Submitted:
20 October 2025
Posted:
22 October 2025
You are already at the latest version
Abstract
As the global threat from climate change continues to increase almost unchecked, the negative impacts of climate change on biodiversity are also increasing, and this could result in the loss of numerous species, habitat fragmentation, and phenological change. Here, we look at the global impact climate change poses to one particular group of birds: Owls (Strigiformes). As apex predators inhabiting every continent and in almost every country of the world, with the exception of Antarctica and some small isolated islands, they are an ideal group to study in relation to negative impacts caused by extreme climatic events globally. Examining multiple published studies from around the world, we determine which species are most susceptible to be negatively impacted by one or multiple events caused by climate change: wildfires, unprecedented levels of precipitation and flooding, rising temperatures, drought, melting of ice and snow, storm events, and rising sea levels. The Tawny Owl Strix aluco offers one of the first evidences that recent climate change can alter natural selection in a wild population leading to a micro-evolutionary response, which demonstrates the ability of wild populations to evolve in response to climate change.
Keywords:
owls
; climate change
; ecosystems
; increasing temperatures
; microevolution
; wildfires
; precipitation
; flooding
; rising sea level
; drought
; snow structure
; thawing
; habitat loss
; nestbox competition
; prey species
1. Introduction
Climate change is one of the greatest ecological and social challenges of the 21st century [1], and while opinion polls continue to show that the increasing threat of climate change is at the top of public concern, continuing geopolitical discord and the reluctance of wealthy industrial nations to vastly reduce their reliance on fossil fuels are only increasing those challenges. According to the United Nations (UN) agency, the World Meteorological Organization (WMO), 2024 was the warmest year on record globally, while one WMO report finds that there is a staggering 86% chance that global average temperatures will exceed 1.5 °C above pre-industrial levels in at least one of the next five years, and a 1% chance of one of those years exceeding 2 °C of warming [2].
Biodiversity and climate are interconnected to each other and, as such, climate change is a continuous threat to wildlife and ecosystems across the world. The basic cause of these climatic changes is human activities. These anthropogenic activities increase the greenhouse gases (such as carbon dioxide, methane, nitrous oxides, and chlorofluorocarbons), which cause the greenhouse effect, where the rising temperature of the atmosphere causes thermal optima to shift towards high altitudes and high latitudes, causing long-term change in weather patterns, including temperature, precipitation, and storm events. Impacts of climate change on biodiversity are continuous, and this could result in the loss of numerous species, habitat fragmentation, and phenological change. Even a slight temperature change will have a complex sequence in terms of species distribution and numbers, affecting behaviour, reproduction, migration, and foraging. Therefore, it is critical to address climate change for biodiversity conservation and ecosystem management [3].
Here, we look at the global threats climate change poses to one particular group of birds: Owls (Strigiformes). Owls are an important study group in terms of the impact climate change has on a wide variety of ecosystems globally. Not only are they apex predators, where their high trophic position allows them to be prime indicators to the health of lower trophic levels and the availability of prey, more importantly, species of owls are found on every continent and in almost every country of the world, with the exception of Antarctica and some small isolated islands, and can thrive in habitats as diverse as frozen tundra, equatorial rainforests, temperate northern forests, and even open grasslands and deserts [4].
Because of their vast geographical range and the incredible diversity of habitats, many owl species are negatively impacted by one or multiple events caused by climate change: wildfires, unprecedented levels of precipitation and flooding, rising temperatures, drought, ice and snow melt, storm events, and rising sea levels. However, it is challenging to separate the impacts of climate change on owl biodiversity from those of other global change drivers such as urbanization, land-use change, and increased long-distance trade [5]. It is also important to remember that avian response to climate change may be species-specific and not all species exhibit changing phenology despite regional climatic changes [6].
Climate change is a global phenomen, but most of our examples and references are from northern regions. Warming takes place in tropical areas as well but its impact to owls is more difficult to recognize in an environment that is initially warm and with large human populations and human activity. Therefore, in an increasingly anthropized world, alpine tundra and boreal forest ecosystems can be seen better ruled by climate change rather than by human disturbance.
2. Methods
In addition to a global literature search, we have used published active nest and fledged broods of owls material from Birdlife Finland (Year books 2008‒2024)[7] in order to see the assumed northward movements of the populations due to climate change [8]. Two seven-year periods have been compared, 2008‒2014 and 2018‒2024. South Finland includes BirdLife Finland societies 1, 11,12, 21‒23, and 31, Central Finland includes 32, 41‒44, 46, 51, 54, 57, 61, 71‒74, and North Finland respectively 81, 82, 91, and 92 (Map 1). Reasoning for the seven-year period is that it should cover two peak vole years and two low vole years in the 3-year vole cycle to reduce the cyclic variation in the average period nest numbers (Table 1). The main purpose was to see if the owl populations have moved north from the southern and central parts of the country, as the southern part of the country covers only the coastal areas and Åland archipelago, and the northern parts include north Ostrobothnia and Kainuu, resembling Lapland more than Central Finland. Table 2 summarizes known climate change impacts on owl species, including data collected from global literature and from this study based on BirdLife Finland yearbook material as presented in Table 1.
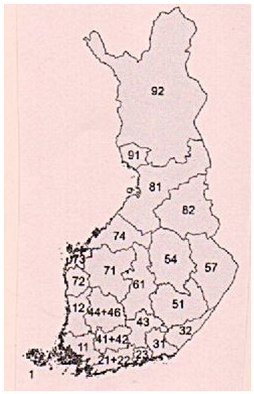 |
| Map 1. BirdLife Finland observation areas [9]. |
3. Results
Table 1 and Table 2 show that in Finland, all owl species except Tawny Owls Strix aluco and Great Grey Owls Strix nebulosa have declining populations based on these two 7-year periods from 2008‒14 and 2018‒24. Tawny and Great Grey Owls have increased nearly 30 and 9 per cent, respectively, while Snowy Owls Bubo scandiacus and Asio-owls have seen the greatest decline (Snowy Owl 88%, Long-eared Owl Asio otus 65% and Short-eared Owl Asio flammeus 63%). Almost similar reductions have taken place with Tengmalm’s Owl Aegolius funereus, Northern Hawk Owl Surnia ulula, and Eurasian Pygmy Owl Glaucidium passerinum, with 59, 59, and 58%. Less decline has taken place in Eurasian Eagle Owls Bubo bubo and Ural Owls Strix uralensis (22% and 25% respectively). Decreasing populations make it difficult to see clearly how much owl populations would have moved northwards, as assumed to happen due to climate change, especially with the boreal forest owls.
It seems that the Short-eared Owl, Tengmalm’s Owl, Great Grey Owl, and Northern Hawk Owl have seen little increase in their nesting in the north. Southern species, Tawny Owl and Long-eared Owl, have increased their population share in the south, but climate change has not yet assisted them in moving further north in Finland. The Eurasian Eagle Owl and the Pygmy Owl has shown only minor shifts between south, central, and north (Table 1).
Climate change is taking place more severely in the Arctic, rapidly affecting Arctic ecosystems [44]. Long-term Arctic breeding surveys suggest that the world population of Snowy Owls is much lower than estimated earlier. The breeding range is 12 million km2, but only 1.3 million of that has a high probability for breeding, i.e., breeding at no more than 3‒9 year intervals. Population size (previously 50,000‒290,000 individuals in [45]) is now calculated to be only 14,000‒28,000 mature individuals, making the owl vulnerable [19]. Roughly half the remaining population lives in the Canadian Arctic. In many countries, like Finland, the owl is already seen as Critically Endangered (Table 1 &Table 2). Global temperatures have risen dramatically in recent years, with the frequency and duration of extreme heat events expected to continue increasing. The Snowy Owl has less and less thermal refugia, which could allow it to escape the extreme heat and adapt more readily to temperature shift [46]. In the Russian Arctic, the increasing June temperatures are already seen to be responsible for the long-term decline in the breeding effort of Snowy Owls [47].
3.1. Microevolution
In many species, markedly different colour morphs exist, and the frequency of these is sometimes climate-related, the pheomelanic reddish-brown morph (hereafter rufous or brown morph) predominates in warm climates while less melanic grey morph dominates in a less humid and colder environment [48]. One polymorphic example is the Eastern Screech Owl Megascops asio, individuals of which exhibit rufous, intermediate, or grey coloration. This species exhibits clinal variation in morph prevalence; rufous Eastern Screech Owls are relatively scarce in northern areas because it appears that they suffer greater mortality than grey ones do during severe winter cold snaps. It has also been noted that rufous females are likely to survive cold spells better than rufous-coloured males [49]. In Europe, skin collections of Eurasian Scops Owls Otus scops, spanning a 137-year period, show a significant increase in the rufous morph corresponding to an increase in rainfall, humidity and temperature over that time [50].
In Finland, numbers of brown and grey morphs of Tawny Owls are also climate-related, as the grey morphs have a denser and more insulative plumage, enabling them to survive better in a cold climate compared to brown ones [51]. The brown morph consistently moults more primary flight feathers than the grey morph [52]. This is interesting when we know that the plumage moult is a costly and crucial somatic maintenance function in owls [53]. The two morphs differ also in several life-history aspects related to immune defence against parasites and somatic maintenance costs, possibly explaining their different sensitivity to winter conditions [54]. Over the last three decades, the warming of the winter climate has produced a microevolutionary response in morph frequency: the brown morph has increased in number at the expense of the grey [55]. Thus, colour polymorphism in owls is an adaptive character likely maintained by the selective advantage of camouflage under different light regimes or in terms of physiological adaptation to environmental conditions via disruptive selection mechanisms. Under this hypothesis, climate change could bring about a dramatic change in the colour polymorphism of some northern species [55,56]. It has already been noted with Tawny Owls in Finland that in winter the grey phase helps avoid avian mobbing and predators more efficiently than the brown morph and therefore has a higher survival rate in snowy environments [57]. However, as winters are getting milder and shorter in this species range due to climate change, the selection periods promoting grey colouration may eventually disappear or shift northward [54,57]. However, the Tawny Owl in Finland offers one of the first evidences that recent climate change can alter natural selection in a wild population leading to a microevolutionary response, which demonstrates the ability of wild populations to evolve in response to climate change.
Plumage traits of Barn Owls have been proven to be strongly heritable and on a continental scale, climatic factors are associated with plumage traits [58]. Western Barn Owls in continental Europe and in the British Isles have larger spots in the colder north-east regions and smaller spots in the warmer southern regions. American Barn Owls Tyto furcata are also displaying larger black feather spots in regions where ambient temperatures were colder. Female spottiness is shown to signal parasite resistance [59].
3.2. Wildfires
Climate change creates warmer and drier conditions globally, lengthening fire seasons and increasing the frequency and severity of wildfires. The severity of forest fires is measured in terms of low-density, mid-density, and high-density, with especially severe high-density fires now being classed as megafires. Global forests are key ecosystem indicators, used to monitor a wide range of ecological factors from climate regulation to biodiversity habitat. Today, forests are under increasing pressure from the combined impacts of climate and land use change. In 2024, global forest fires burned a record-breaking 13.5 million hectares of forest worldwide, surpassing the loss of forests to agricultural use globally for the first time [60].
The immediate effect of a high-density forest fire on forest-dwelling owls is devastating, with the initial loss of habitat, nesting sites, and prey species. However, studies have shown that any long-term negative effect varies from species to species, depending on which ecological niches they occupy within the forest area and on the severity of the fire [61]. In the USA, one species, the Northern Spotted Owl Strix occidentalis caurina, already endangered by past commercial logging and competition from the much larger Barred Owl Strix varia, relies on closed canopy old growth forests and is especially vulnerable to forest fires [62], with one recent study showing that Spotted Owls avoid high-severity burned sites for up to two decades after a fire [61].
Recent studies have also shown that, amongst other forest owl species, the Northern Saw-whet Owl Aegolius acadicus also appears to avoid nearly all burned areas. However, Barred Owls and Great Horned Owls Bubo virginianus have been found to use unburned and low-severity burned areas compared to areas that burned more severely, whereas Northern Pygmy Owls Glaucidium gnoma and Western Screech Owls Megascops kennicottii have shown high use of high-severity burned areas [61 and 63].
While some owl species, like the Spotted Owl, are extremely vulnerable to both immediate and long-term negative effects from forest fires, some species, such as the Great Grey Owl, although impacted by the immediate effect of loss of nesting habitat and prey species, are showing some short-term resilience. A study has shown that after one megafire in California’s Sierra Nevada, many Great Grey Owls remained within the burned area and continued to nest in the newly created snags and open meadows. As Great Grey Owls typically nest in dead trees, fire would kill off large trees, potentially creating new nesting sites, while the newly created meadows would help in the recovery of their meadow-dwelling rodent prey [64].
3.3. Precipitation & Flooding
Global rainfall in 2025 has shown regional variation, with forecasts of above-normal precipitation in the Indian subcontinent, parts of eastern Asia, the Maritime Continent (Southeast Asia), eastern Australia, northwestern South America, northwestern North America, and interior eastern equatorial Africa [2]. The rise in global average temperatures is a key driver of changes in rainfall patterns as a hotter planet leads to more intense rainfall events and a higher risk of flooding [65].
Continuous heavy rainfall is a serious threat to owls, especially to species such as the Western Barn Owl Tyto alba, as their soft, lightly oiled feathers can quickly become waterlogged, making their silent flight extremely difficult and making their ability to hunt by hearing impossible. This inability to hunt effectively can lead to the starvation of both adult owls and chicks, ultimately impacting their ability to breed successfully [66].
Long periods of torrential rain can also lead to flooding in certain circumstances, thus causing additional problems, such as fast-rising river levels bursting their banks and flooding low-lying areas, negatively impacting the habitat of small mammal assemblages and further reducing the food availability for the owl and forcing them to find alternative prey [67].
Ground nesting species, such as the Short-eared Owl, are, of course, vulnerable to extreme rainfall events, with one species in particular, the Burrowing Owl Athene cunicularia, being especially vulnerable. A recent study from Canada recorded that within the study area, Burrowing Owl nest and owlet survival markedly declined due to extreme precipitation and burrow flooding. As the heavy rainfall occurred late in the season, giving the owls little chance of re-nesting, the extreme precipitation resulted in a complete loss of annual reproductive output for the pairs that failed [68].
3.4. Rising Sea Level
Sea levels rise due to climate change. This will reduce coastal wintering habitats of Short-eared Owl as some wintering locations will be lost to the sea and saltification may affect some of the remaining habitats. This would increase competition and reduce the suitability of coastal wintering grounds, reducing owls’ wintering survival and breeding condition upon return from migration [14].
3.5. Drought
Climate change intensifies droughts, making them more frequent, longer, and more severe. As average temperatures have risen because of climate change, higher temperatures dry out soils and vegetation, strain water supplies, and reduce snowpack. Historically dry areas are more likely to see reduced precipitation, and total land area subject to drought is projected to increase.
The effect of drought on prey selection of the Eastern Barn Owl Tyto javanica was studied in north-eastern South Australia. Prolonged dry periods have made Barn Owls predominantly prey on Geckos and other reptiles when small mammals became less available [69]. The owl being able to switch to a primarily reptilian diet may explain why Barn Owls sometimes remain faithful to a single area, and even a single roost/breeding site, for generations.
Owls often have a particularly difficult time during periods of drought. Many owls, like the Barking Owl Ninox connivens in Australia, are very territorial and don’t move to new locations easily. Some breeding pairs seem to stay and defend their patch at all costs. The coincidence of both drought and bushfires has become a ‘double whammy’ [70].
The effects of increasing air temperature and aridity on a Burrowing Owl population have been studied from 1998-2013 in central New Mexico, US [16]. Decreased precipitation and rapid warming caused a multiyear drought, and over a period of 16 years, the owl population declined from 52 pairs to 1 pair. Nest success and productivity declined after an unexpected delay in breeding phenology, most likely causing a mismatch with food resources. Significant changes in body mass in both breeders and nestlings reflect poor breeding as well as wintering habitat quality [16]. Further studies indicated that delays are due to food limitation caused by drought. Drought during migration may also constrain energetic requirements, forcing Burrowing Owls to stop more frequently and for longer periods at stopover sites, resulting in delayed arrival on breeding grounds [17].
3.6. Winter and Snow
Northern Europe has been suffering increasingly warm and wet winters associated with long-term changes in the North Atlantic Oscillation [71]. Shifts in precipitation frequency, intensity, and quantity serve as significant indicators of climate change. There has been a decrease in the frequency alongside an increase in intensity of precipitation [72]. Insects that undergo winter dormancy in soil are directly influenced by concurrent rainfall. Essentially, heavy rainfall can cause flooding and prolonged water stagnation, posing a threat to insect survival and potentially disrupting their dormancy patterns. Also, insect eggs and larvae may be washed away due to heavy rains and flooding [72].
The warming climate in Fennoscandia has induced the reduction and duration of the subnivean space, which has had a negative impact on the survival and reproductive performance of northern rodents, consequently altering their population cycles [73]. When available, the subnivean space provides thermal insulation, access to food plants, and protection from owls and raptors, and other generalist predators. Extension of the available subnivean space has been shown to increase winter survival of the Root Vole Microtus oeconomus [74]. Increasingly warm late winter/early spring and snow conditions are seen as a reason to observe the absence of rodent peak years since 1994. This has led to dramatic declines in northern owls [75], also noted in this study (Table 1).
Climate change impairs owls’ foraging and thus decreases local overwinter survival. The Eurasian Pygmy Owl normally hoards food (small mammals and birds) in nest-boxes for winter, but now this hoarding behaviour is highly susceptible to global warming. In several northern areas, autumns have become warmer and winters milder and rainier. This is likely to continue, ultimately decreasing the length of winter. The more rainy days there are between mid-October and mid-December, the more likely it is that the food hoards of the owl go rotten. Especially during the poor vole years, owls are forced to use rotten food, meaning that the owl either dies or is forced to leave the area [76]. The future will show if the Pygmy Owls are able to adapt to climate change by delaying food hoarding or, more likely, they will suffer further due to the changes caused by the autumn/winter warming.
During the winter, it is normal for Great Grey Owls to move south in search of food. Although literature gives the impression that this is mainly a forest owl, it is by no means confined to the forest when it wanders south in winter. Lately, every winter, Great Grey Owls have entered the capital city, Helsinki, where BirdLife Finland decided to ban publishing the photos, as so many birdwatchers approached the owl, giving it no peace to feed itself [77]. Despite apparently unfavourable climate changes for boreal species, the Great Grey Owl has been expanding its range in Europe to the south and west. Especially in Sweden and Norway, the southward shift in recent decades has been almost total, coupled with drastic declines in the north of their range. For a number of years, no Great Grey Owl nests were found in Norwegian Lapland, but nowadays as many as 140 nests are found annually in Hedmark county, SE Norway, below 62°N. In Sweden, most southern nests occur even below 58°N. In Europe, the most southern nests are in Ukraine and Poland around 51°N, while in North America, it breeds as far south as 37-38°N, which in Europe would take it down to Sicily [78]. It is interesting to note that during the Pleistocene Period (3 million to 10,000 years ago), the Great Grey Owl inhabited areas as far south as Bulgaria and Romania according to fossil findings [79]. Table 1 shows that recently the Great Grey Owl population in Finland has increased by some 8.5%. It seems that the Central Finland population has moved to North Finland (C ‒24% and N + 23.1%). South Finland has gained 0.9%, but no similar movement to the south has happened as in Norway and Sweden [39 and 40].
3.7. Snow Structure
Boreal owl species, wintering in Fennoscandia, are dependent in their winter hunting not only on snow depth, but of the hardness and ice structures in the snow carpet as well [80]. Several abiotic processes can make significant changes to a snow cover, affecting owl hunting success. One important factor might be sudden melting periods or rain episodes followed by cold weather, forming crusts. Repeated cycles of freezing and thawing, named as “frost seesaw effect” [81], create through the winter several ice layers which are buried into the snow layer by new snowfalls [82]. Cold wind alone often hardens the snow cover, especially in higher elevations, making wind the most significant abiotic factor imposing extreme hardness on snow covers in alpine and arctic areas of Fennoscandia [80]. How vulnerable boreal owl species are to snow hardness is not well known, but climate change definitely has changed wind regimes, increasing the frost seesaw effect [81]. This effect is expected to be more significant in the cold and continental areas, lacking more frequent fluxes from maritime influence. Mild winters at least allow the Ural Owl to breed earlier [29]. Also, the Tawny Owl benefits from increasing late winter or early spring temperatures, advancing its breeding at least as much as does high autumn abundance of voles. However, breeding too early or too late may decrease reproductive output if the timing of breeding does not meet the food peak [83].
A dramatic Western Barn Owl population decline has been projected in Switzerland under a scenario of non-linear relationships between climate change and population vital rates [43]. Adult and juvenile owl survival plummeted due to the extended snow cover duration, recorded only twice in a 58-year study period. Under the scenario of non-linear climate change, the extreme snow-rich winters can increase, reducing small mammal prey availability and owls’ long-term growth rates [43].
3.8. Habitat Loss
Climate change habitat models forecast severe, region-wide breeding habitat losses for most montane owls in the Southwestern US by 2090 [10]. The steepest rates of habitat loss were predicted for the Aegolius funereus habitat distribution, which will only consist of isolated areas, perhaps too small to sustain the regional persistence of the species. The near-complete loss of current breeding habitats is also projected for known Flammulated Owl Psiloscops flammeolus populations in Western New Mexico. However, restoration of more open stands of ponderosa pine and mixed conifer forest through reduction of stand densities would benefit the Flammulated Owl [27].
Three more species, the Long-eared Owl, Northern Pygmy Owl, and Northern Saw-Whet Owl, were predicted to lose at least 60% of their current breeding habitats. The Whiskered Screech Owl Megascops trichopsis was forecasted to lose all its current habitat and would only persist in the region if it could track areas that become suitable to the north of its current range. More uncertainty exists for the Great Horned Owl and the Western Screech Owl, both with distributions that extend down to lower elevations [10].
Logging patches of forest was assumed to provide Great Grey Owl foraging areas by creating meadows, but a new study shows that relatively dry openings created by clearings and regeneration harvests are unlikely to provide preferred, long-term foraging habitat [84]. Like for many other species, conserving mature forests, including riparian forests, is critical [85 and 86].
3.9. Climate-Related Nestbox Competition
Climate change is altering the strength of interspecific interactions as rising temperatures are encouraging nestbox competition. In southern Hungary, there are sympatric breeding populations of Western Barn Owls (hereafter Barn Owl) and Tawny Owls co-occurring in the same nestbox during the same breeding season [87]. The onset of Tawny Owl breeding has not changed during the study period, but that of Barn Owl advanced by two weeks in parallel with rising temperatures. Thus, the breeding season has shifted closer to the breeding season of the Tawny Owl. When using the same nestbox, the breeding of Barn Owls was delayed by a month, and second clutches were practically absent. Climate change heightens competition for nest-sites between Barn and Tawny Owls [87]. As the preferred nestboxes are limited, the Barn Owl faces greater disadvantages in this competition with the heavier and more aggressive Tawny Owl; able to kill Barn Owls, at least the young ones [88]. A significant decline in reproductive success of Barn Owls happens due to the absence of second broods [87]. Barn Owls can also exert negative effects on the Tawny Owls by killing their chicks [89].
3.10. The Effect of Climate Change on Prey Species
Small mammals such as rodents and shrews predominate as food items for the majority of larger owls, and insects and other arthropods for the smaller ones. Many species supplement their diet by eating other birds, reptiles, amphibians, fish, crabs, and earthworms. Of the world’s 273 owl species, 40% are mainly insectivorous, 31% are carnivorous, and 3% are piscivorous. The diet and hunting behaviour of the remaining 26% are still unknown [90].
- Fish
Semi-aquatic fish-eating owls use their talons to seize fish from the surface of rivers, or they hunt crabs on shores and in river shallows. Eastern Screech Owls hunting both early and late in the evening tended to hunt for crayfish in the evening (18‒23h) and small mammals later at night (23‒03h). In global terms, there are only nine mainly piscivorous species, 14 species have been recorded to eat occasionally fish, or crustaceans, 11 species, or both, 17 species [90]. Extreme climatic events include unpredictable rainfall patterns causing both droughts and floods, which impact fish-eating owls. Serious drought can force owls to seek new areas with water in ponds and rivers, and flooding makes it difficult for owls to locate fish in deep, muddy waters.
- Insects
Climate change is pounding insects in a wide variety of ways all over the world. It impacts insects by altering habitats, shifting suitable climatic areas, and increasing extreme weather events like heatwaves and droughts, which can increase population mortality, species range shifts, and changes in community composition. Many species are relocating to poleward places with lower temperatures or to higher altitudes. It has been estimated that up to 49% of insects will lose more than half of their historical geographical distribution [91].
In Norway, the number of insects has decreased by 14 per cent over the past four years, from 2020 to 2023. The weather change seems to affect them, and understandably, insects thrive in warm and not too dry summers. More surprising is that cold winters with lots of snow have a positive effect on insects. A thick snow cover insulates well, but climate change brings more dry and mild winters with less snow, and therefore has negative effects on insects in the future [92]. Early spring can cause insects to become active a bit too early, as the return to more winter-like conditions impacts their survival badly.
- Small mammals
The impact of climate change on small mammals can be significant, affecting them directly as well as indirectly. Extreme weather events like droughts and heatwaves can lead to population decline. The low dispersal ability and small body size make them particularly vulnerable. Very few species are able to relocate to areas with more favourable, cooler temperatures and face challenges in highly fragmented landscapes, often resulting from human activities, to find new suitable habitats. Changes in species distribution and population dynamics lead to new inter-specific interactions, including increased competition and changes in community composition [93‒95].
Many owl species are known to have high site tenacity with limited ability to migrate to new areas if their existing habitat becomes unsuitable or after their regular food supply has disappeared. Stochastic demographic modeling predicted that already visible dampening vole cycles could be driving the Northern England Tawny Owl population towards extinction [28]. Western Barn Owl diet analysis in multiple locations in Italy demonstrated a consistent increase of xerothermophilic species (living in hot and dry places) in the small mammal communities since 70s [96]. Also in Spain, the small mammal diversity has declined with time due to climate change [97]. Such a reduction in diversity and abundance of small mammals is alarming for place faithful owl species like Tyto alba, among others, whose diet, breeding success, and other ecological parameters depend on small mammals’ abundance [97].
Nomadic and migratory owls have better chances to relocate to cooler temperatures, but unfortunately, their main prey animals, small mammals, are less able to move to new cooler areas at higher altitudes or poleward.
4. Discussion
Drastic owl population declines are usually attributed to multiple sources, such as the global climate change and habitat loss and degradation, inducing decreased food supply [98]. However, only in the case of relatively few owl species has the interactive effects of fluctuations in abundance of main foods and weather conditions on population densities and reproductive success been well studied. In the case of Tengmalm’s Owl milder and more humid spring and early summer temperatures due to global warming are not able to compensate for the lowered offspring production of owls [99]. The main reason for low productivity is probably loss and degradation of mature and old-growth forests due to clear-felling, which results in loss of coverage of prime habitat for main (Bank Voles Myodes glareolus) and alternative prey (small birds) of owls, inducing a lack of food, and refuges against predators of Tengmalm’s Owls [99].
Some birds have advanced their breeding or migration phenology, and northward density shifts have been observed in Fennoscandian species [100], but not so much in the case of owls. Contrary to this, the Great Grey Owl has moved rather to the south and southwest in Norway and Sweden [39 and 40]. Only the southern Eurasian Scops Owl has shown an opposite tendency to expand its distribution to the north [26].
In Finland, there are some indications that Short-eared Owls Tengmalm’s Owls Great Grey Owl, and Northern Hawk Owls have increased their nesting in the north. However, climate change has not yet assisted southern species, such asTawny and Long-eared Owls, in moving further north in Finland. Eurasian Eagle Owls and Pygmy Owls show only minor shifts between south, central, and north. So, the response of one species to climate change can hardly be used to predict the response of another [101].
Therefore, studying survival and distribution in an apex predator, like the Great Grey owl, requires individual-based data from long-term studies and is complicated by the integration of climatic effects on prey species at lower trophic levels [36 and 37]. According to Mysterud [80], not only are field studies of raptor distribution extensions that take into account the food factor needed, but simultaneous snow-cover studies are very much needed as well. Among the most important ecological winter factors might be the frost seesaw [81]; that is, alternating thaw and freezing cycles and related ice formations blocking hunting and decreasing the food factor.
Tawny Owl and Barn Owl studies have shown that climate change can alter natural selection in a wild population leading to a microevolutionary response. This demonstrates the ability of wild populations to evolve in response to climate change. Even if a species could be able to adapt slowly to climate change, the speed of change has been so fast that a negative impact is unavoidable. This assumption disregards the owl species’ capacity to persist through plasticity or shift their distribution to other regions when confronted with novel environmental conditions. It is also possible that climate change itself could take a new direction at least in the northern hemisphere should the Gulf stream collapse. There are signs that rotating ocean currents south of Greenland have been losing stability since the1950s. This would bring harsh, freezing cold winters back to Europe and North America [102‒104], but could it bring Snowy Owls back remains to be seen.
References
- Dietz, T., Shwom, R. L. & Whitley, C. T. (2020). Climate Change and Society. Annual Review of Sociology 46:135-158. [CrossRef]
- WMO: https://wmo.int/.
- Sattar, Q., Maqbool, M. E., Ehsan, R. & Akhtar, S. (2021). Review on climate change and its effect on wildlife and ecosystem. Open Journal of Environmental Biology 6(1): 008-014. [CrossRef]
- Sieradzki, A. 2022. Designed for Darkness: The Unique Physiology and Anatomy of Owls. Chapter 1: 3‒26. In: Mikkola, H. (ed.), Owls: Clever Survivors. IntechOpen, London. [CrossRef]
- Pautasso, M. (2012). Observed impacts of climate change on terrestrial birds in Europe: an overview. Italian Journal of Zoology 79(2): 296‒314. [CrossRef]
- Vegvari, Z., Bokony, V., Barta, Z. & Kovacs, G. (2010).Life history predicts advancement ofavian spring migration in response to climate change. Global Change Biology 16: 1‒11.
- BirdLife Finland, https://lintulehti.birdlife.fi.
- Virkkala, R., Heikkinen, R. K., Leikola, N. & Luoto, M. (2008). Projected large-scale range reductions of northern-boreal land bird species due to climate change. Biological Conservation 141: 1343-1353. [CrossRef]
- Honkala, J., Lehikoinen, P., Saurola, P. & Valkama, J. (2025). Breeding and population trends of common raptors and owls in Finland in 2024. Linnut-vuosikirja 2024: 64‒79. (in Finnish, English summary).
- Cartron, J.-L. E., Triepke, F.J., Stahlecker, D.W., Arsenault, D.P., Ganey, J.L., Hathcock, C.D., Thompson, H.K., Cartron, M.C. & Calhoun, K.C. (2023). Climate Change Habitat Model Forecasts for Eight Owl Species in the Southwestern US. Animals, 13, 3770. [CrossRef]
- Kopij, G. (2011). Population and range expansion of forest boreal owl (Glaucidium passerinum, Aegolius funereus, Strix uralensis, Strix nebulosa) in East-Central Europe. Vogelwelt 132: 93‒100.
- Kasprzyk, K. & Frątezak, W. (2025). Diet of solitary males Boreal Owls Aegolius funereus in the lowlands of the Bydgoszcz Forest (northern Poland). Ecological Questions 36(2): 1‒15. [CrossRef]
- Booms, T.L., Holroyd, G.L., Gahbauer, M.A., Trefry, H.E., Wiggins, D.A., Holt, D.W., Johnson, J.A., Lewis, S.B., Larson, M.D., Keyes, K.L. & Swengel, S. (2013). Assessing the Status and Conservation Priorities of the Short-eared Owl in North America. The Journal of Wildlife Management 9999. [CrossRef]
- Peak District National Park Authority (2025). Feature Assessment: Wildlife/Short-eared Owl. Reports.peakdistrict.gov.uk/cova/docs/assessment/wildlife/owl.html Accessed 18.09.2025.
- Penagos-López, A.P. & Esquivel Melo (2025). Impact of climate change on Andean owls in Colombia. Ornitologia Colombiana 27(i):49.
- Cruz-McDonnell, K. (2015). Negative effects of rapid warming and drought on reproductive dynamics and population size of an avian predator in the arid southwest. M.Sc. Thesis in the University of New Mexico, Albuquerque, New Mexico. https://digitalrepository.unm.edu/biol_etds/21.
- Porro, C.M., Desmond, M.J., Savidge, J.A., Abadi, F., Cruz-McDonnell, K.K., Davis, J.L., Griebel, R.L., Ekstein, R.T. & Rodríguez, N.H. (2020). Burrowing Owl (Athene cunicularia) nest phenology influenced by drought on nonbreeding grounds. The Auk, 137: 1‒17. [CrossRef]
- Lehtiniemi, T. (2025). The occurrence of some threatened and fairly rare bird species in Finland in 2024. Linnut vuosikirja 2024: 80‒89. (In Finnish, English summary).
- McCabe, R.A., Aarvak, T., Aebischer, A., Bates, K., Bety,J., Bollache, L., Brinker, D., Driscoll. C., Elliott, K.H., Fitzgerald, G., Fuller, M., Gauthier, G., Gilg, O., Gousy-Leblanc, M., Holt, D., Jacobsen, K-O., Johnson, D., Kulikova, O., Lang, J., Lecomte, N., McClure, C., McDonald, T., Menyushina, I., Miller, E., Morozov, V.V., Øien, I.J., Robillard, A., Rolek, B., Sittler, B., Smith, N., Sokolov, A., Sokolova, N., Solheim, R., Soloviev, M., Stoffel, M., Weidensaul, S., Wiebe, K.L., Zazelenchuck, D. &Therrien, J.F. (2024). Status assessment and conservation priorities for a circumpolar raptor: the Snowy Owl Bubo scandiacus. Bird Conservation International 34, e41: 1‒11. [CrossRef]
- Domine, F., Gauthier, G., Vionnet, V., Fauteux, D., Dumont, M. & Barrère, M. (2018). Snow physical properties may be a significant determinant of lemming population dynamics in the high Arctic. Arctic Science 4: 813‒826. [CrossRef]
- Lamarre, V., Legagneux, P., Franke, A., Casajus, N., Currie, D.C., Berteaux, D. et al. (2018). Precipitation and ectoparasitism reduce reproductive success in an arctic-nesting top-predator. Scientific Reports 8, 8530. [CrossRef]
- Solheim, R., Jacobsen, K.-O., Øien, I.J., Aarvak, T. & Polojärvi, P. (2013). Snowy Owl nest failures caused by blackfly attacks on incubating females. Ornis Norvegica 36: 1‒5. [CrossRef]
- Penagos-López, A.P., Jiménez García, D. & Carlos, C.J. (2025). Protected areas as key to identifying and prioritizing climate refugia for the conservation of endemic Atlantic Forest owls. Ornitologia Colombiana 27(i): 53.
- Masoero, G. (2020). Food hoarding of an avian predator under food limitation and climate change. Ph.D. thesis, Annales Universitatis Turkuensis, Finland. Ser AII, Tom. 373.
- Class, B., Masoero, G., Terraube, J. & Korpimäki, E. (2021). Estimating the long-term repeatability of food-hoarding behaviours in an avian predator. Biology Letters 17: 202102286. [CrossRef]
- Martin, J.R. & Mikkola, H. (2019). Distribution trends of the Scops Owl in Britain and northern Europe. Scottish Birds 39(2): 168‒172.
- McKelvey, K. S. & Buotte, P. C. (2018). Effects of Climate Change on Wildlife in the Northern Rockies. In Halofsky, J. E. & Peterson, D. L. [Eds.], Climate Change and Rocky Mountain Ecosystems. Advances in Global Change Research 63: 143‒167. [CrossRef]
- Millon, A., Petty, S.J., Little, B., Gimenez, O., Cornulier, T. & Lambin, X. (2014). Dampening prey cycle overrides the impact of climate change on predator population dynamics: a long-term demographic study on tawny owls. Global Change Biology 20: 1770‒1781. [CrossRef]
- Lehikoinen, A., Ranta, E., Pietiäinen, H., Byholm, P., Saurola, P., Valkama, J., Huitu, O., Henttonen, H., and Korpimäki, E. (2011). The impact of climate and cyclic food abundance on the timing of breeding and brood size in four boreal owl species. Oecologia 165: 349‒355.
- Comay, O., Ezov, E., Yom-Tov, Y. & Dayan, T. (2022). In Its Southern Edge of Distribution, the Tawny Owl (Strix aluco) Is More Sensitive to Extreme Temperatures Than to Rural Development. Animals 12, 641. [CrossRef]
- Peery, M.Z., Guttiérez, R.J., Kirby, R., Ledee, O.E. & Layahe, W. (2011). Climate change and spotted owls: Potentially contrasting responses in the Southwestern United States. Global Change Biology 18: 865‒880.
- Berg, T., Solheim, R., Wernberg, T. & Østby, E. (2011). Lappuglene kom! [Great Grey Owls come! ]. Vår Fuglefauna 34(3): 108‒115. (in Norwegian).
- Keller, M., Chodkiewicz, T. & Woźniak, B. (2011). Great Grey Owl Strix nebulosa ‒ a new breeding species in Poland. Ornis Polonica 52: 150‒158. (In Polish, English summary).
- Ławicki, L., Abramčuk, A.V., Domashevsky, S.V., Paal, U., Solheim, R., Chodkiewicz, T. & Woźniak, B. (2013). Range extension of Great Grey Owl in Europe. Dutch Birding 35: 145‒154.
- Mikkola, H. (2010). Most Southern Great Grey Owl. Tyto; The International Owl Society XIII (4):8‒9.
- Mikkola, H. (2014a). Global Warming and Great Grey Owls. Tyto; The International Owl Society March 2014: 7‒8.
- Mikkola, H. (2014b). Der Einfluss der Erderwärmung auf die Ausbreitung von Bartkäuzen. Kauzbrief 26:22‒23. (in German).
- Mirski, P., Ivanov, A., Kitel, T. & Tumiel, T. (2021). The ranging behaviour of the Great Grey Owl Strix nebulosa: a pilot study using GPS tracking on a nocturnal species. Bird Study. [CrossRef]
- Solheim, R. (2009). Lappugla ‒ en klimaflyktning på vei sydover? [Great Grey Owl ‒ a climate fugitive on the way south?]. Vår Fuglefauna 32: 164‒169 (in Norwegian).
- Solheim, R. (2014). Lappugglan på frammarsch [Great Grey Owl on the march forward]. Vår Fågelvärld 73: 46‒50 (in Swedish).
- Vermouzek, Z., Krenek,D., Czerneková, B. (2005): Increase in numbers of Ural Owls (Strix uralensis) in the Beskydy Mts. (NE Czech Republic). Sylvia 40: 151-155.
- Šotnár, K. (2005): Nesting, diet and expansion of the Ural Owl (Strix uralensis) in the area of Horné Ponitrie (Central Slovakia). Buteo 14: 67‒68.
- Jenouvrier, S. (2013). Impacts of climate change on avian populations. Global Change Biology 19: 2036‒2057.
- Gilg, O., Kovacs, K.M., Aars, J., Fort, J., Gauthier, G., Grémillet, D., Ims, R.A., Meltofte, H., Moreau, J., Post, E., Schmidt, N.M., Yannic, G. & Bollache, L. (2012). Climate change and the ecology and evolution of Arctic vertebrates. Annals of the New York Academy of Science 1249: 166‒190. [CrossRef]
- Solheim, R., Jacobsen, K-O. & Øien, I.J. (2008). Snøuglenes vandringer: Ett år, tre ugler og ny kunnskap [Snowy Owl movements: one year, three owls and new knowledge]. Vår Fuglefauna 31(3): 102‒109. (In Norwegian).
- Mikkola, H. (2022a). Gufo delle nevi Bubo scandiacus. Pp. 192‒203, 391. In: Cauli, F., Galeotti, P. & Genero, F. (Eds.) Rapaci d’Italia e d’Europa. Edizioni Belvedere, Latina. (in Italian).
- Morozov, V.V., Rosenfeld, S.B., Rogova, N.V., Golovnyuk, V.V., Kirtaev, G.V. & Kharitonov, S.P. (2020). What is the number of snowy owls in the Russian Arctic? Ornithologia 44: 18‒25. (In Russian, English summary).
- Gasparini, J., Bize, P., Piault, R., Wakamatsu, K., Blount, J.D, Ducrest, A.L. & Roulin, A. (2009). Strength and cost of an induced immune response are associated with heritable melanin-based colour trait in female tawny owls. Journal of Animal Ecology 78: 608‒616.
- Gehlbach, F.R. (2012). Eastern screech-owl responses to suburban sprawl, warmer climate, and additional avian food in central Texas. Wilson Journal of Ornithology 124(3): 630‒633.
- Weidensaul, S. (2015). Owls of North America and The Caribbean. 333 p. Peterson Field Guides, New York.
- Koskenpato, K., Ahola, K., Karstinen, T. & Karell, P. (2016). Is the denser contour feather structure in pale grey than in pheomelanic brown tawny owls Strix aluco an adaptation to cold environments? Journal of Avian Biology 47(1): 1‒6. [CrossRef]
- Karell, P., Brommer, J.E., Ahola, K. & Karstinen, T. (2013). Brown tawny owls moult more flight feathers than grey ones. Journal of Avian Biology 44(3): 235‒244. [CrossRef]
- Mikkola, H. & Lamminmäki, J. (2014). Moult, ageing and sexing of Finnish Owls. 98 p. BirdLife Suomenselkä (in Finnish, English summary).
- Karell, P., Bensch, S., Ahola, K. & Asghar, M. (2017). Pale and dark morphs of tawny owls show different patterns of telomere dynamics in relation to disease status. – Proceedings of the Royal Society B: Biological Sciences 284 (1859): 1-8. [CrossRef]
- Karell, P., Ahola, K., Karstinen, T., Valkama, J. & Brommer, J.E. (2011). Climate change drives microevolution in a wild bird. Nature Communications 2, 208. [CrossRef]
- Brommer, J.E., Ahola, K. & Karstinen, T. (2005). The colour of fitness: Plumage coloration and lifetime reproductive success in the tawny owl. Proceedings of the Royal Society B272, 1566: 935‒940.
- Koskenpato, K., Lehikoinen, A., Lindstedt, C. & Karell, P. (2020). Gray plumage color is more cryptic than brown in snowy landscapes in a resident color polymorphic bird. Ecology and Evolution 10: 1751‒1761. [CrossRef]
- Roulin, A. (2014). Melanin-based colour polymorphism responding to climate change. Global Change Biology 20: 3344–3350. [CrossRef]
- Roulin, A., Riols, C., Dijkstra, C. & Ducrest, A.L. (2001). Female plumage spottiness signals parasite resistance in the barn owl (Tyto alba). Behavioral Ecology 12(1): 103‒110.
- Potapova, P., Tyukavina, A., Turubanova, S., Hansen, M. C., Giglio, L., Hernandez-Serna, A., Lima, A., Harris, N. & Stolleb, F. 2025. Unprecedentedly high global forest disturbance due to fire in 2023 and 2024. PNAS 122(30), e2505418122. [CrossRef]
- McGinn, K., Zuckerberg, B., Jones, G. M., Wood, C. M., Kahl, S., Kelly, K. G., Whitmore, S. A., Kramer, H. A., Barry, J. M., Ng, E. & Peery, M. Z. (2025). Frequent, heterogenous fire supports a forest owl assemblage. Ecological Applications. 2025;35:e3080. [CrossRef]
- Lesmeister, D.B., Davis, R.J., Sovern, S.G. & Yang, Z. (2021). Northern spotted owl nesting forests as fire refugia: a 30-year synthesis of large wildfires. Fire Ecology 17:32. [CrossRef]
- Duchac, L. S., Lesmeister, D. B., Dugger, K. M. & Davis, R. J. (2021). Differential landscape use by forest owls two years after a mixed-severity wildfire. Ecosphere 12(10): e03770. [CrossRef]
- Siegel, R. B., Eyes, S. A., Tingley, M. W., Wu, J. X., Stock, S. L., Medley, J. R., Kalinowski, R. S., Casas, A., Lima-Baumbach, M. & Rich, A. C. 2019.Short-term resilience of Great Gray Owls to a megafire in California, USA. The Condor, 121: 1-13. [CrossRef]
- metoffice.gov.uk.
- Barn Owl Trust: www.barnowltrust.org.uk/.
- Avotins, A., Avotins, A. Sr., Kerus, V. & Aunins, A. 2023.Numerical Response of Owls to the Dampening of Small Mammal Population Cycles in Latvia. Life, 2023, 13, 572. [CrossRef]
- Fisher, R. J., Wellicome, T. I., Bayne, E. M., Poulin, R. G., Todd, L. D. & Ford, A. T. (2015). Extreme precipitation reduces reproductive output of an endangered raptor. Journal of Applied Ecology 52: 1500‒1508. [CrossRef]
- McDowell, M.C. & Medlin, G.C. (2009). The effects of drought on prey selection of the barn owl (Tyto alba) in the Strzelecki Regional Reserve, north-eastern South Australia. Australian Mammalogy 31: 47‒55. [CrossRef]
- Beemster, M. (2004). Drought and fires threaten the Barking Owls. https://science-health.csu.edu.au/herbarium/woodland-web/barking-owl Accessed18.09.2025.
- Steenseth, N.C., Mysterud, A., Ottersen, G., Hurrell, J.W., Chan, K-S.& Lima, M. (2002). Ecological effects of climate fluctuations. Science 297: 1292‒1296.
- John, A., Riat, A.K., Bhat, K.A., Ganie, S.A., Endarto, O., Nugroho, C., Handoko, H. & Wani, A.K. (2024). Adapting to climate extremes: Implications for insect populations and sustainable solutions. Journal for Nature Conservation 79: 1‒10. [CrossRef]
- Kausrud, K.L., Mysterud, A., Steen, H., Vik, J.O., Østbye et al. (2008). Linking climate change in lemming cycles. Nature 456: 93‒97.
- Korslund, L. & Steen, H. (2006). Small rodent winter survival: snow conditions limit access to food resources. Journal of Animal Ecology 75: 156‒166.
- Ims, R.A., Henden, J.A. & Killengreen, S.T. (2008). Collapsing population cycles. Trends of Ecological Evolution 23: 79‒86.
- Armarego-Marriott, T. (2020). Owls’ hoards rot. Nature Climate Change 10:802. [CrossRef]
- Marttinen, V. (2024). Lapinpöllö lensi Helsinkiin ja someraivo räjähti liekkeihin ‒ lintujärjestö joutui kieltämään pöllökuvien julkaisemisen. https:yle.fi/a/74-20078977?utm_medium=social&utm_source=ema… Accessed 14.03.2024 (in Finnish).
- Mikkola, H. (2022b). Allocco di Lapponia Strix nebulosa. Pp. 240‒251, 395‒396. In: Cauli, F., Galeotti, P. & Genero, F. (Eds.) Rapaci d’Italia e d’Europa. Edizione Belvedere, Latina. (in Italian).
- Boev, Z. & Mikkola, H. (2022). First Pleistocene Record of Great Grey Owl (Strix nebulosa Forster, 1772) in Bulgaria. Comptes rendus de l’Académie bulgare des Sciences 75(5): 680‒685. [CrossRef]
- Mysterud, I. (2016). Range extensions of some boreal owl species: comments on snow cover. Ice crusts and climate change. Arctic, Antarctic and Alpine Research 48: 213‒219. [CrossRef]
- Solonen, T., 2006: Overwinter population change of small mammals in Southern Finland. Annales Zoologici Fennici 43: 295‒302.
- Marsh, P. & Woo, M.K. (1984).Wetting front advance and freezing of meltwater within a snow cover. 1. Observations in the Canadian Arctic. Water Resources Research 20: 1853‒1864.
- Visser, M.E., Both, C. & Lambrechts, M.M. (2004). Global climate change leads to mistimed avian reproduction. Pp. 89‒110. In: Møller, A.P., Fiedler, W. & Berthold, P. (Eds.) Effects of climate change on birds. Elsevier, Amsterdam.
- Gura, K.B., Bedrosian, B., Patla, S. & Chalfoun, A.D. (2025). Variation in habitat selection by male Strix nebulosa (Great Gray Owls) across the diel cycle. Ornithology 142: 1‒14. [CrossRef]
- Bateman, B., L. Taylor, C. Wilsey, J. Wu, G.S. LeBaron, and G. Langham (2020a). North American birds require mitigation and adaptation to reduce vulnerability to climate change. Conservation Science and Practice 2:e242.
- Bateman, B., L. Taylor, C. Wilsey, J. Wu, G. S. LeBaron, and G. Langham (2020b). Risk to North American birds from climate change-related threats. Conservation Science and Practice 2:e243.
- Schneider, Z., Matics, E., Hoffmann, G., Laczi, M., Herczog, G. & Matics, R. (2025). The potential for climate change to intensify nest-site competition between two sympatric owl species. Ibis. [CrossRef]
- Mikkola, H. (1983). Owls of Europe. 397 p. A.D. & T. Poyser, Calton.
- Matics, R., Bank, L., Varga, S., Klein,A. & Hoffmann, G. (2008). Interspecific offspring killing in owls. Biological Journal of the Linnean Society 95: 488‒494.
- Mikkola, H. (2026). Owls of the World. A Photographic Guide. 3rd Revised Edition. Bloomsbury/Christopher Helm, London.
- Wudu, K., Abegaz, A., Ayele, L. & Ybabe, M. (2023). The Impacts of Climate Change on Biodiversity Loss and Its Remedial Measures Using Nature Based Conservation Approach: A Global Perspective. Biodiversity and Conservation 32(12): 3681–3701. [CrossRef]
- Setsaas, T.H. (2024). Insects prefer cold winters with lots of snow. https://partner.sciencenorway.no/climate-change-insects-natural-sci… Accessed 23.9.2025.
- Hansson, L. & Henttonen, H. (1988). Rodent dynamics as community processes. Trends of Ecological Evolution 3: 195‒200. [CrossRef]
- Previtali, M.A., Lima, M., Meserve, P.L., Kelt, D.A. & Gutierrez, J.R. (2009). Population dynamics of two sympatric rodents in a variable environment: rainfall, resource availability, and predation. Ecology 90(7): 1996–2006. [CrossRef]
- Krebs, C.J., Boonstra, R., Gilbert, B.S., Kenney, A.J. & Boutin, S. (2019). Impact of climate change on the small mammal community of the Yukon boreal forest. Integrative Zoology 14: 528‒541. [CrossRef]
- Szpunar, G., Aloise, G., Mazzotti, S., Nieder, I. & Cristaldi, M. (2008). Effects of global climate change on terrestrial small mammal communities in Italy. Fresenius Environ Bulletin 17: 1526‒1533.
- Santoro, S., Sanchez-Suorez, C., Rouco, C., Palomo, L.J., Fernández, M.C., Kufner,M.B. & Moreno, S. (2017). Long-term data from a small mammal community reveal loss of diversity and potential effects of local climate change. Current Zoology 63(5): 515‒523. [CrossRef]
- Sharikov, A.V., Volkov, S.V., Sviridova, T.V. & Buslakov, V. V. (2019). Cumulative Effect of Trophic and Weather ‒ Climatic Factors on the Population Dynamics of the Vole-Eating Birds of Prey in Their Breeding Habitats. Biology Bulletin, 46(9):1097‒1107. [CrossRef]
- Kouba, M., Bartoš, L., Bartošová, J., Hongisto, K. & Korpimäki, E. (2020).Interactive influences of fluctuations of main food resources and climate change on long-term population decline of Tengmalm’s owls in the boreal forest. Scientific Reports 10:20429. [CrossRef]
- Lehikoinen, A., and Virkkala, R. (2016). North by northwest: climate change and directions of density shifts in birds. Global Change Biology 22: 1121‒1129.
- Bretagnolle, V. & Terraube, J. (2019). Predator-prey interactions and climate change. In: Dunn, P.O. & Møller, A.P. (Eds.) Effects of Climate Change on Birds. Second Edition. Oxford University Press. [CrossRef]
- Chadwick, J. (2025). Key North Atlantic current is on the brink of collapsing ‒ plunging Europe into the Ice Age, scientists warn.https://www.dailymail.co.uk/sciencetech/article-15165637/Key-No… Accessed 6.10.1605.
- Arellano Nava, B., Halloran, P.R., Boulton, C.A. & Lenton, T.M. (2024). Clams reveal the North Atlantic subpolar gyre has destabilised over recent decades. EGU General Assembly 2024, Vienna, Austria, 14‒19 April 2024, EGU24-20891. [CrossRef]
- Arellano Nava B,. Lenton, T.M., Boulton, C.A., Holmes, S., Scourse, J., Butler, P.G., Reynolds, D.J., Trofimova, T., Poitevin, P., Roman-Gonzáles, A. & Halloran, P.R. (2025). Recent and early 20th century destabilization of the subpolar North Atlantic recorded in bivalves. Sciences Advances 11: 1‒12. [CrossRef]

Table 1.
Changes in the active nest and fledged broods of owls in Finland. Material adopted from recorded observations in the Year books of Birdlife Finland 2008‒2024 [7]. S= South Finland, C = Central Finland and N = Northern Finland (limits given in the Methods). TN = total number of nests during the period, TA = annual average total/period, TP = percentage difference in total period number of nests and TD = total average annual nest number difference + or – between the periods.
Table 1.
Changes in the active nest and fledged broods of owls in Finland. Material adopted from recorded observations in the Year books of Birdlife Finland 2008‒2024 [7]. S= South Finland, C = Central Finland and N = Northern Finland (limits given in the Methods). TN = total number of nests during the period, TA = annual average total/period, TP = percentage difference in total period number of nests and TD = total average annual nest number difference + or – between the periods.
| Owl species |
2008‒2014 percentage of nests from total in S,C,N |
Average numbers of nests/region |
2018‒2024 percentage of nest from total in S,C,N |
Average numbers of nests/region |
Difference in two period percentages + or - |
Difference in annual average nest numbers |
| Aegolius funereus | S 7.9% C 74.4% N 17.7% TN 3050 |
S 35 C 324 N 77 TA 436 |
S 2.2% C 65.3% N 32.5% TN 1242 |
S 4 C 116 N 58 TA 178 |
S - 5.7% C - 9.1% N + 14.8% TP - 59.3% |
S - 31 C -208 N - 19 TD -258 |
| Asio flammeus | S 1.7% C 79.8% N 18.5% TN 346 |
S 0.9 C 39.4 N 9.1 TA 49.4 |
S 1.6% C 55% N 43.4% TN 129 |
S 0.3 C 10.1 N 8 TA 18.4 |
S - 0.1% C -24.8% N +24.9% TP -62.7% |
S - 0.6 C -29.3 N - 1.1 TD -31 |
| Asio otus | S 33.6% C 65.1% N 1.3% TN 2380 |
S 114.3 C 221.3 N 4.4 TA 340 |
S 51.3% C 47.1% N 1.6% TN 826 |
S 60.6 C 55.6 N 1.9 TA 118.1 |
S + 17.7% C - 18% N -7.5% TP -65.3% |
S - 53.7 C -165.7 N - 2.5 TD - 221.9 |
| Bubo bubo | S 48.6% C 48.9% N 2.5% TN 1215 |
S 84 C 85 N 4 TA 173 |
S 52.9% C 43.6% N 3.5% TN 953 |
S 72 C 59 N 5 TA 136 |
S + 4.3% C - 5.3% N + 1.0% TP - 21.6% |
S - 12 C - 26 N + 1 TD - 37 |
| Glaucidium passerinum | S 6.2% C 91.7% N 2.1% TN 2794 |
S 24.7 C 366.1 N 8.3 TA 399.1 |
S 10.5% C 85.6% N 3.9% TN 1170 |
S 17.6 C 143 N 6.6 TA 167.2 |
S + 4.3% C - 6.1% N + 1.8% TP - 58.1% |
S -7.1 C - 223.1 N - 1.7 TD - 231.9 |
| Strix aluco | S 46% C 54% N 0% TN 3045 |
S 200 C 235 N 0 TA 435 |
S 62.3% C 37.5% N 0% TN 4334 |
S 387 C 232 N 0 TA 619 |
S + 16.3% C - 16.5% N 0 TP + 29.7% |
S + 187 C - 3 N 0 TD + 184 |
| Strix nebulosa | S 2.8% C 81.3% N 15.9% TN 503 |
S 2 C 58.5 N 11.4 TA 71.9 |
S 3.7% C 57.3% N 39 TN 546 |
S 2.9 C 44.7 N 30.4 TA 78 |
S + 0.9% C - 24% N + 23.1% TP + 8.5% |
S + 0.9 C -13.8 N + 19 TD + 6.1 |
| Strix uralensis | S 10% C 85.8% N 4.2% TN 7435 |
S 106 C 912 N 44 TA 1062 |
S 11.6% C 82.4% N 6% TN 5613 |
S 93.1 C 660.7 N 48 TA 801.8 |
S + 1.6% C - 3.4% N + 1.8% TP - 24.5% |
S -12.9 C -251.3 N + 4 TD - 260.2 |
| Surnia ulula | S 0.5% C 15.2% N 84.3% TN 198 |
S 0.1 C 4.3 N 23.9 TA 28.3 |
S 0% C 9.9% N 90.1% TN 81 |
S 0 C 1.2 N 10.4 TA 11.6 |
S - 0.5% C - 5.3% N + 5.8% TP - 59.1% |
S - 0.1 C -3.1 N -13.5 TD - 16.7 |
Table 2.
Known climate change impacts on owls species. Data collected from global literature and from this study based on BirdLife Finland year book material [7] as presented in Table 1.
| Owl species | Climate change impact(s) | Reference(s) |
| Aegolius acadicus | Predicted to lose 60% of its current breeding habitats in the Southwestern US (hereafter SW US) | 10 |
| Aegolius funereus | In Finland, a drastic habitat related decrease (nearly 60%) in population but the reduced numbers have shifted northwards (+ 15%) In SW US projected to experience the steepest climate change related habitat loss, up to 85% Interestingly, in Central Europe one of range increasing owls even in commercially used forests and in Scots pine Pinus sylvestris and not Spruce Picea sp: a keystone species (namely Norway Spruce Picea abies) for the owl in Finland |
Table 1. 10 11 and 12 |
| Asio flammeus | In Finland, second largest loss of the population (nearly 63%) and the largest shift northwards (+25%) Loss, fragmentation, and degradation of grass- and wetlands causing a range-wide long term decline in North America Sea level rising reduces winter habitats |
Table 1. 13 14 |
| Asio otus | In Finland, the largest population loss (over 65%) and remaining owls have shifted to south (nearly 18%) In SW US predicted to lose 70% of its current breeding habitats |
Table 1. 10 |
| Asio stygius | In Colombia, up to 22% habitat loss due to climate change | 15 |
| Athene cunicularia | One well studied population in central New Mexico declined from 52 pairs to 1 pair in 16 years due to increasing air temperature and extreme multiyear drought conditions Changes in phenology linked to climate change in N-America. Delays in breeding are due to food limitation caused by drought. Drought during migration also constrains energetic requirements, forcing owls to stop more frequently and for longer periods at stopover sites, resulting in delayed arrival on breeding grounds |
16 17 |
| Bubo bubo | In Finland, nearly 22% decrease in population but no clear region shifts |
Table 1. |
| Bubo scandiacus | In Finland, Snowy Owls breeds irregularly , average 2 nests per year (min 0 and max 10) ; the population change – 88% calculated by comparing the average during the first four and last four years between 2007‒2024 Global decline, however, is only 30% over the past three generations Milder and wetter climate in the Arctic has been fading lemming population cycles and increasing the risk for detrimental black fly (Simulidae) attacks on nestlings and breeding females |
18 19 20,21 and 22 |
| Bubo virginianus | Predicted to lose 35% of its current breeding habitats in SW US | 10 |
| Glaucidium jardinii | Up to 43% habitat loss due to climate change in Colombia | 15 |
| Glaucidium minutissimum | Climate change will have a negative impact in Brazil | 23 |
| Glaucidium gnoma | Predicted to lose 75% of its current breeding habitats in SW US | 10 |
| Glaucidium passerinum | In Finland, nearly 60% population decline but no clear region shifts Climate change lowers food store quality when autumn rains and mild winters rot food hoards |
Table 1. 24 and 25 |
| Megascops albogularis | Up to 41% habitat loss due to climate change in Colombia | 15 |
| Megascops atricapilla | Climate change will have a negative impact in Brazil | 23 |
| Megascops ingens | In Colombia, significant habitat loss due to climate change but no percentage given | 15 |
| Megascops kennicottii | Predicted to lose 55% of its current breeding habitats in SW US | 10 |
| Megascops sanctaecatarinae | Climate change will have a negative impact in Brazil | 23 |
| Megascops trichopsis | Predicted to lose up to 60% of its current breeding habitats in SW US | 10 |
| Otus scops | Northward moving population in Britain and Europe | 26 |
| Psiloscops flammeolus | As cavity nester associated with large diameter trees, climate change would most likely be through disturbance processes that remove large trees. Shifts to denser forest structure would be a concern, but this is unlikely because drought and wildfire are projected to increase throughout the Northern Rockies In SW US projected to experience the steepest climate change related habitat loss, up to 85% |
27 10 |
| Pulsatrix koeniswaldiana | In Brazil, climate change will have a negative impact but no percentage given | 23 |
| Pulsatrix melanota | Up to 44% habitat loss due to climate change in Colombia | 15 |
| Strix albitarsis | In Colombia, significant habitat loss due to climate change but no percentage given | 15 |
| Strix aluco | Dampening vole cycles may drive this owl towards extinction from Northern England In Finland, advance in breeding dates have been noted Only owl in Finland with clearly increased population (+30%) but this far no northward expansion happening In Israel, climate change, which would increase spring temperatures and decrease rainfall, is a larger threat to this owl than rural development. |
28 29 Table 1. 30 |
| Strix hylophila | In Brazil, climate change will have a negative impact | 23 |
| Strix occidentalis | In SW US negative associations between warm, dry conditions and seemingly less heat-tolerant | 31 |
| Strix nebulosa | In Europe, population moving southward and westward against the climate change expectations In Finland, the second owl with a clear population increase (+ 8.5%) and this material would indicate population move between Central and North Finland (C-24% and N +23%) |
32‒40 Table 1. |
| Strix uralensis | In Finland, less alarming population decrease (less than 25%) and no clear sift to south or north. Climate change may advance the breeding in Finland– deep snow depth has delayed breeding In Slovakia, a positive population and range trend (from the east to the west), but reason may not be climate related |
Table 1. 29 41 and 42 |
| Surnia ulula | In Finland, almost 60% reduction in breeding population, remaining owls moving northwards (+6%) |
Table 1. |
| Tyto alba | Snowrich winters are part of non-linear climate change and can be dramatic to adult and juvenile barn owls in Switzerland | 43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



