
Submitted:
31 October 2025
Posted:
03 November 2025
You are already at the latest version
Abstract
Irritable Bowel Syndrome (IBS) is a common gastrointestinal disorder characterized by chronic abdominal discomfort, pain, and altered bowel habits, as well as extraintestinal manifestations such as fatigue, musculoskeletal pain, gastroesophageal reflux, depression, and anxiety. Despite prevalences of approximately 10% in the general population, its fundamental etiopathogenesis remains unclear. Current theories mainly point toward a multifactorial aetiology crediting psychological stress with a prominent role. The paradigms currently embraced for IBS are mainly based on, and emphasize, the gut-brain axis, visceral hypersensitivity, central sensitization, neuroendocrinology, dysbiosis, motility abnormalities, and post-infectious persistent low-grade mucosal inflammation. Yet, they don’t fully explain its diverse clinical presentations nor IBS’s symptomatic overlap with conditions such as fibrositis/fibromyalgia syndrome. Available treatments are mostly symptomatic and provide limited relief, indicating a still major gap in our mechanistic understanding. This paper proposes a hypothesis for IBS etiopathogenesis as an organic disease of the extracellular matrix driven by myofibroblast overactivity in the gut wall and abdominal soft tissue, along with downstream consequences stemming from tissue stiffness. This process is theorized to mechanically compress visceral structures and nerves, impair synchronized organ motility, and induce substrate stiffness-mediated visceral hypersensitivity. In this theoretical framework the extraintestinal manifestations reflect mechanistic overlap with fibromyalgia, which helps unify functional-psychosomatic syndromes as a medical entity with a shared organic mechanism. This mechanism, along with known mechanisms of gut dysbiosis and the gut-brain axis, helps explain “medically unexplained symptoms” of fibromyalgia-type syndromes and offers testable predictions for future research and suggests new avenues for IBS diagnosis and treatment.
Keywords:
irritable bowel syndrome
; fibromyalgia
; myofibroblast
; extracellular matrix
1. Introduction
Irritable Bowel Syndrome (IBS) is a common gastrointestinal disorder affecting approximately 10 percent of otherwise healthy individuals [1], and comprises 50% of referrals to gastroenterologists [2], characterized by recurrent abdominal discomfort and pain, bloating, and altered bowel habits (constipation, diarrhea, or mixed). Despite its high prevalence and deleterious impact on patient quality of life, the basics of the pathophysiology of IBS remains poorly understood [3]. Once thought of as a purely functional (or “non-organic”) disorder of gut motility, IBS is now viewed as a disorder of gut-brain interaction [1]. Current explanatory models often point toward a complex interplay of factors, including abnormalities of metabolism and the gut-brain axis, visceral hypersensitivity, gut dysbiosis, altered gut motility, and low-grade inflammation [1,3]. Other mechanisms discussed in the literature are increased intestinal permeability, dietary and fermentation, psychological and stress-related responses, neurotransmitter and serotonin imbalance, and bile acid malabsorption [1,3]. Still, these do not seem to fully account for the heterogeneous clinical presentation of IBS, including various extraintestinal manifestations, nor have they consistently guided effective mechanism-based treatments or cure thus far, leaving a major diagnostic and therapeutic gap [3]. If all mechanism-based treatments still disappoint, perhaps medicine is missing something basic about the mechanism of IBS.
Common extraintestinal manifestations of IBS are somatic pain, fatigue, depression, and sleeping disturbances, which overlap considerably with the syndrome of fibromyalgia [4]. In fact, 26–65 percent of IBS patients are reported to have fibromyalgia [4], and in one study of 100 fibromyalgia participants, more than 30% percent fulfilled the Rome criteria for IBS (P<0.001) [5]. In a 2023 study among American population, only 3% (n=8) of IBS patients reported IBS alone without a comorbid functional somatic syndrome [6]. These marked associations and symptomatic overlap have led several researchers to speculate of a shared underlying pathophysiology [7,8,9], emphasizing that IBS and fibromyalgia are syndromes whose definition in nosology isn’t biologically-based nor does its diagnosis at all rely on a specific measurable mechanism [10,11]. Besides, the boundary of a medical entity goes as far as the boundary of the biological process which underlies it.
While several paradigms exist, such as those related to immune system dysregulation, the microbiota-gut-brain axis, epithelial dysfunction, and nociplastic pain and central sensitization, many are under-researched and disconnected from one another. No single theory accounts for the condition comprehensively, explains the various anomalies, or offers effective, disease-modifying treatments or cure. For example, a 2017 study that investigated extracellular matrix (ECM) turnover in IBS found that IBS patients have elevated levels of serum type V pro-collagen when compared to healthy control subjects (p<0.001) [12]. According to TS Kuhn [13], a state of crisis in a scientific field is characterized by multiple competing paradigms where no paradigm may seem adequate — each has flaws or explanatory gaps such that many anomalies persist — scientists argue over fundamentals, and the field is disorganized.
The obscurity that’s seen in the puzzle of IBS is further amplified by its significant overlap with other enigmatic conditions [14,15,16,17]. It’s often grouped within “Chronic Overlapping Pain Conditions” (COPC) / “Functional Somatic Syndromes” (FSS) [14,15,18] such as myofascial pain syndrome and chronic fatigue syndrome which share a baffling array of symptoms and a lack of disease-specific objective findings, leading some researchers to speculate of a shared pathophysiology [7,9,19]. Each of these pain conditions has traditionally belonged to the domain of a different specialty field (e.g., tension-type headache and migraine- neurology. Irritable bowel syndrome- gastroenterology. Myofascial pain- physiotherapy. Post-traumatic stress disorder- psychiatry. Fibromyalgia- rheumatology and neuroscience). Currently, the field of COPC/FSS suffers from siloed disciplines, a fragmented state, multiple competing paradigms, and insufficient treatments. Functional-psychosomatic syndromes such as fibromyalgia, chronic fatigue syndrome, and IBS are being studied in isolation across different medical specialties [20].
Given the “medically unexplained” nature of IBS symptoms and their frequent co-occurrence with fibromyalgia-type syndromes [17], this paper aims to offer a unifying organic mechanistic explanation for IBS pathophysiology that’s also compatible with the extraintestinal manifestations of IBS and overlap with other functional somatic syndromes (namely- fibromyalgia). Any serious theory for IBS must be able to explain its various symptoms and anomalies besides merely abdominal pain or altered bowel habits.
2. Hypothesis
The central hypothesis posited in this work is that a combination of environmental, behavioral, physiological, and genetic factors (e.g., pesticide/herbicide residues, detergents, personal care products and household chemicals, sedentary lifestyle, diet, pharmacology, gut microbiota metabolites, hypermobility syndrome) interact to predispose in combination toward fibroblast-to-myofibroblast trans-differentiation in gut wall and abdominal ECM in susceptible individuals. Consequently, the altered ECM caused by myofibroblast overactivity leads to a disruption of peristalsis, when involving myenteric plexus for instance, and abnormal bowel function. This cellular process is characterized by a local myofibroblast syncytium disturbing the normal synchronized contraction of the gut.
3. Rationale and Supporting Evidence
As part of their normal function, myofibroblasts increase tissue rigidity and mechanical tension via ECM remodelling and stress shielding [21], and they secrete specific cytokines [22] which may influence nerve cell mechanoelectrical activity and visceral sensation. It was previously shown that the biophysical properties of the matrix affect, among several things, embedded nerves and their electrical activity [23]. The effect on autonomic nerves of the enteric nervous system should be similar in this framework. Mechanical tension that develops in the mesentery or fascial sheaths due to contracture of tissue is then dispersed and transmitted to adjacent tissues and anatomical structures. The clinical manifestation is the outcome of the whole mechanism. Myofibroblasts are hypothesized by several authors to play a part in the pathophysiology of conditions such as myofascial pain syndrome [24], endometriosis [25], low back pain [26], asthma [27], malignant hyperthermia [28], fibromyalgia syndrome, and more [29,30]. The following section provides findings for support of the main rationale of the hypothesis.
The proposed hypothesis builds on a recently developed paradigm linking soft tissue myofibroblasts with fibromyalgia-type syndromes and aligns with a multifactorial aetiology [21,22]. Functional somatic syndromes are hypothesized to be a clinical phenomenon with a common pathophysiological mechanism, potentially representing a single heterogenous medical entity manifesting with varying clinical expressions [7,8,9,14,23,24]. The idea of a shared mechanism among functional somatic syndromes is not new [7,8,19]. Supporting evidence for the suggested myofibroblast-based paradigm for fibromyalgia-type syndromes can be found in a recent study [21]. The following sections provide empirical findings from previous studies to substantiate the main hypothesis.
3.1. Myofibroblast Mechanobiology, Tissue Remodelling and Contracture
Myofibroblasts are mechanosensitive cells that have natural mechanoactivity that is often compared to smooth muscle cells [25]. When fibroblast differentiate into myofibroblasts, they secrete and degrade extracellular matrix (ECM) material, remodel their surrounding environment, and lock-in mechanical stress into the surrounding matrix by the help of intracellular alpha-smooth muscle actin (α-SMA) fibres [25]. Myofibroblast-mediated ECM remodelling and contracture formation in tissue occurs in a positive feed-back loop mediated by a cellular signalling pathway that involves integrin and transforming growth-factor beta (TGF-β). Myofibroblasts are induced by various stimuli and are part of the inflammatory and healing process, most known for in scar formation. They orient themselves in tissue by stiffness gradient and arranged themselves in cords or nodules [26,27,28,29].
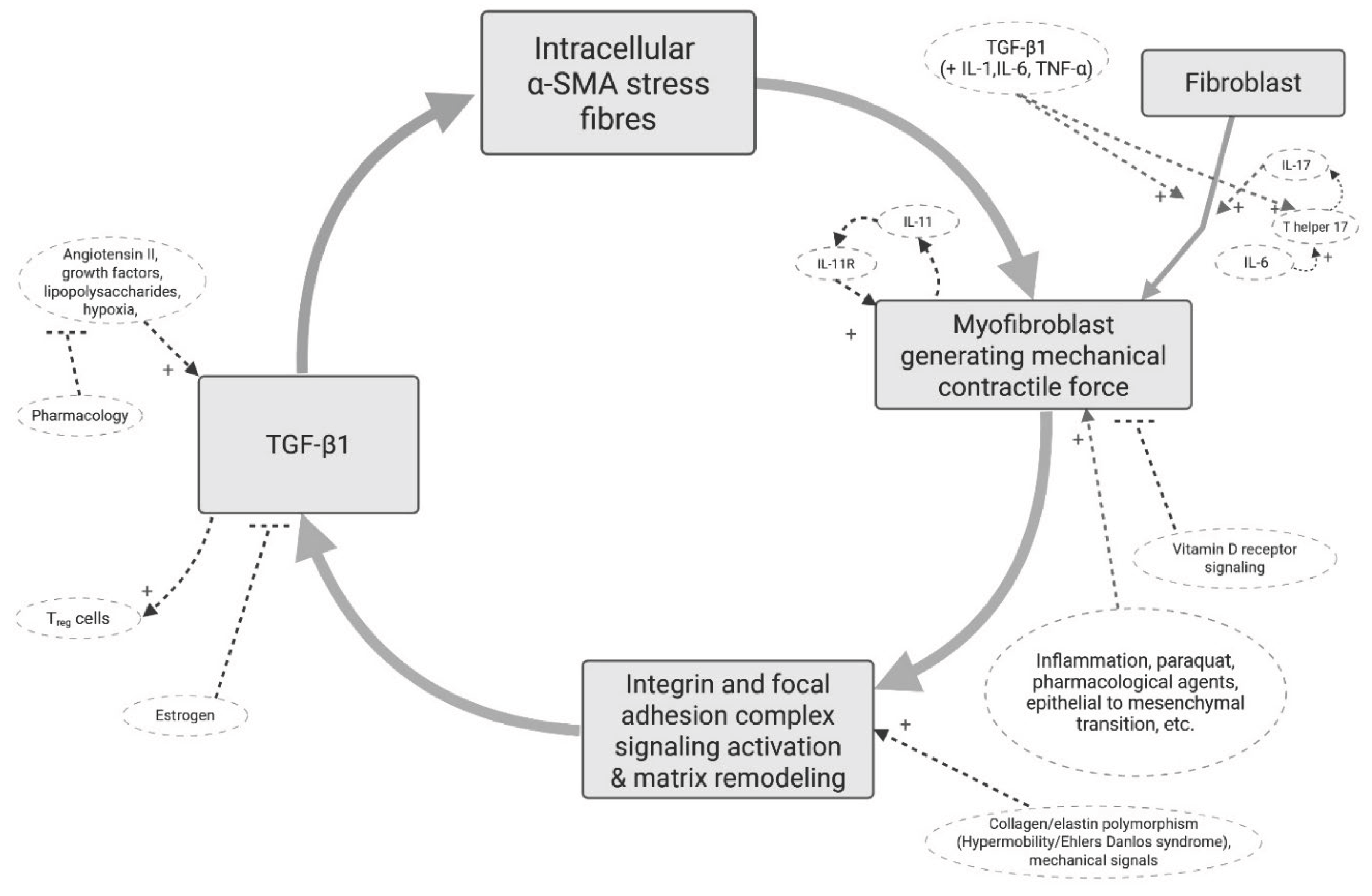
Myofibroblasts are also relevant, though somewhat overlooked, in other conditions including asthma [30], cardiac arrythmias [31], inflammatory bowel disease [32], and during infection and events of systemic inflammation [33]. Figure 1 shows the core cellular pathway of myofibroblasts. Mechanical stress within the local tissue enhances TGF-β signalling, which in turn promotes α-SMA expression and further contraction, creating a self-perpetuating cycle [25,34,35]. While this process is vital for normal physiological functions, its dysregulation can lead to pathological states of chronic tissue tension and fibrosis [36,37]. Examples of factors that can help induce or inhibit the differentiation of fibroblasts to myofibroblasts are immobility, paraquat, collagen microarchitecture, vitamin D, and more [38,39,40,41,42,43,44,45], which provide input into the vicious cycle. Gut microbe-derived metabolites can accelerate fibroblast-to-myofibroblast differentiation [46]. The delicate process of myofibroblast de-differentiation/senescence/apoptosis is important for the health of tissue [37] and is influenced by factors such as fibroblast growth factors, prostaglandins, cellular communication network factor 1 (CCN1), metformin, and more [43,44,45,47,48]. Myofibroblasts are also more easily stimulated in hypermobility syndrome/Ehler’s Danlos syndrome because collagen micro-architecture affects mechanosensitive signalling of myofibroblasts [49,50]. In a mechanism driven by myofibroblasts, factors providing upregulating input into the myofibroblast mechano-active cycle are expected to be part of the aetiology and risk factors. On the other hand, factors that inhibit fibroblast-to-myofibroblast transition or attenuate myofibroblast activity are expected to be protective factors [21].
3.2. Herbicide Residues and Harmful Substances in Food Products
Chronic ingestion of pesticide residues through food is a global public health concern [52]. The presence of herbicide residues and other harmful chemical contaminants in food, even in small amounts [53], has raised growing concern about their potential contribution to the development of chronic diseases and metabolic dysfunctions in humans [54,55,56,57]. A 2022 study investigating environmental factors contributing to the pathogenesis of inflammatory bowel disease identified the herbicide propyzamide as factor that increases inflammation in the small and large intestine [58]. In a 2025 in-vivo study paraquat exposure impaired gut barrier function and affected gut microbial diversity (P < 0.05) [59]. Histopathological alterations in intestinal tissue are induced by herbicide exposure in animal models, including a decrease in colonic tight junction proteins, increased expression levels of inflammatory markers, and an altered diversity and abundance of gut microbiota [57]. A systematic review evaluating the exposed to pesticides in agricultural and food products identified studies that found an amount of carcinogenic organochlorine pesticides higher than the permissible level [60].
Myofibroblasts are induced and stimulated by herbicides such as paraquat. Sun et al. (2020) showed that the herbicide paraquat induces pulmonary fibrosis through myofibroblast differentiation [42]. Exposure to paraquat induces fibroblast-to-myofibroblast transition in vitro, a process involving connective tissue growth factor and the canonical Wnt signaling pathway, α-smooth muscle actin, vimentin, and collagen [61]. Paraquat-Induced epithelial to mesenchymal transition is regulated by TGF-β and inhibited by pretreatment of curcumin in alveolar epithelial cells in vitro [62]. Animal models suggest that in muscle and ileum tissue the gene expression of TGF-β1, connective tissue growth factor, COL1A1, and COL3A1, increase significantly after exposure of cells to propiconazole [63], a fungicide found in meat products.
Taken together, these findings suggest that chronic environmental exposure of herbicides, pesticides and other toxic substances in small amounts in food products may be able to induce myofibroblast cells in human gut.
3.3. Myofibroblasts in Visceral Tissue Influence Organ Function and Integrity
Myofibroblasts can function as a collaborative network of cells in tissue by the coupling of cells via connexin 43 gap junctions and are important for organ function [64,65,66,67]. Myofibroblasts exist in the gut wall of normal individuals [68] and seem to be involved in the pathophysiology of inflammatory bowel disease and its mucosal repair process [69]. Subepithelial cells of the intestine are a distinct cell type - α-SMA positive myofibroblasts - that are immunophenotypically distinct from muscularis mucosae smooth muscle cells. These pericryptal stromal myofibroblast cells are connected with the interstitial, non-pericryptal fibroblasts as a network throughout the lamina propria of the normal colon [68].
Connexin expression is prominent in myofibroblast-like cells, supposedly involved in afferent signalling pathways of the bladder. Up-regulation of Connexin43 in patients with urge incontinence supports the possibility of functional changes in the syncytial properties of detrusor smooth muscle cells in this condition [70]. In addition, the observed increase of Cx45 in the myofibroblast cell layer supports the idea that alterations in sensory signalling are also involved [70].
Electrical coupling of fibroblasts and myocytes is relevant for cardiac propagation and electrophysiology [71,72]. The inherent mechanosensitivity of cardiac fibroblasts could allow them to play a sensory role and to affect cardiac electrophysiology via mechanoelectric feedback. In heart, fibroblasts may act as current sinks, contributing to the formation of unidirectional block or to the delay in atrioventricular conduction [71]. In culture, however, the heterogeneous cell types form functional gap junctions, which can provide a substrate for electrical coupling of distant myocytes, interconnected by fibroblasts only [73].
Lastly, interaction between Th-17 cells and colonic subepithelial myofibroblasts may play an important role in the pathophysiology of IBS and inflammatory bowel disease perhaps by contributing to low-grade inflammation [74].
3.4. Substrate Stiffness Affects Cell Function and Behaviour
Myofibroblasts normally remodel the ECM in a higher stiffness state [21,25]. Substrate stiffness and rigidity influences behaviour and function of nerve cells [75] including synaptic density, electrophysiological activity, and growth dynamics [76]. A 2012 study [77] investigated the influence of substrate stiffness on the behaviour and functions of Schwann cells in culture. Cell attachment and spreading were more evident for Schwann cells grown on a 7.45 kPa substrate. Their cytoskeletal arrangement was markedly different from that of Schwann cells grown on the 4.42 or 12.04 kPa substrate [77]. A 2016 study investigated the role of matrix rigidity on the formation and activity of cortical neuronal networks in vitro [76]. Results showed that the mean maximal number of action potentials was lower on soft matrices than on stiff matrices, suggesting that the intrinsic excitability of cortical neurons is enhanced on stiff matrices.
Of relevance, Piezo channels sense extracellular mechanical stimuli, and Piezo-dependent mechanosensing is essential for intestinal stem cell fate decision and maintenance [78]. In response to mechanical forces enterochromaffin cells release serotonin [79]. Epithelial Piezo2 mechanosensitive ion channels have been noted in regulation of intestinal physiology as a primary mechanotransducer and mechanosensitive serotonin release [80].
4. Extraintestinal Manifestations of IBS: Pain and Fibromyalgia-Type Symptoms
IBS and fibromyalgia are known to overlap considerably in symptoms [2,81]. Extraintestinal manifestations of IBS include back pain, migraine headache, muscle pain, fatigue, and depression [2,4]. Common gastrointestinal manifestations of fibromyalgia include functional bowel disturbances, abdominal discomfort, and bloating [82]. The overlap of IBS and fibromyalgia is noticeable [14,83]. Referred pain from myofascial trigger points can mimic visceral pain syndromes and visceral pain syndromes can induce trigger point development and myofascial pain and dysfunction [84]. In this conceptual framework of myofibroblast-mediated ECM alterations in peripheral tissue, ECM remodelling induces increased tissue stiffness. When myofascial tissue is affected, it’s expected to manifest clinically as a mild-to-moderate chronic exertional compartment-like syndrome [21]. In this situation, starling forces are altered in muscle microcirculation in a state characterized by congestion as blood backtracks and is shunted towards the venous circulation even though the precapillary sphincters remain open. Nitric oxide synthesis can increase as part of the compensation, but it doesn’t correct the pathological cause of the ongoing low-grade ischemia. Post-exertional malaise and chronic fatigue are part of its result. In severe cases, especially if involvement of the diaphragm impairs deep inhalation and breathing patterns, mild hyperventilation would reasonably be seen in a healthy body seeking physiological compensation. Increased respiratory rate and shallow breathing (compared to one’s own baseline) doesn’t characterize a parasympathetic state of relaxation, so sleep disturbances are inferred easily here, and agitation and anxiety are secondary. As a mild-moderate global chronic compartment-like syndrome, single-site or single-time measurements may not be the best choice for its investigation. In this myofibroblast-based paradigm, fibromyalgia and IBS symptomatic overlap reflects a mechanistic overlap. Pain is a symptom of the entity; it isn’t the actual entity itself. Any serious theory for IBS must be able to explain its various symptoms and anomalies besides merely abdominal pain and altered bowel habits.
5. Implications of the Hypothesis
This hypothesis suggests that myofibroblast overactivity could be a common pathway or a critical mediator through which various established factors such as chronic low-grade inflammation, dysbiosis, or stress signals, interact. The hypothesis shifts focus from a psychosocial neurocentric thesis [85] to an organic biophysical basis of the disorder. This potentially opens entirely new avenues for treatment by targeting altered stiffness and composition of the ECM. By identifying a distinct and explicit cellular pathway, this theory could lead to more accurate diagnostic biomarkers and more effective targeted treatments, developing preclinical models that accurately reflect this pathology, and exploring the interplay between the gut microbiome, colonic epithelial cells, immune cells, enteric nervous system, and myofibroblasts to fully map the disease pathology.
6. Ways to Test the Hypothesis
Based on the hypothesis of myofibroblasts induction in IBS, several empirical predictions can be formulated:
- Biophysical tests: atomic force microscopy, optical coherence elastography, dynamic mechanical analysis, of sampled tissue (e.g., abdominal fascia, psoas muscle, gut wall) might be insightful, although these would have to take into account the complexity of the model and possible confounding factors, and control for hypermobility syndrome. Age, sex, pH, temperature, hydration, hyaluronic acid composition, adipocytes, cell phenotype and density, are all variables that may affect the properties of tissue.
- Imaging: Results of magnetic resonance elastography investigations, if sensitive enough, are expected to reflect higher abdominal ECM rigidity and reveal abdominal taut bands or fascial nodules and cords.
- Staining: immunohistochemistry and histological staining of tissue samples for myofibroblast markers would reveal a higher density of myofibroblasts in IBS. Presence of vimentin, α-SMA, and connexin 43, should be increased.
- Response to current medications: the most effective pharmacological treatments for IBS that have so far been identified empirically are thus expected to work mainly through an influence on myofibroblasts and fascia (downregulate myofibroblasts and reduce ECM stiffness). As a disorder in the domain of biophysics, extracorporeal shockwave therapy and vibration therapy should be useful in IBS, though not necessarily curative. If myofascial tension in the low back is transmitted to abdominal myofascial tissue through myofascial links, there should be clinical improvement by treating tension in the low back.
- Issue with systemic pharmacological therapies: A key player in the activation and maintenance of myofibroblasts is the cytokine TGF-β1. At first, based on the myofibroblast theory, systemically targeting the TGF-β pathway pharmacologically seems like a promising therapeutic strategy. However, TGF-β has a multifunctional role in the body. TGF-β is essential for numerous physiological processes, including immune regulation, cell growth, differentiation, and tissue repair. Indiscriminate, systemic inhibition of TGF-β1 can lead to a wide range of severe on-target and off-target adverse effects such as cardiovascular toxicity, immune dysregulation, and more.
Falsification: According to Karl Popper, for a theory to be truly scientific, it must be falsifiable, meaning there must be conceivable empirical observations or experiments that can potentially falsify the theory. This myofibroblast-based paradigm can be falsified in a study that quantifies tissue stiffness and elasticity across multiple samples of abdominal tissue (fascia, intestine, etc) in a large sample size cohort of IBS patients and well-matched controls, if these consistently show no statistically significant difference in specific, predicted mechanical and biophysical properties between IBS patients and controls, or if the observed differences do not correlate with patient-reported symptom burden. If an intervention aiming for prevention successfully inhibits myofibroblast induction and prevents a pathological alteration in ECM, but there is no statistically and clinically significant corresponding reduction in new onset IBS symptoms compared to a credible sham or control intervention, this will weaken the causal link posited by the theory. If there is no difference in ECM properties or myofibroblast density between IBS/fibromyalgia and non-IBS-fibromyalgia-type patients, this will be a clear falsifying counterinstance. Disease burden and myofibroblasts should correlate well.
7. Limitations and Challenges
The proposed hypothesis, while offering a coherent framework, is not without limitations. Much of the supporting evidence is currently correlational and derived from in vitro and in vivo studies, meaning a direct causal link between visceral myofibroblast activity and IBS has yet to be definitively established. The complex and multifactorial nature of IBS means that this mechanism is integrated with other factors such as genetics, gut microbiota, and psychological input. Nevertheless, relying on psychosomatic psychosocial stress seems to be a weak attempt to simplify such a complex problem. Furthermore, measuring the specific biomechanical properties of the visceral fascia in real time, such as tissue stiffness and tension, presents significant technical and methodological challenges. The variability of IBS symptoms between individuals and the lack of standardized measurement protocols make rigorous, large-scale clinical validation difficult to design and execute.
8. Conclusion
Taken together, the findings presented here suggest that induction of myofibroblasts by factors such as pollutants and contaminated food products may lead to their overactivity in the gut wall and abdominal soft tissue, which would affect ECM and organ functioning. Altered ECM biophysical properties in the gut wall and epithelial tissue have downstream consequences on enterochromaffin cells that release serotonin and the enteric nervous system, mainly peristalsis. The gut flora is integrated into the mechanism too. Involvement of serotonin may have implications for psychiatric conditions such as depression.
The myofibroblast-based hypothesis offers a unifying framework that can help explain the diverse and perplexing symptoms of IBS, as well as its overlap with other chronic pain conditions. The downstream consequences of such myofibroblast-mediated ECM stiffness may include central neurophysiological compensation and spinal and brain adaptations. This perspective calls for new research avenues, including the use of advanced biophysics investigational techniques like elastography, optical coherence elastography, and atomic force microscopy, to measure visceral tissue stiffness. If myofibroblasts are involved in the pathogenesis of IBS, this calls for the development of targeted therapies aimed at preventing and reversing myofibroblast over-stimulation and their active contractility and ECM remodelling and opens a new avenue for research on the link between ECM and neurophysiology. This theory clearly needs to be adjusted according to further empirical research. Lastly, if all theory-based treatments are stubbornly disappointing, perhaps the theory should be reconsidered.
Author Contributions
SP was solely responsible for all aspects of the work, including study conception and design, material preparation, data collection, data curation, conceptualization, integration of information, theoretical framework development, formal analysis investigation, methodology, review of literature, visualization, figures and diagrams, resources, writing— original draft, writing—review and editing, and preparing the manuscript for submission.
Funding
The author declares that no funds, grants, or other support were received during the preparation of this manuscript.
Conflicts of Interest
The author has no relevant financial or non-financial interests to disclose.
AI Use Disclosure
During the preparation of this manuscript large-language model GEMINI Flash 2.5 was used to generate a first preliminary draft of the text by giving it input (literature) and requesting specific output as the prompt. Afterwards the text was edited and revised in iterations. The conceptualization was done solely by the author based on the fascia-based conceptual framework for fibromyalgia, and the author takes full responsibility for the final version of the manuscript.
Abbreviations
α-SMA- alpha smooth muscle actin, ECM- extracellular matrix, TGF-β- Transforming growth-factor beta,
References
- Ford AC, Sperber AD, Corsetti M, Camilleri M. Irritable bowel syndrome. Lancet. 2020 Nov 21;396(10263):1675-1688. Epub 2020 Oct 10. PMID: 33049223. n.d. [CrossRef]
- Arendt-Nielsen L, Morlion B, Perrot S, Dahan A, Dickenson A, Kress HG, Wells C, Bouhassira D, Drewes AM. Assessment and manifestation of central sensitisation across different chronic pain conditions. Eur J Pain. 2018 Feb;22(2):216-241. Epub 2017 Nov 5. PMID: 29105941. n.d. [CrossRef]
- Huang KY, Wang FY, Lv M, Ma XX, Tang XD, Lv L. Irritable bowel syndrome: Epidemiology, overlap disorders, pathophysiology and treatment. World J Gastroenterol. 2023 Jul 14;29(26):4120-4135. PMID: 37475846; PMCID: PMC10354571. n.d. [CrossRef]
- Ohlsson B. Extraintestinal manifestations in irritable bowel syndrome: A systematic review. Therap Adv Gastroenterol. 2022 Aug 9;15:17562848221114558. n.d. [CrossRef]
- Kurland JE, Coyle WJ, Winkler A, Zable E. Prevalence of irritable bowel syndrome and depression in fibromyalgia. Dig Dis Sci. 2006 Mar;51(3):454-60. PMID: 16614951. n.d. [CrossRef]
- Wang XJ, Ebbert JO, Loftus CG, Rosedahl JK, Philpot LM. Comorbid extra-intestinal central sensitization conditions worsen irritable bowel syndrome in primary care patients. Neurogastroenterol Motil. 2023 Apr;35(4):e14546. Epub 2023 Feb 19. PMID: 36807964. n.d. [CrossRef]
- Luís M, Pinto AM, Häuser W, Jacobs JW, Saraiva A, Giorgi V, Sarzi-Puttini P, Castelo-Branco M, Geenen R, Pereira da Silva JA. Fibromyalgia and post-traumatic stress disorder: different parts of an elephant? Clin Exp Rheumatol. 2025 Jun;43(6):1146-1160. n.d. [CrossRef]
- Bourke JH, Langford RM, White PD. The common link between functional somatic syndromes may be central sensitisation. J Psychosom Res. 2015 Mar;78(3):228-36. Epub 2015 Jan 9. PMID: 25598410. n.d. [CrossRef]
- Wessely S, Nimnuan C, Sharpe M. Functional somatic syndromes: one or many? Lancet. 1999 Sep 11;354(9182):936-9. PMID: 10489969. n.d. [CrossRef]
- Wolfe F, Walitt B, Rasker JJ, Häuser W. Primary and Secondary Fibromyalgia Are The Same: The Universality of Polysymptomatic Distress. J Rheumatol. 2019 Feb;46(2):204-212. Epub 2018 Jul 15. PMID: 30008459; PMCID: PMC12320112. n.d. [CrossRef]
- Talley NJ. Irritable bowel syndrome: definition, diagnosis and epidemiology. Baillieres Best Pract Res Clin Gastroenterol. 1999 Oct;13(3):371-84. PMID: 10580915. n.d. [CrossRef]
- Mortensen JH, Manon-Jensen T, Jensen MD, Hägglund P, Klinge LG, Kjeldsen J, Krag A, Karsdal MA, Bay-Jensen AC. Ulcerative colitis, Crohn’s disease, and irritable bowel syndrome have different profiles of extracellular matrix turnover, which also reflects disease activity in Crohn’s disease. PLoS One. 2017 Oct 13;12(10):e0185855. PMID: 29028807; PMCID: PMC5640222. n.d. [CrossRef]
- Kuhn TS. The structures of scientific revolutions. 4th edition (50th anniversary edition). London: University of Chicago Press (2012). Chapter 4. pp 39. n.d.
- Goldenberg DL. How to understand the overlap of long COVID, chronic fatigue syndrome/myalgic encephalomyelitis, fibromyalgia and irritable bowel syndromes. Semin Arthritis Rheum. 2024 Aug;67:152455. Epub 2024 May 7. PMID: 38761526. n.d. [CrossRef]
- Maixner W, Fillingim RB, Williams DA, Smith SB, Slade GD. Overlapping Chronic Pain Conditions: Implications for Diagnosis and Classification. J Pain. 2016 Sep;17(9 Suppl):T93-T107. PMID: 27586833; PMCID: PMC6193199. n.d. [CrossRef]
- McKernan LC, Crofford LJ, Kim A, Vandekar SN, Reynolds WS, Hansen KA, Clauw DJ, Williams DA. Electronic Delivery of Pain Education for Chronic Overlapping Pain Conditions: A Prospective Cohort Study. Pain Med. 2021 Oct 8;22(10):2252-2262. PMID: 33871025; PMCID: PMC8677459. n.d. [CrossRef]
- Valencia C, Fatima H, Nwankwo I, Anam M, Maharjan S, Amjad Z, Abaza A, Vasavada AM, Sadhu A, Khan S. A Correlation Between the Pathogenic Processes of Fibromyalgia and Irritable Bowel Syndrome in the Middle-Aged Population: A Systematic Review. Cureus. 2022 Oct 4;14(10):e29923. PMID: 36381861; PMCID: PMC9635936. n.d. [CrossRef]
- Fall EA, Chen Y, Lin JS, Issa A, Brimmer DJ, Bateman L, Lapp CW, Podell RN, Natelson BH, Kogelnik AM, Klimas NG, Peterson DL, Unger ER; MCAM Study Group. Chronic Overlapping Pain Conditions in people with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): a sample from the Multi-site Clinical Assessment of ME/CFS (MCAM) study. BMC Neurol. 2024 Oct 18;24(1):399. PMID: 39425035; PMCID: PMC11488184. n.d. [CrossRef]
- Johnston KJA, Signer R, Huckins LM. Chronic overlapping pain conditions and nociplastic pain. HGG Adv. 2025 Jan 9;6(1):100381. Epub 2024 Nov 4. PMID: 39497418; PMCID: PMC11617767. n.d. [CrossRef]
- Alpers DH. Is irritable bowel syndrome more than just a gastroenterologist’s problem? Am J Gastroenterol. 2001 Apr;96(4):943-5. PMID: 11316208. n.d. [CrossRef]
- Plaut S. Scoping review and interpretation of myofascial pain/fibromyalgia syndrome: An attempt to assemble a medical puzzle. PLoS One 2022;17. [CrossRef]
- Plaut S. Disrupted Biotensegrity in the Fiber Cellular Fascial Network and Neuroma Microenvironment: A Conceptual Framework for “Phantom Limb Pain”. Int J Mol Sci. 2025 Aug 22;26(17):8161. n.d. [CrossRef]
- Goebel A, Andersson D, Shoenfeld Y. The biology of symptom-based disorders - time to act. Autoimmun Rev. 2023 Jan;22(1):103218. Epub 2022 Oct 22. PMID: 36280093. n.d. [CrossRef]
- Malkova AM, Shoenfeld Y. Autoimmune autonomic nervous system imbalance and conditions: Chronic fatigue syndrome, fibromyalgia, silicone breast implants, COVID and post-COVID syndrome, sick building syndrome, post-orthostatic tachycardia syndrome, autoimmune diseases and autoimmune/inflammatory syndrome induced by adjuvants. Autoimmunity reviews. 2023 Jan 1;22(1):103230. n.d.
- Tomasek JJ, Gabbiani G, Hinz B, Chaponnier C, Brown RA. Myofibroblasts and mechano: Regulation of connective tissue remodelling. Nat Rev Mol Cell Biol 2002;3:349–63. [CrossRef]
- Moyer KE, Banducci DR, Graham WP 3rd, Ehrlich HP. Dupuytren’s disease: physiologic changes in nodule and cord fibroblasts through aging in vitro. Plast Reconstr Surg. 2002 Jul;110(1):187-93; discussion 194-6. PMID: 12087251. n.d. [CrossRef]
- Bisson MA, Mudera V, McGrouther DA, Grobbelaar AO. The contractile properties and responses to tensional loading of Dupuytren’s disease--derived fibroblasts are altered: a cause of the contracture? Plast Reconstr Surg. 2004 Feb;113(2):611-21; discussion 622-4. PMID: 14758224. n.d. [CrossRef]
- Ziegler ME, Staben A, Lem M, Pham J, Alaniz L, Halaseh FF, Obagi S, Leis A, Widgerow AD. Targeting Myofibroblasts as a Treatment Modality for Dupuytren Disease. J Hand Surg Am. 2023 Sep;48(9):914-922. Epub 2023 Jul 20. PMID: 37480917. n.d. [CrossRef]
- Zhao F, Zhang M, Nizamoglu M, Kaper HJ, Brouwer LA, Borghuis T, Burgess JK, Harmsen MC, Sharma PK. Fibroblast alignment and matrix remodeling induced by a stiffness gradient in a skin-derived extracellular matrix hydrogel. Acta Biomater. 2024 Jul 1;182:67-80. Epub 2024 May 13. PMID: 38750915. n.d. [CrossRef]
- Jendzjowsky NG, Kelly MM. The Role of Airway Myofibroblasts in Asthma. Chest. 2019 Dec;156(6):1254-1267. Epub 2019 Aug 28. PMID: 31472157. n.d. [CrossRef]
- Johnson RD, Lei M, McVey JH, Camelliti P. Human myofibroblasts increase the arrhythmogenic potential of human induced pluripotent stem cell-derived cardiomyocytes. Cell Mol Life Sci. 2023 Sep 5;80(9):276. Erratum in: Cell Mol Life Sci. 2024 Dec 27;82(1):20. doi:10.1007/s00018-024-05492-w. PMID: 37668685; PMCID: PMC10480244. n.d. [CrossRef]
- Kinchen J, Chen HH, Parikh K, Antanaviciute A, Jagielowicz M, Fawkner-Corbett D, Ashley N, Cubitt L, Mellado-Gomez E, Attar M, Sharma E, Wills Q, Bowden R, Richter FC, Ahern D, Puri KD, Henault J, Gervais F, Koohy H, Simmons A. Structural Remodeling of the Human Colonic Mesenchyme in Inflammatory Bowel Disease. Cell. 2018 Oct 4;175(2):372-386.e17. Epub 2018 Sep 27. PMID: 30270042; PMCID: PMC6176871. n.d. [CrossRef]
- Kruglikov IL, Scherer PE. The Role of Adipocytes and Adipocyte-Like Cells in the Severity of COVID-19 Infections. Obesity 2020;28:1187–90. [CrossRef]
- Martin K, Pritchett J, Llewellyn J, Mullan AF, Athwal VS, Dobie R, et al. PAK proteins and YAP-1 signalling downstream of integrin beta-1 in myofibroblasts promote liver fibrosis. Nat Commun 2016;7. [CrossRef]
- Wang S, Meng X-M, Ng Y-Y, Ma FY, Zhou S, Zhang Y, et al. TGF-β/Smad3 signalling regulates the transition of bone marrow-derived macrophages into myofibroblasts during tissue fibrosis. vol. 7. n.d.
- Stempien-Otero A, Kim DH, Davis J. Molecular networks underlying myofibroblast fate and fibrosis. J Mol Cell Cardiol 2016;97:153–61. [CrossRef]
- Hinz B, Lagares D. Evasion of apoptosis by myofibroblasts: a hallmark of fibrotic diseases. Nat Rev Rheumatol. 2020 Jan;16(1):11-31. Epub 2019 Dec 2. PMID: 31792399; PMCID: PMC7913072. n.d. [CrossRef]
- Hinz B, Mastrangelo D, Iselin CE, Chaponnier C, Gabbiani G. Mechanical tension controls granulation tissue contractile activity and myofibroblast differentiation. American Journal of Pathology 2001;159:1009–20. [CrossRef]
- Sasabe R, Sakamoto J, Goto K, Honda Y, Kataoka H, Nakano J, et al. Effects of joint immobilization on changes in myofibroblasts and collagen in the rat knee contracture model. Journal of Orthopaedic Research 2017;35:1998–2006. [CrossRef]
- Seo BR, Chen X, Ling L, Song YH, Shimpi AA, Choi S, et al. Collagen microarchitecture mechanically controls myofibroblast differentiation. Proc Natl Acad Sci U S A. 2020;117(21):11387–11398. n.d. [CrossRef]
- Sari E, Oztay F, Tasci AE. Vitamin D modulates E-cadherin turnover by regulating TGF-β and Wnt signalings during EMT-mediated myofibroblast differentiation in A459 cells. Journal of Steroid Biochemistry and Molecular Biology 2020;202. [CrossRef]
- Sun Z, Yang Z, Wang M, Huang C, Ren Y, Zhang W, Gao F, Cao L, Li L, Nie S. Paraquat induces pulmonary fibrosis through Wnt/β-catenin signaling pathway and myofibroblast differentiation. Toxicol Lett. 2020 Oct 15;333:170-183. Epub 2020 Aug 11. PMID: 32795487. n.d. [CrossRef]
- Lee SA, Yang HW, Um JY, Shin JM, Park IH, Lee HM. Vitamin D attenuates myofibroblast differentiation and extracellular matrix accumulation in nasal polyp-derived fibroblasts through smad2/3 signaling pathway. Sci Rep. 2017 Aug 4;7(1):7299. PMID: 28779150; PMCID: PMC5544725. n.d. [CrossRef]
- Olson ER, Naugle JE, Zhang X, Bomser JA, Meszaros JG. Inhibition of cardiac fibroblast proliferation and myofibroblast differentiation by resveratrol. Am J Physiol Heart Circ Physiol. 2005 Mar;288(3):H1131-8. Epub 2004 Oct 21. PMID: 15498824. n.d. [CrossRef]
- Baghdasaryan A, Claudel T, Kosters A, Gumhold J, Silbert D, Thüringer A, Leski K, Fickert P, Karpen SJ, Trauner M. Curcumin improves sclerosing cholangitis in Mdr2-/- mice by inhibition of cholangiocyte inflammatory response and portal myofibroblast proliferation. Gut. 2010 Apr;59(4):521-30. PMID: 20332524; PMCID: PMC3756478. n.d. [CrossRef]
- Yang W, Zhang S, Zhu J, Jiang H, Jia D, Ou T, Qi Z, Zou Y, Qian J, Sun A, Ge J. Gut microbe-derived metabolite trimethylamine N-oxide accelerates fibroblast-myofibroblast differentiation and induces cardiac fibrosis. J Mol Cell Cardiol. 2019 Sep;134:119-130. Epub 2019 Jul 9. PMID: 31299216. n.d. [CrossRef]
- Wu M, Han M, Li J, Xu X, Li T, Que L, Ha T, Li C, Chen Q, Li Y. 17beta-estradiol inhibits angiotensin II-induced cardiac myofibroblast differentiation. Eur J Pharmacol. 2009 Aug 15;616(1-3):155-9. Epub 2009 May 24. PMID: 19470381. n.d. [CrossRef]
- Kheirollahi V, Wasnick RM, Biasin V, Vazquez-Armendariz AI, Chu X, Moiseenko A, Weiss A, Wilhelm J, Zhang JS, Kwapiszewska G, Herold S, Schermuly RT, Mari B, Li X, Seeger W, Günther A, Bellusci S, El Agha E. Metformin induces lipogenic differentiation in myofibroblasts to reverse lung fibrosis. Nat Commun. 2019 Jul 5;10(1):2987. PMID: 31278260; PMCID: PMC6611870. n.d. [CrossRef]
- Zoppi N, Chiarelli N, Binetti S, Ritelli M, Colombi M. Dermal fibroblast-to-myofibroblast transition sustained by αvß3 integrin-ILK-Snail1/Slug signaling is a common feature for hypermobile Ehlers-Danlos syndrome and hypermobility spectrum disorders. Biochim Biophys Acta Mol Basis Dis 2018;1864:1010–23. [CrossRef]
- Chiarelli N, Zoppi N, Ritelli M, Venturini M, Capitanio D, Gelfi C, et al. Biological insights in the pathogenesis of hypermobile Ehlers-Danlos syndrome from proteome profiling of patients’ dermal myofibroblasts. Biochim Biophys Acta Mol Basis Dis 2021;1867. [CrossRef]
- Johnson LA, Sauder KL, Rodansky ES, Simpson RU, Higgins PD. CARD-024, a vitamin D analog, attenuates the pro-fibrotic response to substrate stiffness in colonic myofibroblasts. Exp Mol Pathol. 2012 Aug;93(1):91-8. Epub 2012 Apr 18. PMID: 22542712; PMCID: PMC6443252. n.d. [CrossRef]
- da Silva TM, Seabra LMJ, Colares LGT, de Araújo BLPC, Pires VCDC, Rolim PM. Risk assessment of pesticide residues ingestion in food offered by institutional restaurant menus. PLoS One. 2024 Dec 18;19(12):e0313836. PMID: 39693277; PMCID: PMC11654986. n.d. [CrossRef]
- Magulova K, Priceputu A. Global monitoring plan for persistent organic pollutants (POPs) under the Stockholm Convention: Triggering, streamlining and catalyzing global POPs monitoring. Environ Pollut. 2016 Oct;217:82-4. Epub 2016 Jan 18. PMID: 26794340. n.d. [CrossRef]
- Wei D, Shi J, Chen Z, Xu H, Wu X, Guo Y, Zen X, Fan C, Liu X, Hou J, Huo W, Li L, Jing T, Wang C, Mao Z. Unraveling the pesticide-diabetes connection: A case-cohort study integrating Mendelian randomization analysis with a focus on physical activity’s mitigating effect. Ecotoxicol Environ Saf. 2024 Sep 15;283:116778. Epub 2024 Jul 26. PMID: 39067072. n.d. [CrossRef]
- Ospina M, Schütze A, Morales-Agudelo P, Vidal M, Wong LY, Calafat AM. Temporal trends of exposure to the herbicide glyphosate in the United States (2013-2018): Data from the National Health and Nutrition Examination Survey. Chemosphere. 2024 Sep;364:142966. Epub 2024 Jul 27. PMID: 39074666; PMCID: PMC11590049. n.d. [CrossRef]
- McGwin G Jr, Griffin RL. An ecological study regarding the association between paraquat exposure and end stage renal disease. Environ Health. 2022 Dec 12;21(1):127. PMID: 36503540; PMCID: PMC9743741. n.d. [CrossRef]
- Wang K, Zhang C, Zhang B, Li G, Shi G, Cai Q, Huang M. Gut dysfunction may be the source of pathological aggregation of alpha-synuclein in the central nervous system through Paraquat exposure in mice. Ecotoxicol Environ Saf. 2022 Nov;246:114152. Epub 2022 Oct 3. PMID: 36201918. n.d. [CrossRef]
- Sanmarco LM, Chao CC, Wang YC, Kenison JE, Li Z, Rone JM, Rejano-Gordillo CM, et al. Identification of environmental factors that promote intestinal inflammation. Nature. 2022 Nov;611(7937):801-809. n.d. [CrossRef]
- Chen C, Qi M, Zhang W, Chen F, Sun Z, Sun W, Tang W, Yang Z, Zhao X, Tang Z. Taurine alleviated paraquat-induced oxidative stress and gut-liver axis damage in weaned piglets by regulating the Nrf2/Keap1 and TLR4/NF-κB signaling pathways. J Anim Sci Biotechnol. 2025 Aug 18;16(1):117. PMID: 40826133; PMCID: PMC12359926. n.d. [CrossRef]
- Sadighara P, Mahmudiono T, Marufi N, Yazdanfar N, Fakhri Y, Rikabadi AK, Khaneghah AM. Residues of carcinogenic pesticides in food: a systematic review. Rev Environ Health. 2023 Jun 6;39(4):659-666. PMID: 37272608. n.d. [CrossRef]
- Yang Z, Wang M, Cao L, Liu R, Ren Y, Li L, Zhang Y, Liu C, Zhang W, Nie S, Sun Z. Interference with connective tissue growth factor attenuated fibroblast-to-myofibroblast transition and pulmonary fibrosis. Ann Transl Med. 2022 May;10(10):566. PMID: 35722387; PMCID: PMC9201195. n.d. [CrossRef]
- Tyagi N, Singh DK, Dash D, Singh R. Curcumin Modulates Paraquat-Induced Epithelial to Mesenchymal Transition by Regulating Transforming Growth Factor-β (TGF-β) in A549 Cells. Inflammation. 2019 Aug;42(4):1441-1455. PMID: 31028577. n.d. [CrossRef]
- Jeong JY, Kim B, Ji SY, Baek YC, Kim M, Park SH, Kim KH, Oh SI, Kim E, Jung H. Effect of Pesticide Residue in Muscle and Fat Tissue of Pigs Treated with Propiconazole. Food Sci Anim Resour. 2021 Nov;41(6):1022-1035. Epub 2021 Nov 1. PMID: 34796328; PMCID: PMC8564320. n.d. [CrossRef]
- Nguyen TP, Xie Y, Garfinkel A, Qu Z, Weiss JN. Arrhythmogenic consequences of myofibroblastmyocyte coupling. Cardiovasc Res 2012;93:242–51. [CrossRef]
- Grafstein B, Liu S, Cotrina ML, Goldman SA, Nedergaard M. Meningeal Cells Can Communicate with Astrocytes by Calcium Signaling. vol. 47. 2000.
- Sui G-P, Wu C, Roosen A, Ikeda Y, Kanai AJ, Fry CH. Modulation of bladder myofibroblast activity: implications for bladder function. Am J Physiol Renal Physiol 2008;295:688–97. [CrossRef]
- Follonier L, Schaub S, Meister JJ, Hinz B. Myofibroblast communication is controlled by intercellular mechanical coupling. J Cell Sci. 2008 Oct 15;121(Pt 20):3305-16. Epub 2008 Sep 30. PMID: 18827018. n.d. [CrossRef]
- Adegboyega PA, Mifflin RC, DiMari JF, Saada JI, Powell DW. Immunohistochemical study of myofibroblasts in normal colonic mucosa, hyperplastic polyps, and adenomatous colorectal polyps. Arch Pathol Lab Med. 2002 Jul;126(7):829-36. PMID: 12088453. n.d. [CrossRef]
- Andoh A, Fujino S, Okuno T, Fujiyama Y, Bamba T. Intestinal subepithelial myofibroblasts in inflammatory bowel diseases. J Gastroenterol. 2002 Nov;37 Suppl 14:33-7. PMID: 12572863. n.d. [CrossRef]
- Neuhaus J, Pfeiffer F, Wolburg H, Horn LC, Dorschner W. Alterations in connexin expression in the bladder of patients with urge symptoms. BJU Int. 2005 Sep;96(4):670-6. PMID: 16104929. n.d. [CrossRef]
- Kohl P, Camelliti P, Burton FL, Smith GL. Electrical coupling of fibroblasts and myocytes: relevance for cardiac propagation. J Electrocardiol. 2005 Oct;38(4 Suppl):45-50. PMID: 16226073. n.d. [CrossRef]
- Camelliti P, Green CR, LeGrice I, Kohl P. Fibroblast network in rabbit sinoatrial node: structural and functional identification of homogeneous and heterogeneous cell coupling. Circ Res. 2004 Apr 2;94(6):828-35. Epub 2004 Feb 19. PMID: 14976125. n.d. [CrossRef]
- Camelliti P, Green CR, Kohl P. Structural and functional coupling of cardiac myocytes and fibroblasts. Adv Cardiol. 2006;42:132-149. PMID: 16646588. n.d. [CrossRef]
- Andoh A, Ogawa A, Bamba S, Fujiyama Y. Interaction between interleukin-17-producing CD4+ T cells and colonic subepithelial myofibroblasts: what are they doing in mucosal inflammation? J Gastroenterol. 2007 Jan;42 Suppl 17:29-33. PMID: 17238023. n.d. [CrossRef]
- Kayal C, Moeendarbary E, Shipley RJ, Phillips JB. Mechanical Response of Neural Cells to Physiologically Relevant Stiffness Gradients. Adv Healthc Mater. 2020 Apr;9(8):e1901036. Epub 2019 Dec 2. PMID: 31793251; PMCID: PMC8407326. n.d. [CrossRef]
- Lantoine J, Grevesse T, Villers A, Delhaye G, Mestdagh C, Versaevel M, Mohammed D, Bruyère C, Alaimo L, Lacour SP, Ris L, Gabriele S. Matrix stiffness modulates formation and activity of neuronal networks of controlled architectures. Biomaterials. 2016 May;89:14-24. Epub 2016 Feb 26. PMID: 26946402. n.d. [CrossRef]
- Gu Y, Ji Y, Zhao Y, Liu Y, Ding F, Gu X, Yang Y. The influence of substrate stiffness on the behavior and functions of Schwann cells in culture. Biomaterials. 2012 Oct;33(28):6672-81. Epub 2012 Jun 25. PMID: 22738780. n.d. [CrossRef]
- Baghdadi MB, Houtekamer RM, Perrin L, Rao-Bhatia A, Whelen M, Decker L, Bergert M, Pérez-Gonzàlez C, Bouras R, Gropplero G, Loe AKH, Afkhami-Poostchi A, Chen X, Huang X, Descroix S, Wrana JL, Diz-Muñoz A, Gloerich M, Ayyaz A, Matic Vignjevic D, Kim TH. PIEZO-dependent mechanosensing is essential for intestinal stem cell fate decision and maintenance. Science. 2024 Nov 29;386(6725):eadj7615. Epub 2024 Nov 29. PMID: 39607940. n.d. [CrossRef]
- Wang F, Knutson K, Alcaino C, Linden DR, Gibbons SJ, Kashyap P, Grover M, Oeckler R, Gottlieb PA, Li HJ, Leiter AB, Farrugia G, Beyder A. Mechanosensitive ion channel Piezo2 is important for enterochromaffin cell response to mechanical forces. J Physiol. 2017 Jan 1;595(1):79-91. Epub 2016 Aug 13. PMID: 27392819; PMCID: PMC5199733. n.d. [CrossRef]
- Alcaino C, Knutson KR, Treichel AJ, Yildiz G, Strege PR, Linden DR, Li JH, Leiter AB, Szurszewski JH, Farrugia G, Beyder A. A population of gut epithelial enterochromaffin cells is mechanosensitive and requires Piezo2 to convert force into serotonin release. Proc Natl Acad Sci U S A. 2018 Aug 7;115(32):E7632-E7641. Epub 2018 Jul 23. PMID: 30037999; PMCID: PMC6094143. n.d. [CrossRef]
- Sperber AD, Atzmon Y, Neumann L, Weisberg I, Shalit Y, Abu-Shakrah M, Fich A, Buskila D. Fibromyalgia in the irritable bowel syndrome: studies of prevalence and clinical implications. Am J Gastroenterol. 1999 Dec;94(12):3541-6. PMID: 10606316. n.d. [CrossRef]
- Slim M, Calandre EP, Rico-Villademoros F. An insight into the gastrointestinal component of fibromyalgia: clinical manifestations and potential underlying mechanisms. Rheumatol Int. 2015 Mar;35(3):433-44. Epub 2014 Aug 14. PMID: 25119830. n.d. [CrossRef]
- Wang XJ, Ebbert JO, Loftus CG, Rosedahl JK, Philpot LM. Comorbid extra-intestinal central sensitization conditions worsen irritable bowel syndrome in primary care patients. Neurogastroenterol Motil. 2023 Apr;35(4):e14546. Epub 2023 Feb 19. PMID: 36807964. n.d. [CrossRef]
- Doggweiler-Wiygul R. Urologic myofascial pain syndromes. Curr Pain Headache Rep. 2004 Dec;8(6):445-51. PMID: 15509457. n.d. [CrossRef]
- Drossman DA. Do psychosocial factors define symptom severity and patient status in irritable bowel syndrome? Am J Med. 1999 Nov 8;107(5A):41S-50S. PMID: 10588172. n.d. [CrossRef]
Figure 1.
the basic molecular pathway of the suggested hypothesis with various inputs into the positive feedback loop of myofibroblast activity. A vitamin D analogue was shown to attenuate the pro-fibrotic response to substrate stiffness in colonic myofibroblasts [51].
Figure 1.
the basic molecular pathway of the suggested hypothesis with various inputs into the positive feedback loop of myofibroblast activity. A vitamin D analogue was shown to attenuate the pro-fibrotic response to substrate stiffness in colonic myofibroblasts [51].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



