Submitted:
21 November 2025
Posted:
24 November 2025
You are already at the latest version
Abstract
We recently published that phycocyanin, a phycobiliprotein which accounts for up to 20% of Arthrospira platensis dry weight, has a powerful anti-aging effect, greatly extending the chronological life span (CLS) of yeast cells grown in synthetic-defined medium, both under caloric restriction (CR) conditions (0.2% glucose) or under non-CR conditions (2% glucose). In this study, to explore the molecular mechanisms underlying the effects of phycocyanin, we investigated its impact on key signalling pathways involved in aging. Specifically, we performed CLS experiments using ras2Δ and snf1Δ yeast mutants. The Snf1 pathway is known to promote longevity (anti-aging), whereas the Ras2/PKA pathway accelerates aging (pro-aging). We show that, while in the snf1 mutant the anti-aging effect of phycocyanin was still evident, in the ras2Δ mutant phycocyanin did not appear to exert any anti-aging activity, suggesting that the Ras2/PKA pathway may be essential for mediating the anti-aging effect of phycocyanin. To evaluate the activity of phycocyanin under different nutritional conditions, we performed the CLS experiment in a YPDA rich medium. We show that in this medium phycocyanin accelerated the chronological aging of yeast cells, greatly decreasing the CLS, both when glucose was present at low (0.2%) or at high (2%) concentration. Our data suggest that Saccharomyces cerevisiae could serve as a model not only to investigate the anti-aging properties and targets of phycocyanin, but also its potential side effects, possibly present in higher eukaryotes under certain conditions.
Keywords:
ROS
; aging
; chronological life span
; Yeast
; S. cerevisiae
; nutrient sensing
; A. platensis
; Spirulina
1. Introduction
Aging is a complex and inevitable process, marked by a progressive decline in cellular and tissue function. This deterioration increases the risk of developing chronic diseases, including cancer, cardiovascular conditions, and neurodegenerative disorders. [1,2]. In recent years, there has been growing attention to develop strategies aimed at counteracting the effects of aging, with particular interest in marine-derived compounds [3]. In this context, Arthrospira platensis, more commonly known as Spirulina, has attracted growing scientific and commercial interest. This cyanobacterium is an exceptionally rich source of bioactive compounds, many of which are already known for their potential health-promoting effects, while others have yet to be fully identified and characterized. Phycocyanin is a complex composed of proteins belonging to the phycobiliprotein family and water-soluble pigments of photosynthesis, the phycocyanobilins, and it is particularly abundant in the cyanobacterium A. platensis, constituting approximately 20% of its dry weight [4]. Phycocyanin is an oligomeric protein composed of α and β subunits with a molecular weight of 16.3 kD and 18.9 kD, respectively [4]. Several scientific studies suggest that A. platensis may have various applications in both medicinal and food industries [5] and phycocyanin is incorporated into diets in various forms such as powders, tablets, extracts, and supplements. In particular, phycocyanin is believed to play a key role in the anti-cancer, anti-inflammatory, and antioxidant effects of A. platensis [6,7,8]. It has been also demonstrated that phycocyanin reduces oxidative stress in both yeast and mammalian systems [9,10].
Recently we investigated the effect of phycocyanin on the survival of Saccharomyces cerevisiae cells growing in synthetic-defined medium and showed that phycocyanin has proven to possess anti-aging properties, greatly extending the chronological life span of yeast cells regardless of whether the glucose was present at low (0.2%) or high (2%) concentration [11]. Aging, together with its wide range of age-associated conditions, is a complex, multifactorial process influenced by both genetic and environmental factors, involving many interconnected pathways and regulatory networks. In this regard, the yeast Saccharomyces cerevisiae serves as a valuable model organism for studying eukaryotic cells and offers important insights into the biology of human aging. This is because the key molecular pathways governing aging are evolutionarily conserved between yeast and higher eukaryotes [12,13,14,15]. In this study, we examined how phycocyanin influences key signalling pathways associated with aging in yeast to understand the molecular mechanisms behind its anti-aging properties. We show data suggesting that the Ras2/PKA pathway may be essential for mediating the anti-aging effect of phycocyanin. Moreover, we show that in rich medium, unlike what happens in synthetic-defined medium, phycocyanin accelerated the chronological aging of yeast cells, leading to a marked reduction in their chronological life span, indicating that S. cerevisiae may be a valuable model not only for studying the anti-aging effects and molecular targets of phycocyanin, but also its potential side effects, possibly present in higher eukaryotic systems under certain conditions.
2. Results and Discussion
2.1. Effect of Phycocyanin on Saccharomyces Cerevisiae snf1Δ and ras2Δ Mutants
Saccharomyces cerevisiae, commonly known as budding yeast, is a key model organism for studying eukaryotic cells and offers valuable insights into the biological mechanisms underlying human aging [13,14]. We recently demonstrated that phycocyanin exerts a potent anti-aging effect by significantly extending the chronological lifespan (CLS) of yeast cells cultured in synthetic-defined medium (SD medium), under both calorie restriction (CR) conditions (0.2% glucose) or non-CR conditions (2% glucose) [11]. In this study, we compared the survival of the wild-type strain and two mutant strains (snf1Δ and ras2Δ mutants) to explore the influence of phycocyanin on signal transduction pathways associated with longevity and to identify its physiologically relevant targets. The Snf1/AMPK pathway (sucrose-non-fermenting/AMP-activated protein kinase) is an anti-aging pathway [16], while the Ras2/PKA (Rat sarcoma/protein kinase A) pathway is a pro-aging one [17,18]. We inoculated wild-type cells and the snf1Δ and ras2Δ mutants in SD medium containing 0.2% glucose or 2% glucose, in the absence and presence of phycocyanin, respectively 4.3 mg/mL (±0.1 mg/mL) and 3.9 mg/mL (±0.4 mg/mL) (day -3 of experiment). When the carbon source present in the culture medium is completely consumed, cells enter the stationary phase, do not carry out any cell division and reach the maximum cell concentration (day 0 CLS). We monitored the progressive loss of viability of these stationary phase cells kept in their culture medium from day 0 until day 12 (SD medium containing 0.2% glucose) or day 8 (SD medium containing 2% glucose) and used them as an index of chronological aging [19] (Figure 1 and Figure 2). In the wild-type strain grown under conditions of caloric restriction (0.2% glucose), phycocyanin showed a potent anti-aging effect, confirming the previously published data [11]. Cell survival increased significantly on day 12 of CLS, where the viability of cells treated with phycocyanin was approximately double that of untreated cells (Figure 1). The anti-aging effect of phycocyanin was also clear in the snf1Δ strain (Figure 1). In particular, on day 6 of CLS, all cells in the untreated sample were non-viable, while the presence of phycocyanin allowed the survival of a small fraction of cells. Furthermore, as expected, the snf1Δ strain was less long-lived than the wild-type strain (Figure 1). The ras2Δ strain, known for being longer-lived compared to the wild-type strain [20], as also confirmed by our data (Figure 1), did not show significant differences between phycocyanin-treated and untreated samples (Figure 1). At both time points (days 6 and 12), the survival rate was indeed comparable. As previously published [11], phycocyanin showed a potent anti-aging activity also when the wild-type strain was grown in SD medium containing 2% glucose (non-caloric restriction conditions) (Figure 2). Moreover, as expected, in this strain grown in the absence of phycocyanin, cell survival was lower than in CR conditions (Figure 1 and Figure 2). Like in the wild type strain, also in the snf1Δ strain, the presence of phycocyanin increased cell survival, demonstrating the anti-aging activity of the extract in this mutant (Figure 2). Furthermore, in the absence of phycocyanin, cell survival was higher than that observed under CR conditions (Figure 1 and Figure 2), consistent with the known role of the Snf1 protein being involved in metabolic pathways activated under conditions of low glucose availability [16]. Finally, in the ras2Δ strain, phycocyanin did not appear to exert any anti-aging activity (Figure 2) also in this nutritional condition. This data suggests that the Ras2/PKA pathway may be essential for mediating the anti-aging effect of phycocyanin, suggesting a possible direct involvement of this signal transduction pathway in the molecular mechanism of action of this compound. Furthermore, as expected, the ras2Δ strain was longer-lived than the wild-type strain (Figure 2).
2.2. Effect of Phycocyanin on Sacchoromyces Cerevisiae Reactive Oxygen Species Accumulation in snf1Δ and ras2Δ Mutants
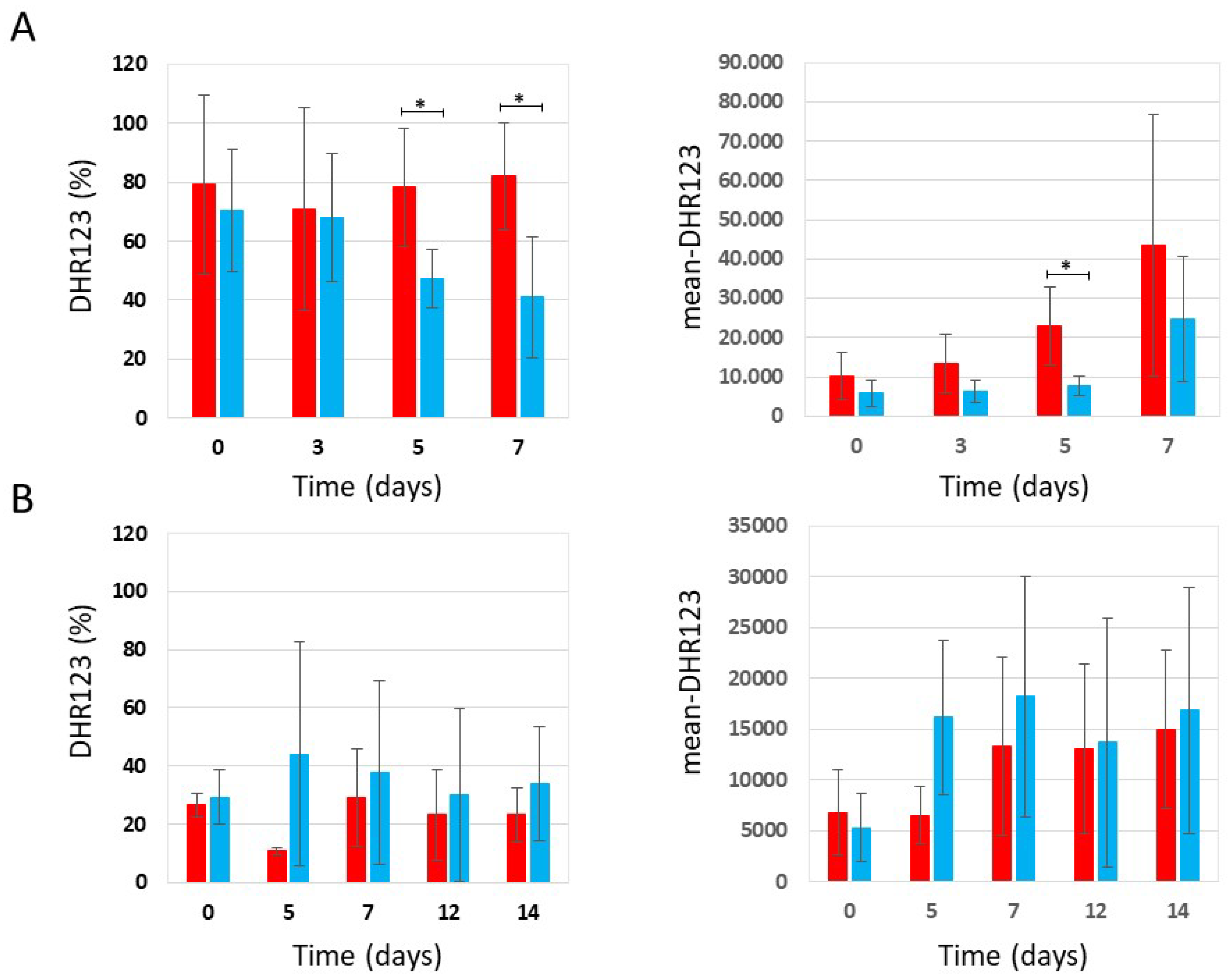
Since oxidative stress and the accumulation of reactive oxygen species (ROS) play a key role in aging [21], we determined the accumulation of ROS in snf1Δ and ras2Δ mutants in the context of CLS experiments. Therefore, as described previously (see Figure 1 and Figure 2), the presence of phycocyanin in the medium significantly increased the chronological aging of wild type and snf1Δ cells, while in the ras2Δ strain, phycocyanin did not appear to exert any anti-aging activity. In parallel, using a cytofluorimeter, we determined the accumulation of ROS by staining the chronologically aged cells with dihydrorhodamine123 (DHR123) (Supplementary Figure S1). Flow cytometry data were analysed using CytExpert and Floreada.io software. The results obtained from the cytofluorimetric analysis conducted on the wild type strain grown under conditions of caloric restriction (0.2% glucose) confirmed the data recently published by Nova et al. [11]. In particular, we observed that wild type cells treated with phycocyanin, although more viable than untreated controls (Figure 1), exhibited a higher percentage of DHR123-positive cells (Figure 3A) and an increased mean fluorescence intensity (mean-DHR123) within the population (Figure 3B). These findings are consistent with previously published data [11] and suggest that phycocyanin may exert a protective effect by promoting cell survival through elevated ROS levels. This increase in ROS likely triggers an adaptive response to oxidative and other forms of stress, known as hormesis, which renders cells more resistant to subsequent, otherwise harmful, exposures to the same stressor [22,23]. Similar to what was observed for wild type cells, snf1Δ cells treated with phycocyanin —although longer-lived than the untreated ones (Figure 1)—also displayed a higher percentage of DHR123-positive cells (Figure 3A) and an increased mean fluorescence intensity (mean-DHR123) within the population (Figure 3B) compared to untreated cells. These findings suggest that, in this strain as well, phycocyanin may promote cell survival through hormesis-related mechanisms. Finally, also in ras2Δ cells, although no differences in cell viability were observed between samples treated or untreated with phycocyanin (Figure 1), treatment with this compound led to an increased percentage of DHR123-positive cells and a higher mean fluorescence intensity (mean-DHR123) within the population (Figure 3A and Figure 3B).
Under non-CR conditions (2% glucose), flow cytometry analysis showed that wild-type cells treated with phycocyanin, which were more viable than untreated cells (Figure 2), exhibited a lower percentage of DHR123-positive cells (Figure 4A) and a reduced mean fluorescence intensity (mean DHR123) (Figure 4B) compared with untreated cells. These findings suggest that under these conditions phycocyanin may exert a protective effect and promote cell survival through a mechanism independent of hormesis. As observed in wild-type cells, snf1Δ cells treated with phycocyanin also exhibited greater longevity compared with untreated cells (Fig. 2). In these cells, the percentage of DHR123-positive cells was similar to that found in untreated controls (Figure 4A). However, the mean fluorescence intensity (mean DHR123) within the population was lower than that of untreated cells (Figure 4B), suggesting that, as in the previous strain, phycocyanin may confer a protective effect and promote cell survival through a mechanism distinct from hormesis. Finally, in ras2Δ cells, no differences in viability were observed between samples treated and untreated with phycocyanin (Figure 2) and both the percentage of DHR123-positive cells and the mean fluorescence intensity (mean-DHR123) in the population between phycocyanin-treated and untreated cells was comparable (Figure 4A and 4B). Overall, our findings suggest that, in both the wild-type and the snf1Δ mutant, phycocyanin exerts a protective effect and promotes cell survival through hormesis when the cells are grown in SD medium containing 0.2% glucose. Conversely, in SD medium containing 2% glucose, phycocyanin appears to promote cell survival through a mechanism other than hormesis.
2.3. Phycocyanin Accelerates the Chronological Aging of Yeast Cells Grown in Rich Medium
It is known that the composition of the culture medium in which Saccharomyces cerevisiae cells are grown plays a significant role in regulating various parameters, including growth and proliferation, stress tolerance, and chronological aging [24,25,26]. To investigate this aspect, we performed the chronological aging experiments in YPDA medium, a medium particularly rich in nutrients (amino acids, peptides, nucleosides, etc.). Yeast cells were grown in YPDA medium under CR (0.2% glucose) or non-CR (2% glucose) conditions, in the absence and presence of phycocyanin, respectively 4.6 mg/mL (±0.4 mg/mL) and 4.6 mg/mL (±0.2 mg/mL) (day -3 of experiment), and their colony-forming units (CFU) ability was measured over time. Our results showed that in rich medium phycocyanin did not exhibit an anti-aging effect, instead significantly reduced their chronological lifespan at both low (0.2%) and high (2%) glucose concentrations (Figure 5).
As previously shown by Wu et al., cells grown in rich medium containing 2% glucose displayed increased longevity than those cultured in the same medium containing a lower % of glucose [27]. Here we show that this phenotype is independent of the presence or absence of phycocyanin, suggesting that caloric restriction confers an anti-aging effect only in the context of the SD medium. However, phycocyanin under these rich nutritional conditions behaves in an opposite way and reduces the chronological lifespan.
In parallel with the CLS experiment, we determined the accumulation of reactive oxygen species by staining the chronologically aged cells with DHR123. Our results showed that in YPDA medium containing 0.2% glucose, phycocyanin-treated cells showed a lower percentage of DHR123-positive cells and a lower mean fluorescence intensity (mean-DHR123) in the population than untreated cells (Figure 6A), while in YPDA medium containing 2% glucose no difference between treated and untreated cells was observed (Figure 6B).
3. Materials and Methods
3.1. Yeast Strains and Media
Strains used in this study: W303-1A (MATa ade2-1 can1-100 his3-11,15 leu2-3112 trp1-1 ura3-1) [28]; W303-1A with ras2::URA3 (our laboratory); W303-1A with snf1:HIS3 (kindly provided by J. Winderickx, KU Leuven). Synthetic-defined medium (SD) contained either 0.2% glucose or 2% glucose, 6.7 g/l YNB w/o amino acids (supplied by ForMedium™, Swaffham, UK) and the selective drop-out CSM (Complete Supplement Mixture, supplied By ForMedium™, Swaffham, UK) -HIS-LEU-TRP-URA; in addition, 100 mg/L adenine and 50 mg/L histidine, leucine, tryptophan and uracil were added. The rich medium (YPDA) consisted of 1% (w/v) yeast extract, 2% (w/v) peptone, 100 mg/L adenine and either 0.2% or 2% (w/v) glucose. YPDA plates contained 2%wv−1 glucose, 2%wv−1 peptone, 1%wv−1 yeast extract, 100mg/L adenine and 2%wv−1 agar. Culture density was measured with a Coulter Counter (Coulter mod. Z2) on mildly sonicated, diluted samples.
Phycocyanin (Organic Spirulina Extract, kindly provided by Algavista, Chennai, India; for more details see Nova et al. [11]) was dissolved in the proper medium (the concentration slightly varies among experiments) and sterilized by filtration through 0.22 μm PES filters (Biosigma). Given the difficulty in filtering the culture medium containing phycocyanin and since a deposit of material on the filter was observed during the filtration step, as previously described, a calibration curve was used to determine whether phycocyanin loss occurred during this step and to determine the quantity of phycocyanin actually present in the medium [11]. On average, the concentration of phycocyanin after filtration was approximately 80% of the phycocyanin dissolved in the fresh medium. The pH of the fresh medium was 5.7 regardless of the presence of phycocyanin.
3.2. Aging Experiments and Cell Viability
We used one of the established procedures to measure chronological life span (CLS), as described by Fabrizio and Longo [19]. Assessment of clonogenicity using the CLS assay is the most used method to quantify cell viability, which reflects a cell’s ability to divide. It should be emphasized that compromised proliferation does not necessarily lead to cell death; however, the literature suggests that clonogenic capacity represents an excellent indicator of cell death in a wide range of contexts, i.e. it represents a valid approximation to quantify survival in yeast populations [29,30,31,32]. The yeast cells were grown in either SD medium or YPDA medium (containing either 0.2% glucose or 2% glucose) at 30 °C on a shaker at 160 rpm and prolonged incubation in their original exhaust medium. Chronological aging was monitored after arrest into stationary phase (day 0 of CLS) as progressive loss of cell viability. In particular, cells were plated on YPDA agar plates in triplicate and colony forming units (CFUs) were used to monitor viable cells after 3 days of growth at 30° C.
3.3. Staining with Dihydrorhodamine 123 (DHR123)
ROS were detected with DHR123 (Sigma Aldrich) essentially as described by Madeo et al. [33]. DHR123 was added directly to the culture medium at the final concentration of 5 µg/mL (from a 2.5 mg/mL stock solution in ethanol) and cells were incubated for 2 h at 30 ° C with shaking in the dark. At the end of this period, cells were diluted in 50 mM TrisHCl pH 7.5 to 107 cell/mL and analysed using a cytofluorimeter (CytoFLEX©, Beckman Coulter, Inc., Brea, CA, USA), with excitation and emission settings of 488 and 525–550 nm (filter FL 1-H). A total of 20.000 events were acquired for each sample and data were processed using CytExpert software (Version 2.6) (Beckman) and Floreada.io software (a free web-based tool for flow cytometry analysis; https://floreada.io/).
3.4. Statistical Analysis
All the experiments were conducted at least in triplicate, and the Mean and Standard Deviation were shown. The Student’s t test was used for assessing the significance of the experimental data. The experimental data were elaborated with Excel TM.
4. Conclusions
Phycocyanin, a complex consisting of proteins belonging to the phycobiliprotein family and of a pigment, phycocyanobilin, that can account for up to 20% of Spirulina’s dry weight, is considered one of the main contributors to the functional properties attributed to this cyanobacterium: numerous studies report antioxidant, anti-inflammatory, and even anticancer activities, making it a highly promising candidate for nutraceutical, pharmacological, and biotechnological applications [6,7]. In this paper we used Saccharomyces cerevisiae as a model organism to study the anti-aging activity of phycocyanin purified from A. platensis (formerly Spirulina platensis). We showed that growth conditions and nutrient sensing modulate the anti-aging activity of phycocyanin in budding yeast. In particular, we showed that phycocyanin has a powerful anti-aging effect, greatly extending the chronological life span of yeast cells grown in SD medium both under calorie restriction (CR) conditions (0.2% glucose) or non-CR conditions (2% glucose), while in rich medium (YPDA) phycocyanin does not exhibit an anti-aging activity, instead significantly reduces yeast chronological lifespan at both low (0.2%) and high (2%) glucose concentrations. Additionally, our data support prior studies [27] demonstrating that cells grown in rich medium containing 2% glucose exhibit greater longevity than those cultured in the same medium containing a lower % of glucose. We further showed that this effect occurs regardless of the presence or absence of phycocyanin, indicating that caloric restriction promotes anti-aging benefits only within the context of the synthetic medium. Taken together our results support the idea that nutrient balance, in addition to caloric restriction, is a key factor for longevity of yeast.
Understanding the mechanism of action of molecules is crucial for several reasons. Identifying the specific biological targets they interact with helps to explain the effects they produce. Furthermore, knowledge of the mechanism allows for better prediction of potential side effects and toxicity. In order to find physiologically relevant phycocyanin targets, we studied the effect of this compound in mutants affecting longevity, in particular the snf1∆ and the ras2∆ mutants, and found that that the Ras2/PKA pathway may be essential for mediating the anti-aging effect of phycocyanin, suggesting a possible direct involvement of this signal transduction pathway in the molecular mechanism of action of phycocyanin.
Since oxidative stress and the accumulation of reactive oxygen species are considered important factors causing aging and since data in literature attribute an antioxidant role to phycocyanin [6,7,21], in parallel with the chronological life span experiments, we determined the accumulation of reactive oxygen species by staining the chronologically aged cells with DHR123. Our findings suggest that, in both the wild-type and the snf1∆ mutant, phycocyanin exerts a protective effect and promotes cell survival through hormesis [22,23], an adaptive response to a variety of stresses according to which low doses of ROS act as essential signalling molecules to promote metabolic health and longevity, when the cells are grown in SD medium containing 0.2% glucose. Conversely, in SD medium containing 2% glucose, phycocyanin appears to promote cell survival through a mechanism other than hormesis.
In summary, our findings suggest that the anti-aging action of phycocyanin is strongly dependent on the metabolism of S. cerevisiae and that this microorganism may represent a useful model organism to investigate the anti-aging properties and targets of substances, but also their potential side effects, possibly present in higher eukaryotes under certain conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, S.C. and E.M.; Investigation and data acquisition, D.C. and St.C.; Data Curation, S.C. and E.M.; Writing – Original Draft Preparation, S.C.; Writing – Review & Editing, S.C. and E.M.; Supervision, S.C and E.M.; Project Administration, S.C.; Funding Acquisition, S.C. and E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by FA – University of Milano Bicocca grants to S.C. and E.M.
Acknowledgments
We thank Algavista, Chennai, India for providing Phycocyanin Powder.
Conflicts of interest
The authors declare that there is no conflict of interest regarding the publication of this manuscript.
References
- Hou, Y.; Dan, X.; Babbar, M.; et al. Ageing as a risk factor for neurodegenerative disease. Nat Rev Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef]
- Costantino, S.; Paneni, F.; Cosentino, F. Ageing, metabolism and cardiovascular disease. J Physiol. 2016, 594, 2061–2073. [Google Scholar] [CrossRef]
- Haque, N.; Parveen, S.; Tang, T.; Wei, J.; Huang, Z. Marine Natural Products in Clinical Use. Mar. Drugs 2022, 20, 528. [Google Scholar] [CrossRef]
- Bannu, S.A.; Lomada, D.; Gulla, S.; Chandrasekhar, T.; Reddanna, P.; Reddy, M.C. Potential Therapeutic Applications of C-Phycocyanin. Curr. Drug Metab 2019, 20, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Maddiboyina, B.; Vanamamalai, H.K.; Roy, H.; Gandhi, R.S.; et al. Food and drug industry applications of microalgae Spirulina platensis: A review. J Basic Microbiol. 2023, 63, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Romay, C.; Armesto, J.; Remirez, D.; Gonzalez, R.; Ledon, N.; Garcıa, I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch Toxicol 2016, 90, 1817–1840. [Google Scholar] [CrossRef]
- Li, Y. The Bioactivities of Phycocyanobilin from Spirulina. J. Immunol. Res. 2022, 2022, 4008991. [Google Scholar] [CrossRef]
- Rogova, A.G.; Sergeeva, Y.E.; Sukhinov, D.V.; Ivaschenko, M.V.; Kuvyrchenkova, A.P.; Vasilov, R.G. The Effect of Phycocyanin Isolated from Arthrospira platensis on the Oxidative Stress in Yeasts. Nanobiotechnology Reports 2023, 18, 126–131. [Google Scholar] [CrossRef]
- Pleonsila, P.; Soogarunb, S.; Suwanwong, Y. Anti-oxidant activity of holo- and apo-c-phycocyanin and their protective effects on human erythrocytes. Int. J. Biol. Macromol. 2013, 60, 393–398. [Google Scholar] [CrossRef]
- Nova, M.; Citterio, S.; Martegani, E.; Colombo, S. Unraveling the Anti-Aging Properties of Phycocyanin from the Cyanobacterium Spirulina (Arthrospira platensis). Int. J. Mol. Sci. 2024, 25, 4215. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Longo, V.D.; Fabrizio, P. Chronological aging in Saccharomyces cerevisiae. Subcell Biochem 2012, 57, 101–121. [Google Scholar] [CrossRef]
- Sampaio-Marques, B.; Burhans, W.C.; Ludovico, P. Yeast at the forefront of research on ageing and age-related diseases. Prog Mol Subcell Biol 2019, 58, 217–242. [Google Scholar] [CrossRef]
- Dahiya, R.; Mohammad, T.; Alajmi, M.F.; Rehman, M.T.; Hasan, G.M.; Hussain, A.; Hassan, M.I. Insights into the Conserved Regulatory Mechanisms of Human and Yeast. Aging. Biomolecules 2020, 10, 882. [Google Scholar] [CrossRef] [PubMed]
- Wierman, M.B.; Maqani, N.; Strickler, E.; Li, M.; Smith, J.S. Caloric Restriction Extends Yeast Chronological Life Span by Optimizing the Snf1 (AMPK) Signaling Pathway. Mol. Cell. Biol. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span—From yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Shadel, G.S.; Kaeberlein, M.; Kennedy, B. Replicative and chronological aging in Saccharomyces cerevisiae. Cell Metab 2012, 16, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, P.; Longo, V.D. The chronological life span of Saccharomyces cerevisiae. Methods Mol Biol 2007, 371, 89–95. [Google Scholar] [CrossRef]
- Fabrizio, P.; Pletcher, S.D.; Minois, N.; Vaupel, J.W.; Longo, V.D. Chronological aging-independent replicative life span regulation by Msn2/Msn4 and Sod2 in Saccharomyces cerevisiae. FEBS Lett. 2004, 557, 136–142. [Google Scholar] [CrossRef]
- De la Torre-Ruiz, M.A.; Pujol, N.; Sundaran, V. Coping with oxidative stress. The yeast model. Curr. Drug Targets 2015, 16, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Nascarella, M.; Pressman, P.; et al. Hormesis determines lifespan. Ageing Research Reviews 2024, 94, 102181. [Google Scholar] [CrossRef] [PubMed]
- Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C.; Traverso, N. Hormesis and Oxidative Distress: Pathophysiology of Reactive Oxygen Species and the Open Question of Antioxidant Modulation and Supplementation. Antioxidants 2022, 11, 1613. [Google Scholar] [CrossRef]
- Aris, J.P.; Fishwick, L.K.; Marraffini, M.L.; Seo, A.Y.; Leeuwenburgh, C.; Dunn, W.A. Jr Amino acid homeostasis and chronological longevity in Saccharomyces cerevisiae. Sub-cellular biochemistry 2012, 57, 161–186. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wei, M.; Mirzaei, H.; Madia, F.; Mirisola, M.; Amparo, C.; Chagoury, S.; Kennedy, B.; Longo, V.D. Tor-Sch9 deficiency activates catabolism of the ketone body-like acetic acid to promote trehalose accumulation and longevity. Aging cell 2014, 13, 457–467. [Google Scholar] [CrossRef]
- Jiang, J.C.; Jaruga, E.; Repnevskaya, M.V.; Jazwinski, S.M. An intervention resembling caloric restriction prolongs life span and retards aging in yeast. The FASEB Journal 2000, 14, 2135–2137. [Google Scholar] [CrossRef]
- Wu, Z.; Quan, S.; Huang, L.D. Dietary Restriction Depends on Nutrient Composition to Extend Chronological Life span in Budding Yeast Saccharomyces cerevisiae. PLoS ONE 2013, 8, e64448. [Google Scholar] [CrossRef]
- Thomas, B.J.; Rothstein, R. Elevated recombination rates in transcriptionally active DNA. Cell 1989, 56, 619–630. [Google Scholar] [CrossRef]
- Madeo, F.; Herker, E.; Maldener, C.; Wissing, S.; Lächelt, S.; Herlan, M.; Fehr, M.; Lauber, K.; Sigrist, S.J.; Wesselborg, S.; Fröhlich, K.U. A caspase-related protease regulates apoptosis in yeast. Mol Cell 2002, 9, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Büttner, S.; Eisenberg, T.; Carmona-Gutierrez, D.; Ruli, D.; Knauer, H.; Ruckenstuhl, C.; Sigrist, C.; Wissing, S.; Kollroser, M.; Fröhlich, K.U.; Sigrist, S.; Madeo, F. Endonuclease G regulates budding yeast life and death. Mol Cell 2007, 25, 233–246. [Google Scholar] [CrossRef]
- Carmona-Gutiérrez, D.; Bauer, M.A.; Ring, J.; Knauer, H.; Eisenberg, T.; Büttner, S.; Ruckenstuhl, C.; Reisenbichler, A.; Magnes, C.; Rechberger, G.N.; Birner-Gruenberger, R.; Jungwirth, H.; Fröhlich, K.U.; Sinner, F.; Kroemer, G.; Madeo, F. The propeptide of yeast cathepsin D inhibits programmed necrosis. Cell Death Dis 2011, 2, e161. [Google Scholar] [CrossRef]
- Ruckenstuhl, C.; Netzberger, C.; Entfellner, I.; Carmona-Gutierrez, D.; Kickenweiz, T.; Stekovic, S.; Gleixner, C.; Schmid, C.; Klug, L.; Sorgo, A.G.; Eisenberg, T.; Büttner, S.; Mariño, G.; Koziel, R.; Jansen-Dürr, P.; Fröhlich, K.U.; Kroemer, G.; Madeo, F. Lifespan Extension by Methionine Restriction Requires Autophagy-Dependent Vacuolar Acidification. PLoS Genet 2014, 10, e1004347. [Google Scholar] [CrossRef]
- Madeo, F.; Fröhlich, E.; Ligr, M.; Grey, M.; Sigrist, S.J.; Wolf, D.H.; Fröhlich, K.-U. Oxygen stress: A regulator of apoptosis in yeast. J. Cell Biol. 1999, 145, 757–767. [Google Scholar] [CrossRef]
Figure 1.
Cell survival in W303-1A, snf1Δ and ras2Δ cells grown in SD medium containing 0.2% glucose in the presence and absence of phycocyanin (given at the moment of inoculation). Cell viability of W303-1A, snf1Δ and ras2Δ cells, either untreated (red) or treated (blue) with 4.3 mg/ml (+/-0.1 mg/ml) phycocyanin, was analysed by measuring colony-forming units (CFU) after 3 days of growth at 30 °C. Cell survival is expressed as % to the CFU at time zero. The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
Figure 1.
Cell survival in W303-1A, snf1Δ and ras2Δ cells grown in SD medium containing 0.2% glucose in the presence and absence of phycocyanin (given at the moment of inoculation). Cell viability of W303-1A, snf1Δ and ras2Δ cells, either untreated (red) or treated (blue) with 4.3 mg/ml (+/-0.1 mg/ml) phycocyanin, was analysed by measuring colony-forming units (CFU) after 3 days of growth at 30 °C. Cell survival is expressed as % to the CFU at time zero. The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
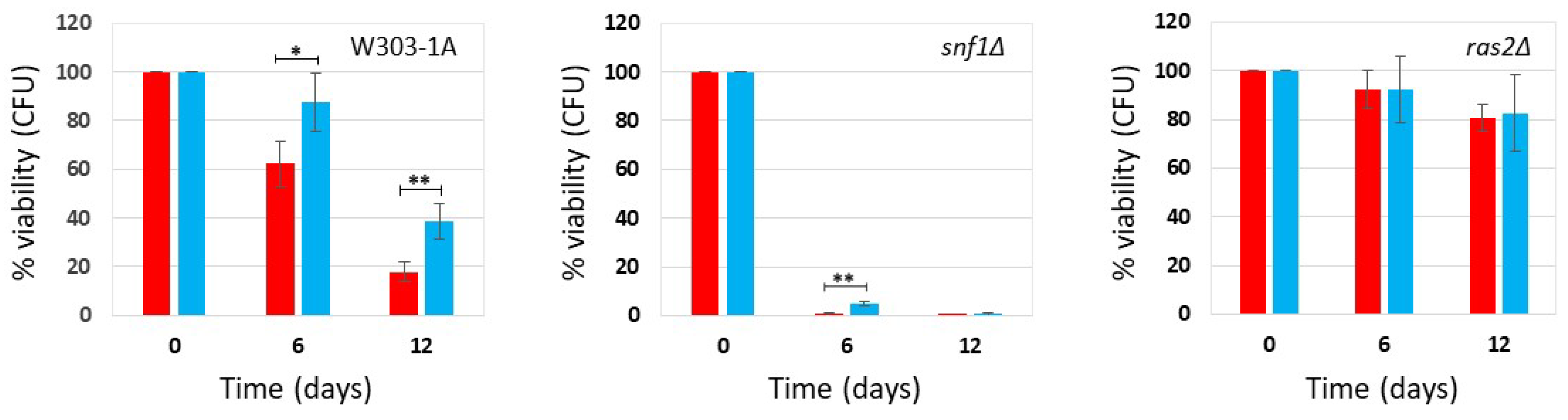
Figure 2.
Cell survival in W303-1A, snf1Δ and ras2Δ cells grown in SD medium containing 2% glucose in the presence and absence of phycocyanin (given at the moment of inoculation). Cell viability of W303-1A, snf1Δ and ras2Δ cells, either untreated (red) or treated (blue) with 3.9 mg/ml (+/-0.4 mg/ml) phycocyanin, was analysed by measuring colony-forming units (CFU) after 3 days of growth at 30 °C. Cell survival is expressed as % to the CFU at time zero. The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
Figure 2.
Cell survival in W303-1A, snf1Δ and ras2Δ cells grown in SD medium containing 2% glucose in the presence and absence of phycocyanin (given at the moment of inoculation). Cell viability of W303-1A, snf1Δ and ras2Δ cells, either untreated (red) or treated (blue) with 3.9 mg/ml (+/-0.4 mg/ml) phycocyanin, was analysed by measuring colony-forming units (CFU) after 3 days of growth at 30 °C. Cell survival is expressed as % to the CFU at time zero. The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
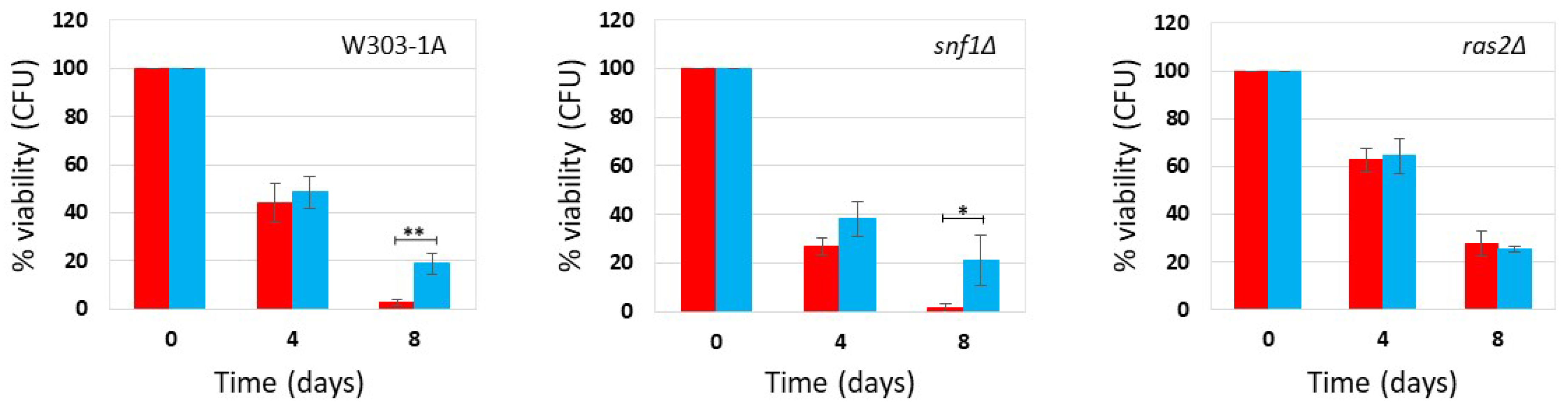
Figure 3.
Flow cytometric analysis of W303-1A, snf1Δ and ras2Δ cells grown in synthetic medium containing 0,2% glucose, either untreated (red bars) or treated (blue bars) with 4.3 mg/ml (+/-0.1 mg/ml) phycocyanin. Dihydrorhodamine 123 (DHR123) was used to assay ROS accumulation (for more details, see Supplementary Figures S1 and S2). (A) Percentage of cells positive for DHR123 staining; (B) Mean fluorescence intensity (mean-DHR123). The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
Figure 3.
Flow cytometric analysis of W303-1A, snf1Δ and ras2Δ cells grown in synthetic medium containing 0,2% glucose, either untreated (red bars) or treated (blue bars) with 4.3 mg/ml (+/-0.1 mg/ml) phycocyanin. Dihydrorhodamine 123 (DHR123) was used to assay ROS accumulation (for more details, see Supplementary Figures S1 and S2). (A) Percentage of cells positive for DHR123 staining; (B) Mean fluorescence intensity (mean-DHR123). The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.

Figure 4.
Flow cytometric analysis of W303-1A, snf1Δ and ras2Δ cells grown in synthetic medium containing 2% glucose, either untreated (red bars) or treated (blue bars) with 3.9 mg/ml (+/-0.4 mg/ml) phycocyanin. Dihydrorhodamine 123 (DHR123) was used to assay ROS accumulation (for more details, see Supplementary Figures S1 and S2). (A) Percentage of cells positive for DHR123 staining; (B) Mean fluorescence intensity (mean-DHR123). The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
Figure 4.
Flow cytometric analysis of W303-1A, snf1Δ and ras2Δ cells grown in synthetic medium containing 2% glucose, either untreated (red bars) or treated (blue bars) with 3.9 mg/ml (+/-0.4 mg/ml) phycocyanin. Dihydrorhodamine 123 (DHR123) was used to assay ROS accumulation (for more details, see Supplementary Figures S1 and S2). (A) Percentage of cells positive for DHR123 staining; (B) Mean fluorescence intensity (mean-DHR123). The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
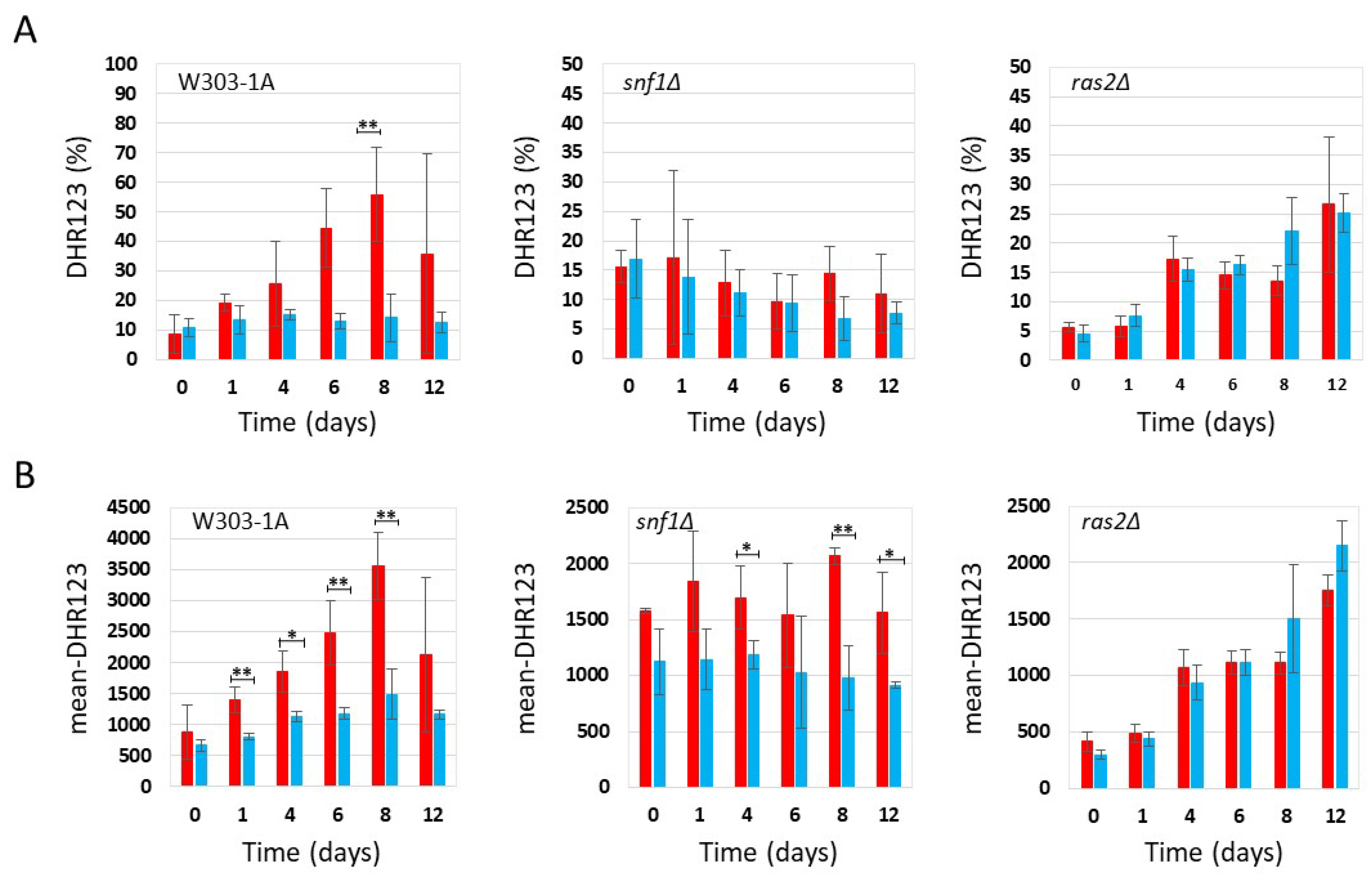
Figure 5.
Cell survival in W303-1A cells grown in YPDA medium containing 0.2% glucose (A) and 2% glucose (B) in the presence and absence of phycocyanin (given at the moment of inoculation). Cell viability of W303-1A cells, either untreated (red) or treated (blue) with 4.6 mg/mL (±0.4 mg/mL) (A) and 4.6 mg/mL (±0.2 mg/mL) (B) phycocyanin, was analysed by measuring colony-forming units (CFU) after 3 days of growth at 30 °C. Cell survival is expressed as % to the CFU at time zero. The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
Figure 5.
Cell survival in W303-1A cells grown in YPDA medium containing 0.2% glucose (A) and 2% glucose (B) in the presence and absence of phycocyanin (given at the moment of inoculation). Cell viability of W303-1A cells, either untreated (red) or treated (blue) with 4.6 mg/mL (±0.4 mg/mL) (A) and 4.6 mg/mL (±0.2 mg/mL) (B) phycocyanin, was analysed by measuring colony-forming units (CFU) after 3 days of growth at 30 °C. Cell survival is expressed as % to the CFU at time zero. The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
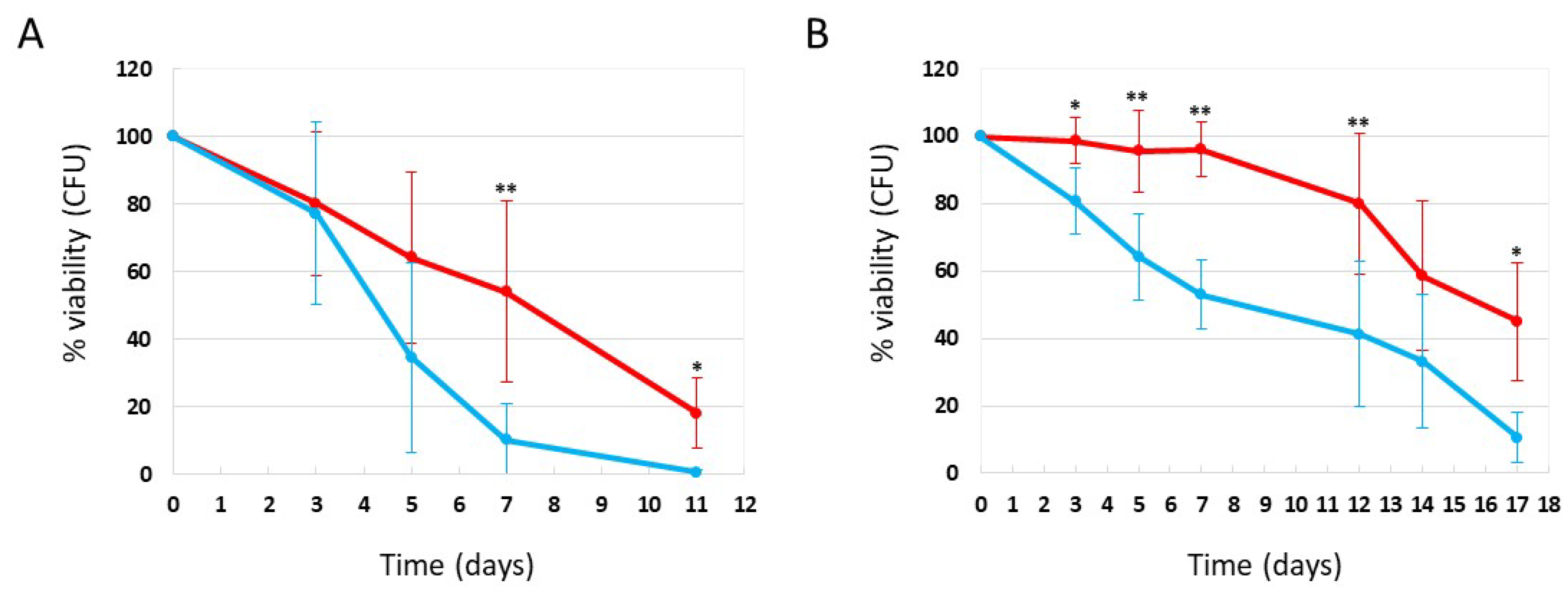
Figure 6.
Flow cytometric analysis of W303-1A cells grown in YPDA medium containing 0,2% glucose (A) or 2% glucose (B), either untreated (red bars) or treated (blue bars) with 4.6 mg/mL (±0.4 mg/mL) (A) and 4.6 mg/mL (±0.2 mg/mL) (B) phycocyanin. Dihydrorhodamine 123 (DHR123) was used to assay ROS accumulation (for more details, see Supplementary Figures S1 and S2). The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
Figure 6.
Flow cytometric analysis of W303-1A cells grown in YPDA medium containing 0,2% glucose (A) or 2% glucose (B), either untreated (red bars) or treated (blue bars) with 4.6 mg/mL (±0.4 mg/mL) (A) and 4.6 mg/mL (±0.2 mg/mL) (B) phycocyanin. Dihydrorhodamine 123 (DHR123) was used to assay ROS accumulation (for more details, see Supplementary Figures S1 and S2). The means of 3 independent experiments with standard deviations are reported. Student’s t-test ∗P < 0.05 and ∗∗P < 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



