
Submitted:
19 November 2025
Posted:
20 November 2025
You are already at the latest version
Abstract
The incretin hormones glucose-dependent insulinotropic polypeptide (GIP) and gluca-gon-like peptide-1 (GLP-1) play central roles in metabolic and cardiovascular regulation. GLP-1 receptor agonists (GLP-1RAs) are established therapies for type 2 diabetes mellitus (T2DM) and obesity because of their insulinotropic effects, weight reduction, and proven cardiovascular benefit. In contrast, GIP was historically overlooked due to reduced β-cell responsiveness in T2DM. The development of dual GIP/GLP-1 receptor agonists has reshaped this view. Tirzepatide, the first-in-class co-agonist, provides superior glycemic control and weight loss compared with selective GLP-1RAs, demonstrating synergistic actions between the two incretin pathways.
This review summarizes key physiology, pathophysiology, and therapeutic evidence in incretin biology. We describe secretion patterns, receptor distributions, and distinct ac-tions of GIP and GLP-1, as well as alterations in incretin signaling in T2DM and obesity. Cardiovascular protective mechanisms are outlined, including improvements in lipid metabolism, reductions in blood pressure, enhanced endothelial nitric oxide activity, suppression of macrophage inflammation, decreased foam-cell formation, and stabiliza-tion of atherosclerotic plaques. Emerging directions—such as dual and triple ago-nists—and unresolved questions regarding long-term vascular effects of GIP and the potential for genotype-guided incretin therapy are also discussed. Collectively, these findings highlight a shift toward integrated incretin-axis modulation for metabolic and cardiovascular disease.
Keywords:
incretin hormones
; GLP-1 receptor agonists
; GIP signaling
; tirzepatide
; dual agonists
; insulin secretion
; atherosclerosis
; cardiovascular protection
; metabolic disease
; obesity treatment
1. Introduction
1.1. The Evolving Story of Incretins
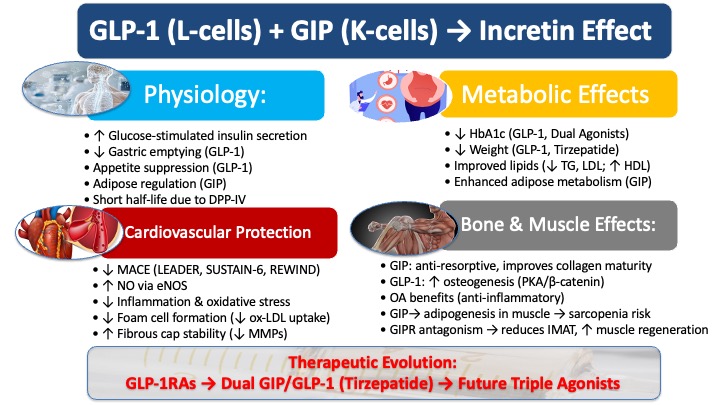
Incretin biology has become increasingly central to modern metabolic therapeutics, reshaping not only how glucose regulation is understood but also how obesity, cardiovascular disease, and multisystem metabolic dysfunction are treated. The two primary incretin hormones—glucose-dependent insulinotropic polypeptide (GIP) and glucagon-like peptide-1 (GLP-1)—are secreted from enteroendocrine cells of the small intestine following nutrient ingestion. Together, they orchestrate the “incretin effect”, a fundamental physiological mechanism that amplifies glucose-stimulated insulin secretion after oral glucose intake compared with intravenous administration [1,2]. This effect accounts for roughly 50–70% of total postprandial insulin release in healthy individuals and has served as a cornerstone for understanding gut–pancreas communication in energy and glucose homeostasis. For decades, research and drug development overwhelmingly focused on GLP-1. Its ability to stimulate insulin secretion was found to be largely preserved in patients with type 2 diabetes (T2DM), providing a clear rationale for its development as a major therapeutic target [3,4]. Beyond its glucose-dependent enhancement of insulin secretion, GLP-1 was shown to suppress inappropriate glucagon release, slow gastric emptying, promote satiety through central appetite pathways, and favor weight reduction—properties that aligned seamlessly with clinical goals for diabetes and obesity management. This led to a highly successful class of drugs—GLP-1 receptor agonists (GLP-1RAs)—that established a new benchmark for metabolic therapy, creating a high bar for any subsequent innovations [5]. The clinical success of GLP-1RAs also broadened the scope of incretin biology, revealing important cardioprotective effects that extended their relevance far beyond glycemic control.
In contrast, GIP was historically relegated to a secondary role, often considered the "neglected incretin" [1]. Although initially identified as a potent stimulator of glucose-dependent insulin secretion, its action was found to be markedly diminished in patients with T2DM, leading to the assumption that GIP was rendered functionally irrelevant in the diabetic state [3]. This apparent “GIP resistance” resulted in the hormone being largely sidelined from therapeutic consideration for many years. Early clinical studies consistently showed blunted β-cell responsiveness to GIP in T2DM, reinforcing the belief that the hormone lacked translational potential and was unlikely to contribute meaningfully to metabolic therapy [6]. However, this long-held dogma has been dramatically overturned by the development of dual GIP/GLP-1 receptor co-agonists, most notably tirzepatide. Clinical trials have demonstrated that tirzepatide achieves superior reductions in both HbA1c and body weight compared to even the most potent selective GLP-1RAs [7,8]. This remarkable efficacy has not only reignited scientific interest in GIP but has also revealed a more complex, synergistic relationship between these two hormones, prompting a fundamental re-evaluation of their integrated roles in metabolic regulation [1].
This review synthesizes the current understanding of these two pivotal hormones, beginning with their foundational physiology.
1.2. Fundamental Physiology of the Incretin System
A thorough understanding of the fundamental physiology of GIP and GLP-1 is essential to fully appreciate their roles in metabolic homeostasis and their therapeutic manipulation in disease. Although both hormones participate in the incretin response, they arise from distinct enteroendocrine cell populations, are secreted under different nutritional contexts, and engage separate receptors with unique tissue distributions. These differences shape their individual physiological actions and also determine how they interact when both pathways are stimulated simultaneously.
GIP, secreted primarily from K cells in the proximal small intestine, responds robustly to dietary fats and carbohydrates, while GLP-1, derived from L cells predominantly located in the distal ileum and colon, is released following nutrient transit through the gut [9,10]. Their receptors are widely distributed across pancreatic islets, adipose tissue, the gastrointestinal tract, the cardiovascular system, bone, and the central nervous system, allowing each hormone to influence a broad array of metabolic processes beyond insulin secretion [9,10].
Together, these characteristics form the biological foundation for their complementary roles in glucose regulation, appetite and energy balance, lipid handling, and cardiovascular function. Understanding these physiological principles is critical not only for interpreting normal metabolic regulation but also for understanding why pharmacologic targeting of these pathways—individually or in combination—has produced such profound therapeutic effects. This section provides a detailed examination of these core physiological mechanisms to establish the groundwork for later discussion of their roles in disease modulation and therapeutic innovation.
2. Physiology of GIP and GLP-1
2.1. Secretion and Metabolism
GIP originates from enteroendocrine K-cells predominantly situated in the duodenum and jejunum, whereas GLP-1 is mainly released from L-cells, which become more abundant toward the ileum and colon [11,12,13]. Their release is triggered primarily by the ingestion and absorption of nutrients, particularly carbohydrates and fats, and to a lesser extent, proteins [1,14]. Following secretion into the bloodstream, both hormones are characterized by extremely short half-lives. Natural GLP-1, for example, has a half-life of only about two minutes [15]. This rapid clearance is due to enzymatic degradation by dipeptidyl peptidase-IV (DPP-IV), an enzyme widely distributed throughout the body that efficiently inactivates both hormones [2,15].
2.2. Receptor Distribution
The biological effects of GIP and GLP-1 are mediated through their specific G protein-coupled receptors, the GIP receptor (GIPR) and GLP-1 receptor (GLP-1R). The widespread distribution of these receptors throughout the body accounts for the pleiotropic effects of these hormones, extending far beyond their primary role in glucose regulation. The table below (Table 1) synthesizes their presence in key tissues relevant to metabolic and cardiovascular health.
This broad receptor distribution is strategically significant. It underlies the capacity of incretin hormones to influence a network of organ systems, including the pancreas, heart, vasculature, and central nervous system. The presence of both GIP and GLP-1 receptors in the heart, blood vessels, and adipose tissue provides a direct anatomical and molecular basis for the cardiovascular benefits observed in clinical trials, suggesting these effects are not solely secondary to metabolic improvements but may also involve direct tissue-level signaling [16].
2.3. Core Biological Actions
While both GIP and GLP-1 share the primary function of stimulating insulin secretion, they possess distinct and sometimes opposing actions, particularly concerning glucagon secretion and gastrointestinal function.
2.3.1. Regulation of Glucose Homeostasis
The defining action of both GIP and GLP-1 is their ability to augment glucose-stimulated insulin secretion from pancreatic β-cells, a process that forms the cornerstone of the incretin effect [17]. However, their influence on the counter-regulatory hormone glucagon differs significantly [1]:
GLP-1 potently suppresses glucagon secretion from pancreatic α-cells when glucose levels are elevated, contributing to its glucose-lowering effect [18].
2.3.2. Gastrointestinal and Appetite Regulation
GLP-1 and GIP also exert markedly different effects on the gastrointestinal tract and central regulation of appetite.
GLP-1 significantly slows gastric emptying, which retards the delivery of nutrients into circulation and helps blunt postprandial glucose excursions [22,23]. At pharmacological concentrations, it also acts on the central nervous system to reduce appetite, decrease food intake, and promote satiety, which are key mechanisms underlying its efficacy in weight management [5].
GIP has no effect on gastric emptying [24]. While animal studies suggest a potential role for GIP in appetite regulation, this has not been confirmed in human studies, where its effects on food intake remain unsubstantiated [1,25].
Having established their functions in a healthy physiological state, it is crucial to examine how their roles are altered in the context of metabolic disease.
3. Pathophysiological Role in Type 2 Diabetes and Obesity
The therapeutic utility of incretin-based drugs stems directly from the altered function and response of the incretin system in metabolic diseases. The key pathophysiological changes observed in T2DM and obesity explain why GLP-1 became a successful drug target while GIP was initially overlooked.
3.1. The Diminished Incretin Effect in T2DM: The "GIP Resistance" Phenomenon
A hallmark of T2DM is a significantly reduced incretin effect, meaning that oral glucose fails to elicit the robust insulin response seen in healthy individuals [26]. While GLP-1 secretion in T2DM is largely preserved or slightly diminished, reports on GIP secretion are inconsistent, with studies showing responses ranging from normal to somewhat increased [1]. The primary driver of this dysfunction is a profound blunting of the insulinotropic action of GIP [1,27]. This phenomenon, often referred to as "GIP resistance," means that even high concentrations of GIP fail to effectively stimulate insulin secretion from the diabetic pancreas [3,28].
In stark contrast, the ability of GLP-1 to stimulate insulin secretion is largely preserved in individuals with T2DM [3,4]. This crucial difference provides the foundational rationale for the development of GLP-1RAs as a highly effective therapeutic class; they effectively "replace" the lost incretin effect by activating a signaling pathway that remains functional in the disease state. The precise molecular mechanisms underlying this GIP resistance, and whether this state can be pharmacologically reversed to restore GIP's insulinotropic efficacy, remain areas of intense clinical investigation.
3.2. Impact on Adipose Tissue and Lipid Metabolism
Beyond glucose control, incretin hormones play a role in lipid metabolism and adipose tissue function, where their effects diverge.
GIP has an anabolic role in white adipose tissue. It promotes the clearance of triglycerides from the circulation and their subsequent storage in adipocytes by enhancing the activity of lipoprotein lipase [1,29,30,31,32]. This function has fueled a long-standing debate about whether GIP may have a potentially "obesogenic" role by promoting fat deposition [33,34]. Crucially, while GIP's insulinotropic effect is blunted in T2DM, its actions on adipose tissue may be preserved, potentially contributing to altered lipid partitioning and fueling the debate over its role in the pathophysiology of metabolic disease.
GLP-1RAs, conversely, are associated with improvements in the overall lipid profile. Clinical studies have shown that they consistently reduce fasting total cholesterol, triglycerides (TG), and LDL-cholesterol while moderately increasing HDL-cholesterol [35].
These differing roles in metabolic disease set the stage for understanding their equally important and largely beneficial implications for cardiovascular health.
4. Cardiovascular Implications and Anti-Atherosclerotic Effects
Beyond their established benefits for glycemic and weight control, a major advantage of incretin-based therapies—particularly GLP-1RAs—is their proven ability to provide cardiovascular protection. The mechanisms responsible for this benefit are multifaceted, involving both indirect improvements in systemic risk factors and direct, vasoprotective effects on the arterial wall, as extensively reviewed by Wang et al. [16].
4.1. Clinical Evidence from Cardiovascular Outcome Trials (CVOTs)
Large-scale, long-term cardiovascular outcome trials (CVOTs) have provided definitive evidence that several GLP-1RAs reduce the risk of major adverse cardiac events (MACE) in patients with T2DM [16]. Across multiple outcome trials, GLP-1RAs consistently lowered major adverse cardiovascular events. Liraglutide improved cardiovascular outcomes in LEADER, semaglutide showed significant stroke and MI reductions in SUSTAIN-6, and dulaglutide demonstrated similar benefits in REWIND.
LEADER Trial: Liraglutide demonstrated a 13% reduction in the risk of MACE (a composite of cardiovascular death, nonfatal myocardial infarction, and nonfatal stroke) and a significant 22% reduction in cardiovascular death compared to placebo [16,36].
4.2. Indirect Cardioprotective Mechanisms: Modulating Systemic Risk Factors
A substantial portion of the cardiovascular benefit derived from incretin-based therapies stems from their positive impact on systemic cardiometabolic risk factors.
Improvement in Dyslipidemia: As noted previously, GLP-1RAs and dual GIP/GLP-1RAs effectively improve lipid profiles by reducing levels of triglycerides and LDL-cholesterol while raising HDL-cholesterol, which helps mitigate a key driver of atherosclerosis [16,39]. This improvement extends to postprandial lipemia, where GLP-1RAs have been shown to blunt the rise in triglycerides and atherogenic chylomicron remnants, a key contributor to cardiovascular risk that is often overlooked in fasting lipid panels [40].
Reduction in Hypertension: Clinical trials have consistently shown that GLP-1RAs and the dual agonist tirzepatide significantly reduce systolic blood pressure (SBP) [41,42]. The underlying mechanisms are thought to involve direct renal effects that promote urinary sodium excretion (natriuresis) as well as vasodilation of blood vessels [43,44,45,46].
4.3. Direct Anti-Atherosclerotic Mechanisms
In addition to improving systemic risk factors, incretins exert direct protective effects on the vasculature that are independent of their metabolic actions. These cellular mechanisms, synthesized from recent reviews [16], directly counteract the key processes involved in the formation and progression of atherosclerotic plaques.
4.3.1. Preserving Endothelial Function
Incretin receptor agonists support vascular health by promoting endothelial nitric oxide synthesis, limiting inflammatory signaling, reducing lipid uptake by macrophages, and curbing maladaptive smooth-muscle proliferation—mechanisms that collectively contribute to plaque stability.
Promoting Vasodilation: They enhance the production of nitric oxide (NO), a potent vasodilator, by activating the enzyme endothelial nitric oxide synthase (eNOS) [47,48].
4.3.2. Modulating Macrophage Activity and Plaque Inflammation
IRAs intervene at critical steps of plaque development by:
Inhibiting Foam Cell Formation: They reduce the formation of macrophage-derived foam cells—a hallmark of early atherosclerotic lesions—by inhibiting the uptake of oxidized LDL (ox-LDL) and up-regulating cholesterol efflux transporters like ABCA1 that remove excess cholesterol from cells [54,55,56,57].
4.3.3. Stabilizing Vascular Smooth Muscle Cells (VSMCs)
GLP-1RAs contribute to the stability of advanced atherosclerotic plaques, making them less prone to rupture, by:
Suppressing Abnormal VSMC Behavior: They inhibit the abnormal proliferation, migration, and phenotypic switching of VSMCs from a stable contractile state to an unstable synthetic state [61,62].
Strengthening the Fibrous Cap: They increase the collagen content within the plaque's protective fibrous cap while reducing the expression of matrix metalloproteinases (MMPs), which are enzymes that degrade the cap and increase rupture risk [63,64].
This deep understanding of incretin biology has paved the way for an evolution in therapeutic strategies, from targeting single receptors to harnessing dual-hormone action.
4.4. Dual Agonists Effect
A nuanced understanding of the dual agonists requires examining the individual contributions of GIP and GLP-1 to these vascular effects. While GIP is known to regulate the physiology of blood vessels, such as controlling blood flow rate in the hepatic portal veins [65], evidence suggests that the direct cardioprotective action on endothelial cells is primarily mediated by GLP-1 [66]. One study suggested that GLP-1, but not GIP, provides protective effects on endothelial function linked to increased levels of the second messenger, cyclic adenosine monophosphate (cAMP) [65]. Therefore, the immediate, anti-atherosclerotic benefits reported in short-term studies are largely driven by the GLP-1 component. GIP’s major contribution to superior clinical outcomes in dual agonists is consequently mediated more through its powerful systemic effects, specifically the enhanced management of total body and ectopic fat, which reduces the overall burden on the cardiovascular system.
5. The Therapeutic Landscape: From GLP-1RAs to Dual and Triple Agonists
The deepening understanding of incretin pathophysiology has directly fueled an evolution in drug development. This journey has progressed from leveraging the benefits of a single hormone to unlocking the superior potential of co-agonism, fundamentally changing the standards of care for T2DM and obesity. The advent of unimolecular dual GIP/GLP-1 receptor agonists, often termed "twincretins," marks a significant advancement by leveraging the synergistic potential of both incretin pathways. Tirzepatide, a prominent example of this class, activates both the GLP-1 and GIP receptors simultaneously, leading to enhanced metabolic effectiveness [67]. The U.S. Food and Drug Administration (FDA) recognizes Tirzepatide (Zepbound) as a first-in-class medication for weight loss, demonstrating its unique position in the therapeutic landscape.
5.1. The Established Role of GLP-1 Receptor Agonists
For over a decade, GLP-1RAs have become a cornerstone in the management of T2DM and obesity. Their success is built on a solid foundation: a preserved insulinotropic effect in T2DM, robust glucose-lowering efficacy, significant weight reduction benefits, and proven cardiovascular protection in large-scale clinical trials [68,69,70]. Their established profile has made them a preferred choice for patients with T2DM and co-existing cardiovascular disease or obesity.
5.2. The Re-Emergence of GIP: Synergies in Dual GIP/GLP-1 Receptor Agonism
The therapeutic landscape was transformed by the arrival of tirzepatide, the first-in-class dual GIP/GLP-1 receptor agonist. Clinical trial data have shown that tirzepatide achieves unprecedented efficacy, with average HbA1c reductions of approximately 2% and body weight reductions often exceeding 10 kg—results that are superior to those seen with even the most potent selective GLP-1RAs [7,8,39].
These powerful findings have challenged the long-held view that GIP possesses no therapeutic value in T2DM [1]. The superior outcomes strongly suggest an additive or synergistic interaction between GIP and GLP-1 receptor signaling pathways [71,72]. This has sparked a new wave of research aimed at elucidating the precise mechanisms by which GIP agonism, in the context of GLP-1 receptor activation, overcomes the apparent GIP resistance seen in T2DM to produce these enhanced metabolic benefits.
6. Clinical Evidence for GLP-1 RAs in Non-Diabetic Patients (Secondary Prevention)
6.1. The SELECT Trial: The Paradigm Shift
The SELECT trial provided the definitive, robust evidence for the cardiovascular benefits of GLP-1 RAs in non-diabetic patients. This randomized controlled trial evaluated Semaglutide versus placebo over an average follow-up of nearly 40 months in 17,604 non-diabetic patients (aged ≥45 years) who had established Atherosclerotic Cardiovascular Disease (ASCVD) and a BMI ≥27 kg/m2 [73].
The primary outcome, MACE—a composite of cardiovascular death, nonfatal myocardial infarction (MI), or nonfatal stroke —was significantly reduced by Semaglutide [73]. The trial established that Semaglutide reduced the risk of MACE by 20% among non-diabetic patients with obesity and pre-existing CVD [66,74]. This benefit confirmed that the mechanism of action extends beyond blood sugar regulation, representing a potential paradigm shift in cardiovascular risk management [74].
6.2. Subgroup Analysis: Efficacy in Highest-Risk Cohorts
The MACE reduction benefit demonstrated in SELECT was remarkably consistent across the entire patient population, regardless of their history of established CVD [75]. A detailed prespecified analysis focused on patients with a history of Coronary Artery Bypass Grafting (CABG), a marker for high-risk, severe ASCVD [73].
Patients with prior CABG were found to have inherently higher baseline risks for MACE and all-cause mortality. However, Semaglutide consistently reduced the MACE risk compared with placebo in both patients with a CABG history (Hazard Ratio: 0.72; 95% Confidence Interval [CI]: 0.54–0.95) and those without (HR: 0.82; 95% CI: 0.73–0.92). A critical finding for clinical implementation lies in the Absolute Risk Reduction (ARR). While the relative benefit was consistent, the ARR with Semaglutide versus placebo over the approximately 3.5-year follow-up was numerically greater in patients with a history of CABG (2.3%) than in those without (1.0%). This differential ARR demonstrates that in patients with the highest pre-existing risk of recurrent events, the intervention yields the most substantial absolute clinical reward, providing a clear rationale for prioritizing this therapy in the most vulnerable non-diabetic ASCVD patients.
6.3. Impact on Secondary CV Outcomes and Mortality
Beyond the primary MACE endpoint, Semaglutide demonstrated consistent reductions in secondary outcomes across the SELECT cohort, lowering the risks of cardiovascular death and all-cause mortality regardless of CABG status.
Further analyses on the broader class of GLP-1 RAs confirm significant benefits. These agents reduced all-cause mortality (RR 0.88), cardiovascular death (RR 0.88), and the incidence of stroke (RR 0.86). Importantly, the magnitude of MACE and mortality effects in overweight/obese patients without diabetes was similar to the effects seen in patients with diabetes, highlighting the weight-independent nature of some protective pathways [76].
The risk of hospitalization for heart failure was notably reduced by 10% overall (RR 0.90) in GLP-1 RA trials [76]. This benefit was statistically significant and particularly pronounced in the subgroup of overweight/obese non-diabetic patients (RR 0.73; 95% CI: 0.56–0.95), reinforcing the specific utility of this class for heart failure prevention in obesity [76]. Furthermore, GLP-1 RAs were associated with a 22% reduction in the incidence of coronary revascularization (RR 0.78; 95% CI: 0.69 to 0.87) compared with placebo in overweight/obese patients without diabetes [77].
6.4. Awaiting Definitive Evidence: The SURMOUNT-MMO Trial
To provide definitive, prospective evidence regarding the clinical benefits of dual agonism in non-diabetic populations, the global, randomized SURMOUNT-MMO trial (estimated completion 2027) is currently underway [78].
This pivotal trial is specifically designed to investigate the impact of once-weekly Tirzepatide versus placebo on morbidity and mortality in adults with obesity or overweight, without diabetes, who are either at risk of or have established cardiovascular disease. Its scope is unique in that it assesses both primary and secondary cardiovascular disease prevention within the same outcome trial for an incretin medication in a non-diabetic population. The comprehensive primary endpoint includes a five-component composite outcome: nonfatal myocardial infarction, nonfatal stroke, coronary revascularization, heart failure events, or death from any cause. The results of SURMOUNT-MMO are necessary to fully integrate dual agonists into cardiovascular guidelines for non-diabetic individuals.
7. GLP-1 and GIP Influence on Bone Health and Remodeling
GLP-1 and GIP play important roles in skeletal homeostasis and bone remodeling. Evidence from GLP-1R and GIPR knockout mouse models demonstrates deterioration of bone microarchitecture and increased skeletal fragility, indicating that both incretin pathways contribute to maintaining bone health [2].
7.1. Bone Resorption and Formation
Both GLP-1 and GIP support bone formation and suppress bone resorption [16,79]. GIP appears to exert the more potent anti-resorptive effect, accounting for approximately 22–25% of the postprandial reduction in bone resorption in healthy adults [79,80]. Mechanistically, GIP acts directly on human osteoclasts to inhibit bone resorption, reduce osteoclast survival, and delay actin ring formation [81]. GIP also acutely stimulates bone formation and rapidly uncouples bone resorption from formation [80].
GLP-1RAs, similarly, promote bone formation through several intracellular mechanisms. They enhance osteogenic differentiation of bone marrow stromal cells (BMSCs) via the PKA/β-catenin signaling pathway [82,83], and may also activate cAMP/PKA and MAPK signaling in mature osteoblasts, further contributing to anabolic bone effects [82].
7.2. Bone Material Properties
Beyond cellular activity, GIP influences the intrinsic material properties of bone. Experimental studies show that GIP improves collagen maturity and reduces collagen fibril diameter in osteoblast cultures [82]. Novel multi-agonist approaches also support bone quality: for example, the dual GIP/GLP-2 analog GL-0001 enhances bone tissue material properties and increases biomechanical strength in preclinical studies [84].
7.3. Fracture Risk and Musculoskeletal Disorders
GLP-1RAs are considered safe with respect to fracture risk and do not increase skeletal events in patients with T2DM [85,86]. Some meta-analyses report significant reductions in fracture incidence with GLP-1RA therapy compared with other antidiabetic treatments over 52 weeks [85,87,88]. Clinically, low circulating GLP-1 levels correlate with osteoporosis, suggesting a physiological link between GLP-1 deficiency and impaired bone health [85].
GLP-1RAs also show promise in musculoskeletal disorders beyond osteoporosis. Their weight-reducing, anti-inflammatory, and immunomodulatory effects support potential therapeutic roles in osteoarthritis (OA) [89].
7.4. Effect of GIP on Sarcopenia
However, GIP has non-negligible side effects on muscle cells, and GIPR antagonism could be used as a new treatment for sarcopenia. In a study of GIP receptor-knockout mice, researchers discovered that GIP drives the differentiation of fibroadipogenic muscle precursors into adipocytes, which is thought to be the pathological cause of sarcopenia. GIPR antagonism functions to reduce intramuscular adipose tissue and regenerate new muscle [90]. This research revealed that GLP-1/GIP co-agonists, such as tirzepatide, can have side effects such as causing sarcopenia. For the drug development industry, developing drugs to prevent muscle wasting and collaborating with the successful obesity management drug GLP-1/GIP agonists is very promising [91].
Conversely, the effects of GIP on muscle tissue are complex. GIP may promote adipogenic differentiation of muscle precursor cells, contributing to sarcopenia; thus, GIPR antagonism has been proposed as a potential therapeutic approach for sarcopenia management [90].
8. Conclusions and Future Directions
Insights into incretin physiology have shifted the therapeutic landscape from single-receptor approaches to multi-agonist strategies that integrate metabolic and cardiovascular regulation. While GLP-1 has proven to be a critical pharmacological tool, with its actions on gastric emptying, appetite suppression, and a preserved insulinotropic effect in T2DM forming the basis of a highly successful class of therapeutics, GIP stands as the dominant physiological incretin in healthy individuals. The success of GIP/GLP-1 receptor co-agonists has reconciled this dichotomy, confirming that harnessing the synergy between these two pathways can unlock a new therapeutic frontier with metabolic benefits that surpass what can be achieved by targeting GLP-1 alone.
Author Contributions
Conceptualization, D.Y.; methodology, D.Y.; validation, D.Y.; formal analysis, D.Y.; investigation, D.Y.; resources, D.Y.; data curation, D.Y.; writing—original draft preparation, D.Y.; writing—review and editing, D.Y.; visualization, D.Y.; supervision, D.Y.; project administration, D.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable. This study did not involve human subjects or identifiable patient data.
Data Availability Statement
No new datasets were generated or analyzed during the current study. All data discussed are from previously published sources cited within the manuscript.
Acknowledgments
During the preparation of this manuscript, the author used ChatGPT (OpenAI, GPT-5.1) for assistance with language refinement, and formatting. The author reviewed and edited all generated content and takes full responsibility for the final version of this publication.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Abbreviation | Full Term |
| ACAT-1 | Acyl-CoA:cholesterol acyltransferase-1 |
| ApoE−/− | Apolipoprotein E knockout mouse |
| ASCVD | Atherosclerotic Cardiovascular Disease |
| ARR | Absolute risk reduction |
| BMI | Body mass index |
| BMSC | Bone marrow stromal cell |
| CABG | Coronary artery bypass grafting |
| cAMP | Cyclic adenosine monophosphate |
| CVD | Cardiovascular disease |
| CVOT | Cardiovascular outcome trial |
| DPP-IV | Dipeptidyl peptidase-IV |
| eNOS | Endothelial nitric oxide synthase |
| FDA | Food and Drug Administration |
| GIP | Glucose-dependent insulinotropic polypeptide |
| GIPR | Glucose-dependent insulinotropic polypeptide receptor |
| GLP-1 | Glucagon-like peptide-1 |
| GLP-1RA(s) | GLP-1 receptor agonist(s) |
| HDL | High-density lipoprotein cholesterol |
| HFpEF | Heart failure with preserved ejection fraction |
| IRA(s) | Incretin receptor agonist(s) |
| LDL | Low-density lipoprotein |
| MACE | Major adverse cardiovascular events |
| MAPK | Mitogen-activated protein kinase |
| MI | Myocardial infarction |
| MMP(s) | Matrix metalloproteinase(s) |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NO | Nitric oxide |
| NT-proBNP | N-terminal pro–B-type natriuretic peptide |
| OA | Osteoarthritis |
| ox-LDL | Oxidized low-density lipoprotein |
| PKA | Protein kinase A |
| SBP | Systolic blood pressure |
| T2DM | Type 2 diabetes mellitus |
| TG | Triglycerides |
| TNF-α | Tumor necrosis factor-alpha |
| VLDL | Very low-density lipoprotein |
| VSMC(s) | Vascular smooth muscle cell(s) |
References
- Nauck, M. A.; Quast, D. R.; Wefers, J.; Pfeiffer, A. F. H. The evolving story of incretins (GIP and GLP-1) in metabolic and cardiovascular disease: A pathophysiological update. Diabetes Obes Metab 2021, 23 Suppl 3, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Li, G.; Liu, P.; Ding, P.; Gao, Y. GLP-1 and GIP: Magic bullet for musculoskeletal diseases? J Adv Res 2025. [Google Scholar] [CrossRef]
- Nauck, M. A.; Heimesaat, M. M.; Orskov, C.; Holst, J. J.; Ebert, R.; Creutzfeldt, W. Preserved incretin activity of glucagon-like peptide 1 [7-36 amide] but not of synthetic human gastric inhibitory polypeptide in patients with type-2 diabetes mellitus. J Clin Invest 1993, 91, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M. A.; Kleine, N.; Orskov, C.; Holst, J. J.; Willms, B.; Creutzfeldt, W. Normalization of fasting hyperglycaemia by exogenous glucagon-like peptide 1 (7-36 amide) in type 2 (non-insulin-dependent) diabetic patients. Diabetologia 1993, 36, 741–744. [Google Scholar] [CrossRef]
- Flint, A.; Raben, A.; Astrup, A.; Holst, J. J. Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. J Clin Invest 1998, 101, 515–520. [Google Scholar] [CrossRef]
- Bastin, M.; Andreelli, F. Dual GIP-GLP1-Receptor Agonists In The Treatment Of Type 2 Diabetes: A Short Review On Emerging Data And Therapeutic Potential. Diabetes Metab Syndr Obes 2019, 12, 1973–1985. [Google Scholar] [CrossRef] [PubMed]
- Garvey, W. T.; Frias, J. P.; Jastreboff, A. M.; le Roux, C. W.; Sattar, N.; Aizenberg, D.; Mao, H.; Zhang, S.; Ahmad, N. N.; Bunck, M. C.; Benabbad, I.; Zhang, X. M.; investigators, S.-. Tirzepatide once weekly for the treatment of obesity in people with type 2 diabetes (SURMOUNT-2): a double-blind, randomised, multicentre, placebo-controlled, phase 3 trial. Lancet 2023, 402, 613–626. [Google Scholar] [CrossRef]
- Tall Bull, S.; Nuffer, W.; Trujillo, J. M. Tirzepatide: A novel, first-in-class, dual GIP/GLP-1 receptor agonist. J Diabetes Complications 2022, 36, 108332. [Google Scholar] [CrossRef]
- Seino, Y.; Maekawa, R.; Ogata, H.; Hayashi, Y. Carbohydrate-induced secretion of glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1. J Diabetes Investig 2016, 7 (Suppl 1), 27–32. [Google Scholar] [CrossRef]
- Liu, Q. K. Mechanisms of action and therapeutic applications of GLP-1 and dual GIP/GLP-1 receptor agonists. Front Endocrinol (Lausanne) 2024, 15, 1431292. [Google Scholar] [CrossRef]
- Buchan, A. M.; Polak, J. M.; Capella, C.; Solcia, E.; Pearse, A. G. Electronimmunocytochemical evidence for the K cell localization of gastric inhibitory polypeptide (GIP) in man. Histochemistry 1978, 56, 37–44. [Google Scholar] [CrossRef]
- Buffa, R.; Polak, J. M.; Pearse, A. G.; Solcia, E.; Grimelius, L.; Capella, C. Identification of the intestinal cell storing gastric inhibitory peptide. Histochemistry 1975, 43, 249–255. [Google Scholar] [CrossRef]
- Jorsal, T.; Rhee, N. A.; Pedersen, J.; Wahlgren, C. D.; Mortensen, B.; Jepsen, S. L.; Jelsing, J.; Dalboge, L. S.; Vilmann, P.; Hassan, H.; Hendel, J. W.; Poulsen, S. S.; Holst, J. J.; Vilsboll, T.; Knop, F. K. Enteroendocrine K and L cells in healthy and type 2 diabetic individuals. Diabetologia 2018, 61, 284–294. [Google Scholar] [CrossRef]
- Adriaenssens, A. E.; Reimann, F.; Gribble, F. M. Distribution and Stimulus Secretion Coupling of Enteroendocrine Cells along the Intestinal Tract. Compr Physiol 2018, 8, 1603–1638. [Google Scholar] [CrossRef]
- Manandhar, B.; Ahn, J. M. Glucagon-like peptide-1 (GLP-1) analogs: recent advances, new possibilities, and therapeutic implications. J Med Chem 2015, 58, 1020–1037. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Qi, X.; Fan, G.; Zhou, L.; Peng, Z.; Yang, J. Anti-atherosclerotic effect of incretin receptor agonists. Front Endocrinol (Lausanne) 2024, 15, 1463547. [Google Scholar]
- Ahren, B.; Yamada, Y.; Seino, Y. The Incretin Effect in Female Mice With Double Deletion of GLP-1 and GIP Receptors. J Endocr Soc 2020, 4, bvz036. [Google Scholar] [PubMed]
- Pederson, R. A.; Brown, J. C. Interaction of gastric inhibitory polypeptide, glucose, and arginine on insulin and glucagon secretion from the perfused rat pancreas. Endocrinology 1978, 103, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Pederson, R. A.; Brown, J. C. The insulinotropic action of gastric inhibitory polypeptide in the perfused isolated rat pancreas. Endocrinology 1976, 99, 780–785. [Google Scholar] [CrossRef]
- Christensen, M.; Vedtofte, L.; Holst, J. J.; Vilsboll, T.; Knop, F. K. Glucose-dependent insulinotropic polypeptide: a bifunctional glucose-dependent regulator of glucagon and insulin secretion in humans. Diabetes 2011, 60, 3103–3109. [Google Scholar]
- Gasbjerg, L. S.; Bergmann, N. C.; Stensen, S.; Christensen, M. B.; Rosenkilde, M. M.; Holst, J. J.; Nauck, M.; Knop, F. K. Evaluation of the incretin effect in humans using GIP and GLP-1 receptor antagonists. Peptides 2020, 125, 170183. [Google Scholar] [CrossRef]
- Wettergren, A.; Schjoldager, B.; Mortensen, P. E.; Myhre, J.; Christiansen, J.; Holst, J. J. Truncated GLP-1 (proglucagon 78-107-amide) inhibits gastric and pancreatic functions in man. Dig Dis Sci 1993, 38, 665–673. [Google Scholar] [CrossRef]
- Nauck, M. A.; Niedereichholz, U.; Ettler, R.; Holst, J. J.; Orskov, C.; Ritzel, R.; Schmiegel, W. H. Glucagon-like peptide 1 inhibition of gastric emptying outweighs its insulinotropic effects in healthy humans. Am J Physiol 1997, 273, E981–E988. [Google Scholar] [CrossRef]
- Meier, J. J.; Goetze, O.; Anstipp, J.; Hagemann, D.; Holst, J. J.; Schmidt, W. E.; Gallwitz, B.; Nauck, M. A. Gastric inhibitory polypeptide does not inhibit gastric emptying in humans. Am J Physiol Endocrinol Metab 2004, 286, E621–E625. [Google Scholar] [CrossRef]
- Bergmann, N. C.; Lund, A.; Gasbjerg, L. S.; Meessen, E. C. E.; Andersen, M. M.; Bergmann, S.; Hartmann, B.; Holst, J. J.; Jessen, L.; Christensen, M. B.; Vilsboll, T.; Knop, F. K. Effects of combined GIP and GLP-1 infusion on energy intake, appetite and energy expenditure in overweight/obese individuals: a randomised, crossover study. Diabetologia 2019, 62, 665–675. [Google Scholar] [CrossRef]
- Nauck, M.; Stockmann, F.; Ebert, R.; Creutzfeldt, W. Reduced incretin effect in type 2 (non-insulin-dependent) diabetes. Diabetologia 1986, 29, 46–52. [Google Scholar] [CrossRef]
- Krarup, T.; Saurbrey, N.; Moody, A. J.; Kuhl, C.; Madsbad, S. Effect of porcine gastric inhibitory polypeptide on beta-cell function in type I and type II diabetes mellitus. Metabolism 1987, 36, 677–682. [Google Scholar] [CrossRef]
- Mentis, N.; Vardarli, I.; Kothe, L. D.; Holst, J. J.; Deacon, C. F.; Theodorakis, M.; Meier, J. J.; Nauck, M. A. GIP does not potentiate the antidiabetic effects of GLP-1 in hyperglycemic patients with type 2 diabetes. Diabetes 2011, 60, 1270–1276. [Google Scholar] [CrossRef]
- Getty-Kaushik, L.; Song, D. H.; Boylan, M. O.; Corkey, B. E.; Wolfe, M. M. Glucose-dependent insulinotropic polypeptide modulates adipocyte lipolysis and reesterification. Obesity (Silver Spring) 2006, 14, 1124–1131. [Google Scholar] [CrossRef]
- Eckel, R. H.; Fujimoto, W. Y.; Brunzell, J. D. Gastric inhibitory polypeptide enhanced lipoprotein lipase activity in cultured preadipocytes. Diabetes 1979, 28, 1141–1142. [Google Scholar] [CrossRef]
- Song, D. H.; Getty-Kaushik, L.; Tseng, E.; Simon, J.; Corkey, B. E.; Wolfe, M. M. Glucose-dependent insulinotropic polypeptide enhances adipocyte development and glucose uptake in part through Akt activation. Gastroenterology 2007, 133, 1796–1805. [Google Scholar] [CrossRef]
- Kim, S. J.; Nian, C.; McIntosh, C. H. Activation of lipoprotein lipase by glucose-dependent insulinotropic polypeptide in adipocytes. A role for a protein kinase B, LKB1, and AMP-activated protein kinase cascade. J Biol Chem 2007, 282, 8557–8567. [Google Scholar] [CrossRef]
- Ahlqvist, E.; Osmark, P.; Kuulasmaa, T.; Pilgaard, K.; Omar, B.; Brons, C.; Kotova, O.; Zetterqvist, A. V.; Stancakova, A.; Jonsson, A.; Hansson, O.; Kuusisto, J.; Kieffer, T. J.; Tuomi, T.; Isomaa, B.; Madsbad, S.; Gomez, M. F.; Poulsen, P.; Laakso, M.; Degerman, E.; Pihlajamaki, J.; Wierup, N.; Vaag, A.; Groop, L.; Lyssenko, V. Link between GIP and osteopontin in adipose tissue and insulin resistance. Diabetes 2013, 62, 2088–2094. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, A. F. H.; Keyhani-Nejad, F. High Glycemic Index Metabolic Damage - a Pivotal Role of GIP and GLP-1. Trends Endocrinol Metab 2018, 29, 289–299. [Google Scholar] [CrossRef]
- Sun, F.; Chai, S.; Li, L.; Yu, K.; Yang, Z.; Wu, S.; Zhang, Y.; Ji, L.; Zhan, S. Effects of glucagon-like peptide-1 receptor agonists on weight loss in patients with type 2 diabetes: a systematic review and network meta-analysis. J Diabetes Res 2015, 2015, 157201. [Google Scholar] [CrossRef]
- Marso, S. P.; Daniels, G. H.; Brown-Frandsen, K.; Kristensen, P.; Mann, J. F.; Nauck, M. A.; Nissen, S. E.; Pocock, S.; Poulter, N. R.; Ravn, L. S.; Steinberg, W. M.; Stockner, M.; Zinman, B.; Bergenstal, R. M.; Buse, J. B.; Committee, L. S.; Investigators, L. T. Liraglutide and Cardiovascular Outcomes in Type 2 Diabetes. N Engl J Med 2016, 375, 311–322. [Google Scholar] [CrossRef]
- Marso, S. P.; Bain, S. C.; Consoli, A.; Eliaschewitz, F. G.; Jodar, E.; Leiter, L. A.; Lingvay, I.; Rosenstock, J.; Seufert, J.; Warren, M. L.; Woo, V.; Hansen, O.; Holst, A. G.; Pettersson, J.; Vilsboll, T.; Investigators, S.-. Semaglutide and Cardiovascular Outcomes in Patients with Type 2 Diabetes. N Engl J Med 2016, 375, 1834–1844. [Google Scholar] [CrossRef]
- Gerstein, H. C.; Colhoun, H. M.; Dagenais, G. R.; Diaz, R.; Lakshmanan, M.; Pais, P.; Probstfield, J.; Riesmeyer, J. S.; Riddle, M. C.; Ryden, L.; Xavier, D.; Atisso, C. M.; Dyal, L.; Hall, S.; Rao-Melacini, P.; Wong, G.; Avezum, A.; Basile, J.; Chung, N.; Conget, I.; Cushman, W. C.; Franek, E.; Hancu, N.; Hanefeld, M.; Holt, S.; Jansky, P.; Keltai, M.; Lanas, F.; Leiter, L. A.; Lopez-Jaramillo, P.; Cardona Munoz, E. G.; Pirags, V.; Pogosova, N.; Raubenheimer, P. J.; Shaw, J. E.; Sheu, W. H.; Temelkova-Kurktschiev, T.; Investigators, R. Dulaglutide and cardiovascular outcomes in type 2 diabetes (REWIND): a double-blind, randomised placebo-controlled trial. Lancet 2019, 394, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M. A.; D'Alessio, D. A. Tirzepatide, a dual GIP/GLP-1 receptor co-agonist for the treatment of type 2 diabetes with unmatched effectiveness regrading glycaemic control and body weight reduction. Cardiovasc Diabetol 2022, 21, 169. [Google Scholar] [CrossRef] [PubMed]
- Meier, J. J.; Gethmann, A.; Gotze, O.; Gallwitz, B.; Holst, J. J.; Schmidt, W. E.; Nauck, M. A. Glucagon-like peptide 1 abolishes the postprandial rise in triglyceride concentrations and lowers levels of non-esterified fatty acids in humans. Diabetologia 2006, 49, 452–458. [Google Scholar] [CrossRef]
- Dahl, D.; Onishi, Y.; Norwood, P.; Huh, R.; Bray, R.; Patel, H.; Rodriguez, A. Effect of Subcutaneous Tirzepatide vs Placebo Added to Titrated Insulin Glargine on Glycemic Control in Patients With Type 2 Diabetes: The SURPASS-5 Randomized Clinical Trial. JAMA 2022, 327, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Wu, S.; Guo, S.; Yu, K.; Yang, Z.; Li, L.; Zhang, Y.; Quan, X.; Ji, L.; Zhan, S. Impact of GLP-1 receptor agonists on blood pressure, heart rate and hypertension among patients with type 2 diabetes: A systematic review and network meta-analysis. Diabetes Res Clin Pract 2015, 110, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Moreno, C.; Mistry, M.; Roman, R. J. Renal effects of glucagon-like peptide in rats. Eur J Pharmacol 2002, 434, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Skov, J.; Dejgaard, A.; Frokiaer, J.; Holst, J. J.; Jonassen, T.; Rittig, S.; Christiansen, J. S. Glucagon-like peptide-1 (GLP-1): effect on kidney hemodynamics and renin-angiotensin-aldosterone system in healthy men. J Clin Endocrinol Metab 2013, 98, E664–E671. [Google Scholar] [CrossRef]
- Jensen, E. P.; Moller, S.; Hviid, A. V.; Veedfald, S.; Holst, J. J.; Pedersen, J.; Orskov, C.; Sorensen, C. M. GLP-1-induced renal vasodilation in rodents depends exclusively on the known GLP-1 receptor and is lost in prehypertensive rats. Am J Physiol Renal Physiol 2020, 318, F1409–F1417. [Google Scholar] [CrossRef]
- Helmstadter, J.; Frenis, K.; Filippou, K.; Grill, A.; Dib, M.; Kalinovic, S.; Pawelke, F.; Kus, K.; Kroller-Schon, S.; Oelze, M.; Chlopicki, S.; Schuppan, D.; Wenzel, P.; Ruf, W.; Drucker, D. J.; Munzel, T.; Daiber, A.; Steven, S. Endothelial GLP-1 (Glucagon-Like Peptide-1) Receptor Mediates Cardiovascular Protection by Liraglutide In Mice With Experimental Arterial Hypertension. Arterioscler Thromb Vasc Biol 2020, 40, 145–158. [Google Scholar] [CrossRef]
- Le, Y.; Wei, R.; Yang, K.; Lang, S.; Gu, L.; Liu, J.; Hong, T.; Yang, J. Liraglutide ameliorates palmitate-induced oxidative injury in islet microvascular endothelial cells through GLP-1 receptor/PKA and GTPCH1/eNOS signaling pathways. Peptides 2020, 124, 170212. [Google Scholar] [CrossRef]
- Zhong, Q.; Bollag, R. J.; Dransfield, D. T.; Gasalla-Herraiz, J.; Ding, K. H.; Min, L.; Isales, C. M. Glucose-dependent insulinotropic peptide signaling pathways in endothelial cells. Peptides 2000, 21, 1427–1432. [Google Scholar] [CrossRef]
- Dai, Y.; Mehta, J. L.; Chen, M. Glucagon-like peptide-1 receptor agonist liraglutide inhibits endothelin-1 in endothelial cell by repressing nuclear factor-kappa B activation. Cardiovasc Drugs Ther 2013, 27, 371–380. [Google Scholar] [CrossRef]
- Gaspari, T.; Liu, H.; Welungoda, I.; Hu, Y.; Widdop, R. E.; Knudsen, L. B.; Simpson, R. W.; Dear, A. E. A GLP-1 receptor agonist liraglutide inhibits endothelial cell dysfunction and vascular adhesion molecule expression in an ApoE-/- mouse model. Diab Vasc Dis Res 2011, 8, 117–124. [Google Scholar] [CrossRef]
- Tang, S. T.; Tang, H. Q.; Su, H.; Wang, Y.; Zhou, Q.; Zhang, Q.; Wang, Y.; Zhu, H. Q. Glucagon-like peptide-1 attenuates endothelial barrier injury in diabetes via cAMP/PKA mediated down-regulation of MLC phosphorylation. Biomed Pharmacother 2019, 113, 108667. [Google Scholar] [CrossRef]
- Cai, X.; She, M.; Xu, M.; Chen, H.; Li, J.; Chen, X.; Zheng, D.; Liu, J.; Chen, S.; Zhu, J.; Xu, X.; Li, R.; Li, J.; Chen, S.; Yang, X.; Li, H. GLP-1 treatment protects endothelial cells from oxidative stress-induced autophagy and endothelial dysfunction. Int J Biol Sci 2018, 14, 1696–1708. [Google Scholar] [CrossRef]
- Zhan, Y.; Sun, H. L.; Chen, H.; Zhang, H.; Sun, J.; Zhang, Z.; Cai, D. H. Glucagon-like peptide-1 (GLP-1) protects vascular endothelial cells against advanced glycation end products (AGEs)-induced apoptosis. Med Sci Monit 2012, 18, BR286–BR291. [Google Scholar] [CrossRef]
- Tashiro, Y.; Sato, K.; Watanabe, T.; Nohtomi, K.; Terasaki, M.; Nagashima, M.; Hirano, T. A glucagon-like peptide-1 analog liraglutide suppresses macrophage foam cell formation and atherosclerosis. Peptides 2014, 54, 19–26. [Google Scholar] [CrossRef]
- Dai, Y.; Dai, D.; Wang, X.; Ding, Z.; Li, C.; Mehta, J. L. GLP-1 agonists inhibit ox-LDL uptake in macrophages by activating protein kinase A. J Cardiovasc Pharmacol 2014, 64, 47–52. [Google Scholar] [CrossRef]
- Nagashima, M.; Watanabe, T.; Terasaki, M.; Tomoyasu, M.; Nohtomi, K.; Kim-Kaneyama, J.; Miyazaki, A.; Hirano, T. Native incretins prevent the development of atherosclerotic lesions in apolipoprotein E knockout mice. Diabetologia 2011, 54, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Yashima, H.; Mori, Y.; Saito, T.; Shiraga, Y.; Kawakami, R.; Ohara, M.; Fukui, T.; Hirano, T.; Yamada, Y.; Seino, Y.; Yamagishi, S. I. Glucose-Dependent Insulinotropic Polypeptide Suppresses Foam Cell Formation of Macrophages through Inhibition of the Cyclin-Dependent Kinase 5-CD36 Pathway. Biomedicines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Mei, A.; Liu, X.; Braunstein, Z.; Wei, Y.; Wang, B.; Duan, L.; Rao, X.; Rajagopalan, S.; Dong, L.; Zhong, J. Glucagon-Like Peptide-1 Receptor Regulates Macrophage Migration in Monosodium Urate-Induced Peritoneal Inflammation. Front Immunol 2022, 13, 772446. [Google Scholar] [CrossRef] [PubMed]
- Vinue, A.; Navarro, J.; Herrero-Cervera, A.; Garcia-Cubas, M.; Andres-Blasco, I.; Martinez-Hervas, S.; Real, J. T.; Ascaso, J. F.; Gonzalez-Navarro, H. The GLP-1 analogue lixisenatide decreases atherosclerosis in insulin-resistant mice by modulating macrophage phenotype. Diabetologia 2017, 60, 1801–1812. [Google Scholar] [CrossRef]
- Bruen, R.; Curley, S.; Kajani, S.; Crean, D.; O'Reilly, M. E.; Lucitt, M. B.; Godson, C. G.; McGillicuddy, F. C.; Belton, O. Liraglutide dictates macrophage phenotype in apolipoprotein E null mice during early atherosclerosis. Cardiovasc Diabetol 2017, 16, 143. [Google Scholar] [CrossRef]
- Shi, L.; Ji, Y.; Jiang, X.; Zhou, L.; Xu, Y.; Li, Y.; Jiang, W.; Meng, P.; Liu, X. Liraglutide attenuates high glucose-induced abnormal cell migration, proliferation, and apoptosis of vascular smooth muscle cells by activating the GLP-1 receptor, and inhibiting ERK1/2 and PI3K/Akt signaling pathways. Cardiovasc Diabetol 2015, 14, 18. [Google Scholar] [CrossRef]
- Torres, G.; Morales, P. E.; Garcia-Miguel, M.; Norambuena-Soto, I.; Cartes-Saavedra, B.; Vidal-Pena, G.; Moncada-Ruff, D.; Sanhueza-Olivares, F.; San Martin, A.; Chiong, M. Glucagon-like peptide-1 inhibits vascular smooth muscle cell dedifferentiation through mitochondrial dynamics regulation. Biochem Pharmacol 2016, 104, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Burgmaier, M.; Liberman, A.; Mollmann, J.; Kahles, F.; Reith, S.; Lebherz, C.; Marx, N.; Lehrke, M. Glucagon-like peptide-1 (GLP-1) and its split products GLP-1(9-37) and GLP-1(28-37) stabilize atherosclerotic lesions in apoe(-)/(-) mice. Atherosclerosis 2013, 231, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Lei, Y.; Inoue, A.; Piao, L.; Hu, L.; Jiang, H.; Sasaki, T.; Wu, H.; Xu, W.; Yu, C.; Zhao, G.; Ogasawara, S.; Okumura, K.; Kuzuya, M.; Cheng, X. W. Exenatide mitigated diet-induced vascular aging and atherosclerotic plaque growth in ApoE-deficient mice under chronic stress. Atherosclerosis 2017, 264, 1–10. [Google Scholar] [CrossRef]
- Lim, D. M.; Park, K. Y.; Hwang, W. M.; Kim, J. Y.; Kim, B. J. Difference in protective effects of GIP and GLP-1 on endothelial cells according to cyclic adenosine monophosphate response. Exp Ther Med 2017, 13, 2558–2564. [Google Scholar] [CrossRef] [PubMed]
- Yang, H. M. GLP-1 Agonists in Cardiovascular Diseases: Mechanisms, Clinical Evidence, and Emerging Therapies. J Clin Med 2025, 14. [Google Scholar] [CrossRef]
- Cho, Y. K.; La Lee, Y.; Jung, C. H. The Cardiovascular Effect of Tirzepatide: A Glucagon-Like Peptide-1 and Glucose-Dependent Insulinotropic Polypeptide Dual Agonist. J Lipid Atheroscler 2023, 12, 213–222. [Google Scholar] [CrossRef]
- Holst, J. J. On the physiology of GIP and GLP-1. Horm Metab Res 2004, 36, 747–754. [Google Scholar] [CrossRef]
- Knop, F. K.; Vilsboll, T.; Larsen, S.; Hojberg, P. V.; Volund, A.; Madsbad, S.; Holst, J. J.; Krarup, T. Increased postprandial responses of GLP-1 and GIP in patients with chronic pancreatitis and steatorrhea following pancreatic enzyme substitution. Am J Physiol Endocrinol Metab 2007, 292, E324–E330. [Google Scholar] [CrossRef]
- Owens, D. R.; Monnier, L.; Hanefeld, M. A review of glucagon-like peptide-1 receptor agonists and their effects on lowering postprandial plasma glucose and cardiovascular outcomes in the treatment of type 2 diabetes mellitus. Diabetes Obes Metab 2017, 19, 1645–1654. [Google Scholar] [CrossRef]
- Frias, J. P.; Nauck, M. A.; Van, J.; Kutner, M. E.; Cui, X.; Benson, C.; Urva, S.; Gimeno, R. E.; Milicevic, Z.; Robins, D.; Haupt, A. Efficacy and safety of LY3298176, a novel dual GIP and GLP-1 receptor agonist, in patients with type 2 diabetes: a randomised, placebo-controlled and active comparator-controlled phase 2 trial. Lancet 2018, 392, 2180–2193. [Google Scholar] [CrossRef]
- Thomas, M. K.; Nikooienejad, A.; Bray, R.; Cui, X.; Wilson, J.; Duffin, K.; Milicevic, Z.; Haupt, A.; Robins, D. A. Dual GIP and GLP-1 Receptor Agonist Tirzepatide Improves Beta-cell Function and Insulin Sensitivity in Type 2 Diabetes. J Clin Endocrinol Metab 2021, 106, 388–396. [Google Scholar] [CrossRef]
- Verma, S.; Colhoun, H. M.; Dicker, D.; Hovingh, G. K.; Kahn, S. E.; Kautzky-Willer, A.; Lingvay, I.; Plutzky, J.; Rasmussen, S.; Rathor, N.; Hoff, S. T.; Lincoff, A. M. Semaglutide Effects on Cardiovascular Outcomes in People With Overweight or Obesity (SELECT): Outcomes by Sex. J Am Coll Cardiol 2024, 84, 1678–1682. [Google Scholar] [CrossRef] [PubMed]
- Odigwe, C.; Mulyala, R.; Malik, H.; Ruiz, B.; Riad, M.; Sayiadeh, M. A.; Honganur, S.; Parks, A.; Rahman, M. U.; Lakkis, N. Emerging role of GLP-1 agonists in cardio-metabolic therapy - Focus on Semaglutide. Am Heart J Plus 2025, 52, 100518. [Google Scholar] [CrossRef]
- Turkistani, Y. Glucagon-like peptide-1 receptor agonists: a review from a cardiovascular perspective. Front Cardiovasc Med 2025, 12, 1535134. [Google Scholar] [CrossRef] [PubMed]
- Hosseinpour, A.; Sood, A.; Kamalpour, J.; Zandi, E.; Pakmehr, S.; Hosseinpour, H.; Sood, A.; Agrawal, A.; Gupta, R. Glucagon-Like Peptide-1 Receptor Agonists and Major Adverse Cardiovascular Events in Patients With and Without Diabetes: A Meta-Analysis of Randomized-Controlled Trials. Clin Cardiol 2024, 47, e24314. [Google Scholar] [CrossRef]
- Kelkar, R.; Barve, N. A.; Kelkar, R.; Kharel, S.; Khanapurkar, S.; Yadav, R. Comparison of glucagon-like peptide-1 receptor agonists vs. placebo on any cardiovascular events in overweight or obese non-diabetic patients: a systematic review and meta-analysis. Front Cardiovasc Med 2024, 11, 1453297. [Google Scholar] [CrossRef]
- Lam, C. S. P.; Rodriguez, A.; Aminian, A.; Ferrannini, E.; Heerspink, H. J. L.; Jastreboff, A. M.; Laffin, L. J.; Pandey, A.; Ray, K. K.; Ridker, P. M.; Sanyal, A. J.; Yki-Jarvinen, H.; Mason, D.; Strzelecki, M.; Bartee, A. K.; Cui, C.; Hurt, K.; Linetzky, B.; Bunck, M. C.; Nissen, S. E. Tirzepatide for reduction of morbidity and mortality in adults with obesity: rationale and design of the SURMOUNT-MMO trial. Obesity (Silver Spring) 2025, 33, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Helsted, M. M.; Gasbjerg, L. S.; Lanng, A. R.; Bergmann, N. C.; Stensen, S.; Hartmann, B.; Christensen, M. B.; Holst, J. J.; Vilsboll, T.; Rosenkilde, M. M.; Knop, F. K. The role of endogenous GIP and GLP-1 in postprandial bone homeostasis. Bone 2020, 140, 115553. [Google Scholar] [CrossRef]
- Hansen, M. S.; Soe, K.; Christensen, L. L.; Fernandez-Guerra, P.; Hansen, N. W.; Wyatt, R. A.; Martin, C.; Hardy, R. S.; Andersen, T. L.; Olesen, J. B.; Hartmann, B.; Rosenkilde, M. M.; Kassem, M.; Rauch, A.; Gorvin, C. M.; Frost, M. GIP reduces osteoclast activity and improves osteoblast survival in primary human bone cells. Eur J Endocrinol 2023, 188. [Google Scholar] [CrossRef]
- Torekov, S. S.; Harslof, T.; Rejnmark, L.; Eiken, P.; Jensen, J. B.; Herman, A. P.; Hansen, T.; Pedersen, O.; Holst, J. J.; Langdahl, B. L. A functional amino acid substitution in the glucose-dependent insulinotropic polypeptide receptor (GIPR) gene is associated with lower bone mineral density and increased fracture risk. J Clin Endocrinol Metab 2014, 99, E729–E733. [Google Scholar] [CrossRef]
- Meng, J.; Ma, X.; Wang, N.; Jia, M.; Bi, L.; Wang, Y.; Li, M.; Zhang, H.; Xue, X.; Hou, Z.; Zhou, Y.; Yu, Z.; He, G.; Luo, X. Activation of GLP-1 Receptor Promotes Bone Marrow Stromal Cell Osteogenic Differentiation through beta-Catenin. Stem Cell Reports 2016, 6, 579–591. [Google Scholar] [CrossRef]
- Luo, G.; Liu, H.; Lu, H. Glucagon-like peptide-1(GLP-1) receptor agonists: potential to reduce fracture risk in diabetic patients? Br J Clin Pharmacol 2016, 81, 78–88. [Google Scholar] [CrossRef]
- Gobron, B.; Couchot, M.; Irwin, N.; Legrand, E.; Bouvard, B.; Mabilleau, G. Development of a First-in-Class Unimolecular Dual GIP/GLP-2 Analogue, GL-0001, for the Treatment of Bone Fragility. J Bone Miner Res 2023, 38, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Hu, Y.; Li, Y. Y.; Cao, X.; Bai, N.; Lu, T. T.; Li, G. Q.; Li, N.; Wang, A. N.; Mao, X. M. Glucagon-like peptide-1 receptor agonists and risk of bone fracture in patients with type 2 diabetes: A meta-analysis of randomized controlled trials. Diabetes Metab Res Rev 2019, 35, e3168. [Google Scholar] [CrossRef] [PubMed]
- Hidayat, K.; Du, X.; Shi, B. M. Risk of fracture with dipeptidyl peptidase-4 inhibitors, glucagon-like peptide-1 receptor agonists, or sodium-glucose cotransporter-2 inhibitors in real-world use: systematic review and meta-analysis of observational studies. Osteoporos Int 2019, 30, 1923–1940. [Google Scholar] [CrossRef]
- Bergmann, N. C.; Lund, A.; Gasbjerg, L. S.; Jorgensen, N. R.; Jessen, L.; Hartmann, B.; Holst, J. J.; Christensen, M. B.; Vilsboll, T.; Knop, F. K. Separate and Combined Effects of GIP and GLP-1 Infusions on Bone Metabolism in Overweight Men Without Diabetes. J Clin Endocrinol Metab 2019, 104, 2953–2960. [Google Scholar] [CrossRef]
- Chen, A. S.; Batsis, J. A. Treating Sarcopenic Obesity in the Era of Incretin Therapies: Perspectives and Challenges. Diabetes 2025. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.; Megyeri, S.; Nuffer, W.; Trujillo, J. M. The potential role of GLP-1 receptor agonists in osteoarthritis. Pharmacotherapy 2025, 45, 177–186. [Google Scholar] [CrossRef]
- Takahashi, Y.; Fujita, H.; Seino, Y.; Hattori, S.; Hidaka, S.; Miyakawa, T.; Suzuki, A.; Waki, H.; Yabe, D.; Seino, Y.; Yamada, Y. Gastric inhibitory polypeptide receptor antagonism suppresses intramuscular adipose tissue accumulation and ameliorates sarcopenia. J Cachexia Sarcopenia Muscle 2023, 14, 2703–2718. [Google Scholar] [CrossRef]
- Arnold, C. After obesity drugs' success, companies rush to preserve skeletal muscle. Nat Biotechnol 2024, 42, 351–353. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Distribution of GLP-1R and GIPR.
| Tissue/Organ | GLP-1 Receptor (GLP-1R) Presence & Details | GIP Receptor (GIPR) Presence & Details |
| Endocrine Pancreas | β-cells: +++ (Abundantly expressed) α-cells: -/+ (Present in a small proportion of α-cells) |
β-cells: +++ (Abundantly expressed) α-cells: ++ (Present) |
| Heart | + (Present in all four chambers, particularly the sinoatrial node) | + (Present in all four chambers) |
| Blood Vessels | + (Present, including in endothelial cells) | + (Present in endothelial cells) |
| Adipose Tissue | + (Present, primarily in vascular cells; debated on adipocytes) | ++ (Present, though unclear if on adipocytes or stromal-vascular cells) |
| Bone | -/+ (Absent in cultured osteoblasts but present in bone marrow stromal cells) | ++ (Present in osteoblasts and osteocytes) |
| Brain | ++ (Present in key areas for appetite regulation like the hypothalamus and brainstem) | + (Present in various regions including hippocampus and cortex) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



