Submitted:
07 November 2025
Posted:
10 November 2025
You are already at the latest version
Abstract
Background: Systemic lupus erythematosus is a multi-faceted auto-immune disease. Growing evidence points to gut permeability and microbiota as key players in the devel-opment of the disease. Cinnamomum cassia is gaining attention as a potential modifier of the gut and liver health. We aim in this study to explore the effect of cinnamon on key ele-ments of gut-liver axis in imiquimod-induced lupus. Methods: Female C57BL/6J mice were stratified into five experimental groups: sham, sham plus cinnamon, lupus, lupus with cinnamon treatment, and lupus with pre- and post-induction cinnamon treatment. Lupus was induced through application of 1.25 mg of 5% imiquimod cream to the right ear, three times per week over six weeks. Cinnamomum cassia was given orally at 200 mg/kg, five days weekly. High-Throughput Sequencing of Bacterial 16S rRNA Gene was used on fecal samples with subsequent bioinformatic analysis of microbiota. Western blot and antibody array were used to measure E.coli translocation, and hepatic inflammatory, oxidative and apoptotic markers. Results: Cinnamon treatment mitigated the trend toward a lower Fir-micutes/Bacteroidota ratio in the lupus mice. While not statistically significant, cinnamon also led to a decrease in Lachnospiraceae abundance. Interestingly, sham mice given cin-namon had more Lactobacillus and Limosilactobacillus. Furthermore, cinnamon effectively reversed the increase of E.coli protein in the liver and normalized the enhanced expression of TLR-7, p-NFκB/NFκB, SOD1 and SOD2 induced by lupus. Conclusions: Cinnamomum cassia modulates key players of the gut-liver axis in lupus, offering a new perspective on lupus pathogenesis and potential nutritional interventions.
Keywords:
lupus
; microbiota
; cinnamon
; liver
; oxidative
1. Introduction
Systemic lupus erythematosus is the prototype of auto-immune diseases, with multiple organ damage. Its pathogenesis is complex and integrates many immunologic, metabolic and oxidative pathways [1]. Growing evidence points to gut permeability and microbiota as key players in the initiation and development of the disease, with a gut-liver axis dynamics characterized by bacterial translocation and activation of TLR4-NFκB in the liver [2].
The human gut microbiota is a complex ecosystem with more than one hundred trillion different microbial organisms [3]. Within this diverse microbial community, more than one thousand bacterial species inhabit the human gut, belonging mostly to four phyla—Firmicutes, Bacteroidota, Proteobacteria and Actinobacterita. Bacteroidota and Firmicutes dominate the gut microbiota in healthy adults, maintaining a symbiotic relationship with the human host [4]. This ecosystem connects environmental factors with the immune effectors and modulate the immunologic response. When dysbiosis with gut permeability dysfunction occurs, inflammation and immune system sensitization may follow, leading to chronic inflammation and autoimmune diseases [5]. Dietary modification, antibiotics, probiotics supplementation, and fecal microbiota transplantation are emerging as new paradigms to treat or prevent autoimmune or chronic inflammatory diseases.
Cinnamomum cassia, a worldwide used spice, rich in bioactive phytochemicals, is gaining attention as a potential nutritional modifier of the gut microbial structure and gut barrier function [6]. Beneficial effects of cinnamon in lupus have been suggested by recent studies [7] but has never been linked to modulating dysbiosis and liver inflammation in experimental lupus models.
Therefore, this study aims to assess the cinnamon supplementation effect on gut microbiome, activation of hepatic inflammatory pathways and liver oxidative stress in imiquimod-induced lupus in mice.
2. Materials and Methods
2.1. Ethical Statement
The study was conducted with the highest ethical standards for animal care. It was approved by the Ethical Committee of Saint Joseph University (CEHDF 1762, 2 December 2020). The research followed established animal care protocols, including: the Guiding Principles in the Care and Use of Animals by the American Physiological Society, the NIH’s “Guide for the Care and Use of Laboratory Animals” (NIH Publication no. 85–23, revised 1996), and the European Parliament Directive 2010/63 EU, which ensures ethical and humane treatment of animals.
2.2. Animals and Study Groups
Fifty adult female C57BL/6J mice were used in this study. Animals were kept in an environment with a constant temperature of 25 °C and humidity of 50 ± 5%, and they experienced a 12:12 h light–dark cycle. They had access to standard rodent food and tap water ad libitum. Lupus was induced by skin application on the right ear with 1.25 mg of 5% imiquimod cream three times per week for six weeks. Cinnamomum cassia powder (Solgar, Leonia, NJ, USA, powder of cinnamon bark and bark extract) was orally administered at 200 mg/kg/day to each animal individually on a small piece of chow slightly moistened with water to retain the cinnamon powder. Furthermore, the experimenter verified each time that the mouse had completely ingested this piece of chow. Accordingly, the animals were divided into five groups of ten mice per group: Sham, Sham treated with cinnamon (Sham Cinna), Lupus, lupus treated with cinnamon (Lupus Cinna or LC), and lupus treated with cinnamon starting two weeks before and continuously after induction ( Cinna Lupus Cinna or CLC). At the end of the protocol, the animals were anesthetized with a combination of ketamine (75 mg/kg; Interchemie, Waalre, The Netherlands) and xylazine (10 mg/kg; RotexMedica, Trittau, Germany). Once they showed no response to toe pinching, indicating deep anesthesia, they were euthanized for subsequent tissue collection.
2.4. Extraction and Purification of Total DNA from Feces
Fecal samples were collected (100 mg to 150 mg) from rectal content to investigate changes in GM composition at the species level. It was notable that all mice, irrespective of the study group, were sacrificed at the same time of day. DNA extraction was performed using the Zymo Quick-DNA™ Fecal/Soil Microbe Miniprep (Zymo Research, Irvine, CA, USA) according to the manufacturer’s instructions. The bead-beating step was achieved using a homogenizer (Biospec, Bartlesville, OK, USA) twice for 2 min. The concentration and quality of DNA was assessed by Nanodrop D100 Spectrophotometer (Nanodrop Technology, Wilmington, DE, USA) and measured using a Qubit dsDNA BR Assay Kit with a Qubit 4.0 Fluorometer (Thermo Fisher Scientific, Inc.), following the manufacturer's instructions.
2.5. High-Throughput Sequencing of Bacterial 16S rRNA Gene
The microbial community was assessed via the high-throughput sequencing the V3-V4 region of the bacterial 16S rRNA gene. We used for library preparation the Zymo Research Quick-16S™ NGS Library Prep Kit (Zymo Research, USA) according to manufacturer’s protocol. The resulting PCR products were purified and loaded onto Illumina MiSeq Reagent Kit v3 (San Diego, CA, USA) and sequenced on an Illumina MiSeq instrument with 2 × 300 paired end according to the manufacturer’s instructions. The quality of the run was checked internally using PhiX, and then each pair-end sequence was assigned to its samples with the help of the previously integrated index. Each pair-end sequence was assembled using Flash software v1.2.6 (Magoc 2011) using an at least 10 bp overlap between the forward and reverse sequences, allowing 10% mismatch. The lack of contamination was checked with a negative control during the PCR, using water as a template. The quality of the stitching procedure was controlled using the Zymobiomics Microbial community DNA Standard with 8 bacterial species that were run in parallel to the current samples.
2.6. Bioinformatics Analysis
Sequences were analyzed and normalized with the pipeline FROGS (Find Rapidly Operational Taxonomic Units (OTUs) with Galaxy Solution) [8]. PCR primers were removed, and sequences with sequencing errors in the primers were excluded. Reads were clustered into OTUs using the Swarm clustering method. Chimeras were removed, and 988 OTUs were assigned at different taxonomic levels (from phylum to species) using the RDP classifier and NCBI Blast+ on the 16S SILVA 138.1 database.
A total of 81,439 reads were randomly selected for each sample to normalize the data. The sequences were aligned using Clustal Omega 1.1.0 via the profile alignment option in Sea View 4.5 [9]. Neighbor joining trees, as well as maximum-likelihood trees using PhyML 3.1, were built to assess identifications [10].
2.6. Western Blot and Antibody Array
Liver tissue was homogenized and lysed in an assay lysis buffer composed of 150 mM NaCl, 50 mM Tris-OH (pH 7.5), 95 mM EDTA, and 0.5% Triton X-100, supplemented with protease and phosphatase inhibitors to extract total protein content. Protein concentrations were determined using the Bradford protein assay (Bio-Rad, Marnes-la-Coquette, France). Samples were then mixed with Laemmli loading buffer (Bio-Rad) containing 10% β-mercaptoethanol (Sigma-Aldrich) and incubated at 37 °C for 20 minutes for denaturation. Proteins were separated on 12% SDS-PAGE gels and transferred to polyvinylidene fluoride (PVDF) membranes (Bio-Rad). Membranes were blocked in TBS–Tween buffer containing 5% BSA, followed by overnight incubation at 4 °C with the appropriate primary antibodies: TLR4 (1/1000; ab22048; Abcam, Cambridge, UK ), TLR7 (1/1000; ab24184; Abcam, Cambridge, UK), NFkB (1/1000; 8242; Abcam, Cambridge, UK ) and p-NFkB (1/1000; 3033; Abcam, Cambridge, UK).
After being washed with TBS–Tween, the membranes were incubated for one hour at room temperature with goat anti-rabbit and anti-mouse antibodies (1/3000, Bio-Rad Laboratories) and rabbit anti-Goat (1/5000. ab6741; Abcam, Cambridge, UK) as secondary antibodies.
To perform the antibody array, 5 µg of protein was immobilized onto a PVDF membrane using dot blot equipment (Cleaver Scientific, Rugby, UK). This was followed by a blocking step and incubation with the primary antibody. The primary antibodies were: anti-FOXO3 (NBP2-16521), anti-FOXO3 (pSer253) (NBP2-67521), anti-NRF2 (NBP1-32822), anti-NRF2 (pSer40) (NBP2-67465) (Novus Biologicals, Bio-Techne, MN, USA), NOS3 (sc-654), SOD1 (sc-101523), SOD2 (sc133134) (Santa Cruz Biotechnology, Dallas, TX, USA), NOS3 (Ser1177) (#9571) (Cell Signaling Technology, Danvers, MA, USA), anti-Caspase 3 (ab13847; Abcam, Cambridge, UK), anti-Caspase 8 (ab108333; Abcam, Cambridge, UK), anti-BID (ab62469; Abcam, Cambridge, UK), t-BID (ab10640; Abcam, Cambridge, UK) and E.coli protein (1/1000; ab25823; Abcam, Cambridge, UK).
The same secondary antibodies used in the Western blot experiments were also used here.
Both Western blots and antibody arrays were visualized using enhanced chemiluminescence. Signal detection was carried out with an Omega Lum G imaging system (Aplegen, Gel Company, San Francisco, CA, USA), which is equipped with a CCD camera. Quantification of the signals was then performed using Licor Image Studio Lite version 5.2. Three western blots were analyzed for each condition.
2.7. Statistical Analysis
The gut microbiota of the mice in all groups was analyzed using high-throughput sequencing (average number of reads ± SEM = 152,411 ± 47828). Microbial diversity analyses were performed by clustering sequence tags into groups of defined sequence variation. α-Diversity measurements (observed OTUs, Chao 1, Shannon diversity index or SDI and inverted Simpson index) and β-diversity measurements (Jaccard, Bray–Curtis, UniFrac and weighted UniFrac) were analyzed using a blocked analysis of variance. The relative abundance of bacteria was compared with a MULTINOVA using the Jaccard and unweighted UniFrac similarity measures to construct distance metrics using QIIME 2 (Quantitative Insights Into Microbial Ecology 2) v 2019.7. All analyses were conducted using the R programming language in FROGS and GraphPad Prism 9 where Ordinary one-way Anova and Student’s T tests were employed to determine statistical significance among experimental groups. A p value less than 0.05 was considered statistically significant.
3. Results
3.1. Gut Microbiota
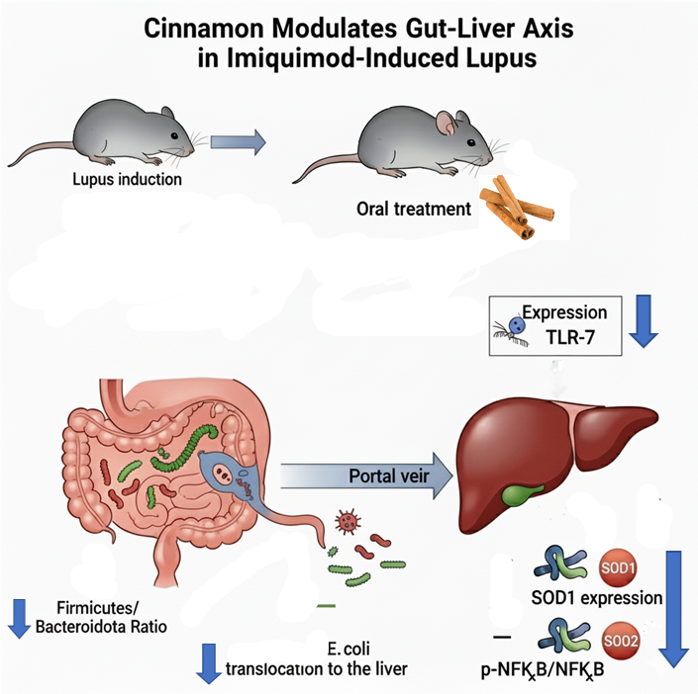
The mice gut microbiota was dominated by three phyla: Firmicutes, Bacteroidota and Actinobacterita (Figure 1). The Firmicutes/Bacteroidota ratio decreased in lupus mice but was restored in LC and CLC groups without reaching statistical significance (Figure 2).
At the family level, the gut microbiota was mainly colonized by Lactobacillaceae, Lachnospiraceae, Erysipelotrichaceae, Muribaculaceae, Bacteroidaceae, Prevotellaceae and Bifidobacteriaceae (Figure 3). Erysipelotrichaceae seems to be more represented in the control group (S) than in lupus group. Although statistically not significant, cinnamon supplementation seems to decrease Lachnospiraceae. In addition, the obtained results showed Lactobacillaceae was significantly lower in lupus mice in comparison with control group, while significantly increasing in control group supplemented with cinnamon (Figure 4). Eubacterium coprostanoligenes group was significantly decreased in lupus group. Saccharimonodaceae was significantly increased by cinnamon in control groups. Carnobacteriaceae, Planococcaceae, Aerococcaceae, Bacillaceae, Flavobacteriaceae, Corynebacteriaceae, Alcaligenaceae, Aeromonaceae, and Micrococcaceae were significantly increased in CLC group in comparison to lupus group.
At the genus level, Sham mice supplemented with Cinnamon were more colonized by Lactobacillus (27,9%) compared to non-supplemented mice (6,7%) and mice with induced lupus (13,6%) (Figure 5 and Table 1). The level of Lactobacillus was restored in mice with induced lupus supplemented with preventive cinnamon (Cinna Lupus Cinna 21,2%) (Figure 5 and Table 1). The genus Limosilactobacillus followed a very similar pattern to Lactobacillus (Figure 5 and Table 1).
Furthermore, Faecalibacterium was the highest in Sham mice when compared to other groups (Table 1). On the other hand, mice with induced lupus were more colonized by Staphylococcus and Bacteroides than mice without lupus or with induced lupus supplemented with cinnamon. In addition, Bifidobacterium was mainly detected in Sham mice regardless of lupus induction and cinnamon supplementation (Figure 6).
3.2. Effect of Lupus and Cinnamon on Microbial Diversity
Samples collected from the five groups were analyzed and compared in terms of α-diversity metrics (observed OTU richness, Chao-1, Shannon diversity index and inverted Simpson). All the metrics did not differ significantly between Sham mice, and mice supplemented with cinnamon with or without induced lupus (supplementary material).
These results show that the number of species did not variate significantly in supplemented mice or mice with induced lupus. However, cinnamon intake increased slightly bacterial richness and diversity when compared to Sham mice.
3.3. Effect of Lupus and Cinnamon on Microbial Communities
Following binning of the sequences into operational taxonomic units (OTUs) based on 97% sequence identity, comparisons were made using principal coordinates analysis (PCoA) based on weighted UniFrac distances (supplementary material). Each sample corresponding to microbial communities from sham mice with or without cinnamon supplementation and lupus induction clustered tightly and separated on the second principal axis (P2). Samples clustered tightly but did not separate on the first principal axis (P1).
Continued analysis of the microbial communities from these mice did not show the observed divergence of the lupus group when compared to controls, particularly on the first principal axis (P1). Unweighted UniFrac analysis and Jaccard analysis did not reveal any difference in β-diversity by inducing lupus and by supplementing with cinnamon, showing interindividual diversity (supplementary material).
3.4. KEGG Bioinformatic Analysis
Cinnamon significantly enhances glycolytic pathway in comparison to sham and lupus mice (supplementary figure 3). It also increases fatty acid degradation pathway in comparison to sham group. Lupus mice supplemented by cinnamon for a longer period (Cinna Lupus Cinna) exhibits significantly increased fatty acid biosynthesis versus lupus mice (Figure 7).
3.5. Escherichia coli Protein Expression in Liver Tissue:
Microarray studies reveal significantly increased Escherichia coli (E. coli) relative protein level in lupus mice liver when comparing to sham; this increase was significantly reversed in cinnamon supplemented lupus groups (Figure 8).
3.5. TLR-4, TLR-7 and NFκB in the Liver
Protein expression of TLR-7 and p-NFκB/ NFκB in the liver tissue of lupus mice was significantly higher than that of sham group. Preventive cinnamon supplementation (Cinna Lupus Cinna) significantly restored the expression of TLR-7 and p-NFκB/ NFκB. Protein expression of TLR4 was not significantly different between groups (Figure 9).
3.6. Oxidative Stress and Apoptosis Markers in the Liver:
SOD1/ β-actin and SOD2 / β-actin were significantly increased in liver tissue of lupus mice. Supplementation by cinnamon (LC and CLC) reduced significantly this increase (Figure 10). Expression of p-NOS3/p-NOS3, p-FOXO3a/p-FOXO3a, and p-NRF2/NRF2 did not differ significantly between groups. Furthermore, no significant change was found in apoptosis markers (Caspase 3, Caspase 8, t-BID/BID expression) (data not shown).
4. Discussion
The obtained results show, for the first time, that the cinnamon supplementation modulates intestinal microbiota, prevents E. coli liver translocation, calms down inflammatory pathways activation in liver tissue and exhibits ameliorative effect on hepatic oxidative stress in a imiquimod-induced lupus.
Imiquimod-induced lupus is a well-established and widely accepted model for inducing systemic autoimmunity that mirrors key features of human lupus. As demonstrated by Yokogawa et al. (2014), the topical application of IMQ in mice leads to the development of systemic manifestations, including splenomegaly, lymphadenopathy, circulating autoantibodies (anti-ssDNA and anti-Sm/RNP), and a systemic type I interferon signature [11].
Our study is an extension of our previous investigations of this model with documented renal and brain lupus-related lesions and dysfunction, and documented ameliorative effect of cinnamon [7,12].
The oral cinnamon dosing of 200 mg/kg was chosen as it is well-supported by prior research to be a non-toxic and therapeutically effective concentration for eliciting both metabolic and anti-inflammatory benefits [13,14]. This evidence-based approach provides a solid foundation for its use in our study. Besides, although the precise intake of cinnamon was not monitored by pharmacokinetic analysis of its active metabolites in plasma, the administration method aligns with common practices in nutritional studies and was chosen to minimize animal stress[15,16]. To note, that every mouse was observed for the complete consumption of the daily dose. Future research could incorporate such analyses to better define the dose-response relationship.
Since no single experimental model of lupus encompasses all aspects of the disease, various models exhibit distinct disease phenotypes. Likewise, the role of the microbiota in lupus seems to vary depending on the specific mouse model studied.
Firmicutes and Bacteroidota, the two dominant bacterial phyla in the gastrointestinal tract, have gained significant attention in recent years in various diseases. The Firmicutes/Bacteroidota (F/B) ratio is recognized as a crucial factor in maintaining intestinal homeostasis. An altered F/B ratio is considered a marker of dysbiosis, with an increase commonly linked to obesity and a decrease associated with inflammatory bowel disease [17].
A lower Firmicutes/Bacteroidota (F/B) ratio in MRL/lpr and NZBWF1 mice is linked to early disease initiation [18,19,20] . In human patients, lower F/B ratio is observed in lupus in comparison to healthy subjects[21,22]. Besides, active lupus patients show lower F/B ratio [23,24]. In our experimental conditions, a tendency towards decreased F/B ratio was observed in lupus mice but reversed by cinnamon supplementation especially in preventive group.
MRL/lpr mice have reduced Lactobacillaceae and increased Lachnospiraceae [6]. Lachnospiraceae is associated with more severe lupus symptoms in this spontaneous model [25]. In our study, Lactobacillaceae was significantly lower in lupus mice in comparison with control group, while significantly increasing in control group supplemented with cinnamon. Furthermore, cinnamon supplementation favors a shift toward decreased Lachnospiraceae in sham and lupus treated mice.
At the genus level, cinnamon supplementation seems to enhance Lactobacillus abundance; this is in line with prebiotic potential of cinnamon bark on Lactobacillus [26]. Several Lactobacilli are able to utilize phenolic acids (by decarboxylation and/or reduction reactions) and tolerate their presence much more than other members of the GM like Clostridium and Bacteroides members [27]. The relationship between systemic lupus erythematosus and Lactobacillus is complex and strain dependent. Reviews show that Lactobacillus genera can be either decreased or increased in SLE cohorts and animal models, and different Lactobacillus strains may have protective immunomodulatory effects or, in some contexts, exacerbate autoimmunity [28]. Our results suggest a clear timing effect: cinnamon given before lupus induction maintains higher Lactobacillus abundance (21.2%), which was decreased when cinnamon is given after lupus induction (5.4%). This fits a plausible mechanism seen in the literature: cinnamon can shape the baseline microbiota and mucosal barrier [29], promoting colonization resistance and supporting beneficial taxa when applied prophylactically [30], but its antimicrobial constituents or altered host environment post-disease induction may differ; while preventive cinnamon alters gut environment (pH, mucus, metabolites), favoring Lactobacillus expansion prior to immune challenge, post-induction cinnamon probably interacts with an inflamed mucosa or altered community and exerts antimicrobial effects that disproportionately reduce Lactobacillus in that pathological context.
Limosilactobacillus is a genus of lactic acid bacteria that is both thermophilic and heterofermentative; it was established in 2020 as an independent genus following its separation from the genus Lactobacillus [31]. Data on Limosilactobacillus reuteri effect in lupus is limited and may seem contradictory [32]. In NZB/WF1 mice pre-treatment with L. reuteri exhibits ameliorative effect on survival, cardiovascular lesions and liver proinflammatory cytokines IL-1 β, IL-6, and TNF-α [33,34]. In an opposite way, L. reuteri exacerbates lupus-like disease in TLR7-dependant model[35]; however the pathogenic effect of L. reuteri in the latter study was observed under specific-pathogen free or germ free conditions, and thus is not comparable to the effect of L. reuteri in the real complex intestinal environment. In our study, Limosilactobacillus was found more abundant in Sham mice supplemented with Cinnamon than in non-supplemented mice and mice with induced lupus. Identifying Lactobacillus and Limosilactobacillus at species or strain level by shotgun metagenomics or species-specific qPCR would be useful to clarify these heterogeneous results across studies, and to determine whether the expanded taxa are immunoregulatory strains (e.g., L. rhamnosus, L. reuteri).
On the other hand, Staphylococcus and Bacteroides were increased in lupus group in our study similarly to other human and animal studies [36].
Our experimental model did not reveal changes in alpha and beta diversity attributed to lupus induction or cinnamon supplementation. While some authors documented significant diversity shifts in imiquimod induced model [18,37], others did not find differences in the ecological parameters in this same model [18]. Sample size, environmental variability and other factors may contribute to this discrepancy.
Cinnamon effect on intestinal microbiota diversity is heterogeneous across studies depending on formulation, specific extraction and the animal or clinical conditions: while cinnamon oil and cinnamaldehyde microcapsules modulate diversity and richness of intestinal microbiota in mice [38,39] and cinnamon water extract increases alpha diversity in humans with diarrhea [40], cinnamaldehyde has not shown impact on alpha and beta diversity in early weaned rats [30] and consumption of dietary polyphenols (including cinnamon compounds) in humans does not modify diversity measures [41]. In our study, cinnamon supplementation did not result in diversity modification. This suggest that cinnamon's prebiotic effects might be more targeted towards specific microbial taxa rather than causing a broad modification in the richness of the gut microbiota. Future studies with a larger sample size may be needed to verify these findings.
Cinnamon effect on gut microbiota was reflected also by metabolic pathways modification in KEGG bioinformatic analysis. Increased glycolytic pathway may indicate positive impact of cinnamon on bacterial populations relying heavily on glycolysis, as Lactobacillus [42]. Effect on fatty acid degradation may be related to cinnamon's favoring of short-chain fatty acid-producing bacteria [43]. Increasing fatty acid biosynthesis in lupus mice supplemented by cinnamon for a long period is more intriguing and necessitates further investigation in future studies.
These intestinal microbiota modifications related to lupus and cinnamon supplementation were accompanied by alteration in gut-liver axis key components. We previously demonstrated gut permeability dysfunction with liver E. coli translocation and increased TLR4, TLR7, and pNFκB/NFκB liver expression in imiquimod-induced lupus [2]. Our study's primary objective was not to demonstrate that bacterial translocation causes or exacerbates lupus; this link has already been compellingly established in landmark studies that identified Enterococcus gallinarum translocation as a trigger for autoimmunity in genetically predisposed mice [44]. Instead, our goal was to leverage the validated model of systemic inflammation induced by imiquimod to investigate the effects of cinnamon on the gut-liver axis. Moreover, our previous work showed that intestinal tight junctions’ expression is decreased in this model and rescued by cinnamon supplementation [37]. Our present study confirms the presence of a gut-liver axis in this lupus model and adds new data on the protective effect of cinnamon on gut permeability and liver activation of TLR7 and NFκB. This consolidates the beneficial effect of cinnamon on gut barrier function observed in other metabolic or inflammatory conditions [30]. The reduction in bacterial translocation cannot be solely attributed to a direct effect of specific taxa modification (like Lactobacillus) by cinnamon. Instead, it is likely the result of a multifaceted mechanism involving both global shifts in the microbiota and the direct reinforcement of the gut epithelial barrier by cinnamon
Very few studies investigated liver specific oxidative stress in lupus. Mitochondrial dysfunction was described in lupus-prone mice as an early event [45]. Our microarray screening of key antioxidant markers in the liver revealed increased SOD1 and SOD2 expression in lupus mice. Upregulation of these enzymes may reflect compensatory mechanisms of coping with high oxidative stress [46,47]. Reversing of this increased expression with cinnamon may be a surrogate marker of the antioxidant effect of cinnamon and its modulating effect on TLR7-mediated oxidative stress [48].
Taken together, these data highlight the potential of cinnamon supplementation in modulating the gut microbiota in lupus and the associated hepatic inflammation and oxidative stress. Our study paves the way to a better understanding of gut-liver axis in lupus pathophysiology and translation to better prevention and management in human studies.
5. Conclusions
This section is not mandatory but can be added to the manuscript if the discussion is unusually long or complex.
Author Contributions
GM & TI: Conceptualization; Data curation; Formal analysis; Investigation; Methodology; Writing original draft. NF: Conceptualization; Investigation; Formal analysis; Funding acquisition; Supervision; Methodology; Visualization; Writing-Reviewing and Editing.
Funding
This work was supported by the Research Council of the Saint Joseph University - Faculty of Medicine. FM 382 & FM 419.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Ms Anna-Maria Baramili for technical support.
Conflicts of Interest
None.
References
- Arnaud, L.; Chasset, F.; Martin, T. Immunopathogenesis of Systemic Lupus Erythematosus: An Update. Autoimmun Rev 2024, 23, 103648. [CrossRef]
- Maalouly, G.; Hajal, J.; Noujeim, C.; Choueiry, M.; Nassereddine, H.; Smayra, V.; Saliba, Y.; Fares, N. New Insights in Gut-Liver Axis in Wild-Type Murine Imiquimod-Induced Lupus. Lupus 2021, 30, 926–936. [CrossRef]
- Honda, K.; Littman, D.R. The Microbiome in Infectious Disease and Inflammation. Annu Rev Immunol 2012, 30, 759–795. [CrossRef]
- Manos, J. The Human Microbiome in Disease and Pathology. APMIS 2022, 130, 690–705. [CrossRef]
- Yao, K.; Xie, Y.; Wang, J.; Lin, Y.; Chen, X.; Zhou, T. Gut Microbiota: A Newly Identified Environmental Factor in Systemic Lupus Erythematosus. Front Immunol 2023, 14, 1202850. [CrossRef]
- Zhang, K.; Ji, J.; Li, N.; Yin, Z.; Fan, G. Integrated Metabolomics and Gut Microbiome Analysis Reveals the Efficacy of a Phytochemical Constituent in the Management of Ulcerative Colitis. Mol Nutr Food Res 2024, 68, e2200578. [CrossRef]
- Maalouly, G.; Martin, C.-M.-A.; Baz, Y.; Saliba, Y.; Baramili, A.-M.; Fares, N. Antioxidant and Anti-Apoptotic Neuroprotective Effects of Cinnamon in Imiquimod-Induced Lupus. Antioxidants (Basel) 2024, 13, 880. [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol Biol Evol 2010, 27, 221–224. [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst Biol 2010, 59, 307–321. [CrossRef]
- Yokogawa, M.; Takaishi, M.; Nakajima, K.; Kamijima, R.; Fujimoto, C.; Kataoka, S.; Terada, Y.; Sano, S. Epicutaneous Application of Toll-like Receptor 7 Agonists Leads to Systemic Autoimmunity in Wild-Type Mice: A New Model of Systemic Lupus Erythematosus. Arthritis & Rheumatology (Hoboken, N.J.) 2014, 66, 694–706. [CrossRef]
- Maalouly, G.; Saliba, Y.; Hajal, J.; Zein-El-Din, A.; Fakhoury, L.; Najem, R.; Smayra, V.; Nassereddine, H.; Fares, N. Cinnamomum Cassia Alleviates Neuropsychiatric Lupus in a Murine Experimental Model. Nutrients 2025, 17, 1820. [CrossRef]
- Niazmand, S.; Mirzaei, M.; Hosseinian, S.; Khazdair, M.R.; Gowhari Shabgah, A.; Baghcheghi, Y.; Hedayati-Moghadam, M. The Effect of Cinnamomum Cassia Extract on Oxidative Stress in the Liver and Kidney of STZ-Induced Diabetic Rats. J Complement Integr Med 2022, 19, 311–321. [CrossRef]
- Lim, J.-O.; Kim, Y.H.; Lee, I.S.; Kim, W.-I.; Lee, S.-J.; Pak, S.-W.; Shin, I.-S.; Kim, T. Cinnamomum Cassia (L.) J.Presl Alleviates Allergic Responses in Asthmatic Mice via Suppression of MAPKs and MMP-9. Front Pharmacol 2022, 13, 906916. [CrossRef]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol Improves Mitochondrial Function and Protects against Metabolic Disease by Activating SIRT1 and PGC-1alpha. Cell 2006, 127, 1109–1122. [CrossRef]
- Shao, W.; Yu, Z.; Chiang, Y.; Yang, Y.; Chai, T.; Foltz, W.; Lu, H.; Fantus, I.G.; Jin, T. Curcumin Prevents High Fat Diet Induced Insulin Resistance and Obesity via Attenuating Lipogenesis in Liver and Inflammatory Pathway in Adipocytes. PLoS One 2012, 7, e28784. [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel Disease. Microorganisms 2020, 8, 1715. [CrossRef]
- de la Visitación, N.; Robles-Vera, I.; Toral, M.; Gómez-Guzmán, M.; Sánchez, M.; Moleón, J.; González-Correa, C.; Martín-Morales, N.; O’Valle, F.; Jiménez, R.; et al. Gut Microbiota Contributes to the Development of Hypertension in a Genetic Mouse Model of Systemic Lupus Erythematosus. Br J Pharmacol 2021, 178, 3708–3729. [CrossRef]
- Robles-Vera, I.; Visitación, N.D.L.; Toral, M.; Sánchez, M.; Gómez-Guzmán, M.; O’valle, F.; Jiménez, R.; Duarte, J.; Romero, M. Toll-like Receptor 7-Driven Lupus Autoimmunity Induces Hypertension and Vascular Alterations in Mice. J Hypertens 2020, 38, 1322–1335. [CrossRef]
- Wang, H.; Wang, G.; Banerjee, N.; Liang, Y.; Du, X.; Boor, P.J.; Hoffman, K.L.; Khan, M.F. Aberrant Gut Microbiome Contributes to Intestinal Oxidative Stress, Barrier Dysfunction, Inflammation and Systemic Autoimmune Responses in MRL/Lpr Mice. Front Immunol 2021, 12, 651191. [CrossRef]
- He, Z.; Shao, T.; Li, H.; Xie, Z.; Wen, C. Alterations of the Gut Microbiome in Chinese Patients with Systemic Lupus Erythematosus. Gut Pathog 2016, 8, 64. [CrossRef]
- Toumi, E.; Goutorbe, B.; Plauzolles, A.; Bonnet, M.; Mezouar, S.; Militello, M.; Mege, J.-L.; Chiche, L.; Halfon, P. Gut Microbiota in Systemic Lupus Erythematosus Patients and Lupus Mouse Model: A Cross Species Comparative Analysis for Biomarker Discovery. Front Immunol 2022, 13, 943241. [CrossRef]
- Gerges, M.A.; Esmaeel, N.E.; Makram, W.K.; Sharaf, D.M.; Gebriel, M.G. Altered Profile of Fecal Microbiota in Newly Diagnosed Systemic Lupus Erythematosus Egyptian Patients. Int J Microbiol 2021, 2021, 9934533. [CrossRef]
- Widhani, A.; Djauzi, S.; Suyatna, F.D.; Dewi, B.E. Changes in Gut Microbiota and Systemic Inflammation after Synbiotic Supplementation in Patients with Systemic Lupus Erythematosus: A Randomized, Double-Blind, Placebo-Controlled Trial. Cells 2022, 11, 3419. [CrossRef]
- Zhang, H.; Liao, X.; Sparks, J.B.; Luo, X.M. Dynamics of Gut Microbiota in Autoimmune Lupus. Appl Environ Microbiol 2014, 80, 7551–7560. [CrossRef]
- De Giani, A.; Pagliari, S.; Zampolli, J.; Forcella, M.; Fusi, P.; Bruni, I.; Campone, L.; Di Gennaro, P. Characterization of the Biological Activities of a New Polyphenol-Rich Extract from Cinnamon Bark on a Probiotic Consortium and Its Action after Enzymatic and Microbial Fermentation on Colorectal Cell Lines. Foods 2022, 11, 3202. [CrossRef]
- Sánchez-Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Structure-Function Relationships of the Antibacterial Activity of Phenolic Acids and Their Metabolism by Lactic Acid Bacteria. J Appl Microbiol 2011, 111, 1176–1184. [CrossRef]
- Wang, W.; Fan, Y.; Wang, X. Lactobacillus: Friend or Foe for Systemic Lupus Erythematosus? Front Immunol 2022, 13, 883747. [CrossRef]
- Kim, M.S.; Kim, J.Y. Cinnamon Subcritical Water Extract Attenuates Intestinal Inflammation and Enhances Intestinal Tight Junction in a Caco-2 and RAW264.7 Co-Culture Model. Food Funct 2019, 10, 4350–4360. [CrossRef]
- Qi, L.; Mao, H.; Lu, X.; Shi, T.; Wang, J. Cinnamaldehyde Promotes the Intestinal Barrier Functions and Reshapes Gut Microbiome in Early Weaned Rats. Front Nutr 2021, 8, 748503. [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int J Syst Evol Microbiol 2020, 70, 2782–2858. [CrossRef]
- Luo, Z.; Chen, A.; Xie, A.; Liu, X.; Jiang, S.; Yu, R. Limosilactobacillus Reuteri in Immunomodulation: Molecular Mechanisms and Potential Applications. Front Immunol 2023, 14, 1228754. [CrossRef]
- Hsu, T.-C.; Huang, C.-Y.; Liu, C.-H.; Hsu, K.-C.; Chen, Y.-H.; Tzang, B.-S. Lactobacillus Paracasei GMNL-32, Lactobacillus Reuteri GMNL-89 and L. Reuteri GMNL-263 Ameliorate Hepatic Injuries in Lupus-Prone Mice. Br. J. Nutr. 2017, 117, 1066–1074. [CrossRef]
- Manirarora, J.N.; Kosiewicz, M.M.; Alard, P. Feeding Lactobacilli Impacts Lupus Progression in (NZBxNZW)F1 Lupus-Prone Mice by Enhancing Immunoregulation. Autoimmunity 2020, 53, 323–332. [CrossRef]
- Zegarra-Ruiz, D.F.; El Beidaq, A.; Iñiguez, A.J.; Lubrano Di Ricco, M.; Manfredo Vieira, S.; Ruff, W.E.; Mubiru, D.; Fine, R.L.; Sterpka, J.; Greiling, T.M.; et al. A Diet-Sensitive Commensal Lactobacillus Strain Mediates TLR7-Dependent Systemic Autoimmunity. Cell Host Microbe 2019, 25, 113-127.e6. [CrossRef]
- Mohd, R.; Chin, S.-F.; Shaharir, S.S.; Cham, Q.S. Involvement of Gut Microbiota in SLE and Lupus Nephritis. Biomedicines 2023, 11, 653. [CrossRef]
- Moleón, J.; González-Correa, C.; Miñano, S.; Robles-Vera, I.; de la Visitación, N.; Barranco, A.M.; Gómez-Guzmán, M.; Sánchez, M.; Riesco, P.; Guerra-Hernández, E.; et al. Protective Effect of Microbiota-Derived Short Chain Fatty Acids on Vascular Dysfunction in Mice with Systemic Lupus Erythematosus Induced by Toll like Receptor 7 Activation. Pharmacol Res 2023, 198, 106997. [CrossRef]
- Ma, T.; Tang, B.; Wang, Y.; Shen, M.; Ping, Y.; Wang, L.; Su, J. Cinnamon Oil Solid Self-Microemulsion Mediates Chronic Mild Stress-Induced Depression in Mice by Modulating Monoamine Neurotransmitters, Corticosterone, Inflammation Cytokines, and Intestinal Flora. Heliyon 2023, 9, e17125. [CrossRef]
- Xiao, Y.; Zhang, F.; Xu, H.; Yang, C.; Song, X.; Zhou, Y.; Zhou, X.; Liu, X.; Miao, J. Cinnamaldehyde Microcapsules Enhance Bioavailability and Regulate Intestinal Flora in Mice. Food Chem X 2022, 15, 100441. [CrossRef]
- Park, S.-Y.; Kim, Y.D.; Kim, M.S.; Kim, K.-T.; Kim, J.Y. Cinnamon (Cinnamomum Cassia) Water Extract Improves Diarrhea Symptoms by Changing the Gut Environment: A Randomized Controlled Trial. Food Funct 2023, 14, 1520–1529. [CrossRef]
- Vita, A.A.; Roberts, K.M.; Gundersen, A.; Farris, Y.; Zwickey, H.; Bradley, R.; Weir, T.L. Relationships between Habitual Polyphenol Consumption and Gut Microbiota in the INCLD Health Cohort. Nutrients 2024, 16, 773. [CrossRef]
- Wolfe, A.J. Glycolysis for Microbiome Generation. Microbiol Spectr 2015, 3. [CrossRef]
- Li, A.-L.; Ni, W.-W.; Zhang, Q.-M.; Li, Y.; Zhang, X.; Wu, H.-Y.; Du, P.; Hou, J.-C.; Zhang, Y. Effect of Cinnamon Essential Oil on Gut Microbiota in the Mouse Model of Dextran Sodium Sulfate-Induced Colitis. Microbiol Immunol 2020, 64, 23–32. [CrossRef]
- Manfredo Vieira, S.; Hiltensperger, M.; Kumar, V.; Zegarra-Ruiz, D.; Dehner, C.; Khan, N.; Costa, F.R.C.; Tiniakou, E.; Greiling, T.; Ruff, W.; et al. Translocation of a Gut Pathobiont Drives Autoimmunity in Mice and Humans. Science 2018, 359, 1156–1161. [CrossRef]
- Oaks, Z.; Winans, T.; Caza, T.; Fernandez, D.; Liu, Y.; Landas, S.K.; Banki, K.; Perl, A. Mitochondrial Dysfunction in the Liver and Antiphospholipid Antibody Production Precede Disease Onset and Respond to Rapamycin in Lupus-Prone Mice. Arthritis & Rheumatology (Hoboken, N.J.) 2016, 68, 2728–2739. [CrossRef]
- Flynn, J.M.; Melov, S. SOD2 in Mitochondrial Dysfunction and Neurodegeneration. Free Radic Biol Med 2013, 62, 4–12. [CrossRef]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several Lines of Antioxidant Defense against Oxidative Stress: Antioxidant Enzymes, Nanomaterials with Multiple Enzyme-Mimicking Activities, and Low-Molecular-Weight Antioxidants. Arch Toxicol 2024, 98, 1323–1367. [CrossRef]
- Lee, S.-C.; Wang, S.-Y.; Li, C.-C.; Liu, C.-T. Anti-Inflammatory Effect of Cinnamaldehyde and Linalool from the Leaf Essential Oil of Cinnamomum Osmophloeum Kanehira in Endotoxin-Induced Mice. J Food Drug Anal 2018, 26, 211–220. [CrossRef]
Figure 1.
Gut microbiota colonization comparison at the phylum level of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinnamon), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinnamon), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinnamon Lupus Cinnamon).
Figure 1.
Gut microbiota colonization comparison at the phylum level of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinnamon), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinnamon), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinnamon Lupus Cinnamon).
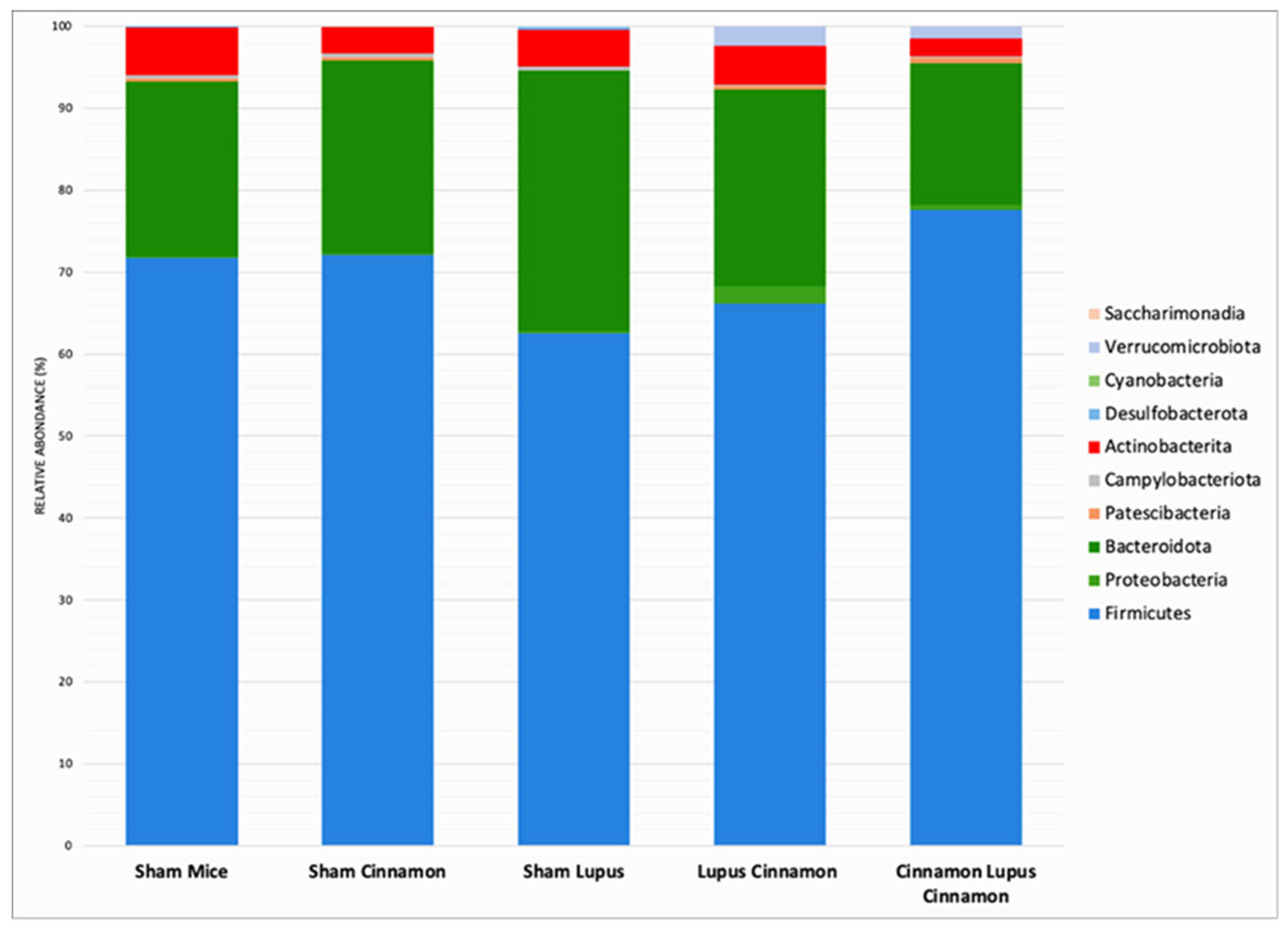
Figure 2.
Firmicutes (A) and Bacteroidota (B) abundance with Firmicutes/Bacteroidota ratio level (C) of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). No statistically difference was observed between groups (p>0.05).
Figure 2.
Firmicutes (A) and Bacteroidota (B) abundance with Firmicutes/Bacteroidota ratio level (C) of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). No statistically difference was observed between groups (p>0.05).
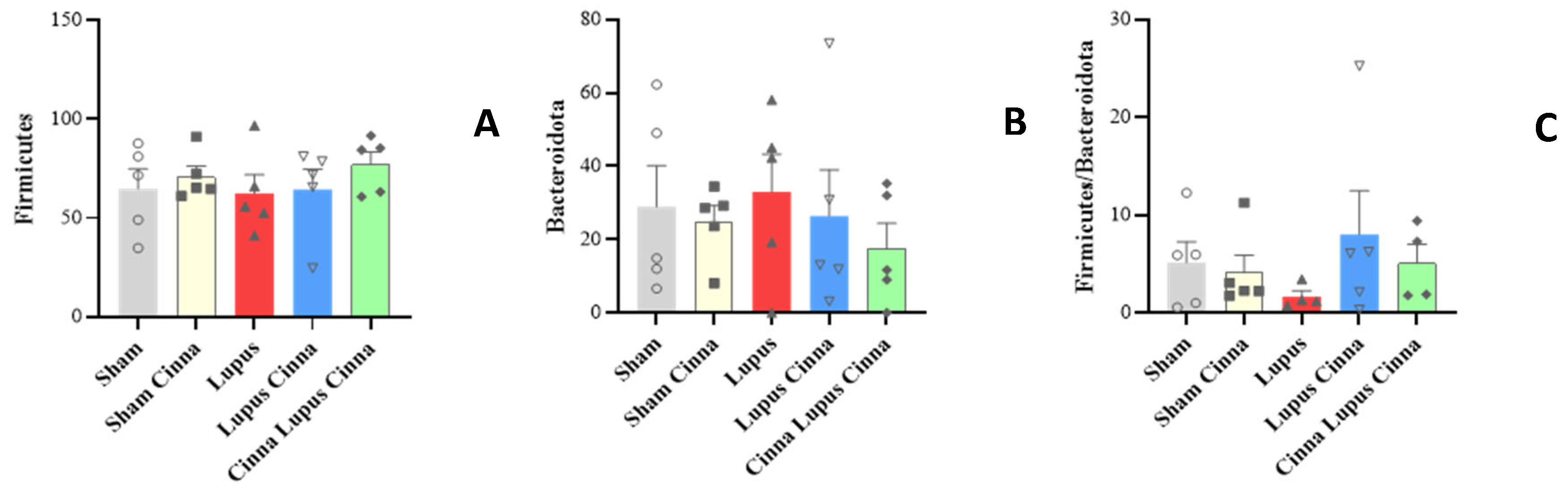
Figure 3.
Gut microbiota colonization comparison at the family level of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinnamon), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinnamon), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinnamon Lupus Cinnamon).
Figure 3.
Gut microbiota colonization comparison at the family level of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinnamon), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinnamon), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinnamon Lupus Cinnamon).
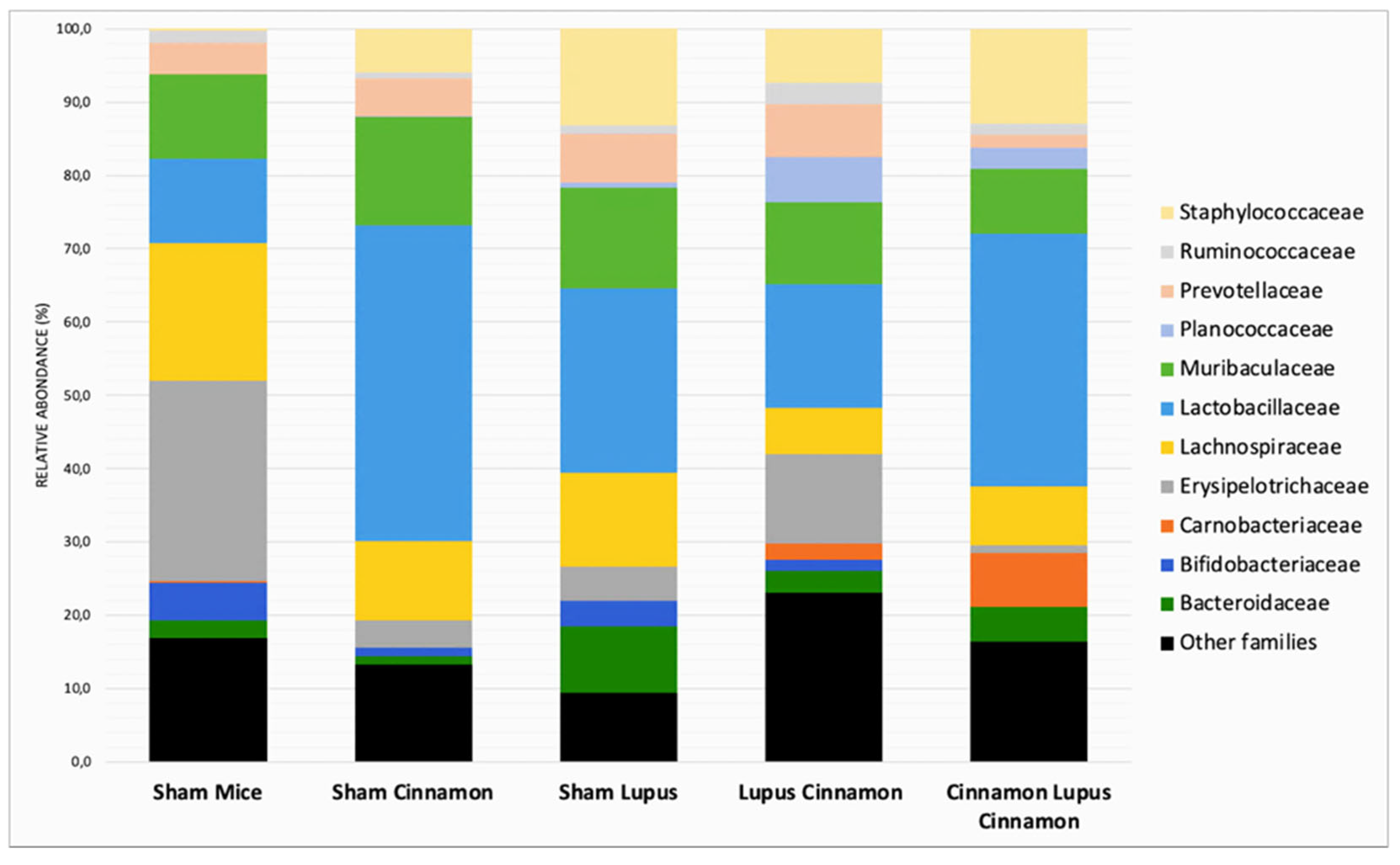
Figure 4.
Relative abundance at the family level in the different groups: normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). Only statistically significant differences are shown. *: p< 0.05.
Figure 4.
Relative abundance at the family level in the different groups: normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). Only statistically significant differences are shown. *: p< 0.05.
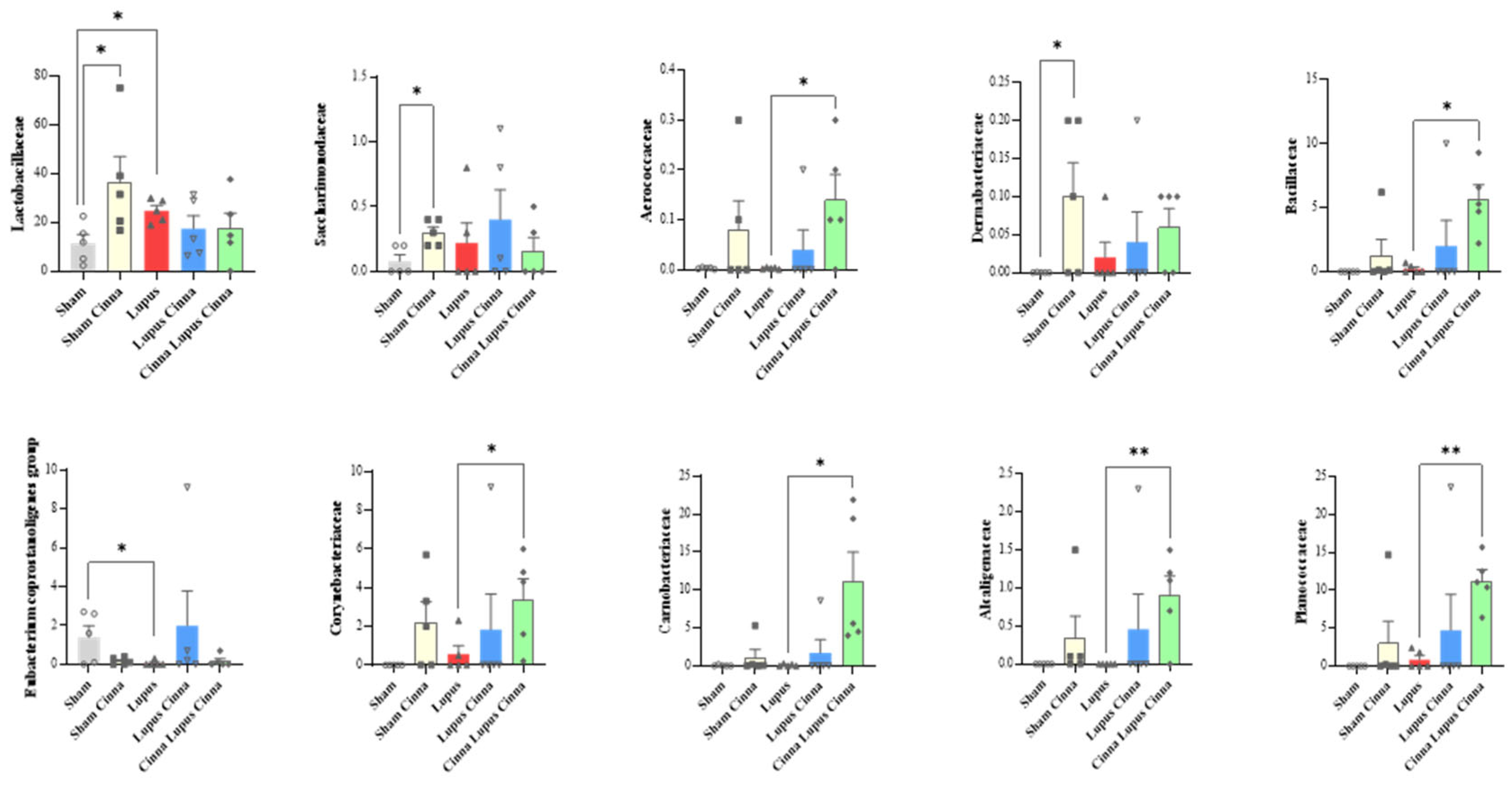
Figure 5.
Gut microbiota colonization comparison at the genus level of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinnamon), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinnamon), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinnamon Lupus Cinnamon).
Figure 5.
Gut microbiota colonization comparison at the genus level of normal mice (Sham Mice), mice treated with cinnamon (Sham Cinnamon), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinnamon), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinnamon Lupus Cinnamon).
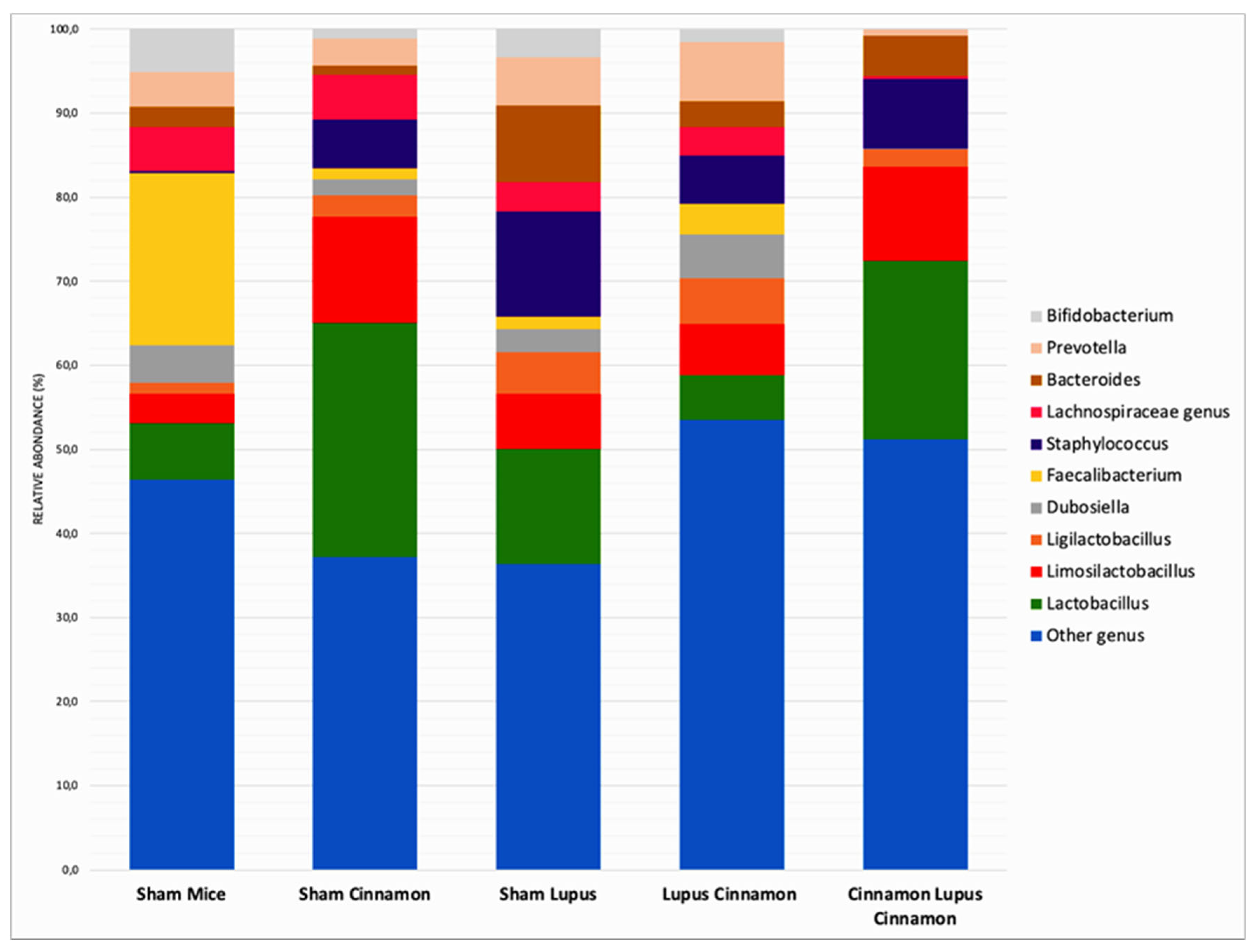
Figure 6.
Relative abundance at the genus level (A: Lactobacillus; B: Limosilactobacillus; C: Bacteroides) in the different groups: normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). Only statistically significant differences are shown. *: p< 0.05.
Figure 6.
Relative abundance at the genus level (A: Lactobacillus; B: Limosilactobacillus; C: Bacteroides) in the different groups: normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). Only statistically significant differences are shown. *: p< 0.05.
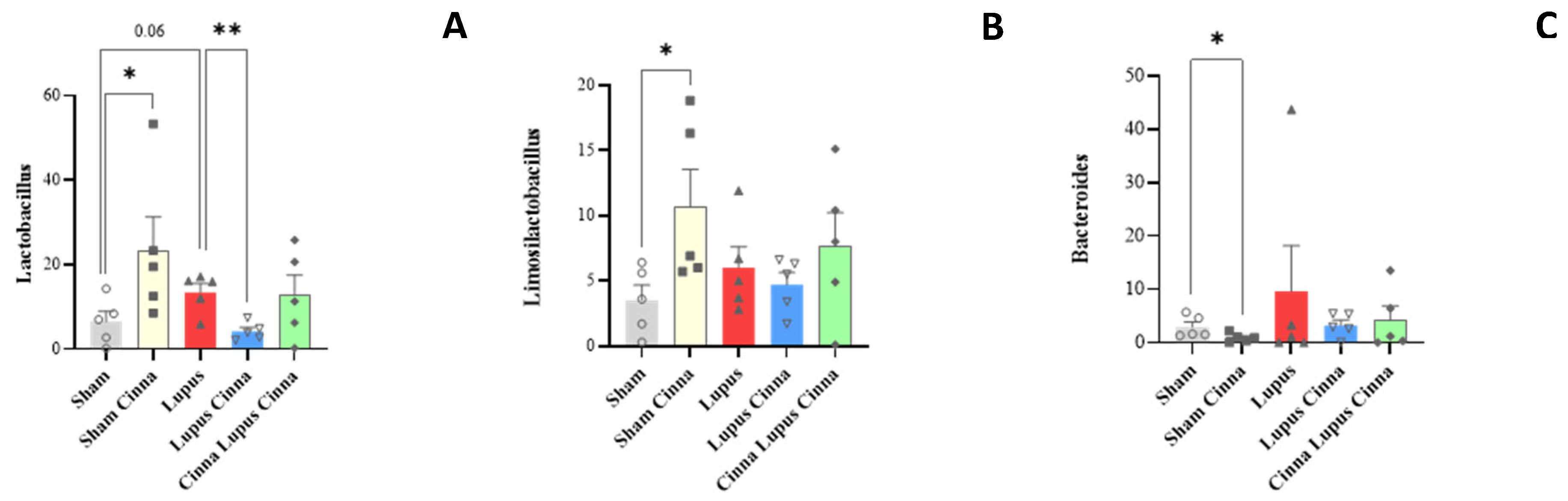
Figure 7.
Relative percentage of the pathway’s distribution KEGG (A: Glycolysis; B: Fatty acid biosynthesis; C: Fatty acid degradation) in the different groups: normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). Only statistically significant differences are shown. *: p< 0.05.
Figure 7.
Relative percentage of the pathway’s distribution KEGG (A: Glycolysis; B: Fatty acid biosynthesis; C: Fatty acid degradation) in the different groups: normal mice (Sham Mice), mice treated with cinnamon (Sham Cinna), mice with induced lupus (Sham Lupus), mice with induced lupus treated with cinnamon (Lupus Cinna), and mice supplemented with cinnamon prior to inducing lupus, then treated with cinnamon (Cinna Lupus Cinna). Only statistically significant differences are shown. *: p< 0.05.
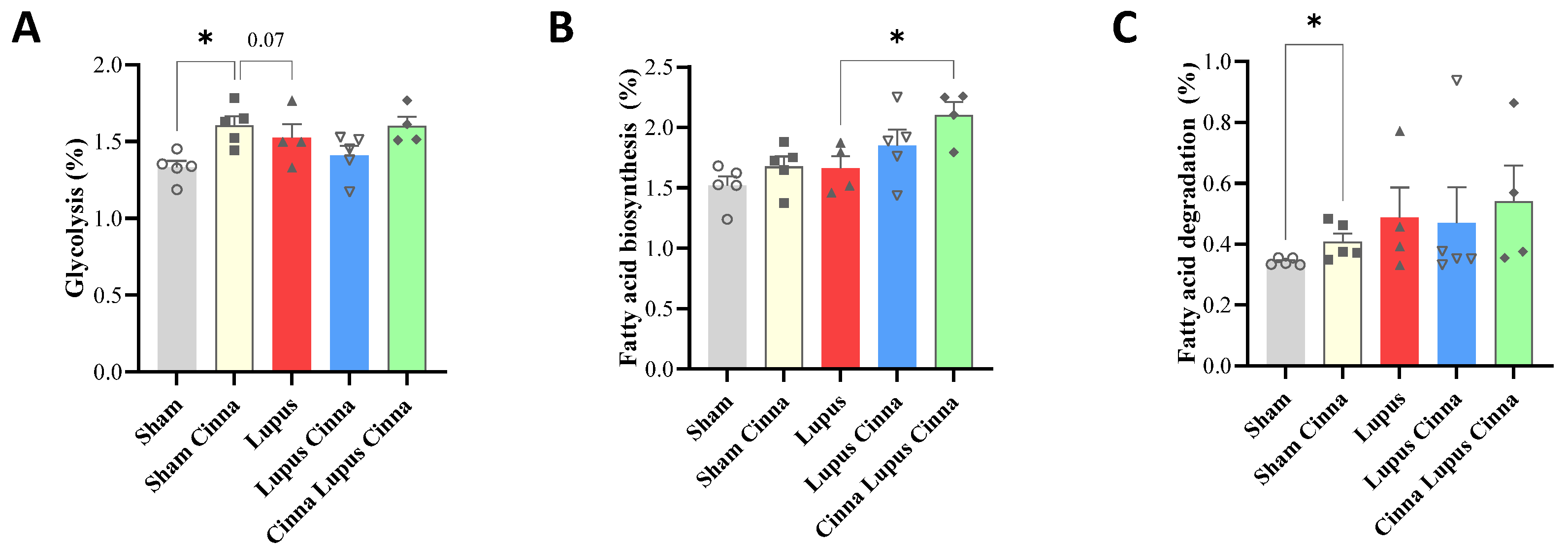
Figure 8.
Antibody array and quantification of E. coli protein with β-actin in liver tissue by microarray measurement in different groups. *: p< 0.05.
Figure 8.
Antibody array and quantification of E. coli protein with β-actin in liver tissue by microarray measurement in different groups. *: p< 0.05.

Figure 9.
Western blot and quantification of TLR4, TLR7 and p-NFκB / NFκB with β-actin in liver tissue of different groups. *: p< 0.05.
Figure 9.
Western blot and quantification of TLR4, TLR7 and p-NFκB / NFκB with β-actin in liver tissue of different groups. *: p< 0.05.
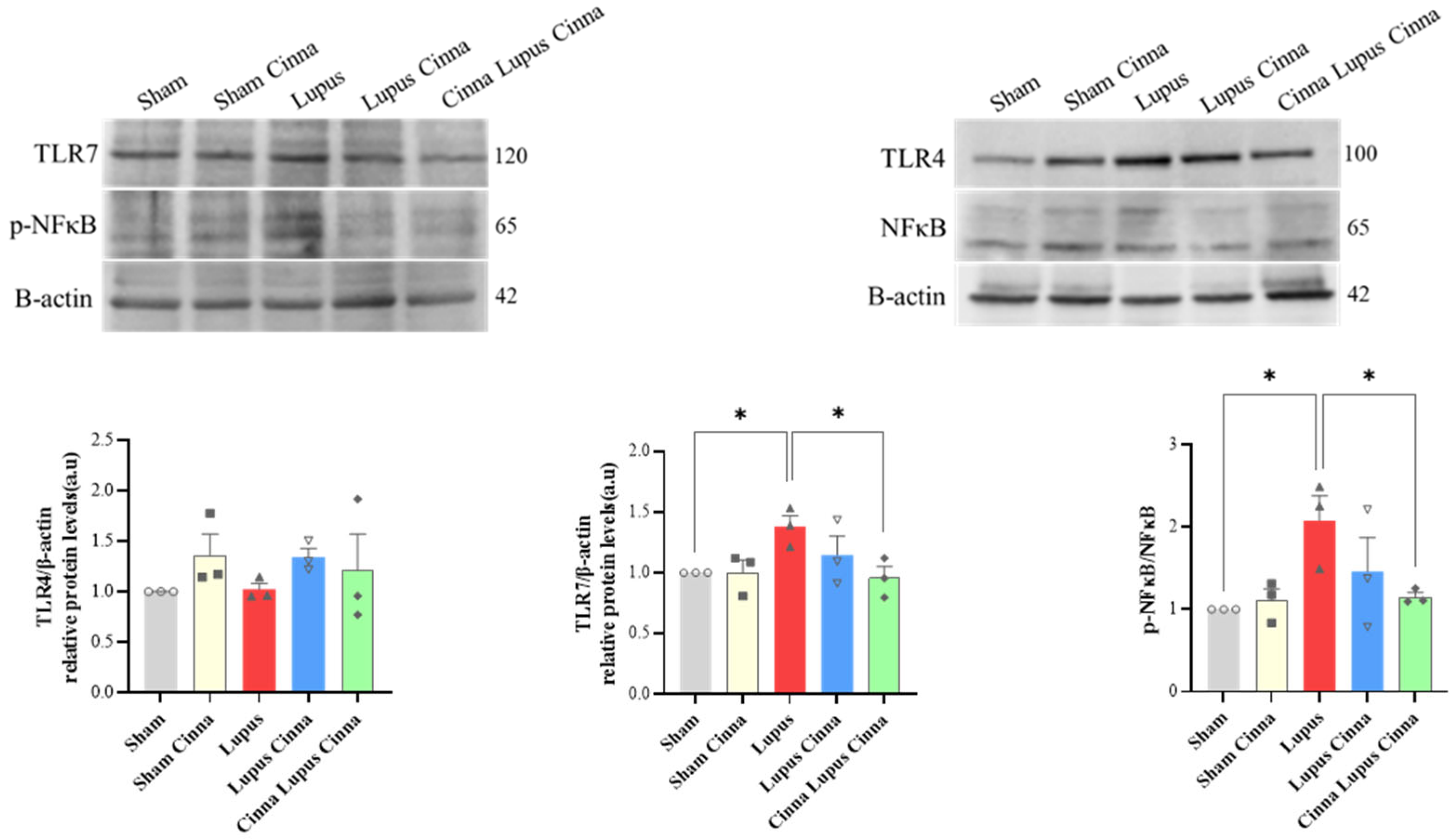
Figure 10.
Antibody array quantification of p-NOS3/p-NOS3, p-FOXO3a/p-FOXO3a, p-NRF2/NRF2, SOD1 and SOD2 with βactin in liver tissue of the different groups. *: p< 0.05.
Figure 10.
Antibody array quantification of p-NOS3/p-NOS3, p-FOXO3a/p-FOXO3a, p-NRF2/NRF2, SOD1 and SOD2 with βactin in liver tissue of the different groups. *: p< 0.05.
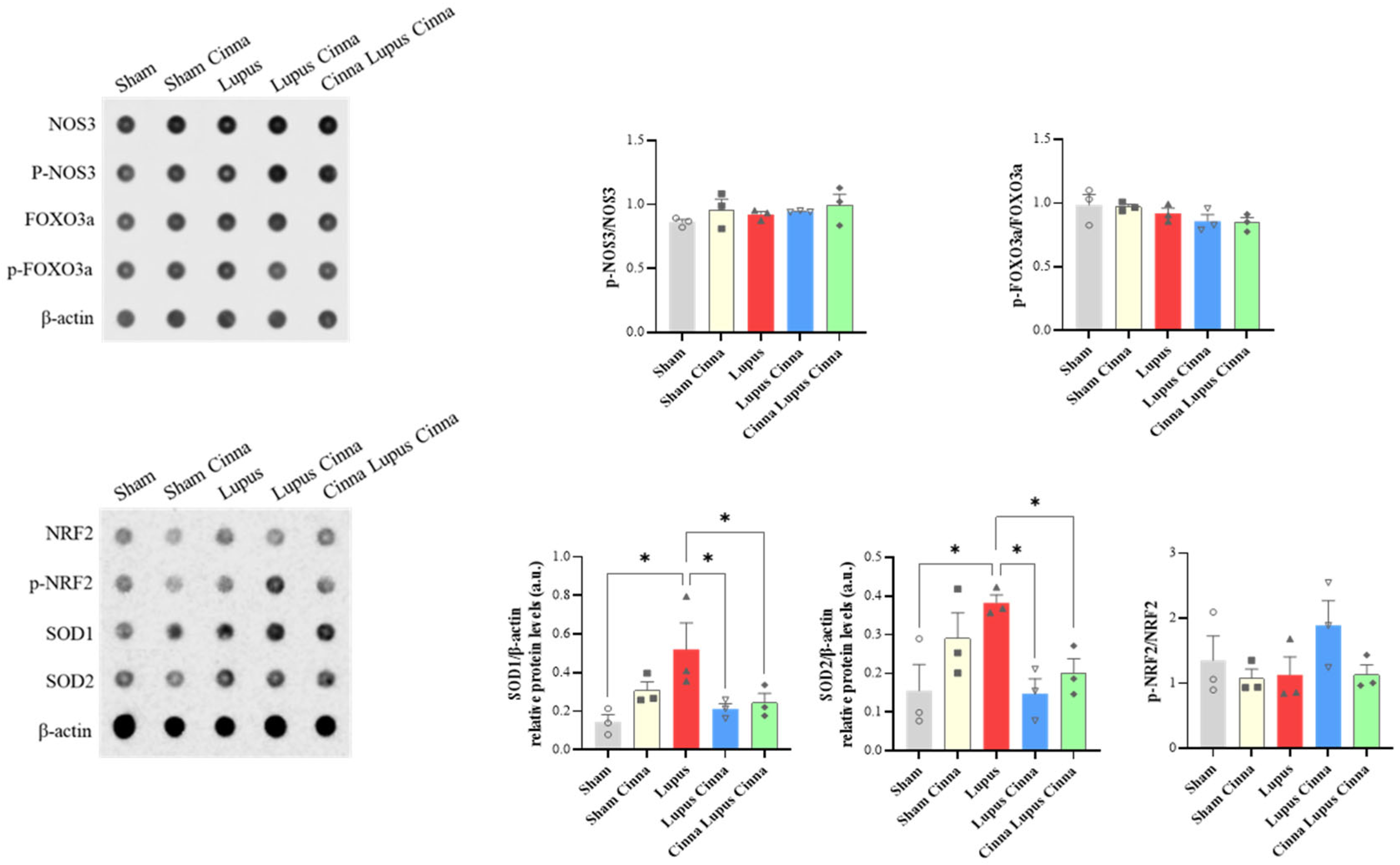

Table 1.
Relative abondance of intestinal microbiota at the genus level.
| Sham Mice | Sham Cinnamon | Sham Lupus | Lupus Cinnamon | Cinnamon Lupus Cinnamon | |
|---|---|---|---|---|---|
| Other genus | 46.4 | 37.2 | 36.4 | 53.5 | 51.2 |
| Lactobacillus | 6.7 | 27.9 | 13.6 | 5.4 | 21.2 |
| Limosilactobacillus | 3.5 | 12.6 | 6.6 | 6.0 | 11.2 |
| Ligilactobacillus | 1.3 | 2.5 | 4.9 | 5.5 | 2.0 |
| Dubosiella | 4.5 | 1.9 | 2.8 | 5.2 | 0.1 |
| Faecalibacterium | 20.5 | 1.4 | 1.5 | 3.6 | 0.0 |
| Staphylococcus | 0.3 | 5.8 | 12.5 | 5.8 | 8.2 |
| Lachnospiraceae genus | 5.2 | 5.3 | 3.5 | 3.4 | 0.4 |
| Bacteroides | 2.4 | 1.1 | 9.1 | 3.0 | 4.7 |
| Prevotella | 4.1 | 3.2 | 5.7 | 7.1 | 0.8 |
| Bifidobacterium | 5.1 | 1.2 | 3.4 | 1.5 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



