and may attenuate CAF’s performance-enhancing effects in particular tasks. The interaction between CAF and TAU requires further research to understand its impact on performance outcomes |
|
Seidl et al. [40] |
N=10 graduate students (23.9 years old ±2.5) 6 female, 4 male Regular CAF consumers n=5 Non-CAF consumers n=5 All healthy, non-smokers |
To evaluate the combined effects of CAF, TAU, and glucuronolactone (CTG) on cognitive performance and mood (replicated quantities of Red Bull drink). To test whether cognitive and mood effects of these ingredients occur at night, when participants are expected to be more fatigued. |
Double-blind, placebo-controlled, crossover, repeated-measures design. Two test sessions separated by at least one week. Participants were randomly assigned to receive either CTG or PLA (wheat-bran capsules) and then switched for the other session. All participants had abstained from CAF and alcohol for at least 24 hours before the test. |
Single dose : 1g TAU, 80 mg CAF + 600mg glucuronolactone across 7 capsules Capsules taken with 250ml water |
D2 test of attention P300 ERP wave |
Basler-Befindlichkeitsbogen Questionnaire |
CAF, TAU, glucuronolactone combination improved RT and D2 attention scores vs PLA P300 latency slowed in PLA. CAF, TAU, glucuronolactone group showed non-significant shorter P300 latencies in comparison with pretreatment. PLA experienced significant decline in well-being, vitality, and social extroversion by the end of the session. The active intervention group did not show this decline. |
No – CAF-user vs non-user status recorded; no broader habitual diet or SCAA measures | The combination of CAF, TAU, and glucuronolactone (CTG) significantly improves cognitive performance and mood, especially during periods of fatigue (late night). These effects are not merely due to reversing CAF withdrawal, as non-CAF users benefited similarly to CAF users. |
*TAU = Taurine **CAF = Caffeine ***PLA = Placebo
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



