Submitted:
01 November 2025
Posted:
03 November 2025
You are already at the latest version
Abstract
Enlarged perivascular spaces (EPVS) within the perivascular unit (PVU) provide a holding space, which provides a crossroad intersection. These EPVS provide a space for multiple cell cell contact interactions and communications for the six cells discussed in this narrative review. These six cells include the brain endothelial cells, mural supportive pericytes, and the supportive perivascular astrocyte endfeet (pvACef) to provide the 1st barrier of the blood-brain barrier of the neurovascular unit and the resident perivascular macrophage of the EPVS along with the outermost barrier pvACef cells of the PVU that provide the 2nd and final barrier to prevent neuroinflammation and the delivery of excess fluids and neurotoxins to the interstitium of the neural parenchymal cells. The final two cells are represented by the interrogating and/or reactive microglia and neurons along with their synapses to provide brain homeostasis. When these cells become damaged or dysfunctional due to multiple injurious species, they contribute to EPVS that are associated with many neurological diseases including both neurodevelopmental and neurodegenerative disorders that are associated with small vessel disease. The use of multiple transmission electron microscopic images along with illustrations are utilized to better understand the different functional and aberrant structural remodeling mechanisms. Indeed, the neuroglia perivascular and perisynaptic astrocytes are at the very center of providing not only the mesmerizing image but also play an important and critical role in neurodevelopmental and neurodegenerative diseases. The primary goal of this narrative review is to increase our holistic understanding of ultrastructural remodeling changes and functional mechanistic knowledge of these cells and spaces.
Keywords:
blood-brain barrier
; brain endothelial cells
; enlarged perivascular spaces
; glymphatic system
; late-onset Alzheimer’s disease
; neurodegeneration
; pericytes
; perivascular macrophages
; reactive perivascular astroglia
; small vessel disease
1. Introduction
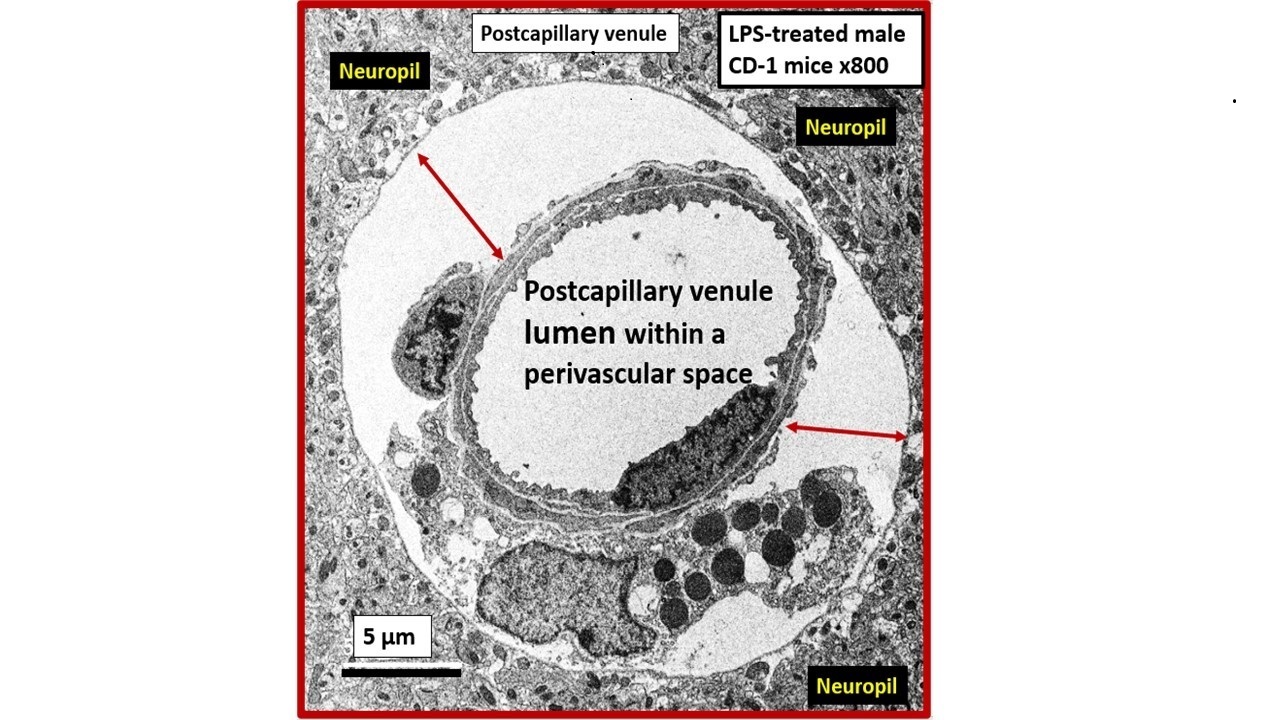
Transmission Electron Microscopy (TEM) remains a powerful tool to investigate cellular and tissue remodeling changes in disease and various treatment models [1]. Indeed, author has examined hundreds of cortical, frontal layer III capillary neurovascular unit(s) (NVU) in diet-induced obesity, insulin resistance, metabolic syndrome, genetically induced female, obese, diabetes models (db/db and BTBR ob/ob), streptozotocin-induced diabetes in male, and control preclinical mouse models. These capillary NVUs demonstrated that the glial protoplasmic perivascular astrocytes endfeet were tightly adherent to the mural cells (endothelial and pericytes) shared basement membrane. Thus, the finding of a NVU with a definite sharply demarcated space separating the NVU capillary mural cells and their basal lamina/basement membrane from the surrounding-encompassing protoplasmic perivascular astrocyte endfeet basal lamina within the neuropil was indeed remarkable and mesmerizing as in the following observed original image from the neuroinflammatory lipopolysaccharide (LPS)-treated model as compared to control models (Figure 1) [2].
Notably, LPS-induced neuroinflammation in CD-1 male mice also depicted multiple response to injury ultrastructural remodeling changes including BECs activation supported by plasma membrane ruffling, aberrant mitochondria, increased macro-micropinocytosis (transcytosis), and increased extracellular vesicle formation. Pericyte(s) (Pc) nuclear rounding with retraction of Pc foot processes-endfeet (Pcfp) with increased lysosome, vacuoles, and vesicles numbers. Additionally, there were increased reactive amoeboid microglia cell attractions to the NVU note shown in Figure 1B [2]
The cellular components of the capillary NVU consists of brain endothelial cell(s) (BEC), Pcs and artery/arteriole vascular smooth muscle cells, protoplasmic perivascular astrocytes and their endfeet (pvACef), neurons, and interrogating microglial cell(s) (iMGC) as depicted (Figure 1) [3,4,5,6,7]. The recently described perivascular unit (PVU) consists of benign perivascular spaces (PVS) and the pathologically dilated or enlarged perivascular spaces (EPVS) identified in neuroinflammatory LPS-treated mice models (Figure 1B) [3,8] The PVS are bound by the innermost boundary created by NUV shared basement membrane of the BECs and Pcs with the outermost boundary formed by the compacted protoplasmic perivascular endfeet basement membrane also known as the glia limitans [8,9]. Importantly. resident perivascular macrophages represent the sixth cell that are known to reside within PVS/EPVS (Figure 1B) [10,11,12].
Descending pial arteries and arterioles PVS are important in delivering the cerebrospinal fluid (CSF) to the brain interstitial spaces (ISS) and interstitial fluid (ISF) of the parenchyma while the postcapillary ascending venular PVS are responsible for the efflux of the interstitial fluid (ISF) and its contents including some residual CSF, metabolic waste, solutes, and excess fluids/water to the subarachnoid space (Figure 2) [3,13,14,15].
Indeed, the PVS of the postcapillary venules, venules, and veins are responsible for the efflux of ISF with its metabolic/phagocytic waste and excess water to the subarachnoid space and hence to the peripheral circulation in addition to providing a conduit for the glymphatic system (GS) in the brain [16,17,18]. PVS are currently thought to form and mature postnatally under the influence of perivascular astrocytes along with other cellular components of the NVU. Perivascular astrocytes play a crucial role in shaping and maintaining the integrity and function of not only the NVU but also PVS. However, the complete mechanisms regarding the formation of PVS still remain an area of ongoing understanding and research [17,18].
It is important to define the GS since this narrative review focuses primarily on the post-capillary venule PVS as the existing anatomical conduit for the GS. Some have simply described the GS as the brain’s waste clearance system. However, the GS may be considered to be comprised by a brain-wide network of perivascular spaces/channels/conduits and their adjoining vascular luminal shared BEC and Pc basement membranes in postcapillary venules; vascular smooth muscle cell basement membranes, interstitium, and adventitia in arteries and arterioles of the intramural periarterial drainage (IPAD) system and specifically the outermost delimiting pvACef compacted basement membranes or glia limitans of the parenchymal components, which function to facilitate the exchange and the eventual clearance of the brain’s metabolic waste solutes carried in the interstitial fluid [18,19]. Further, the GS facilitates the exchange of fluid and solutes between the CSF and interstitial compartments organized along perivascular pathways and supported by arterial pulsation, vasomotion, respiration, and facilitated by pvACef AQP4 water channels [16,17,18,19]. Thus, this definition allows for not only the glymphatic system efflux hypothesis but also the retrograde IPAD efflux and mixing hypotheses within these PVS to be included. Importantly, the GS is thought to be related to two structures: the PVS and its outermost barrier/boundary formed by pvACef with their aquaporin 4 (AQP4) water channels [20].
When PVS (Virchow-Robin spaces) are large enough to be identified with magnetic resonance imaging (MRI), they are termed MRI-visible enlarged perivascular space (MVEPVS) that measure one to three millimeters in diameter that are usually symmetrically located within the basal ganglia or the centrum semiovale regions of brain MRIs (Figure 3) [3,21].
Importantly, EPVS/MVEPVS are known to be biomarkers for cerebral small vessel disease (SVD) that associate with multiple (at least a dozen or more) clinical neurologic diseases as follows: Alzheimer’s disease, Parkinson’s disease, Hunignton’s disease, multiple sclerosis, encephalitis (infectious or traumatic), traumatic brain injuries, post-traumatic stress disorder (still remaining to be fully established), autism spectrum disorders (still remaining to be fully established), migraine headaches, LPS infusions related to neuroinflammatory models or clinically related LPS excess due to gut dysbiosis [2,3], hypertension, and neurovascular clinical diseases that include: stroke (ischemic or hemorrhagic, transient ischemic attacks/events (TIAs), vascular dementia (VaD), vascular contributions to cognitive impairment and dementia (VCID), cerebral amyloid angiopathy (CAA), cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy (CADASIL), neuromyelitis optica and/or neuromyelitis optica spectrum disorder, and normal aging [3,22].
SVD consists of a variety of aberrant structural and functional changes involving small cortical perforating arterioles, precapillary arterioles, capillaries, and ascending postcapillaries venules and veins in the brain [3]. The two most common pathological changes in SVD include cerebral arteriolosclerosis (hyalinosis and lipohyalinosis) and CAA. These changes may be divided into multiple categories identifiable by non-invasive MRI, which include enlarged perivascular spaces, cerebral microbleeds, small recent cortical infarcts, lacunes, and white matter hyperintensities (Box 1) [3,15,21,22,24,25,26,27,28,29].
The development of EPVS is complex and involves multiple functional and cellular mechanisms that are highly interconnected. These mechanisms include but are not limited to the following: 1). Brain endothelial cell activation/dysfunction (BECact/dys) with increased 2). Vascular inflammation; 3). Oxidative stress; 4). Neuroinflammation with decreased BEC protective nitric oxide and increased permeability of fluids (including ions, water, proteins including hormones, and polypeptides) and cells including leukocytes into the PVS due to endothelial blood-brain barrier dysfunction and disruption (BBBdd); 5). Microvessel stiffening especially in the arterial system with a loss of pulsatility that is important to drive the glymphatic system flow of waste and water clearance; 6). Astrocyte and pvACef dysfunction and or loss with decreased or dysfunctional aquaporin 4 (AQP4) water channel activity due to translocation from the plasma membrane or damage; 7). Neuroinflammation; 8). Oxidative stress with increased reactive oxygen, nitrogen, sulfur species; 9). Leukocyte accumulation within the PVS/PVU/GS; 10). Phagocytic debris accumulation of proteins and protein aggregation (specifically amyloid beta and tau), which disrupt GS drainage that result in EPVS; 11). Brain atrophy [3,9,15,22,24].
There are multiple reasons as to why PVS are important and at least eight core reasons are presented (Box 2) [3].
Even though there may exist an ongoing debate by some regarding whether these PVS are true spaces or only potential spaces, the current anatomical concepts suggest that PVS are indeed compartments between the outermost basement membranes of pvACef and innermost NVU mural cell (BEC and Pc) basement membranes. Regardless, author feels that the importance of EPVS will continue to grow along with the evolving role of the GS because current papers regarding these two mechanisms are growing exponentially in the literature as illustrated by the increasing number of published papers regarding these two interrelated systems identified in the National Institutes of Health (NIH) search engine PubMed [30].
The primary aim of this narrative is to increase the holistic understanding of the cells involved in the development of EPVS and SVD, while exploring the interrelationships of neurovascular, neuroimmune, and neurodegenerative mechanisms.
2. Cellular Involvement of the Neurovascular and Perivascular Unit
2.1. Brain Endothelial Cell(s) (BEC)
BECs are the first barrier between the blood and the brain. They provide the semipermeable interface-blood-brain barrier (BBB) (transfer and diffusion of solutes and fluids), trophic (metabolic homeostasis), tonic (vascular hemodynamics), and trafficking (vascular permeability, coagulation, and leukocyte extravasation) functions to provide brain homeostasis [31,32]. Further, BECs have a paucity of pinocytic vesicles, an absence of fenestrae, and the unique expression of paracellular tight and adherens junctions as compared to systemic endothelial cells [31,32,33,34]. BECs are constantly exposed to numerous injurious stimuli and are known to respond to injury via an innate response to injury mechanism (Figure 4) [31,35].
Indeed, the central theme of brain injury involves gene activation and transcription of various factors in a response to injury wound healing mechanisms, which include hemostasis, inflammation, proliferation growth (re-growth), hypertrophy, differentiation, development, remodeling, and limited neural regeneration. If the injury is brief there will be regeneration; however, if the injurious stimuli are chronic there will be aberrant remodeling of cells and tissue to result in dysfunction and disease instead of regeneration and functional restoration (Figure 5) [35].
For example, in the case of LPS-treated models the BEC are injured and respond accordingly via TLR4 and NF-kB signaling to result in BECact/dys and BBBdd to result in increased permeability, neuroinflammation, and the formation of EPVS (Figure 6) [2,35,36,37,38].
Thus, injurious species to BECs of the NVU result in BECact/dys and BBBdd with increased permeability of neurotoxins and leukocytes into the adjacent PVU to allow multiple cellular crosstalk with resident and reactive antigen presenting perivascular macrophage(s) (rPVMΦ) and pvACef. This multicellular crosstalk within the PVS of the PVU results in increased oxidative stress, activation of matrix metalloproteinases (MMPs) to digest the outermost protective barrier of the pvACef glia limitans in the two-step process of neuroinflammation described by Owens et al [10]. Further, this allows BECact/dys-BBBdd to propagate neuroinflammation along with reactive atMGCs to increase cnsCC that may set in motion a self-perpetuating vicious cycle to result in EPVS and impaired waste and water clearance via the GS [3,8,9,10,11,12,14,17,22,29,30,39].
Importantly, the BEC glycocalyx plays a crucial role in protecting the capillary NVU and its BBB. The glycocalyx provides a surface coating, which consists of a complex interwoven polysaccharide mesh-like network of membrane-bound proteoglycans, glycoproteins, and glycosaminoglycans covering the apical luminal layer of the brain endothelial cells [40ecGCx]. It acts as the first barrier to circulating blood in the tripartite BBB consisting of (1) BEC glycocalyx; (2) BECs with their basement membranes; (3) an extravascular compartment of pericytes, the extracellular matrix, and perivascular astrocytes and their basement membranes [40,41]. Perturbations of this glycocalyx barrier (multiple injurious species including LPS) allow for increased permeability in the postcapillary venule that will be permissive to fluids, solutes, and proinflammatory peripherally derived leukocytes into the PVS which result in EPVS as well as increased neuroinflammation and neurodegeneration. LPS administration is known to cause endothelial glycocalyx injury (Figure 7) [42].
2.2. Capillary Neurovascular Unit Pericyte(s) (Pc)
Pcs along with BECs are vascular mural cells embedded in the basement membrane of blood microvessels and provide support to the capillary NVU. They contribute to the formation of the BBB tight and adherens junctions as well as to the shared basement membrane of BECs. Also, Pcs are uniquely positioned within the NVU to interact with BECs and pvACef to integrate, coordinate, and process signals from their neighboring cells to generate diverse functional responses that are critical for CNS functions in health and disease (Figure 1) [43,53]. These responses and interactions include the regulation of the blood-brain barrier permeability, angiogenesis, clearance of toxic metabolites, capillary hemodynamic responses (NVU coupling with vasorelaxation and constriction), neuroinflammation, and mesenchymal stem cell activity (Figure 8) [43,44,45,46,47,48,49,50,51,52,53,54].
Notably, some have elevated the roles of the BECs to be termed the ‘gatekeepers’ and/or the ‘sentinel’ cells, while the Pcs have been given the role of being the ‘guardian angel’ because of their critical support and protection they provide to BECs and NVUs. Further, these mural cells (BECs and Pcs) are co-dependent in that the BECs supply nitric oxide and platelet-derived growth factor beta (PDGFβ) for the Pc, while the Pc provides the BECs with VEGF for angiogenesis and also the receptor for PDGFβ [43,52].
2.3. Perivascular Astrocyte Endfeet (pvACef)
Astrocytes-astroglia are the most abundant glial cells and may be considered the master communicating, connecting, and creating cells within the brain (Figure 9) [55].
Cortical grey matter protoplasmic astrocyte(s) (AC) have many important homeostatic functions in the brain including neurovascular coupling to match cerebral blood flow to neuronal activity. Additionally, ACs are a major supplier of energy to neurons (via glycogen storage and glycolysis to produce glucose and lactate) and are a rich source of antioxidants such as glutathione and superoxide dismutase. They also provide growth factors such as brain-derived growth factor, transforming growth factor beta and glial-derived growth factor [56,57,58,59]. ACs also define many aspects of synapse formation, plasticity, provide protective functions, synaptic maintenance and elimination [60]. Further, ACs are responsible for controlling the fluid volume of the brain including the interstitial fluid, interstitial fluid spaces, and PVS via the highly polarized plasma membrane water channel AQP4. This bidirectional water transport function of AQP4 in the plasma membrane of pvACef that faces the brain spaces including PVS is essential for the homeostasis of water balance in the brain [56]. Unfortunately, when these pvACef become reactive they lose their polarity of the bidirectional water channel resulting in dysfunction and fluid retention and expansion of spaces [61]. Notably findings in rodent models may not always translate well to human studies since human ACs are more complex, larger in size, and extend their cytoplasmic processes much farther when compared to rodent model brains [62].
Protoplasmic pvACef are known to tightly adhere to the basement membranes of the NVU shared by the BECs and Pcs of the capillary NVU as in Figure 1A [55,63,64]. However, LPS-treated models demonstrated that the pvACef became detached and separated from the NVU basement membranes [2], which would impair NVU coupling, similar to the findings associated with obesity, insulin resistance, and hyperglycemia as in the diabetic db/db female models [35]. pvACef are communicating cells via gap junction connexins Cx40, Cx43 and create a syncytium, connecting cells as in Figure 8, coupling cells as in NVU coupling and PVU coupling, and continuing and creating cells, such that they become continuous at the true capillary as it transitions to the postcapillary venule with normal PVUs, and pvACef that are continuous and creative in the development of the PVU, PVS/EPVS (Figure 9) [30,55].
It is important to note that the initial barrier of the PVU/PVS is the BEC BBB and the 2nd and final barrier of the PVS is the glia limitans or compacted basement membrane of the outermost pvACef. This places the pvACef as the critical barrier to prevent the extravasation of proinflammatory leukocytes into the parenchyma of the brain that results in the damaging role of neuroinflammation and its relation to neurodegeneration and impaired cognition [10Owens, 55HaypvACs]. Additionally, the outermost pvACef and their AQP4 water channels are crucial for the proper function of GS waste and water removal system [9,30,55,56]. Indeed, the PVU provides a crossroad and intersection holding area for the dynamic interactions of the vascular, immune and neuronal mechanisms to respond to injury that initially occurs at the BECs of the NVU in addition to the important glymphatic system of waste and excess water removal [8,9,27,65,66]
2.4. Perisynaptic Astrocyte Endfeet (psACef)
Perisynaptic astrocyte endfeet (psACef) provide synaptic plasticity, connectivity, and are important in information processing. They are responsible for forming the tripartite synapse including the presynaptic and postsynaptic terminals along with the cradling psACef, which are essential for synaptic maturation, isolative protection, synaptogenesis, and maintenance of synapses. Further, these functions provide for the synaptic fundamental mechanisms of not only connectivity and plasticity but also informational processing in the brain [35,55,56,58,60]
psACef may become detached similar to their detachment from the capillary NVU as a result of brain injuries that associate with and result from inflammation, oxidative stress with activation of regional proteases (MMP-2, 9) that allow for damage and dysfunction of the anchoring dystroglycans to allow for the psACef to detach and separate from the pre- and postsynaptic terminals of the tripartite synapse [35,55,56,58,60]. Detached and separated psACef will create a perisynaptic space (PSS) that may be referred to as the enlarged perisynaptic spaces (EPSS) similar to EPVS Additionally, these reactive detached and separated psACef could result in the translocation of the bidirectional water channel AQP4 to be translocated and dysfunctional with the possibility of excessive fluid contributing to even further psACef separation from the pre- and postsynaptic terminals (Figure 10) [35,55,60,67,68].
psACef support tripartite formation and function of synapses (synaptic transmission), neurotransmitter recycling, BBB, and the support of oligodendrocytes, which produce myelin [66Mz]. Indeed, if injury mechanisms (injurious species as in Figure 1) to either the NVU or the dendritic synapses are chronic there would be loss of NVU coupling with resulting regional hypoxia and/or hypometabolism with ensuing neuronal dysfunction and neurodegeneration with impaired cognition, especially in the very young or very old vulnerable individuals to result in multiple diseases such as autism spectrum disorder and LOAD respectively. Similarly, synaptic psACef detachment and separation could interfere with glutamate uptake at the involved synapses and the glutamate–glutamine cycle and thus, could contribute to glutamate excitotoxicity to further increase regional injury to perpetuate a chronic cycle of synaptic and synaptic plasticity - dysfunction and/or loss (synaptopathy) [55,69,70,71].
2.5. Resident Perivascular Macrophages (PVMΦ)
NVUs and PVUs/PVS provide a crossroad and an opportunity for multicellular crosstalk that eventually allow for the breech of the outermost glia limitans of the reactive pvACef and reactive PVMΦs (rPVMΦs) with subsequent neuroinflammation via its antigen presenting cell (APC) capabilities (Figure 11) [8,10,30,55,72].
Importantly, PVMΦs are only found in the PVS that lie immediately adjacent to the NVUs, which allow them the ability to have intimate contacts with the mural cells (BECs and Pcs) as well as the outermost pvACef boundary cells of the PVS in addition to their direct exposure and interactions with the CSF and ISF within PVS/EPVS [8,10,72]. Additionally, rPVMΦs are capable of interacting with the leukocytes that are delivered to the PVS once the BBB has been compromised by multiple injurious species due to BECact/dys and BBBdd as previously presented (Figure 5; section 2.1.) [8,10,72,73,74]. Also, PVMΦ are yolk sac-derived innate immune cells that are important for the development of neuroinflammation [73,74,75,76]. Clinically, previous publications have shown that PVMΦs are involved in the pathology of AD [77], PD [78], stress [79], obesity [80s], stroke [81], hypertension [82], and metainflammation associated with insulin resistance, MetS, and diabetes [72]
PVMΦs are considered as a specialized subset of border-associated macrophages (BAMs), which also include border-associated macrophages as also identified in the meninges and the choroid plexus (epiplexis and Kolmer cells) [8,10,83,84]. Indeed, the brain’s border interfaces are important hubs for not only the uptake and efflux of solutes, ions, and cells but also provide the necessary and essential hubs for neuroimmune regulation (PVS/EPVS, meninges, choroid plexuses, and circumventricular organs).
3. Perivascular Spaces (PVS) of the Perivascular Unit (PVU), EPVS, and Interstitial Fluid (ISF)
According to the ancient Chinese Taoist/Daoist philosophy a space is not an absence or a void but acts as a dynamic and essential element of existence, harmony, and homeostasis [85]. To this end, PVS provide not only a crossroad and a specific space for multicellular crosstalk but also provide for the natural flow of fluids (pial arterial CSF influx and postcapillary venular ISF efflux) including water as well as metabolic and phagocytic waste of the GS within this enclosure. When PVS are healthy they are seldom observed by MRH; however, in multiple neurologic pathologies these PVS undergo aberrant structural remodeling to become EPVS/ MVEPVS. Importantly, PVS function as an efflux system or GS for waste and excess water removal in postcapillary venules [2,8,9,17,30].
PVMΦs normally reside within the confines of PVS/EPVS (section 2.4.) [10]. These PVMΦs constantly surveille the PVU spaces and are capable of not only being an antigen presenting cell but also capable of undergoing reactive changes to stressful stimuli as occurs with BECact/dys and BBBdd. They typically stain positive for CD206 and CD163 that are seldom stained in border-associated microglia cells. These PVMΦs allow the PVS their ability to act as a unit to function as a neuromodulator within the brain to prime the neuroimmune system.
The development of EPVS is complex and involves multiple functional and cellular mechanisms that are highly interconnected. These mechanisms include but are not limited to the following: 1). BECact/dys due to multiple injurious species within precapillary arterioles, capillary NVU’s, postcapillary venules, and the subsequent development of increased 2). Vascular inflammation; 3). Oxidative stress; 4). Neuroinflammation with decreased BEC protective nitric oxide (BEC dysfunction) and increased permeability of fluids (including ions, water, proteins including hormones, and polypeptides) and cells including leukocytes into the PVS due to endothelial blood-brain barrier dysfunction and disruption (BBBdd); 5). Microvessel stiffening due to increased pulsatile pressure resulting in microvessel remodeling with further stiffening in the arterial system resulting in impaired pulsatile flow that is important to drive the glymphatic system flow to promote waste and water clearance from the perivascular spaces. 6). Glial pvACef dysfunction and or loss with decreased or dysfunctional AQP4 activity; 7). Neuroinflammation; 8). Oxidative stress with increased reactive oxygen (ROS), nitrogen, sulfur species (RONSS); 9). Leukocyte accumulation within the PVS/PVU/GS; 10). Phagocytic debris accumulation of proteins, misfolded proteins, and protein aggregation (specifically amyloid beta and tau), which disrupt GS waste and excess water removal resulting in stalled flow that further dilates the PVS to remodel into EPVS proximal to regions of stalled flow. Thus, the EPVS structural crossroad and holding space allow for interactions between the neurovascular, neuroimmune, and neuronal mechanisms that relate to clinically relevant neurodevelopmental and neurodegenerative disorders [3,8,9,10,15,23,24,27,30,35,39,55]. In addition to the previous 10 mechanisms, one overarching theme seemed to reoccur in the development and progression of EPVS. This theme involved the response to injury that occurred at the capillary NVU BEC initial injury and the subsequent response to injury aberrant remodeling. The overarching finding of the NVU BECs to develop BECact/dys with subsequent BBBdd with increased permeability was noted to be the primary instigator in the mechanistic evolution of EPVS and SVD [3,8,9,10,30,31,35,40]. The other notable finding was that obstruction to the PVS via excessive metabolic waste, phagocytic debris, phagocytized misfolded and soluble proteins such as amyloid beta and tau, neuroinflammatory cells, or dysfunction of the abluminal perivascular astrocyte endfeet resulting in the dysfunction or loss of their polarized aquaporin-4 water channels might allow for the remodeling obstructive or stalled flow resulting in the proximal dilation of the PVS within the PVU to become EPVS [8,9,10,15,30,86,87,88,89].
The recognition of PVS/EPVS has also brought forth an increased interest in the brain’s interstitial space(s) (ISS) and how they empty into the PVS of the postcapillary venules glymphatic pathway (GS). Even though the ISS diameter is only nanometers in size (e.g. approximately 20–50 nanometers in mouse brains), its total volume is known to account for up to 20% of the total brain volume and is crucial for interstitial fluid (ISF) solute and fluid transport, signal transmission, and communication amongst neuronal parenchyma within the neuropil (Figure 12) [90].
4. Glia and PVS/EPVS Ultrastructural Remodeling in LPS Neuroinflammatory, Glucotoxicity, and Hyperhomocysteinemia (HHCY) Models
In LPS-treated neuroinflammatory models, MGCs have been demonstrated to be attracted to the NVU along with the formation of EPVS [3]. Additionally, MGCs appeared to be invasive to the capillary NUV that was associated with the detachment and separation of pvACef in obese diabetic db/db female models at 20-weeks of age (Figure 13) [91].
Indeed, reactive astrocytes (macroglia) and microglia play an important role in the remodeling of various animal models. Remodeling changes range from attracted MGCs of the NVU in female LPS-treated models [2], to invasive and destructive effects of MGCs that envelope the NVU with subsequent detachment and separation of pvACef in the female obese, insulin resistant, and diabetic db/db models in Figure 13 [91], to the damaged BEC glycocalyx and formation of postcapillary venular enlarged perivascular spaces as identified in the female obese diabetic BTBR.Cg-Lepob/WiscJ ob/ob mouse (BTBR ob/ob) models, glia are actively involved in brain ultrastructural remodeling changes (Figure 14) [92].
In diabetes, elevated brain glucose (neural glucotoxicity) not only undergoes autoxidation to generate ROS but also is capable of forming complex non-enzymatic reactions between reducing sugars and the amino groups of proteins advanced glycation end-products (AGEs), which reacts with the BEC receptor for AGE (RAGE) to generate even larger amounts of ROS along with oxidative damage to proteins, lipids and nucleotides. This glucotoxicity also promotes increased mitochondrial ROS (mtROS) [91]. Importantly, diabetes is known to be associated with increased SVD and EPVS [93,94,95,96,97]. Notably, homocysteine (Hcy) is another injurious species in addition to LPS and glucotoxicity to the BECs of the NVU as in figure. Homocysteine (Hcy) is a physiologic sulfur/sulfhydryl-containing amino acid that is liberated upon protein degradation and breakdown that may lead to a pathological condition called hyperhomocysteinemia (HHCY) especially if there is a nutrient deficiency or impaired uptake of vitamin B cofactors (B6, B9, and B12) that are important for running the folate-methionine cycles of folate one carbon metabolism (FOCM) (Figure 15) [98,99].

When Hcy becomes elevated as in HHCY, it undergoes autoxidation (similar to glucose autoxidation) via the following set of reactions [98,99]: Two homocysteines (2Hcy-SH) plus oxygen yeilds → homocystine: oxidized disulfide, plus two hydrogens. plus two electrons (HcyS-SHcy +2 H+ and 2e-) to generate ROS such as superoxide, hydroxyl radical, and/or hydrgen peroxide. Additionally, formation of mixed disulfides may contribute to the additional formation of ROS such as: Hcy-SH + R-SH ---- O2 -------→ Hcy-S-SR + H2O→ reactive oxygen species or Hcy-SH + R1-S-S-R2 ← → R1-S-S-Hcy + R2-SH → reactive oxygen species where R = any organic compound in the plasma with a thiol group (-SH) accessible to react with Hcy, such as proteins, cysteine, glutathionine, gamma-glutamylcysteine, or cystinylglycine. Further, HHCY may undergo complicated rearrangements to form Hcy thiolactone (a cyclic thioester), which is chemically reactive and acylates free amino groups such as the side-chain lysine groups of proteins (termed N-homocysteinylation), forms an amide bond with the lysine residue and introduces a free thiol group, which can lead to protein damage and is implicated not only cardiovascular disease but also in brain microvessel dysfunction and disease. Further, the process of N-homocysteinylation promotes further ROS/oxidative stress in addition to the homocysteinylated proteins that may become damaged or dysfunctional to result in a loss of their biological activity [98,99,100,101,102].
HHCY is known to be associated with multiple neurological diseases and/or disorders including cognitive impairment [103Elias], cardiovascular disease [104,105,106,107,108,109,110,111,112,113], stroke (ischemic) [102,114], vascular dementia (VaD) [102,115], LOAD [99,102,116], Parkinson’s disease [102,117], schizophrenia, major depressive, and bipolar disorders [102,118], epilepsy and antiepileptic therapies [102,119], and autism spectrum disorder(s) (ASD) [120,121]. Further, it is interesting to note that LPS treatment and HHCY may co-occur and interact synergistically regarding neuroinflammation to result in the interactive vicious cycle between inflammation and oxidative stress [122]
5. Conclusions and Future Directions
Ruoli Chen (guest editor) of this special series in the Neuroglia Journal [https://www.mdpi.com/journal/neuroglia/special_issues/3750Q69655] has put forth the concept that neuroglia (astrocytes and microglia) are at a crossroad from their unifying role of maintaining support and homeostasis in the brain to that of exploring how neuroglia might contribute to disease onset, progression, and potential recovery in multiple neurological diseases and disorders. Indeed, the neuroglia cells are at a definite crossroad along with BECs, Pcs, neurons along with their synapses in addition to neurovascular and perivascular units (NVU and PVU) as discussed in this narrative review.
The PVS within the PVU provide a holding space or a crossroad intersection for multiple cell-cell communications that are bounded by the innermost shared basement membranes of mural cells (BECs and Pcs) and the outmost abluminal basement membranes (glia limitans) of the pvACef. The initial or 1st barrier of PVU consists of the BBB, while the outermost highly protective pvACef basement membrane-glia limitans provide the 2nd barrier. This 2nd barrier created by the pvACef glia limitans prevents excessive fluids, neurotoxins, and proinflammatory cells from entering the interstitial spaces of the brain as in figures 1B,9, 14C, D (Figure 16) [3,8,9,10,30,55].
There are multiple factors responsible for the continuous flow of the admixture of CSF, ISF, and metabolic waste within the PVS including: the continuous production of CSF via the choroid plexuses within the ventricles [14,123], the adjacent pia arterial pulsations [124,125], respiratory motion [126], pvACef-AQP4 water channel proteins [125], and all important sleep-wake cycle (since most glymphatic flow occurs during sleep [14,17,127,128]. Additionally, the psACef also provide a protective barrier to support and protect the synaptic transmission of the tripartite synapses [35,55,60,67,68] as well as the transfer of information throughout multiple informational networks including the default mode network (thought and reflection), central executive network (attention and working memory), and the salience network (important in processing internal and external stimuli) in the brain [129,130,131].
There are certain limitations to any narrative review as they reflect the interests of the author that assist with the knowledge of the topic at hand. Also, this narrative relies heavily upon the use of rodent models to explain and increase the understanding of ultrastructural remodeling changes, which may not always directly translate to human individuals, especially since astrocyte morphology in human individuals are much more complex than in rodent models [62]. Additionally, images were derived primarily from the frontal grey matter cortex and represent primarily layer III and the transition to only the immediate subcortical regions of the brain. Future directions include the need for greater MRI resolution to better identify and understand the venular side of the capillary NVU, which include EPVS and impaired glymphatic waste clearance from the ISF contents of the brain such as the use of 7Tesla instead of 1-3Tesla currently in use [3Why, 9A closer look,]. Also, the venular system with its PVU and PVS are essential for waste removal including the effective removal of neurotoxins including soluble oligomeric amyloid beta, tau, other neurotoxic misfolded proteins, and metabolic waste to slow or prevent neurovascular, neuroinflammatory, and neurodegenerative diseases discussed in this narrative review via the GS that is dependent on the normal functioning of the postcapillary venular PVS as it is known to be the structural conduit for waste removal within the PVU. Additionally, EPVS can now be identified and quantitated via algorithms that have been created for artificial intelligence and deep machine learning, which reduce time and effort, and increase specificity in contrast to earlier visual quantifications to better understand the relationships between EPVS, SVD and impaired cognitive abilities in large, combined cohorts [132].
Regarding future directions, it will be necessary to better understand the development and progression of the recent findings of EPVS in neurodevelopmental disorders such as autism spectrum disorders (ASD). ASD may be broadly defined as a heterogeneous group of neurodevelopmental disorders that are characterized by impaired social interaction and/or communication that are associated with restrictive or repetitive patterns of behavior [133,134]. The recent findings of EPVS in the neurodevelopmental disorder ASD [135,136,137] and the recent increase of ASD have alarmed clinicians, neurologists, neuroscientists (specifically pediatric neuroscientists), the public, and recently governmental agencies [138]. Therefore, it is absolutely necessary to evaluate and better understand why there are increased EPVS in ASD. Not only is ASD associated with EPVS but also Hcy is elevated and known to associate with increased EPVS and is also elevated in ASD as well as being a biomarker for impaired FOCM [99,120,139]. Thus, deficient systemic and brain folate induced either by nutritional deficiency or by the development of autoantibodies to the folate receptor alpha that will impair brain FOCM and result in HHCY as in Figure 15, which acts as an injurious stimulus to BECs due to the excessive autoxidation of Hcy and oxidative stress be examined. Once injured the BECs will undergo both structural and functional changes with the development of BECact/dys and BBBdd with increasing neuroinflammation with the development of EPVS as in figures 9 and 16. These functional and structural remodeling changes to the BECs and EPVS will result in impaired glymphatic system clearance of neurotoxins and metabolic waste to produce an ongoing vicious self-perpetuating cycle of injury and response to injury remodeling. Additionally, HHCY will impair the necessary FOCM donation of methyl groups, which are critically necessary for supporting the rapid neural expansion with neurogenesis, synaptic plasticity. and neural network connectivity in infants and young children due to their positive effects on biosynthesis of nucleic acids, proteins, and lipids required for growth and epigenetic modifications important for regulating gene expression [140,141,142].
While this narrative review has focus primarily on LPS and glucose excess, it is important to note that nutritional deficiencies (deficient folate) or genetic abnormalities including the methylenetetrahydrofolate reductase (MTHFR) mutation gene polymorphism (677C>T) enzyme plays an important role in the folate cycle, and autoantibody formation against the folate receptor alpha also impair FOCM donation of one-carbon units to neurons with resultant neurodevelopmental disorders. Additionally, multiple nutrients are important for proper neurodevelopment [141,142]. Thes nutrients consist of lipids-polarlipids, minerals, vitamins (A, D, B6. B9(folate), B-12), proteins and amino acids, tryptophane, tyrosine, phenylalanine, branched chain amino acids, pro-prebiotics [141].
Treatment for lowering Hcy consists of folic acid or folate supplements (0.5-5 mg folic acid and 0.5mg of vitamin B-12 [143] and/or the newer folinic acid treatments, should there be autoantibodies to the folate receptor alpha [143]. When there are autoantibodies to folate receptor alpha this may be treated with the use of folinic acid (5-formyltetrahydrofolate) that is physiologically converted enzymatically to 5MTHF that can enter the CSF via the reduced folate carrier on the choroid epithelial cells at the choroid plexus [144]. Also, folinic acid is capable of displacement of the blocking autoantibodies to folate receptor alpha in addition to increased diffusion when the plasma level of folinic acid is increased [144]. Interestingly, Beversdorf et al. have recently proposed a precision medicine approach for heterogeneous conditions with autism spectrum disorders [145]. Indeed, Beversdorf Phase 2m proposal might consider the findings of EPVS on MRI as biomarkers for impaired (FOCM) such as elevations in Hcy, deficient nutrient intake of folate, autoantibodies for folate receptor alpha, oxidative stress/ROS due to autoxidation of Hcy, BBBact/dys, BBBdd, decreased GSH, and increased GSSG. Not only will these findings serve as biomarkers for ASD but also will serve to follow the response to various future treatment paradigms [145].
While there are multiple injurious species (neurotoxins) to the BECs of the NVU as in Figure 5, this narrative overview has focused primarily on LPS-induced neuroinflammation, hyperglycemia of diabetes with elevated glucose and the AGE/RAGE interaction to increase ROS, Hcy/HHCY due to impaired FOCM, pC/C due to metainflammation as a result of visceral obesity and gut dysbiosis, and ROS due to multiple causes that are all capable of inducing mesmerizing images. Regardless of the multiple neurotoxins, there seems to be an overarching mechanism of BECact/dys resulting in inflammation, oxidative stress, and decreased nitric oxide that also associates with BBBdd to increase permeability and leaky brains resulting in EPVS. Further clarification of the prognostic values in the development and progression of EPVS and SVD in neurodegenerative diseases such as LOAD in aging and neurodevelopmental and neuropsychiatric disorders such as ASD in youth need to be studied in greater depth in animal models and human individuals. Importantly, the astroglia cells (inclusive of pvACef and psACef) are playing an important and crucial role in the development of EPVS.
Funding
This manuscript received no external funding; however, the original research was funded by the VA Puget Sound Healthcare System as listed in chapter 1.
Institutional Review Board Statement
The tissues provided for the representative electron microscopic images utilized in this manuscript were all approved in advance by the IRB of the VA Puget Sound Healthcare System (protocol 0909). All animal studies were performed under the protocols approved by the VA animal care and use committee in accordance with the IACUC guidelines. These studies were conducted in an AAALAC-accredited facility. Additionally, all animals were cared for in accordance with the National Institutes of Health guidelines and the guidelines from the Institutional Animal Care and Use Committees, and this study conformed to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data and materials will be provided upon reasonable request.
Acknowledgments
The author would like to acknowledge Tatyana Shulyatnikova for the contribution of artistic illustrations and editing. The author would also like to acknowledge DeAna Grant Research Specialist of the Electron Microscopy Core Facility at the Roy Blunt NextGen Precision Health Research Center, University of Missouri, Columbia, Missouri. The author also acknowledges the kind support of the William A Banks Lab including Michele A Erickson who provided the control male CD-1 and LPS treated mouse brain tissues for these original studies from the VA Puget Sound Healthcare System, Seattle, Washington.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
| AC | astrocyte; ACef, astrocyte endfeet; APC(s), antigen presenting cell(s); AQP4, aquaporin-4 water channel; ASD, autism spectrum disorder(s); BBB, blood–brain barrier; BBBact/dys, BBB activation/dysfunction; BBBdd, BBB dysfunction/disruption; BEC(s), brain endothelial cell(s); becGCx, brain endothelial cell glycocalyx; BM, basement membrane; BTBR ob/ob mice, obese hyperphagic, diabetic, brown and tan, brachyuric mice with leptin-deficiency mutation ; CAA, cerebral amyloid angiopathy; CADASIL, cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy; CC 4.0. creative commons 4.0 permission to republish with appropriate references; CL, capillary lumen; CSF, cerebrospinal fluid; db/db mice, hyperphagic obese diabetic mouse models homozygous for the diabetes spontaneous mutation of the leptin receptor (Leprdb); EPVS, enlarged perivascular spaces; FOCM, folate one-carbon metabolism; GHS, glutathione; GS, glymphatic system; Hcy, homocysteine, HHCY, hyperhomocysteinemia; iMGC, interrogating microglia cell; IPAD, intramural peri-arterial drainage; ISF, interstitial fluid; ISS, interstitial space(s); LOAD, late-onset Alzheimer’s disease; LPS, lipopolysaccharide; MetS, metabolic syndrome; MRI, magnetic resonance image-imaging; MTHFR, methylenetetrahydrofolate reductase gene; MVEPVS, MRI visible EPVS; NVU, neurovascular unit; Pc, pericyte; pCC, peripheral cytokines/chemokines; Pcef, pericyte endfeet; Pcfp, pericyte foot process; pvACef, perivascular astrocyte endfeet; psACef, protoplasmic perisynaptic astrocyte endfeet; PVS, perivascular spaces; PVS/EPVS, perivascular space/enlarged perivascular space; rPVMΦ, resident perivascular macrophages; ROS, reactive oxygen species; RTIWH, response to injury wound healing; SAS, subarachnoid space; -SH, thiol group(s); SVD, cerebral small vessel disease; TEM, transmission electron microscopy; TIA(s), transient ischemic attack(s); T2DM, type 2 diabetes mellitus; VaD, vascular dementia; VCID, vascular contributions to cognitive impairment and dementia; WMH, white matter hyperintensities. |
References
- Malatesta, M. Transmission Electron Microscopy as a Powerful Tool to Investigate the Interaction of Nanoparticles with Subcellular Structures. Int. J. Mol. Sci. 2021, 22, 12789. [Google Scholar] [CrossRef]
- Erickson, M.A.; Shulyatnikova, T.; Banks, W.A.; Hayden, M.R. Ultrastructural Remodeling of the Blood-Brain Barrier and Neurovascular Unit by Lipopolysaccharide-Induced Neuroinflammation. Int. J. Mol. Sci. 2023, 24, 1640. [Google Scholar] [CrossRef] [PubMed]
- Shulyatnikova, T.; Hayden, M.R. Why Are Perivascular Spaces Important? Medicina (Kaunas) 2023, 59, 917. [Google Scholar] [CrossRef]
- McConnell, H.L.; Kersch, C.N.; Randall, L.; Woltjer, R.L.; Neuwelt, E.A. The Translational Significance of the Neurovascular Unit. J. Biol. Chem. 2017, 292, 762–770. [Google Scholar] [CrossRef]
- McConnell, H.L.; Mishra, A. Cells of the Blood-Brain Barrier: An Overview of the Neurovascular Unit in Health and Disease. Methods Mol. Biol. 2022, 2492, 3–24. [Google Scholar] [CrossRef]
- Iadecola, C. Neurovascular regulation in the normal brain and in Alzheimer’s disease. Nat. Rev. Neurosci. 2004, 5, 347–360. [Google Scholar] [CrossRef]
- Zlokovic, B.J. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat. Rev. Neurosci. 2011, 12, 723–738. [Google Scholar] [CrossRef]
- Troili, F.; Cipollini, V.; Moci, M.; Morena, E.; Palotai, M.; Rinaldi, V.; et.al., *!!! REPLACE !!!*. Perivascular Unit: This Must Be the Place. The Anatomical Crossroad between the Immune, Vascular and Nervous System. Front. Neuroanat. 2020, 14, 17. [Google Scholar] [CrossRef]
- Hayden, M.R. A Closer Look at the Perivascular Unit in the Development of Enlarged Perivascular Spaces in Obesity, Metabolic Syndrome, and Type 2 Diabetes Mellitus. Biomedicines. 2024, 12, 96. [Google Scholar] [CrossRef]
- Owens, T.; Bechmann, I.; Engelhardt, B. Perivascular spaces and the two steps to neuroinflammation. J. Neuropathol. Exp. Neurol. 2008, 67, 1113–1121. [Google Scholar] [CrossRef]
- Wen, W.; Cheng, J.; Tan, Y. Brain perivascular macrophages: Current understanding and future prospects. Brain. 2024, 147, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.; Wang, S.; Bèchet, N.; Gouras, G.; Wen, G. Perivascular macrophages in the central nervous system: Insights into their roles in health and disease. Cell Death Dis. 2025, 16, 350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.T.; Inman, B.E.; Weller, R.O. Interrelationships of the pia mater and the perivascular (Virchow-Robin) spaces in the human cerebrum. J. Anat. 1990, 170, 111–123. [Google Scholar] [PubMed]
- Yu, L.; He, X.; Li, H.; Zhao, Y. Perivascular Spaces, Glymphatic System and MR. Front. Neurol. 2022, 13, 84493. [Google Scholar] [CrossRef]
- Brown, R.; Benveniste, H.; Black, S.E.; Charpak, S.; Dichgans, M.; Joutel, A.; et al. Understanding the role of the perivascular space in cerebral small vessel disease. Cardiovasc. Res. 2018, 114, 1462–1473. [Google Scholar] [CrossRef]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef]
- Iliff, J.J.; Lee, H.; Yu, M.; Feng, T.; Logan, J.; Nedergaard, M.; Benveniste, H. Brain-wide pathway for waste clearance captured by contrast-enhanced MRI. J. Clin. Invest. 2013, 123, 1299–1309. [Google Scholar] [CrossRef]
- Bohr, T.; Hjorth, P.G.; Holst, S.C.; Hrabětová, S.; Kiviniemi, V.; Lilius, T.; et al. The glymphatic system: Current understanding and modeling. iScience 2022, 25, 104987. [Google Scholar] [CrossRef]
- Hladky, S.B.; Barrand, M.A. The glymphatic hypothesis: The theory and the evidence. Fluids Barriers CNS. 2022, 19, 9. [Google Scholar] [CrossRef]
- Ding, Z.; Fan, X.; Zhang, Y.; Yao, M.; Wang, G.; Dong, Y.; Liu, J.; Song, W. The glymphatic system: A new perspective on brain diseases. Front. Aging Neurosci. 2023, 15, 1179988. [Google Scholar] [CrossRef]
- Wardlaw JM, Smith EE, Biessels GJ, Cordonnier C, Fazekas F, Frayne, R. ; et al. Neuroimaging standards for research into small vessel disease and its contribution to ageing and neurodegeneration. Lancet Neurol. 2023, 22, 602–618. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, M.; Luan, M.; Song, X.; Wang, Y.; Xu, L.; Zhong, M.; Zheng, X. Enlarged Perivascular Spaces and Age-Related Clinical Diseases. Clin. Interv. Aging. 2023, 18, 855–867. [Google Scholar] [CrossRef]
- Heier, L.A.; Bauer, C.J.; Schwartz, L.; Zimmerman, R.D.; Morgello, S.; Deck, M.D. Large Virchow-Robin spaces: MR-clinical correlation. Am. J. Neuroradiol. 1989, 10, 929–936. [Google Scholar]
- Doubal, F.N.; Maclullich, A.M.J.; Ferguson, K.J.; Dennis, M.S.; Wardlaw, J.M. Enlarged Perivascular Spaces on MRI Are a Feature of Cerebral Small Vessel Disease. Stroke. 2010, 41, 450–454. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Kisler, K.; Montagne, A.; Toga, A.W.; Zlokovic, B.V. The role of brain vasculature in neurodegenerative disorders. Nat. Neurosci. 2018, 21, 1318–1331. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, P.; Trippier, S.; Lawrence, A.J.; Lambert, C.; Zeestraten, E.; Williams, O.A.; et al. Lacunar Infarcts, but Not Perivascular Spaces, Are Predictors of Cognitive Decline in Cerebral Small-Vessel Disease. Stroke 2018, 49, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Arba, F.; Quinn, T.J.; Hankey, G.J.; Lees, K.R.; Wardlaw, J.M.; Ali, M.; Inzitari, D. , on behalf of the VISTA Collaboration. Enlarged perivascular spaces and cognitive impairment after stroke and transient ischemic attack. Int. J. Stroke. 2018, 13, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, J.M.; Smith, C.; Dichgans, M. Mechanisms of sporadic cerebral small vessel disease: Insights from neuroimaging. Lancet Neurol. 2013, 12, 483–497. [Google Scholar] [CrossRef]
- Bown, C.W.; Carare, R.O.; Schrag, M.S.; Jefferson, A.L. Physiology and Clinical Relevance of Enlarged Perivascular Spaces. Neurology 2022, 98, 107–117. [Google Scholar] [CrossRef]
- Hayden, M.R.; Tyagi, N. The Integrity of Perivascular Spaces Is Absolutely Essential for Proper Function of the Glymphatic System Waste and Excess Water Removal from the Brain. Histol Histopathol. 2025, 18952. [Google Scholar] [CrossRef]
- Hayden, M.R. Brain Endothelial Cells Play a Central Role in the Development of Enlarged Perivascular Spaces in the Metabolic Syndrome. Medicina (Kaunas). 2023, 59, 1124. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.A.; Banks, W.A. Transcellular routes of blood-brain barrier disruption. Exp. Biol. Med. 2022, 247, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. From blood-brain barrier to blood-brain interface: New opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 2916, 15, 275–292. [Google Scholar] [CrossRef]
- Hayden, M.R. Brain Injury: Response to Injury Wound-Healing Mechanisms and Enlarged Perivascular Spaces in Obesity, Metabolic Syndrome, and Type 2 Diabetes Mellitus. Medicina (Kaunas). 2023, 59, 1337. [Google Scholar] [CrossRef]
- Banks, W.A.; Gray, A.M.; Erickson, M.A. Salameh TS, Damodarasamy M; et al. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflammation. 2015, 12, 223. [Google Scholar] [CrossRef]
- Erickson, M.A.; Banks, W.A. Transcellular routes of blood–brain barrier disruption. Exp. Biol. Med. 2022, 247, 788–796. [Google Scholar] [CrossRef]
- Erickson, M.A.; Liang, W.S.; Fernandez, E.G.; Bullock, K.M.; Thysell, J.A.; Banks, W.A. Genetics and sex influence peripheral and central innate immune responses and blood-brain barrier integrity. PLoS ONE 2018, 13, e0205769. [Google Scholar] [CrossRef]
- Zheng, L.; Guo, Y.; Zhai, X.; Zhang, Y.; Chen, W.; Zhu, Z.; Xuan, W.; Li, P. Perivascular macrophages in the CNS: From health to neurovascular diseases. CNS Neurosci. Ther. 2022, 28, 1908–1920. [Google Scholar] [CrossRef]
- Hayden, M.R. The Brain Endothelial Cell Glycocalyx Plays a Crucial Role in the Development of Enlarged Perivascular Spaces in Obesity, Metabolic Syndrome, and Type 2 Diabetes Mellitus. Life 2023, 13, 1955. [Google Scholar] [CrossRef]
- Kutuzov, N.; Flyvbjerg, H.; Lauritzen, M. Contributions of the glycocalyx, endothelium, and extravascular compartment to the blood–brain barrier. Proc. Natl. Acad. Sci. USA 2018, 115, E9429–E9438. [Google Scholar] [CrossRef]
- Sampei, S.; Okada, H.; Tomita, H.; Takada, C.; Suzuki, K.; Kinoshita, T.; et al. Endothelial Glycocalyx Disorders May Be Associated With Extended Inflammation During Endotoxemia in a Diabetic Mouse Model. Front. Cell Dev. Biol. 2021, 9, 623582. [Google Scholar] [CrossRef] [PubMed]
- Attwell, D.; Mishra, A.; Hall, C.N.; O’Farrell, F.M.; Dalkara, T. What is a pericyte? J. Cereb. Blood Flow. Metab. 2016, 36, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Dalkara, T.; Gursoy-Ozdemir, Y.; Yemisci, M. Brain microvascular pericytes in health and disease. Acta Neuropathol. 2011, 122, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.D.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the neurovascular unit: Key functions and signaling pathways. Nat. Neurosci. 2016, 19, 771–783. [Google Scholar] [CrossRef]
- Winkler, E.A.; Bell, R.D.; Zlokovic, B.V. Central nervous system pericytes in health and disease. Nat. Neurosci. 2011, 14, 1398–1405. [Google Scholar] [CrossRef]
- Dalkara, T.; Østergaard, L.; Heusch, G.; Attwell, D. Pericytes in the brain and heart: Functional roles and response to ischaemia and reperfusion. Cardiovasc. Res. 2025, 120, 2336–2348. [Google Scholar] [CrossRef]
- Jansson, D.; Rustenhoven, J.; Feng, S.; Hurley, D.; ROldfield, R.L.; Bergin, P.S.; et al. +-. J. Neuroinflammation. 2014, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Rustenhoven, J.; Jansson, D.; Smyth, L.C.; Dragunow, M. Brain Pericytes As Mediators of Neuroinflammation. Trends Pharmacol. Sci. 2017, 38, 291–304. [Google Scholar] [CrossRef]
- Iadecola, C.; Nedergaard, M. Glial regulation of the cerebral microvasculature. Nat. Neurosci. 2007, 10, 1369–1376. [Google Scholar] [CrossRef]
- Hall, C.N.; Reynell, C.; Gesslein, B.; Hamilton, N.B.; Mishra, A.; Sutherland, B.A.; et al. Capillary pericytes regulate cerebral blood flow in health and disease. Nature 2014, 508, 55–60. [Google Scholar] [CrossRef]
- Hayden, M.R.; Yang, Y.; Habibi, J.; Bagree, S.V.; Sowers, J.R. Pericytopathy: Oxidative stress and impaired cellular longevity in the pancreas and skeletal muscle in metabolic syndrome and type 2 diabetes. Oxid. Med. Cell Longev. 2010, 3, 290–303. [Google Scholar] [CrossRef]
- Winkler, E.A.; Sagare, A.P.; Zlokovic, B.V. The Pericyte: A Forgotten Cell Type with Important Implications for Alzheimer’s Disease? Brain Pathol. 2014, 24, 371–386. [Google Scholar] [CrossRef]
- Dore-Duffy, P.; Andre Katychev, A.; Xueqian Wang, X.; Van Buren, E. CNS microvascular pericytes exhibit multipotential stem cell activity. J. Cereb. Blood Flow. 2006, 26, 613–624. [Google Scholar] [CrossRef]
- Hayden, M.R. Protoplasmic Perivascular Astrocytes Play a Crucial Role in the Development of Enlarged Perivascular Spaces in Obesity, Metabolic Syndrome, and Type 2 Diabetes Mellitus. Neuroglia. 2023, 4, 307–328. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Butt, A.M. Neuroglia: Function and Pathology, 1st ed.; Academic Press: London, UK, 2023. [Google Scholar]
- Verkhratsky, A.; Nedergaard, M.; Hertz, L. Why are astrocytes important? . Neurochem. Res. 2015, 40, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Parpura, V.; Li, B.; Scuderi, C. Astrocytes: The Housekeepers and Guardians of the CNS. Adv. Neurobiol. 2021, 26, 21–53. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Nedergaard, M. Astroglial cradle in the life of the synapse. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130595. [Google Scholar] [CrossRef]
- Szu, J.I.; Binder, D.K. The Role of Astrocytic Aquaporin-4 in Synaptic Plasticity and Learning and Memory. Front. Integr. Neurosci. 2016, 10, 8. [Google Scholar] [CrossRef]
- Verkhratsky, A.; Bush, N.A.O.; Nedergarrd, M.; Butt, A.M. The Special Case of Human Astrocytes. Neuroglia 2018, 1, 21–29. [Google Scholar] [CrossRef]
- Li, L.; Cui, X.; Zhu, B.; Zhou, L.; Gao, Y.; Lin, T.; Shi, Y. Role of astrocytes in the pathogenesis of perinatal brain injury. Mol. Med. 2025, 31, 277. [Google Scholar] [CrossRef]
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.; Qu, Z.; Hashimoto, H.; et al. Molecular basis of astrocyte diversity and morphology across the CNS in health and disease. Science 2022, 378, eadc9020. [Google Scholar] [CrossRef]
- Engelhardt, B.; Carare, R.O.; Bechmann, I.; Flügel, A.; Laman, J.D.; Weller, R.O. Vascular, glial, and lymphatic immune gateways of the central nervous system. Acta Neuropathol. 2016, 132, 317–338. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.K.; Wang, F.; Wang, W.; Luo, Y.; Wu, P.F.; Jun-Li Xiao, J.L.; et al. Aquaporin-4 deficiency impairs synaptic plasticity and associative fear memory in the lateral amygdala: Involvement of downregulation of glutamate transporter-1 expression. Neuropsychopharmacology 2012, 37, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Nagelhus, E.A.; Ottersen, O.P. Physiological roles of aquaporin-4 in brain. Physiol. Rev. 2013, 93, 1543–1562. [Google Scholar] [CrossRef] [PubMed]
- Mazaré, N.; Oudart, M.; Cohen-Salmon, M.; Cohen-Salmon, M. Local translation in perisynaptic and perivascular astrocytic processes - a means to ensure astrocyte molecular and functional polarity? J Cell Sci. 2021, 134, jcs251629. [Google Scholar] [CrossRef]
- Muñoz de León-López, C.A.; Navarro-Lobato, I.; Khan, Z.U. The Role of Astrocytes in Synaptic Dysfunction and Memory Deficits in Alzheimer’s Disease. Biomolecules. 2025, 15, 910. [Google Scholar] [CrossRef]
- Pozo, K.; Goda, Y. Unraveling mechanisms of homeostatic synaptic plasticity. Neuron. 2010, 66, 337–351. [Google Scholar] [CrossRef]
- Vitureira, N.; Letellier, M.; Goda, Y. Homeostatic synaptic plasticity: From single synapses to neural circuits. Curr. Opin. Neurobiol. 2012, 22, 516–521. [Google Scholar] [CrossRef]
- Hayden, M.R. Pericytes and Resident Perivascular Macrophages Play a Key Role in the Development of Enlarged Perivascular Spaces in Obesity, Metabolic Syndrome and Type 2 Diabetes Mellitus. J. Alzheimers Neurodegener. Dis. 2023, 9, 062. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Guo, Y.; Zhai, X.; Zhang, Y.; Chen, W.; Zhu, Z.; Xuan, W.; Li, P. CNS Perivascular macrophages in the CNS: From health to neurovascular diseases. Neurosci. Ther. 2022, 28, 1908–1920. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Guo, R.; Zhang, F. Brain perivascular macrophages: Recent advances and implications in health and diseases. CNS Neurosci. 2019, 25, 1318–1328. [Google Scholar] [CrossRef]
- Ide, S.; Yahara, Y.; Kobayashi, Y.; Strausser, S.A.; Ide, K.; Watwe, A.; et al. Yolk-sac-derived macrophages progressively expand in the mouse kidney with age. eLife 2020, 9, e51756. [Google Scholar] [CrossRef]
- Zhan, X.; Wang, S.; Bèchet, N.; Gouras, G.; Wen, G. Perivascular macrophages in the central nervous system: Insights into their roles in health and disease. Cell Death Dis. 2025, 16, 350. [Google Scholar] [CrossRef]
- Wu, X.; Saito, T.; Saido, T.C.; Barron, A.M.; Ruedl, C. Microglia and CD206(+) border-associated mouse macrophages maintain their embryonic origin during Alzheimer’s disease. Elife 2021, 10, e71879. [Google Scholar] [CrossRef]
- Frosch, M.; -Amann, L.; Prinz, M. CNS-associated macrophages shape the inflammatory response in a mouse model of Parkinson’s disease. Nat. Commun. 2023, 14, 3753. [Google Scholar] [CrossRef]
- Sayd, A.; Vargas-Caraveo, A.; Perea-Romero, I.; Robledo-Montana, J.; Caso, J.R.; Madrigal, J.L.M.; et al. Depletion of brain perivascular macrophages regulates acute restraint stress-induced neuroinflammation and oxidative/nitrosative stress in rat frontal cortex. Eur. Neuropsychopharmacol. 2020, 34, 50–64. [Google Scholar] [CrossRef]
- Mendes, N.F.; Velloso, L.A. Perivascular macrophages in high-fat diet-induced hypothalamic inflammation. J. Neuroinflamm. 2022, 19, 136. [Google Scholar] [CrossRef]
- Pedragosa, J.; Salas-Perdomo, A.; Gallizioli, M.; Cugota, R.; Miro-Mur, F.; Brianso, F.; et al. CNS-border associated macrophages respond to acute ischemic stroke attracting granulocytes and promoting vascular leakage. Acta Neuropathol. Commun. 2018, 6, 76. [Google Scholar] [CrossRef]
- Faraco, G.; Sugiyama, Y.; Lane, D.; Garcia-Bonilla, L.; Chang, H.; Santisteban, M.M.; et al. Perivascular macrophages mediate the neurovascular and cognitive dysfunction associated with hypertension. J. Clin. Invest. 2016, 126, 4674–4689. [Google Scholar] [CrossRef]
- Sun, R.; Jiang, H. Border-associated macrophages in the central nervous system. J. Neuroinflammation. 2024, 21, 67. [Google Scholar] [CrossRef]
- Vara-Pérez, M.; Movahedi, K. Border-associated macrophages as gatekeepers of brain homeostasis and immunity. Immunity. 2025, 58, 1085–1100. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H. Space and Place: A Daoist Perspective. In The Philosophy of Geography. Springer Geography; Tambassi, T., Tanca, M., Eds.; Springer: Cham, 2021; Volume 6, pp. 95–115. [Google Scholar] [CrossRef]
- Mestre, H.; Kostrikov, S.; Mehta, R.I.; Nedergaard, M. Perivascular spaces, glymphatic dysfunction, and small vessel disease. Clin Sci (Lond). 2017, 131, 2257–2274. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. The glymphatic pathway in neurological disorders. Lancet Neurol. 2018, 17, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. Fluid transport in the brain. Physiol. Rev. 2022, 102, 1025–1151. [Google Scholar] [CrossRef]
- Nedergaard, M.; Goldman, S.A. Glymphatic failure as a final common pathway to dementia. Science 2020, 370, 50–56. [Google Scholar] [CrossRef]
- Lei, Y.; Han, H.; Yuan, F.; Javeed, A.; Zhao, Y. The brain interstitial system: Anatomy, modeling, in vivo measurement, and applications. ProgNeurobiol. 2017, 157, 230–246. [Google Scholar] [CrossRef]
- Hayden, M.R.; Grant, D.G.; Aroor, A.R.; DeMarco, V.G. Empagliflozin Ameliorates Type 2 Diabetes-Induced Ultrastructural Remodeling of the Neurovascular Unit and Neuroglia in the Female db/db Mouse. Brain Sci. 2019, 9, 57. [Google Scholar] [CrossRef]
- Banks, W.A.; Hayden, M.R. Deficient Leptin Cellular Signaling Plays a Key Role in Brain Ultrastructural Remodeling in Obesity and Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 5427. [Google Scholar] [CrossRef]
- Munis, Ö.B. Association of Type 2 Diabetes Mellitus With Perivascular Spaces and Cerebral Amyloid Angiopathy in Alzheimer’s Disease: Insights From MRI Imaging. Dement. Neurocogn Disord. 2023, 22, 87–99. [Google Scholar] [CrossRef]
- Hansen, T.P.; Cain, J.; Thomas, O.; Jackson, A. Dilated perivascular spaces in the Basal Ganglia are a biomarker of small-vessel disease in a very elderly population with dementia. AJNR Am. J. Neuroradiol. 2015, 36, 893–898. [Google Scholar] [CrossRef]
- Zebarth, J.; Kamal, R.; Perlman, G.; Ouk, M.; Xiong, L.Y.; Yu, D.; Lin, W.Z.; Ramirez, J.; et al. Perivascular spaces mediate a relationship between diabetes and other cerebral small vessel disease markers in cerebrovascular and neurodegenerative diseases. J. Stroke Cerebrovasc. Dis. 2023, 32, 107273. [Google Scholar] [CrossRef]
- de la Torre, J.C. Cardiovascular risk factors promote brain hypoperfusion leading to cognitive decline and dementia. Cardiovasc. Psychiatry Neurol. 2012, 2012, 367516. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Zhang, D.; Li, J.; Tang, M.; Yan, X.; Zhang, X.; et al. The enlarged perivascular spaces in the hippocampus is associated with memory function in patients with type 2 diabetes mellitus. Sci. Rep. 2025, 15, 3644. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.R.; Tyagi, S.C. Homocysteine and reactive oxygen species in metabolic syndrome, type 2 diabetes mellitus, and atheroscleropathy: The pleiotropic effects of folate supplementation. Nutr. J. 2004, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.R.; Tyagi, S.C. Impaired Folate-Mediated One-Carbon Metabolism in Type 2 Diabetes, Late-Onset Alzheimer’s Disease and Long COVID. Medicina (Kaunas). 2021, 58, 16. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2010, 21, 103–115. [Google Scholar] [CrossRef]
- Kaur, U.; Banerjee, P.; Bir, A.; Sinha, M.; Biswas, A.; Chakrabarti, S. Reactive oxygen species, redox signaling and neuroinflammation in Alzheimer’s disease: The NF-κB connection. Curr. Top. Med. Chem. 2015, 15, 446–457. [Google Scholar] [CrossRef]
- Cordaro, M.; Siracusa, R.; Fusco, R.; Cuzzocrea, S.; Paola, R.D.; Impellizzeri, D. Involvements of Hyperhomocysteinemia in Neurological Disorders. Metabolites. 2021, 11, 37. [Google Scholar] [CrossRef]
- Elias, M.F.; Sullivan, L.M.; D’Agostino, R.B.; Elias, P.K.; Paul, F. Jacques PF, Selhub J; et al. Homocysteine and Cognitive Performance in the Framingham Offspring Study: Age Is Important. Stroke 2004, 35, 404–409. [Google Scholar] [CrossRef] [PubMed]
- McCully, K.S. Vascular pathology of homocysteinemia: Implications for the pathogenesis of arteriosclerosis. Am. J. Pathol. 1969, 56, 111–128. [Google Scholar] [PubMed]
- Stampfer, M.J.; Malinow, M.R.; Willett, W.C.; Newcomer, L.; Upson, B.; Ullmann, D.; Tishler, P. ; Hennekens CH: Aprospective study of plasma homocyst(e)ine risk of myocardial infarction in USphysicians. JAMA 1992, 268, 877–881. [Google Scholar] [CrossRef]
- Arnesen, E.; Refsum, H.; Bonaa, K.H.; Ueland, P.M.; Forde, O.H.; Nordrehaug, J.E. Serum total homocysteine and coronary heart disease. Int. J. Epidemiol. 1995, 24, 704–709. [Google Scholar] [CrossRef]
- Boushey, C.J.; Beresford, S.A.; Omenn, G.S. ; Motulsky AG: Aquantitative assessment of plasma homocysteine as a risk factor for vascular disease Probable benefits of increasing folic acid intakes. JAMA 1995, 274, 1049–1057. [Google Scholar] [CrossRef]
- Graham, I.M.; Daly, L.E.; Refsum, H.M.; Robinson, K.; Brattstrom, L.E. ; Ueland PM: Plasma homocysteine as a risk factor for vascular disease The European Concerted Action Project. JAMA 1997, 277, 1775–1781. [Google Scholar] [CrossRef]
- Wald, N.J.; Watt, H.C.; Law, M.R.; Weir, D.G.; McPartlin, J.; Scott, J.M. Homocysteine and ischemic heart disease: Results of a prospective study with implications regarding prevention. Arch. Intern. Med. 1998, 158, 862–867. [Google Scholar] [CrossRef]
- Hoogeveen, E.K.; Kostense, P.J.; Beks, P.J.; Mackaay, A.J.; Jakobs, C.; Bouter, L.M.; Heine, R.J.; Stehouwer, C.D. Hyperhomocysteinemia is associated with an increased risk of cardiovascular disease, especially in non-insulin-dependent diabetes mellitus: A population-based study. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 133–138. [Google Scholar] [CrossRef]
- Welch, G.N.; Loscalzo, J. Homocysteine and atherothrombosis. N. Engl. J. Med. 1998, 338, 1042–1050. [Google Scholar] [CrossRef]
- Stehouwer, C.D.; Weijenberg, M.P.; van den Berg, M.; Jakobs, C.; Feskens, E.J.; Kromhout, D. Serum homocysteine and risk of coronary heart disease and cerebrovascular disease in elderly men: A 10-year follow-up. Arterioscler Thromb Vasc Biol. 1998, 18, 1895–1901. [Google Scholar] [CrossRef]
- Boers, G.H. Mild hyperhomocysteinemia is an independent risk factor of arterial vascular disease. Semin. Thromb. Hemost. 2000, 26, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Pinzon, R.T.; Wijaya, V.O.; Veronica, V. The role of homocysteine levels as a risk factor of ischemic stroke events: A systematic review and meta-analysis. Neurol. 2023, 14, 1144584. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Refsum, H.; Bottiglieri, T.; Fenech, M.; Hooshmand, B.; McCaddon, A.; et al. Homocysteine and Dementia: An International Consensus Statement. J. Alzheimers Dis. 2018, 62, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, J.M.; Wang, H.; Pratico, D. Is hyperhomocysteinemia an Alzheimer’s disease (AD) risk factor, an AD marker, or neither? Trends Pharmacol. Sci. 2011, 32, 562–571. [Google Scholar] [CrossRef]
- Zhou, L. Homocysteine and Parkinson’s disease. CNS Neurosci. Ther. 2023, 30, e14420. [Google Scholar] [CrossRef]
- Moustafa, A.A.; Hewedi, D.H.; Eissa, A.M.; Frydecka, D.; Misiak, B. Homocysteine levels in schizophrenia and affective disorders—Focus on cognition. Front. Behav. Neurosci. 2014, 8, 343. [Google Scholar] [CrossRef]
- Baldelli, E.; Leo, G.; Andreoli, N.; Fuxe, K.; Biagini, G.; Luigi FAgnati, L.F. Homocysteine potentiates seizures and cell loss induced by pilocarpine treatment. Neuromolecular Med. 2010, 12, 248–259. [Google Scholar] [CrossRef]
- Li, B.; Xu, Y.; Pang, D.; Zhao, Q.; Zhang, L.; Li, M.; Li, W.; Duan, G.; Zhu, C. Interrelation between homocysteine metabolism and the development of autism spectrum disorder in children. Front. Mol. Neurosci. 2022, 15, 947513. [Google Scholar] [CrossRef]
- Garic, D.; McKinstry, R.C.; Rutsohn, J.; Slomowitz, R.; Wolff, J.; MacIntyre, L.C.; et al. Enlarged Perivascular Spaces in Infancy and Autism Diagnosis, Cerebrospinal Fluid Volume, and Later Sleep Problems. JAMA Netw. Open. 2023, 6, e2348341. [Google Scholar] [CrossRef]
- Gao, S.; Wang, L.; Liu, W.; Wu, Y.; Yuan, Z. The synergistic effect of homocysteine and lipopolysaccharide on the differentiation and conversion of raw264.7 macrophages. 7 macrophages. J Inflamm (Lond). 2014, 11, 13. [Google Scholar] [CrossRef]
- Jessen, N.A.; Munk, A.S.; Lundgaard, I.; Nedergaard, M. The Glymphatic System: A beginner’s guide. Neurochem. Res. 2015, 40, 2583–2599. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Thrippleton, M.J.; Blair, G.W.; Dickie, D.A.; Marshall, I.; Hamilton, I.; et al. Small vessel disease is associated with altered cerebrovascular pulsatility but not resting cerebral blood flow. J. Cereb. Blood Flow. Metab. 2020, 40, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A.M.; et al. Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. Nat. Commun. 2018, 9, 4878. [Google Scholar] [CrossRef] [PubMed]
- Dreha-Kulaczewski, S.; Joseph, A.A.; Merboldt, K.D.; Ludwig, H.C.; Gärtner, J.; Frahm, J. Identification of the upward movement of human CSF in vivo and its relation to the brain venous system. J. Neurosci. 2017, 37, 2395–2402. [Google Scholar] [CrossRef]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef]
- Hablitz, L.M.; Plá, V.; Giannetto, M.; Vinitsky, H.S.; Stæger, F.F.; Metcalfe, T.; et al. Circadian control of brain glymphatic and lymphatic fluid flow. Nat. Commun. 2020, 11, 4411. [Google Scholar] [CrossRef]
- Mazaré, N.; Oudart, M.; Moulard, J.; Cheung, G.; Tortuyaux, R.; Mailly, P.; et al. Local Translation in Perisynaptic Astrocytic Processes Is Specific and Changes after Fear Conditioning. Cell Rep. 2020, 32, 108076. [Google Scholar] [CrossRef]
- Colón-Ramos, D.A. Synapse formation in developing neural circuits. Curr. Top. Dev. Biol. 2009, 87, 53–79. [Google Scholar] [CrossRef]
- Südhof, T.C.; Malenka, R.C. Understanding Synapses: Past, Present, and Future. Neuron. 2008, 60, 469–476. [Google Scholar] [CrossRef]
- Ecker, C.; Suckling, J.; Deoni, S.C.; Lombardo, M.V.; Bullmore, E.T.; Baron-Cohen, S.; et al. Brain anatomy and its relationship to behavior in adults with autism spectrum disorder: A multicenter magnetic resonance imaging study. Arch. Gen. Psychiatry. 2012, 69, 195–209. [Google Scholar] [CrossRef]
- Quaak, I.; Brouns, M.R.; Van de Bor, M. The dynamics of autism spectrum disorders: How neurotoxic compounds and neurotransmitters interact. Int. J. Environ. Res. Public. Health. 2013, 10, 3384–3408. [Google Scholar] [CrossRef] [PubMed]
- Lauritsen, M.B. Autism spectrum disorders. Eur. Child. Adolesc. Psychiatry. 2013, 22, S37–S42. [Google Scholar] [CrossRef] [PubMed]
- Garic, D.; McKinstry, R.C.; Rutsohn, J.; Slomowitz, R.; Wolff, J.; MacIntyre, L.C.; et al. Enlarged Perivascular Spaces in Infancy and Autism Diagnosis, Cerebrospinal Fluid Volume, and Later Sleep Problems. JAMA Netw. Open. 2023, 6, e2348341. [Google Scholar] [CrossRef]
- Sotgiu, M.A.; Jacono, L.A.; Barisano, G.; Saderi, L.; Cavassa, V.; Montella, A.; Crivelli, P.; Carta, A.; Sotgiu, S. Brain perivascular spaces and autism: Clinical and pathogenic implications from an innovative volumetric MRI study. Front. Neurosci. 2023, 17, 1205489. [Google Scholar] [CrossRef]
- González-Castro, V.; Valdes-Hernandez, M.D.C.; Chappell, F.; Armitage, P.A.; Makin, S.; Wardlaw, J.M. Reliability of an automatic classifier for brain enlarged perivascular spaces burden and comparison with human performance. Clin. Sci. 2017, 131, 1465–1481. [Google Scholar] [CrossRef]
- Zeidan, J.; Fombonne, E.; Scorah, J.; Ibrahim, A.; Durkin, M.S.; Saxena, S.; et al. Global prevalence of autism: A systematic review update. Autism Res. 2022, 15, 778–790. [Google Scholar] [CrossRef]
- Frye, R.E.; Rossignol, D.A.; Scahill, L.; McDougle, C.J.; Huberman, H.; Quadros, E.V. Treatment of Folate Metabolism Abnormalities in Autism Spectrum Disorder. Semin. Pediatr. Neurol. 2020, 35, 100835. [Google Scholar] [CrossRef]
- Mattson, M.P.; Shea, T.B. Folate and homocysteine metabolism in neural plasticity and neurodegenerative disorders. Trends Neurosci. 2003, 26, 137–146. [Google Scholar] [CrossRef]
- Cohen Kadosh, K.; Muhardi, L.; Parikh, P.; Basso, M.; Jan Mohamed, H.J.; Prawitasari, T.; et al. Nutritional Support of Neurodevelopment and Cognitive Function in Infants and Young Children—An Update and Novel Insights. Nutrients 2021, 13, 199. [Google Scholar] [CrossRef]
- Korsmo, H.W.; Jiang, X. One Carbon Metabolism and Early Development: A Diet-Dependent Destiny. Trends Endocrinol. Metab. 2021, 32, 579–593. [Google Scholar] [CrossRef]
- Homocysteine Lowering Trialists’ Collaboration Lowering blood homocysteine with folic acid based supplements: Meta-analysis of randomised trials Homocysteine Lowering Trialists’ Collaboration. BMJ 1998, 316, 894–898. [CrossRef]
- Ramaekers, V.T.; Rothenberg, S.P.; JSequeira, J.M.; Opladen, T.; Blau, N.; Quadros, E.V.; Selhub, J. Autoantibodies to folate receptors in the cerebral folate deficiency syndrome. N. Engl. J. Med. 2005, 352, 1985–1991. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Anagnostou, E.; Hardan, A.; Wang, P.; Erickson, C.A.; Frazier, T.W.; Veenstra-VanderWeele, J. Editorial: Precision medicine approaches for heterogeneous conditions such as autism spectrum disorders (The need for a biomarker exploration phase in clinical trials - Phase 2m). Front. Psychiatry 2023, 13, 1079006. [Google Scholar] [CrossRef]
Figure 1.
Comparing normal capillary neurovascular Unit(s) (NVUs) without perivascular spaces (PVS) in control models to those with enlarged perivascular spaces (EPVS) identified in LPS-treated neuroinflammatory models. (A) demonstrates the normal appearance of the NVU in the control cortical C57BL6 male mice at 6-8-weeks of age and note how the perivascular astrocyte endfeet (ACef-pseudo-colored yellow) are tightly abutted to the brain endothelial cells (BECs) and pericyte endfeet (PcP-Pcef) basement membrane (black open arrows). Note the interrogating microglia cell (iMGC - white solid arrow lower left). (B) depicts the detachment and separation of the ACef (pseudo-colored yellow) from the mural cells (BEC and Pc) combined basement membranes to form a space that is known to be the PVS that is enlarged in this postcapillary venular image to result in an enlarged perivascular space(s) (EPVS) (red double arrows) in an LPS-treated CD-1 male mouse model at 6 to 8-weeks of age. Notably, the ACef basement membrane or glia limitans (membrana limitans gliae perivascularis) forms the outermost boundary of these PVS and EPVS. Note the red line marked through an aquaporin 4 water channel (AQP4) labeled astrocyte endfeet to indicate that these reactive astrocyte endfeet have lost their polarity and are associated with dysfunction and or loss of AQP4 water channel that may contribute to increased fluid/water within the PVS. Importantly, PVS form the conduits for the glymphatic system to provide clearance of metabolic waste, phagocytic debris, accumulated neurotoxins, and excess water accumulation in the brain. Note that the reactive perivascular macrophage (rMΦ/rPVMΦ) has an increase in lysosomes (Lys) and vacuoles and vesicles with a retracted non-elliptical nucleus suggesting a reactive morphologic state. Modified image provided with permission by CC 4.0 [2]. Scale bar = 0.5μm and 5μm (panels A and B respectively). ACef, astrocyte endfeet; AQP4, aquaporin 4 water channel; BEC, brain endothelial cell; CD-1, commonly utilized control mouse model; Dys, dysfunctional; CKC, control mouse model; EC, brain endothelial cell; EPVS, enlarged perivascular space(s) double arrows; iMGC, interrogating microglia cell; LPS, lipopolysaccharide; Lys, lysosome; N, nucleus; Neuropil, a mesh-like intertwined network of mostly unmyelinated axons within the grey matter of the brain; Pc, pericyte; Pcfp, pericyte foot process-endfeet; pm, plasma membrane; rMΦ, resident perivascular macrophage.
Figure 1.
Comparing normal capillary neurovascular Unit(s) (NVUs) without perivascular spaces (PVS) in control models to those with enlarged perivascular spaces (EPVS) identified in LPS-treated neuroinflammatory models. (A) demonstrates the normal appearance of the NVU in the control cortical C57BL6 male mice at 6-8-weeks of age and note how the perivascular astrocyte endfeet (ACef-pseudo-colored yellow) are tightly abutted to the brain endothelial cells (BECs) and pericyte endfeet (PcP-Pcef) basement membrane (black open arrows). Note the interrogating microglia cell (iMGC - white solid arrow lower left). (B) depicts the detachment and separation of the ACef (pseudo-colored yellow) from the mural cells (BEC and Pc) combined basement membranes to form a space that is known to be the PVS that is enlarged in this postcapillary venular image to result in an enlarged perivascular space(s) (EPVS) (red double arrows) in an LPS-treated CD-1 male mouse model at 6 to 8-weeks of age. Notably, the ACef basement membrane or glia limitans (membrana limitans gliae perivascularis) forms the outermost boundary of these PVS and EPVS. Note the red line marked through an aquaporin 4 water channel (AQP4) labeled astrocyte endfeet to indicate that these reactive astrocyte endfeet have lost their polarity and are associated with dysfunction and or loss of AQP4 water channel that may contribute to increased fluid/water within the PVS. Importantly, PVS form the conduits for the glymphatic system to provide clearance of metabolic waste, phagocytic debris, accumulated neurotoxins, and excess water accumulation in the brain. Note that the reactive perivascular macrophage (rMΦ/rPVMΦ) has an increase in lysosomes (Lys) and vacuoles and vesicles with a retracted non-elliptical nucleus suggesting a reactive morphologic state. Modified image provided with permission by CC 4.0 [2]. Scale bar = 0.5μm and 5μm (panels A and B respectively). ACef, astrocyte endfeet; AQP4, aquaporin 4 water channel; BEC, brain endothelial cell; CD-1, commonly utilized control mouse model; Dys, dysfunctional; CKC, control mouse model; EC, brain endothelial cell; EPVS, enlarged perivascular space(s) double arrows; iMGC, interrogating microglia cell; LPS, lipopolysaccharide; Lys, lysosome; N, nucleus; Neuropil, a mesh-like intertwined network of mostly unmyelinated axons within the grey matter of the brain; Pc, pericyte; Pcfp, pericyte foot process-endfeet; pm, plasma membrane; rMΦ, resident perivascular macrophage.
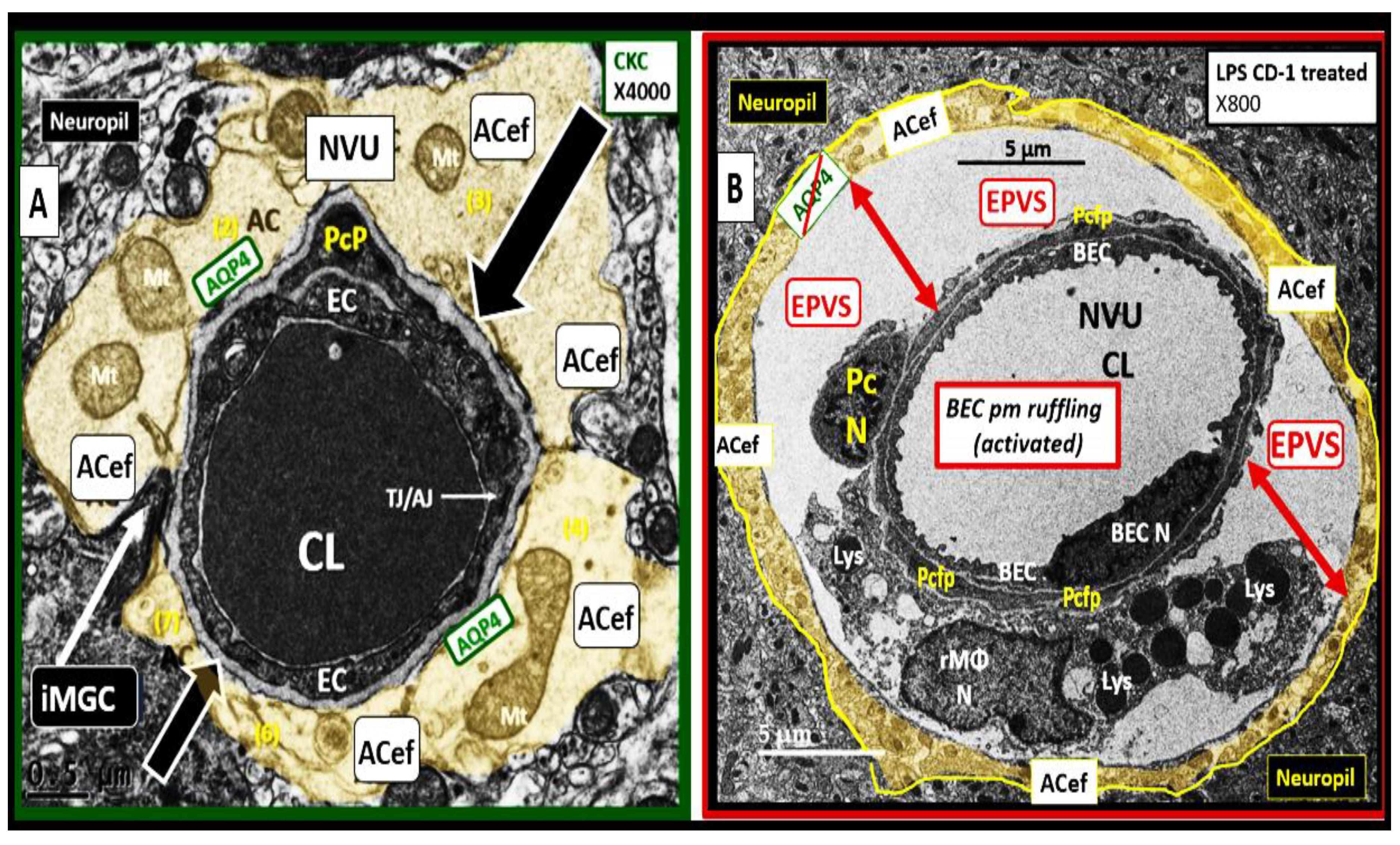
Figure 2.
Illustrations of precapillary arteriole influx, postcapillary venule efflux via perivascular spaces (PVS). (A) illustrates the perivascular spaces (PVS), which are bound by the arterial and venous endothelial/pericyte basement membranes (BMs) and the pia mater/astrocyte endfeet (ACef) (glia limitans) BMs that are responsible for the influx of cerebrospinal fluid (CSF) to the interstitial fluid (ISF) spaces. Likewise, the postcapillary venules, venules, and veins PVS are responsible for delivery of the ISF admixed with metabolic waste to the subarachnoid space (SAS) and eventually to the systemic circulation via arachnoid granulations, the meningeal lymphatics vessels along with the dural sinuses and lymphatics. The outermost pia mater abruptly stops at the true capillary and does not exist in the postcapillary venules and veins. (B) illustrates the important role of the arachnoid villus and its granulations for exchange of ISF/CSF and metabolic waste with the dural venous sinus blood and dura meningeal lymphatics–meningeal lymphatic vessels (MLVs) (cyan color) and-or the perineuronal spaces and paranasal lymphatics and sinuses (not shown) to reach the systemic peripheral circulation for disposal. (C) illustrates NVU and the perivascular astrocyte endfeet (pvACef with blue coloring) barrier with a few 20nm gaps (yellow circles) illustrates a rate-limiting barrier for water and solute exchange. Notably, the pvACef contain polarized aquaporin 4 (AQP4) water channels, which are known to be important in fluid and solute exchange in addition to the transfer of metabolic waste to the CSF. Note the red downward arrow representing the CSF influx as well as the retrograde flow of the IPAD system (upward purple arrow) and the blue upward arrow that depicts the efflux of the ISF that also serves as the space for the functional glymphatic system (GS) (blue upward dashed arrow). True capillary refers specifically to the NVU capillary where solutes, ions, and fluids are exchanged from the capillary lumen to the interstitial spaces of the brain. Also, note the key in within this image. Modified image provided with permission by CC 4.0 [3]. ACef, astrocyte endfeet; AG, arachnoid granulations; CL, capillary lumen; RBC, red blood cell.
Figure 2.
Illustrations of precapillary arteriole influx, postcapillary venule efflux via perivascular spaces (PVS). (A) illustrates the perivascular spaces (PVS), which are bound by the arterial and venous endothelial/pericyte basement membranes (BMs) and the pia mater/astrocyte endfeet (ACef) (glia limitans) BMs that are responsible for the influx of cerebrospinal fluid (CSF) to the interstitial fluid (ISF) spaces. Likewise, the postcapillary venules, venules, and veins PVS are responsible for delivery of the ISF admixed with metabolic waste to the subarachnoid space (SAS) and eventually to the systemic circulation via arachnoid granulations, the meningeal lymphatics vessels along with the dural sinuses and lymphatics. The outermost pia mater abruptly stops at the true capillary and does not exist in the postcapillary venules and veins. (B) illustrates the important role of the arachnoid villus and its granulations for exchange of ISF/CSF and metabolic waste with the dural venous sinus blood and dura meningeal lymphatics–meningeal lymphatic vessels (MLVs) (cyan color) and-or the perineuronal spaces and paranasal lymphatics and sinuses (not shown) to reach the systemic peripheral circulation for disposal. (C) illustrates NVU and the perivascular astrocyte endfeet (pvACef with blue coloring) barrier with a few 20nm gaps (yellow circles) illustrates a rate-limiting barrier for water and solute exchange. Notably, the pvACef contain polarized aquaporin 4 (AQP4) water channels, which are known to be important in fluid and solute exchange in addition to the transfer of metabolic waste to the CSF. Note the red downward arrow representing the CSF influx as well as the retrograde flow of the IPAD system (upward purple arrow) and the blue upward arrow that depicts the efflux of the ISF that also serves as the space for the functional glymphatic system (GS) (blue upward dashed arrow). True capillary refers specifically to the NVU capillary where solutes, ions, and fluids are exchanged from the capillary lumen to the interstitial spaces of the brain. Also, note the key in within this image. Modified image provided with permission by CC 4.0 [3]. ACef, astrocyte endfeet; AG, arachnoid granulations; CL, capillary lumen; RBC, red blood cell.
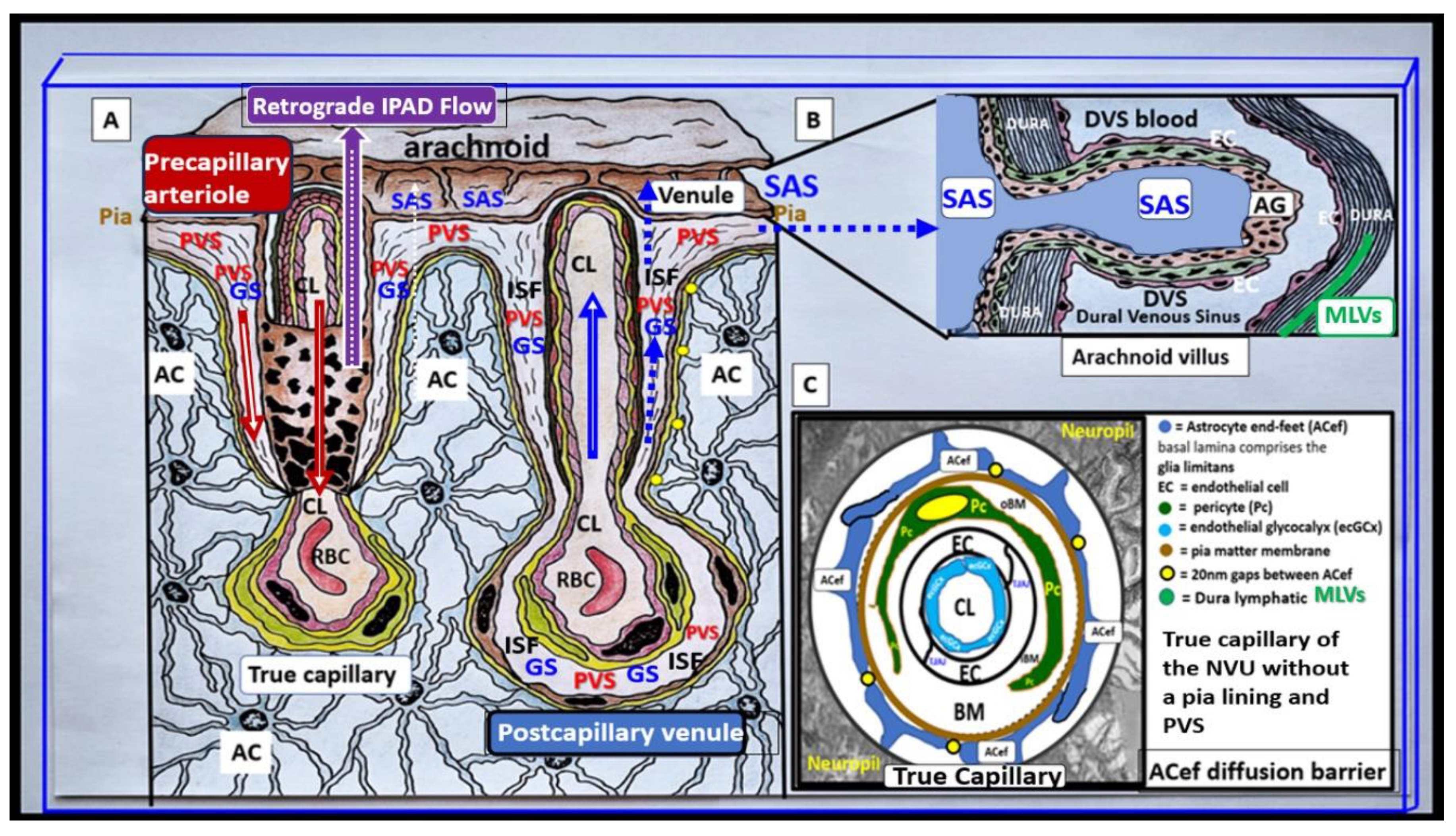
Figure 3.
Comparisons of enlarged perivascular spaces (EPVSs) in basal ganglia (BG) to centrum semiovale (CSO) by magnetic resonance imaging (MRI). (A) depicts the paired EPVSs within the BG (traced in open circles on the left and masked yellow circles on the right BG). Additionally, note the faint white spaces within the dashed lines just above the paired BG structures also depicting EPVS. This MRI image is from a 75 y/o male status post-stroke, recovered with small vessel disease. (B) depicts the paired elongated oval structures outlined by yellow dashed lines to enclose multiple white enlarged perivascular spaces with MRI lucency similar to the cerebral spinal fluid. Note the open white arrows outlined in red pointing to prominent EPVSs. MRI image from a 79 y/o female with history of transient ischemic attacks. Importantly, note that BG EPVS strongly associate with cerebral small vessel disease (SVD) in (A) and CSO EPVS strongly associate with late-onset Alzheimer’s disease and cerebral amyloid angiopathy (CAA) in (B). Modified image provided with permission by CC 4.0 [3,9].
Figure 3.
Comparisons of enlarged perivascular spaces (EPVSs) in basal ganglia (BG) to centrum semiovale (CSO) by magnetic resonance imaging (MRI). (A) depicts the paired EPVSs within the BG (traced in open circles on the left and masked yellow circles on the right BG). Additionally, note the faint white spaces within the dashed lines just above the paired BG structures also depicting EPVS. This MRI image is from a 75 y/o male status post-stroke, recovered with small vessel disease. (B) depicts the paired elongated oval structures outlined by yellow dashed lines to enclose multiple white enlarged perivascular spaces with MRI lucency similar to the cerebral spinal fluid. Note the open white arrows outlined in red pointing to prominent EPVSs. MRI image from a 79 y/o female with history of transient ischemic attacks. Importantly, note that BG EPVS strongly associate with cerebral small vessel disease (SVD) in (A) and CSO EPVS strongly associate with late-onset Alzheimer’s disease and cerebral amyloid angiopathy (CAA) in (B). Modified image provided with permission by CC 4.0 [3,9].
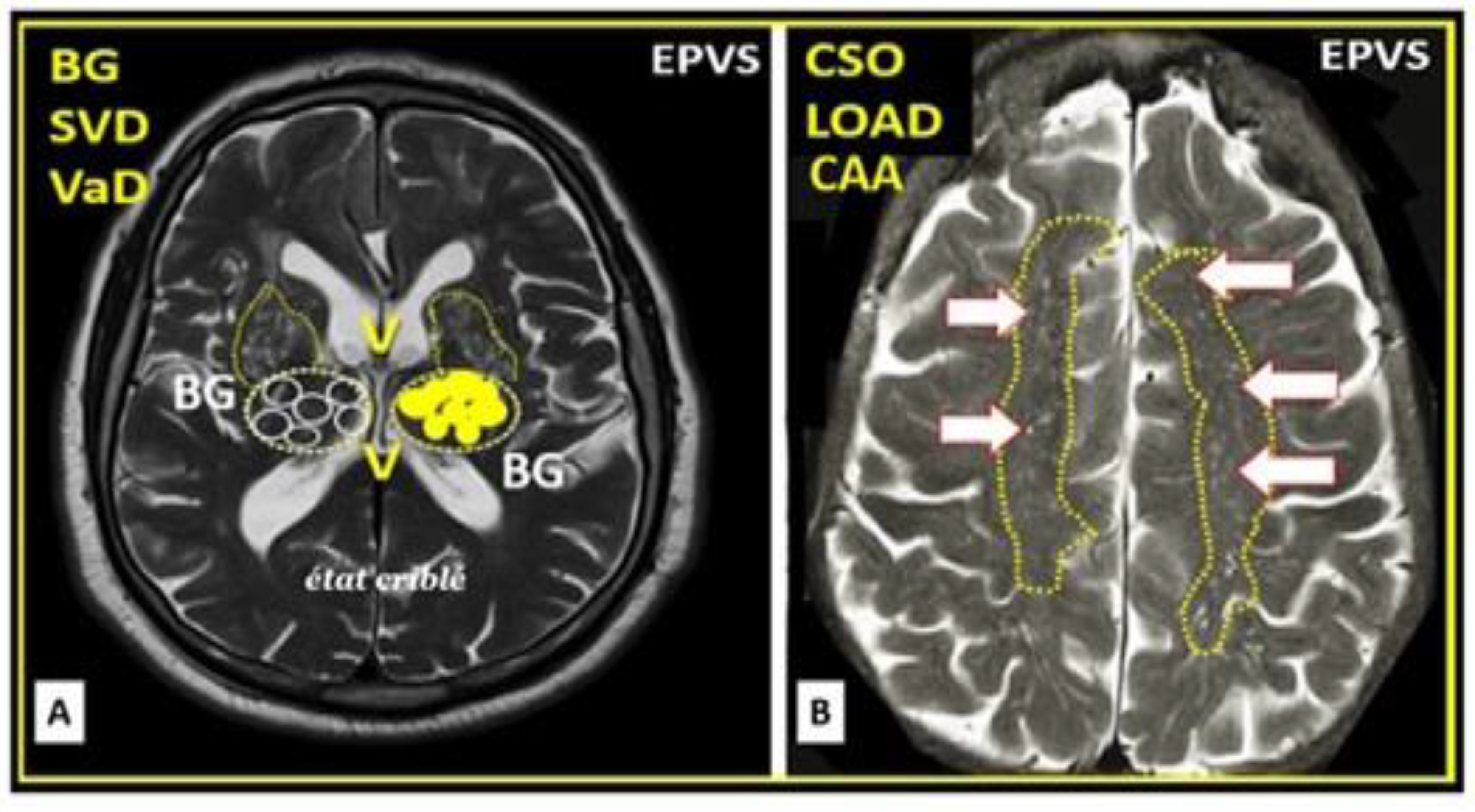
Figure 4.
Brain injury and the response to injury wound healing (RTIWH) mechanism. This image depicts the RTIWH mechanisms via dual signaling by peripheral cytokines/chemokines (pCC), soluble lipopolysaccharide (sLPS), and LPS-enriched extracellular exosomes (lpsEVexos). This dual signaling of the brain endothelial cell(s) (BECs) results in BEC activation and dysfunction (BECact/dys) and blood-brain barrier dysfunction/disruption (BBBdd), which in turn results in central nervous system (CNS) neuroinflammation with increased cnsCC, reactive polarized microglia, and reactive astroglia—reactive astrogliosis. This results in astrogliosis scarring instead of fibrosis scarring, since the brain does not have fibrocytes/fibroblasts in its parenchyma. Importantly, this RTIWH mechanism contributes to maladaptive cellular remodeling, development of enlarged perivascular spaces (EPVSs), neuroinflammation, neurodegeneration. Figure provided with permission by CC 4.0 [35].
Figure 4.
Brain injury and the response to injury wound healing (RTIWH) mechanism. This image depicts the RTIWH mechanisms via dual signaling by peripheral cytokines/chemokines (pCC), soluble lipopolysaccharide (sLPS), and LPS-enriched extracellular exosomes (lpsEVexos). This dual signaling of the brain endothelial cell(s) (BECs) results in BEC activation and dysfunction (BECact/dys) and blood-brain barrier dysfunction/disruption (BBBdd), which in turn results in central nervous system (CNS) neuroinflammation with increased cnsCC, reactive polarized microglia, and reactive astroglia—reactive astrogliosis. This results in astrogliosis scarring instead of fibrosis scarring, since the brain does not have fibrocytes/fibroblasts in its parenchyma. Importantly, this RTIWH mechanism contributes to maladaptive cellular remodeling, development of enlarged perivascular spaces (EPVSs), neuroinflammation, neurodegeneration. Figure provided with permission by CC 4.0 [35].

Figure 5.
Multiple injurious species affect brain endothelial cells (BECs) of the neurovascular unit (NVU). These injurious species contribute to BEC activation (proinflammatory) and dysfunction with decreased protective nitric oxide (NO) (BECact/dys) and blood–brain barrier dysfunction/disruption (BBBdd) with increased permeability via increased transcellular- transcytosis and paracellular leakiness. BECact/dys and BBBdd are bidirectional biomarkers for the development of enlarged perivascular spaces (EPVS) and cerebral small vessel disease (SVD). Note the red-dashed line at the top of this image, which designates the EC plasmalemma location of the multiple injurious species that are responsible for initial BEC injury in multiple clinical diseases and structural remodeling abnormalities including SVD and EPVS. BEC insulin resistance is associated with increased glucose, and the AGE receptor (RAGE) interaction is also important in the development of BECact/dys, BBBdd, and neurodegeneration. Also, note that angiotensin II (AngII) denotes the activation of the renin-angiotensin-aldosterone system (RAAS) including the endothelial cell mineralocorticoid receptor (ECMR) for aldosterone and that both play a role in the development of vascular arterial stiffening. Importantly, note that this image does not illustrate the endothelial glycocalyx. Modified image provided with permission by CC 4.0 [35RTIWH]. AGE, advanced glycation end products; aMMPs, activated matrix metalloproteinases; Ang II, angiotensin two; BBB, blood–brain barrier; BEC, brain endothelial cell; BBBdd, blood–brain barrier dysfunction and disruption; BECact/dys, brain endothelial cell activation/dysfunction; BH4, tetrahydrobiopterin; CCL2, chemokine (C-C motif) ligand 2; Cox-2, cyclo-oxygenase-2; Cox-2/PGE2 axis, cyclo-oxygenase-2/prostaglandin E2; downward arrows, arrowheads, denote call outs; ecGCx, endothelial glycocalyx; ICAM-1, intercellular adhesion molecule-1; IL-1β, interleukin-1β; IL-6, interleukin-6; JAMs, junctional adhesion molecules; LDL, low-density lipoprotein cholesterol; LPa, lipoprotein little a; LPS, lipopolysaccharide; MCP-1, monocyte chemotactic protein-1; NO, nitric oxide; Nox2, nicotinamide adenine dinucleotide phosphate oxidase (NADPH Ox); ONOO-, peroxinitrite; pCC, peripheral systemic cytokines/chemokines; pp, pulsatile pulse pressure;, NVU, neurovascular unit; RBC, red blood cell; Red arrows, denote increase; RONSS, reactive oxygen, nitrogen, sulfur species; ROS, reactive oxygen species; RSI, reactive species interactome; T, transcytosis; TJ/AJ, tight and adherens junctions; TNFα, tumor necrosis factor alpha; underlining, denotes emphasis; upward arrows, denote increase; VAS, vascular arterial stiffness; VCAM-1, vascular cellular adhesion molecule-1; WBC, white blood cell.
Figure 5.
Multiple injurious species affect brain endothelial cells (BECs) of the neurovascular unit (NVU). These injurious species contribute to BEC activation (proinflammatory) and dysfunction with decreased protective nitric oxide (NO) (BECact/dys) and blood–brain barrier dysfunction/disruption (BBBdd) with increased permeability via increased transcellular- transcytosis and paracellular leakiness. BECact/dys and BBBdd are bidirectional biomarkers for the development of enlarged perivascular spaces (EPVS) and cerebral small vessel disease (SVD). Note the red-dashed line at the top of this image, which designates the EC plasmalemma location of the multiple injurious species that are responsible for initial BEC injury in multiple clinical diseases and structural remodeling abnormalities including SVD and EPVS. BEC insulin resistance is associated with increased glucose, and the AGE receptor (RAGE) interaction is also important in the development of BECact/dys, BBBdd, and neurodegeneration. Also, note that angiotensin II (AngII) denotes the activation of the renin-angiotensin-aldosterone system (RAAS) including the endothelial cell mineralocorticoid receptor (ECMR) for aldosterone and that both play a role in the development of vascular arterial stiffening. Importantly, note that this image does not illustrate the endothelial glycocalyx. Modified image provided with permission by CC 4.0 [35RTIWH]. AGE, advanced glycation end products; aMMPs, activated matrix metalloproteinases; Ang II, angiotensin two; BBB, blood–brain barrier; BEC, brain endothelial cell; BBBdd, blood–brain barrier dysfunction and disruption; BECact/dys, brain endothelial cell activation/dysfunction; BH4, tetrahydrobiopterin; CCL2, chemokine (C-C motif) ligand 2; Cox-2, cyclo-oxygenase-2; Cox-2/PGE2 axis, cyclo-oxygenase-2/prostaglandin E2; downward arrows, arrowheads, denote call outs; ecGCx, endothelial glycocalyx; ICAM-1, intercellular adhesion molecule-1; IL-1β, interleukin-1β; IL-6, interleukin-6; JAMs, junctional adhesion molecules; LDL, low-density lipoprotein cholesterol; LPa, lipoprotein little a; LPS, lipopolysaccharide; MCP-1, monocyte chemotactic protein-1; NO, nitric oxide; Nox2, nicotinamide adenine dinucleotide phosphate oxidase (NADPH Ox); ONOO-, peroxinitrite; pCC, peripheral systemic cytokines/chemokines; pp, pulsatile pulse pressure;, NVU, neurovascular unit; RBC, red blood cell; Red arrows, denote increase; RONSS, reactive oxygen, nitrogen, sulfur species; ROS, reactive oxygen species; RSI, reactive species interactome; T, transcytosis; TJ/AJ, tight and adherens junctions; TNFα, tumor necrosis factor alpha; underlining, denotes emphasis; upward arrows, denote increase; VAS, vascular arterial stiffness; VCAM-1, vascular cellular adhesion molecule-1; WBC, white blood cell.
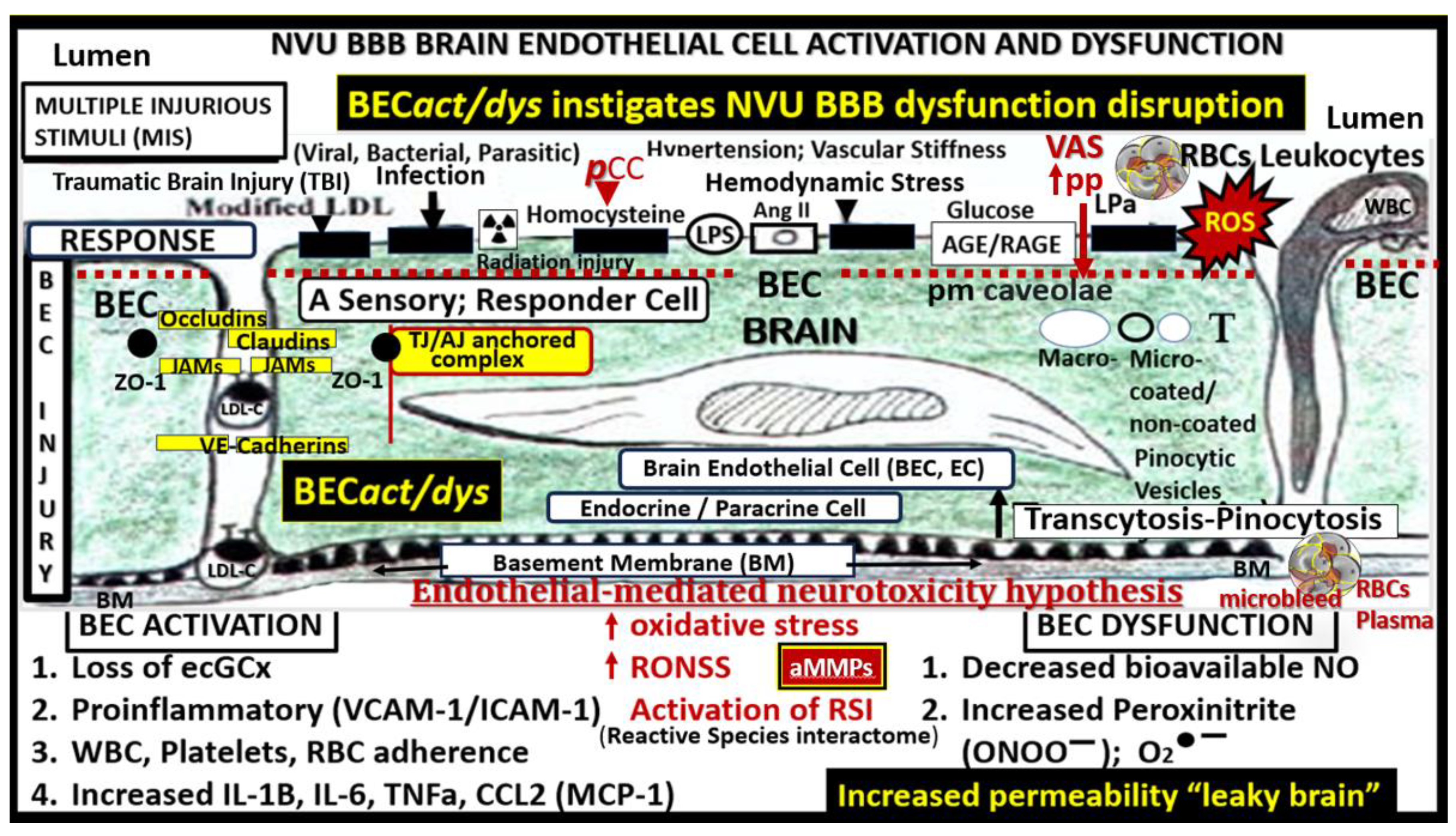
Figure 6.
Brain endothelial cell (BEC) signaling by peripheral cytokines/chemokines (pCC) and soluble lipopolysaccharide (sLPS) and LPS-enriched small extracellular vesicle exosome (lpsEVexos), to result in brain endothelial activation and dysfunction (BECact/dys). Note the insert lower-left, which depicts a small EVexo liberating multiple LPS-enriched vesicles and the metabolic signaling of BECs to result in central nervous system cytokines chemokines (cnsCC) and attracted microglia cells (atMGCs) via CCL5 (RANTES). BECact/dys results in neuroinflammation, blood–brain barrier (BBB) disruption, and enlarged perivascular spaces (EPVSs). Modified image provided with permission by CC4.0 [31,35]. atMGC, attracted microglia cell; CCL5, chemokine (C-C motif) ligand 5 (chemoattractant for microglia cells); CD14, cluster of differentiation 14; CCR5, -C-C chemokine receptor type 5, also known as CD195; CL, capillary lumen; EC, brain endothelial cell, brain; ecGCx, endothelial glycocalyx; EV, extracellular vesicle(s); EVMp, EVmicroparticles or microvesicles; EVexo, EVexosomes; IL1-β, -interleukin-1 beta; IL-6, interleukin-6; LPS, lipopolysaccharide; LBP, lipopolysaccharide-binding protein; MAPK, mitogen-activated protein kinase; MD2, myeloid differentiation protein 2; Mp, microparticles; N, nucleus; MtROS, mitochondrial reactive oxygen species; NADPH-Ox, nicotinamide adenine dinucleotide phosphate oxidase; NF-κB, nuclear factor-kappa B; NVU, neurovascular unit; PI3K/AKT = phosphatidylinositol 3-kinase/protein kinase B; PM, plasma membrane; RANTES, regulated on activation, normal T cell expressed and secreted; TLR4, toll-like receptor 4; TNFα, tumor necrosis alpha.
Figure 6.
Brain endothelial cell (BEC) signaling by peripheral cytokines/chemokines (pCC) and soluble lipopolysaccharide (sLPS) and LPS-enriched small extracellular vesicle exosome (lpsEVexos), to result in brain endothelial activation and dysfunction (BECact/dys). Note the insert lower-left, which depicts a small EVexo liberating multiple LPS-enriched vesicles and the metabolic signaling of BECs to result in central nervous system cytokines chemokines (cnsCC) and attracted microglia cells (atMGCs) via CCL5 (RANTES). BECact/dys results in neuroinflammation, blood–brain barrier (BBB) disruption, and enlarged perivascular spaces (EPVSs). Modified image provided with permission by CC4.0 [31,35]. atMGC, attracted microglia cell; CCL5, chemokine (C-C motif) ligand 5 (chemoattractant for microglia cells); CD14, cluster of differentiation 14; CCR5, -C-C chemokine receptor type 5, also known as CD195; CL, capillary lumen; EC, brain endothelial cell, brain; ecGCx, endothelial glycocalyx; EV, extracellular vesicle(s); EVMp, EVmicroparticles or microvesicles; EVexo, EVexosomes; IL1-β, -interleukin-1 beta; IL-6, interleukin-6; LPS, lipopolysaccharide; LBP, lipopolysaccharide-binding protein; MAPK, mitogen-activated protein kinase; MD2, myeloid differentiation protein 2; Mp, microparticles; N, nucleus; MtROS, mitochondrial reactive oxygen species; NADPH-Ox, nicotinamide adenine dinucleotide phosphate oxidase; NF-κB, nuclear factor-kappa B; NVU, neurovascular unit; PI3K/AKT = phosphatidylinositol 3-kinase/protein kinase B; PM, plasma membrane; RANTES, regulated on activation, normal T cell expressed and secreted; TLR4, toll-like receptor 4; TNFα, tumor necrosis alpha.
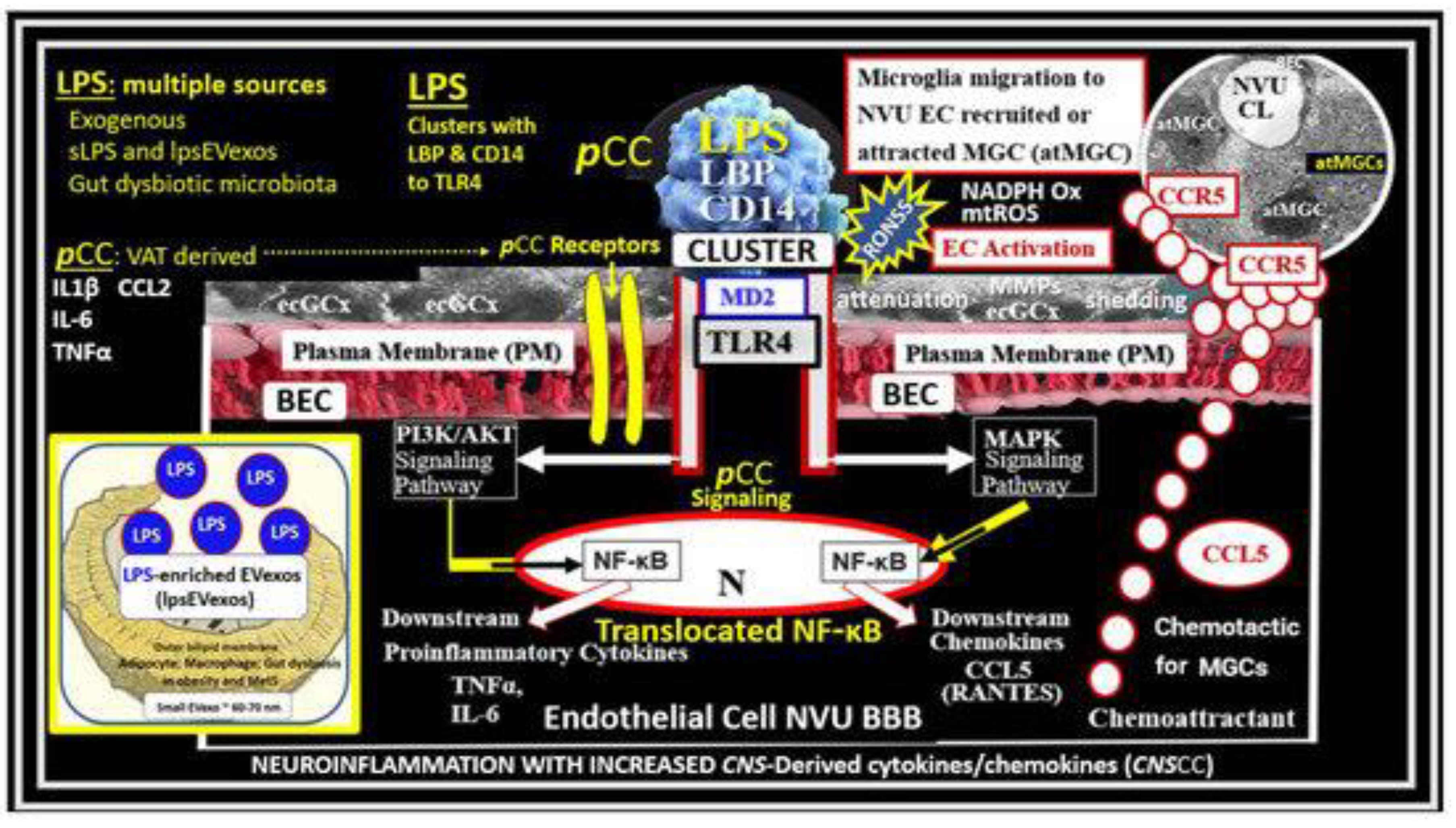
Figure 7.
Brain endothelial cell (BEC) glycocalyx (ecGCx) in lipopolysaccharide (LPS)-treated control male CD-1 mice at seven weeks of age with lanthanum nitrite (LAN) staining from frontal cortex layer III. (A and C) demonstrate the normal continuous electron-dense positive LAN staining of the ecGCx in control non-LPS treated, LAN-perfused mice. (B and D) male age-matched LPS-treated CD-1 models that depict attenuation, loss, or discontinuous ecGCx, which would allow for neurovascular unit (NVU) BEC activation and dysfunction and increased vulnerability of BEC with blood–brain barrier (BBB) disruption with increased permeability of fluids, cytokines and chemokines, cells including proinflammatory leukocytes. This increased permeability could result in dilated or enlarged perivascular spaces (EPVS). Notably the increased fluid and proinflammatory cellular entry could dilate the PVS to the point of enlargement. Also, note that LAN staining interferes with normal staining of the surrounding membranes in some images. Scale bars = 2 μm. Image provided with permission by CC 4.0 [40], Asterisk, basement membrane of brain endothelial cell; CL, capillary lumen; EC, brain endothelial cell; PcN, pericyte nucleus; VSMC, vascular smooth muscle cell.
Figure 7.
Brain endothelial cell (BEC) glycocalyx (ecGCx) in lipopolysaccharide (LPS)-treated control male CD-1 mice at seven weeks of age with lanthanum nitrite (LAN) staining from frontal cortex layer III. (A and C) demonstrate the normal continuous electron-dense positive LAN staining of the ecGCx in control non-LPS treated, LAN-perfused mice. (B and D) male age-matched LPS-treated CD-1 models that depict attenuation, loss, or discontinuous ecGCx, which would allow for neurovascular unit (NVU) BEC activation and dysfunction and increased vulnerability of BEC with blood–brain barrier (BBB) disruption with increased permeability of fluids, cytokines and chemokines, cells including proinflammatory leukocytes. This increased permeability could result in dilated or enlarged perivascular spaces (EPVS). Notably the increased fluid and proinflammatory cellular entry could dilate the PVS to the point of enlargement. Also, note that LAN staining interferes with normal staining of the surrounding membranes in some images. Scale bars = 2 μm. Image provided with permission by CC 4.0 [40], Asterisk, basement membrane of brain endothelial cell; CL, capillary lumen; EC, brain endothelial cell; PcN, pericyte nucleus; VSMC, vascular smooth muscle cell.
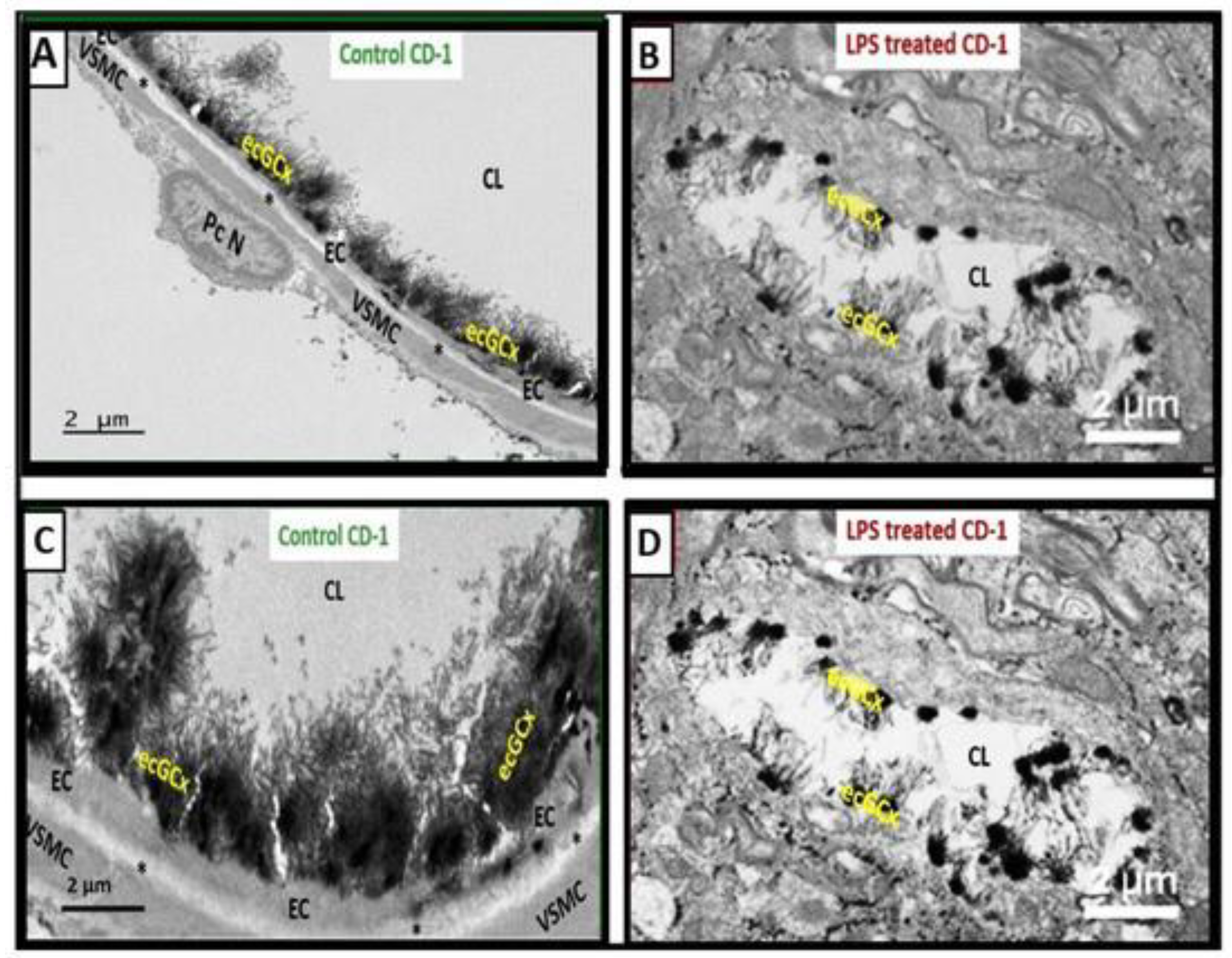
Figure 8.
Pericyte-endothelial morphology. (A) demonstrates an intra pancreatic islet circumferential pericyte (Pc) and its process enveloping a capillary endothelial cell (EC) with its cytoplasmic pericyte processes (PcP) that are representative of the brain NVU ECs and Pcs. Note that PcP are in intimate contact with EC at specific sites termed peg sockets (PS) and adherens junctions (Aj) arrows). Additionally, note the loose areolar interstitium (int) surround these two cells. The contact points between these two cells are illustrated in greater detail in panels B and D. Magnification x15,000; Bar = 500nm. (B) depicts an exploded image of A and the Pc has been pseudo-darkened to highlight and depict the communication cell-cell connections. (C) illustrates both types of EC-Pc communication and cell-cell connections. Note the peg socket connection (arrowhead) and the adherens junction (heterotypic N-cadherins, arrows) along with gap junctions connexin 43 (Cx43). Magnification x50,000; bar = 100nm. (D) depicts the PS connection between the EC and Pc at higher magnification and also shows the presence of a caveolae (arrows), which also provides communication between these two cells. Importantly, the ECs and Pcs are codependent on one another, in that, ECs supply nitric oxide and platelet derived growth factor beta (PDGFβ) to the enveloping Pc. and the Pcs supply vascular endothelial growth factor (VEGF) and the receptor for PDGFβ (not illustrated). Additionally, Pcs have contractile proteins (alpha smooth muscle actin), which allow vasoregulation (contraction or relaxation to control regional cerebral blood flow) and are known to stain positive for neuroglia 2 (NG2) in contrast to ECs (not shown). Magnification x150,000; bar =50nm. Insert (d) is an exploded image of the adherens junction in panel C. Image provided with permission by CC 4.0 [49]. .
Figure 8.
Pericyte-endothelial morphology. (A) demonstrates an intra pancreatic islet circumferential pericyte (Pc) and its process enveloping a capillary endothelial cell (EC) with its cytoplasmic pericyte processes (PcP) that are representative of the brain NVU ECs and Pcs. Note that PcP are in intimate contact with EC at specific sites termed peg sockets (PS) and adherens junctions (Aj) arrows). Additionally, note the loose areolar interstitium (int) surround these two cells. The contact points between these two cells are illustrated in greater detail in panels B and D. Magnification x15,000; Bar = 500nm. (B) depicts an exploded image of A and the Pc has been pseudo-darkened to highlight and depict the communication cell-cell connections. (C) illustrates both types of EC-Pc communication and cell-cell connections. Note the peg socket connection (arrowhead) and the adherens junction (heterotypic N-cadherins, arrows) along with gap junctions connexin 43 (Cx43). Magnification x50,000; bar = 100nm. (D) depicts the PS connection between the EC and Pc at higher magnification and also shows the presence of a caveolae (arrows), which also provides communication between these two cells. Importantly, the ECs and Pcs are codependent on one another, in that, ECs supply nitric oxide and platelet derived growth factor beta (PDGFβ) to the enveloping Pc. and the Pcs supply vascular endothelial growth factor (VEGF) and the receptor for PDGFβ (not illustrated). Additionally, Pcs have contractile proteins (alpha smooth muscle actin), which allow vasoregulation (contraction or relaxation to control regional cerebral blood flow) and are known to stain positive for neuroglia 2 (NG2) in contrast to ECs (not shown). Magnification x150,000; bar =50nm. Insert (d) is an exploded image of the adherens junction in panel C. Image provided with permission by CC 4.0 [49]. .
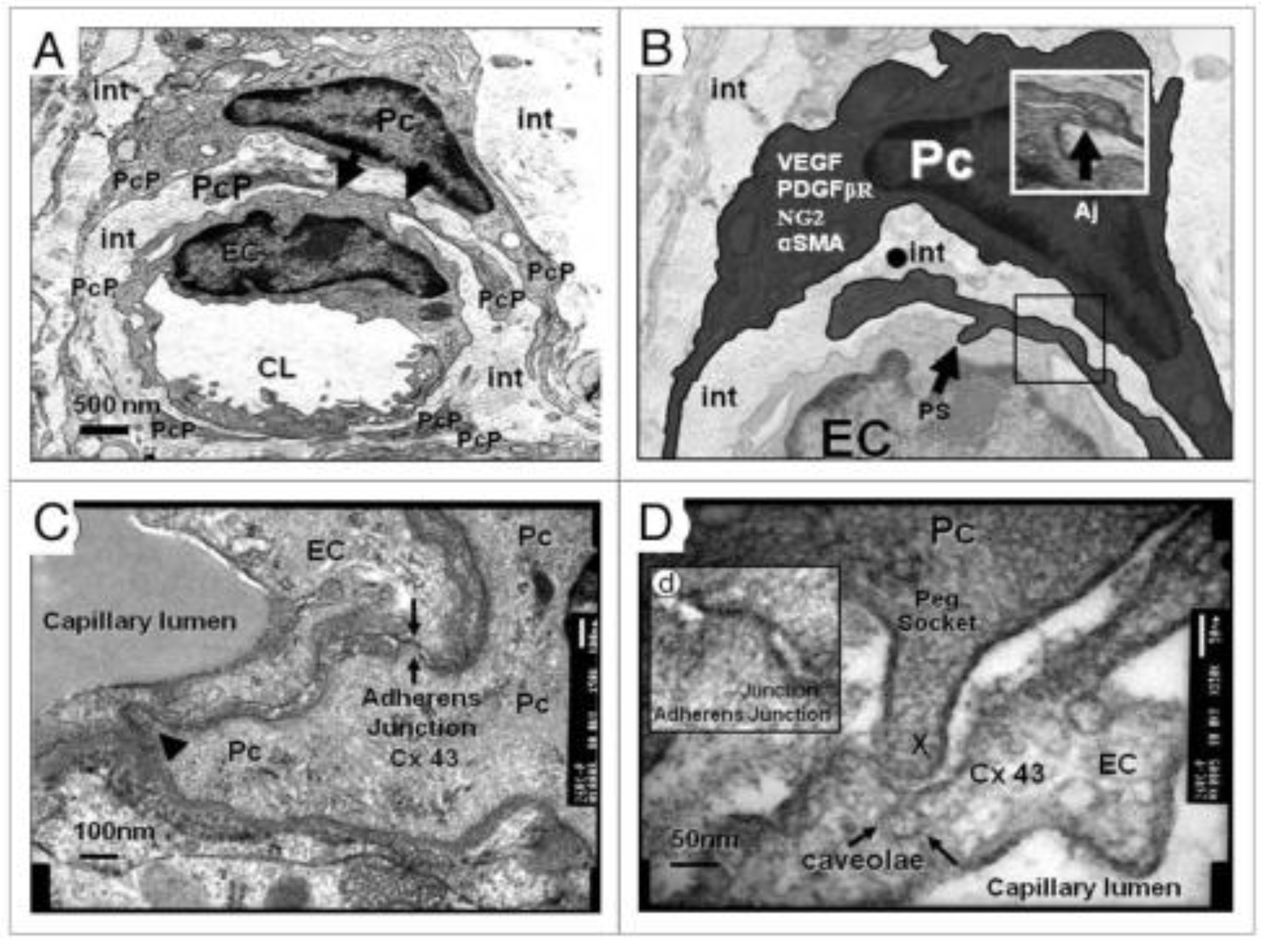
Figure 8.
Astrocyte(s) (ACs) connection/communication with other astrocytes and multiple cells within the brain universe. ACs are ectodermal, neuroepithelial-derived, and star-shaped cells that provide protection, defense, and homeostasis of the brain and spinal cord. This montage illustration of hand-drawn images and transmission electron micrographs allows one to observe the resident non-reactive protoplasmic AC soma, as seen in the center of this image that connects to virtually every cell within the brain universe. ACs may thus be considered the master connecting cells of the central nervous system (CNS) against a background of distant white dots representing even more ACs stars within the brain universe from control C57BL/6J models (hand-drawn computer-assisted illustration from toluidine blue stained images). Importantly, the AC-to-AC connections via gap junction connexins such as Cx43 allow them to form a syncytium to coordinate brain function and proper cognition (insert number 5). Insert 1 depicts the protoplasmic perivascular astrocyte endfeet (pvACef) processes (pseudo-colored golden) connecting to the neurovascular unit (NVU) capillary, which allows for neurovascular coupling in frontal cortex layer III in a non-diabetic control female C57BL/6J mouse model at 20 weeks of age. Insert 2 illustrates the protoplasmic perisynaptic astrocyte endfeet (psACef) cradling of pre- and post-synaptic neurons and emphasizes its importance to the tripartite cradle synapse in cortical layer III. Insert 3 illustrates the connection of the fibrous ACs (pseudo-colored yellow-gold and the formation of myelin) to myelinated axons with axoplasm (pseudo-colored cyan) of neurons in white matter in a control at 20 weeks C57BL/6J. Insert 4 illustrates the lost connections between a reactive microglial cell (rMGC) (pseudo-colored blue) and multiple reactive detached and separated ACs (pseudo-colored red) adjacent to a neurovascular unit (NVU) with a single intact non-reactive ACs (pseudo-colored yellow) in diabetic db/db model cortical layer III at 20 weeks of age. Insert 5 demonstrates AC-to-AC connections in cortical layer III in control models (hand-drawn computer-assisted illustration of light microscopic toluidine blue stained images from control C57BL/6J models) via gap junction connexins. Only Inserts 1–4 have scale bars of 0.5 μm, 100 nm, 1 μm, and 2 μm, respectively. The background is also a hand-drawn computer-assisted image derived from control C57BL/6J models toluidine blue stained models and does not have a scale bar. Modified image provided with permission from CC 4.0 [55]. ACfp, protoplasmic astrocyte endfeet; ACPVef, astrocyte perivascular endfeet; Cap L, capillary lumen; EC, brain endothelial cell; iMGC, interrogating microglial cell; Mt, mitochondria; N, nucleus; Pc, pericyte; PSD, post-synaptic density; PVACef, perivascular astrocyte endfeet; rMGC, reactive microglia cell; psACef, perisynaptic astrocyte endfeet.
Figure 8.
Astrocyte(s) (ACs) connection/communication with other astrocytes and multiple cells within the brain universe. ACs are ectodermal, neuroepithelial-derived, and star-shaped cells that provide protection, defense, and homeostasis of the brain and spinal cord. This montage illustration of hand-drawn images and transmission electron micrographs allows one to observe the resident non-reactive protoplasmic AC soma, as seen in the center of this image that connects to virtually every cell within the brain universe. ACs may thus be considered the master connecting cells of the central nervous system (CNS) against a background of distant white dots representing even more ACs stars within the brain universe from control C57BL/6J models (hand-drawn computer-assisted illustration from toluidine blue stained images). Importantly, the AC-to-AC connections via gap junction connexins such as Cx43 allow them to form a syncytium to coordinate brain function and proper cognition (insert number 5). Insert 1 depicts the protoplasmic perivascular astrocyte endfeet (pvACef) processes (pseudo-colored golden) connecting to the neurovascular unit (NVU) capillary, which allows for neurovascular coupling in frontal cortex layer III in a non-diabetic control female C57BL/6J mouse model at 20 weeks of age. Insert 2 illustrates the protoplasmic perisynaptic astrocyte endfeet (psACef) cradling of pre- and post-synaptic neurons and emphasizes its importance to the tripartite cradle synapse in cortical layer III. Insert 3 illustrates the connection of the fibrous ACs (pseudo-colored yellow-gold and the formation of myelin) to myelinated axons with axoplasm (pseudo-colored cyan) of neurons in white matter in a control at 20 weeks C57BL/6J. Insert 4 illustrates the lost connections between a reactive microglial cell (rMGC) (pseudo-colored blue) and multiple reactive detached and separated ACs (pseudo-colored red) adjacent to a neurovascular unit (NVU) with a single intact non-reactive ACs (pseudo-colored yellow) in diabetic db/db model cortical layer III at 20 weeks of age. Insert 5 demonstrates AC-to-AC connections in cortical layer III in control models (hand-drawn computer-assisted illustration of light microscopic toluidine blue stained images from control C57BL/6J models) via gap junction connexins. Only Inserts 1–4 have scale bars of 0.5 μm, 100 nm, 1 μm, and 2 μm, respectively. The background is also a hand-drawn computer-assisted image derived from control C57BL/6J models toluidine blue stained models and does not have a scale bar. Modified image provided with permission from CC 4.0 [55]. ACfp, protoplasmic astrocyte endfeet; ACPVef, astrocyte perivascular endfeet; Cap L, capillary lumen; EC, brain endothelial cell; iMGC, interrogating microglial cell; Mt, mitochondria; N, nucleus; Pc, pericyte; PSD, post-synaptic density; PVACef, perivascular astrocyte endfeet; rMGC, reactive microglia cell; psACef, perisynaptic astrocyte endfeet.
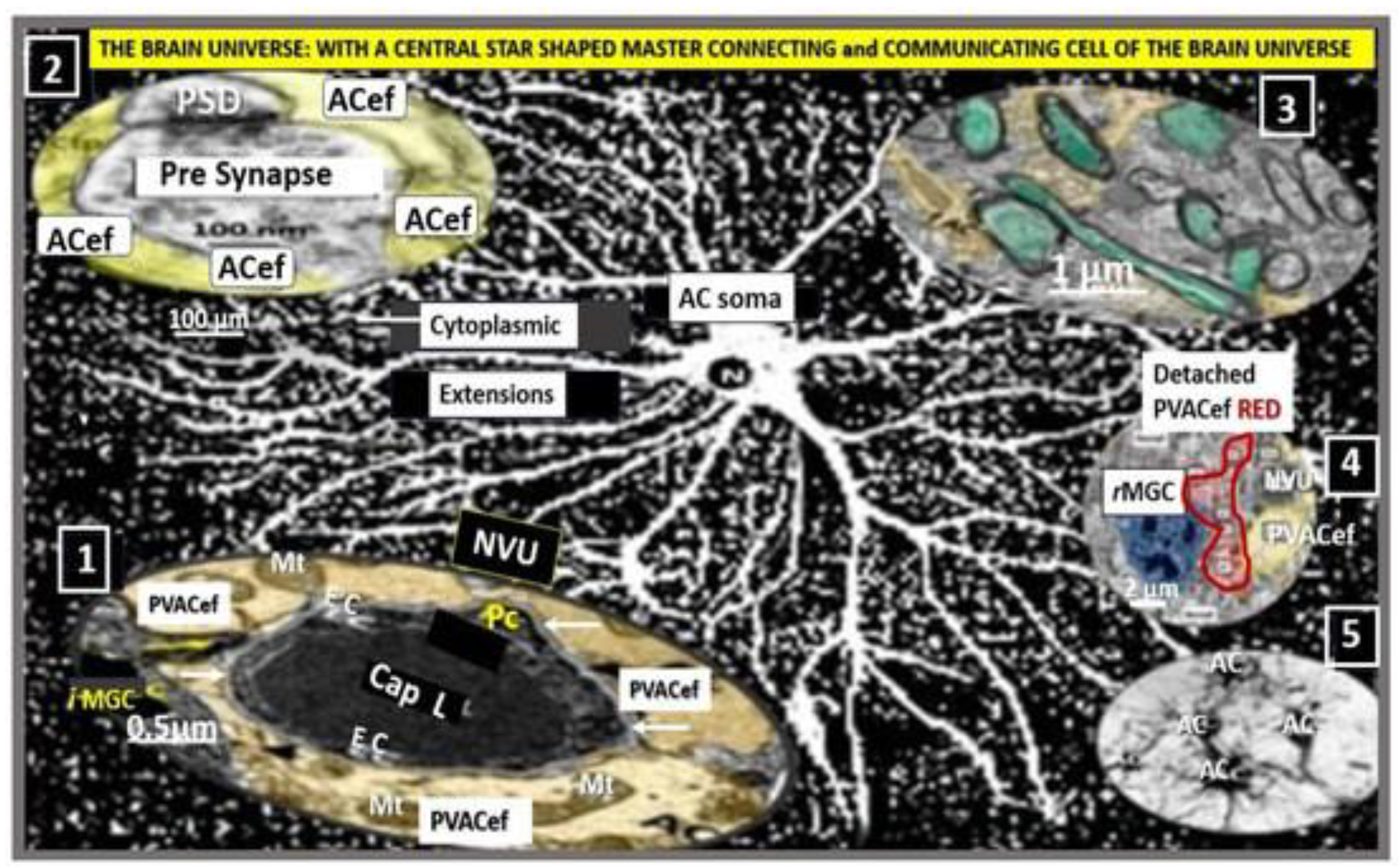
Figure 9.
Normal Capillary Neurovascular Unit (NVU), Perivascular Unit (PVU), Perivascular spaces (PVS), and Enlarged Perivascular Spaces (EPVS). (A) illustrates a hand-drawn representative image of the normal healthy capillary NVU blood-brain barrier (BBB) consisting of the brain endothelial glycocalyx (ecGCx, aqua color), brain endothelial cell(s) (EC white) with their tight and adherence junction(s) (TJ/AJ asterisks), pericytes and their endfeet (Pc, green color), and the protoplasmic perivascular astrocyte endfeet (ACef, blue color). Note the innate immune system neutrophil (N) within the lumen of the capillary NVU and step 1, which depicts the initial injury to the NVU that allows the N to breach the NVU BBB. (B) depicts the PVU, which lies immediately adjacent to the NVU in this longitudinal illustration that contains both the normal PVS (green color) and the EPVS (red color) within the postcapillary venule. Note the blue ACef that are a continuous and creating the outermost boundary cells of the PVS within the PVU in panel B. Also, note how the normal blue ACef become the purple reactive ACef as the PVS remodel to become EPVS that are associated with PVS inflammation (proinflammatory neutrophils, monocytes, and lymphocytes where there exists a crossroad for the multiple cellular crosstalk between the proinflammatory leukocytes and the resident reactive antigen presenting perivascular macrophage (PVMΦ) (pseudo-colored red). Notably, as these pvACef become reactive they have now detached and retracted from the combined basement membranes of the BECs and Pcs as depicted in Figure 1 to form the outermost cell of the perivascular space (blue color) to form the reactive pvACef (purple color). Importantly, note that the interactions between the proinflammatory leucocytes and the reactive PVMΦs resulting in increased cytokines/chemokines (promoting neuroinflammation) and oxidative stress (ROS), which activate local MMPs to degrade the pvACef basal lamina-glia limitans to result in step 2 (breeching of the PVS) to result in neuroinflammation as described by Owens et al. in his 2-step process of neuroinflammation [10]. (C) depicts how the EPVS are associated with dysfunctional reactive purple pvACef, activated BECs with plasma membrane ruffling and increased transcytosis, and rounding of the pericyte nuclei (PcN) with retraction of pericyte endfeet (Pcef) in LPS-treated CD1 male mice. Importantly, the now reactive resident perivascular macrophages (PVMΦ) as a result of step 1 result in their important role as an antigen presenting cells to affect the recruitment of reactive lymphocytes of the adaptive immune system with resulting neuroinflammation. Modified image provided with permission by CC 4.0 [30,55]. ACef, perivascular astrocyte endfeet; dAQP4, dysfunctional aquaporin 4 water channel; BEC, brain endothelial cell; Dys, dysfunctional; EC, brain endothelial cell; EPVS, enlarged perivascular space(s) (double arrows); LPS, lipopolysaccharide; Lys, lysosome; MMP(s), matrix metalloproteinases; N, neutrophile and nucleus; Pc, pericyte; PcN, pericyte nucleus; Pcfp, pericyte foot process endfeet; pm, plasma membrane; rpvACef and pvMΦ, resident/reactive perivascular macrophage; ROS, reactive oxygen species.
Figure 9.
Normal Capillary Neurovascular Unit (NVU), Perivascular Unit (PVU), Perivascular spaces (PVS), and Enlarged Perivascular Spaces (EPVS). (A) illustrates a hand-drawn representative image of the normal healthy capillary NVU blood-brain barrier (BBB) consisting of the brain endothelial glycocalyx (ecGCx, aqua color), brain endothelial cell(s) (EC white) with their tight and adherence junction(s) (TJ/AJ asterisks), pericytes and their endfeet (Pc, green color), and the protoplasmic perivascular astrocyte endfeet (ACef, blue color). Note the innate immune system neutrophil (N) within the lumen of the capillary NVU and step 1, which depicts the initial injury to the NVU that allows the N to breach the NVU BBB. (B) depicts the PVU, which lies immediately adjacent to the NVU in this longitudinal illustration that contains both the normal PVS (green color) and the EPVS (red color) within the postcapillary venule. Note the blue ACef that are a continuous and creating the outermost boundary cells of the PVS within the PVU in panel B. Also, note how the normal blue ACef become the purple reactive ACef as the PVS remodel to become EPVS that are associated with PVS inflammation (proinflammatory neutrophils, monocytes, and lymphocytes where there exists a crossroad for the multiple cellular crosstalk between the proinflammatory leukocytes and the resident reactive antigen presenting perivascular macrophage (PVMΦ) (pseudo-colored red). Notably, as these pvACef become reactive they have now detached and retracted from the combined basement membranes of the BECs and Pcs as depicted in Figure 1 to form the outermost cell of the perivascular space (blue color) to form the reactive pvACef (purple color). Importantly, note that the interactions between the proinflammatory leucocytes and the reactive PVMΦs resulting in increased cytokines/chemokines (promoting neuroinflammation) and oxidative stress (ROS), which activate local MMPs to degrade the pvACef basal lamina-glia limitans to result in step 2 (breeching of the PVS) to result in neuroinflammation as described by Owens et al. in his 2-step process of neuroinflammation [10]. (C) depicts how the EPVS are associated with dysfunctional reactive purple pvACef, activated BECs with plasma membrane ruffling and increased transcytosis, and rounding of the pericyte nuclei (PcN) with retraction of pericyte endfeet (Pcef) in LPS-treated CD1 male mice. Importantly, the now reactive resident perivascular macrophages (PVMΦ) as a result of step 1 result in their important role as an antigen presenting cells to affect the recruitment of reactive lymphocytes of the adaptive immune system with resulting neuroinflammation. Modified image provided with permission by CC 4.0 [30,55]. ACef, perivascular astrocyte endfeet; dAQP4, dysfunctional aquaporin 4 water channel; BEC, brain endothelial cell; Dys, dysfunctional; EC, brain endothelial cell; EPVS, enlarged perivascular space(s) (double arrows); LPS, lipopolysaccharide; Lys, lysosome; MMP(s), matrix metalloproteinases; N, neutrophile and nucleus; Pc, pericyte; PcN, pericyte nucleus; Pcfp, pericyte foot process endfeet; pm, plasma membrane; rpvACef and pvMΦ, resident/reactive perivascular macrophage; ROS, reactive oxygen species.
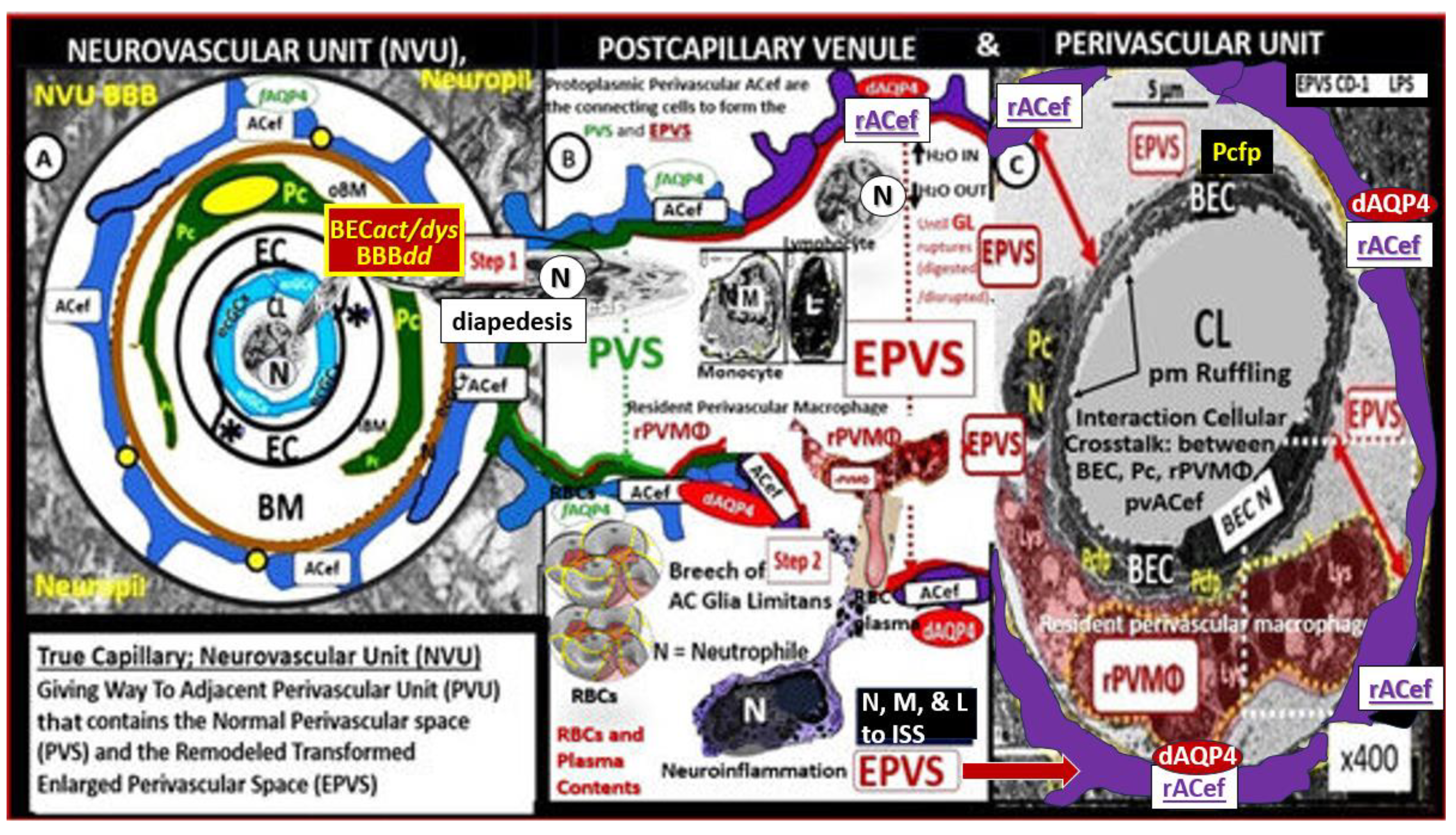
Figure 10.
Comparing perivascular astrocyte endfeet (pvACef) and the cradling perisynaptic astrocyte endfeet (psACef) detachment and separation. These similarities implicate damaged or dysfunction aquaporin-4 (AQP4) either due to activated proteases such as matrix metalloproteinases (MMP-2, 9) or a loss of polarization of the bidirectional water channel AQP4 from the plasma membranes facing the synapse resulting in impaired synaptic transmission and impaired cognition or the timing of arrival of incoming information to disturb networks of information processing. (A, C) are female age-matched controls and (B, D) are from 20-week-old diabetic db/db models with tissues obtained from the frontal cortex, cortical layer III, and depict detachment and separation of pvACef in (B) and psACef in panel D. Note that this detachment and separation create a perivascular space (PVS) (B) and a perisynaptic space (D) that may continue to become enlarged with dysfunctional dystroglycan (dysDG) and dysfunctional aquaporin-4 (dysAQP4) as in (panels B, D). Note that the cyan-green line denoting the glia limitans in (panel B) is not present in (D). Blue arrows represent detachment and separation of pvACef. Red dotted arrows represent detachment and separation with emphasis. Faint yellow numbers indicate numbering of pvACef (1-7). (A, B, D) represent cleaned and cut original images in order to demonstrate separation. Image reproduced with permission by CC 4.0 [55]. Scale bars =100 nm (D, E). BBB = blood–brain barrier; CL = capillary lumen; dys = dysfunctional; DG = dystroglycans; EC = brain endothelial cell; NVU = neurovascular unit; PcP = pericyte process; PSD = post synaptic density; TJ/AJ = tight and adherens junctions.
Figure 10.
Comparing perivascular astrocyte endfeet (pvACef) and the cradling perisynaptic astrocyte endfeet (psACef) detachment and separation. These similarities implicate damaged or dysfunction aquaporin-4 (AQP4) either due to activated proteases such as matrix metalloproteinases (MMP-2, 9) or a loss of polarization of the bidirectional water channel AQP4 from the plasma membranes facing the synapse resulting in impaired synaptic transmission and impaired cognition or the timing of arrival of incoming information to disturb networks of information processing. (A, C) are female age-matched controls and (B, D) are from 20-week-old diabetic db/db models with tissues obtained from the frontal cortex, cortical layer III, and depict detachment and separation of pvACef in (B) and psACef in panel D. Note that this detachment and separation create a perivascular space (PVS) (B) and a perisynaptic space (D) that may continue to become enlarged with dysfunctional dystroglycan (dysDG) and dysfunctional aquaporin-4 (dysAQP4) as in (panels B, D). Note that the cyan-green line denoting the glia limitans in (panel B) is not present in (D). Blue arrows represent detachment and separation of pvACef. Red dotted arrows represent detachment and separation with emphasis. Faint yellow numbers indicate numbering of pvACef (1-7). (A, B, D) represent cleaned and cut original images in order to demonstrate separation. Image reproduced with permission by CC 4.0 [55]. Scale bars =100 nm (D, E). BBB = blood–brain barrier; CL = capillary lumen; dys = dysfunctional; DG = dystroglycans; EC = brain endothelial cell; NVU = neurovascular unit; PcP = pericyte process; PSD = post synaptic density; TJ/AJ = tight and adherens junctions.
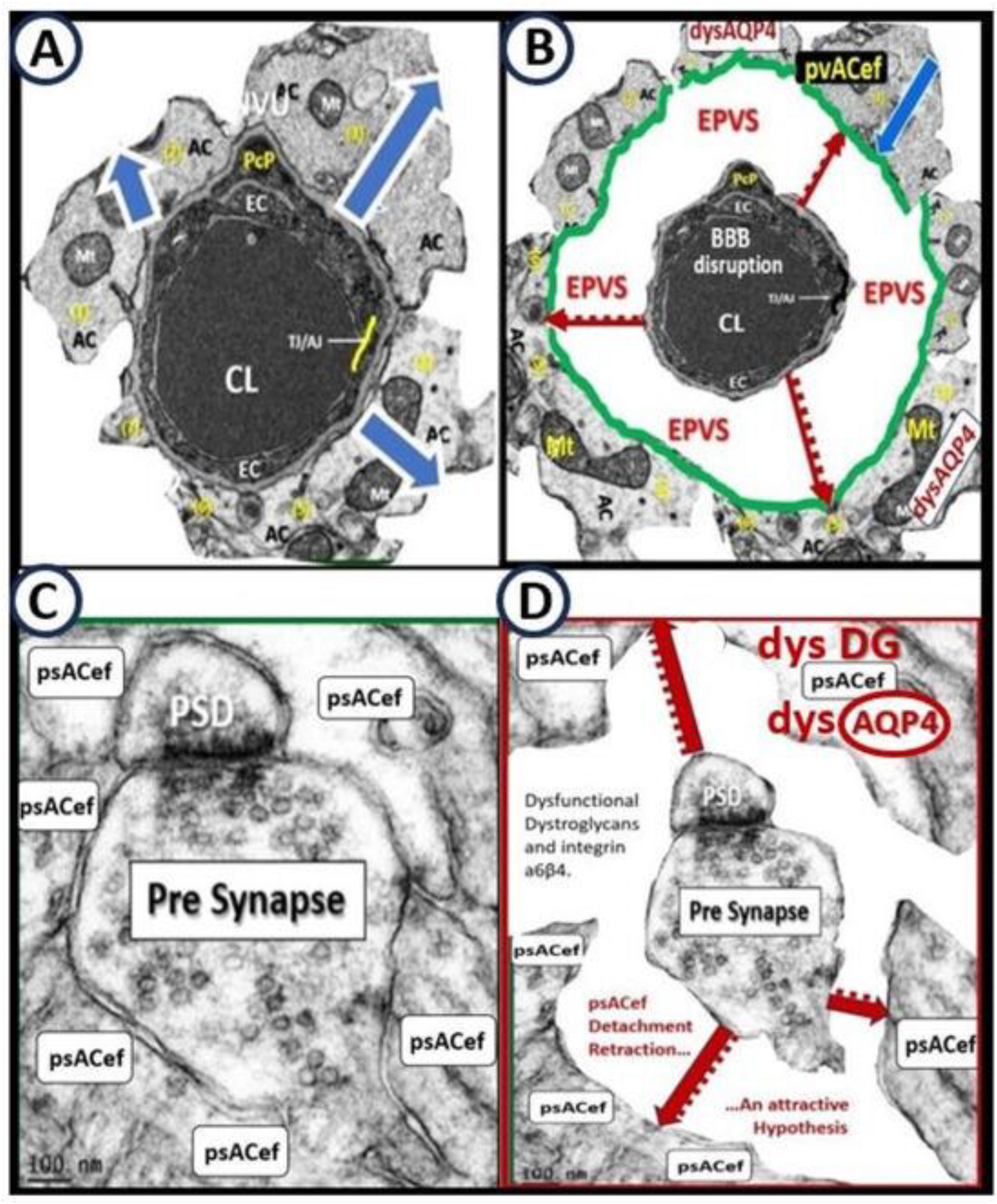
Figure 11.
Multicellular cell-cell contact and communication in the neurovascular and perivascular units. Perivascular spaces (PVS) and enlarged PVS (EPVS) provide a specialized holding space, which serves as a crossroad for multicellular crosstalk between brain endothelial cell(s) (BECs), pericyte(s) (Pcs) and pericyte foot processes-endfeet (Pcfps), resident reactive perivascular macrophage(s) (rPVMΦs), leukocytes, and reactive perivascular astrocyte endfeet (ACef). (A) demonstrates a healthy capillary neurovascular unit (NVU). Note the BEC and encircling Pc and its Pc foot processes (Pcfp) have a unique cell-cell physical contact for cell-cell communication via the peg socket morphology and phenotype along with its N-cadherin junctions and gap junction protein connexin 43 (Cx43). (B) depicts the PVS remodeled EPVS with its cellular contents of a rPVMΦ. Importantly note that there are three close intimate cell-cell contact regions for cellular crosstalk including (1). BEC/rPVMΦ (yellow boxed-in dashed lines and asterisk); (2). Pcfp/rPVMΦ (red boxed-in dashed lines; (3). rPVMΦ/ACef (white boxed-in dashed lines). Thus, this figure identifies extensive crosstalk communication between NVU BECs, Pc-Pcef, rPVMΦs, and ACef. Double arrows depict this cell-cell crosstalk communication. Modified image provided with permission by CC 4.0 [72]. .
Figure 11.
Multicellular cell-cell contact and communication in the neurovascular and perivascular units. Perivascular spaces (PVS) and enlarged PVS (EPVS) provide a specialized holding space, which serves as a crossroad for multicellular crosstalk between brain endothelial cell(s) (BECs), pericyte(s) (Pcs) and pericyte foot processes-endfeet (Pcfps), resident reactive perivascular macrophage(s) (rPVMΦs), leukocytes, and reactive perivascular astrocyte endfeet (ACef). (A) demonstrates a healthy capillary neurovascular unit (NVU). Note the BEC and encircling Pc and its Pc foot processes (Pcfp) have a unique cell-cell physical contact for cell-cell communication via the peg socket morphology and phenotype along with its N-cadherin junctions and gap junction protein connexin 43 (Cx43). (B) depicts the PVS remodeled EPVS with its cellular contents of a rPVMΦ. Importantly note that there are three close intimate cell-cell contact regions for cellular crosstalk including (1). BEC/rPVMΦ (yellow boxed-in dashed lines and asterisk); (2). Pcfp/rPVMΦ (red boxed-in dashed lines; (3). rPVMΦ/ACef (white boxed-in dashed lines). Thus, this figure identifies extensive crosstalk communication between NVU BECs, Pc-Pcef, rPVMΦs, and ACef. Double arrows depict this cell-cell crosstalk communication. Modified image provided with permission by CC 4.0 [72]. .
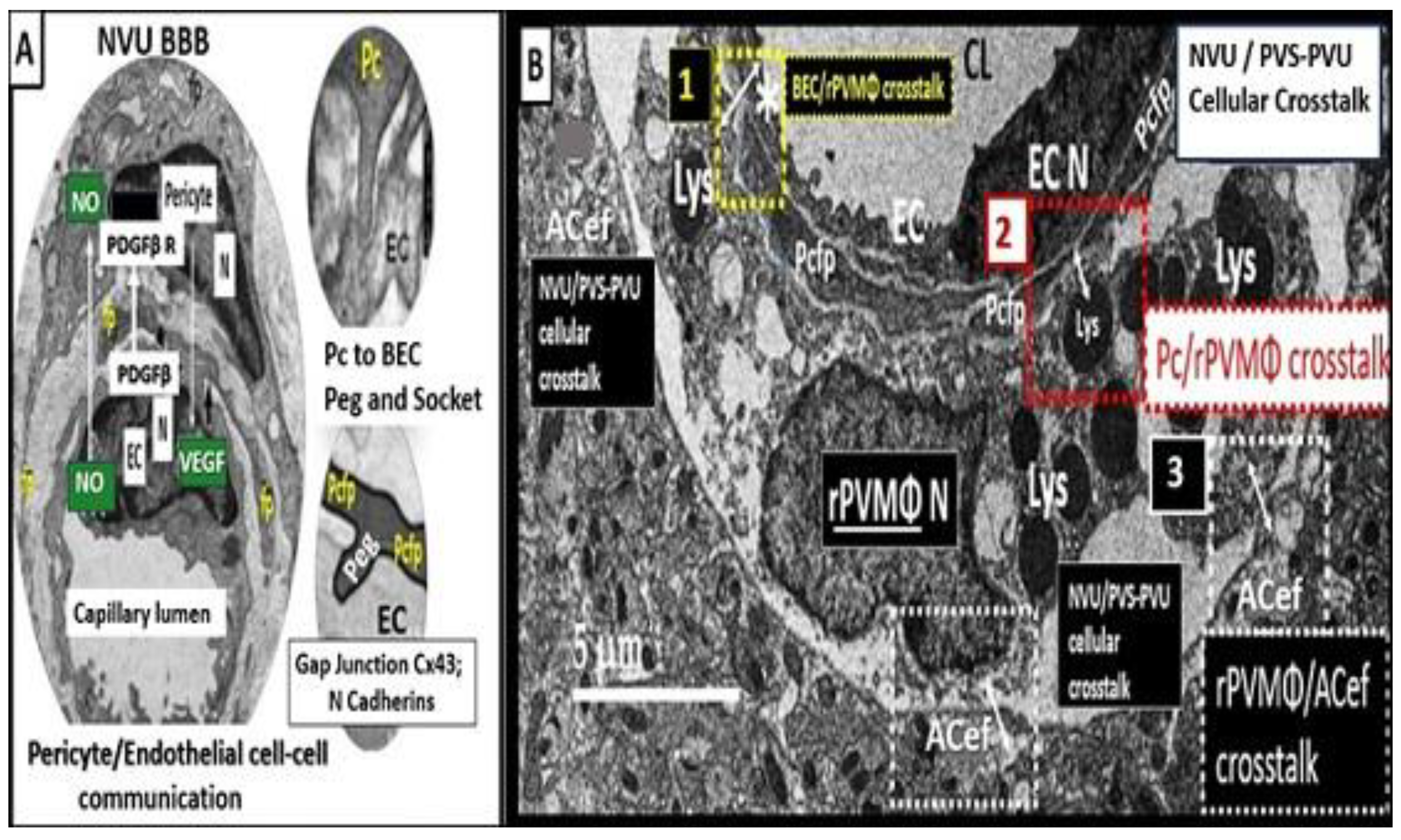
Figure 12.
Ultrastructural image of interstitial spaces (ISS) and illustration of interstitial fluid (ISF) flow to the perivascular spaces (PVS) and the glymphatic system (GS). ISS are nanometer structures (pseudo-colored blue with red outlines); however, they combine to create an approximate total of 20 percent of total brain volume and empty their interstitial fluid contents and metabolic wastes into the PVS – GS to be delivered to the subarachnoid space (SAS) and the cerebrospinal fluid (CSF) to the periphery. (A) demonstrates the nanosized diameter of ISS containing the ISS in a control 20-week-old mouse model from the frontal cortical grey matter. (B) depicts how the ISS containing the ISF, solutes (black dots), and metabolic waste (purple star) empties into the postcapillary venule perivascular space (PVS) or the conduit of the GS, which is responsible for toxic waste and excess water clearance. AC, astrocyte; ACef, astrocyte endfeet; EC, brain endothelial cell; CL = capillary lumen.
Figure 12.
Ultrastructural image of interstitial spaces (ISS) and illustration of interstitial fluid (ISF) flow to the perivascular spaces (PVS) and the glymphatic system (GS). ISS are nanometer structures (pseudo-colored blue with red outlines); however, they combine to create an approximate total of 20 percent of total brain volume and empty their interstitial fluid contents and metabolic wastes into the PVS – GS to be delivered to the subarachnoid space (SAS) and the cerebrospinal fluid (CSF) to the periphery. (A) demonstrates the nanosized diameter of ISS containing the ISS in a control 20-week-old mouse model from the frontal cortical grey matter. (B) depicts how the ISS containing the ISF, solutes (black dots), and metabolic waste (purple star) empties into the postcapillary venule perivascular space (PVS) or the conduit of the GS, which is responsible for toxic waste and excess water clearance. AC, astrocyte; ACef, astrocyte endfeet; EC, brain endothelial cell; CL = capillary lumen.
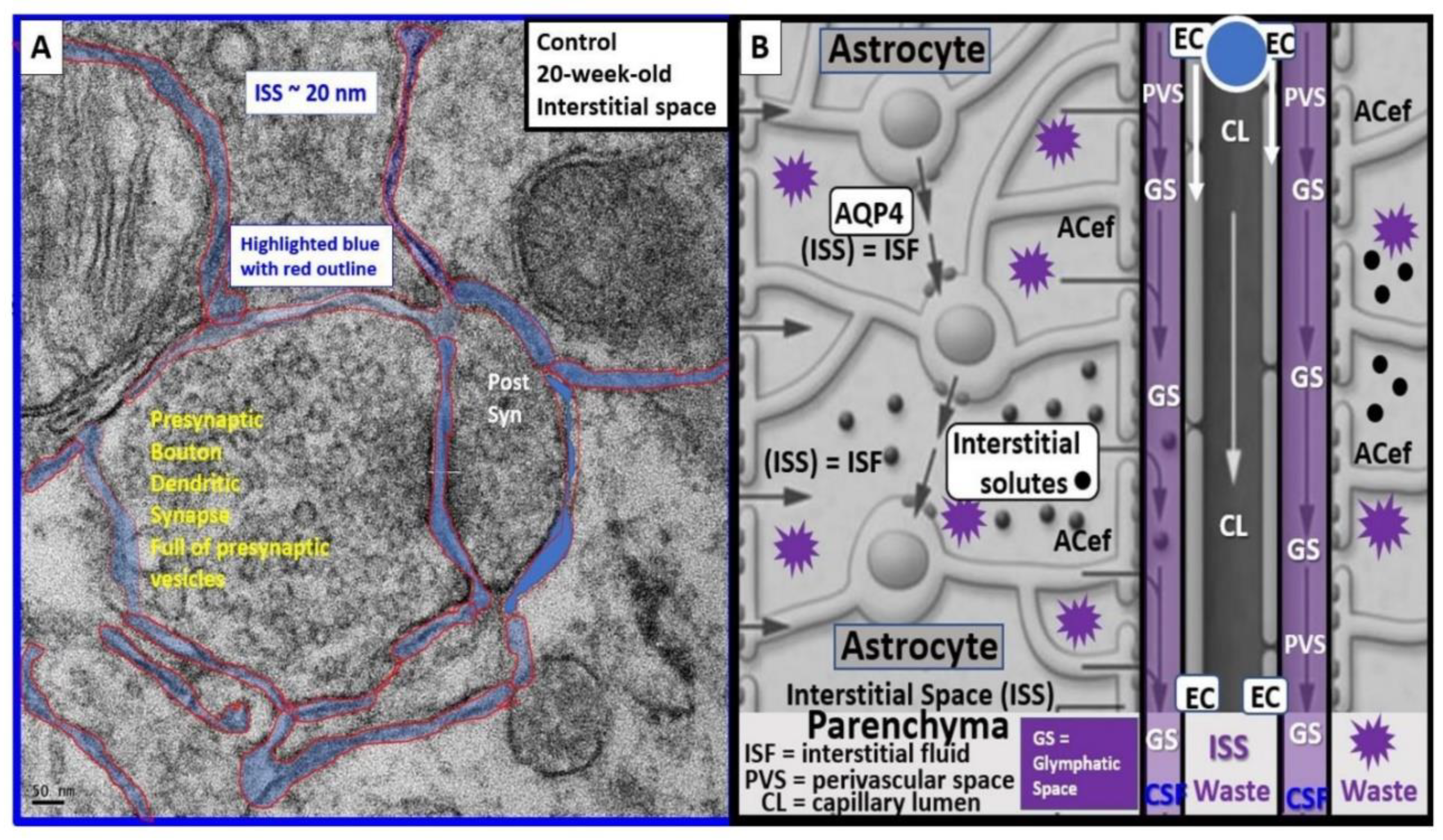
Figure 13.
Reactive microglia cell (aMGC) invasion of the neurovascular unit (NVU) with detachment and separation of perivascular astrocyte endfeet (pvACef) are protected with empagliflozin. (A) left-hand side of image depicts the invasion of a reactive microglia cell aMGC (pseudo-colored red) that appears to totally engulf the capillary NVU with detachment and retraction of perivascular astrocyte endfeet (drAC; pseudo-colored blue). Also, note the aberrant mitochondria (aMt) (pseudo-colored yellow with red outline) in DBC (20-week-old female db/db obese diabetic models that are responsible for reactive oxygen species (ROS) production and leakage. Dashed arrows depict the detachment and retraction of pvACef that are associated with reactive aMGC invasion of the NVU. (B) illustrates how empagliflozin protects the NVU from aberrant remodeling, which appeared similar to control models. Note the intact pvACef (pseudo-colored golden) and that the mitochondria in DBE are electron dense and not aberrant as in the DBC. Image provided with permission by CC 4.0 [91]. Magnification x2500, scale bar = 1 µm. aMGC, reactive microglia cell; aMt, aberrant mitochondria; drAC, detached retracted pvACef; EC, brain endothelial cell; CL, NVU capillary lumen; Pc, pericyte foot process; RBC, red blood cell.
Figure 13.
Reactive microglia cell (aMGC) invasion of the neurovascular unit (NVU) with detachment and separation of perivascular astrocyte endfeet (pvACef) are protected with empagliflozin. (A) left-hand side of image depicts the invasion of a reactive microglia cell aMGC (pseudo-colored red) that appears to totally engulf the capillary NVU with detachment and retraction of perivascular astrocyte endfeet (drAC; pseudo-colored blue). Also, note the aberrant mitochondria (aMt) (pseudo-colored yellow with red outline) in DBC (20-week-old female db/db obese diabetic models that are responsible for reactive oxygen species (ROS) production and leakage. Dashed arrows depict the detachment and retraction of pvACef that are associated with reactive aMGC invasion of the NVU. (B) illustrates how empagliflozin protects the NVU from aberrant remodeling, which appeared similar to control models. Note the intact pvACef (pseudo-colored golden) and that the mitochondria in DBE are electron dense and not aberrant as in the DBC. Image provided with permission by CC 4.0 [91]. Magnification x2500, scale bar = 1 µm. aMGC, reactive microglia cell; aMt, aberrant mitochondria; drAC, detached retracted pvACef; EC, brain endothelial cell; CL, NVU capillary lumen; Pc, pericyte foot process; RBC, red blood cell.
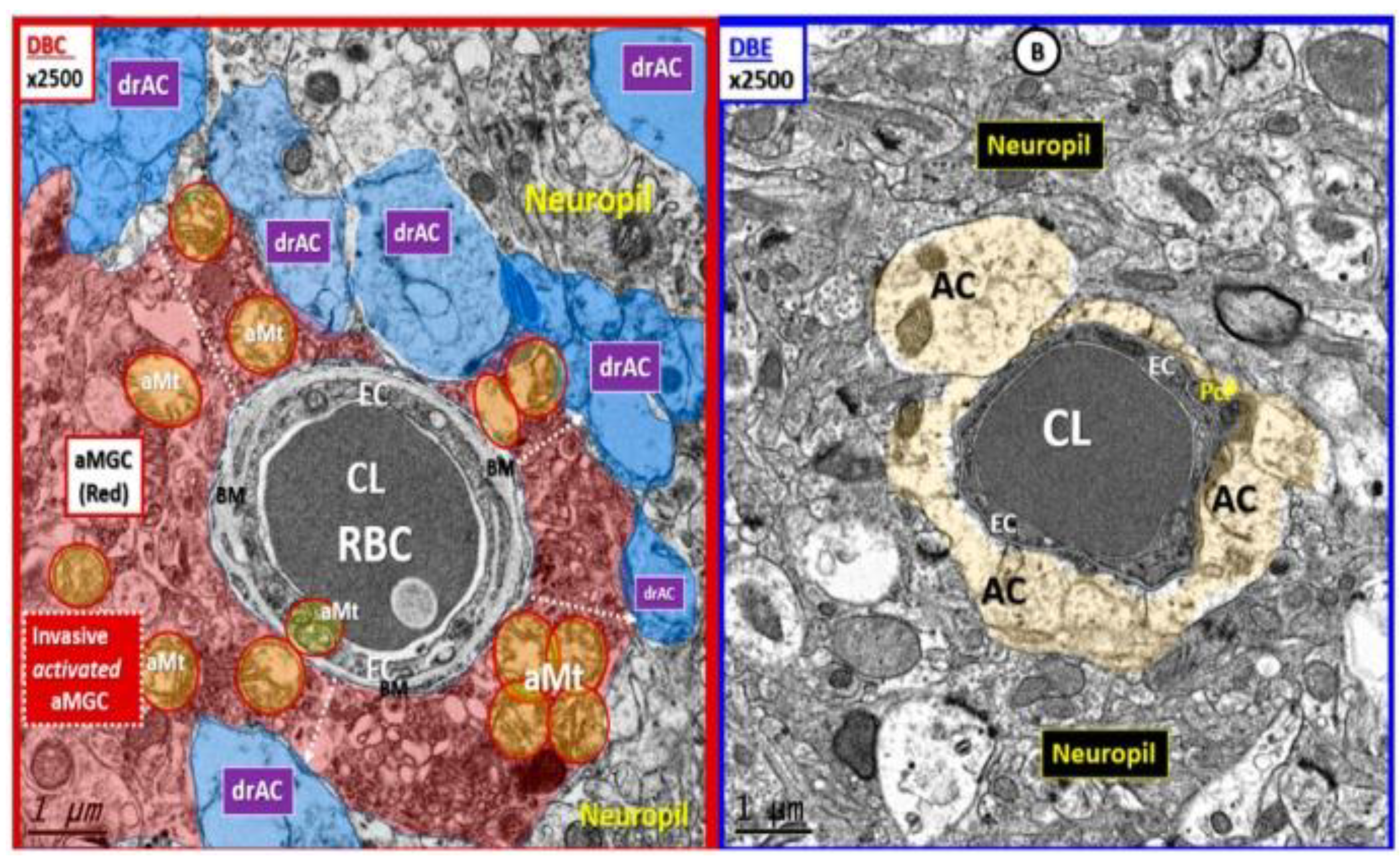
Figure 14.
Capillary neurovascular unit (NVU) and precapillary arterioles as compared to postcapillary venules with enlarged perivascular spaces (EPVSs). (A) demonstrates a capillary NVU with its astrocyte endfeet (ACef) (pseudo-colored yellow) tightly abutting the shared basement membrane of the brain endothelial cell and pericyte (asterisk and closed arrows) without a pia mater membrane. (B) demonstrates a precapillary arteriole with only a nanometer thinned perivascular space (PVS) (pseudo-colored green). Note that the abluminal perivascular space is still ensheathed by the pia matter and basal lamina of the adjacent ACef. (C, D) each depict two different postcapillary venules with enlarged perivascular spaces (EPVS) between 2 and 6.5 μm (double red arrows). Image provided with permission by CC 4.0 [35]. Magnification x1000, x2000, scale bars = 2 and 1 μm in Panels (C, D) respectively. AC, astrocyte; ACef, astrocyte endfeet; AQP 4, aquaporin 4; Cl, capillary lumen; DTBRob/ob, Black and Tan Brachyuric (BTBR) mice with the ob/ob mutation that are leptin deficient; EC, brain endothelial cell; Pc, pericyte; Pc N, pericyte nucleus; Pcfp, pericyte foot process—endfeet; RBC, red blood cell.
Figure 14.
Capillary neurovascular unit (NVU) and precapillary arterioles as compared to postcapillary venules with enlarged perivascular spaces (EPVSs). (A) demonstrates a capillary NVU with its astrocyte endfeet (ACef) (pseudo-colored yellow) tightly abutting the shared basement membrane of the brain endothelial cell and pericyte (asterisk and closed arrows) without a pia mater membrane. (B) demonstrates a precapillary arteriole with only a nanometer thinned perivascular space (PVS) (pseudo-colored green). Note that the abluminal perivascular space is still ensheathed by the pia matter and basal lamina of the adjacent ACef. (C, D) each depict two different postcapillary venules with enlarged perivascular spaces (EPVS) between 2 and 6.5 μm (double red arrows). Image provided with permission by CC 4.0 [35]. Magnification x1000, x2000, scale bars = 2 and 1 μm in Panels (C, D) respectively. AC, astrocyte; ACef, astrocyte endfeet; AQP 4, aquaporin 4; Cl, capillary lumen; DTBRob/ob, Black and Tan Brachyuric (BTBR) mice with the ob/ob mutation that are leptin deficient; EC, brain endothelial cell; Pc, pericyte; Pc N, pericyte nucleus; Pcfp, pericyte foot process—endfeet; RBC, red blood cell.
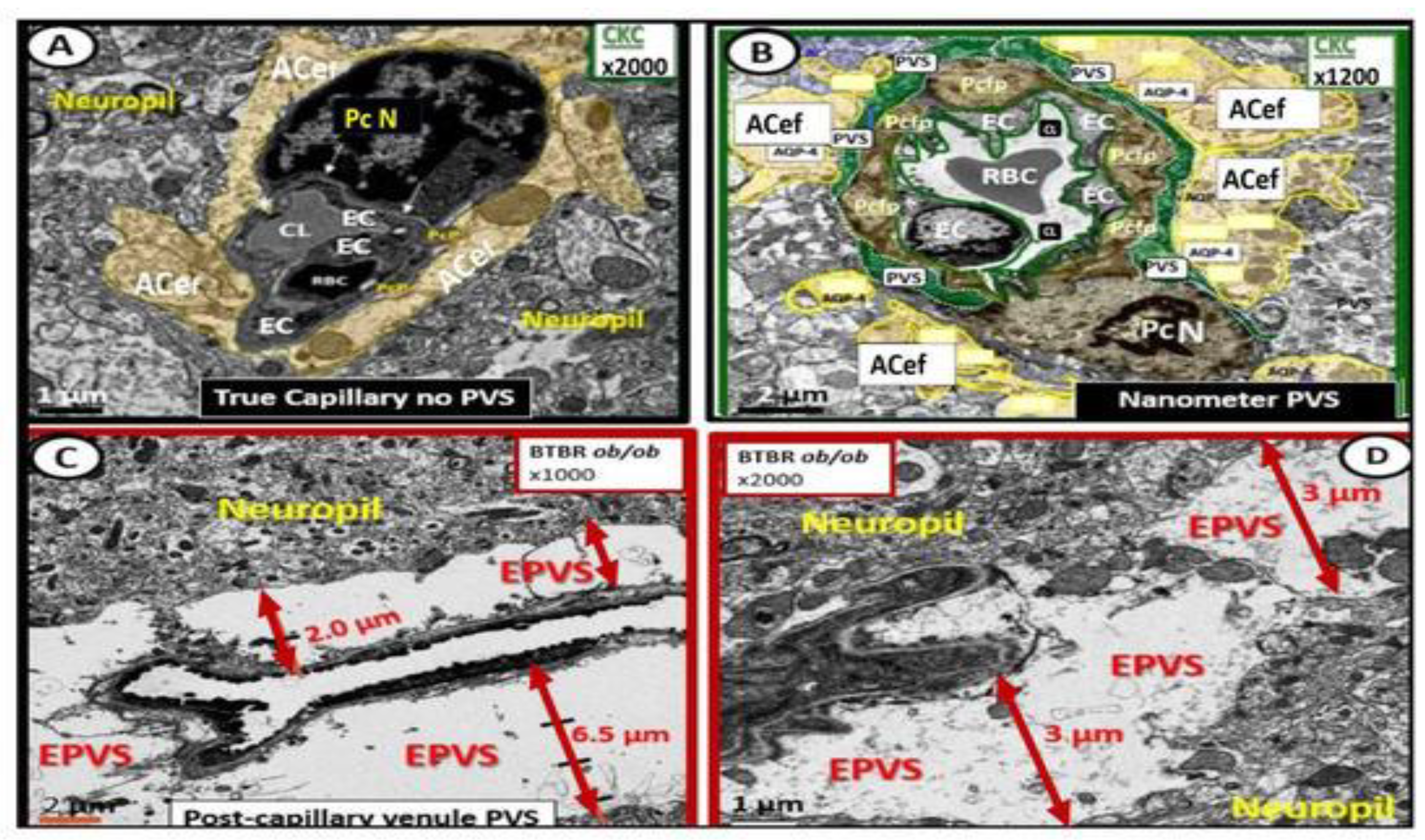
Figure 15.
Folate-Mediated One-Carbon Metabolism (FOCM). This figure illustrates both the folate and methionine interdependent cycles and supports the importance of the methyl donor S-adenosylmethionine (SAM) as well as demonstrating the importance of the essential B vitamins (B6, B9, and B12). Importantly, note that methionine and tetrahydrofolate (THF) are derived primarily through dietary intake to supply the methionine and folate cycles and that the enzyme methionine synthase (MS) and its essential cofactor vitamin B12 are placed in a central position of the interconnected folate and methionine cycles. FOCM comprises a network of interconnected folate-dependent metabolic pathways responsible for serine and glycine interconversion, de novo purine synthesis, de novo thymidylate synthesis and homocysteine remethylation to methionine as well as providing antioxidant defense via glutathione (GSH) production via the transsulfuration pathway. Note that the encircled methylenetetrahydrofolate reductase (MTHFR) enzyme plays an important role in the folate cycle. The most common genetic variant in MTHFR gene to date is the 677C > T polymorphism, which results in elevated levels of Hcy especially if there is deficient folate. Once Hcy is synthesized through multiple steps in the methionine cycle, it may then undergo remethylation to methionine to support methyl groups as a donor or be eliminated through the transsulfuration pathway. Additionally, thymidylate synthase (TYMS) converts deoxyuridine monophosphate (dUMP) to deoxythymidine monophosphate (dTMP) (not shown) in a 5,10-methylene-THF-dependent reaction. Importantly, cystathionine beta synthase (CBS) and cystathionine gamma lyase (CSE-CGL) do not only contribute to generate GSH (antioxidant) in the transsulfuration pathway but also are important for the endothelial cell generation of hydrogen sulfide (H2S), a known gaso-transmitter and vasodilator to the vessels including brain endothelial cells of the neurovascular unit (NVU) (red dashed-in insert lower left). Elevation of Hcy from the methionine cycle may result in hyperhomocysteinemia (HHCY), which is an independent risk factor for cerebro-cardiovascular diseases, accelerated atherosclerosis, thromboembolism, hypoxemia and stroke, plus neurodegenerative disease such a late-onset Alzheimer’s disease (LOAD), emerging neurodevelopmental, and neuropsychiatric diseases/disorders (boxed in red dashed lines lower left) that are associated with decreased hydrogen sulfide (H2S). Modified image provided with permission by CC 4.0 [99]. B6, pyridoxal 5′-phosphate; B12, cobalamin; CAT, cysteine aminotransferase; CBS, cystathionine-beta-synthase; CSE, cystathionine gamma lyase (CGL); GCS, glutamate cysteine ligase (gamma-glutamylcysteine synthetase); GS, glutathione synthase; GSH, glutathione; 3MST, 3-mercaptopyruvate sulfurtransferase; MTHFR, methylenetetrahydrofolate; MS, methionine synthase; SAM, S-adenosylmethionine; THF, tetrahydrofolate.
Figure 15.
Folate-Mediated One-Carbon Metabolism (FOCM). This figure illustrates both the folate and methionine interdependent cycles and supports the importance of the methyl donor S-adenosylmethionine (SAM) as well as demonstrating the importance of the essential B vitamins (B6, B9, and B12). Importantly, note that methionine and tetrahydrofolate (THF) are derived primarily through dietary intake to supply the methionine and folate cycles and that the enzyme methionine synthase (MS) and its essential cofactor vitamin B12 are placed in a central position of the interconnected folate and methionine cycles. FOCM comprises a network of interconnected folate-dependent metabolic pathways responsible for serine and glycine interconversion, de novo purine synthesis, de novo thymidylate synthesis and homocysteine remethylation to methionine as well as providing antioxidant defense via glutathione (GSH) production via the transsulfuration pathway. Note that the encircled methylenetetrahydrofolate reductase (MTHFR) enzyme plays an important role in the folate cycle. The most common genetic variant in MTHFR gene to date is the 677C > T polymorphism, which results in elevated levels of Hcy especially if there is deficient folate. Once Hcy is synthesized through multiple steps in the methionine cycle, it may then undergo remethylation to methionine to support methyl groups as a donor or be eliminated through the transsulfuration pathway. Additionally, thymidylate synthase (TYMS) converts deoxyuridine monophosphate (dUMP) to deoxythymidine monophosphate (dTMP) (not shown) in a 5,10-methylene-THF-dependent reaction. Importantly, cystathionine beta synthase (CBS) and cystathionine gamma lyase (CSE-CGL) do not only contribute to generate GSH (antioxidant) in the transsulfuration pathway but also are important for the endothelial cell generation of hydrogen sulfide (H2S), a known gaso-transmitter and vasodilator to the vessels including brain endothelial cells of the neurovascular unit (NVU) (red dashed-in insert lower left). Elevation of Hcy from the methionine cycle may result in hyperhomocysteinemia (HHCY), which is an independent risk factor for cerebro-cardiovascular diseases, accelerated atherosclerosis, thromboembolism, hypoxemia and stroke, plus neurodegenerative disease such a late-onset Alzheimer’s disease (LOAD), emerging neurodevelopmental, and neuropsychiatric diseases/disorders (boxed in red dashed lines lower left) that are associated with decreased hydrogen sulfide (H2S). Modified image provided with permission by CC 4.0 [99]. B6, pyridoxal 5′-phosphate; B12, cobalamin; CAT, cysteine aminotransferase; CBS, cystathionine-beta-synthase; CSE, cystathionine gamma lyase (CGL); GCS, glutamate cysteine ligase (gamma-glutamylcysteine synthetase); GS, glutathione synthase; GSH, glutathione; 3MST, 3-mercaptopyruvate sulfurtransferase; MTHFR, methylenetetrahydrofolate; MS, methionine synthase; SAM, S-adenosylmethionine; THF, tetrahydrofolate.
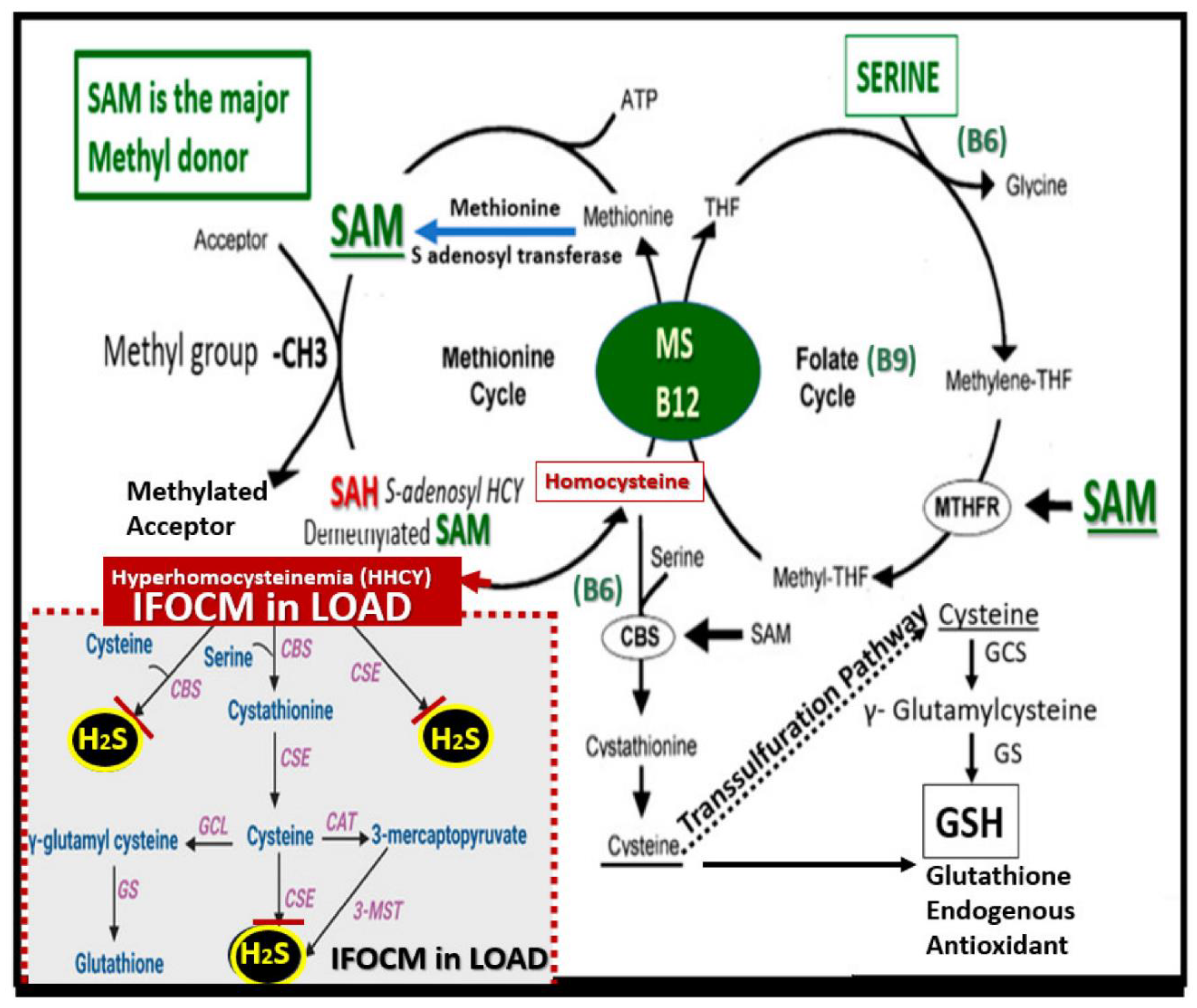
Figure 16.
Enlarged perivascular spaces (EPVS) morphologically resemble Erlenmeyer flasks at the pia precapillary arterioles/arteries and the postcapillary venules/vein’s location immediately adjacent to the centrally located capillary neurovascular unit(s) (NVU) in+ longitudinal views. Incidentally, the neck of these flasks represents the normal PVS, while the conical body represents the gradual enlargement of the PVS to form EPVS. (A) depicts the pre- and postcapillary perivascular spaces (PVS) and the enlarged perivascular spaces (EPVS) of the perivascular unit (PVU) that are morphologically shaped like Erlenmeyer flasks. Please note that the oval-shaped subarachnoid spaces (SAS) are not accurately depicted, as they are significantly larger and this illustration is not too scale. AGE, advanced glycation end-products; Asterisk, illustrates the vicious cycles between systemic inflammation and oxidative-redox stress/reactive oxygen species and brain endothelial cells that is dependent on NFkappaB; BBBdd, blood-brain barrier dysfunction/disruption; BECact/dys, brain endothelial cell activation/dysfunction; DTBRob/ob, Black and Tan Brachyuric (BTBR) mice with the ob/ob mutation that are leptin deficient; CSF, cerebrospinal fluid; GS, glymphatic system of waste removal; HCY, homocysteine; ISF, interstitial space(s); LPS, lipopolysaccharide; MMPs, matrix metalloproteinases, mtROS, mitochondrial ROS; NFkappaB, nuclear factor kappa B; pCC, peripheral cytokines/chemokines; PVS, perivascular spaces; PVU, perivascular unit; RAGE, receptor for AGE; ROS, reactive oxygen species; SAS, subarachnoid space(s); µm, micrometer.
Figure 16.
Enlarged perivascular spaces (EPVS) morphologically resemble Erlenmeyer flasks at the pia precapillary arterioles/arteries and the postcapillary venules/vein’s location immediately adjacent to the centrally located capillary neurovascular unit(s) (NVU) in+ longitudinal views. Incidentally, the neck of these flasks represents the normal PVS, while the conical body represents the gradual enlargement of the PVS to form EPVS. (A) depicts the pre- and postcapillary perivascular spaces (PVS) and the enlarged perivascular spaces (EPVS) of the perivascular unit (PVU) that are morphologically shaped like Erlenmeyer flasks. Please note that the oval-shaped subarachnoid spaces (SAS) are not accurately depicted, as they are significantly larger and this illustration is not too scale. AGE, advanced glycation end-products; Asterisk, illustrates the vicious cycles between systemic inflammation and oxidative-redox stress/reactive oxygen species and brain endothelial cells that is dependent on NFkappaB; BBBdd, blood-brain barrier dysfunction/disruption; BECact/dys, brain endothelial cell activation/dysfunction; DTBRob/ob, Black and Tan Brachyuric (BTBR) mice with the ob/ob mutation that are leptin deficient; CSF, cerebrospinal fluid; GS, glymphatic system of waste removal; HCY, homocysteine; ISF, interstitial space(s); LPS, lipopolysaccharide; MMPs, matrix metalloproteinases, mtROS, mitochondrial ROS; NFkappaB, nuclear factor kappa B; pCC, peripheral cytokines/chemokines; PVS, perivascular spaces; PVU, perivascular unit; RAGE, receptor for AGE; ROS, reactive oxygen species; SAS, subarachnoid space(s); µm, micrometer.

Box 1.
Comparison of cerebral small vessel disease (SVD) as identified by magnetic resonance image(s) (MRI). Incidentally, cerebral amyloid arteriopathy (CAA) is considered a SVD (not included). Also, small recent cortical infarcts are considered to be another SVD but occur more superficially in cortical regions as compared to deeper lacunes (not included). Modified box image provided with permission by CC 4.O [3]. GLY Dys, glymphatic dysfunction; mm, millimeter; MRI, magnetic resonance image(s); T2*, T2-weighted gradient recall echo magnetic resonance images.
Box 1.
Comparison of cerebral small vessel disease (SVD) as identified by magnetic resonance image(s) (MRI). Incidentally, cerebral amyloid arteriopathy (CAA) is considered a SVD (not included). Also, small recent cortical infarcts are considered to be another SVD but occur more superficially in cortical regions as compared to deeper lacunes (not included). Modified box image provided with permission by CC 4.O [3]. GLY Dys, glymphatic dysfunction; mm, millimeter; MRI, magnetic resonance image(s); T2*, T2-weighted gradient recall echo magnetic resonance images.
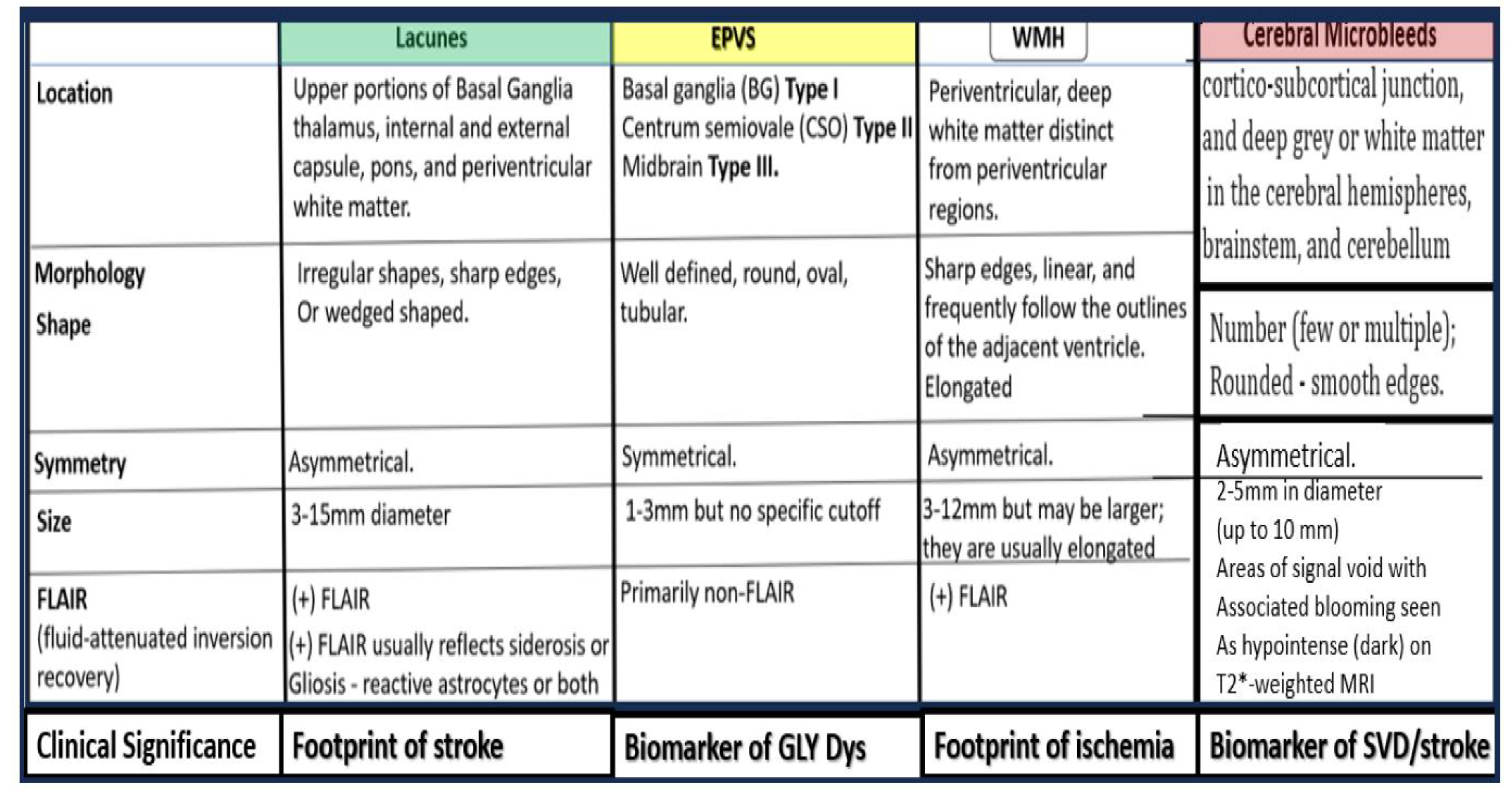
Box 2.
Why are perivascular spaces (PVS) important? box image provided with permission by CC 4.O [3]. GLY Dys, glymphatic dysfunction; mm, millimeter; MRI, magnetic resonance image(s); T2*, T2-weighted gradient recall echo magnetic resonance images.
Box 2.
Why are perivascular spaces (PVS) important? box image provided with permission by CC 4.O [3]. GLY Dys, glymphatic dysfunction; mm, millimeter; MRI, magnetic resonance image(s); T2*, T2-weighted gradient recall echo magnetic resonance images.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



