
Submitted:
21 October 2025
Posted:
24 October 2025
You are already at the latest version
Abstract
The development of advanced biomaterials for corneal applications requires robust trans-lational platforms that faithfully replicate human characteristics. Porcine corneas are in-creasingly recognized for ophthalmic research. Their unique combination of anatomical similarity, biomechanical comparability, and accessibility make them highly suitable for preclinical evaluation of innovative therapies, bridging the gap between preclinical dis-covery and clinical application. This review outlines the utility of porcine corneal models in validating advanced biomaterials, particularly in ex vivo settings, focusing on current methodologies, while addressing challenges and future directions. We aim to underscore the potential of porcine corneal models to accelerate the translation of next-generation biomaterials into clinically relevant corneal therapies.
Keywords:
porcine corneal model
; ex vivo model
; translational platform
; ophthalmic research
; cell viability
; drug permeability
1. Anatomy of the Human Eye
The visual system is a highly specialized sensory network responsible for acquiring and processing visual information from the environment. Light first enters the eye through the cornea, then passes through successive ocular media until it reaches the retina, where the photoreceptors convert it into neural signals. These signals are transmitted via complex neural pathways to the brain, where they are decoded into coherent visual perception [1].
The human eyeball is not a perfect sphere; instead, it has an oblate spheroid shape [2]. It contains three chambers: the anterior, posterior, and vitreous chambers. The anterior chamber, between the cornea and the iris, holds about 220 μL of aqueous humor and averages 3.11 mm in depth. The posterior chamber, located between the iris and lens, contains around 60 μL of aqueous humor [2]. These chambers are separated through the pupil, and the aqueous humor is produced by the ciliary body and nourishes the cornea and lens [1]. The vitreous chamber, the largest compartment, lies behind the lens adjacent to the retina and contains 5 to 6 mL of gel-like vitreous humor. The total volume of the adult eye is approximately 6.5–7.0 mL [2] (Table 1).
Together with these internal chambers, the eye is also structurally organized into three concentric layers. The outer fibrous layer, consisting of the cornea and sclera, provides protection and maintains the shape of the globe. The transparent cornea lets light enter and contributes to most of the eye’s refractive power, while the opaque sclera offers structural support. The middle vascular layer includes the iris, ciliary body, and choroid. The iris regulates light entry through the pupil. The ciliary body produces aqueous humor and controls lens accommodation. The choroid, located posteriorly, is rich in blood vessels and supplies nutrients to the outer retina. The inner neural layer - the retina - contains photoreceptors and neural elements that convert light into electrical signals. These signals travel through the retina and exit the eye via the optic nerve, ultimately reaching the brain for visual processing [1,2].
The proper functioning of the eye relies on the close integration of its anatomical structures. Each layer contributes to a shared goal, which is visual clarity, and the loss of integrity in any of these components can significantly impair vision. In addition to their optical roles, structures such as the corneal epithelium and the tear film act as selective barriers, regulating molecular transport and protecting against external pathogenic agents. These barriers are particularly important in the context of ocular drug delivery, as they influence the ability of therapeutic agents to reach specific target tissues, such as the cornea.
1.1. Cornea
The cornea is the principal refracting component of the eye (43.25 diopters (D) out of a total of 58.60 D) [2,3]. Its transparency and avascularity provide optimal light transmittance.
From anterior to posterior, the five layers which compose the cornea are the epithelium, the Bowman’s layer, the stroma, the Descemet’s membrane, and the endothelium (Figure 1).
The corneal epithelium is a lipophilic, nonkeratinized, stratified squamous epithelium consisting of 4–6 cell layers and typically 40–50 μm thick [2] (Figure 2).
The superficial layer, which is 1 to 2 cells thick, consists of flattened squamous cells with tight junctions between these cells that form a crucial barrier that prevents intercellular passage of substances and controls fluid uptake from the tear film, creating a selective semipermeable membrane. Desmosomes provide additional adhesion, reinforcing this barrier. The middle layer consists of 2 to 3 layers of wing cells, interconnected by desmosomes and gap junctions. The basal layer is a single layer of columnar cells attached to the basement membrane through hemidesmosomes. These basal cells secrete the basement membrane and serve as the germinal layer where mitosis occurs, renewing the epithelium. Epithelial renewal involves the upward migration and differentiation of basal cells into wing cells and then superficial cells, with a full turnover approximately every 7 days. Minor epithelial injuries heal rapidly within hours to overnight, but damage to the basement membrane can require several months for full repair.
The Bowman’s layer is the second layer of the cornea, measuring approximately 8 to 14 μm in thickness. It consists of a dense, acellular sheet of randomly arranged collagen fibrils (20–25 nm in diameter), serving as a transition zone between the epithelium and the stroma. Although its exact function remains unclear, it is believed to contribute to the biomechanical rigidity and shape of the cornea. It does not regenerate after injury. Despite this, it is highly resistant to shearing, penetration, and infection. Its removal, such as during photorefractive keratoplasty, has not been associated with significant long-term visual effects.
The corneal stroma, measuring 470–500 μm, accounts for approximately 90% of the total corneal thickness [2]. It is primarily composed of collagen fibrils, keratocytes, and extracellular ground substance. Collagen fibrils, mainly type I with some type V, have a uniform diameter (25–35 nm) and are arranged in regularly spaced, parallel lamellae (about 200–300 in total) extending from limbus to limbus. This precise organization is essential to maintain corneal transparency [3].
The Descemet’s membrane is continuously secreted by endothelial cells and gradually thickens with age, measuring about 3–4 μm at birth, and reaching 10–12 μm in adulthood. This layer is particularly rich in type IV collagen.
The corneal endothelium is a single layer of flattened, typically hexagonal cells located adjacent to the anterior chamber, with a thickness of 4–6 μm. This hexagonal arrangement, known as the endothelial mosaic, maximizes surface coverage and structural efficiency. Endothelial cells do not proliferate significantly in humans, so their number gradually declines with age (from 3000–4000 cells/mm2 in children to 1000–2000 cells/mm2 by the age of 80) due to cell loss and disintegration [2,3].
1.2. Ocular Barriers
The presence of ocular barriers is the key challenge for therapeutics in terms of reaching the target site and remaining there for a sufficient duration.
The tear film is a thin, transparent layer that plays a crucial role in ocular surface protection and drug absorption. It consists of three layers: an outer lipid layer, a central aqueous layer, and an inner mucin layer (Figure 3).
The tear film constitutes a dynamic barrier to drug administration, not only due to its multilayered composition but also because of continuous tear turnover and nasolacrimal drainage. Each layer contributes to the tear film’s barrier function: the lipid layer primarily prevents tear evaporation and forms a barrier to hydrophilic drugs, the aqueous layer removes hydrophobic drugs, the mucin layer, consisting of negatively charged macromolecules, influences drug absorption through electrostatic interactions, either attracting or repelling drug molecules. Following topical drug administration, tear turnover increases significantly, promoting rapid clearance of the drug via nasolacrimal drainage often within one to two minutes. Due to the limited ocular surface area most of the solution (typically ~30 µL) is quickly lost. It is estimated that around 60% of the drug is eliminated within the first 2 min. After 8 min, the drug concentration on the ocular surface may drop to 0.1%, and, after 15 to 25 min, nearly all active components are cleared [4]. This dynamic clearance substantially reduces drug bioavailability and limits therapeutic efficacy in ocular treatments.
The cornea, located directly beneath the tear film, constitutes a significant anatomical barrier to the penetration of topically applied drugs into intraocular structures. As the outermost corneal layer, the epithelium is the first to interface with exogenous substances and represents one of the most substantial obstacles to drug absorption. It consists of 4 to 6 layers of tightly packed epithelial cells interconnected by various intercellular junctions: tight junctions, adherens junctions, desmosomes, and gap junctions. Among these, tight junctions, which are located predominantly in the superficial epithelial layer, are particularly critical to maintain the structural and functional integrity of the corneal barrier. They are composed of transmembrane proteins, such as occludin and members of the claudin family, which are anchored intracellularly by scaffold proteins like zonula occludens-1 (ZO-1) [5] (Figure 4). This complex and dynamic architecture plays a pivotal role in preserving epithelial homeostasis and protecting the underlying ocular tissues. As a result, the presence of tight junctions constitutes a major challenge to effective topical delivery, often resulting in limited bioavailability.
2. Experimental Models in Corneal Research
The study of the cornea and its pathologies depend on reliable experimental models which enable the investigation of physiological mechanisms, the testing of novel therapeutic formulations, and the assessment of the safety and efficacy of biomaterials or ophthalmic drugs before clinical application. Due to the complex and stratified nature of the cornea the choice of an appropriate experimental model is essential to ensure relevant and translational results. In vision research, in vitro, ex vivo, and in vivo approaches are all employed, with animal models playing a central role in each one of these approaches [6].
2.1. In Vitro, Ex Vivo, In Vivo Corneal Models
Over the past decades, various in vitro, ex vivo, and in vivo models have been developed to reproduce, in a controlled manner, the physiological conditions of the human cornea.
In vitro corneal models are based on cultured cells or reconstructed tissues that provide a controlled environment to study cellular behavior, molecular mechanisms, and tissue responses. These models allow for precise manipulation of experimental conditions and are highly reproducible, making them valuable tools for investigating corneal physiology, drug permeability, toxicity, and wound healing processes [7,8,9]. Recently, advances in 3D cultures and corneal organoids have significantly improved their physiological relevance [10].
Corneal cell monolayer models represent one of the simplest and most widely used in vitro approaches. In these models, a single layer of corneal epithelial, stromal, or endothelial cells is cultured on a suitable substrate, allowing researchers to analyze cellular behavior under controlled conditions [11,12]. A study by Travers et al. [13] investigated the role of molecular coatings in human corneal endothelial cell monolayers. Zhang et al. [7] utilized monolayer cultures of corneal epithelial cells to study the effects of the ZEB1 gene on cell migration, demonstrating that ZEB1 plays a crucial role in promoting epithelial cell migration, which is vital for corneal wound healing.
3D corneal models and organoids have been developed to better recapitulate the structural and functional complexity of the human cornea. These models involve single or multiple corneal cell types arranged in three-dimensional architecture, often using scaffolds, hydrogels, or bioprinting techniques to mimic the natural extracellular matrix [10]. Organotypic cultures consisting of epithelial cells have been developed to study the effects of topical drugs on the corneal epithelium. In these models, primary epithelial cells are cultured on membrane inserts and exposed to an air–liquid interface to mimic physiological conditions. The reconstructed human corneal epithelial model (EpiOcular™, MatTek Life Sciences) consists of human corneal epithelial cells grown in 3D cultures forming a stratified squamous epithelium [9]. Da Silva et al. [8] used this model as a screening tool for assessment of eye toxicity potential of isolated chemicals and botanical mixtures. These 3D and organoid models provide several advantages over monolayers, including more physiologically relevant cell–cell and cell–matrix interactions, the preservation of tissue stratification, and the ability to simulate wound healing, drug penetration and disease more accurately [10]. However, they are more technically demanding and less reproducible than simple monolayer cultures, and standardization across laboratories remains a challenge [14].
Ex vivo corneal models serve as an essential intermediate between simplified in vitro systems and complex in vivo studies. These models make use of intact corneal tissues obtained from human donors, animals, or abattoir sources that preserve the natural architecture, cellular stratification, and biochemical composition of the cornea. By maintaining the physiological organization of corneal layers, ex vivo models allow for the assessment of drug permeability, wound healing, and tissue responses under conditions that closely mimic the in vivo environment. Several recent studies have explored a wide range of ex vivo platforms for ophthalmic research [15,16].
Barbalho et al. [17] developed a dynamic ex vivo porcine eye model with simulated tear flow to evaluate the penetration of ophthalmic drug formulations, providing more realistic pharmacokinetic data. Pescina et al. [18] developed an ex vivo porcine corneal model to evaluate the transcorneal permeation of drugs, combining histological assessment and permeability testing to better understand how physicochemical properties (hydrophilicity of the compounds) influence corneal transport. Shi et al. [19] utilized an ex vivo porcine cornea model to study Acanthamoeba infections, demonstrating its applicability to infectious disease research. Okurowska et al. [20] established a porcine ex vivo cornea model to investigate antimicrobial therapies against Pseudomonas aeruginosa-induced bacterial keratitis, providing a reproducible system for assessing treatment efficacy under physiologically relevant conditions. Tsai et al. [21] established a human corneal endothelium wound model using donor corneas maintained in an anterior chamber culture system, allowing for continuous perfusion and precise monitoring of corneal thickness. This model enables the evaluation of endothelial wound healing and the effects of cell-based or pharmacological therapies. Similarly, Castro et al. [22], investigated stromal and epithelial regeneration by culturing porcine corneas after anterior keratectomy, providing valuable insight into fibrosis and myofibroblast activation mechanisms. Netto et al. [23] incubated porcine corneas at low humidity to reproduce the inflammatory and structural changes characteristic of dry eye disease, demonstrating the potential of ex vivo platforms for studying ocular surface disorders. In another study, Rouhbakhshzaeri et al. [24] designed a new ex vivo porcine model to mimic endothelial injury caused by phacoemulsification and tested the therapeutic potential of mesenchymal stromal cell secretome, highlighting its regenerative and protective effects on the corneal endothelium. Moreover, Rodrigues da Penha et al. [25] developed a novel ex vivo model using bovine corneas for chemical ocular toxicity assessment, successfully distinguishing irritant compounds and confirming its applicability for cosmetic product safety testing. Sarfraz et al. [26] employed ex vivo porcine eyes to assess the performance of a twin nanoparticulate system designed to enhance dexamethasone absorption and prolong ocular retention, confirming the relevance of ex vivo approaches in preclinical screening of advanced drug delivery systems.
Ex vivo corneal models present several advantages that make them valuable tools in ophthalmic research. They preserve the native architecture of the cornea, maintaining realistic barrier properties and tissue responses. This structural integrity enables the evaluation of drug permeability, wound healing, infection dynamics, and tissue toxicity under controlled but physiologically relevant conditions. Furthermore, ex vivo models, particularly those using porcine or bovine corneas from abattoir sources, are cost-effective and ethically preferable alternatives to animal testing [16]. However, despite their translational relevance, these systems also present significant limitations. Tissue viability decreases over time due to the absence of systemic circulation, limiting the duration of experiments to a few days [27]. Variability among donor tissues and difficulties in reproducing immune or vascular responses further constrain their predictive power for long-term or systemic effects [20,28]. Thus, while ex vivo corneal models bridge the gap between in vitro and in vivo experimentation, they are best suited for short-term studies of drug permeability, toxicity, or early-stage therapeutic screening.
In vivo corneal models use live animals to investigate corneal physiology, disease mechanisms, therapeutic efficacy, and safety in a complete biological context. These models are critical for assessing integrated responses which ex vivo or in vitro systems cannot fully recapitulate, including immune response, vascularization and systemic pharmacokinetics. The primary purpose of an animal model is not to replicate human conditions in every detail, but rather to reproduce the specific aspects that are critical for understanding a disease or testing an intervention. The vertebrate species used can include rodents, rabbits, pigs, primates, felines, and canines, while invertebrates can include flies and nematodes. The selection of an appropriate model depends on the research question, as differences in ocular anatomy, physiology, and drug response can significantly affect experimental outcomes. For instance, variations in corneal thickness, eye axial length, and ocular surface properties between species can influence drug absorption, distribution, and overall therapeutic efficacy (Table 2). Understanding these interspecies differences is therefore essential for interpreting experimental results and translating findings to human applications [6].
In vivo corneal models have been widely used to investigate therapeutic delivery, drug bioavailability, and molecular mechanisms of wound healing. For instance, Peterson et al. [34] employed a rabbit corneal suture model to evaluate the sustained delivery of bevacizumab using biodegradable densomere microparticles, illustrating how in vivo models can be applied to assess long-term drug administration strategies. Chauchat et al. [35] utilized pigmented rabbits to compare the ocular bioavailability of different latanoprost formulations, demonstrating the value of in vivo systems to study pharmacokinetics and the influence of formulation excipients. Similarly, Chen et al. [36] used a C57BL/6 mouse corneal model to explore epithelial wound healing and the role of Interleukin-36 receptor signaling, highlighting the application of in vivo models for investigating molecular pathways and tissue repair processes. Akpek et al. [37] used a rabbit model to evaluate the integration and functional performance of a second-generation synthetic cornea, demonstrating how in vivo systems can assess biocompatibility and tissue integration under physiological conditions. Similarly, Yamashita et al. [38] employed rabbit models to study corneal endothelial dysfunction, providing insights into disease mechanisms and enabling the preclinical evaluation of potential therapeutic strategies. Additionally, Sun et al. [39] used in vivo models for studying specific corneal pathologies, such as Fuchs’ endothelial corneal dystrophy, allowing researchers to investigate disease progression and test interventions in a whole-organism context. Koseoglu et al. [40] used advanced vivo imaging techniques, combined with deep learning to diagnose neuropathic corneal pain, highlighting the role of in vivo models in developing and validating novel diagnostic tools. These examples emphasize the versatility of in vivo corneal models in preclinical ophthalmic research, from drug delivery and pharmacokinetics to mechanistic studies of corneal physiology and pathology in a biologically relevant environment.
Despite their relevance to translational research, in vivo corneal models present several limitations. Anatomical and physiological differences between animal models and the human eye can affect the extrapolation of findings, particularly regarding corneal thickness and endothelial density [6]. Ethical concerns and regulatory restrictions also limit the use of live animals, especially in large-scale or long-term studies [41]. Variability in individual animal responses can also increase data heterogeneity, which leads to the need for larger sample sizes to achieve statistical significance. Furthermore, in vivo experiments are costly, time-consuming, and require specialized facilities and expertise, which may constrain experimental design and reproducibility [42].
2.2. Porcine Corneal Models
The porcine eye is widely used as a model in vision science research due to its anatomical similarity to the human eye [32]. It shares several anatomical features with the human eye, such as overall size, general morphology, retinal vasculature, and the absence of a tapetum lucidum (a reflective layer of the retina), where retinal pigment epithelium (RPE) cells are not pigmented (unlike other farm animals like cows and sheep) [33]. It also contains cone photoreceptors in the outer retina [43]. A compilation of more specific parameters that have been compared among porcine and human eyes can be consulted in Table 3.
Porcine eyes can be employed both as in vivo and ex vivo models; however, they are mostly used in ex vivo corneal research due to practical and ethical considerations. The use of live pigs in ophthalmic studies presents several challenges, including their rapid growth rate, which complicates long-term experimental designs, and the limited orbital space, which makes surgical procedures involving extraocular devices difficult. Additionally, housing, handling, and maintenance costs further restrict the feasibility of in vivo porcine studies. For these reasons, ex vivo porcine corneal models have become a preferred alternative, offering substantial economic and logistical advantages over live animal experimentation [6]. Another important reason is that ex vivo porcine eyes are readily available as by-products of the food industry, thereby substantially reducing both the economic cost and ethical concerns associated with animal use. As they are collected post-mortem from slaughterhouses, their use does not involve the deliberate sacrifice of animals for research purposes. This makes the porcine model particularly suitable for ex vivo studies, offering a cost-effective, accessible, and ethically responsible alternative [43].
Porcine eye models have been applied in various contexts, including studies of the neurosensory retina, cataract surgery research, corneal transplantation, aberrometry, and transscleral drug delivery [32,43,46]. In particular, ex vivo porcine corneal models allow for the evaluation of the safety and potential toxicity of chemical and pharmaceutical compounds before proceeding to in vivo or clinical testing [47]. Due to their anatomical similarity to the human eye and wide availability, they also represent a valuable platform for surgical training. Indeed, porcine eyes are routinely used by ophthalmology residents to practice and refine surgical techniques, and many corneal procedures and innovations have been first developed and optimized using ex vivo porcine ocular tissues [48,49].
Despite their numerous advantages, ex vivo porcine corneal models also present certain limitations that must be considered when interpreting results. One of the main drawbacks is the absence of physiological factors such as blood flow, tear film dynamics, and immune responses, which can significantly influence drug absorption, metabolism, and tissue repair in vivo. Furthermore, post-mortem tissue degradation and variability in the time between eye collection and experimentation can affect corneal integrity and cellular viability, potentially altering results. The lack of standardized collection and preservation protocols across studies further contributes to experimental variability. Nevertheless, porcine models remain highly valuable translational tools to bridge the gap between in vitro experimentation and human ophthalmic applications.
3. Current Methodologies
3.1. Cell Viability
Assessing cell viability is a crucial step in validating and supporting ocular therapy research. The evaluation of cell viability provides valuable insights into tissue health following experimental manipulation, drug exposure, surgical procedures, or storage conditions [50,51]. It also serves as a key indicator of safety and biocompatibility before advancing to in vivo studies or clinical trials. Therefore, assessing the cytotoxic potential of formulations, biomaterials, or surgical interventions in ex vivo porcine corneas yields essential preclinical data that contributes to the refinement of ocular therapies under physiologically relevant yet ethically sustainable conditions. In ex vivo porcine corneas, viability is commonly assessed in specific layers depending on the research purpose. Epithelial, endothelial and keratocyte viability is typically evaluated to examine the effects of formulations under research, toxic or mechanical injuries.
A wide range of studies has been applied to quantify and visualize viable and non-viable cells in porcine ex vivo models. To evaluate corneal epithelial cell viability, the most used approach involves vital dye staining with trypan blue. Following the experimental procedure, the whole cornea is immersed in a 0.4% trypan blue solution for approximately 2 min, with the epithelial surface facing downward. The tissue is then gently rinsed in phosphate-buffered saline (PBS) to remove excess dye. Subsequently, the cornea is carefully dissected to isolate it, ensuring minimal mechanical damage (Figure 5). Microscopic examination is performed to quantify the number of trypan blue–stained cells, which correspond to non-viable epithelial cells [52,53]. This method provides a rapid and reliable estimation of epithelial integrity after topical drug exposure or mechanical stress.
To assess endothelial cell viability in ex vivo porcine corneal models, several vital staining methods have been applied, most commonly using trypan blue and/or alizarin red S. In one study, the endothelium was exposed to 0.2% trypan blue, and the proportion of blue-stained non-viable cells was quantified relative to the total endothelial surface [54]. Another study employed alizarin red S to evaluate endothelial morphology and detect alterations in cell membranes [55]. In a different approach, both trypan blue and alizarin red S were combined to enhance the visualization of cell borders and improve the assessment of endothelial morphology and viability [50] (Figure 6). Alternatively, fluorescence-based Live/Dead assays using calcein-acetoxymethyl ester (AM) and ethidium homodimer-I was used to simultaneously identify viable and non-viable cells [56]. Collectively, these techniques provide reproducible information on endothelial integrity, which is a critical parameter to validate the safety of ocular therapies.
To evaluate keratocyte viability in an ex vivo porcine corneal model, keratocytes were labelled with the fluorescent probe 5-chloromethylfluorescein diacetate (CMFDA) and visualized by confocal laser scanning microscopy, allowing for high-resolution imaging of viable stromal cells and assessment of tissue integrity [57]. This approach provided detailed morphological information.
3.2. Drug Permeability
The evaluation of corneal drug permeability is a crucial step in the preclinical development of ophthalmic formulations, as it directly influences the therapeutic efficacy of ocular treatments. Porcine eyes are one of the most suitable models for drug diffusion and permeability studies because of their close resemblance to human eyes in terms of globe size, vascular anatomy, histological features, physiological properties, and stromal collagen bundle organization. The porcine cornea presents a comparable overall thickness and an identical endothelial thickness to that of humans. However, notable structural differences exist: the porcine corneal epithelium and sclera are roughly twice as thick as those in humans, and the corneal stroma is about 30% thicker [58]. These variations must be carefully considered when interpreting results from permeation experiments.
Franz diffusion cells represent the most widely used setup for ex vivo corneal permeability studies. A conventional Franz diffusion system is composed of two compartments: a donor and a receptor chamber separated by a membrane, which in this case corresponds to the porcine cornea with the epithelial surface facing the donor chamber to simulate topical drug application (Figure 7). The donor chamber receives the formulation under investigation, while the receptor chamber, connected to a sampling port, allows periodic collection of samples for quantitative analysis. The receptor compartment is typically filled with a physiologically relevant buffer solution, such as PBS or simulated tear fluid, maintained at 37 °C and continuously stirred with a magnetic stirrer to ensure homogeneity. At predefined intervals, aliquots are collected from the receptor chamber, and the amount of permeated drug is quantified using analytical methods such as high-performance liquid chromatography (HPLC) or ultraviolet–visible (UV–Vis) spectrophotometry [58].
Several studies have employed Franz diffusion cells with ex vivo porcine corneas to investigate drug permeability across ocular tissues. For instance, this model has been used to assess the permeation of erythropoietin, desmoteplase, and carprofen, as well as to evaluate novel ophthalmic formulations such as ciprofloxacin polymeric films and riboflavin-5′-phosphate enhanced with vitamin E D-α-tocopheryl polyethylene glycol succinate (TPGS) [59,60,61,62,63]. These examples highlight the versatility of the Franz diffusion cell setup for studying transcorneal drug diffusion and comparing the permeability profiles of different compounds and formulations. Despite their widespread use in corneal permeability studies, Franz diffusion cells present several limitations. The system only supports vertical diffusion and fails to reproduce the natural curvature and physiological environment of the cornea. Also, unintentional corneal damage can be generated in this model. Furthermore, prolonged exposure of the endothelium to the receptor medium may also cause stromal swelling due to excessive hydration, further affecting the accuracy of permeability measurements [64].
Whole-eye models use intact porcine eyes to study corneal drug permeability under more physiologically relevant conditions than isolated corneal setups such as Franz diffusion cells. By preserving the natural curvature and anatomical relationships of the eye, these models allow for drug penetration to be assessed in a context that closely mimics in vivo conditions. Drug formulations are applied to the corneal surface, and samples are collected from the aqueous humor or other compartments to determine penetration. Although the basic setup is the same, experimental protocols can be adapted depending on the drug, the detection method or the specific goals of the study. For instance, Barbalho et al. [65] developed a dynamic model with simulated lacrimal flow to evaluate the performance of pharmaceutical drug products. Similar, Bhujbal et al. [66] assessed the ocular drug penetration using ex vivo porcine whole-eye model with simulated tear flow. Sun et al. [67] investigated the permeation dynamics of organosilica nanoparticles across porcine corneal barriers for glaucoma drug delivery. Sarfraz et al. [68] also carried out an ex vivo whole eye permeation study. While these models provide more realistic diffusion dynamics, they are limited by variability between donor eyes, tissue viability over time, and the absence of systemic circulation.
A novel ex vivo corneal permeability model has been developed using CellCrown™ inserts in a 96 well plate format, enabling high-throughput testing while reducing the number of porcine eyes required [69]. 5 mm corneal punches are placed into the base of the outer insert with the epithelium facing upwards. The inner insert is positioned on top of the corneal disc to create a water-tight seal without deforming the tissue. The assembled inserts are placed into the well of a 96 well plate containing PBS, with parafilm or a similar cover used to prevent evaporation. To assess the permeability the study substance was pipetted into the inner insert (Figure 8). Despite allowing for high throughput testing, this ex vivo model has several limitations. The use of small corneal punches fails to reproduce the natural curvature of the cornea. Variability between porcine eyes and challenges in maintaining tissue viability can also impact reproducibility.
Even though corneal permeability has been extensively investigated in numerous studies, the underlying causes of increased permeability are often not explored in detail. Understanding why permeability changes occur is essential for elucidating the molecular mechanisms that take place in the ocular surface following the application of ophthalmic therapies. Ophthalmic therapies, such as drug formulations or contact lenses may alter corneal barrier properties, but without studying the specific pathways involved, it remains unclear whether these effects are transient, reversible, or associated with epithelial damage. Therefore, identifying the cellular and molecular basis behind changes in permeability is a critical step toward developing safer ocular treatments.
Tight junctions play a key role in maintaining the structural and functional integrity of the corneal barriers. Tight junctions are composed of transmembrane proteins, such as occludin and members of the claudin family, which are anchored intracellularly by scaffold proteins, such as ZO-1 [5]. Theoretically, an increase in corneal permeability is directly related to a loss of tight junction integrity and, consequently, disruption of the epithelial layer [70]. Assessing tight junctions’ integrity thus provides a mechanistic link between observed permeability changes and cellular barrier function. Some studies have investigated tight junction alterations using immunofluorescence labelling in ex vivo rabbit and mouse corneal models [71,72,73,74]. However, no equivalent data currently exists for porcine models. Considering the structural and physiological similarities between porcine and human corneas, performing such analyses in the porcine model would provide valuable insights into drug-induced barrier modulation and enhance translational relevance in preclinical testing.
4. Critical Evaluation and Future Perspectives
Porcine ex vivo corneal models represent a highly promising translational platform for preclinical ophthalmic research. Their anatomical and physiological similarity to human corneas, combined with their wide availability from abattoir-derived tissues, provides an ethically advantageous, cost-effective, and reproducible alternative to traditional in vivo models. Despite these advantages, research using porcine ex vivo corneas remain relatively limited, and methodological gaps persist. In particular, standardized protocols to assess key corneal features, such as tight junction integrity, are scarce. Critical proteins like ZO-1 and occludin, which play essential roles in epithelial barrier function, have not been evaluated in porcine ex vivo corneas, representing a notable knowledge gap. We identified this critical gap and successfully developed a protocol to address it. We are currently preparing a manuscript presenting this protocol, along with additional methodologies using porcine ex vivo corneas, to facilitate broader adoption and standardization in future studies.
Looking forward, porcine ex vivo corneas represent an exceptionally valuable translational model that, due to their structural and physiological similarity to human eyes, versatility, and wide availability, should be more widely utilized in preclinical ophthalmic research. This model allows for the integration of diverse techniques, including permeability assays, cell viability assessments of epithelium, endothelium, and keratocytes, as well as evaluations of tight junction integrity. Standardizing these methodologies can greatly enhance reproducibility across laboratories and facilitate broader adoption. By capitalizing on these inherent advantages, researchers can not only bridge existing knowledge gaps but also accelerate the development of safe and effective ocular therapies, while minimizing reliance on live animals. Given these unique features, we strongly advocate for increased use of porcine ex vivo corneas as a robust, practical, and translationally relevant platform in future ophthalmic studies.
Author Contributions
Conceptualization, P.N. and E.J.C.; methodology, P.N. and E.J.C.; validation, P.N. and E.J.C.; investigation, P.N.; resources, E.J.C.; writing—original draft preparation, P.N.; writing—review and editing, P.N., M.F. and E.J.C.; supervision, E.J.C.; project administration, E.J.C.; funding acquisition, E.J.C. All authors have read and agreed to the published version of the manuscript.
Funding
M.F. is financially supported by Foundation for Science and Technology through the Institutional Scientific Employment program (CEECIND/CP1651/CT0016), the Laboratory of Physics for Materials and Emerging Technologies (LA/P/0095/2020), and Research Unit CeFEMA, Centre of Physics, Engineering and Advanced Materials (UID/04540). E.J.C. is financially supported by Foundation for Science and Technology through the Institutional Scientific Employment program 2nd edition (CEECINST/00038/2021/CP2781/CT005), and the financial support to the Research Unit CEReS, Chemical Engineering and Renewable Resources for Sustainability (UIDB/00102/2020, UIDP/00102/2020, UID/00102/2025, UID/PRR/00102/2025).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
During the preparation of this manuscript, the authors used the GPT-5 and GPT-4.5 language models developed by OpenAI (2025) for grammar checking, typo correction, and clarity enhancement. The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AM | Acetoxymethyl ester |
| CMFDA | 5-chloromethylfluorescein diacetate |
| D | Diopter |
| HPLC | High-performance liquid chromatography |
| PBS | Phosphate-buffered saline |
| RPE | Retinal pigment epithelium |
| TPGS | D-α-tocopheryl polyethylene glycol succinate |
| UV–Vis | Ultraviolet–visible |
| ZO-1 | Zonula occludens-1 |
References
- Remington, L.A. Visual System. Clinical Anatomy and Physiology of the Visual System 2012, 1–9. [Google Scholar] [CrossRef]
- American Academy of Ophthalmology Fundamentals and Principles of Ophthalmology; 2024.
- Remington, L.A. Cornea and Sclera. Clinical Anatomy and Physiology of the Visual System 2012, 10–39. [Google Scholar] [CrossRef]
- Li, S.; Chen, L.; Fu, Y. Nanotechnology-Based Ocular Drug Delivery Systems: Recent Advances and Future Prospects. Journal of Nanobiotechnology 2023 21:1 2023, 21, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.Y.; Tong, L. Barrier Function in the Ocular Surface: From Conventional Paradigms to New Opportunities. Ocular Surface 2015, 13, 103–109. [Google Scholar] [CrossRef]
- Loiseau, A.; Raîche-Marcoux, G.; Maranda, C.; Bertrand, N.; Boisselier, E. Animal Models in Eye Research: Focus on Corneal Pathologies. International Journal of Molecular Sciences 2023, Vol. 24, Page 16661 2023, 24, 16661. [Google Scholar] [CrossRef]
- Zhang, Y.; Do, K.K.; Wang, F.; Lu, X.; Liu, J.Y.; Li, C.; Ceresa, B.P.; Zhang, L.; Dean, D.C.; Liu, Y. Zeb1 Facilitates Corneal Epithelial Wound Healing by Maintaining Corneal Epithelial Cell Viability and Mobility. Commun Biol 2023, 6, 1–12. [Google Scholar] [CrossRef]
- da Silva, A.C.G.; Chialchia, A.R.; de Castro, E.G.; e Silva, M.R.L.; Arantes, D.A.C.; Batista, A.C.; Kitten, G.T.; Valadares, M.C. A New Corneal Epithelial Biomimetic 3D Model for in Vitro Eye Toxicity Assessment: Development, Characterization and Applicability. Toxicology in Vitro 2020, 62, 104666. [Google Scholar] [CrossRef]
- Kaluzhny, Y.; Kinuthia, M.W.; Truong, T.; Lapointe, A.M.; Hayden, P.; Klausner, M. New Human Organotypic Corneal Tissue Model for Ophthalmic Drug Delivery Studies. Invest Ophthalmol Vis Sci 2018, 59, 2880–2898. [Google Scholar] [CrossRef]
- Shiju, T.M.; Carlos de Oliveira, R.; Wilson, S.E. 3D in Vitro Corneal Models: A Review of Current Technologies. Exp Eye Res 2020, 200, 108213. [Google Scholar] [CrossRef]
- Bandeira, F.; Grottone, G.T.; Covre, J.L.; Cristovam, P.C.; Loureiro, R.R.; Pinheiro, F.I.; Casaroli-Marano, R.P.; Donato, W.; Gomes, J.Á.P. A Framework for Human Corneal Endothelial Cell Culture and Preliminary Wound Model Experiments with a New Cell Tracking Approach. International Journal of Molecular Sciences 2023, Vol. 24, Page 2982 2023, 24, 2982. [Google Scholar] [CrossRef]
- Erkoc-Biradli, F.Z.; Ozgun, A.; Öztürk-Öncel, M.Ö.; Marcali, M.; Elbuken, C.; Bulut, O.; Rasier, R.; Garipcan, B. Bioinspired Hydrogel Surfaces to Augment Corneal Endothelial Cell Monolayer Formation. J Tissue Eng Regen Med 2021, 15, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Travers, G.; Coulomb, L.; Aouimeur, I.; He, Z.; Bonnet, G.; Ollier, E.; Gavet, Y.; Moisan, A.; Gain, P.; Thuret, G.; et al. Investigating the Role of Molecular Coating in Human Corneal Endothelial Cell Primary Culture Using Artificial Intelligence-Driven Image Analysis. Sci Rep 2025, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Balters, L.; Reichl, S. 3D Bioprinting of Corneal Models: A Review of the Current State and Future Outlook. J Tissue Eng 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- De Hoon, I.; Boukherroub, R.; De Smedt, S.C.; Szunerits, S.; Sauvage, F. In Vitro and Ex Vivo Models for Assessing Drug Permeation across the Cornea. Mol Pharm 2023, 20, 3298–3319. [Google Scholar] [CrossRef]
- Diebold, Y.; García-Posadas, L. Ex Vivo Applications of Porcine Ocular Surface Tissues: Advancing Eye Research and Alternatives to Animal Studies. Histol Histopathol 2025, 40, 1139–1151. [Google Scholar] [CrossRef]
- Barbalho, G.N.; Falcão, M.A.; Lopes, J.M.S.; Lopes, J.M.; Contarato, J.L.A.; Gelfuso, G.M.; Cunha-Filho, M.; Gratieri, T. Dynamic Ex Vivo Porcine Eye Model to Measure Ophthalmic Drug Penetration under Simulated Lacrimal Flow. Pharmaceutics 2023, 15, 2325. [Google Scholar] [CrossRef]
- Pescina, S.; Govoni, P.; Potenza, A.; Padula, C.; Santi, P.; Nicoli, S. Development of a Convenient Ex Vivo Model for the Study of the Transcorneal Permeation of Drugs: Histological and Permeability Evaluation. J Pharm Sci 2015, 104, 63–71. [Google Scholar] [CrossRef]
- Shi, M.-D.; Sung, K.-C.; Huang, J.-M.; Chen, C.-H.; Wang, Y.-J.; Shi, M.-D.; Sung, K.-C.; Huang, J.-M.; Chen, C.-H.; Wang, Y.-J. Development of an Ex Vivo Porcine Eye Model for Exploring the Pathogenicity of Acanthamoeba. Microorganisms 2024, Vol. 12, Page 1161 2024, 12, 1161. [Google Scholar] [CrossRef]
- Okurowska, K.; Roy, S.; Thokala, P.; Partridge, L.; Garg, P.; Macneil, S.; Monk, P.N.; Karunakaran, E. Establishing a Porcine Ex Vivo Cornea Model for Studying Drug Treatments against Bacterial Keratitis. J Vis Exp 2020, 2020. [Google Scholar] [CrossRef]
- Tsai, M.C.; Kureshi, A.; Daniels, J.T. Establishment of an Ex Vivo Human Corneal Endothelium Wound Model. Transl Vis Sci Technol 2025, 14, 24–24. [Google Scholar] [CrossRef]
- Castro, N.; Gillespie, S.R.; Bernstein, A.M. Ex Vivo Corneal Organ Culture Model for Wound Healing Studies. J Vis Exp 2019, 2019. [Google Scholar] [CrossRef]
- Netto, A.R.T.; Hurst, J.; Bartz-Schmidt, K.-U.; Schnichels, S.; Rocha, A.; Netto, T.; Hurst, J.; Bartz-Schmidt, K.-U.; Schnichels, S. Porcine Corneas Incubated at Low Humidity Present Characteristic Features Found in Dry Eye Disease. International Journal of Molecular Sciences 2022, Vol. 23, Page 4567 2022, 23, 4567. [Google Scholar] [CrossRef]
- Rouhbakhshzaeri, M.; Rabiee, B.; Azar, N.; Ghahari, E.; Putra, I.; Eslani, M.; Djalilian, A.R. New Ex Vivo Model of Corneal Endothelial Phacoemulsification Injury and Rescue Therapy with Mesenchymal Stromal Cell Secretome. J Cataract Refract Surg 2019, 45, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues da Penha, J.; Garcia da Silva, A.C.; de Ávila, R.I.; Valadares, M.C. Development of a Novel Ex Vivo Model for Chemical Ocular Toxicity Assessment and Its Applicability for Hair Straightening Products. Food and Chemical Toxicology 2022, 170. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, M.; Behl, G.; Rani, S.; O’Reilly, N.; McLoughlin, P.; O’Donovan, O.; Lynch, J.; Fitzhenry, L.; Sarfraz, M.; Reynolds, A.L.; et al. Development and in Vitro and Ex Vivo Characterization of a Twin Nanoparticulate System to Enhance Ocular Absorption and Prolong Retention of Dexamethasone in the Eye: From Lab to Pilot Scale Optimization. Nanoscale Adv 2025, 7, 3125–3142. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.S.; Shrestha, D.; Rupenthal, I.D. Evaluation of 2 Ex Vivo Bovine Cornea Storage Protocols for Drug Delivery Applications. Ophthalmic Res 2019, 61, 204–209. [Google Scholar] [CrossRef]
- Shehab, A.; Gram, N.; Ivarsen, A.; Hjortdal, J. The Importance of Donor Characteristics, Post-Mortem Time and Preservation Time for Use and Efficacy of Donated Corneas for Posterior Lamellar Keratoplasty. Acta Ophthalmol 2022, 100, 269–276. [Google Scholar] [CrossRef]
- Wilson, S.E. Bowman’s Layer in the Cornea– Structure and Function and Regeneration. Exp Eye Res 2020, 195, 108033. [Google Scholar] [CrossRef]
- Thomasy, S.M.; Eaton, J.S.; Timberlake, M.J.; Miller, P.E.; Matsumoto, S.; Murphy, C.J. Species Differences in the Geometry of the Anterior Segment Differentially Affect Anterior Chamber Cell Scoring Systems in Laboratory Animals. https://home.liebertpub.com/jop 2016, 32, 28–37. [Google Scholar] [CrossRef]
- Vézina, M. Comparative Ocular Anatomy in Commonly Used Laboratory Animals. Molecular and Integrative Toxicology 2012, 1–21. [Google Scholar] [CrossRef]
- Sanchez, I.; Martin, R.; Ussa, F.; Fernandez-Bueno, I. The Parameters of the Porcine Eyeball. Graefes Arch Clin Exp Ophthalmol 2011, 249, 475–482. [Google Scholar] [CrossRef]
- Schnichels, S.; Paquet-Durand, F.; Löscher, M.; Tsai, T.; Hurst, J.; Joachim, S.C.; Klettner, A. Retina in a Dish: Cell Cultures, Retinal Explants and Animal Models for Common Diseases of the Retina. Prog Retin Eye Res 2021, 81, 100880. [Google Scholar] [CrossRef]
- Peterson, J.S.; Chen, W.; Libin, B.M.; S. boyer, D.; Kaiser, P.K.; Liebmann, J.M. Sustained Ocular Delivery of Bevacizumab Using Densomeres in Rabbits: Effects on Molecular Integrity and Bioactivity. Transl Vis Sci Technol 2023, 12, 28–28. [Google Scholar] [CrossRef]
- Chauchat, L.; Guerin, C.; Kaluzhny, Y.; Renard, J.P. Comparison of In Vitro Corneal Permeation and In Vivo Ocular Bioavailability in Rabbits of Three Marketed Latanoprost Formulations. Eur J Drug Metab Pharmacokinet 2023, 48, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gao, N.; Yu, F.S. Interleukin-36 Receptor Signaling Attenuates Epithelial Wound Healing in C57BL/6 Mouse Corneas. Cells 2023, Vol. 12, Page 1587 2023, 12, 1587. [Google Scholar] [CrossRef] [PubMed]
- Akpek, E.K.; Aldave, A.J.; Amescua, G.; Colby, K.A.; Cortina, M.S.; Cruz, J.; Parel, J.M.A.; Li, G. Twelve-Month Clinical and Histopathological Performance of a Novel Synthetic Cornea Device in Rabbit Model. Transl Vis Sci Technol 2023, 12, 9–9. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, K.; Hatou, S.; Inagaki, E.; Higa, K.; Tsubota, K.; Shimmura, S. A Rabbit Corneal Endothelial Dysfunction Model Using Endothelial-Mesenchymal Transformed Cells. Sci Rep 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Sun, F.; Xi, L.W.Q.; Luu, W.; Enkhbat, M.; Neo, D.; Mehta, J.S.; Peh, G.S.L.; Yim, E.K.F. Preclinical Models for Studying Fuchs Endothelial Corneal Dystrophy. Cells 2025, Vol. 14, Page 505 2025, 14, 505. [Google Scholar] [CrossRef]
- Koseoglu, N.D.; Chen, E.; Tuwani, R.; Kompa, B.; Cox, S.M.; Cuneyt Ozmen, M.; Massaro-Giordano, M.; Beam, A.L.; Hamrah, P. Development and Validation of a Deep Learning Model for Diagnosing Neuropathic Corneal Pain via in Vivo Confocal Microscopy. NPJ Digit Med 2025, 8, 1–8. [Google Scholar] [CrossRef]
- Kiani, A.K.; Pheby, D.; Henehan, G.; Brown, R.; Sieving, P.; Sykora, P.; Marks, R.; Falsini, B.; Capodicasa, N.; Miertus, S.; et al. Ethical Considerations Regarding Animal Experimentation. J Prev Med Hyg 2022, 63, E255. [Google Scholar] [CrossRef]
- Mann, B.K.; Stirland, D.L.; Lee, H.K.; Wirostko, B.M. Ocular Translational Science: A Review of Development Steps and Paths. Adv Drug Deliv Rev 2018, 126, 195–203. [Google Scholar] [CrossRef]
- Middleton, S. Porcine Ophthalmology. Veterinary Clinics of North America: Food Animal Practice 2010, 26, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Menduni, F.; Davies, L.N.; Madrid-Costa, D.; Fratini, A.; Wolffsohn, J.S. Characterisation of the Porcine Eyeball as an In-Vitro Model for Dry Eye. Contact Lens and Anterior Eye 2018, 41, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Moral, M.; García-Posadas, L.; López-García, A.; Diebold, Y. Histological and Immunohistochemical Characterization of the Porcine Ocular Surface. PLoS One 2020, 15, e0227732. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.H.; Choi, H.J.; Kim, M.K. Corneal Xenotransplantation: Where Are We Standing? Prog Retin Eye Res 2021, 80. [Google Scholar] [CrossRef]
- Peynshaert, K.; Devoldere, J.; De Smedt, S.C.; Remaut, K. In Vitro and Ex Vivo Models to Study Drug Delivery Barriers in the Posterior Segment of the Eye. Adv Drug Deliv Rev 2018, 126, 44–57. [Google Scholar] [CrossRef]
- Loewen, N.; Fallano, K.; Bussel, I.; Kagemann, L.; Lathrop, K.L. Training Strategies and Outcomes of Ab Interno Trabeculectomy with the Trabectome. F1000Res 2017, 6. [Google Scholar] [CrossRef]
- Brunette, I.; Rosolen, S.G.; Carrier, M.; Abderrahman, M.; Nada, O.; Germain, L.; Proulx, S. Comparison of the Pig and Feline Models for Full Thickness Corneal Transplantation. Vet Ophthalmol 2011, 14, 365–377. [Google Scholar] [CrossRef]
- Rouhbakhshzaeri, M.; Rabiee, B.; Azar, N.; Ghahari, E.; Putra, I.; Eslani, M.; Djalilian, A.R. New Ex Vivo Model of Corneal Endothelial Phacoemulsification Injury and Rescue Therapy with Mesenchymal Stromal Cell Secretome. J Cataract Refract Surg 2019, 45, 361–366. [Google Scholar] [CrossRef]
- Foja, S.; Heinzelmann, J.; Hünniger, S.; Viestenz, A.; Rüger, C.; Viestenz, A. Drug-Dependent Inhibitory Effects on Corneal Epithelium Structure, Cell Viability, and Corneal Wound Healing by Local Anesthetics. International Journal of Molecular Sciences 2024, Vol. 25, Page 13074 2024, 25, 13074. [Google Scholar] [CrossRef]
- Choy, E.P.Y.; To, T.S.S.; Cho, P.; Benzie, I.F.F.; Choy, C.K.M. Viability of Porcine Corneal Epithelium Ex Vivo and Effect of Exposure to Air: A Pilot Study for a Dry Eye Model. Cornea 2004, 23, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.Y.; Cho, P.; Boost, M. Corneal Epithelial Cell Viability of an Ex Vivo Porcine Eye Model. Clin Exp Optom 2014, 97, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Rodella, U.; Bosio, L.; Ferrari, S.; Gatto, C.; Giurgola, L.; Rossi, O.; Ciciliot, S.; Ragazzi, E.; Ponzin, D.; Tóthová, J.D. Porcine Cornea Storage Ex Vivo Model as an Alternative to Human Donor Tissues for Investigations of Endothelial Layer Preservation. Transl Vis Sci Technol 2023, 12, 24. [Google Scholar] [CrossRef]
- Avila, M.Y.; Gerena, V.A.; Navia, J.L. Corneal Crosslinking with Genipin, Comparison with UV-Riboflavin in Ex-Vivo Model. Mol Vis 2012, 18, 1068. [Google Scholar] [PubMed]
- Kim, K.W.; Shin, Y.J.; Lee, S.C.S. Novel ROCK Inhibitors, Sovesudil and PHP-0961, Enhance Proliferation, Adhesion and Migration of Corneal Endothelial Cells. Int J Mol Sci 2022, 23, 14690. [Google Scholar] [CrossRef]
- Poole, C.A.; Brookes, N.H.; Clover, G.M. Confocal Imaging of the Keratocyte Network in Porcine Cornea Using the Fixable Vital Dye 5-Chloromethylfluorescein Diacetate. Curr Eye Res 1996, 15, 165–174. [Google Scholar] [CrossRef]
- De Hoon, I.; Boukherroub, R.; De Smedt, S.C.; Szunerits, S.; Sauvage, F. In Vitro and Ex Vivo Models for Assessing Drug Permeation across the Cornea. Mol Pharm 2023, 20, 3298–3319. [Google Scholar] [CrossRef]
- Resende, A.P.; Silva, B.; Braz, B.S.; Nunes, T.; Gonçalves, L.; Delgado, E. Ex Vivo Permeation of Erythropoietin through Porcine Conjunctiva, Cornea, and Sclera. Drug Deliv Transl Res 2017, 7, 625–631. [Google Scholar] [CrossRef]
- Brütsch, D.R.; Hunziker, P.; Pot, S.; Tappeiner, C.; Voelter, K. Corneal and Scleral Permeability of Desmoteplase in Different Species. Vet Ophthalmol 2020, 23, 785–791. [Google Scholar] [CrossRef]
- Gómez-Segura, L.; Parra, A.; Calpena, A.C.; Gimeno, Á.; Boix-Montañes, A. Carprofen Permeation Test through Porcine Ex Vivo Mucous Membranes and Ophthalmic Tissues for Tolerability Assessments: Validation and Histological Study. Veterinary Sciences 2020, Vol. 7, Page 152 2020, 7, 152. [Google Scholar] [CrossRef]
- Guillot, A.J.; Petalas, D.; Skondra, P.; Rico, H.; Garrigues, T.M.; Melero, A. Ciprofloxacin Self-Dissolvable Soluplus Based Polymeric Films: A Novel Proposal to Improve the Management of Eye Infections. Drug Deliv Transl Res 2021, 11, 608–625. [Google Scholar] [CrossRef]
- Ostacolo, C.; Caruso, C.; Tronino, D.; Troisi, S.; Laneri, S.; Pacente, L.; Del Prete, A.; Sacchi, A. Enhancement of Corneal Permeation of Riboflavin-5′-Phosphate through Vitamin E TPGS: A Promising Approach in Corneal Trans-Epithelial Cross Linking Treatment. Int J Pharm 2013, 440, 148–153. [Google Scholar] [CrossRef]
- Cardoso, T.P. de A.; Kishishita, J.; Souza, A.T.M.; Vieira, J.R.C.; Melo, C.M.L. de; Santana, D.P. de; Leal, L.B. A Proposed Eye Ex Vivo Permeation Approach to Evaluate Pesticides: Case Dimethoate. Toxicology in Vitro 2020, 66, 104833. [Google Scholar] [CrossRef] [PubMed]
- Barbalho, G.N.; Falcão, M.A.; Lopes, J.M.S.; Lopes, J.M.; Contarato, J.L.A.; Gelfuso, G.M.; Cunha-Filho, M.; Gratieri, T. Dynamic Ex Vivo Porcine Eye Model to Measure Ophthalmic Drug Penetration under Simulated Lacrimal Flow. Pharmaceutics 2023, 15, 2325. [Google Scholar] [CrossRef] [PubMed]
- Bhujbal, S.; Rupenthal, I.D.; Agarwal, P. Evaluation of Ocular Tolerability and Bioavailability of Tonabersat Transfersomes Ex Vivo. Drug Deliv Transl Res 2025, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Mei, H.; Cui, H.; Song, M.; Qiao, Y.; Chen, J.; Lei, Y. Permeation Dynamics of Organic Moiety-Tuned Organosilica Nanoparticles across Porcine Corneal Barriers: Experimental and Mass Transfer Analysis for Glaucoma Drug Delivery. Biomed Phys Eng Express 2025, 11, 045015. [Google Scholar] [CrossRef]
- Sarfraz, M.; Behl, G.; Rani, S.; O’Reilly, N.; McLoughlin, P.; O’Donovan, O.; Lynch, J.; Fitzhenry, L.; Sarfraz, M.; Reynolds, A.L.; et al. Development and in Vitro and Ex Vivo Characterization of a Twin Nanoparticulate System to Enhance Ocular Absorption and Prolong Retention of Dexamethasone in the Eye: From Lab to Pilot Scale Optimization. Nanoscale Adv 2025, 7, 3125–3142. [Google Scholar] [CrossRef]
- Begum, G.; Leigh, T.; Courtie, E.; Moakes, R.; Butt, G.; Ahmed, Z.; Rauz, S.; Logan, A.; Blanch, R.J. Rapid Assessment of Ocular Drug Delivery in a Novel Ex Vivo Corneal Model. Sci Rep 2020, 10. [Google Scholar] [CrossRef]
- Cauwe, B.; Steen, P.E.V. Den; Opdenakker, G. The Biochemical, Biological, and Pathological Kaleidoscope of Cell Surface Substrates Processed by Matrix Metalloproteinases. Crit Rev Biochem Mol Biol 2007, 42, 113–185. [Google Scholar] [CrossRef]
- Chen, W.; Li, Z.; Hu, J.; Zhang, Z.; Chen, L.; Chen, Y.; Liu, Z. Corneal Alternations Induced by Topical Application of Benzalkonium Chloride in Rabbit. PLoS One 2011, 6, e26103. [Google Scholar] [CrossRef]
- Guo, P.; Li, N.; Fan, L.; Lu, J.; Liu, B.; Zhang, B.; Wu, Y.; Liu, Z.; Li, J.; Pi, J.; et al. Study of Penetration Mechanism of Labrasol on Rabbit Cornea by Ussing Chamber, RT-PCR Assay, Western Blot and Immunohistochemistry. Asian J Pharm Sci 2019, 14, 329–339. [Google Scholar] [CrossRef]
- Oliveira, S.; Guimarães, P.; Campos, E.J.; Fernandes, R.; Martins, J.; Castelo-Branco, M.; Serranho, P.; Matafome, P.; Bernardes, R.; Ambrósio, A.F. Retinal OCT-Derived Texture Features as Potential Biomarkers for Early Diagnosis and Progression of Diabetic Retinopathy. Invest Ophthalmol Vis Sci 2025, 66. [Google Scholar] [CrossRef]
- Mohamed, Y.H.; Uematsu, M.; Kusano, M.; Inoue, D.; Tang, D.; Suzuki, K.; Kitaoka, T. A Novel Technique for Corneal Transepithelial Electrical Resistance Measurement in Mice. Life 2024, Vol. 14, Page 1046 2024, 14, 1046. [Google Scholar] [CrossRef]
Figure 1.
Histologic section showing the 5 layers of the cornea: the epithelium (40–50 μm), the Bowman layer (8–15 μm), the stroma (470–500 μm), the Descemet membrane (10–12 μm), and the endothelium (4–6 μm). B = Bowman layer; D = Descemet membrane; En = endothelium; Ep = epithelium; S = stroma [2].
Figure 1.
Histologic section showing the 5 layers of the cornea: the epithelium (40–50 μm), the Bowman layer (8–15 μm), the stroma (470–500 μm), the Descemet membrane (10–12 μm), and the endothelium (4–6 μm). B = Bowman layer; D = Descemet membrane; En = endothelium; Ep = epithelium; S = stroma [2].
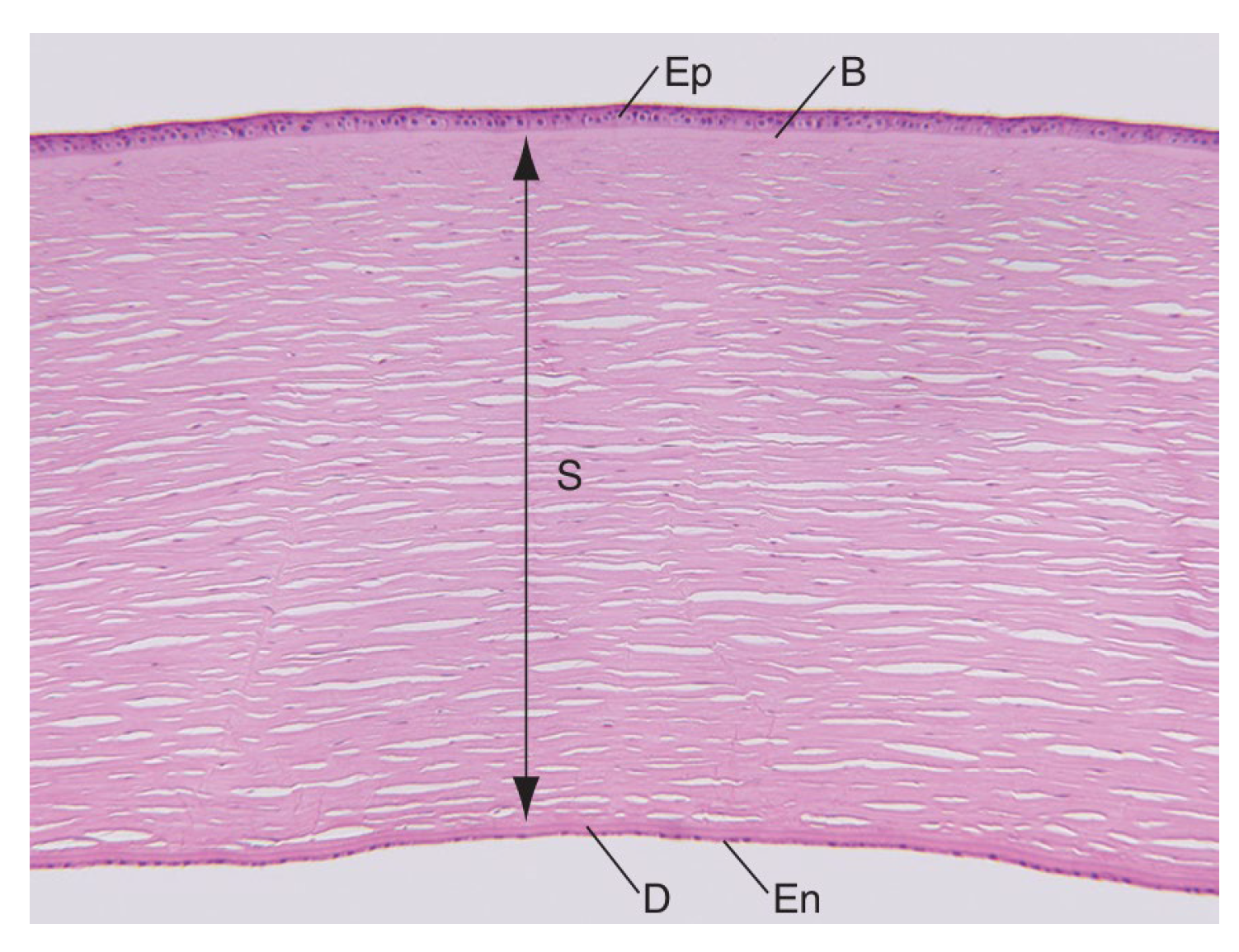
Figure 2.
Schematic image of the corneal epithelium demonstrating adhesion between cells and to the under lying basal lamina (purple) and the Bowman layer via hemidesmosomes [2].
Figure 2.
Schematic image of the corneal epithelium demonstrating adhesion between cells and to the under lying basal lamina (purple) and the Bowman layer via hemidesmosomes [2].
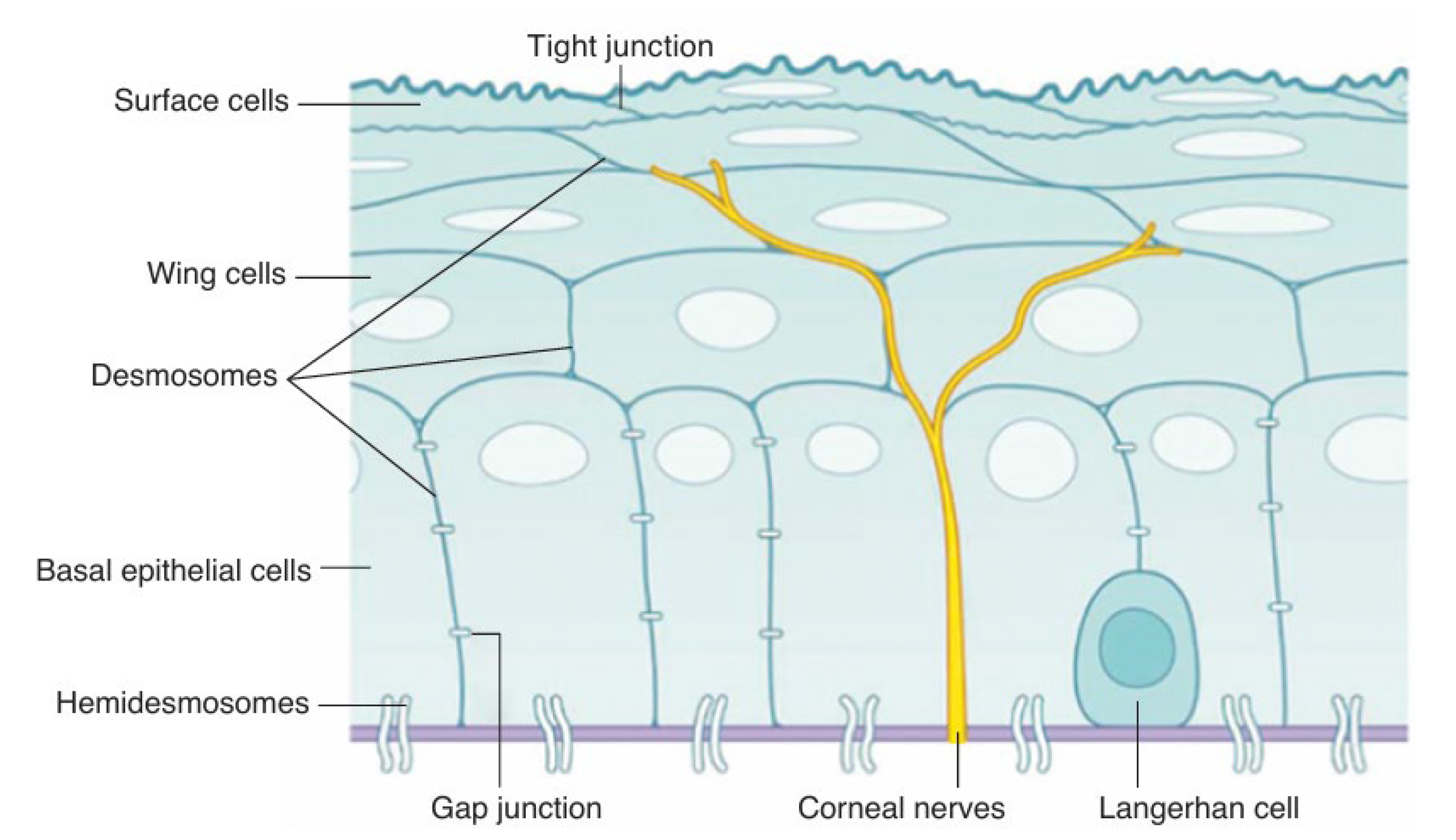
Figure 3.
Schematic representation of the tear film (Adapted from[3]). Illustration created with BioRender.
Figure 3.
Schematic representation of the tear film (Adapted from[3]). Illustration created with BioRender.
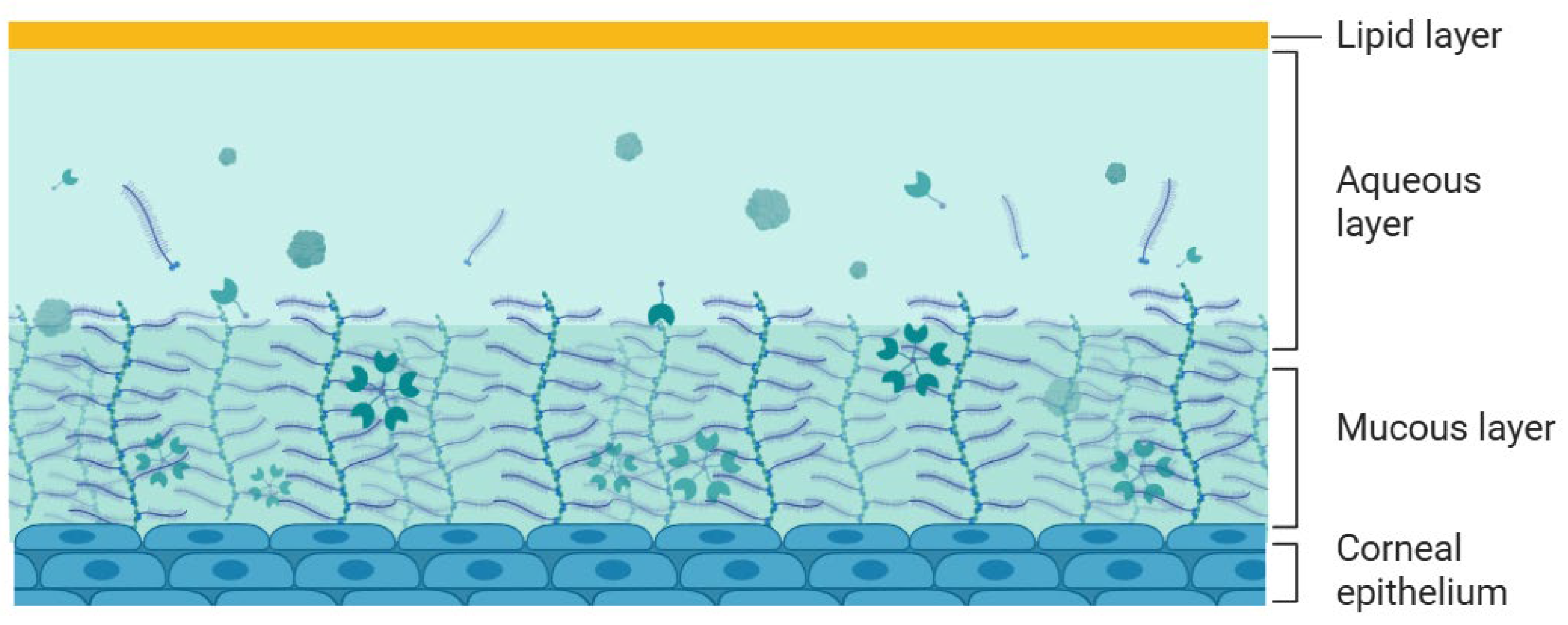
Figure 4.
Schematic representation of the corneal epithelium, highlighting the structure of tight junctions in the superficial epithelial layer. The zoomed-in area illustrates the intercellular junctional complex composed of transmembrane proteins, such as occludin and claudins, and the cytoplasmic scaffold protein ZO-1, which anchors the complex to the actin cytoskeleton. These proteins are essential to maintain the epithelial barrier function and regulate paracellular permeability. Illustration created with BioRender.
Figure 4.
Schematic representation of the corneal epithelium, highlighting the structure of tight junctions in the superficial epithelial layer. The zoomed-in area illustrates the intercellular junctional complex composed of transmembrane proteins, such as occludin and claudins, and the cytoplasmic scaffold protein ZO-1, which anchors the complex to the actin cytoskeleton. These proteins are essential to maintain the epithelial barrier function and regulate paracellular permeability. Illustration created with BioRender.
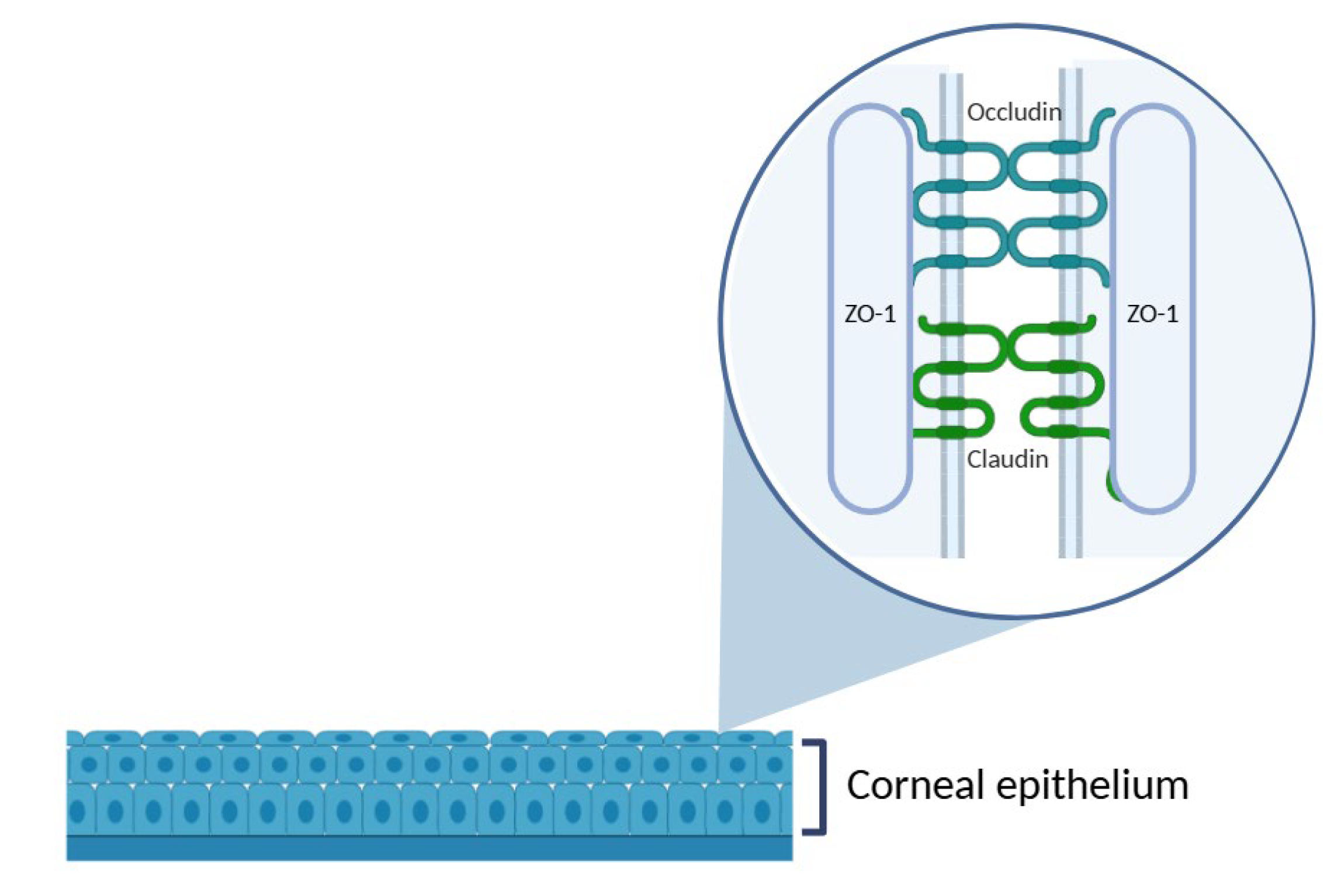
Figure 5.
Schematic representation of the procedure used to assess epithelial cell viability in ex vivo porcine corneal models, using trypan blue.
Figure 5.
Schematic representation of the procedure used to assess epithelial cell viability in ex vivo porcine corneal models, using trypan blue.
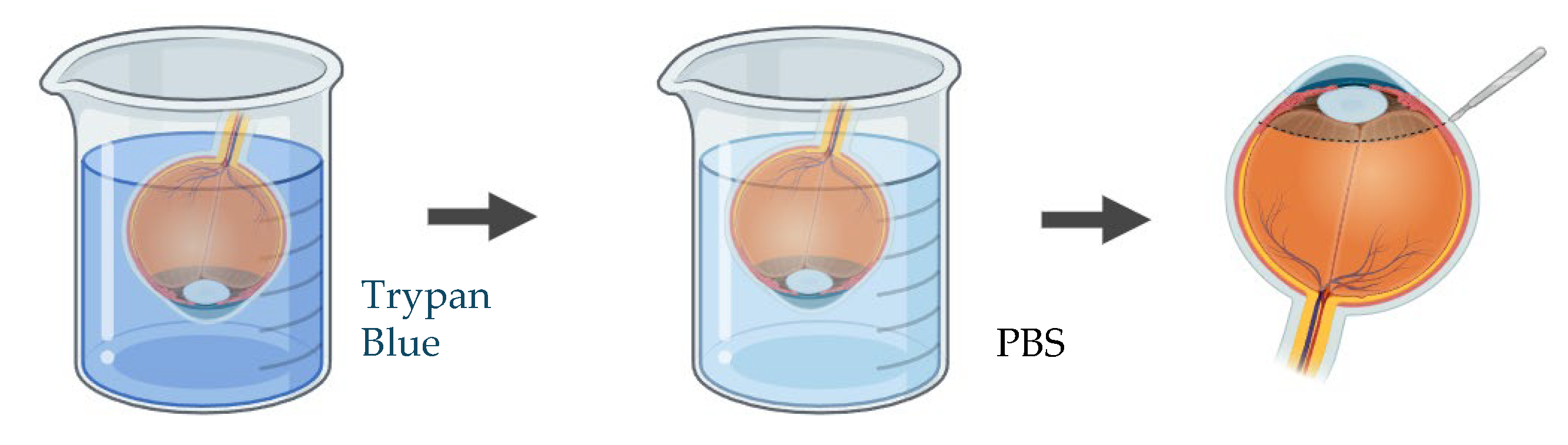
Figure 6.
Schematic representation of the procedure used to assess endothelial cell viability in ex vivo porcine corneal models, using trypan blue and alizarin red S staining.
Figure 6.
Schematic representation of the procedure used to assess endothelial cell viability in ex vivo porcine corneal models, using trypan blue and alizarin red S staining.
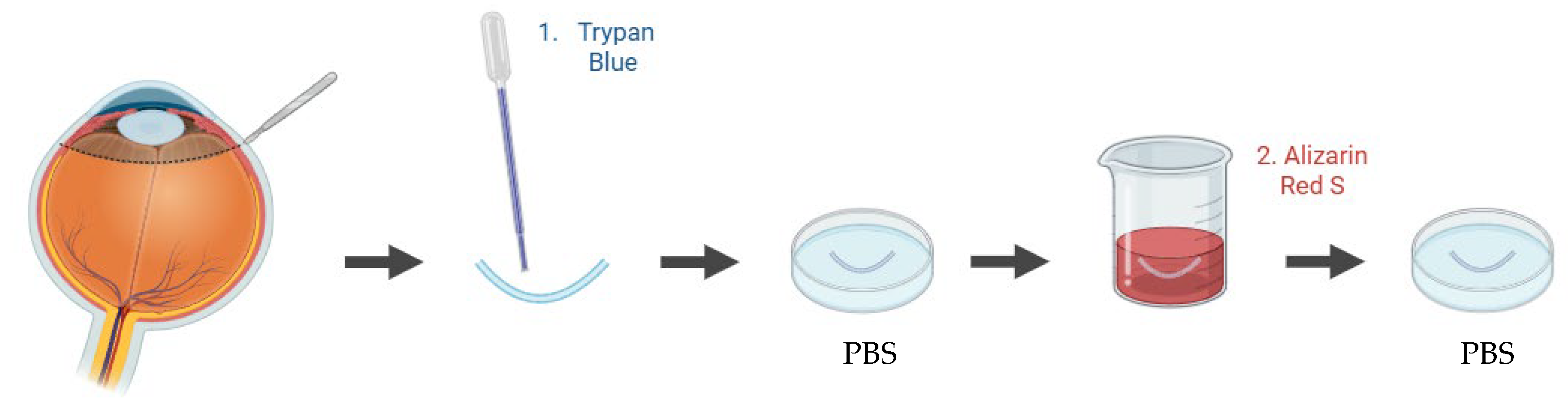
Figure 7.
Schematic representation of a Franz diffusion cell setup using an ex vivo porcine cornea for drug permeability assessment. Illustration created with BioRender.
Figure 7.
Schematic representation of a Franz diffusion cell setup using an ex vivo porcine cornea for drug permeability assessment. Illustration created with BioRender.
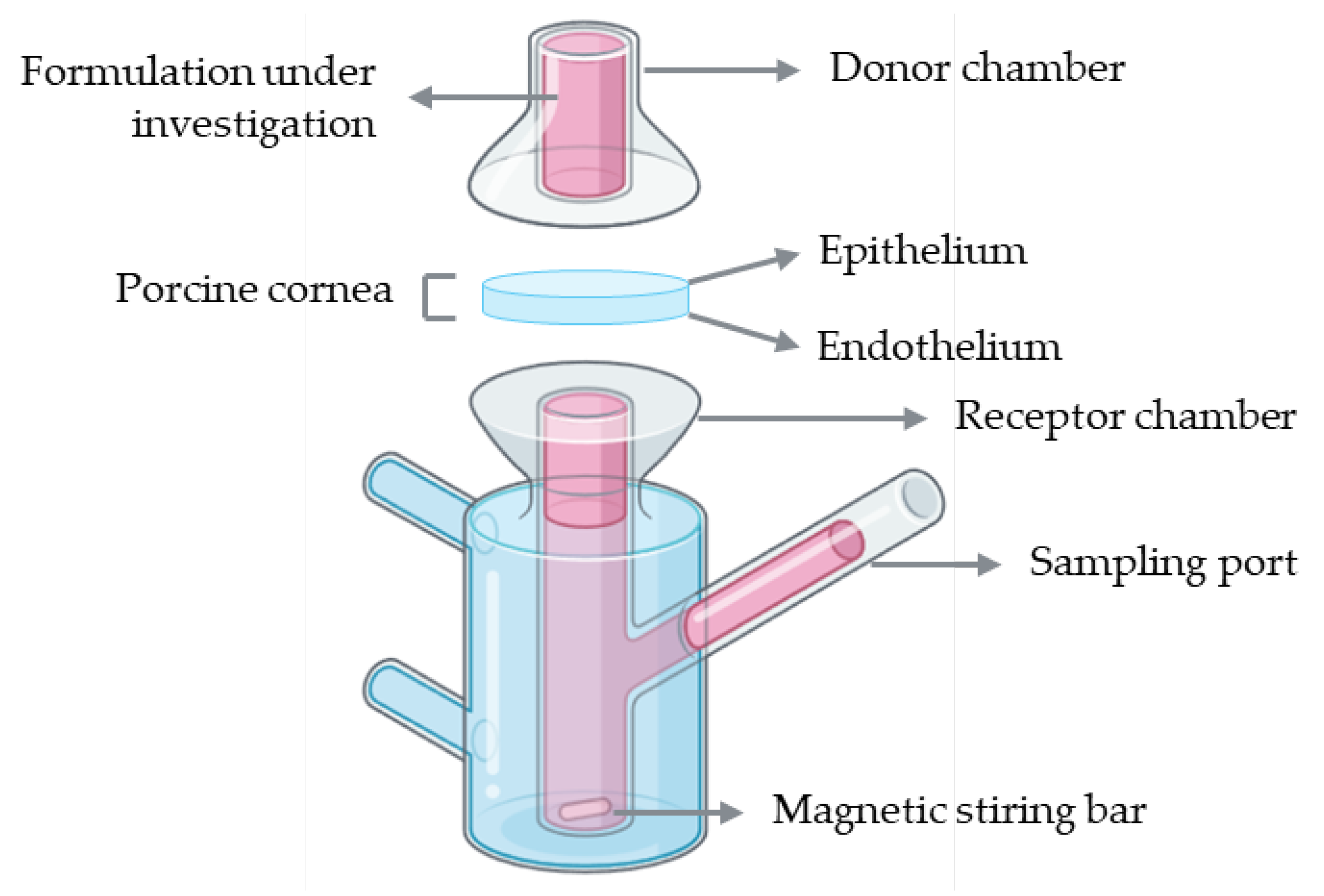
Figure 8.
Schematic representation of the procedure used to assess corneal permeability to a formulation under investigation using the ex vivo cell insert model.
Figure 8.
Schematic representation of the procedure used to assess corneal permeability to a formulation under investigation using the ex vivo cell insert model.
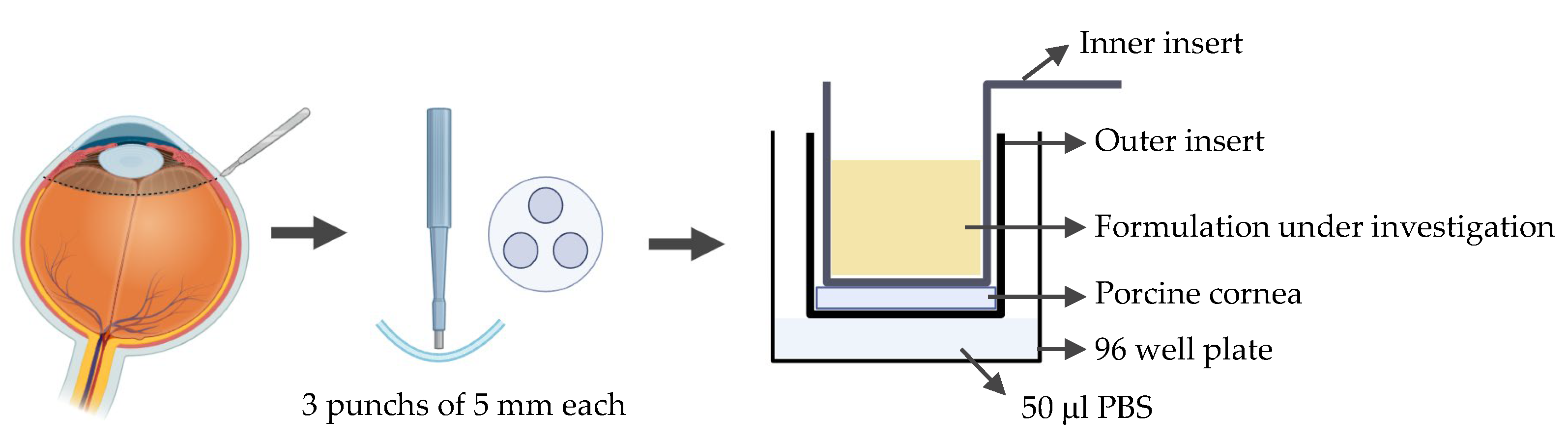

Table 1.
Dimensions and Parameters of the Human Adult Eye.
| Anterior chamber | Posterior chamber | Vitreous | Whole eye | |
|---|---|---|---|---|
| Average depth (emmetropic eye) | 3.11 mm | 0.52 mm | 16.5 mm | 23-25 mm |
| Volume | 220 μL | 60 μL | 5-6 mL | 6.5-7 mL |
| Content | Aqueous | Aqueous | Vitreous |
Table 2.
Comparison of ocular parameters between the human and relevant laboratory animal species. The values presented are average measurements reported in the literature. It is worth noting that several factors, such as age, physiological condition, and the measurement method, can significantly influence these parameters.
Table 2.
Comparison of ocular parameters between the human and relevant laboratory animal species. The values presented are average measurements reported in the literature. It is worth noting that several factors, such as age, physiological condition, and the measurement method, can significantly influence these parameters.
| Mouse | Rat | Rabbit | Porcine | Human | |
|---|---|---|---|---|---|
| Average eye dimension in volume (cm3) | 0.025 | 0.1 | 2.6 | 6.5 | 7.2 |
| Average eye dimension (axial length in mm) | 3.4 | 6.0 | 17.1 | 23.9 | 24 |
| Corneal horizontal diameter (mm) | 3.15 | 5.1 | 13.4 | 14.3 | 11.81 |
| Corneal thickness (µm) | 0.089-0.123 | 0.16-2 | 0.36 | 543-797 | 530-710 |
| Cornea shape | flat | flat | dome | dome | dome |
| Bowman’s membrane | No | No | No | No | Yes |
| Time between eye blinks | 5 min | 5 min | 6 min | 20-30 s | 5 s |
| References | [6,29,30,31] | [6,29,30,31,32] | [1,6,29,30,31,33] | ||
Table 3.
Comparison of ocular parameters between the human and porcine eye. The values presented are average measurements reported in the literature. It is important to note that several factors, such as age, physiological condition, and the measurement method, can significantly influence these parameters.
Table 3.
Comparison of ocular parameters between the human and porcine eye. The values presented are average measurements reported in the literature. It is important to note that several factors, such as age, physiological condition, and the measurement method, can significantly influence these parameters.
| Porcine | Human | |
|---|---|---|
| Corneal curvature | 7.85-8.28 mm [44] | 6.5-7.8 mm [3] |
| Corneal epithelium thickness | 80 μm [32] | 50 μm [3] |
| Corneal epithelium cell layers | 6-8 layers [45] | 4-6 layers [2] |
| Corneal endothelium cell density | 3250 cell/mm2 [44] | 2496.9-4049.5 cell/mm2 [44] |
| Retinal thickness | 300 μm [33] | 310 μm [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



