
Submitted:
11 November 2025
Posted:
12 November 2025
You are already at the latest version
Abstract
Background: Histologic healing is increasingly recognized as a sensitive marker of disease remission in ulcerative colitis (UC). However, the dynamics of mucosal T lymphocytes and pro-inflammatory cytokines during healing remain incompletely understood.
Methods: In this observational, cross-sectional study, paired colonic biopsies from 20 adult UC patients were analyzed during active inflammation and subsequent histologic healing. Immunohistochemistry was performed for CD3, CD4, CD8, and IL-6. Lymphocyte densities were quantified in intraepithelial and lamina propria compartments, while IL-6 expression was scored semi-quantitatively. Histologic activity was assessed using the Geboes score.
Results: Intraepithelial CD4⁺ T cells significantly decreased during histologic healing (mean 6.8 → 3.75 cells/HPF, p < 0.05), whereas lamina propria CD4⁺ cells remained variably persistent, suggesting ongoing immune regulation. Intraepithelial CD8⁺ cells increased during remission, indicating a potential reparative or surveillance role. IL-6 expression markedly declined in epithelial and stromal compartments during healing, reflecting resolution of mucosal inflammation. Correlation analyses revealed enhanced coordination between CD4⁺ and CD8⁺ cells in the healing phase, consistent with immune homeostasis.
Conclusions: Histologic healing in UC involves compartment-specific shifts in T lymphocyte populations and a marked reduction of IL-6 expression, reflecting coordinated immune regulation beyond clinical remission. These findings highlight the potential of combined cellular and cytokine biomarkers to monitor mucosal healing and guide immunomodulatory therapies.
Keywords:
ulcerative colitis
; histologic healing
; CD4 T cells
; CD8 T cells
; IL-6
; mucosal immunity
1. Introduction
Inflammatory bowel diseases (IBD) including ulcerative colitis (UC) and Crohn’s disease, represent a global problem with a significant impact on the quality of life of affected patients, characterized by continuous mucosal inflammation of the colon [1].It is expected that by 2030 the incidence and prevalence of IBD will significantly increase [2]. Traditionally, clinical and endoscopic remission have been the primary therapeutic goals in UC management. However, accumulating evidence suggests that histologic healing represents a deeper and more durable state of remission, associated with a lower risk of relapse, hospitalization, colorectal neoplasia, and colectomy [3]. Consequently, histologic healing has emerged as an important endpoint in both clinical trials and daily practice.
Despite its growing relevance, the immunological mechanisms underpinning histologic healing remain incompletely understood. The intestinal mucosa is a highly dynamic immune environment, in which T lymphocytes play pivotal roles in orchestrating inflammation and repair. Among them, CD4+ helper T cells and CD8+ cytotoxic T cells contribute to both tissue injury and immune regulation [4]. In the active phase of UC, CD4+ T cells promote cytokine release and therecruitment of inflammatory cells [5], whereas CD8+ T cells may mediate epithelial cytotoxicity or, alternatively, support mucosal barrier restoration during resolution [6]. Understanding the spatial distribution and functional balance of these T-cell subsets across the epithelial and lamina propria compartments is essential for understanding immune regulation during mucosal healing [7].
Interleukin-6 (IL-6) is another critical player in IBD pathogenesis [8]. As a multifunctional pro-inflammatory cytokine [9], IL-6 promotes T-cell activation, B-cell differentiation, and epithelial proliferation [10,11]. Elevated IL-6 expression has been documented in active CD mucosa, correlating with disease severity and systemic inflammation [12]. Conversely, downregulation of IL-6 may reflect mucosal recovery and immune quiescence [13]. However, the compartmental expression of IL-6 across different cellular and structural contexts (epithelium, stroma and endothelium) during histologic healing has not been systematically characterized.
The mucosal immune landscape is further shaped by interactions among epithelial cells interconnected through tight junctions [14], Paneth-like cells, goblet cells and the cytokine milieu. Dysregulation at this level represents a central mechanism in the pathogenesis of IBD. Inflammation arises from genetic variants associated with the disease, such as NOD2 and ATG16L1 mutations that impair Paneth cell function [15,16], as well as from defects in tight junctions, which lead to increased intestinal permeability and bacterial translocation into the lamina propria [17] and the deficiency of MUC2 mucin secreted by goblet cells which change the composition of the mucus [18],resulting in the impairment of the mucosal protective barrier [19,20]. Epithelial restitution after inflammation involves a shift from immune activation toward resolution, marked by changes inthe lymphocyte composition and cytokine signaling. Characterizing these transitions offers insights into the persistence of subclinical immune activity that may predisposeone to relapse even after apparent histologic healing.
Therefore, the aim of this study was to evaluate the dynamics of CD4+ and CD8+ lymphocytes and IL-6 expression in colonic biopsies from UC patients during active inflammation and histologic healing. By analyzing matched biopsy sets from the same individuals at both disease stages, we sought to elucidate how mucosal immune cell distribution and cytokine expression evolve during healing, and to identify potential immunologic signatures of sustained remission.
1.1. Cellular and Molecular Mechanism
1.1.1. Local Immunity: Macrophages, Mast Cells, and Eosinophils
The intestinal mucosal immune system maintains a delicate balance between tolerance and inflammation. Resident immune cells within the lamina propria continuously interact with the epithelium and microbiota, and disruption of these interactions contributes to the chronic inflammation characteristic of IBD [21]. Intestinal macrophages exhibit functional plasticity, oscillating between the proinflammatory M1 and the reparative M2 phenotypes [22]. In IBD, predominant M1 activation leads to increased secretion of IL-1β, TNF-α, and IL-6, amplifying inflammatory infiltration and fibrogenesis [23]. Mast cells, abundant in the intestinal mucosa, interact with the enteric nervous system and release histamine, tryptase, and TNF-α, thereby increasing epithelial permeability and modulating neuroimmune responses [24]. Eosinophils infiltrate the mucosa even during clinical remission, releasing cytotoxic granule proteins such as Major Basic Protein (MBP) and Eosinophil Cationic Protein (ECP), which can damage epithelial cells and sustain inflammation [25,26]. Altogether, macrophages, mast cells, and eosinophils not only reflect the inflammatory status but actively participate in the disease progression and chronicity.
1.1.2. Cell–Cell Interactions and the Role of the Mesenchymal Compartment
Beyond epithelial and immune cells, the intestinal mesenchymal compartment plays a pivotal role in the pathogenesis of IBD through dynamic interactions with the inflammatory microenvironment [27]. Fibroblasts and myofibroblasts act as major effectors of tissue remodeling. Under the influence of proinflammatory mediators (TNF-α, TGF-β, IL-17), they proliferate and secrete extracellular matrix (ECM) components, promoting fibrosis and stricture formation, particularly in Crohn’s disease [28,29]. Mesenchymal stromal cells normally support mucosal homeostasis but adopt a proinflammatory phenotype during chronic inflammation [30,31]. Single-cell RNA sequencing studies have identified fibroblast subsets involved in T-cell recruitment and resistance to anti-TNF therapy [32]. These fibroblast–immune–epithelial interactions contribute to pathological angiogenesis, ECM remodeling, and perpetuation of the inflammatory cycle. Thus, the mesenchymal compartment is an active participant rather than a passive bystander, representing an emerging therapeutic target in IBD.
1.1.3. Cytokines and Molecular Signaling
Cytokines orchestrate immune responses and tissue remodeling in IBD. Dysregulation of proinflammatory and antifibrotic signaling pathways drives both chronic inflammation and fibrotic complications. The TGF-β/Smad pathway has a dual role: physiologically maintaining immune tolerance [33], and in inflammatory settings inducing fibroblast-to-myofibroblast transition and excessive ECM deposition.Overexpression of Smad7, which interferes with transforming growth factor-β1 (TGF-β1) signaling, is a characteristic feature of inflammatory bowel disease [34].In addition, Smad7 inhibition has shown therapeutic potential in experimental models [35]. IL-17A, produced by Th17 cells, promotes collagen and heat shock protein 47 (HSP47) expression in fibroblasts [36]; however, clinical blockade of IL-17 has proven ineffective, highlighting its context-dependent functions [37]. TNF-α enhances TGF-β and tissue inhibitor of metallproteinase-1 (TIMP-1) expression, contributing to fibrosis [38]; anti-TNF agents reduce inflammation but fail to prevent strictures [39]. Th2 cytokines (IL-4, IL-13) which are overexpressed in fibrotic IBD, also drive collagen synthesis [40], whereas Th1 cytokines, particularly IFN-γ, exert antifibrotic effects by inhibiting TGF-β signaling [41,42,43]. Collectively, cytokine signaling in IBD represents a fragile balance between inflammatory and fibrogenic pathways, differing between Th1/Th17-dominant Crohn’s disease and Th2-skewed ulcerative colitis.
1.1.4. Immunologic Receptors and Cellular Responses
Genetic susceptibility to IBD is closely linked to alterations in innate immune receptors that detect microbial components. NOD2, a cytosolic receptor for muramyl dipeptide (MDP) [44], is among the most significant genetic risk factors for Crohn’s disease. NOD2 mutations impair Paneth cell defensin production [45] and autophagy [46], leading to defective microbial clearance. TLR5, a Toll-like receptor recognizing bacterial flagellin, maintains microbial homeostasis and epithelial defense; its reduced expression in severe ulcerative colitis and spontaneous colitis in TLR5-deficient mice highlight its protective role [47]. Osteopontin (Opn), a matrix glycoprotein with immunomodulatory activity, is elevated in both the serum [48] and intestinal tissue of IBD patients [49], correlating with disease severity and chronic inflammation [50]. Dysregulation of these innate immune receptors not only initiates aberrant inflammatory responses but also perpetuates mucosal injury, identifying them as promising targets for next-generation immunotherapies.
1.1.5. Microbiota and Receptor-Mediated Signaling
The intestinal microbiota is a key determinant of immune homeostasis, and dysbiosis is a hallmark of IBD. Alterations in the microbial composition influence the local metabolism, including bile acid profiles, thereby affecting immune signaling and epithelial integrity. Bile acid receptors such as G-protein-coupled bile acid receptor 1 (GPBAR1, also known asTGR5) [51] and the nuclear receptor FXR modulate cytokine secretion and barrier function [52]; dysbiosis reduces the natural ligands for these receptors, promoting inflammation and permeability. The transcription factor (RAR-related orphan receptor gamma)RORγt, crucial for Th17 differentiation, is indirectly regulated by microbial metabolites, emphasizing the microbiota–immune axis [53,54]. Beyond compositional shifts, dysbiosis also involves profound changes in the intestinal metabolome. Overall, the microbiome and its metabolites function as an “immunologic organ” shaping the balance between tolerance and inflammation, with distinct microbial and metabolic profiles characterizing Crohn’s disease and ulcerative colitis.
Figure 1.
Cellular and molecular interactions driving inflammation and mucosal healing in ulcerative colitis.
Figure 1.
Cellular and molecular interactions driving inflammation and mucosal healing in ulcerative colitis.
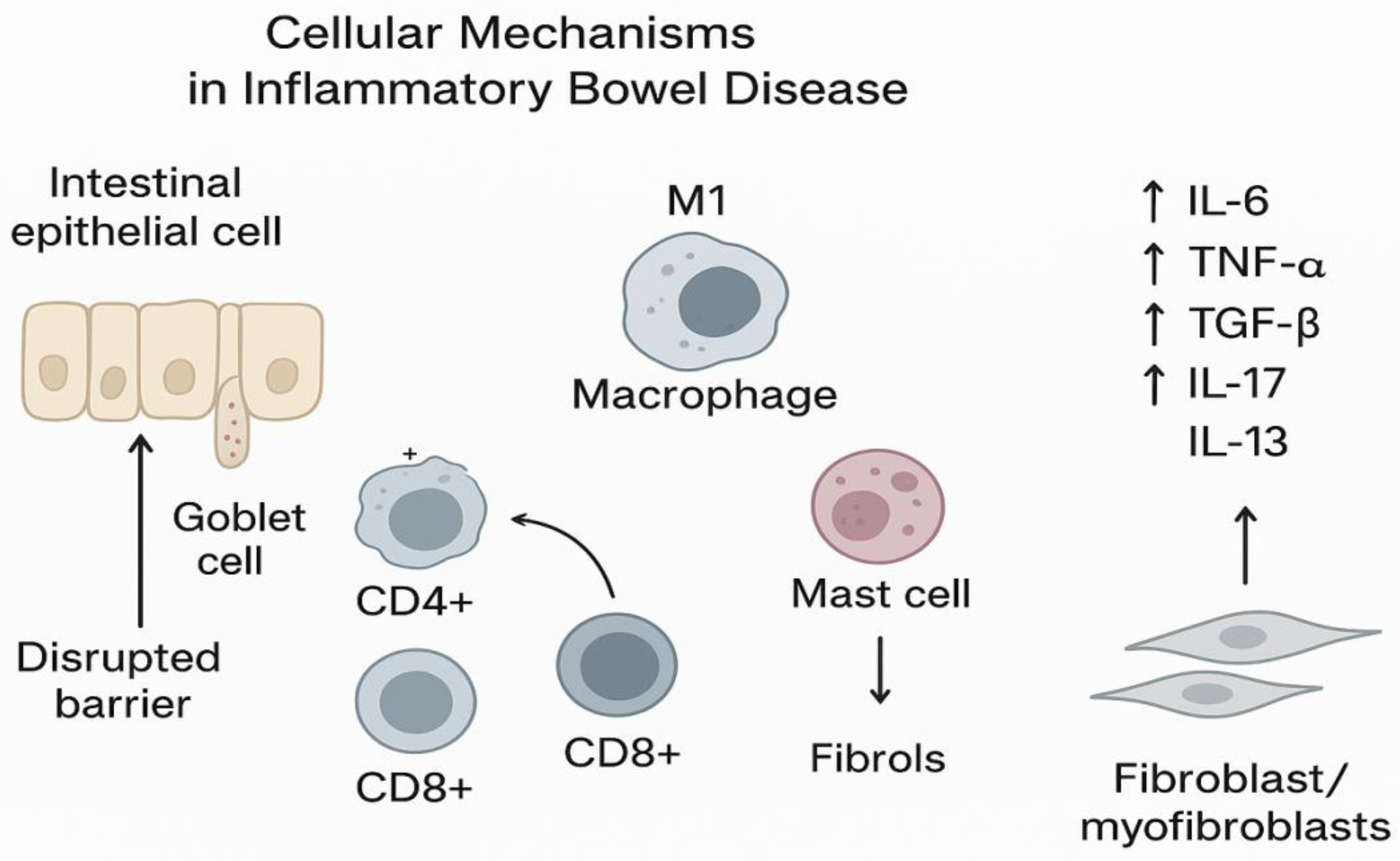
Schematic representation of the intestinal mucosa during active inflammation (left) and histologic healing (right). In active disease, intestinal epithelial cells (IEC) show barrier disruption, goblet cell depletion, and increased expression of proinflammatory cytokines such as interleukin-6 (IL-6). Activated CD4+ T cells accumulate within the lamina propria and intraepithelial compartment, releasing cytokines that amplify inflammation and tissue injury. CD8+ T cells contribute to epithelial cytotoxicity and barrier breakdown.
During histologic healing, epithelial integrity and mucus production are restored. IL-6 expression markedly decreases, while CD8+ cells may persist, contributing to immune surveillance and mucosal repair. CD4+ cells in the lamina propria likely reflect ongoing immune regulation.
Abbreviations: IEC—intestinal epithelial cells; LP—lamina propria; IL-6—interleukin 6; UC—ulcerative colitis.
2. Results
2.1. Histological Disease Activity
All 20 patients included in the study had paired colonic biopsies, one from an active disease phase and one from a subsequent histologic healing phase, using the Geboes score, which assesses the histological activity in ulcerative colitis. This grading system includes the following features: crypt architecture, chronic inflammation and eosinophilis in the lamina propria, intraepithelial neutrophils and lamina propria neutrophils, crypt destruction and surface epithelial injury [55]. The Geboes score ranges from 0 to 5.4. A simplified version indicates an active histological inflammation when the score is ≥3.1 and inactive disease of ulcerative colitis with a score less than 3.1 [56,57].
During the active phase, the Geboes score ranged from 1.2 to 5.4 (median ≈ 4.0), reflecting variable histologic inflammation. In contrast, during histologic healing, scores ranged from 0.3 to 2.0 (median ≈ 1.5), indicating significant mucosal improvement.
While in the active biopsies, the Geboes score ranged widely, the biopsies harvested during histologic healing had a narrow distribution, supporting the robustness of histologic remission as a defined endpoint, overall the difference was statistically significant (p < 0.01) (Figure 2).
Despite theoverall group consistency, one patienthad a notably low Geboes score of 0.3, in the histologic healing phase, potentially indicating a deeper state of mucosal quiescence. Conversely, three patients in the active group had Geboes scores ≤ 1.3, suggesting mild or patchy inflammation.
2.2. Distribution of T Lymphocyte Subsets
2.2.1. CD4+ Lymphocytes
CD4+ T cells were identified in both the intraepithelial compartmentand thelamina propria.
Intraepithelial CD4+ cells number had a median value of 5 during activity and of 4 during histologic healing, a difference that was not significant (p < 0.05).However, we observed a more significant standard deviation for the active phase (6.9 versus 3.2 in the histologic healing phase).
Different to the intraepithelial compartment, the lamina propria CD4+ cell counts didnot consistently decrease during histologic healing: the mean stayed almost the same (30.25 versus 29.25), whilethe median increased significantly (10 to 30), suggesting that there is a possible repopulating or regulatory behavior of CD4+ cells in healing tissue.
These findings indicate that intraepithelial CD4+ cells correlate with active inflammation, while lamina propria CD4+ cells may persist beyond visible healing, possibly reflecting a regulatory or memory phenotype.
2.2.2. CD8+ Lymphocytes
CD8+ T cells were consistently present in both compartments.
CD8+ cells were also found in epithelial and stromal compartments during both phases. Compared to intraepithelial CD4+ cells (mean: 6.8), theCD8+ cells were generally higher in number (11,25) during the active phase.CD8+ cells appear to be more consistently present across patients during active inflammation.Their relatively consistent presence and higher numbers suggest they play a central role in mucosal immunity during active colitis, possibly contributing to epithelial damage or regulating immune responses.
In contrast to the CD4+ intraepithelial cells (which decreased with healing), the CD8+ cells increased significantly (p < 0.05) in histologic healing.The mean nearly doubled, and the median increased by 60%.This pattern suggests that CD8+ lymphocytes may contribute to epithelial repair and immune surveillance during remission, in addition to their cytotoxic role in active disease (Figure 3 and Figure 4).
2.3. IL-6 Expression
IL-6 immunoreactivity was detected in epithelial, stromal, and endothelial compartments of colonic biopsies during both the active inflammation and histologic healing phases. In the active phase, IL-6 staining was generally higher across all compartments, with median scores of 1.0–2.0 observed in the epi the liumand stroma, and lower expression in the endothelium. The epithelial and stromal compartments frequently showed moderate tostrong IL-6 positivity, reflecting active cytokine production during inflammation.
During histologic healing, IL-6 expression significantly decreased in all compartments (p < 0.01). The majority of epithelialandstromalsamplesexhibitedlow or absent IL-6 staining (median scorescloseto 0–1), indicating resolution of active inflammatory signaling. Endothelial IL-6 staining remainedlow in bothphases, with minimal changes.
Overall, these findings demonstrate a marked reduction in IL-6 expression in themucosa during healing comparedto active disease, consistent with decreased inflammatory activity and cytokine production as the tissue recovers.
2.4. Correlation Analysis
We used correlationanalysis to explore the relationships betweenthe CD4+, CD8+, and IL-6 expression patterns (Figure 5).
- In the active phase, correlations between intraepithelial CD4+ and CD8+ counts were weak (Spearman’s ρ ≈ 0.21, p = 0.35), suggesting independent recruitment during inflammation.
- During histologic healing, the correlation strengthened significantly (ρ ≈ 0.62, p< 0.01), reflecting a more coordinated immune cell presence consistent with restoration of mucosal homeostasis.
- IL-6 expression correlated positively with intraepithelial CD4+ density (ρ = 0.55, p< 0.05) during active disease but lost significance in remission, highlighting its role in inflammation-driven immune activation.

Table 1.
Quantitative summary of CD4+, CD8+ lymphocyte counts and IL-6 expression in active and histologic healing phases of ulcerative colitis.
Table 1.
Quantitative summary of CD4+, CD8+ lymphocyte counts and IL-6 expression in active and histologic healing phases of ulcerative colitis.
| Marker | Compartment | Active Disease (Mean ± SD) | Median (IQR) | Histologic Healing (Mean ± SD) | Median (IQR) |
|---|---|---|---|---|---|
| CD4+ | Intraepithelial | 6.8 ± 6.9 | 5 (3–9) | 3.75 ± 3.2 | 4 (2–6) |
| Lamina propria | 30.25 ± — | 10 (6–25) | 29.25 ± — | 30 (20–40) | |
| CD8+ | Intraepithelial | 11.25 ± — | 10 (7–15) | 22.5 ± — | 16 (12–25) |
| Lamina propria | — | — | — | — | |
| IL-6 | Epithelium | 1.5 ± — | 1–2 | 0.5 ± — | 0–1 |
| Stroma | 1.5 ± — | 1–2 | 0.5 ± — | 0–1 | |
| Endothelium | 0.5 ± — | — | 0.5 ± — | — |
Note: Data are presented as mean ± standard deviation (SD) and median (interquartile range, IQR) where available. SD not available for some parameters due to limited sample variability or descriptive evaluation.Cell counts are expressed as number of positive cells per high-power field (HPF).IL-6 staining intensity was semi-quantitatively scored (0 = absent, 1 = mild, 2 = moderate, 3 = strong).
Together, these results suggest a shift from uncoordinated pro-inflammatory T-cell infiltrationtowardregulated immune surveillance and cytokine downregulationas healing progresses.
These histologic and immunologic findings provide insight into the persistence and modulation of mucosal immunity, forming the basis for the discussion below on the immune dynamics and potential therapeutic implications.
3. Discussion
3.1. Histologic and Immunologic Correlates of Mucosal Healing
Histologic activity has been increasingly recognized as a more sensitive indicator of residual disease, and growing evidence supports histologic remission being associated with a lower risk of relapse, hospitalization, and colectomy. These findings are aligned with previous studies suggesting that histologic healing should be considered a therapeutic target in UC management.
We also observed minor overlap at the lower end of the inflammatory scale. A small subset of patients with active disease had Geboes scores below 2.0, and one patient in the histologic healing phase had a score of 0.3, reflecting deep remission. These cases emphasize the heterogeneity of inflammation in UC, the potential for sampling variability, and the limits of clinical-endoscopic correlation. Histologic assessment adds an objective layer to disease evaluation and may be particularly valuable in therapeutic decision-making, especially in cases wherethe symptoms and endoscopic appearance do not align.
CD4+ T cells are helper T-cells involved in immune activation. In the epithelium, they can signal active inflammation and immune surveillance.
In ourgroup, thenumber of intraepithelial CD4+ lymphocyteswas significantly reduced during histologic healing compared to the active phase. The mean count decreased from 6.8 to 3.75 cells per high-powerfield (HPF), with a corresponding decrease in median values (5 vs. 4) and a shift in distribution toward lower counts. A Wilcoxonsigned-rank test confirmed that this reduction was statistically significant (p < 0.05), suggesting that intraepithelial CD4+ lymphocytes are associated with active mucosal inflammation and diminish as histologic healingoccurs.
In contrast, the CD4+ celldensity in lamina propria hot spotsdidnot demonstrate a consistent decrease during healing. Duringthe active phase, the estimated counts ranged widely (mean: 30.25 cells/HPF; median: 10), reflectingheterogeneous immune infiltration. During histologic healing, the mean remained comparable (29.25 cells/HPF), but the median increased to 30 cells/HPF, and the distribution shiftedupward, indicatingpersistence or even repopulation of CD4+ lymphocytes. These findings suggest that while intraepithelial CD4+ cells are closelytiedto active inflammation, lamina propria CD4+ cells may remain during healing and could reflect ongoing immune regulation or residual inflammation.
The higher counts during the active phase reflect the recruitment of CD4+ cells to the inflamed mucosa and an ongoing autoimmune response. The decrease during histologic healing reveals a reduction in mucosal immune activation with partial or full restoration of mucosal homeostasis. However, there is not a complete return to “zero,” as some CD4+ cells persist indicating a possibile immune memory or low-grade residual inflammation.Thesedatasupportthe idea that CD4+ intraepitheliallymphocytes are markers of active inflammation in ulcerativecolitisandmaysubsideduring histologic remission—but notnecessarilyvanish, whichcouldimplytheyhave apotential role in diseaserecurrence.
The analysis of CD8+ lymphocytes in both intraepithelial and lamina propria compartments revealed distinct patterns during active inflammation and histologic healing in ulcerative colitis. Intraepithelial CD8+ cells increased significantly during healing, suggesting a potential role in mucosal repair or immune regulation once active inflammation subsides. Conversely, lamina propria CD8+ cells showed stable mean counts but a decrease in median values, reflecting heterogeneity in their persistence during remission. This sustained presence of CD8+ cells in the lamina propria may indicate ongoing immune surveillance or low-level inflammation despite histologic healing. Overall, CD8+ cells appear to contribute dynamically to the immune environment of ulcerative colitis, with possible roles beyond direct cytotoxicity, including tissue homeostasis and protection.
3.2. Integration of Cellular and Cytokine Findings
The correlation analysis between CD4+ and CD8+ lymphocytepopulations within both the intraepithelial compartment and the lamina propria provides important insights into the immune dynamics in ulcerative colitis. In the active phase, the relationship between intraepithelial CD4+ and CD8+ cells appeared weak or non-significant, suggesting that these T cellsubsets may expand independently during active inflammation. However, during histologic healing, a strongerpositivecorrelationwasobserved, indicating a more coordinatedpresence or regulation of theselymphocytesubsets in mucosalremission. This pattern could reflect a reestablishment of immunehomeostasis, where CD4+ helper T cellsand CD8+ cytotoxic or regulatory T cells functionally interacttomaintainmucosalintegrity and prevent relapse.
In contrast, in the lamina propria, correlations between CD4+ and CD8+ cells were modest and variable across disease phases. The persistent presence of both cell types, with onlypartial correlation, suggests complex and possibly compartment-specific immune mechanisms. The lamina propria mayharbor diverse T cell populations with distinct roles in sustaining chronic inflammation or immune regulation. The lack of strong correlation during active disease may reflect heterogeneous immune activation, while partial correlation in healing could indicate evolving immune interactions as the mucosa recovers.
Overall, the sefindingshigh light that CD4+ and CD8+ T cells do not uniformly change in parallel but may play complementary roles depending on the mucosal compartment and disease stage. Understanding the sequenced relationships could inform targetedimmunomodulatory the rapies aimed at restoring balanced T cell responses in ulcerative colitis.
3.3. IL-6 Expression and Cytokine Modulation
The evaluation of IL-6 expression across epithelial, stromal, and endothelial compartments revealed a significant decrease in IL-6 staining during histologic healing compared to the active phase of ulcerative colitis. This reduction is consistent with the resolution of mucosal inflammation, as IL-6 is a well-known pro-inflammatory cytokine involved in promoting immune cell activation and sustaining inflammatory responses. The higher IL-6 levels observed in the epithelial and stromal compartments during active disease likely reflect the ongoing inflammatory milieu and epithelial stress. Conversely, the low IL-6 expression in the healing phase indicates dampened cytokine signaling, which may contribute to mucosal restoration and maintenance of immune homeostasis. Interestingly, endothelial IL-6 expression was low in both phases, suggesting a lesser role for endothelial cells in IL-6–mediated inflammation in this context. The combined reduction in intraepithelial CD4+ cells, the persistence of lamina propria CD4+ and CD8+ populations, and the marked decrease in IL-6 expression together illustrate a gradual transition from inflammatory activation to controlled immune surveillance. Overall, these findings underscore the importance of IL-6 as a biomarker of active mucosal inflammation and a potential therapeutic target in ulcerative colitis.
3.4. Clinical Implications
From a clinical standpoint, our data support the use of histologic evaluation alongside endoscopic assessment as part of a deeper remission concept in UC. The persistence of immune cell subsets during histologic healing may explain why some patients experience relapse despite endoscopic quiescence. Targeted immunomodulatory strategies that reinforce regulatory CD4+ and reparative CD8+ cell functions while suppressing IL-6–driven inflammation may sustain remission and reducethe relapse risk.
Furthermore, histologic biomarkers such as IL-6 staining patterns and T-cell compartmentalization could complement traditional scoring systems such as the Geboes or Nancy Index, refining therapeutic monitoring and patient stratification. The Janus kinase (JAK) which activates the signal transducer and activator of transcription (STAT) pathway is important to the pathogenesis of IBD [58]. An oral pan-JAK inhibitor (Tofacitinib) has shown clinical efficacy and is approved for the treatment of ulcerative colitis [59,60]. In the last few years, researchers have developed selective JAK 1 inhibitors such as Filgotinib and Upadatinib which showed improvements in moderate to severe UC [61,62].
4. Materials and Methods
4.1. Study Design and Population
To investigate the mucosal immune landscape during disease activity and histologic healing, we performed immunohistochemical analysis of T-cell subsets and IL-6 expression.
This observational, cross-sectional study was conducted to evaluate the relationship between T lymphocyte subsets (CD4+, CD8+) and interleukin-6 (IL-6) expression in the colonic mucosa of patients with ulcerative colitis (UC). Twenty adult patients with a confirmed diagnosis of UC were recruited from the Gastroenterology Department of Colentina University Hospital, Bucharest and all their biopsies were evaluated, for diagnosis and research purposes in the Pathology Department of the same hospital. The study protocol was approved by the Institutional Review Board, and written informed consent was obtained from all participants prior to inclusion.
4.2. Inclusion and Exclusion Criteria
Eligible participants were aged between 18 and 75 years and had a confirmed diagnosis of UC for at least six months, based on standard clinical, endoscopic, and histological criteria. Patients were required to have undergone a recent colonoscopy with available biopsy samples.
The exclusion criteria included: (i) concurrent diagnosis of Crohn’s disease or indeterminate colitis; (ii) history of colonic tuberculosis, colorectal cancer or dysplasia; (iii) recent use of investigational drugs; and (iv) presence of active systemic infection at the time of sampling.
4.3. Study Groups and Sampling Strategy
For each patient, twoindependent sets of biopsies samples were analyzed:
- Active phase samples, obtained during a documented clinical and endoscopic flare (Mayo endoscopic subscore 2–3).
- Histologic healing samples, collected later from the same colonic segments once patients achieved endoscopic remission (Mayo subscore 0) and were clinically asymptomatic.
Histologic healing was defined as a Geboes score < 2.0, corresponding to the absence of neutrophilic infiltration, crypt destruction, or abscesses, erosions, or ulceration. Thus, each patient served as their own control, allowing direct comparison between inflammatory and remission states.
4.4. Clinical and Endoscopic Assessment
Disease activity, in both moments was assessed using the Mayo score, including both clinical parameters and endoscopic findings. Patients with active UC had a Mayo endoscopic subscore of 2 or 3 and reported increased stool frequency, urgency, and rectal bleeding. In the histologic healing moment, they had a Mayo endoscopic subscore of 0 and were asymptomatic at the time of biopsy.
4.5. Biopsy Collection and Processing
Colonic biopsies were obtained during astandard colonoscopy from predefined locations (rectum, sigmoid, and descending colon). At least two mucosal samples per site were collected using cold forceps. The specimens were immediately fixed in 10% neutral buffered formalin and processed for histological evaluation.
4.6. Histopathological Evaluation
Formalin-fixed, paraffin-embedded (FFPE) tissue blocks were sectioned at 3μm thickness and stained with hematoxylin and eosin (H&E) for initial histologic assessment. Two experienced gastrointestinal pathologists, blinded to the clinical data, evaluated the samples using the Geboes score. Any discrepancies in scoring were resolved through joint review and consensus (Figure 6 and Figure 7).
4.7. Immunohistochemical Analysis
In addition to H&E, immunohistochemical (IHC) staining was performed on each biopsy block using the following antibodies:
- CD3 (pan–T-cell marker)
- CD4 (helper T-cell marker)
- CD8 (cytotoxic T-cell marker)
- IL-6 (pro-inflammatory cytokine)
IHC staining was carried out on an automated platform (Leica Bond), according to the manufacturer’s protocols. After deparaffinization and rehydration, antigen retrieval was performed using citrate buffer (pH 6.0) or EDTA (pH 9.0), depending on the antibody. Visualization was achieved using DAB (3,3′-diaminobenzidine) as chromogen, and counterstaining was performed with hematoxylin.
The density of T lymph cells (CD3+, CD4+, CD8+) was scored in the hot spot in the lamina propria (number of positive cells on one high-power field with a diameter of 0,55 mm) and in the intraepithelial compartment (number of inflammatory cells reported at 100 epithelial cells).
IL-6 expression was assessedsemi-quantitatively, in epithelial and endothelial cells, and in the inflammatory infiltrates from the stromal compartment. The results were expressed as categorical scores (0—absent; 1—mild; 2—moderate; 3—strong) staining intensity.
4.8. Ethical Considerations
The study protocol was approved by the Institutional Review Board of Colentina University Hospital. All patients provided written informed consent prior to inclusion. All procedures were conducted in accordance with the Declaration of Helsinki and relevant institutional guidelines.
4.9. Statistical Analysis
Descriptive statistics were used to summarize thedemographic and clinical characteristics. Differences in histological parameters between groups were assessed using non-parametric tests (Mann-Whitney U test for continuous variables; Fisher’s exact test for categorical variables). A p-value < 0.05 was considered statistically significant.
The immunohistochemical characterization of CD4+, CD8+, and IL-6 expression allowed comparative analysis of mucosal immune activity during active inflammation and histologic healing in ulcerative colitis.
5. Study Limitations
This study has several limitations. First, the sample size was relatively small (n = 20), which may limit the statistical power and generalizability of the findings. Second, the cross-sectional design precludes causal inference regarding the dynamic changes in mucosal T-cell populations and IL-6 expression over time. Third, only a limited set of immune markers (CD4, CD8, and IL-6) was analyzed; inclusion of additional cytokines and functional markers (e.g.,TNF-α, TGF-β, FOXP3, IL-17, granzyme B) could provide a more comprehensive understanding of immune regulation in ulcerative colitis. Fourth, inter-patient heterogeneity, despite paired biopsies, may influence immune cell distributions and histologic responses. Finally, the semi-quantitative nature of immunohistochemistry may not capture the full complexity of thecellular and cytokine dynamics, which could be further elucidated using multiplexed immunostaining or spatial transcriptomics.
Despite these limitations, our findings contribute to a growing understanding of immune modulation in UC histologic healing. Future research should integrate immunohistochemistry, transcriptomic, and spatial profiling to map mucosal immune niches in remission. Identifying immune signatures of durable healing could inform personalized therapeutic strategies and novel biomarkers for clinical monitoring.
6. Conclusions and Future Perspectives
Inflammatory bowel disease (IBD) represents a complex interplay of epithelial barrier dysfunction, dysregulated immune responses, mesenchymal activation, and microbiota alterations.
This study evaluated the dynamics of intraepithelial and lamina propria CD4+ and CD8+ lymphocytes, alongside IL-6 expression, in colonic biopsies from patients with ulcerative colitis during active inflammation and histologic healing. CD4+ and CD8+ intraepithelial lymphocytes showed distinct patterns, with CD8+ cells increasing during healing, suggesting a potential regulatory or reparative role. In contrast, lamina propria CD4+ and CD8+ cells remained variably persistent even during remission, indicating possible ongoing immune surveillance or subclinical inflammation.
IL-6 expression was significantly higher in the epithelial and stromal compartments during active disease and decreased markedly during histologic healing, in line with reduced cytokine-driven inflammation. The endothelial IL-6 staining remained low in both phases.
Collectively, these findings highlight that while histologic healing is accompanied by reduced IL-6 expression and changes in lymphocyte populations, certain immune cell subsets may persist beyond the resolution of active inflammation. This suggests that mucosal immune regulation continues during remission and may influence disease recurrence. The combined evaluation of T cell profiles and cytokine expression offers valuable insights into mucosal immune dynamics and may guide future therapeutic strategies targeting immune resolution in ulcerative colitis.
Future research should focus on the following:
- Targeted therapeutics: refining inhibitors of Smad7, MEK [63], and specific cytokines, while assessing their safety and efficacy in patient subpopulations.
- Microbiota modulation: exploring bile acid receptor agonists and microbiome-targeted therapies to restore homeostasis and reduce inflammation [53].
- Single-cell and spatial omics: identifying novel cellular subtypes and their interactions to uncover predictive biomarkers for disease progression and therapeutic response [64].
- Integrative SDOH analysis: evaluating the impact of social determinants of health on treatment adherence, disease severity, and hospitalization rates, to guide holistic preventive strategies [65].
- Translational models: developing patient-derived organoids, co-culture systems, and humanized mouse models for mechanistic studies and preclinical drug testing [66].
In conclusion, a multidimensional understanding of cellular, molecular, and environmental contributors to IBD is essential for developing personalized therapies and preventive strategies. Continued integration of mechanistic insights with clinical and socio-environmental data holds promise for improving patient outcomes and reducing disease burden.
7. Conclusion
This research demonstrates that histologic healing in ulcerative colitis (UC) is accompanied by significant modulation of themucosal immune architecture.These findings indicate that mucosal healing represents not merely the absence of inflammation, but an active process of immune remodeling and balance restoration. The persistence of certain lymphocyte subsets even during remission suggests that mucosal immune regulation continues beyond endoscopic or clinical recovery. This emphasizes the need to integrate histologic and immunologic biomarkers into treatment monitoring and therapeutic endpoints.Integration of cellular and molecular data could ultimately lead to precision-guided immunomodulatory approaches in this disease.Future research should aim to characterize the phenotypic and functional plasticity of mucosal T cellsusing multiplex immunohistochemistry and spatial transcriptomics, to delineate pro-inflammatory versus reparative subpopulations.
Author Contributions
Conceptialization, R.M. and C.P.; methodology: C.P. and A.C.; validation R.M., C.P., A.C. and M.S.; formal analysis C.P., A.C.; investigation and data curation C.P. and A.C., writing—original draft R.M.; preparation supervision R.M. and C.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study protocol was approved by the Institutional Review Board of Colentina University Hospital. The approval code is 39, the approval date is 4.11.2025.
Informed Consent Statement
Written informed consent has been obtained from the patient(s) to publish this paper.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mirică, R.E.; Matură, T.F.; Craciun, E.; Pavel, D. The Importance of Magnetic Resonance Enterography in Monitoring Inflammatory Bowel Disease: A Review of Clinical Significance and Current Challenges. Diagnostics 2025, 15, 1540. [Google Scholar] [CrossRef]
- Coward, S.; Clement, F.; Benchimol, E.I.; Bernstein, C.N.; Avina-Zubieta, J.A.; Bitton, A.; Carroll, M.W.; Hazlewood, G.; Jacobson, K.; Jelinski, S.; et al. Past and future burden of inflammatory bowel diseases based on modeling of population-based data. Gastroenterology 2019, 156, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Jucan, A.E.; Gavrilescu, O.; Dranga, M.; Popa, I.V.; Mihai, I.-R.; Mihai, V.-C.; Stefanescu, G.; Drug, V.L.; CijevschiPrelipcean, C.; Vulpoi, R.-A. Evaluation of Disease Activity in Inflammatory Bowel Disease: Diagnostic Tools in the Assessment of Histological Healing. Biomedicines 2023, 11, 3090. [Google Scholar] [CrossRef] [PubMed]
- Goswami, T.K.; Singh, M.; Dhawan, M.; Mitra, S.; Bin Emran, T.; Rabaan, A.A.; Al Mutair, A.; Al Alawi, Z.; Alhumaid, S.; Dhama, K. Regulatory T cells (Tregs) and their therapeutic potential against autoimmune disorders—Advances and challenges. Hum. Vaccines Immunother. 2022, 18, 2035117. [Google Scholar] [CrossRef]
- Jacobse, J.; Li, J.; Rings, E.H.H.M.; Samsom, J.N.; Goettel, J.A. Intestinal Regulatory T Cells as Specialized Tissue-Restricted Immune Cells in Intestinal Immune Homeostasis and Disease. Front. Immunol. 2021, 12, 716499. [Google Scholar] [CrossRef] [PubMed]
- Van Den Broek, T.; Borghans, J.A.M.; Van Wijk, F. The full spectrum of human naive T cells. Nat. Rev. Immunol. 2018, 18, 363–373. [Google Scholar] [CrossRef]
- Vebr, M.; Pomahačová, R.; Sýkora, J.; Schwarz, J.A. Narrative Review of Cytokine Networks: Pathophysiological and Therapeutic Implications for Inflammatory Bowel Disease Pathogenesis. Biomedicines 2023, 11, 3229. [Google Scholar] [CrossRef]
- Pawłowska-Kamieniak, A.; Krawiec, P.; Pac-Kożuchowska, E. Interleukin 6: Biological significance and role in inflammatory bowel diseases. Adv. Clin. Exp. Med. 2021, 30, 465–469. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Hirano, T.; Matsuda, T.; Hosoi, K.; Okano, A.; Matsui, H.; Kishimoto, T. Absence of antiviral activity in recombinant B cell stimulatory factor 2 (BSF-2). Immunol. Lett. 1988, 17, 41–45. [Google Scholar] [CrossRef]
- Kusugami, K.; Fukatsu, A.; Tanimoto, M.; Shinoda, M.; Haruta, J.; Kuroiwa, A.; Ina, K.; Kanayama, K.; Ando, T.; Matsuura, T. Elevation of interleukin-6 in inflammatory bowel disease is macrophage- and epithelial cell-dependent. Dig. Dis. Sci. 1995, 40, 949–959. [Google Scholar] [CrossRef]
- Lu, Q.; Yang, M.-F.; Liang, Y.-J.; Xu, J.; Xu, H.-M.; Nie, Y.-Q.; Wang, L.-S.; Yao, J.; Li, D.-F. Immunology of Inflammatory Bowel Disease: Molecular Mechanisms and Therapeutics. J. Inflamm. Res. 2022, 15, 1825. [Google Scholar] [CrossRef]
- Shahini, A.; Shahini, A. Role of interleukin-6-mediated inflammation in the pathogenesis of inflammatory bowel disease: Focus on the available therapeutic approaches and gut microbiome. J. Cell Commun. Signal. 2023, 17, 55. [Google Scholar] [CrossRef]
- Zhao, C.; Lin, S. PANoptosis in intestinal epithelium: Its significance in inflammatory bowel disease and a potential novel therapeutic target for natural products. Front. Immunol. 2025, 15, 1507065. [Google Scholar] [CrossRef] [PubMed]
- Travassos, L.H.; Carneiro, L.A.; Ramjeet, M.; Hussey, S.; Kim, Y.G.; Magalhães, J.G.; Yuan, L.; Soares, F.; Chea, E.; Le Bourhis, L.; et al. Nod1 and Nod2 direct autophagy by recruiting ATG16L1 to the plasma membrane at the site of bacterial entry. Nat. Immunol. 2010, 11, 55–62. [Google Scholar] [CrossRef]
- Hoefkens, E.; Nys, K.; John, J.M.; Van Steen, K.; Arijs, I.; Van der Goten, J.; Van Assche, G.; Agostinis, P.; Rutgeerts, P.; Vermeire, S.; et al. Genetic association and functional role of Crohn disease risk alleles involved in microbial sensing, autophagy, and endoplasmic reticulum (ER) stress. Autophagy 2013, 9, 2046–2055. [Google Scholar] [CrossRef]
- Goto, Y.; Kiyono, H. Epithelial barrier: An interface for the cross-communication between gut flora and immune system. Immunol. Rev. 2012, 245, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Park, H.; Choe, B.-H.; Kang, B. The Role and Function of Mucins and Its Relationship to Inflammatory Bowel Disease. Front. Med. 2022, 9, 848344. [Google Scholar] [CrossRef]
- Larsson, J.M.H.; Karlsson, H.; Crespo, J.G.; Johansson, M.E.V.; Eklund, L.; Sjövall, H.; Hansson, G.C. Altered O-glycosylation profile of MUC2 mucin occurs in active ulcerative colitis and is associated with increased inflammation. Inflamm. Bowel Dis. 2011, 17, 2299–2307. [Google Scholar] [CrossRef] [PubMed]
- Gersemann, M.; Becker, S.; Kübler, I.; Koslowski, M.; Wang, G.; Herrlinger, K.R.; Griger, J.; Fritz, P.; Fellermann, K.; Schwab, M.; et al. Differences in goblet cell differentiation between Crohn’s disease and ulcerative colitis. Differentiation 2009, 77, 84–94. [Google Scholar] [CrossRef]
- Quan, T.; Li, R.; Gao, T. The Intestinal Macrophage-Intestinal Stem Cell Axis in Inflammatory Bowel Diseases: From Pathogenesis to Therapy. Int. J. Mol. Sci. 2025, 26, 2855. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Liu, S.; Chen, S.; Zhang, F.; Li, N.; Yin, J.; Peng, Y.; Wu, L.; Liu, G.; Yin, Y.; et al. Dietary L-glutamine supplementation increases Pasteurella multocida burden and the expression of its major virulence factors in mice. Amino Acids 2013, 45, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.J.; Frei, S.M.; Stevens, R.L. The Multifaceted Mast Cell in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2014, 20, 2364–2378. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Hogan, S.P.; Lee, J.J.; Foster, P.S.; Rothenberg, M.E. Fundamental signals that regulate eosinophil homing to the gastrointestinal tract. J. Clin. Invest. 1999, 103, 1719–1727. [Google Scholar] [CrossRef]
- Rothenberg, M.E.; Mishra, A.; Brandt, E.B.; Hogan, S.P. Gastrointestinal eosinophils. Immunol. Rev. 2001, 179, 139–155. [Google Scholar] [CrossRef]
- Fagerholm, S.C. Integrins in Health and Disease. N. Engl. J. Med. 2022, 387, 1519–1521. [Google Scholar] [CrossRef]
- Ito, T.; Kayama, H. Roles of fibroblasts in the pathogenesis of inflammatory bowel diseases and IBD-associated fibrosis. Int. Immunol. 2025, 37, 377–392. [Google Scholar] [CrossRef]
- McLean, L.P.; Shea-Donohue, T.; Cross, R.K. Vedolizumab for the treatment of ulcerative colitis and Crohn’s disease. Immunotherapy 2012, 4, 883–898. [Google Scholar] [CrossRef]
- Degirmenci, B.; Valenta, T.; Dimitrieva, S.; Hausmann, G.; Basler, K. GLI1-expressing mesenchymal cells form the essential Wnt-secreting niche for colon stem cells. Nature 2018, 558, 449–453. [Google Scholar] [CrossRef]
- Biancheri, P.; Di Sabatino, A.; Corazza, G.R.; MacDonald, T.T. Proteases and the gut barrier. Cell Tissue Res. 2013, 351, 269–280. [Google Scholar] [CrossRef]
- Schreurs, R.; Baumdick, M.E.; Sagebiel, A.F.; Kaufmann, M.; Mokry, M.; Klarenbeek, P.L.; Schaltenberg, N.; Steinert, F.L.; van Rijn, J.M.; Drewniak, A.; et al. Human fetal TNF-alpha-cytokine-producing CD4+ effector memory T cells promote intestinal development and mediate inflammation early in life. Immunity 2019, 50, 462–476.e8. [Google Scholar] [CrossRef] [PubMed]
- Marek, A.; Brodzicki, J.; Liberek, A.; Korzon, M. TGF-beta (transforming growth factor-beta) in chronic inflammatory conditions–A new diagnostic and prognostic marker? Med. Sci. Monit. 2002, 8, RA145–RA151. [Google Scholar] [PubMed]
- Monteleone, G.; Kumberova, A.; Croft, N.M.; McKenzie, C.; Steer, H.W.; MacDonald, T.T. Blocking Smad7 restores TGF-beta1 signaling in chronic inflammatory bowel disease. J. Clin. Investig. 2001, 108, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Boirivant, M.; Pallone, F.; Di Giacinto, C.; Fina, D.; Monteleone, I.; Marinaro, M.; Caruso, R.; Colantoni, A.; Palmieri, G.; Sanchez, M.; et al. Inhibition of Smad7 with a specific antisense oligonucleotide facilitates TGF-beta1-mediated suppression of colitis. Gastroenterology 2006, 131, 1786–1798. [Google Scholar] [CrossRef]
- Honzawa, Y.; Nakase, H.; Shiokawa, M.; Yoshino, T.; Imaeda, H.; Matsuura, M.; Kodama, Y.; Ikeuchi, H.; Andoh, A.; Sakai, Y.; et al. Involvement of interleukin-17A-induced expression of heat shock protein 47 in intestinal fibrosis in Crohn’s disease. Gut 2014, 63, 1902–1912. [Google Scholar] [CrossRef]
- Li, J.; Liu, L.; Zhao, Q.; Chen, M. Role of interleukin-17 in pathogenesis of intestinal fibrosis in mice. Dig. Dis. Sci. 2020, 65, 1971–1979. [Google Scholar] [CrossRef]
- Drygiannakis, I.; Valatas, V.; Sfakianaki, O.; Bourikas, L.; Manousou, P.; Kambas, K.; Ritis, K.; Kolios, G.; Kouroumalis, E. Proinflammatory cytokines induce crosstalk between colonic epithelial cells and subepithelial myofibroblasts: Implication in intestinal fibrosis. J. Crohns Colitis. 2013, 7, 286–300. [Google Scholar] [CrossRef]
- Lichtenstein, G.R.; Olson, A.; Travers, S.; Diamond, R.H.; Chen, D.M.; Pritchard, M.L.; Feagan, B.G.; Cohen, R.D.; Salzberg, B.A.; Hanauer, S.B.; et al. Factors associated with the development of intestinal strictures or obstructions in patients with Crohn’s disease. Am. J. Gastroenterol. 2006, 101, 1030–1038. [Google Scholar] [CrossRef]
- Wynn, T.A. Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat. Rev. Immunol. 2004, 4, 583–594. [Google Scholar] [CrossRef]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef]
- Gurujeyalakshmi, G.; Giri, S.N. Molecular mechanisms of antifibrotic effect of interferon gamma in bleomycin-mouse model of lung fibrosis: Downregulation of TGF-beta and procollagen I and III gene expression. Exp. Lung Res. 1995, 21, 791–808. [Google Scholar] [CrossRef]
- Inohara, N.; Ogura, Y.; Fontalba, A.; Gutierrez, O.; Pons, F.; Crespo, J.; Fukase, K.; Inamura, S.; Kusumoto, S.; Hashimoto, M.; et al. Host recognition of bacterial muramyl dipeptide mediated through NOD2. Implications for Crohn’s disease. J. Biol. Chem. 2003, 278, 5509–5512. [Google Scholar] [CrossRef] [PubMed]
- Wehkamp, J.; Harder, J.; Weichenthal, M.; Schwab, M.; Schäffeler, E.; Schlee, M.; Herrlinger, K.R.; Stallmach, A.; Noack, F.; Fritz, P.; et al. NOD2 (CARD15) mutations in Crohn’s disease are associated with diminished mucosal alpha-defensin expression. Gut 2004, 53, 1658–1664. [Google Scholar] [CrossRef]
- Chamaillard, M.; Girardin, S.E.; Viala, J.; Philpott, D.J. Nods, Nalps and Naip: Intracellular regulators of bacterial-induced inflammation. Cell. Microbiol. 2003, 5, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Hardenberg, G.; Yao, Y.; Piccirillo, C.A.; Levings, M.K.; Steiner, T.S. Toll-like receptor 5 deficiency protects from wasting disease in a T cell transfer colitis model in T cell receptor-β-deficient mice. Inflamm. Bowel Dis. 2012, 18, 85–93. [Google Scholar] [CrossRef]
- Sato, T.; Nakai, T.; Tamura, N.; Okamoto, S.; Matsuoka, K.; Sakuraba, A.; Fukushima, T.; Uede, T.; Hibi, T. Osteopontin/Eta-1 upregulated in Crohn’s disease regulates the Th1 immune response. Gut 2005, 54, 1254–1262. [Google Scholar] [CrossRef]
- Masuda, H.; Takahashi, Y.; Asai, S.; Takayama, T. Distinct gene expression of osteopontin in patients with ulcerative colitis. J. Surg. Res. 2003, 111, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Agnholt, J.; Kelsen, J.; Schack, L.; Hvas, C.L.; Dahlerup, J.F.; Sorensen, E.S. Osteopontin, a protein with cytokine-like properties, is associated with inflammation in Crohn’s disease. Scand. J. Immunol. 2007, 65, 453–460. [Google Scholar] [CrossRef]
- Maruyama, T.; Miyamoto, Y.; Nakamura, T.T.T.; Tamai, Y.; Okada, H.; Sugiyama, E.; Itadani, H.; Tanaka, K.; Nakamura, T.T.T.; Itadani, H.; et al. Identification of membrane-type receptor for bile acids (M-BAR). Biochem. Biophys. Res. Commun. 2002, 298, 714–719. [Google Scholar] [CrossRef]
- Renga, B.; Migliorati, M.; Mencarelli, A.; Fiorucci, S. Reciprocal regulation of the bile acid-activated receptor FXR and the interferon-gamma-STAT-1 pathway in macrophages. Biochim. Biophys. Acta 2009, 1792, 564–573. [Google Scholar] [CrossRef]
- Sato, Y.; Atarashi, K.; Plichta, D.R.; Arai, Y.; Sasajima, S.; Kearney, S.M.; Suda, W.; Takeshita, K.; Sasaki, T.; Okamoto, S.; et al. Novel bile acid biosynthetic pathways are enriched in the microbiome of centenarians. Nature 2021, 599, 458–464. [Google Scholar] [CrossRef]
- Tanoue, T.; Atarashi, K.; Honda, K. Development and maintenance of intestinal regulatory T cells. Nat. Rev. Immunol. 2016, 16, 295–309. [Google Scholar] [CrossRef]
- Geboes, K.; Riddell, R.; Öst, A.; Jensfelt, B.; Persson, T.; Löfberg, R. A reproducible grading scale for histological assessment of inflammation in ulcerative colitis. Gut 2000, 47, 404–409. [Google Scholar] [CrossRef]
- Rosenberg, L.; Nanda, K.S.; Zenlea, T.; Gifford, A.; Lawlor, G.O.; Falchuk, K.R.; Wolf, J.L.; Cheifetz, A.S.; Goldsmith, J.D.; Moss, A.C. Histologic markers of inflammation in patients with ulcerative colitis in clinical remission. Clin. Gastroenterol. Hepatol. 2013, 11, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Bessissow, T.; Lemmens, B.; Ferrante, M.; Bisschops, R.; Van Steen, K.; Geboes, K.; Van Assche, G.; Vermeire, S.; Rutgeerts, P.; De Hertogh, G. Prognostic value of serologic and histologic markers on clinical relapse in ulcerative colitis patients with mucosal healing. Am. J. Gastroenterol. 2012, 107, 1684–1692. [Google Scholar] [CrossRef] [PubMed]
- Boland, B.S.; Sandborn, W.J.; Chang, J.T. Update on Janus kinase antagonists in inflammatory bowel disease. Gastroenterol. Clin. North. Am. 2014, 43, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Digby-Bell, J.L.; Atreya, R.; Monteleone, G.; Powell, N. Interrogating host immunity to predict treatment response in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 9–20. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Su, C.; Sands, B.E.; D’Haens, G.R.; Vermeire, S.; Schreiber, S.; Danese, S.; Feagan, B.G.; Reinisch, W.; Niezychowski, W.; et al. Tofacitinib as induction and maintenance therapy for ulcerative colitis. N. Engl. J. Med. 2017, 376, 1723–1736. [Google Scholar] [CrossRef]
- Feagan, B.G.; Danese, S.; Loftus, E.V.; Vermeire, S.; Schreiber, S.; Ritter, T.; Fogel, R.; Mehta, R.; Nijhawan, S.; Kempiński, R.; et al. Filgotinib as induction and maintenance therapy for ulcerative colitis (SELECTION): A phase 2b/3 double-blind, randomised, placebo-controlled trial. Lancet 2021, 397, 2372–2384. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Feagan, B.G.; Loftus, E.V., Jr.; Peyrin-Biroulet, L.; Van Assche, G.; D’Haens, G.; Schreiber, S.; Colombel, J.F.; Lewis, J.D.; Ghosh, S.; et al. Efficacy and safety of upadacitinib in a randomized trial of patients with crohn’s disease. Gastroenterology 2020, 158, 2123–2138.e8. [Google Scholar] [CrossRef]
- Stankey, C.T.; Bourges, C.; Haag, L.M.; Turner-Stokes, T.; Piedade, A.P.; Palmer-Jones, C.; Papa, I.; dos Santos, M.S.; Zhang, Q.; Cameron, A.J.; et al. A disease-associated gene desert directs macrophage inflammation through ETS2. Nature 2024, 630, 447–456. [Google Scholar] [CrossRef]
- Gudiño, V.; Bartolomé-Casado, R.; Salas, A. Single-cell omics in inflammatory bowel disease: Recent insights and future clinical applications. Gut 2025, 74, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Kelley, J.; Amundsen, T.; Coggins, K.; Edwards, A.; Johnson, C.M. Medical and social determinants of health as predictors of adverse outcomes in patients with inflammatory bowel disease. Bayl. Univ. Med. Cent. Proc. 2023, 36, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Acera, M.; Patankar, J.V.; Erkert, L.; Cineus, R.; Gamez-Belmonte, R.; Leupold, T.; Bubeck, M.; Bao, L.-L.; Dinkel, M.; Wang, R.; et al. Integrated multimodel analysis of intestinal inflammation exposes key molecular features of preclinical and clinical IBD. Gut 2025, 74, 1602–1615. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Comparative graphic of Geboes score in the histologic healing phase (left) and in the active phase (right).
Figure 2.
Comparative graphic of Geboes score in the histologic healing phase (left) and in the active phase (right).
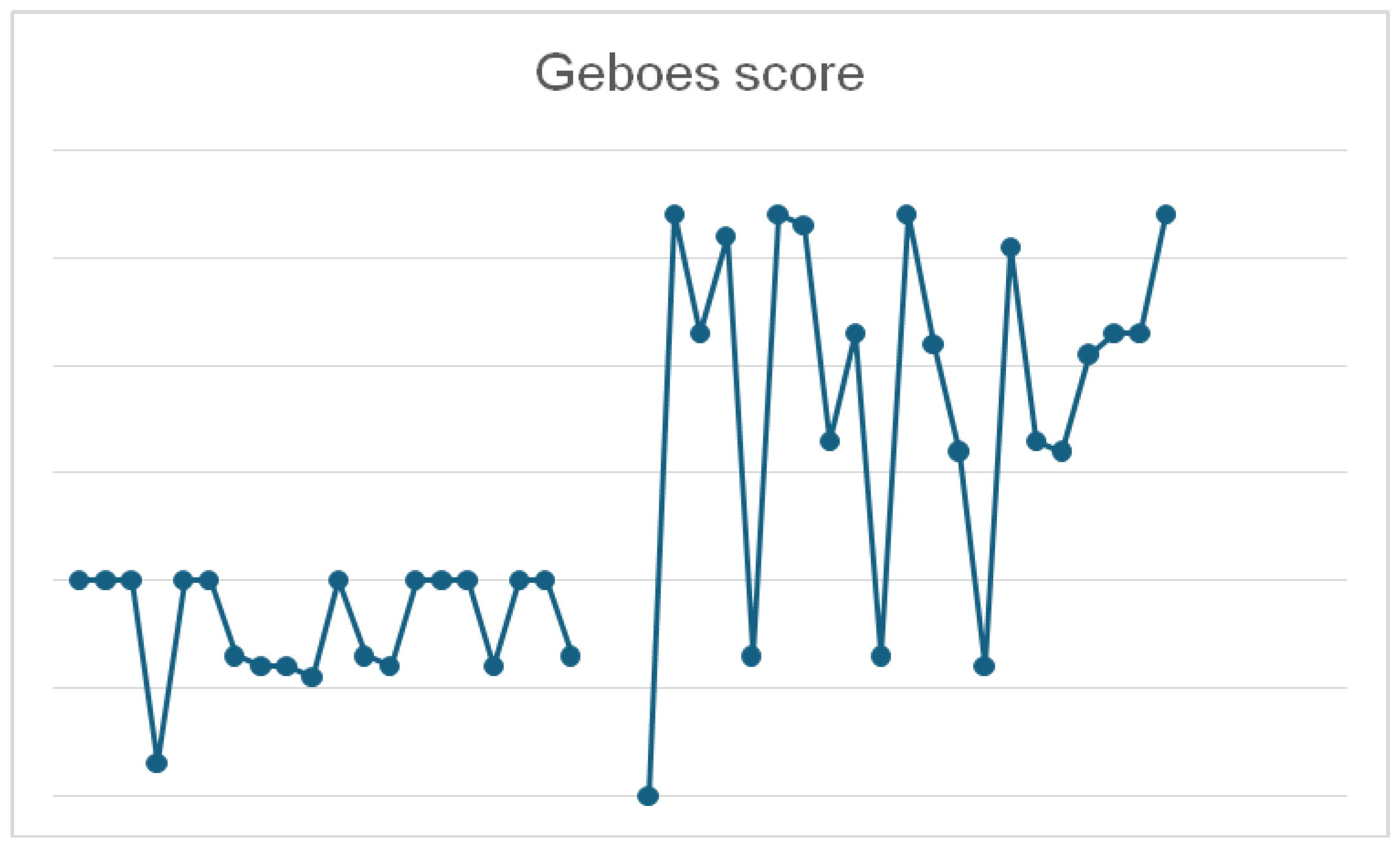
Figure 3.
CD4 (left) and CD8 (right) immunostain in a patient in histologic healing phase. Note rare positive cells in both epithelial and stromal compartments.
Figure 3.
CD4 (left) and CD8 (right) immunostain in a patient in histologic healing phase. Note rare positive cells in both epithelial and stromal compartments.
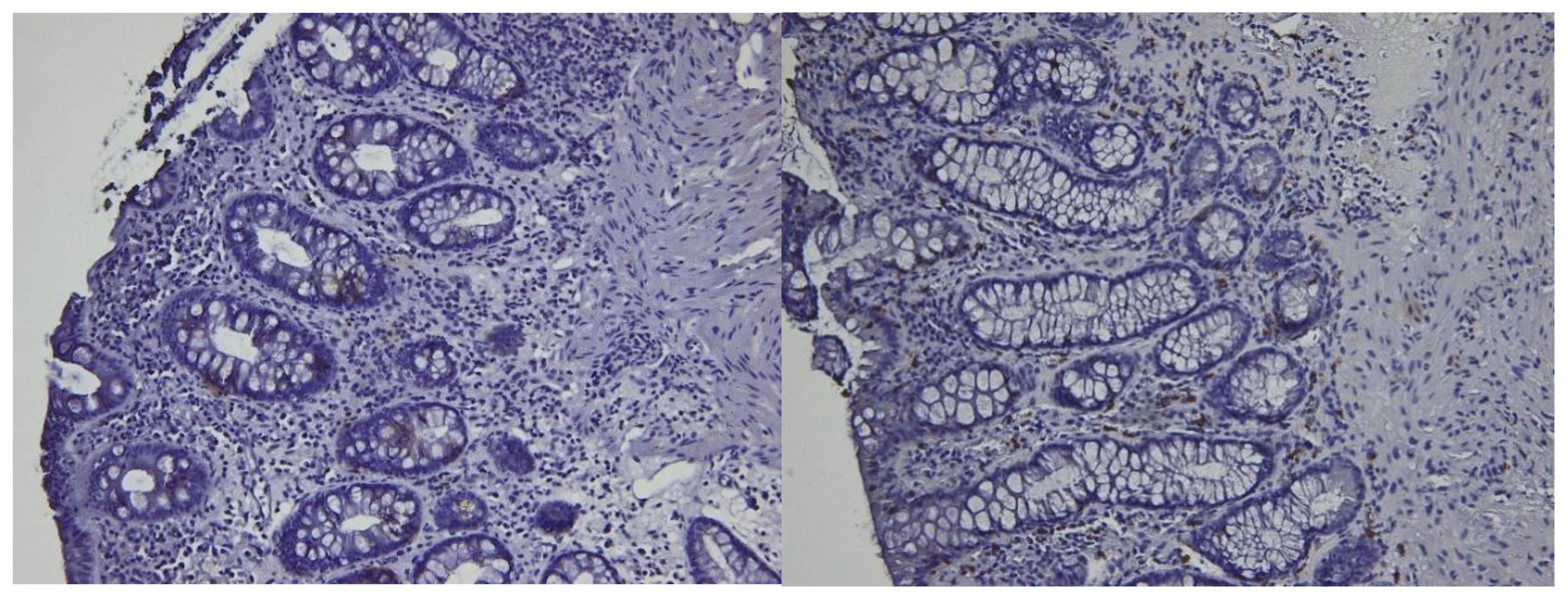
Figure 4.
CD4 (left) and CD8 (right) immunostain in the same patient with active disease. Note more frequent positive cells in both epithelial and stromal compartments.
Figure 4.
CD4 (left) and CD8 (right) immunostain in the same patient with active disease. Note more frequent positive cells in both epithelial and stromal compartments.
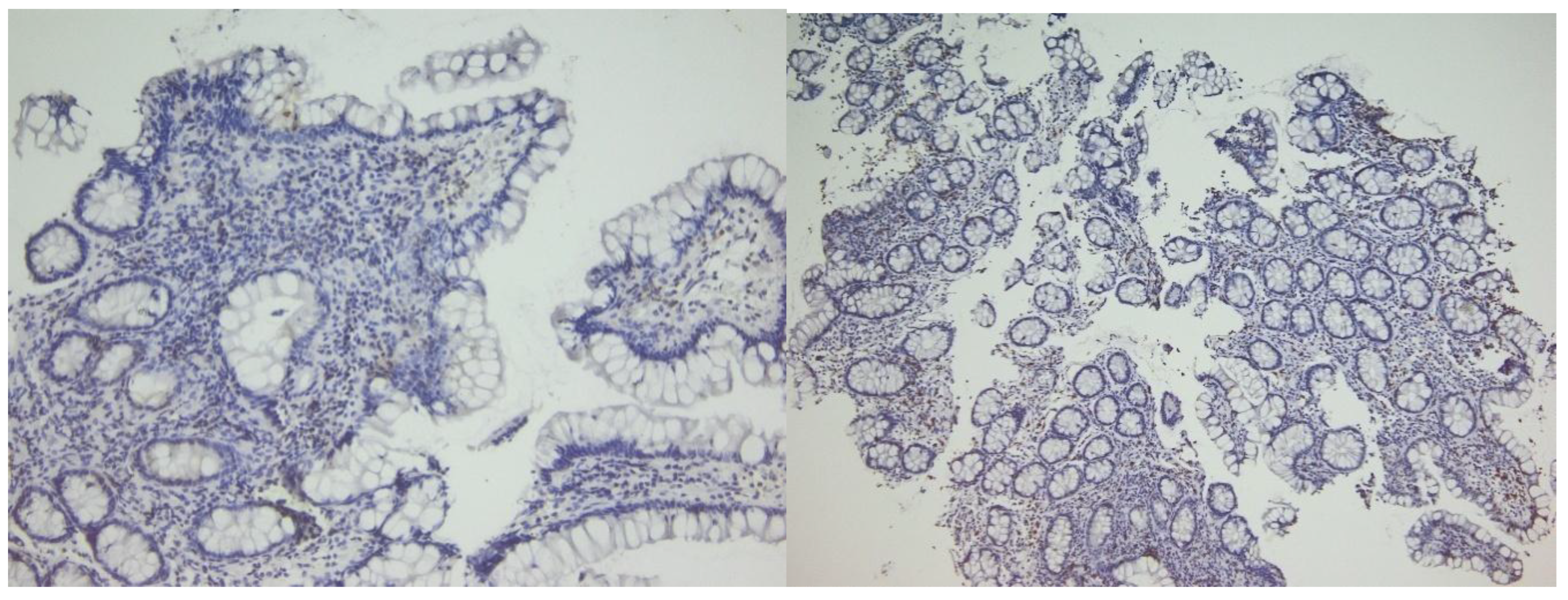
Figure 5.
Correlative analysis between CD4+, CD8+, and IL-6 expression patterns in both active and healing phases.
Figure 5.
Correlative analysis between CD4+, CD8+, and IL-6 expression patterns in both active and healing phases.
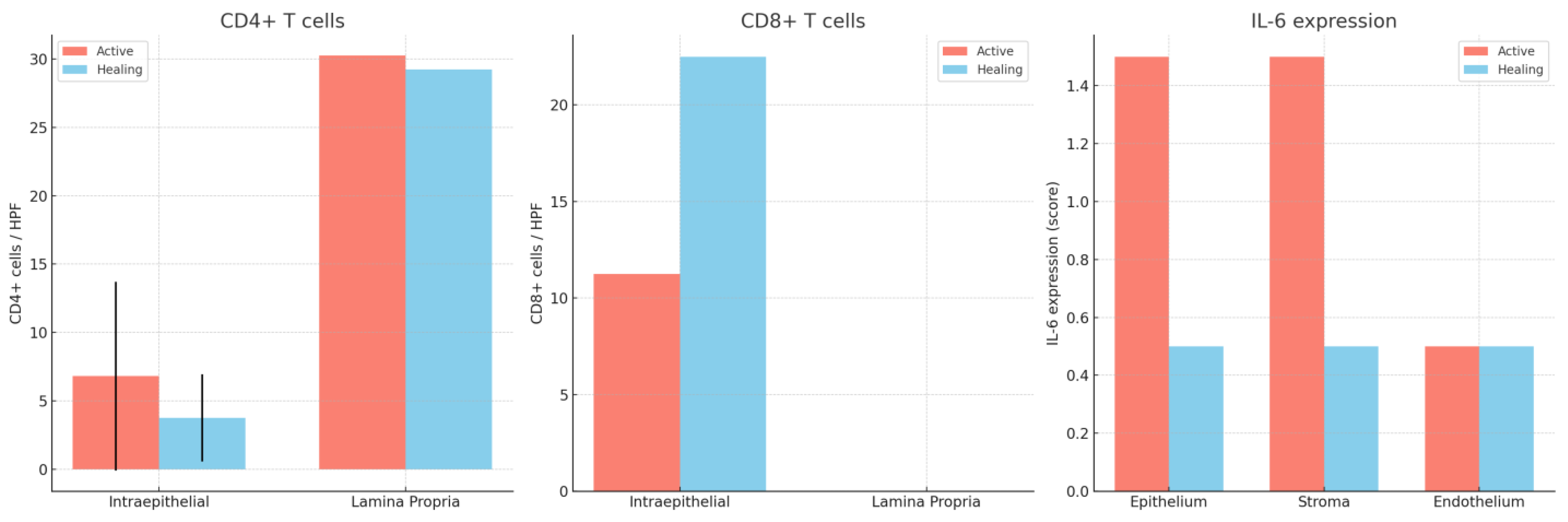
Figure 6.
Histologic healing in a patient with ulcerative colitis. Hematoxylin-eozin 100× (left) and 200× (right). Note the changes in cryptic architecture, but no inflammation, erosion or epithelial regenerative changes can be identified.
Figure 6.
Histologic healing in a patient with ulcerative colitis. Hematoxylin-eozin 100× (left) and 200× (right). Note the changes in cryptic architecture, but no inflammation, erosion or epithelial regenerative changes can be identified.
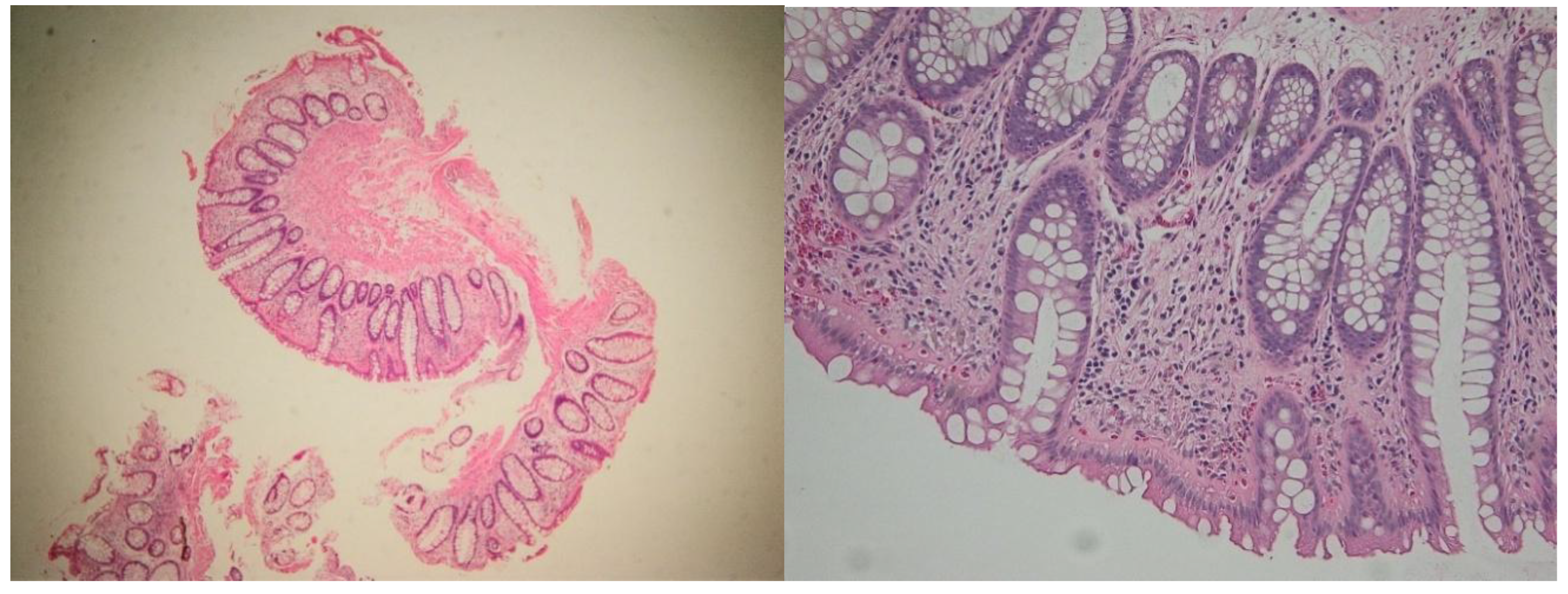
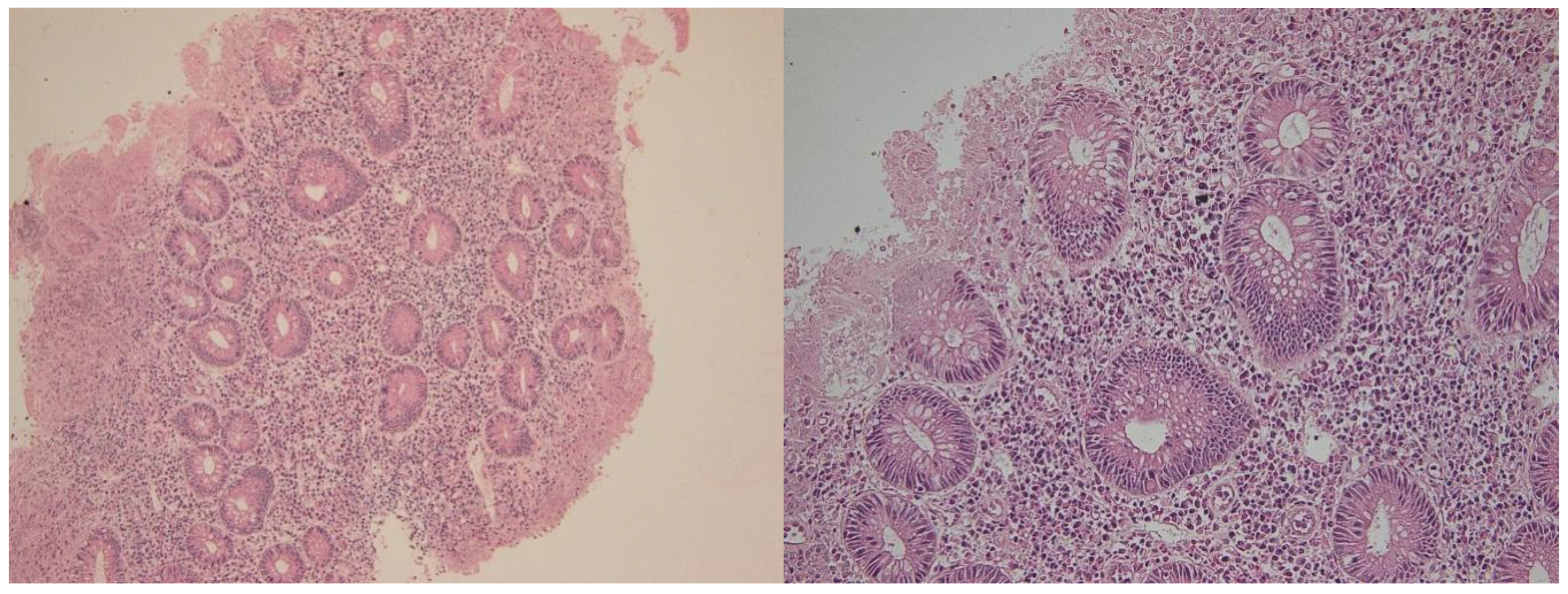
Figure 7.
Samples from the same patient as above, but in the active phase. Note the severe inflammation, with erosions and significant regenerative epithelial changes. Hematoxylin-eosin 100× (left) and 400× (right).
Figure 7.
Samples from the same patient as above, but in the active phase. Note the severe inflammation, with erosions and significant regenerative epithelial changes. Hematoxylin-eosin 100× (left) and 400× (right).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



