Submitted:
05 December 2025
Posted:
08 December 2025
You are already at the latest version
Abstract
The extended application of pesticides has intensified the resistance problem in Liriomyza trifolii within Hainan Province. This study aimed to elucidate the underlying mechanisms contributing to the elevated resistance observed in this pest by employing whole-genome resequencing (WGS) technology. Through the analysis and comparison of WGS data focusing on Voltage-gated sodium channels (VGSC) from diverse regions and sensitive strains of L. trifolii in Hainan Province. A total of six nonsynonymous single nu-cleotide polymorphisms (nsSNPs) and thirty-one synonymous single nucleotide poly-morphisms (sSNPs) were detected in the five field populations MY, TS, DA, TY, and JY. Among the six nsSNPs, three (PyR1: M918T, L1014F, and PyR2: T933I) have been con-firmed as linked to pyrethroids resistance, while one (D IVS6: V1845I) was associated with resistance to indoxacarb. Moreover, the frequency of these four mutations generally in-creases with decreasing latitude. Additionally, under sustained pesticide selection pres-sure, L. trifolii exhibits an evolutionary pattern characterized by a dN/dS ratio (nsSNP/sSNP = 6/31 ≈ 0.19) of less than 1. Among the 31 sSNPs that held an absolute quantitative advantage, the highest occurrence frequency reached 94.44 % (G2033: JY), and this sSNP occurred in all populations. In contrast, among a limited number of 6 nsSNPs, the highest occurrence frequency attained 100% (L1014F: all populations). This study sub-stantiates that the elevated resistance observed in L. trifolii within Hainan Province can be ascribed to the presence of four nsSNPs-M922T, T933I, L1018F, and V1845I in their VGSC. Furthermore, the emergence of cross-resistance between pyrethroids and indoxacarb has been identified. This research offers a novel theoretical foundation for future investigations into the resistance mechanisms of L. trifolii.
Keywords:
Liriomyza trifolii
; whole-genome resequencing
; voltage-gated sodium channels
; sSNPs
; resistance
1. Introduction
Liriomyza trifolii (Burgess), an invasive pest species in China, initiated its global dissemination from Florida, USA, in 1968 [1]. Currently, the distribution of the L. trifolii is primarily concentrated in the southeastern coastal regions of China, encompassing Hainan, Guangdong, Guangxi, Shanghai, Fujian, Zhejiang, Jiangsu, and Taiwan. L. trifolii exhibits a broad host range, impacting crops such as legumes, Solanaceae, Cucurbitaceae, and cruciferous plants [2], with documented evidence of host range expansion [3]. This pest is distinguished by its diminutive size, high reproductive capacity, brief development cycle, and rapid spread dispersal, frequently resulting in overlapping generations, which predisposes it to outbreaks and substantial agricultural damage [4]. In warmer regions, L. trifolii can cause continuous damage throughout the year due to overlapping generations.
Hainan Province, located in southern China, functions as the principal distribution hub for winter fruits and vegetables in the region, with an annual cultivation area exceeding 60,000 hectares. That’s a significant challenge for agriculture in Hainan. The year-round cowpea cultivation creates a perfect storm for L. trifolii populations, essentially providing a continuous host and breeding ground. This, combined with the extended and frequent pesticide exposure, creates a classic scenario for the rapid development of pesticide resistance. Currently, both domestically and internationally, chemical pesticides remain the primary method for controlling the L. trifolii. The widespread and excessive application of pesticides has led to a continuous rise in field population resistance.
Pyrethroids are a significant class of neurotoxic pesticides [5]. Their mode of action involves disrupting the function of the insect’s peripheral and central nervous systems [6]. Specifically, pyrethroids target voltage-gated sodium channels (VGSC) as their main site of activity [7]. Mutations at VGSC sites can lead to knockdown resistance (kdr), the primary mechanism of pyrethroid resistance, by reducing the pesticide’s affinity for its target [8,9]. This same VGSC target is also exploited by other insecticides, including indoxacarb and flufenidin [10,11,12,13]. VGSCs are comprised of four homologous internal domains (I–IV), each containing six transmembrane α-helices (S1–S6) (Figure S1). These domains are interconnected by intracellular linkers, while the helices are linked by either intracellular or extracellular loops [14]. The homologous channel model suggests that pyrethroids and VGSCs possess two distinct binding regions: PyR1 (II L45-II S5-III S6) [15] and PyR2 (I L45-I S5-I S6-IIS6) [16]. Moreover, studies on mammals have demonstrated that the drug Cenobamate binds to the canonical human Nav1.5 ion channels by interacting primarily with S6 in domains II and III, along with a sole residue (Ile 1768) in domain IV, which differs slightly from insects [17]. Research has extensively focused on mutations within these binding regions, as such mutations are likely to diminish the sensitivity of insect VGSCs to pyrethroids, thereby contributing to increased insect resistance to these pyrethroids.
Whole-genome resequencing (WGS) is a process that swiftly and accurately identifies all the “differences” between a specific individual and the “standard map” (reference genome) based on the existing “map” information. WGS has emerged as a prominent application in Next-Generation Sequencing (NGS). Relevant studies have conducted genotyping on populations exhibiting varying levels of pyrethroids resistance in northern Brazil, focusing on genetic polymorphisms within the genomes of Aedes aegypti with distinct characteristics. The allele frequencies across the entire genome were assessed using SNP chips, leading to the identification of single nucleotide polymorphisms (SNPs) directly associated with resistance, as well as one superior SNP pair [18]. WGS of Spodoptera frugiperda from the Indochinese Peninsula to northern China (2019–2023) identified eight stable genomic markers (six SNPs, two InDels) that clearly distinguish eastern and western populations. This work established a reliable molecular toolkit for tracing the origin of individual moths [19].
This study employed WGS to investigate the underlying mechanisms of high resistance in L. trifolii populations from Hainan. By comparing the VGSC across various geographic strains and a susceptible reference, four critical VGSC mutations were identified. These mutations confer high levels of resistance and cross-resistance to pyrethroids and indoxacarb, offering crucial molecular targets for future resistance management strategies.
2. Results
2.1. Complete Genomic Information of VGSC in L. trifolii
Following the analysis of first-generation sequencing results, the complete genomic sequence of the VGSC of L. trifolii was determined to be 36,774 base pairs (bp). This sequence comprises specific lengths for Domain I (6,696 bp), Domain II (10,162 bp), Domain III (11,025 bp), and Domain IV (8,891 bp). Within the entire genomic sequence, the intron and coding sequence (CDS) lengths are 30,360 bp and 6,414 bp, respectively (Figure S2). The focused 6,414 bp CDS segment contains a total of 2,138 amino acids, including the stop codon. This segment encodes 415, 614, 531, and 578 amino acids for Domain I, Domain II, Domain III, and Domain IV, respectively (Figure 1).
2.2. Information of Exonic Synonymous sSNP in L. trifolii
Based on codon degeneracy, the 31 identified sSNPs were classified as L145L, I290I, S319S, Y354Y, A470A, L474L, E496E, E505E, E661E, A685A, L733L, L778L, T814T, Y866Y, N931N, S978S, A1112A, L1231L, D1232D, P1282P, F1372F, I1471I, Y1483Y, H1996H, Q2006Q, R2011R, G2022G, G2033G, A2038A, G2040G, and A2050A. These sSNPs are primarily distributed across the following regions: “D I -S1”, “D I -L-S56”, “L-D I - II”, “D II -S1”, “D II -L-S23”, “D II -S5”, “D II -L-S56”, “L-D II - III”, “D III -S3”, “D III -L-S56” and “L-D IV <“. The positions of these sSNPs within the VGSC, along with the nucleotide alterations and occurrence frequencies, are detailed in Table 1. Notably, 14 sSNPs were found in all five field populations (MY, TS, DA, JY, TY), specifically L145L, I290I, S319S, E496E, E505E, L733L, N931N, S978S, A1112A, P1282P, Q2006Q, R2011R, G2033G, and A2050A. Among these, Q2006Q exhibited the highest occurrence rate at 94.31%, while N931N had the lowest at 7.08%. Furthermore, both the highest and lowest frequencies were observed in the JY population. Of the 31 sSNPs, A470A, L1231L, D1232D, I1471I, Y1483Y, A2038A, and G2040G were found exclusively in a single population, with frequencies of 8.57% (TS), 11.76% (DA), 12.24% (DA), 10.10% (TS), 16.19% (TS), 7.04% (TS), and 8.33% (TS), respectively (Figure 2).
2.3. Information of Exonic nsSNP in L. trifolii
After conducting multiple comparisons and analyses of the CDS region in the genome resequencing data, our results identified 6 nsSNPs in the VGSC of L. trifolii: M922T (918), T933I, L1018F (1014), Q1285H, V1845I (1851), and D2036E. These nsSNPs are primarily situated in specific regions of the VGSC: D II -L-S45 (M922T), D II -S5 (T933I), D II -S6 (L1018F), L-D II - III (Q1285H), D IV -S6 (V1845I), and L-D IV < (D2036E) (Figure 3A). The positions, nucleotide alterations, and frequencies of these nsSNPs in the VGSC are detailed in Table 2. Notably, 3 nsSNPs, namely M922T, L1018F, and V1845I, were present across all five field populations (MY, TS, DA, JY, TY). Among these, L1018F exhibited the highest occurrence rate of 100.00%, which was consistent across all five populations. M922T exhibits the highest incidence rate in JY (90.91%) and the lowest in MY (58.46%). Similarly, V1845I shows the highest frequency in JY (96.52%) and the lowest in MY (36.54%), mirroring the pattern observed for M922T. T933I and D2036E are unique to the TS population among the 6 nsSNPs. The frequencies of Q1285H and V1845I in the DA and JY populations are identical at 80.00% and 96.52%, respectively. Figure 3B illustrates the frequency comparison of various nsSNPs across populations in a north-to-south order based on decreasing latitude.
3. Discussion
The high level of resistance and unique reproductive capabilities are the primary factors contributing to the L. trifolii ‘s status as a significant pest [20]. For decades, deltamethrin has been extensively employed to manage the L. trifolii. However, subsequent studies have demonstrated that deltamethrin is no longer effective against this pest [21]. In 1979, permethrin was introduced in California, United States, for the control of the L. trifolii. Unfortunately, its effectiveness diminished due to the pest’s increased resistance [22,23]. Additionally, over the years, the L. trifolii exhibited varying degrees of resistance to permethrin, deltamethrin, and several other insecticides [24,25]. The sustainable management of crop pests in agricultural fields has been significantly compromised by the emergence of pesticide resistance [26], a challenge that is particularly pronounced in tropical regions [27].
For the identified nsSNPs, M918T (a nsSNP of M922T in L. trifolii), conferred exceptionally high resistance to deltamethrin in rat VGSC [28]. Furthermore, in 2011 and 2012, M918T was confirmed to be linked to resistance against pyrethroid insecticides in Tetranychus evansi and Aphis gossypii [29]. The M918L variant at the same position was identified in Hyelella azteca [30]. Additionally, M918V was discovered in Bemisia tabaci [31]. When examined in Xenopus oocytes, various substitutions can confer differing levels of resistance. Distinct mutant VGSCs offer varying degrees of protection against type I and type II pyrethroids [16]. This phenomenon has been observed at the M918 (T/ L/ V) site, where the M918T substitution conferred the highest level of protection against permethrin and deltamethrin [28]. In this study, only the M918T variant was identified in the L. trifolii, and the mutation frequency exhibited a gradual increase as latitude decreased, ultimately reaching a maximum of 90.91%. This finding suggests that L. trifolii in Hainan Province has developed an exceptionally high resistance to pyrethroids. This explains the widespread emergence of the potent M918T substitution across different populations and its remarkably high frequency. Furthermore, the frequency of these mutations correlates with rising temperatures at lower latitudes. This trend is likely attributed to the shortened generation cycle of L. trifolii at higher temperatures, which accelerating the accumulation of insecticide resistance in their offspring.
Among the six nsSNPs identified in this study, mutations consistently linked to pyrethroids resistance include M918, as well as L1014 (F, C, H, S, or W) (L1018F in L. trifolii) and T933 (I, C, or V). Notably, L1014F is situated in PyR1, similar to M918, whereas T933 is found in PyR2. Therefore, M918 and L1014F within PyR1 will be analyzed in our results firstly, followed by a discussion of T933 located in PyR2.
The first mutation identified and confirmed as kdr-related was L1014F (L1018F in L. trifolii) [32]. L1014F provide variable levels of protection to Type I or Type II pyrethroids or DDT [33]. Prior research has demonstrated that the L1014F single mutation confers resistance to pyrethroids in various species, including Anopheles stephensi [34], A. gossypii [29], Blattella germanica [8,35], Ctenocephalides felis [36], Cydia pomonella [37], Frankliniella occidentalis [38], Leptinotarsa decemlineata [39], Liriomyza huidobrensis [40], Meligethes aeneus [41], Musca domestica [35,42,43], Sitobion avenae [44], Triatoma infestans [45] and Liriomyza sativae [40], etc. The variants L1014S, L1014H, L1014C, and L1014W were identified at position 1014 [40,46,47]. In contrast, only one mutant type, L1014F, was observed in the L. trifolii, exhibiting a frequency of occurrence of 100% across all five populations. This finding indicates that the resistance of the L. trifolii in Hainan Province to pyrethroids has reached an exceptionally high level.
The T933I, corresponding to the amino acid number in the L. trifolii, has been linked to resistance against pyrethroids in various pests, including Thrips palmi [48], Trialeurodes vaporariorum [49], L. decemlineata [50] and Thrips tabaci [51]. Other single mutations at this position include T933C and T933V. Additionally, T929C was identified in F. occidentalis [38], while T933V has been observed in C. felis [36] and F. occidentalis [38]. In this study, only the T933I mutation was identified, and this mutation only occurred at a relatively low frequency of 8.57% in the TS. The outcome could be attributed to TS being a classic “mountain city” enclosed by mountains from all directions, leading to reproductive isolation or minimal gene flow with other populations. In small and secluded populations, particular mutations are prone to enhancing survival chances, despite lacking an immediate advantage. The T933I evidently confers a survival advantage special to the environmental challenges encountered by this population, including the intensity and frequency of local pesticide use and the cold temperatures prevalent in mountainous regions.
Prior research has demonstrated that L. trifolii populations with avermectin tolerance display the highest resistance to cypermethrin, a type II pyrethroid [52]. Consequently, cross-resistance between pyrethroids and other insecticides has surfaced in L. trifolii. Moreover, in the Sanya City field of Hainan Province, China, in 2022, the sensitivity of second-instar L. trifolii larvae to indoxacarb decreased by 776.17 times, with resistance to avermectin exhibiting a consistent upward trajectory [53]. One of the nsSNP identified in this study, V1845I (amino acid position in the L. trifolii), has been confirmed to be associated with resistance to indoxacarb in Plutella xylostella [12]. Subsequently, V1845I resistance mutation were also detected in Tuta absoluta [11]. Furthermore, this mutation has been reported in the L. trifolii [27]. Research has confirmed, using a two-electrode voltage clamp system, that the V1845I mutant channel are more resistant to indoxacarb, DCJW and metaflumizone than wild type channels [10]. Moreover, the resistance mutation V1845I of the VGSC blocker insecticides (SCBIs) was functionally validated in Drosophila through molecular simulation and genomic engineering [54]. These findings suggest that the L. trifolii populations in Hainan Province, China, not only demonstrates exceptionally high resistance to pyrethroids but also shows a relatively elevated level of cross-resistance to indoxacarb. The findings of this study indicate that as latitude decreases, the mutation frequency of V1845I exhibits a general upward trend, peaking at 96.52%. The mutation frequency in the TS region is notably higher than that observed in high-latitude areas. This discrepancy may be attributed to the widespread application of indoxacarb in the agricultural practices of the TS region.
Moreover, research has demonstrated that double mutations, in contrast to single mutations, nearly abolish the sensitivity of Drosophila melanogaster VGSC to deltamethrin [28,39]. This finding suggests that the impact of the double mutation surpasses that of the single mutation. Notably, the mutation Q1285H+V1845I exhibited identical mutation frequencies in the DA and JY populations, at 80.00% and 96.52%, respectively. Due to the sequencing methods and characteristics of WGS, it remains uncertain whether Q1285H+V1845I represents a genuine tandem double mutation. Nevertheless, based on frequency analysis, there is a likelihood of the occurrence of the double mutant Q1285H+V1845I. The functional implications of this dual process necessitate validation through electrophysiology-related experiments in future investigations. The D2036E variant identified in this study is located in the non-domain region of the VGSC of the L. trifolii. Further investigation is necessary to determine its potential association with drug resistance in this species.
In addition to the numerous nsSNPs analyzed above, our results identified a substantial number of sSNPs in the VGSC of the L. trifolii, with their prevalence significantly exceeding that of nsSNPs. Historically referred to as “silent mutations”, sSNPs are now understood to be not entirely silent. In general, sSNPs are still considered benign or minimally harmful. Furthermore, accurately quantifying the number of harmful variations (specifically nsSNPs), and neutral variations (specifically sSNPs), present in a population can elucidate the influence of the population’s developmental history on genetic variation. This assessment can also determine whether a small-scale population is prone to the accumulation of harmful variations, potentially resulting in uncontrolled mutations or even species extinction and outbreaks [55,56]. In evolutionary research, the ratio of nsSNP to sSNP (dN/dS) serves as a crucial indicator for assessing whether a gene is subject to natural selection pressure. When natural selection facilitates alterations in protein sequences, the dN/dS ratio is anticipated to exceed 1. Conversely, when natural selection restricts changes in proteins, the ratio falls below 1 [58,59]. This interpretation of dN/dS is further substantiated by theoretical analyses of the relationship between dN/dS statistics and potential selection pressure within the Wright-Fisher model [60]. In this study, all five populations adhered to the principle of natural selection, as indicated by a dN/dS ratio of less than 1. Our results propose that this observation signifies several important aspects. Firstly, in the VGSC of the L. trifolii, the majority of deleterious mutations (nsSNPs) are purged from the population. Individuals harboring these mutations, which disrupt the original structure and function of proteins, experience significantly reduced survival and reproductive success. Consequently, natural selection persistently eliminates these harmful nsSNPs, resulting in a dN/dS ratio of less than 1. Next, the functionality of VGSC is critically important. Their protein sequences are essential for survival and reproduction, such that any non-synonymous alteration in their amino acid sequences carries substantial implications. Furthermore, the emergence of any nsSNP likely represents a significant site that aids the population in resisting natural selection pressures, such as pesticide resistance. Finally, a dN/dS ratio of less than 1 also suggests that the VGSC sequence has remained highly conserved throughout a prolonged evolutionary period, with minimal changes in its amino acid sequence across different species or generations. Key regions, including structural or functional sites of VGSCs, are subject to considerable selection pressure to preserve their existing functions, thereby limiting the potential for alteration. In Hainan Province, characterized by persistent high temperatures, humidity, and continuous pesticide use, further investigation is warranted regarding the unique evolution of VGSC in L. trifolii and associated nsSNPs.
4. Materials and Methods
4.1. Insects Collection
The sensitive strain of L. trifolii (LT-S) used in this study was provided by Professor Yuzhou Du of Yangzhou University. This strain has been maintained in the laboratory without exposure to insecticides for over two decades. Wild populations selected for WGS were gathered from five locations: Tian-Ya (TY: N 18.31, E 109.48) and Ji-Yang (JY: N 18.29, E 109.54) in Sanya, Mao-Yang (MY: N 18.94, E 109.51) and Tong-Shi (TS: N 18.79, E 109.52) in Wuzhishan, and Da-An in Ledong (DA: N 18.47, E 108.90) (Figure 4). The entire life cycle of the insects was raised at a temperature of 26 ± 1 ℃, the humidity should be maintained at 70% ± 5% and L: D = 16h: 8 h. The adult insects laid eggs and the larvae were raised using cultivated bean seedlings (Baijiali cowpea seeds, USA) that were not exposed to any pesticides.
4.2. Acquisition of Complete Genomic of VGSC in L. trifolii and Sequencing WGS
Genomic DNA extracted from LT-S was single-headed and processed using the Universal Genomic DNA Extraction Kit (Beijing Solarbio Science & Technology Co., Ltd., China). Segmented gene amplification on the complete genomic sequence of VGSC of L. trifolii was conducted utilizing the Phanta-Max Super-Fidelity DNA Polymerase kit (Vazyme Biotech Co., Ltd., China). By integrating the genomic data of the VGSC from the L. trifolii (GenBank: GCA_001014935.1), primers were formulated for cloning to acquire the complete genome sequence of the susceptible strain of the VGSC from the L. trifolii. Details of the primers and annealing temperatures employed for fragment amplification can be found in Table 3. Subsequently, PCR products were submitted to Tsingke Biotechnology Co., Ltd. (China) for first-generation sequencing, and the DNASTAR Lasergene 11 Core Suite software was utilized to assemble the complete genomic sequence of VGSC.
Large quantities of sensitive strains cultivated indoors and resistant populations gathered from the field were sent to the Beijing Genomics Institute (BGI Genomics Co., Ltd.) for WGS. Genomic data analysis was conducted utilizing the GeneAn cloud platform.
4.3. Analysis and Acquisition of sSNP and nsSNP in VGSC of the L. trifolii
The genomic fragment sequences of the VGSC from the LT-S strain were concatenated to derive the complete full-length genomic sequence of the VGSC of L. trifolii. This complete VGSC genomic sequence will be annotated according to the Coding Sequence (CDS) (GenBank: MT648287.1), and the nucleotide position information of the intron and exon segments will be scrutinized. Mutants situated in the exon region were filtered from the outcomes of WGS using the aforementioned nucleotide position information. Subsequently, by correlating the codon information of each mutation in the exon with the CDS position, our results classified the mutations as synonymous single nucleotide polymorphism (sSNP) or non-synonymous single nucleotide polymorphism (nsSNP), determined their positions on the VGSC, and computed their frequencies of occurrence. The approach for distinguishing between sSNPs and nsSNPs is illustrated in Figure S3, where the section highlighted in red elucidates the method for identifying sSNPs as discussed in this study.
Author Contributions
Conducted experiments and wrote the manuscript, Linlin Yuan and Zhiyuan Lei; data curation, Junyi Zhang; writing—review and editing, Fen Li and Shaoying Wu. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D Program of China, grant number 2024YFD1400100, Position Scientist Project of the Department of Agriculture and Rural Affairs of Hainan Province, grant number HNARS-05-G04, China National Natural Science Foundation (32260666 and 32460659), 2025 College Students’ In-novation Training Program of Hainan University S202510589007.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
For confidentiality reasons, the original data supporting the results of this study have not been stored publicly. However, desensitized data may be obtained from the corresponding author upon reasonable request, provided that ethical and confidentiality agreements are upheld.
Acknowledgments
We gratefully acknowledge Professor Yuzhou Du (Yangzhou University) for supplying the sensitive L. trifolii strain (LT-S) used in this research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Scheffer, S.J.; Lewis, M.L. Mitochondrial Phylogeography of the Vegetable Pest Liriomyza Trifolii (Diptera: Agromyzidae): Diverged Clades and Invasive Populations. Ann Entomol. Soc. Am 2006, 99, 991–998. [Google Scholar] [CrossRef]
- Spencer, K.A. Agromyzidae (Diptera) of Economic Importance; Springer Netherlands: Dordrecht, 1973; ISBN 978-90-481-8513-9. [Google Scholar]
- Gao, Y.L.; Reitz, S.; Xing, Zhen Long; Ferguson, S.; Lei, Z.R. A Decade of Leafminer Invasion in China: Lessons Learned. Pest Manag. Sci. 2017, 73, 1775–1779. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Tokumaru, S. Displacement in Two Invasive Species of Leafminer Fly in Different Localities. Biol. Invasions 2008, 10, 951–953. [Google Scholar] [CrossRef]
- Meurer, B. Pyrethrum Flowers. Production, Chemistry, Toxicology, and Uses. Brittonia 1996, 48, 613–614. [Google Scholar] [CrossRef]
- Davies, T.G.E.; Field, L.M.; Usherwood, P.N.R.; Williamson, M.S. DDT, Pyrethrins, Pyrethroids and Insect Sodium Channels. IUBMB Life 2007, 59, 151–162. [Google Scholar] [CrossRef]
- Dong, K.; Du, Y.Z.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.; Silver, K.; Zhorov, B.S. Molecular Biology of Insect Sodium Channels and Pyrethroid Resistance. Insect Biochem. Mol. Biol. 2014, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dong, K. Insect Sodium Channels and Insecticide Resistance. Invert Neurosci. 2007, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, D.M.; Bloomquist, J.R. Molecular Mechanisms of Insecticide Resistance. In Pesticide Resistance in Arthropods; Roush, R.T., Tabashnik, B.E., Eds.; Springer US: Boston, MA, 1990; pp. 58–96. ISBN 978-1-4684-6431-3. [Google Scholar]
- Jiang, D.X.; Du, Y.Z.; Nomura, Y.; Wang, X.L.; Wu, Y.D.; Zhorov, B.S.; Dong, K. Mutations in the Transmembrane Helix S6 of Domain IV Confer Cockroach Sodium Channel Resistance to Sodium Channel Blocker Insecticides and Local Anesthetics. Insect Biochem. Mol. Biol. 2015, 66, 88–95. [Google Scholar] [CrossRef]
- Roditakis, E.; Mavridis, K.; Riga, M.; Vasakis, E.; Morou, E.; Rison, J.L.; Vontas, J. Identification and Detection of Indoxacarb Resistance Mutations in the Para Sodium Channel of the Tomato Leafminer, Tuta Absoluta. Pest Manag. Sci. 2017, 73, 1679–1688. [Google Scholar] [CrossRef]
- Wang, X.L.; Su, W.; Zhang, J.H.; Yang, Y.H.; Dong, K.; Wu, Y.D. Two Novel Sodium Channel Mutations Associated with Resistance to Indoxacarb and Metaflumizone in the Diamondback Moth, Plutella Xylostella. Insect Sci. 2016, 23, 50–58. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Du, Y.Z.; Jiang, D.X.; Behnke, C.; Nomura, Y.; Zhorov, B.S.; Dong, K. The Receptor Site and Mechanism of Action of Sodium Channel Blocker Insecticides. J Biological Chem. 2016, 291, 20113–20124. [Google Scholar] [CrossRef]
- Catterall, W.A. From Ionic Currents to Molecular Mechanisms: The Structure and Function of Voltage-Gated Sodium Channels. Neuron 2000, 26, 13–25. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, A.O.; Khambay, B.P.S.; Williamson, M.S.; Field, L.M.; WAllace, B.A.; Davies, T.G.E. Modelling Insecticide-Binding Sites in the Voltage-Gated Sodium Channel. Biochem. J 2006, 396, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.Z.; Nomura, Y.; Satar, G.; Hu, Z.N.; Nauen, R.; He, S.Y.; Zhorov, B.S.; Dong, K. Molecular Evidence for Dual Pyrethroid-Receptor Sites on a Mosquito Sodium Channel. P Natl. A. Sci. 2013, 110, 11785–11790. [Google Scholar] [CrossRef]
- Şulea, T.A.; Draga, S.; Mernea, M.; Corlan, A.D.; Radu, B.M.; Petrescu, A.-J.; Amuzescu, B. Differential Inhibition by Cenobamate of Canonical Human Nav1.5 Ion Channels and Several Point Mutants. IJMS 2025, 26, 358. [Google Scholar] [CrossRef] [PubMed]
- Cosme, L.V.; Lima, J.B.P.; Powell, J.R.; Martins, A.J. Genome-Wide Association Study Reveals New Loci Associated with Pyrethroid Resistance in Aedes Aegypti. Front. Genet. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Fu, P.; Chen, Y.; Wan, F.; Hu, G.; Gui, F. Employing Genome-Wide Association Studies and Machine Learning to Accurately Identify Eastern and Western Migratory Pathways of Spodoptera Frugiperda in China via Key Molecular Markers. Ecol. Inf. 2025, 92, 103490. [Google Scholar] [CrossRef]
- Gao, Y.L.; Lei, Z.R.; Abe, Y.; Reitz, S.R. Species Displacements Are Common to Two Invasive Species of Leafminer Fly in China, Japan, and the United States. J Econ Entomol 2011, 104, 1771–1773. [Google Scholar] [CrossRef]
- Abeeluck, D.; Dunhawoor, C.; Unmole, L. Development and Implementation of Integrated Pest Management in Mauritius an Overview. University of Mauritius Research Journal 2009, 15. [Google Scholar]
- Keil, C.B.; Parrella, M.P.; Morse, J.G. Method for Monitoring and Establishing Baseline Data for Resistance to Permethrin by Liriomyza Trifolii (Burgess). J Econ Entomol 1985, 78, 419–422. [Google Scholar] [CrossRef]
- Mason, G.A.; Johnson, M.W.; Tabashnik, B.E. Susceptibility of Liriomyza Sativae and L. Trifolii (Diptera: Agromyzidae) to Permethrin and Fenvalerate. J Econ Entomol 1987, 80, 1262–1266. [Google Scholar] [CrossRef]
- Macdonald, O.C. Responses of the Alien Leaf Miners Liriomyza Trifolii and Liriomyza Huidobrensis (Diptera: Agromyzidae) to Some Pesticides Scheduled for Their Control in the UK. Crop Prot. 1991, 10, 509–513. [Google Scholar] [CrossRef]
- Parrella, M.P.; Trumble, J.T. Decline of Resistance in Liriomyza Trifolii (Diptera: Agromyzidae) in the Absence of Insecticide Selection Pressure. J Econ Entomol 1989, 82, 365–368. [Google Scholar] [CrossRef]
- Sparks, T.C.; Nauen, R. IRAC: Mode of Action Classification and Insecticide Resistance Management. Pestic. Biochem. Physiol. 2015, 121, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Gong, X.Y.; Yuan, L.L.; Pan, X.L.; Jin, H.F.; Lu, R.C.; Wu, S.Y. Indoxacarb Resistance-Associated Mutation of Liriomyza Trifolii in Hainan, China. Pestic. Biochem. Physiol. 2022, 183, 105054. [Google Scholar] [CrossRef] [PubMed]
- Vais, H.; Atkinson, S.; Eldursi, N.; Devonshire, A.L.; Williamson, M.S.; Usherwood, P.N.R. A Single Amino Acid Change Makes a Rat Neuronal Sodium Channel Highly Sensitive to Pyrethroid Insecticides. FEBS Letters 2000, 470, 135–138. [Google Scholar] [CrossRef]
- Clark, J.M.; Symington, S.B. Advances in the Mode of Action of Pyrethroids. Pyrethroids: From Chrysanthemum to Modern Industrial Insecticide 2012, 49–72. [Google Scholar] [CrossRef]
- Weston, D.P.; Poynton, H.C.; Wellborn, G.A.; Lydy, M.J.; Blalock, B.J.; Sepulveda, M.S.; Colbourne, J.K. Multiple Origins of Pyrethroid Insecticide Resistance across the Species Complex of a Nontarget Aquatic Crustacean, Hyalella Azteca. P Natl. A. Sci. 2013, 110, 16532–16537. [Google Scholar] [CrossRef] [PubMed]
- Morin, S.; Williamson, M.S.; Goodson, S.J.; Brown, J.K.; Tabashnik, B.E.; Dennehy, T.J. Mutations in the Bemisia Tabaci Para Sodium Channel Gene Associated with Resistance to a Pyrethroid plus Organophosphate Mixture. Insect Biochem. Mol. Biol. 2002, 32, 1781–1791. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Du, Y.Z.; Dong, K. Diversity and Convergence of Sodium Channel Mutations Involved in Resistance to Pyrethroids. Pestic. Biochem. Physiol. 2013, 106, 93–100. [Google Scholar] [CrossRef]
- Burton, M.J.; Mellor, I.R.; Duce, I.R.; Davies, T.G.E.; Field, L.M.; Williamson, M.S. Differential Resistance of Insect Sodium Channels with Kdr Mutations to Deltamethrin, Permethrin and DDT. Insect Biochem. Mol. Biol. 2011, 41, 723–732. [Google Scholar] [CrossRef]
- Enayati, A.A.; Vatandoost, H.; Ladonni, H.; Townson, H.; Hemingway, J. Molecular Evidence for a Kdr-like Pyrethroid Resistance Mechanism in the Malaria Vector Mosquito Anopheles Stephensi. Med Vet Entomol 2003, 17, 138–144. [Google Scholar] [CrossRef]
- Miyazaki, M.; Ohyama, K.; Dunlap, D.Y.; Matsumura, F. Cloning and Sequencing of Thepara-Type Sodium Channel Gene from Susceptible Andkdr-Resistant German Cockroaches (Blattella Germanica) and House Fly (Musca Domestica). Molec. Gen. Genet. 1996, 252, 61–68. [Google Scholar] [CrossRef]
- Bass, C.; Schroeder, I.; Turberg, A.; M Field, L.; S Williamson, M. Identification of Mutations Associated with Pyrethroid Resistance in the Para-Type Sodium Channel of the Cat Flea, Ctenocephalides Felis. Insect Biochem. Mol. Biol. 2004, 34, 1305–1313. [Google Scholar] [CrossRef]
- Brun-Barale, A.; Bouvier, J.-C.; Pauron, D.; Bergé, J.-B.; Sauphanor, B. Involvement of a Sodium Channel Mutation in Pyrethroid Resistance in Cydia Pomonella L, and Development of a Diagnostic Test. Pest Manag. Sci. 2005, 61, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Forcioli, D.; Frey, B.; Frey, J.E. High Nucleotide Diversity in the Para-like Voltage-Sensitive Sodium Channel Gene Sequence in the Western Flower Thrips (Thysanoptera: Thripidae). J Econ Entomol 2002, 95, 838–848. [Google Scholar] [CrossRef]
- Hyeock Lee, S.; Smith, T.; C. Knipple, D.; Soderlund, D. Mutations in the House Fly Vssc1 Sodium Channel Gene Associated with Super-Kdr Resistance Abolish the Pyrethroid Sensitivity of Vssc1/tipE Sodium Channels Expressed in Xenopus Oocytes. Insect Biochem. Mol. Biol. 1999, 29, 185–194. [Google Scholar] [CrossRef]
- Davies, T.G.E.; Field, L.M.; Usherwood, P.N.R.; Williamson, M.S. A Comparative Study of Voltage-Gated Sodium Channels in the Insecta: Implications for Pyrethroid Resistance in Anopheline and Other Neopteran Species. Insect Mol. Biol. 2007, 16, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Zimmer, C.T.; Andrews, M.; Slater, R.; Bass, C.; Ekbom, B.; Gustafsson, G.; Hansen, L.M.; Kristensen, M.; Zebitz, C.P.W. Target-Site Resistance to Pyrethroids in European Populations of Pollen Beetle, Meligethes Aeneus F. Pestic. Biochem. Physiol. 2012, 103, 173–180. [Google Scholar] [CrossRef]
- Ingles, P.J.; Adams, P.M.; Knipple, D.C.; Soderlund, D.M. Characterization of Voltage-Sensitive Sodium Channel Gene Coding Sequences from Insecticide-Susceptible and Knockdown-Resistant House Fly Strains. Insect Biochem. Mol. Biol. 1996, 26, 319–326. [Google Scholar] [CrossRef]
- Williamson, M.S.; Martinez-Torres, D.; Hick, C.A.; Devonshire, A.L. Identification of Mutations in the Housefly Para-Type Sodium Channel Gene Associated with Knockdown Resistance (Kdr) to Pyrethroid Insecticides. Molec. Gen. Genet. 1996, 252, 51–60. [Google Scholar] [CrossRef]
- Foster, S.P.; Paul, V.L.; Slater, R.; Warren, A.; Denholm, I.; Field, L.M.; Williamson, M.S. A Mutation (L1014F) in the Voltage-Gated Sodium Channel of the Grain Aphid, Sitobion Avenae, Is Associated with Resistance to Pyrethroid Insecticides. Pest Manag. Sci. 2014, 70, 1249–1253. [Google Scholar] [CrossRef]
- Fabro, J.; Sterkel, M.; Capriotti, N.; Mougabure-Cueto, G.; Germano, M.; Rivera-Pomar, R.; Ons, S. Identification of a Point Mutation Associated with Pyrethroid Resistance in the Para-Type Sodium Channel of Triatoma Infestans, a Vector of Chagas’ Disease. Infect Genet Evol. 2012, 12, 487–491. [Google Scholar] [CrossRef]
- Verhaeghen, K.; Van Bortel, W.; Trung, H.D.; Sochantha, T.; Keokenchanh, K.; Coosemans, M. Knockdown Resistance in “Anopheles Vagus”, “An. Sinensis”, “An. Paraliae” and “An. Peditaeniatus” Populations of the Mekong Region. Parasites Vectors 2010, 3, 59. [Google Scholar] [CrossRef] [PubMed]
- Lol, J.C.; Castellanos, M.E.; Liebman, K.A.; Lenhart, A.; Pennington, P.M.; Padilla, N.R. Molecular Evidence for Historical Presence of Knock-down Resistance in Anopheles Albimanus, a Key Malaria Vector in Latin America. Parasites Vectors 2013, 6, 268. [Google Scholar] [CrossRef]
- Bao, W.X.; Sonoda, S. Resistance to Cypermethrin in Melon Thrips, Thrips Palmi (Thysanoptera: Thripidae), Is Conferred by Reduced Sensitivity of the Sodium Channel and CYP450-Mediated Detoxification. Appl. Entomol. Zool 2012, 47, 443–448. [Google Scholar] [CrossRef]
- Karatolos, N.; Gorman, K.; Williamson, M.S.; Denholm, I. Mutations in the Sodium Channel Associated with Pyrethroid Resistance in the Greenhouse Whitefly, Trialeurodes Vaporariorum. Pest Manag. Sci. 2012, 68, 834–838. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Su, C.; Lazo, T.A.; Hawthorne, D.J.; Tingey, W.M.; Naimov, S.; Scott, J.G. Multiple Evolutionary Origins of Knockdown Resistance (Kdr) in Pyrethroid-Resistant Colorado Potato Beetle, Leptinotarsa Decemlineata. Pestic. Biochem. Physiol. 2012, 104, 192–200. [Google Scholar] [CrossRef]
- Toda, S.; Morishita, M. Identification of Three Point Mutations on the Sodium Channel Gene in Pyrethroid-Resistant Thrips Tabaci (Thysanoptera: Thripidae). J Econ Entomol 2009, 102, 2296–2300. [Google Scholar] [CrossRef]
- Wang, Y.C.; Chang, Y.W.; Gong, W.R.; Hu, J.; Du, Y.Z. The Development of Abamectin Resistance in Liriomyza Trifolii and Its Contribution to Thermotolerance. Pest Manag. Sci. 2024, 80, 2053–2060. [Google Scholar] [CrossRef]
- Gong, X.Y.; Chen, Y.; Dong, W.B.; Li, F.; Wu, S.Y. Toxicity of Indoxacarb to the Population of Liriomyza Trifolii (Diptera: Agromyzidae) in Sanya (China), and the Effects of Temperature and Food on Its Biological Characteristics. Tropical Plants 2024, 3. [Google Scholar] [CrossRef]
- Samantsidis, G.; O’Reilly, A.O.; Douris, V.; Vontas, J. Functional Validation of Target-Site Resistance Mutations against Sodium Channel Blocker Insecticides (SCBIs) via Molecular Modeling and Genome Engineering in Drosophila. Insect Mol. Biol. 2019, 104, 73–81. [Google Scholar] [CrossRef]
- Schubert, M.; Jónsson, H.; Chang, D.; Der Sarkissian, C.; Ermini, L.; Ginolhac, A.; Albrechtsen, A.; Dupanloup, I.; Foucal, A.; Petersen, B.; et al. Prehistoric Genomes Reveal the Genetic Foundation and Cost of Horse Domestication. P Natl. A. Sci. 2014, 111, 5661–5669. [Google Scholar] [CrossRef] [PubMed]
- Marsden, C.D.; Ortega-Del Vecchyo, D.; O’Brien, D.P.; Taylor, J.F.; Ramirez, O.; Vilà, C.; Marques-Bonet, T.; Schnabel, R.D.; Wayne, R.K.; Lohmueller, K.E. Bottlenecks and Selective Sweeps during Domestication Have Increased Deleterious Genetic Variation in Dogs. P Natl. A. Sci. 2016, 113, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Henn, B.M.; Botigué, L.R.; Peischl, S.; Dupanloup, I.; Lipatov, M.; Maples, B.K.; Martin, A.R.; Musharoff, S.; Cann, H.; Snyder, M.P.; et al. Distance from Sub-Saharan Africa Predicts Mutational Load in Diverse Human Genomes. P Natl. A. Sci. 2016, 113, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. Preponderance of Synonymous Changes as Evidence for the Neutral Theory of Molecular Evolution. Nature 1977, 267, 275–276. [Google Scholar] [CrossRef]
- Yang, Z.H.; Bielawski, J.P. Statistical Methods for Detecting Molecular Adaptation. Trends Ecol. Evol. 2000, 15, 496–503. [Google Scholar] [CrossRef]
- Nielsen, R.; Yang, Z.H. Estimating the Distribution of Selection Coefficients from Phylogenetic Data with Applications to Mitochondrial and Viral DNA. Mol. Biol. Evol. 2003, 20, 1231–1239. [Google Scholar] [CrossRef]
Figure 1.
Amino acid information map of the complete CDS region, including four domains and each helical fragment of the VGSC from the sensitive strain of L. trifolii.
Figure 1.
Amino acid information map of the complete CDS region, including four domains and each helical fragment of the VGSC from the sensitive strain of L. trifolii.

Figure 2.
Heat maps of the occurrence and frequency distribution of 31 sSNPs in five populations.
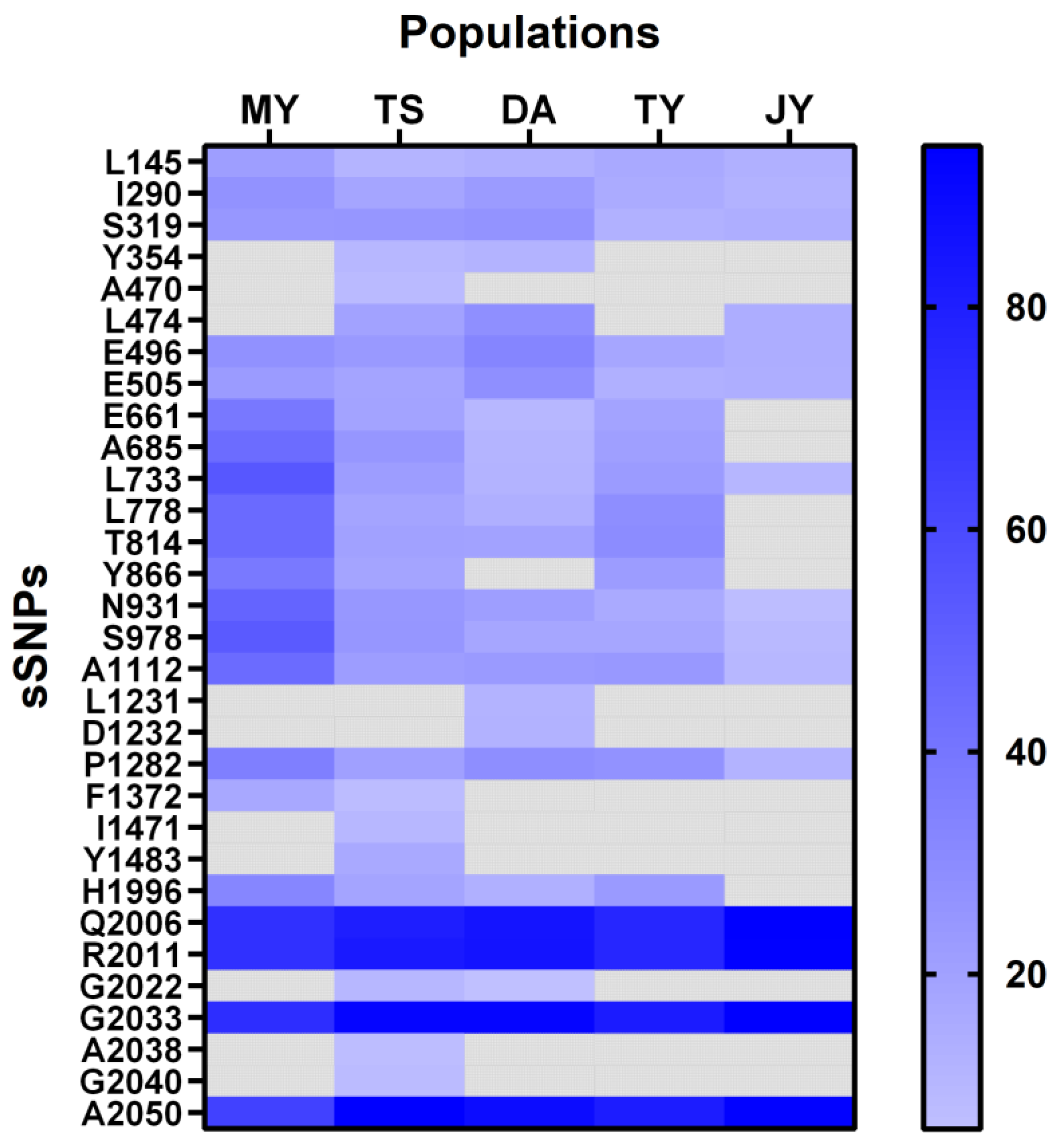
Figure 3.
The positions of six nsSNPs on the VGSC topological map. The (A) occurrence and (B) frequency distribution heat maps in the five populations.
Figure 3.
The positions of six nsSNPs on the VGSC topological map. The (A) occurrence and (B) frequency distribution heat maps in the five populations.
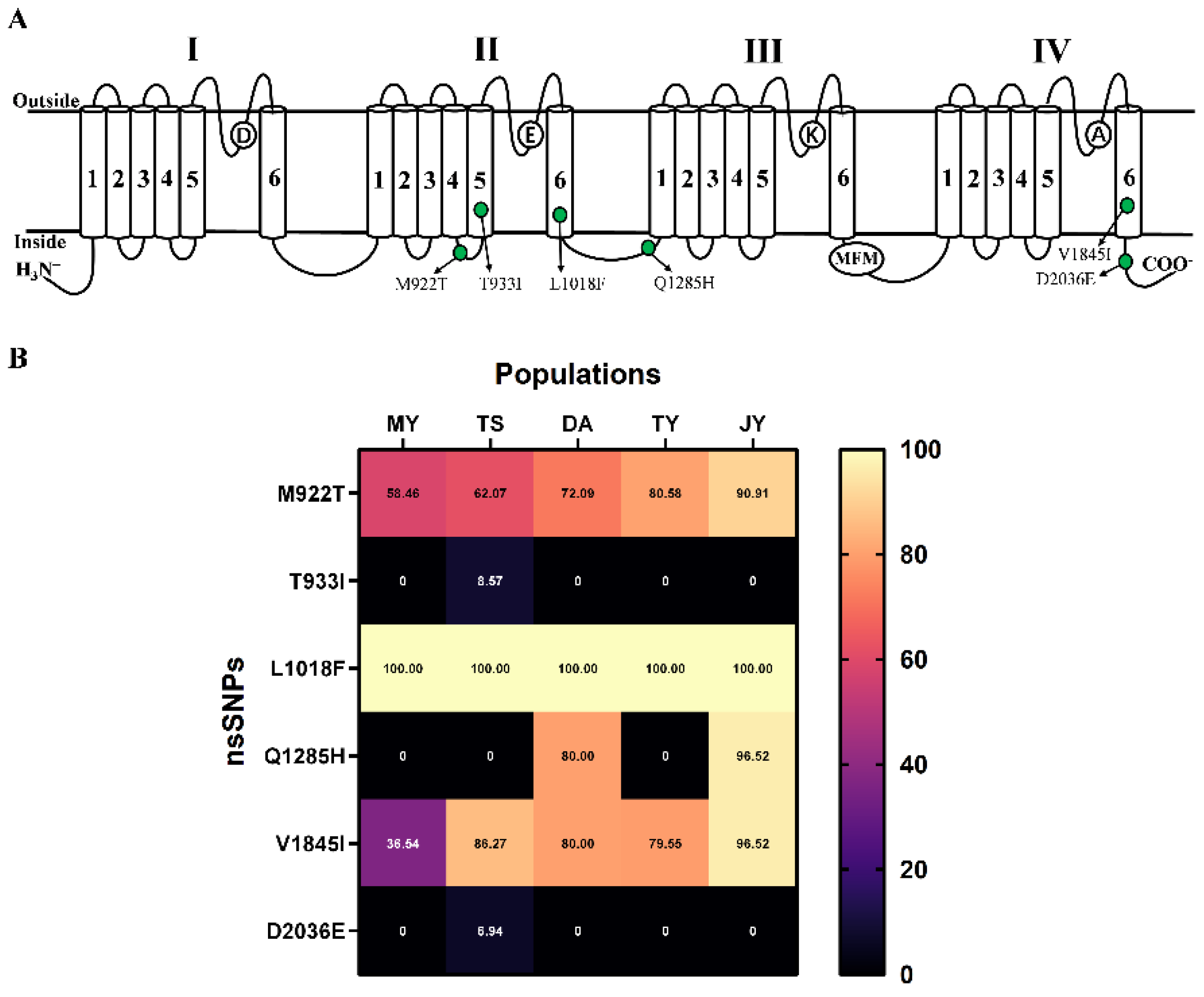
Figure 4.
Origin information of population samples for whole-genome resequencing (WGS).
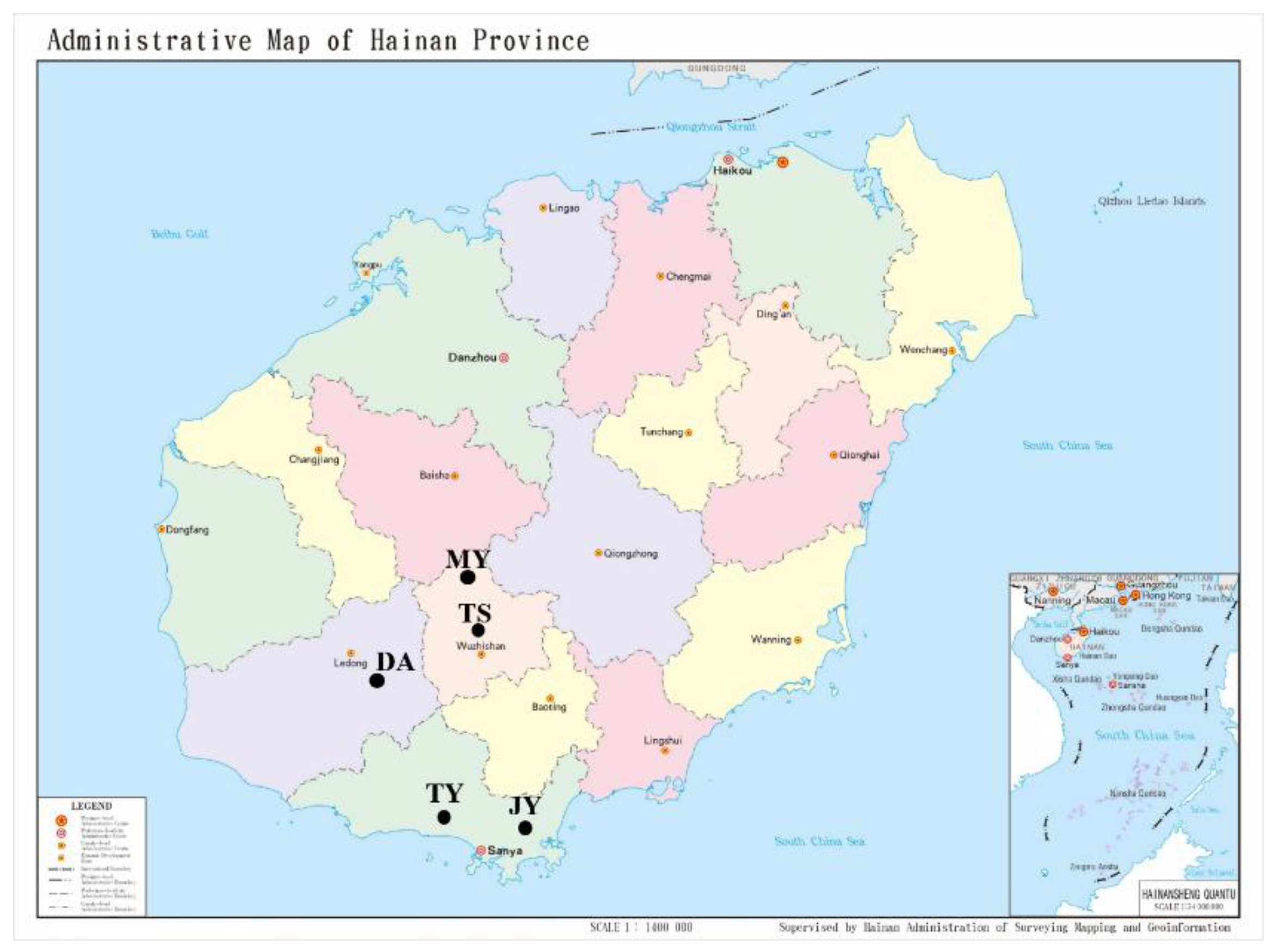

Table 1.
Positions, variations and frequencies of 31 sSNPs in the VGSC of L. trifolii.
| Position d | sSNPs | Nucleotide alteration | Frequency/ %e (f, g) | ||||
| MY | TS | DA | TY | JY | |||
| D Ⅰ-S1 | L145 | TTA>TTG | 21.33 (59, 16) | 11.36 (78, 10) | 13.11 (53, 8) | 16.22 (62, 12) | 13.11 (53, 8) |
| D Ⅰ-L-S56 | I290 | ATT>ATC | 27.05 (89, 33) | 18.03 (100, 22) | 23.08 (60, 18) | 15.65 (97, 18) | 12.07 (102,14) |
| S319 | TCT>TCA | 24.77 (82, 27) | 25.00 (72, 24) | 26.47 (50, 18) | 12.90 (81, 12) | 14.00 (86, 14) | |
| Y354 | TAT>TAC | - | 9.89 (82, 9) | 11.96 (81, 11) | - | - | |
| L-D Ⅰ-Ⅱ | A470 | GCG>GCT | - | 8.57 (64, 6) | - | - | - |
| L474 | TTA>TTG | - | 19.70 (53, 13) | 28.24 (61, 24) | - | 14.55 (94, 16) | |
| E496 | GAG>GAA | 27.27 (48, 18) | 23.91 (35, 11) | 32.94 (57, 28) | 17.72 (65, 14) | 14.46 (71, 12) | |
| E505 | GAA>GAG | 22.95 (47, 14) | 18.75 (39, 9) | 28.57 (55, 22) | 13.33 (52, 8) | 14.08 (61, 10) | |
| E661 | GAG>GAA | 39.18 (59, 38) | 19.18 (59, 14) | 10.00 (81, 9) | 18.95 (77, 18) | - | |
| A685 | GCC>GCA | 44.33 (54, 43) | 25.00 (51, 17) | 11.24 (79, 10) | 20.93 (68, 18) | - | |
| L733 | TTG>TTA | 54.22 (38, 45) | 21.95 (64, 18) | 11.59 (61, 8) | 22.69 (92, 27) | 10.24 (114, 13) | |
| L778 | CTC>CTT | 45.53 (67, 56) | 18.63 (83, 19) | 13.49 (109, 17) | 28.91 (91, 37) | - | |
| D Ⅱ-S1 | T814 | ACA>ACG | 45.28 (58, 48) | 20.00 (80, 20) | 19.82 (89, 22) | 29.91 (75, 32) | - |
| D Ⅱ-L-S23 | Y866 | TAC>TAT | 38.46 (56, 35) | 18.68 (74, 17) | - | 22.23 (70, 20) | - |
| D Ⅱ-S5 | N931 | AAC>AAT | 48.00 (39, 36) | 24.64 (52, 17) | 21.43 (66, 18) | 16.16 (83, 16) | 7.08 (105, 8) |
| D Ⅱ-L-S56 | S978 | TCT>TCG | 52.87 (41, 46) | 25.00 (54, 18) | 17.71 (79, 17) | 18.00 (82, 18) | 9.16 (119, 12) |
| L-D Ⅱ-Ⅲ | A1112 | GCT>GCA | 44.87 (43, 35) | 21.88 (50, 14) | 23.38 (59, 18) | 24.44 (68, 22) | 9.76 (74, 8) |
| L1231 | CTA>CTT | - | - | 11.76 (90, 12) | - | - | |
| D1232 | GAT>GAC | - | - | 12.24 (86, 12) | - | - | |
| P1282 | CCT>CCG | 35.71 (72, 40) | 20.72 (88, 23) | 28.70 (77, 31) | 27.17 (67, 25) | 11.72 (113, 15) | |
| D Ⅲ-S3 | F1372 | TTT>TTC | 16.67 (30, 6) | 7.50 (37, 3) | - | - | - |
| D Ⅲ-L-S56 | I1471 | ATC>ATT | - | 10.10 (89, 10) | - | - | - |
| Y1483 | TAT>TAC | - | 16.19 (88, 17) | - | - | - | |
| L-D Ⅳ < | H1996 | CAC>CAT | 32.43 (75, 36) | 18.52 (66, 15) | 13.21 (92, 14) | 23.15 (83, 25) | - |
| Q2006 | CAA>CAG | 72.32 (31, 81) | 80.28 (14, 57) | 85.15 (15, 86) | 76.77 (23, 76) | 94.31 (7, 116) | |
| R2011 | CGA>CGT | 72.22 (30, 78) | 82.54 (11, 52) | 85.29 (15, 87) | 76.84 (22, 73) | 93.97 (7, 109) | |
| G2022 | GGT>GGC | - | 10.00 (54, 6) | 6.06 (93, 6) | - | - | |
| G2033 | GGG>GGT | 73.63 (24, 67) | 91.94 (5, 57) | 91.86 (7, 79) | 81.94 (13, 59) | 94.44 (6, 102) | |
| A2038 | GCG>GCT | - | 7.04 (66, 5) | - | - | - | |
| G2040 | GGA>GGT | - | 8.33 (66, 6) | - | - | - | |
| A2050 | GCC>GCT | 64.41 (21, 38) | 94.29 (4, 66) | 88.57 (8, 62) | 81.36 (11, 48) | 93.40 (7, 99) | |
Note: The meanings represented by each symbol in position “d” are as follows: “<“ indicates being located “behind” a certain position; “D” represents “Domain”; “L” represents “Linker”; “S” represents “Helix Segment”; “Ⅰ”, “Ⅱ”, “Ⅲ” and “Ⅳ” represent the naming of four domains respectively; “1”, “2”, “3”, “4”, “5” and “6” represent six “Hydrophobic transmembrane helix”. “f” represents the number of times that no nucleotide change occurred at a specific sSNP position during the Whole Genome Re-sequencing (WGS), while “g” represents the number of times that a nucleotide change occurred at that sSNP position. The calculation method for “e” is: e = g / (f + g).
Table 2.
Positions, variations and frequencies of 6 nsSNPs in the VGSC of L. trifolii.
| Position d | nsSNPs | Nucleotide alteration | Frequency/ % h (i, j) | ||||
| MY | TS | DA | TY | JY | |||
| D Ⅱ-L-S45 | M922T | ATG>ACG | 58.46 (27, 38) | 62.07 (22, 36) | 72.09 (24, 62) | 80.58 (20, 83) | 90.91 (11, 110) |
| D Ⅱ-S5 | T933I | ACA>ATA | - | 8.57 (64, 6) | - | - | - |
| D Ⅱ-S6 | L1018F | CTT>TTT | 100.00 (0, 64) | 100.00 (0, 71) | 100.00 (0, 90) | 100.00 (0, 102) | 100.00 (0, 126) |
| L-D Ⅱ-Ⅲ | Q1285H | CAA>CAC | - | - | 80.00 (18,72) | - | 96.52 (4,111) |
| D Ⅳ-S6 | V1845I | GTT>ATA | 36.54 (99,57) | 86.27 (14,88) | 80.00 (18,72) | 79.55 (18,70) | 96.52 (4,111) |
| L-D Ⅳ < | D2036E | GAC>GAA | - | 6.94 (67,5) | - | - | - |
Note: The meanings represented by each symbol in position “d” are as follows: “<“ indicates being located “behind” a certain position; “D” represents “Domain”; “L” represents “Linker”; “S” represents “Helix Segment”; “Ⅰ”, “Ⅱ”, “Ⅲ” and “Ⅳ” represent the naming of four domains respectively; “1”, “2”, “3”, “4”, “5” and “6” represent six “Hydrophobic transmembrane helix”. “i” represents the number of times that no nucleotide change occurred at a specific nsSNP position during the Whole Genome Re-sequencing (WGS), while “j” represents the number of times that a nucleotide change occurred at that nsSNP position. The calculation method for “h” is: h = j / (i + j).
Table 3.
Primers Information and annealing temperatures for VGSC in L. trifolii.
| Primers | Sequence | Product length a (b, c) (bp) |
Annealing Temperature ( ℃) |
| LT-1F | ATGACAGAAGATTCCGACTCGA | 288 (215, 73) | 47.5 |
| LT-1R | TTCCGGCGGGAAGCTGCCCTGCAAT | ||
| LT-2F | GGTCCACAACCGGATCCTAC | 1671 (188, 1483) | 39.0 |
| LT-2R | TGGCTACACGACGTATTGGA | ||
| LT-3F | TCCAATACGTCGTGTAGCCA | 2811 (236, 2575) | 41.5 |
| LT-3R | AAGTCCAGCCAATTCCATGCA | ||
| LT-4F | GTGATGGCACGAGGTTTCAT | 1982 (713, 1269) | 40.0 |
| LT-4R | TCGTCATACGACATGGCAAC | ||
| LT-5F | CGAATTGCAAAAGAAAGCCGA | 3701 (565, 3136) | 41.5 |
| LT-5R | TTCAGACATCGGTGTGACGG | ||
| LT-6F | CCGTCACACCGATGTCTGAA | 4563 (610, 3953) | 44.5 |
| LT-6R | TCGAAGACAATTAATGACACCCA | ||
| LT-7F | TGGGTGTCATTAATTGTCTTCGA | 1898 (697, 1201) | 45.0 |
| LT-7R | TGGACAAAAGCAAGGCCAAG | ||
| LT-8F | TGGCCTTGCTTTTGTCCAATT | 4357 (796, 3561) | 42.0 |
| LT-8R | AAATTGCCCCATCCTTGCCA | ||
| LT-9F | GGCAATTTACGACTGAAAACTTTTCA | 1008 (225, 783) | 49.0 |
| LT-9R | ATACACCGCAAACCCGAGAG | ||
| LT-10F | CTGCCGCAAAGACCCATACT | 4363 (301, 4062) | 39.0 |
| LT-10R | GAACCAGCGCATTAACGACG | ||
| LT-11F | CGTCGTTAATGCGCTGGTTC | 4544 (584, 3960) | 39.5 |
| LT-11R | ACTATTGCTTGTGGTCGCCA | ||
| LT-12F | CGACCACAAGCAATAGTTTTTGA | 3583 (327, 3256) | 44.5 |
| LT-12R | CAGCACACGACCGACTTTTG | ||
| LT-13F | GTGTGGTACGTGTGGCAAAA | 2247 (1274, 973) | 38.5 |
| LT-13R | TCAGACATCCGCCGTGCGTG |
Note: “a” represents the total length of the PCR (Polymerase Chain Reaction) product, “b” is the length of exons in the total length, and “c” is the length of introns.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



