Submitted:
09 October 2025
Posted:
10 October 2025
You are already at the latest version
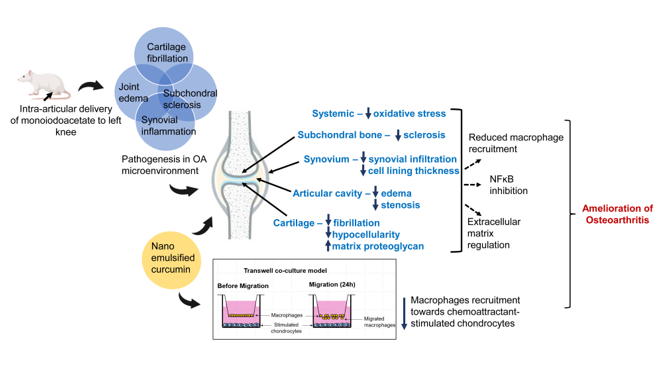
Abstract
The pathogenesis of knee osteoarthritis (OA) is multifaceted and involves the complete joint microenvironment. Despite beneficial evidence of curcumin, the mechanistic insights of nanoemulsified curcumin (n-Cur) delivery to the knee-OA microenvironment are limited. A detailed histological change that occurs in the knee joint of the OA model after localized delivery of curcumin was examined. n-Cur was prepared using a neutral dietary oil and a surfactant. Adult (5 mo) male SD rats were intra-articularly delivered 40 mg/ml of monoiodoacetate (MIA) to induce OA in the left knee and further treated with n-Cur (30 mg/ml). The effect of n-Cur on macrophage recruitment was evaluated using a co-culture model of CHON 001 and RAW 264.7 cells. In the MIA-model, localized delivery of n-Cur significantly reduced knee-joint edema, and articular cavity stenosis in the target site. Curcumin ameliorated cartilage degeneration by reducing fibrillation, hypocellularity, and restoring matrix proteoglycan, as evidenced by histology. Reduced synovial inflammation displays the effect of curcumin on the synovium, possibly by lowering the recruitment of macrophages in chemoattractant-stimulated chondrocytes. Thus, curcumin nanoemulsion can act as a chondroprotective agent, modulating the OA microenvironment by reducing joint edema, synovial inflammation, and oxidative stress in the OA model.
Keywords:
curcumin nanoemulsion
; osteoarthritis
; inflammation
; chondroprotective
; intra‐articular
; macrophage recruitment
1. Introduction
Knee osteoarthritis (OA) is a degenerative disease of the knee joints characterized by cartilage damage, synovial infiltration, and inflammation, which currently has no cure. Available pharmacological treatments and lifestyle modifications only provide symptomatic relief [1]. A major challenge to treatment options is the avascular nature of cartilage, which limits its ability to regenerate after wear and tear. Further, systemically administered drugs have shown limited therapeutic ability due to poor penetration. Compared to traditional drug delivery modes, intra-articular drug delivery is gaining traction due to its several benefits [2]. Despite this, intra-articular delivery of small molecules is prone to rapid clearance from the joint, resulting in less retention [3]. In this regard, complementary to site-specific delivery, nanoformulations are also known to increase solubility, enhance local bioavailability, and improve the compound’s overall absorption.
It is becoming increasingly evident that osteoarthritis is not just a cartilage degenerative disease but encompasses the complete joint microenvironment, including the subchondral bone, articular cavity, and the synovium. Pathological changes, including cartilage degeneration [4], joint edema [5], articular cavity stenosis, subchondral sclerosis [6], and synovial inflammation [7], occur in the knee joint during the progression of osteoarthritis. Transcriptome analysis of primary articular chondrocytes derived from different pathological grades of osteoarthritis has revealed significant alterations in gene expression responsible for immune response, cell adhesion, and extracellular matrix production, thus validating the involvement of the complete joint environment [8]. Despite the avascularity of cartilage, the vascular microenvironment in the surrounding tissues of the joint may contribute towards restorative pathways in OA.
In osteoarthritis pathogenesis, NFᴋB is a major regulator that modulates inflammation and degeneration [9]. NFᴋB is involved in the regulation of matrix-degrading enzymes and cartilage degeneration via the activation of various factors [10]. As an inflammatory modulator, NFᴋB plausibly exerts significant effects on the synovial membrane as well. Despite evidence in this regard [9], the majority of studies have explored the mechanistic changes of NF-κB only in relation to cartilage or chondrocytes [11,12]. The osteoarthritic synovium is characterized by the presence of fibroblasts in the lining layer, as well as macrophages and other infiltrated immune cells, including T and B lymphocytes [13]. Of these, macrophages are the prime mediators of inflammation, and macrophage-targeting is increasingly being reported for the alleviation of osteoarthritis [9,14]. Activated macrophages in the synovial membrane of the osteoarthritic microenvironment play a pivotal role in OA pathogenesis [15] and have been highlighted as possible therapeutic targets [16]. Recent evidences suggest cellular crosstalk between synovial cells and chondrocytes [17,18]. Despite these preliminary findings, studies investigating macrophage recruitment in OA are limited [19].
Recently, studies have demonstrated the chondroprotective properties of curcumin, a well-known polyphenol, in addition to its well-established anti-inflammatory effects [20,21]. The roles of curcumin in ameliorating osteoarthritis have been highlighted extensively [22]. Furthermore, formulating nanotherapeutic strategies for curcumin may enhance its bioactive effects and improve the chondroprotective effects [23]. Several preclinical studies have demonstrated the efficacy of various curcumin nanoformulations when administered intra-articularly, indicating anti-inflammatory and chondroprotective properties. Approaches such as curcumin-encapsulated microspheres [24], a core-brush nanoplatform containing curcumin [25], curcumin nanoparticles [26], or the synergistic delivery of curcumin and metformin nanoparticles [27] consistently demonstrate anti-inflammatory and anti-apoptotic effects in mitigating OA. For cartilage repair and regeneration, microenvironment-responsive nanomedicines show promising potential [28].
Despite reporting the efficacy and mechanistic evidence of curcumin on osteoarthritis pathogenesis [26,29,30], the mechanistic evidence of its effects on the OA microenvironment is limited. Macrophage recruitment is a central feature of synovial inflammation during osteoarthritis. The presence of infiltrated cells is explained by the activation state of macrophages (M1/M2), which constitute 43% of all infiltrated immune cells in the knee joint synovium during osteoarthritis [31]. Dysregulated cartilage has been shown to promote an inflammatory response in the synovium [7,32]. Synovial fibroblasts, inflammatory macrophages (M1), immune regulatory macrophages (M2), and dendritic cells abundantly constitute the synovium and produce major pro-inflammatory cytokines, including IL1β [33]. Few studies have investigated the impact of curcumin on synovial inflammation [26,29,30]; however, the roles of infiltrated macrophages in relation to chondrocytes have not been reported with nanoemulsified curcumin.
This study investigated the mechanisms occurring in the joint microenvironment by scoring histopathological changes in osteoarthritis that could be modulated by nanocurcumin. The monoiodoacetate (MIA)-induced model of osteoarthritis closely mimics human osteoarthritis in terms of biochemical, histological, and inflammatory changes [34]. Mimicking the osteoarthritis microenvironment, this study employed a co-culture model to evaluate the chemotactic effect of IL1β-stimulated chondrocyte-derived factors on the migration of macrophages. Curcumin nanoemulsion was prepared in a micellar form using a dietary oil formulated with curcuminoids.
This study provides the first report on the effects of nanoemulsified curcumin on OA-induced cartilage fibrillation in rats and inflammatory modulating activities in the microenvironment associated with osteoarthritis.
2. Results
2.1. Curcumin Nanoemulsion Reduced MIA-Induced Knee Joint Edema and Articular Cavity Stenosis in Osteoarthritic Rats
The formulated curcumin nanoemulsion was characterized with a size of 196.94 ± 12.65 nm, a negative zeta potential of 33.30 ± 0.70 mV, and a PDI (%) of 29.85 ± 1.35 (Table S1). Knee joint edema due to inflammation is a key characteristic symptom of osteoarthritis. To understand its effect on joint edema in an MIA-induced osteoarthritis model, joint diameter was measured. One week after the MIA injection, both anteroposterior and mediolateral joint diameter of the affected knee increased significantly and remained above baseline throughout the study duration, indicating OA-related edema (Figure S1). Intra-articular injection of nanocurcumin for four weeks significantly reduced edema of the knee joint, with joint diameter being comparable to that of the control at the end of the study duration (Figure 1B,D). Joint edema and the subsequent effect of nanocurcumin were site-specific and did not alter anteroposterior or mediolateral knee joint diameter of the unstimulated knee (Figure 1A,C).
Knee joints of control rats were observed to have smooth bone contours with a defined joint space (Figure 2A.i, iv). The most prominent MIA-induced structural alterations of the knee joint included joint effusion as indicated by increased opacity (Figure 2A.v), articular cavity erosion and stenosis indicated by roughened surfaces of bicondyles, subchondral bone sclerosis, and possible osteophytes (Figure 2A.ii). Intra-articular injections of curcumin nanoemulsion led to substantial improvement in articular cavity stenosis and joint effusion (Figure 2A.iii, vi). Slight surface irregularities were observed in addition to possible osteophytic lipping (Figure 2A.iii). Curcumin nanoemulsion significantly mitigated MIA-induced osteoarthritis [MIA-induced vs. Cur nanoemulsion: 3.33 ± 0.21 vs. 1.33 ± 0.21 (a.u.), arbitrary units, n=3/group] by reducing effusion and joint space narrowing (Figure 2B) based on the Kellgren-Lawrence classification system for osteoarthritis grading of knee joints (Table S2).
2.2. Curcumin Nanoemulsion Alleviated MIA-Induced Cartilage Fibrillation, Reduced Systemic Oxidative Stress, and Synovial Inflammation in the Knee Joint
Osteoarthritic histology exhibited deep fibrillation of cartilage, distinct erosion and loss of tidemark in the femorotibial joint (Figure 3A.ii, v, viii). In contrast, normal cellularity, surface, and intact tidemark were observed in the control (Figure 3A. i, iv, vii). Safranin O/fast green and toluidine blue staining revealed depletion of matrix proteoglycans, matrix atrophy and loss of chondrocytes compared to control. In contrast, curcumin nanoemulsion restored matrix proteoglycans, reduced cartilage erosion, fibrillation, and matrix atrophy (Figure 3A.iii, xi, ix). Histological findings were scored based on the Modified Mankin score (Table S3). Further, nanoemulsion significantly decreased modified Mankin score (MIA-induced vs. Cur nanoemulsion: 9.70 ± 0.40 vs. 4.05 ± 0.66, n=3/group) by reducing chondrocyte loss and restoring tidemark (Figure 3B).
In addition, H&E staining of the synovial membrane in osteoarthritic rats revealed increased number of cell layers in the synovial lining (Table S4) which significantly reduced post treatment with nanoemulsion (Figure 4A-B). Nanoemulsion effectively decreased infiltration of inflammatory cells and proliferation of the sub-synovial tissue compared to OA (MIA-induced vs. Cur nanoemulsion: 3.75 ± 0.25 vs. 2.25 ± 0.25 a.u, n=3/group). In addition, upregulated MDA levels in OA rats were significantly reduced with curcumin nanoemulsion injections (MIA-induced vs. Cur nanoemulsion: 0.92 ± 0.04 vs. 0.59 ± 0.03 a.u, n=3/group), implicating a decrease in systemic oxidative stress (Figure 4C).
2.3. Curcumin Nanoemulsion Reduced In vitro Macrophage Migration Towards Chondrocytes in the Presence of IL1β
Synovial inflammation and associated macrophage infiltration are key to the pathogenesis of osteoarthritis. To mimic the osteoarthritis microenvironment in vitro, we employed an indirect transwell co-culture system to check macrophage response to the release of chemokines and soluble factors from chondrocytes. Two cell lines, chondrocytes (CHON-001) and macrophages (RAW264.7) were employed. A dose-response assay was performed using the MTT assay to evaluate cell viability when cells were exposed to different concentrations (0.1 to 100 μM) of curcumin or curcumin nanoemulsion (Figure 5A-B). Compared to native curcumin, nanoemulsion maintained cell viability even at high concentrations in both chondrocytes and macrophages. Further, in chondrocytes, curcumin nanoemulsion treatment retains viable cells even at 20-100μM concentrations. For all subsequent experiments, concentrations of 5 or 10 μM were used for both curcumin and the nanoemulsion.
To assess the potential of nanocurcumin in mitigating the macrophage response and migration towards inflamed chondrocytes, a chemotactic gradient was established using IL1β. Chondrocytes were added to the lower chamber, while unstimulated macrophages were incubated on inserts. The co-culture setup prevented direct cell-to-cell contact and facilitated only the transfer of soluble factors, including chemokines, while also aiding the migration of macrophages in the presence of a strong chemotactic signal from chondrocytes. Chondrocytes exposed to IL1β promoted chemotactic migration of macrophages (Figure 5C. ii), which was reduced significantly upon treatment with curcumin. Both curcumin and nanoemulsion reduced macrophage migration despite the presence of IL1β (Figure 5D). Basal migration of macrophages towards unstimulated chondrocytes was present but minimal.
2.4. Modulation of Pro-Inflammatory Mediators and Matrix Regulators in Chondrocytes Exposed to Curcumin Nanoemulsion
We further analyzed the effect of curcumin nanoemulsion on the expression of pro-inflammatory modulators and matrix regulators in chondrocytes. Foremost, stability values for a panel of endogenously expressed genes (Table S5) were assessed, and TBP was chosen as an endogenous control due to its highest stability for the given experimental conditions in CHON-001 cells (data not presented). NFKB expression was upregulated significantly upon IL1β stimulation (Table 1), while nanoemulsion downregulated the expression of NFKB considerably. In contrast, NFKBIB was upregulated upon exposure to nanoemulsion (Table 1). TGFB1 was significantly downregulated by ~2-fold upon IL1β stimulation, while curcumin nanoemulsion restored TGFB1 expression. Upregulated levels of TNFA by IL1β were significantly reduced in cells exposed to nanoemulsion. Bone morphogenetic protein 2 (BMP 2) was upregulated considerably following IL1β stimulation. It was further upregulated by curcumin and curcumin nanoemulsion despite no significant difference between curcumin and IL1β. Both curcumin and nanoemulsion upregulated SMAD1; only nanoemulsion significantly downregulated SMAD5 expression. Expression of osteonectin (SPARC) was significantly downregulated by IL1β, which was further reduced in cells exposed to nanoemulsion. The relative mRNA expression of MMP2 did not change significantly with curcumin and nanoemulsion; however, curcumin, but not the nanoemulsion, downregulated MMP-9. Significant upregulation in expression of MMP14 and TIMP1 was observed in curcumin and nanoemulsion-exposed cells.
2.5. Curcumin Nanoemulsion Lowered IL1β-Induced Expression of NFκB in Macrophages (RAW264.7)
Macrophages were stimulated with IL1β to mimic osteoarthritis-related inflammatory responses in vitro and then treated with curcumin or a nanoemulsion. In macrophages, both curcumin and curcumin nanoemulsion displayed dose-dependent decrease in the expression of NFκB (Figure 6A). However, nanoemulsion at 5μM concentration was significantly more effective than native curcumin in reducing the expression (Figure 6B). Further, we evaluated the localization of NFκB in macrophages using immunocytochemistry. For this, cells were treated with either IL1β or a combination of IL1β and curcumin nanoemulsion (10 μM). An upregulation in overall cellular expression of NFκB (Figure 6C) in cells exposed to IL1β (10ng/ml) was observed. Curcumin nanoemulsion (10μM) was found to significantly downregulate both nuclear and cytoplasmic expression of NFκB (Figure 6D).
3. Discussion
This study demonstrated that targeted delivery of curcumin nanoemulsion by intra-articular administration ameliorates osteoarthritis pathogenesis in vivo by modulating stenosis, cartilage degeneration, and synovial inflammation. This study provides novel evidence that nanoemulsified curcumin reduces osteoarthritis-induced cartilage fibrillation in rats and mitigates macrophage recruitment towards chemoattractant-stimulated chondrocytes, suggesting its inflammatory-modulating activities in the microenvironment associated with osteoarthritis. Curcumin nanoemulsion attenuated structural alterations of osteoarthritis, including joint edema, articular cavity stenosis, cartilage erosion, and subchondral sclerosis by reducing inflammation and oxidative stress in the joint microenvironment.
Extensive thickening of the synovial membrane lining and infiltration of inflammatory cells were observed in OA rats in this study, suggesting synovial inflammation [35]. In this study, curcumin nanoemulsion reduced the thickening of synovial lining despite reduced but persistent infiltration. Persistent infiltration can be attributed to the presence of macrophages in the synovium [31]. Similar to the synovium, the subchondral bone and the joint cavity contribute to different mechanisms in promoting osteoarthritis pathogenesis. Significant changes in the subchondral bone, increased joint edema, and effusion observed in this study might contribute towards cartilage degeneration [6,36]. The reduction of edema and effusion in the joint after treatment with nanoemulsion, as also reported previously [26,27], underscores its function in mitigating osteoarthritis-related inflammation. Curcumin may also modulate alterations in the subchondral bone by suppressing osteoclastogenesis [37].
Further, articular cavity stenosis and possible osteophytes were observed in the OA model. Articular cavity stenosis, a characteristic feature of the MIA model, is associated with both pain and inflammation [38]. The reduction of stenosis due to nanocurcumin delivery suggests its beneficial effects in alleviating pain [39,40]. In the OA model, rough surfaces of the bicondyles indicated cartilage erosion, which was lessened with curcumin nanoemulsion, demonstrating its chondroprotective role. Additionally, synovial inflammation and structural changes are closely linked to pain responses in osteoarthritis [38]. The inflammatory microenvironment of osteoarthritis significantly contributes to oxidative stress [41]. Consistent with earlier findings [40,42], lower MDA levels, which assess oxidative damage in the affected tissues, indicate curcumin’s role in restoring oxidative balance.
Enhanced migration of macrophages toward OA-like chondrocytes observed in this study highlights the role of both IL1β as a chemoattractant and soluble factors secreted by the chondrocytes that facilitate crosstalk with macrophages [43,44]. Conversely, the curcumin nanoemulsion significantly decreased the chemotactic migration of macrophages, emphasizing the role of the chondrocyte secretome in macrophage recruitment. These findings offer new mechanistic insights into how curcumin affects chondrocytes in an inflammatory environment, contributing to macrophage recruitment and synovial inflammation.
In line with previous reports [4,45], this work also observed that monoiodoacetate led to a prominent degeneration of the articular cartilage, accompanied by erosion, fibrillation, chondrocyte loss, and reduced proteoglycan staining. This can be explained by alterations in the expression of matrix metalloproteinases [46] and TGFβ [47,48]. In this study, a curcumin nanoemulsion was found to reduce cartilage erosion, fibrillation, and hypocellularity. These structural alterations can be attributed to curcumin’s potential impact on the extracellular matrix and cartilage homeostasis [49]. In an explant OA model, curcumin has been previously reported to inhibit the degradation of the extracellular matrix and the release of glycosaminoglycans [50].
Previous studies on curcumin nanoemulsion have not explained the mechanisms behind the restoration of cartilage damage. The observed reduction in cartilage fibrillation and promotion of proteoglycan in histology, along with increased BMP2 expression due to curcumin nanoemulsion, suggests that BMP2 is involved in these processes. Bone morphogenetic protein-2, which is essential for bone and cartilage development, reduced cartilage damage and encouraged chondrocyte regeneration [51]. Typically, BMP2 transmits signals through the SMAD-dependent pathway [51]. The upregulation of BMP2 may result from the activation and phosphorylation of the BMP-specific receptor SMAD1, which was increased in this study. Furthermore, the role of BMP2 in the synthesis and maintenance of proteoglycans is well established [52]. Interestingly, overexpression of TGFβ has been linked to promoting an anti-inflammatory osteoarthritis (OA) microenvironment [53]. Curcumin nanoemulsion increased TGFβ expression in this study, emphasizing its role in both inflammation regulation and cartilage homeostasis. Curcumin nanoemulsion could promote ECM synthesis and chondrocyte proliferation via TGFβ [54]. A recent study showed that a deficiency of MMP14 expression led to inflammatory arthritis [55]. The increased expression of MMP14 by curcumin nanoemulsion in this study may be part of a restorative mechanism.
The reduced fibrillation of cartilage by curcumin nanoemulsion may also be due to its known anti-inflammatory activity. In line with several other studies [56-59], our results also show downregulation of NFᴋB and TNFα expression in chondrocytes. Previously, NFᴋB has been shown to regulate BMP2 expression, indicating its role in chondrogenesis [11]. NFᴋB is a key transcriptional regulator in synovial inflammation, modulating the expression of pro-inflammatory cytokines [9]. Curcumin has been shown to inhibit NFᴋB activity in osteoarthritis [56,59]. Upregulation of NFᴋB in OA-like macrophages was significantly reduced in the presence of curcumin nanoemulsion (Figure 6). Therefore, curcumin helps reduce the inflammatory environment associated with osteoarthritis by decreasing synovial inflammation, possibly by controlling macrophage recruitment through NF-κB inhibition.
A comparative assessment with native curcumin in vivo and further analysis of cartilage histology could not be achieved due to strict ethical obligation for providing novel data only using nanoemulsion, as the effects of curcumin are already established. Further, studying the impact of nanocurcumin on macrophage polarization in a cellular model would give better insights into its mechanism of action.
4. Materials and Methods
4.1. Preparation of Nanoemulsion
Curcumin nanoemulsion was prepared as a micellar form using an oil-in-water approach by mixing 10mM curcumin (#C7727, Sigma) with neutral triglyceride oil, Tween-20 (#P7949, Sigma) as surfactant, and deionized water. Oil and surfactant were taken in 1:1 ratio at ≤ 1% (v/v). The solution was sonicated at room temperature to aid emulsification and centrifuged. The prepared nanoemulsion was evaluated for its biophysical properties using a Zetasizer (DelsaTM Nano, Beckman Coulter, USA). Characterization was based on dynamic light scattering (nm) for size measurements, negative zeta potential for optical stability in solution, and polydispersity index (PDI) as a heterogeneity index of the particle size. A homogeneous micellar solution that fulfilled the biophysical characterization criteria (size < 200nm; zeta potential < -30 mV; and PDI < 0.3%) was used in all subsequent experiments.
4.2. Animal Experiment
The animal study and involved protocols were conducted in accordance with the institutional guidelines and approval of the Institutional Animal Ethics Committee (#ICMR-NIN/IAEC/2024-I/002). In brief, 5-month-old male SD rats (n=9) were procured from the animal facility, National Institute of Nutrition (NIN). Rats were housed individually in cages with a 12-hour light/dark cycle at a temperature of 22±2℃ and a relative humidity of 45-55%. Rats were given a standard chow diet (AIN93M) and water ad libitum. Sodium monoiodoacetate (MIA) (#I2512, Sigma) was used to induce osteoarthritis in rats according to the previously described protocol [60]. After one week of acclimatization, rats were randomly divided into three groups: control group (saline, vehicle), MIA-induced OA group (MIA, vehicle), and curcumin nanoemulsion group (MIA, treated with curcumin nanoemulsion, 30mg/ml [61]). To induce osteoarthritis in rats, the left knee was shaved and cleaned with alcohol for disinfection. Except for the control group, a single dose of 2mg MIA dissolved in 50μl sterile saline was injected intra-articularly using a 31G needle into the left knee. Rats in the control group were injected with sterile saline. After 4 weeks of disease induction using MIA rats, the rats in the curcumin nanoemulsion group received intra-articular injections of the nanoemulsion once a week for an additional 4 weeks, while the others were administered vehicle (sterile water). All intra-articular injections were given under mild anaesthesia.
4.3. X-ray Imaging and Scoring for Osteoarthritis
Radiographic images of the rat’s affected knee joint in anteroposterior and lateral views were obtained. Briefly, SD rats were anesthetized using ketamine (70mg/kg)/xylazine (2.5mg/kg) injection (intraparitoneal) and imaged under MRad 5.0 (BPL Medical Technologies, India) using manual mode with phase voltage – 240V, kVp – 70, mA – 40, mAs – 0.001, focal length – 80cm and developed using AGFA Musica. Images obtained were reviewed for OA-related changes following the Kellgren-Lawrence classification [62].
4.4. Knee Diameter Measurement
The diameter of the ipsilateral and contralateral knee joints was measured once a week throughout the study duration using digital vernier callipers (Mitutoyo) to monitor MIA-induced edema. Measurements of knee diameter (mm) were taken in anteroposterior and mediolateral directions as described [63].
4.5. Collection of Blood and Knee Joints
Blood was collected in clot-activator vacutainers from rats at three time points via retro-orbital puncture, and the separated serum was stored at -80 °C for further analysis. At the end of the experiment, rats were sacrificed by carbon dioxide asphyxiation. Intact knee joints were harvested and stored in 10% neutral buffered formalin for histopathology.
4.6. Serum Oxidative Stress
Oxidative stress in rats was assessed by measuring lipid peroxidation using the thiobarbituric acid reactive substances (TBARS) assay. Briefly, the serum was incubated with 0.3 M Tris-HCl buffer, 20% trichloroacetic acid (#Q28444, Qualigens), and 0.67% thiobarbituric acid (#T5500-25G, Sigma). Then, it was boiled and centrifuged to precipitate the protein. The supernatant was used to measure absorbance at 532nm against malonaldehyde (MDA) (#108383, Sigma) as the standard using a 96-well microplate reader (Biotek, PowerWave XS).
4.7. Knee Joint Histology and Staining
Left knee joints fixed in 10% formalin were decalcified using 5% formic acid for 2 weeks and embedded in paraffin as described previously [64]. Specimen blocks were cut into 4μm sections. The sections were deparaffinized, rehydrated, and stained using Haematoxylin and Eosin (H&E). Additionally, knee joint sections were also stained using safranin O (#16593, SRL) or toluidine blue (#22134, SRL) and counter-stained using fast green (#60339, SRL) as previously described [65]. The sections were visualized and imaged under a light microscope at 10x and 40x magnification (Eclipse TE2000U, Nikon, Japan). The histopathological changes were determined quantitatively using the modified Mankin scoring system [66,67].
4.8. Reagents and Cell Culture
CHON-001 chondrocyte cell line (#CRL_2846) and RAW 264.7 macrophage cell line (#TIB-71) were obtained from the American Type Culture Collection (ATCC), USA. IL1β (#201-LB-005) was procured from R&D Systems. Fetal Bovine Serum (FBS) (#10270106), Trypsin-EDTA (#25300062), and penicillin/streptomycin solution (#15140122) were obtained from Thermo Fischer Scientific, USA. Dulbecco's Modified Eagle Medium/High Glucose (DMEM) (#SH30249.01) was acquired from Cytiva, HyClone laboratories. F12 nutrient-rich medium (#21127-022) was obtained from Gibco. Curcumin (#C7727), Methyl thiazolyl diphenyl-tetrazolium bromide (MTT) (#5655), Dimethyl sulfoxide (DMSO) (#D2650) were procured from Sigma Aldrich, Germany.
CHON-001 cells were cultured in DMEM/F12 supplemented with 10% heat-inactivated FBS and 1% Penicillin/Streptomycin. RAW 264.7 cells were maintained in DMEM supplemented with 10% FBS and 1% antibiotics (penicillin/streptomycin). Cells were maintained at 37 °C in a 5% CO2 incubator. The cells were seeded in 6-well plates and serum-starved for 6 hours, followed by incubation with IL1β (10 ng/ml) for 24 hours to induce OA-like cells. Curcumin was dissolved in DMSO to prepare stock solution and further diluted with assay media keeping DMSO at 0.01% (v/v). Nanoemulsion was prepared as described earlier. Curcumin or its nanoemulsion (5μM,10μM) were administered to cells in the presence of IL1β.
4.9. Cell Viability by MTT Assay
Cells were seeded at a density of 1 × 10^4 cells in 96-well plates and serum-starved for 6 hours. Following this, the cells were exposed to different concentrations of native curcumin and its nanoemulsion. After 24 hours, the assay media were removed, and MTT (5mg/ml) was added. The cells were then incubated for an additional 4 hours to generate formazan crystals. The resulting crystals were solubilized with DMSO (100 μL), and the absorbance of the samples was determined using a microplate reader at 562nm (BioTek, PowerWave XS).
4.10. Immunoblotting
Immunoblotting in cell lysates was performed as described previously [68] to assess protein expression. The nitrocellulose membranes (#1620112, Bio-Rad) were incubated with primary antibody NF-κB (#SC-8008, Santa Cruz Biotechnology) at a 1:1000 dilution. HRP-conjugated anti-mouse antibody (#31430, Invitrogen) or anti-rabbit antibody (#31460, Invitrogen) was used for secondary incubation, followed by chemiluminescent imaging (iBright FL1500, Invitrogen). The blots were quantified and analyzed using ImageJ software, version 1.50i (NIH, Bethesda, MD, USA).
4.11. Quantitative Real-Time PCR
RNA isolation was performed from chondrocytes (CHON-001) using TRIzol (#T9424, Sigma-Aldrich). cDNA synthesis and quantitative real-time PCR were performed using a CFX-96 well real-time PCR system (Bio-Rad, USA) as previously described [68]. KiCqstart primers (Sigma) were used as mentioned (Table S5). A panel of candidate housekeeping genes was assessed, and stability values were derived using the NormFinder algorithm to identify the optimal endogenous control. Relative quantification of gene expression was calculated using ddCt method.
4.12. Detection of NFκB Localization by Immunocytochemistry
Cells cultured on coverslips were fixed using 4% paraformaldehyde for 20 minutes. After permeabilization with 0.25% Triton-X 100 (#RM845, Himedia) and being rinsed with phosphate-buffered saline (PBS), cells were blocked with 5% BSA and incubated with primary antibody NF-κB (#SC-8008, SantaCruz Biotechnology) with a dilution of 1:250 at 4℃ overnight. Next day, cells were incubated with secondary antibody conjugated with AlexaFluor 594 (#A11032, Invitrogen) for 2 hours after rinsing with PBS. The nuclei were counterstained, and coverslips were mounted onto slides with fluoromount containing DAPI (#ab104139, Abcam). Slides were imaged using a confocal microscope (TCS SP5, Leica Microsystems, Germany) at 40x magnification.
4.13. Transwell Co-Culture Chemotaxis Assay
Mimicking the OA microenvironment, this study employed a co-culture model of chondrocyte (CHON 001) and macrophage (RAW 264.7), to evaluate the chemotactic effect of IL1β-stimulated chondrocyte-derived factors on the migration of macrophages. Chondrocytes or macrophages were stimulated with IL1β to mimic osteoarthritis-related inflammatory responses in vitro and treated with curcumin or nanoemulsion for comparative evaluation. Chemotactic migration of macrophages was performed using a previously described protocol with slight modifications [19]. Briefly, CHON-001 cells (1x105) were seeded in the lower chamber of a 24-well plate, serum starved and treated with curcumin or curcumin nanoemulsion (10μM) in the presence of IL1β (10ng/ml) for 24 hours. After 24 hours, serum-starved RAW 264.7 macrophages (2 × 10^4) were added to the transwell hanging inserts (#37224, SPL Life Sciences) with 8.0μm pore size. The inserts were positioned into the wells containing stimulated chondrocytes and incubated for another 24 hours. Spent media was aspirated, and non-migratory macrophages from the apical side of the transwell insert were gently removed using a swab. Macrophages on the underside of the insert membrane were stained with 5μM calcein AM dye (#C1430, Invitrogen) for 30 minutes and visualized using fluorescent inverted microscope, 10x magnification (Eclipse TE2000U, Nikon, Japan). Mean migrated cells were analyzed using three random fields per image [69].
4.14. Statistical Analysis
Statistical analysis was performed with the GraphPad Prism v.8 platform. Each experiment was conducted independently and replicated multiple times, as described in the text or figure legends, with values being represented as mean ± SEM. Analysis of repeated measurements was carried out using a repeated measures mixed-effects ANOVA, and one-way ANOVA was used to analyze differences between multiple groups, followed by post-hoc Tukey’s test for multiple comparisons. p < 0.05 was considered statistically significant, with letters representing a significant difference.
5. Conclusions
Overall, the present study shows that the localized delivery of dietary oil-formulated curcumin nanoemulsion displayed comprehensive anti-inflammatory effects across the osteoarthritis microenvironment. In particular, curcumin mitigated cartilage degeneration by reducing fibrillation, hypocellularity, and restoring staining of proteoglycans, which could be due to reduced expression of NFᴋB in chondrocytes. Curcumin nanoemulsion also reduced synovial inflammation, potentially by suppression of NFᴋB in macrophages and mitigated macrophage recruitment indicating its anti-inflammatory role. These inhibitory effects of curcumin nanoemulsion suggest its potential as an NFᴋB-supressing therapeutic compound.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Representative images of joint edema in MIA-induced rats. (A) Anteroposterior and (B) Mediolateral diameter (in mm) of the affected knee (left) measured using digital vernier callipers at week 9. MIA-induced swelling was significantly reduced post-treatment with nanocurcumin via intra-articular injection at week 9.; Figure S2: Raw images of NFᴋB immunoblots and their respective β-actin blots in triplicate; Table S1: Characterization data of curcumin nanoemulsion; Table S2: Grading of X-ray radiographs of the knee joints of rats based on Kellgren-Lawrence classification; Table S3: Modified Mankin scoring for osteoarthritis based on histological findings in the knee joints of rats; Table S4: Scoring of synovial membrane inflammation based on histological findings in the knee joints of rats; Table S5: List of predesigned KiCqStart® SYBR® Green I pre-designed primers for gene expression analysis;.
Author Contributions
Investigation, Experiment, Methodology, Data curation, Writing- Original drafting, review and editing; K.S.N.H.; Investigation, Experiment, S.S.R.; Experiment, S.V.; Investigation, Experiment, N.S.B.; Experiment, A.M.; P.B.P.; M.S.; Critical comments, Editing, A.K.D.; Conceptualization, Project administration, Supervision, Drafting, Analysis, Writing—review and editing, Interpretation, Funding acquisition, S.B. All authors have read and agreed to the published version of the manuscript.
Funding
The work is part of a research project sponsored by the ICMR National Institute of Nutrition (Grant number: 23-BS08).
Institutional Review Board Statement
The study was conducted in accordance with the guidelines of the Helsinki Declaration and was approved by the Institutional Animal Ethical Committee of the ICMR National Institute of Nutrition (#ICMR-NIN/IAEC/2024-I/002).
Informed Consent Statement
Not applicable.
Data Availability Statement
All of the data generated or analyzed during this study are available from the corresponding author upon request.
Acknowledgments
K.S.N.H. was supported by the DST Inspire Fellowship, Department of Science and Technology, Government of India. We appreciate the efforts of Syeda Farhana in carrying out the animal trial and Raviprakash in imaging.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gregori, D.; Giacovelli, G.; Minto, C.; Barbetta, B.; Gualtieri, F.; Azzolina, D.; Vaghi, P.; Rovati, L.C. Association of Pharmacological Treatments With Long-term Pain Control in Patients With Knee Osteoarthritis: A Systematic Review and Meta-analysis. Jama 2018, 320, 2564–2579. [Google Scholar] [CrossRef]
- Huang, H.; Lou, Z.; Zheng, S.; Wu, J.; Yao, Q.; Chen, R.; Kou, L.; Chen, D. Intra-articular drug delivery systems for osteoarthritis therapy: shifting from sustained release to enhancing penetration into cartilage. Drug delivery 2022, 29, 767–791. [Google Scholar] [CrossRef]
- Cao, Y.; Ma, Y.; Tao, Y.; Lin, W.; Wang, P. Intra-Articular Drug Delivery for Osteoarthritis Treatment. Pharmaceutics 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Naveen, S.V.; Ahmad, R.E.; Hui, W.J.; Suhaeb, A.M.; Murali, M.R.; Shanmugam, R.; Kamarul, T. Histology, glycosaminoglycan level and cartilage stiffness in monoiodoacetate-induced osteoarthritis: comparative analysis with anterior cruciate ligament transection in rat model and human osteoarthritis. International journal of medical sciences 2014, 11, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Nam, D.; Kim, J. Pathological Characteristics of Monosodium Iodoacetate-Induced Osteoarthritis in Rats. Tissue engineering and regenerative medicine 2023, 20, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Guzman, R.E.; Evans, M.G.; Bove, S.; Morenko, B.; Kilgore, K. Mono-iodoacetate-induced histologic changes in subchondral bone and articular cartilage of rat femorotibial joints: an animal model of osteoarthritis. Toxicologic pathology 2003, 31, 619–624. [Google Scholar] [CrossRef]
- Li, J.; Gui, T.; Yao, L.; Guo, H.; Lin, Y.L.; Lu, J.; Duffy, M.; Zgonis, M.; Mauck, R.; Dyment, N.; et al. Synovium and infrapatellar fat pad share common mesenchymal progenitors and undergo coordinated changes in osteoarthritis. Journal of bone and mineral research : the official journal of the American Society for Bone and Mineral Research 2024, 39, 161–176. [Google Scholar] [CrossRef]
- Lewallen, E.A.; Bonin, C.A.; Li, X.; Smith, J.; Karperien, M.; Larson, A.N.; Lewallen, D.G.; Cool, S.M.; Westendorf, J.J.; Krych, A.J.; et al. The synovial microenvironment of osteoarthritic joints alters RNA-seq expression profiles of human primary articular chondrocytes. Gene 2016, 591, 456–464. [Google Scholar] [CrossRef]
- Ostojic, M.; Zevrnja, A.; Vukojevic, K.; Soljic, V. Immunofluorescence Analysis of NF-kB and iNOS Expression in Different Cell Populations during Early and Advanced Knee Osteoarthritis. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8. [Google Scholar] [CrossRef]
- Wu, S.; Morrison, A.; Sun, H.; De Luca, F. Nuclear factor-kappaB (NF-kappaB) p65 interacts with Stat5b in growth plate chondrocytes and mediates the effects of growth hormone on chondrogenesis and on the expression of insulin-like growth factor-1 and bone morphogenetic protein-2. The Journal of biological chemistry 2011, 286, 24726–24734. [Google Scholar] [CrossRef]
- Caron, M.M.; Emans, P.J.; Surtel, D.A.; Cremers, A.; Voncken, J.W.; Welting, T.J.; van Rhijn, L.W. Activation of NF-κB/p65 facilitates early chondrogenic differentiation during endochondral ossification. PLoS One 2012, 7, e33467. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Bhutani, N. Profiling joint tissues at single-cell resolution: advances and insights. Nature reviews. Rheumatology 2024, 20, 7–20. [Google Scholar] [CrossRef]
- Liu, Y.; Hao, R.; Lv, J.; Yuan, J.; Wang, X.; Xu, C.; Ma, D.; Duan, Z.; Zhang, B.; Dai, L.; et al. Targeted knockdown of PGAM5 in synovial macrophages efficiently alleviates osteoarthritis. Bone Res 2024, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.L.; Harasymowicz, N.S.; Klimak, M.A.; Collins, K.H.; Guilak, F. The role of macrophages in osteoarthritis and cartilage repair. Osteoarthritis and cartilage 2020, 28, 544–554. [Google Scholar] [CrossRef]
- Ummarino, A.; Gambaro, F.M.; Kon, E.; Torres Andón, F. Therapeutic Manipulation of Macrophages Using Nanotechnological Approaches for the Treatment of Osteoarthritis. Nanomaterials (Basel, Switzerland) 2020, 10. [Google Scholar] [CrossRef]
- Hamasaki, M.; Terkawi, M.A.; Onodera, T.; Homan, K.; Iwasaki, N. A Novel Cartilage Fragments Stimulation Model Revealed that Macrophage Inflammatory Response Causes an Upregulation of Catabolic Factors of Chondrocytes In Vitro. Cartilage 2021, 12, 354–361. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Kubota, K.; Asawa, Y.; Hoshi, K.; Hikita, A. M1-like macrophage contributes to chondrogenesis in vitro. Scientific Reports 2021, 11, 21307. [Google Scholar] [CrossRef]
- Olivotto, E.; Minguzzi, M.; D’Adamo, S.; Astolfi, A.; Santi, S.; Uguccioni, M.; Marcu, K.B.; Borzì, R.M. Basal and IL-1β enhanced chondrocyte chemotactic activity on monocytes are co-dependent on both IKKα and IKKβ NF-κB activating kinases. Scientific Reports 2021, 11, 21697. [Google Scholar] [CrossRef]
- Jin, Z.; Chang, B.; Wei, Y.; Yang, Y.; Zhang, H.; Liu, J.; Piao, L.; Bai, L. Curcumin exerts chondroprotective effects against osteoarthritis by promoting AMPK/PINK1/Parkin-mediated mitophagy. Biomed Pharmacother 2022, 151, 113092. [Google Scholar] [CrossRef] [PubMed]
- Csaki, C.; Mobasheri, A.; Shakibaei, M. Synergistic chondroprotective effects of curcumin and resveratrol in human articular chondrocytes: inhibition of IL-1beta-induced NF-kappaB-mediated inflammation and apoptosis. Arthritis Res Ther 2009, 11, R165–R165. [Google Scholar] [CrossRef]
- Basak, S.; Hridayanka, K.S.N.; Duttaroy, A.K. Bioactives and their roles in bone metabolism of osteoarthritis: evidence and mechanisms on gut-bone axis. Frontiers in Immunology 2024, 14. [Google Scholar] [CrossRef]
- Hridayanka, K.S.N.; Duttaroy, A.K.; Basak, S. Bioactive Compounds and Their Chondroprotective Effects for Osteoarthritis Amelioration: A Focus on Nanotherapeutic Strategies, Epigenetic Modifications, and Gut Microbiota. Nutrients 2024, 16. [Google Scholar] [CrossRef]
- Ratanavaraporn, J.; Soontornvipart, K.; Shuangshoti, S.; Shuangshoti, S.; Damrongsakkul, S. Localized delivery of curcumin from injectable gelatin/Thai silk fibroin microspheres for anti-inflammatory treatment of osteoarthritis in a rat model. Inflammopharmacology 2017, 25, 211–221. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, Z.; Wang, X.; Liang, J.; Zhao, L.; Zhang, Y.; Ren, J.; Zhang, S.; Liu, Y. A Core-Brush Nanoplatform with Enhanced Lubrication and Anti-Inflammatory Properties for Osteoarthritis Treatment. Advanced science (Weinheim, Baden-Wurttemberg, Germany) 2024, 11, e2406027. [Google Scholar] [CrossRef] [PubMed]
- Hamdalla, H.M.; Ahmed, R.R.; Galaly, S.R.; Naguib, I.A.; Alghamdi, B.S.; Ahmed, O.M.; Farghali, A.; Abdul-Hamid, M. Ameliorative Effect of Curcumin Nanoparticles against Monosodium Iodoacetate-Induced Knee Osteoarthritis in Rats. Mediators of inflammation 2022, 2022, 8353472. [Google Scholar] [CrossRef] [PubMed]
- El-Haddad, M.E.; El-Refaie, W.M.; Hammad, G.O.; El-Massik, M.A. Intra-articular metformin-curcumin cationic PLGA nanoparticles rejuvenate articular structure in MIA induced osteoarthritis model via modulating the crosstalk between miR93, TNFAIP3/TLR/NF-κB and AMPK/SIRT1 trajectories. International journal of biological macromolecules 2025, 315, 144482. [Google Scholar] [CrossRef]
- Xiong, Y.; Mi, B.B.; Shahbazi, M.A.; Xia, T.; Xiao, J. Microenvironment-responsive nanomedicines: a promising direction for tissue regeneration. Military Medical Research 2024, 11, 69. [Google Scholar] [CrossRef]
- Yabas, M.; Orhan, C.; Er, B.; Tuzcu, M.; Durmus, A.S.; Ozercan, I.H.; Sahin, N.; Bhanuse, P.; Morde, A.A.; Padigaru, M.; et al. A Next Generation Formulation of Curcumin Ameliorates Experimentally Induced Osteoarthritis in Rats via Regulation of Inflammatory Mediators. Front Immunol 2021, 12, 609629. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, C.K.; Song, S.H.; Yun, J.H.; Lee, A.; Park, H.J. Highly bioavailable curcumin powder suppresses articular cartilage damage in rats with mono-iodoacetate (MIA)-induced osteoarthritis. Food Sci Biotechnol 2020, 29, 251–263. [Google Scholar] [CrossRef]
- Mimpen, J.Y.; Hedley, R.; Ridley, A.; Baldwin, M.J.; Windell, D.; Bhalla, A.; Ramos-Mucci, L.; Buckley, C.D.; Coles, M.C.; Alvand, A.; et al. Cellular characterisation of advanced osteoarthritis knee synovium. Arthritis Res Ther 2023, 25, 154. [Google Scholar] [CrossRef] [PubMed]
- Roebuck, M.M.; Jamal, J.; Lane, B.; Wood, A.; Santini, A.; Wong, P.F.; Bou-Gharios, G.; Frostick, S.P. Cartilage debris and osteoarthritis risk factors influence gene expression in the synovium in end stage osteoarthritis. The Knee 2022, 37, 47–59. [Google Scholar] [CrossRef]
- Chou, C.H.; Jain, V.; Gibson, J.; Attarian, D.E.; Haraden, C.A.; Yohn, C.B.; Laberge, R.M.; Gregory, S.; Kraus, V.B. Synovial cell cross-talk with cartilage plays a major role in the pathogenesis of osteoarthritis. Sci Rep 2020, 10, 10868. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.H.; Ghosh, D.; Attari, S.; Ude, C.C.; Laurencin, C.T. Animal Models of Osteoarthritis: Updated Models and Outcome Measures 2016-2023. Regenerative engineering and translational medicine 2024, 10, 127–146. [Google Scholar] [CrossRef]
- Takahashi, I.; Ikeshima, K.; Matsuzaki, T.; Hoso, M. Histopathological changes in joint capsule and synovial membrane in a rat model of osteoarthritis. Osteoarthritis and cartilage 2016, 24, S398. [Google Scholar] [CrossRef]
- Holzer, L.A.; Kraiger, M.; Talakic, E.; Fritz, G.A.; Avian, A.; Hofmeister, A.; Leithner, A.; Holzer, G. Microstructural analysis of subchondral bone in knee osteoarthritis. Osteoporosis international : a journal established as result of cooperation between the European Foundation for Osteoporosis and the National Osteoporosis Foundation of the USA 2020, 31, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Liu, G.; Yan, J.; Zhang, Q.; Meng, F.; Wang, L. Curcumin alleviates osteoarthritis in mice by suppressing osteoclastogenesis in subchondral bone via inhibiting NF-κB/JNK signaling pathway. PLoS One 2024, 19, e0309807. [Google Scholar] [CrossRef]
- Nwosu, L.N.; Mapp, P.I.; Chapman, V.; Walsh, D.A. Relationship between structural pathology and pain behaviour in a model of osteoarthritis (OA). Osteoarthritis and cartilage 2016, 24, 1910–1917. [Google Scholar] [CrossRef]
- Mende, L.K.; Kuthati, Y.; Wong, C.S. Curcumin and Vitamin D Supplement Attenuates Knee Osteoarthritis Progression in ACLT + MMx Rat Model: Effect on Cartilage Protection and Pain Reduction. Nutrients 2025, 17. [Google Scholar] [CrossRef]
- Jhun, J.; Lee, D.; Na, H.S.; Cho, K.H.; Lee, S.Y.; Lee, J.S.; Lee, Y.J.; Kim, S.J.; Park, S.H.; Cho, M.L. Curcumin and omega-3 ameliorate experimental osteoarthritis progression in terms of joint pain and mitochondrial dysfunction. Journal of inflammation (London, England) 2025, 22, 27. [Google Scholar] [CrossRef]
- Altindag, O.; Erel, O.; Aksoy, N.; Selek, S.; Celik, H.; Karaoglanoglu, M. Increased oxidative stress and its relation with collagen metabolism in knee osteoarthritis. Rheumatology international 2007, 27, 339–344. [Google Scholar] [CrossRef]
- Chen, B.; He, Q.; Chen, C.; Lin, Y.; Xiao, J.; Pan, Z.; Li, M.; Li, S.; Yang, J.; Wang, F.; et al. Combination of curcumin and catalase protects against chondrocyte injury and knee osteoarthritis progression by suppressing oxidative stress. Biomed Pharmacother 2023, 168, 115751. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Zhu, Y.; Wei, K.; Pu, H.; Peng, R.; Xiao, J.; Liu, C.; Sun, X. Metformin Attenuates the Inflammatory Response via the Regulation of Synovial M1 Macrophage in Osteoarthritis. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Z.; Chen, C.; Wei, W.; Li, Z.; Zhou, H.; He, W.; Xia, J.; Li, B.; Yang, Y. SRGN promotes macrophage recruitment through CCL3 in osteoarthritis. Connective tissue research 2024, 65, 330–342. [Google Scholar] [CrossRef]
- Sasaki, Y.; Kijima, K.; Yoshioka, K. Validity evaluation of a rat model of monoiodoacetate-induced osteoarthritis with clinically effective drugs. BMC musculoskeletal disorders 2024, 25, 975. [Google Scholar] [CrossRef] [PubMed]
- Bryk, M.; Chwastek, J.; Mlost, J.; Kostrzewa, M.; Starowicz, K. Sodium Monoiodoacetate Dose-Dependent Changes in Matrix Metalloproteinases and Inflammatory Components as Prognostic Factors for the Progression of Osteoarthritis. Front Pharmacol 2021, 12, 643605. [Google Scholar] [CrossRef]
- van Beuningen, H.M.; van der Kraan, P.M.; Arntz, O.J.; van den Berg, W.B. Transforming growth factor-beta 1 stimulates articular chondrocyte proteoglycan synthesis and induces osteophyte formation in the murine knee joint. Laboratory investigation; a journal of technical methods and pathology 1994, 71, 279–290. [Google Scholar]
- Yang, X.; Chen, L.; Xu, X.; Li, C.; Huang, C.; Deng, C.X. TGF-beta/Smad3 signals repress chondrocyte hypertrophic differentiation and are required for maintaining articular cartilage. The Journal of cell biology 2001, 153, 35–46. [Google Scholar] [CrossRef]
- Zhou, Y.; Ming, J.; Deng, M.; Li, Y.; Li, B.; Li, J.; Ma, Y.; Chen, Z.; Wang, G.; Liu, S. Chemically modified curcumin (CMC2.24) alleviates osteoarthritis progression by restoring cartilage homeostasis and inhibiting chondrocyte apoptosis via the NF-κB/HIF-2α axis. Journal of molecular medicine (Berlin, Germany) 2020, 98, 1479–1491. [Google Scholar] [CrossRef]
- Clutterbuck, A.L.; Mobasheri, A.; Shakibaei, M.; Allaway, D.; Harris, P. Interleukin-1beta-induced extracellular matrix degradation and glycosaminoglycan release is inhibited by curcumin in an explant model of cartilage inflammation. Ann N Y Acad Sci 2009, 1171, 428–435. [Google Scholar] [CrossRef]
- Wu, M.; Wu, S.; Chen, W.; Li, Y.-P. The roles and regulatory mechanisms of TGF-β and BMP signaling in bone and cartilage development, homeostasis and disease. Cell Research 2024, 34, 101–123. [Google Scholar] [CrossRef]
- Luyten, F.P.; Yu, Y.M.; Yanagishita, M.; Vukicevic, S.; Hammonds, R.G.; Reddi, A.H. Natural bovine osteogenin and recombinant human bone morphogenetic protein-2B are equipotent in the maintenance of proteoglycans in bovine articular cartilage explant cultures. The Journal of biological chemistry 1992, 267, 3691–3695. [Google Scholar] [CrossRef]
- Lee, H.; Kim, H.; Seo, J.; Choi, K.; Lee, Y.; Park, K.; Kim, S.; Mobasheri, A.; Choi, H. TissueGene-C promotes an anti-inflammatory micro-environment in a rat monoiodoacetate model of osteoarthritis via polarization of M2 macrophages leading to pain relief and structural improvement. Inflammopharmacology 2020, 28, 1237–1252. [Google Scholar] [CrossRef]
- Lin, S.; Li, H.; Wu, B.; Shang, J.; Jiang, N.; Peng, R.; Xing, B.; Xu, X.; Lu, H. TGF-β1 regulates chondrocyte proliferation and extracellular matrix synthesis via circPhf21a-Vegfa axis in osteoarthritis. Cell Communication and Signaling 2022, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.D.; Gill, G.; Lin, H.; Roth, D.M.; Gu, H.M.; Wang, X.J.; Su, F.Y.; Alabi, A.; Alexiou, M.; Zhang, Z.; et al. Global, but not chondrocyte-specific, MT1-MMP deficiency in adult mice causes inflammatory arthritis. Matrix biology : journal of the International Society for Matrix Biology 2023, 122, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Xu, X.; Yi, P.; Hao, Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J Cell Mol Med 2020, 24, 10855–10865. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Tanzil, G.; Mobasheri, A.; Sendzik, J.; John, T.; Shakibaei, M. Effects of curcumin (diferuloylmethane) on nuclear factor kappaB signaling in interleukin-1beta-stimulated chondrocytes. Ann N Y Acad Sci 2004, 1030, 578–586. [Google Scholar] [CrossRef]
- Yang, Q.; Wu, S.; Mao, X.; Wang, W.; Tai, H. Inhibition effect of curcumin on TNF-α and MMP-13 expression induced by advanced glycation end products in chondrocytes. Pharmacology 2013, 91, 77–85. [Google Scholar] [CrossRef]
- Buhrmann, C.; Brockmueller, A.; Mueller, A.-L.; Shayan, P.; Shakibaei, M. Curcumin Attenuates Environment-Derived Osteoarthritis by Sox9/NF-kB Signaling Axis. International journal of molecular sciences 2021, 22, 7645. [Google Scholar] [CrossRef]
- Pitcher, T.; Sousa-Valente, J.; Malcangio, M. The Monoiodoacetate Model of Osteoarthritis Pain in the Mouse. JoVE 2016, e53746. [Google Scholar] [CrossRef]
- Nakahata, A.; Ito, A.; Nakahara, R.; Kishimoto, A.; Imaizumi, A.; Hashimoto, T.; Mukai, S.; Nakagawa, Y.; Kuroki, H. Intra-Articular Injections of Curcumin Monoglucuronide TBP1901 Suppresses Articular Cartilage Damage and Regulates Subchondral Bone Alteration in an Osteoarthritis Rat Model. Cartilage 2021, 13, 153s–167s. [Google Scholar] [CrossRef]
- Kohn, M.D.; Sassoon, A.A.; Fernando, N.D. Classifications in Brief: Kellgren-Lawrence Classification of Osteoarthritis. Clinical orthopaedics and related research 2016, 474, 1886–1893. [Google Scholar] [CrossRef]
- Horváth Á, I.; Bölcskei, K.; Szentes, N.; Borbély, É.; Tékus, V.; Botz, B.; Rusznák, K.; Futácsi, A.; Czéh, B.; Mátyus, P.; et al. Novel multitarget analgesic candidate SZV-1287 demonstrates potential disease-modifying effects in the monoiodoacetate-induced osteoarthritis mouse model. Front Pharmacol 2024, 15, 1377081. [Google Scholar] [CrossRef] [PubMed]
- Gerwin, N.; Bendele, A.M.; Glasson, S.; Carlson, C.S. The OARSI histopathology initiative - recommendations for histological assessments of osteoarthritis in the rat. Osteoarthritis and cartilage 2010, 18 Suppl 3, S24–34. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, N.; Laverty, S.; Kraus, V.B.; Aigner, T. Basic methods in histopathology of joint tissues. Osteoarthritis and cartilage 2010, 18 Suppl 3, S113–116. [Google Scholar] [CrossRef] [PubMed]
- Mankin, H.J.; Dorfman, H.; Lippiello, L.; Zarins, A. Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips. II. Correlation of morphology with biochemical and metabolic data. The Journal of bone and joint surgery. American volume 1971, 53, 523–537. [Google Scholar]
- Takahashi, I.; Matsuzaki, T.; Kuroki, H.; Hoso, M. Joint unloading inhibits articular cartilage degeneration in knee joints of a monosodium iodoacetate-induced rat model of osteoarthritis. Osteoarthritis and cartilage 2019, 27, 1084–1093. [Google Scholar] [CrossRef]
- Varma, S.; Molangiri, A.; Mudavath, S.; Ananthan, R.; Rajanna, A.; Duttaroy, A.K.; Basak, S. Exposure to BPA and BPS during pregnancy disrupts the bone mineralization in the offspring. Food and chemical toxicology : an international journal published for the British Industrial Biological Research Association 2024, 189, 114772. [Google Scholar] [CrossRef]
- Venter, C.; Niesler, C.U. Rapid quantification of cellular proliferation and migration using ImageJ. BioTechniques 2019, 66, 99–102. [Google Scholar] [CrossRef]
Figure 1.
The effect of curcumin nanoemulsion on joint edema in osteoarthritis rats. Knee diameter in rats was measured at different time points using digital vernier callipers to monitor joint edema. (A-B) Anteroposterior knee diameter (in mm) of unstimulated (A) and MIA-induced knee (B); (C-D) Mediolateral knee diameter (in mm) of unstimulated (C) and MIA-induced knee (D), respectively. Data were analyzed using a repeated-measures mixed-effects model with Tukey’s multiple comparison tests and represented as mean ± SEM (n = 3/group). Values with unlike superscript letters were considered significant with p < 0.05 vs. control.
Figure 1.
The effect of curcumin nanoemulsion on joint edema in osteoarthritis rats. Knee diameter in rats was measured at different time points using digital vernier callipers to monitor joint edema. (A-B) Anteroposterior knee diameter (in mm) of unstimulated (A) and MIA-induced knee (B); (C-D) Mediolateral knee diameter (in mm) of unstimulated (C) and MIA-induced knee (D), respectively. Data were analyzed using a repeated-measures mixed-effects model with Tukey’s multiple comparison tests and represented as mean ± SEM (n = 3/group). Values with unlike superscript letters were considered significant with p < 0.05 vs. control.
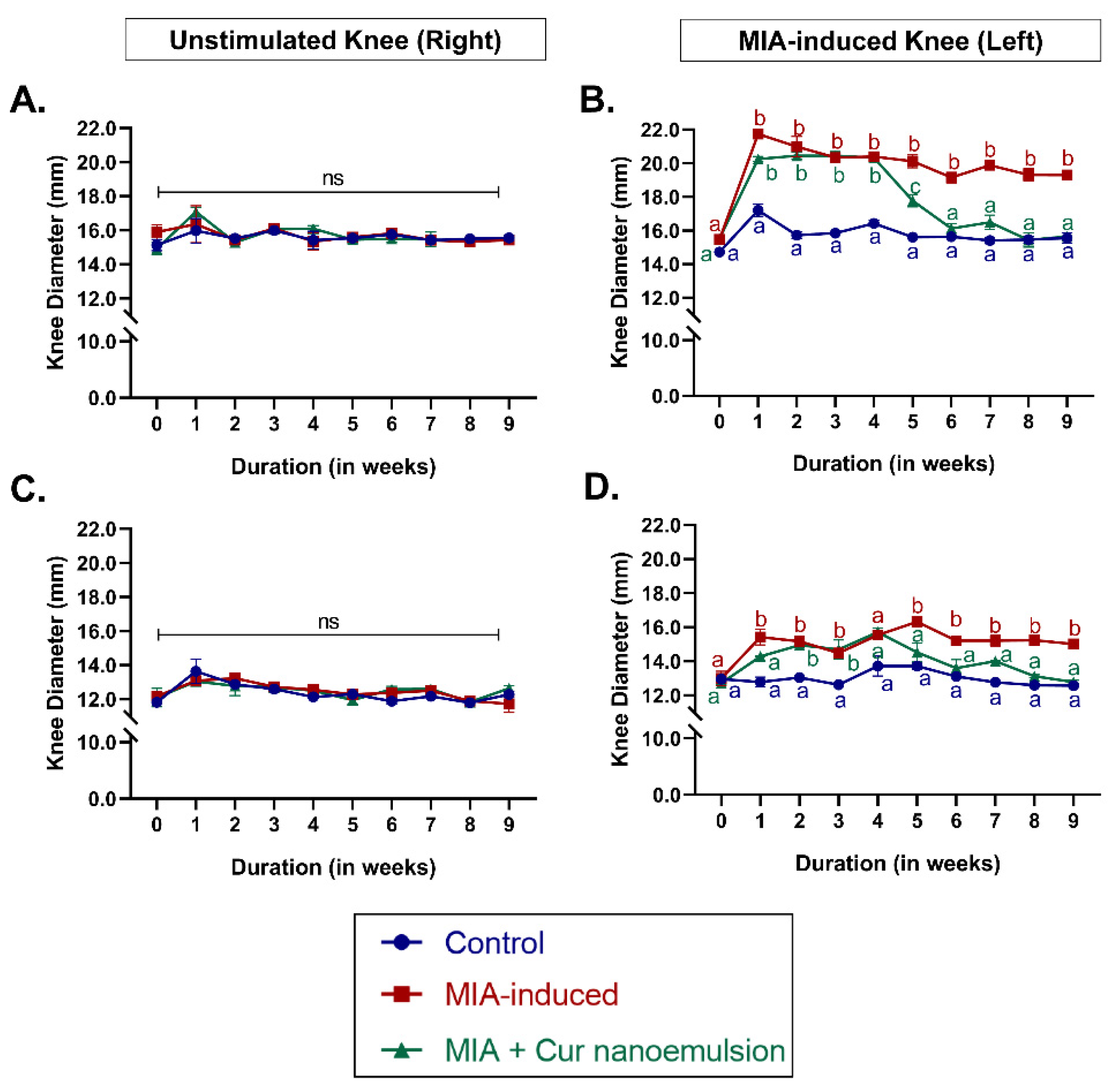
Figure 2.
X-ray radiographs of rat knee joints induced with monoiodoacetate and subsequently treated with nanocurcumin. (A) X-ray radiographs of the knee joint in anteroposterior (i-iii) and lateral (iv-vi) planes. (i, iv) – The control rat showed a well-defined joint space with uniform and smooth bone contours. (ii, v) - MIA-induced rats revealed stenosis of the articular cavity (1), roughened surfaces of bicondyles (2), increased opacity indicating joint effusion (3), subchondral sclerosis (4), and possible osteophytes (5). (iii, vi) - Treatment with curcumin nanoemulsion restored articular cavity stenosis (1) and reduced joint effusion (3), despite slight surface irregularity (2) and possible osteophytic lipping (5). (B) – Mean osteoarthritis grade using the Kellgren-Lawrence classification. Data are presented as mean ± SEM (n = 3/group) and analyzed using one-way ANOVA followed by post-hoc Tukey’s test for multiple comparisons. Unlike letters represent significance at p < 0.05 compared to the control.
Figure 2.
X-ray radiographs of rat knee joints induced with monoiodoacetate and subsequently treated with nanocurcumin. (A) X-ray radiographs of the knee joint in anteroposterior (i-iii) and lateral (iv-vi) planes. (i, iv) – The control rat showed a well-defined joint space with uniform and smooth bone contours. (ii, v) - MIA-induced rats revealed stenosis of the articular cavity (1), roughened surfaces of bicondyles (2), increased opacity indicating joint effusion (3), subchondral sclerosis (4), and possible osteophytes (5). (iii, vi) - Treatment with curcumin nanoemulsion restored articular cavity stenosis (1) and reduced joint effusion (3), despite slight surface irregularity (2) and possible osteophytic lipping (5). (B) – Mean osteoarthritis grade using the Kellgren-Lawrence classification. Data are presented as mean ± SEM (n = 3/group) and analyzed using one-way ANOVA followed by post-hoc Tukey’s test for multiple comparisons. Unlike letters represent significance at p < 0.05 compared to the control.
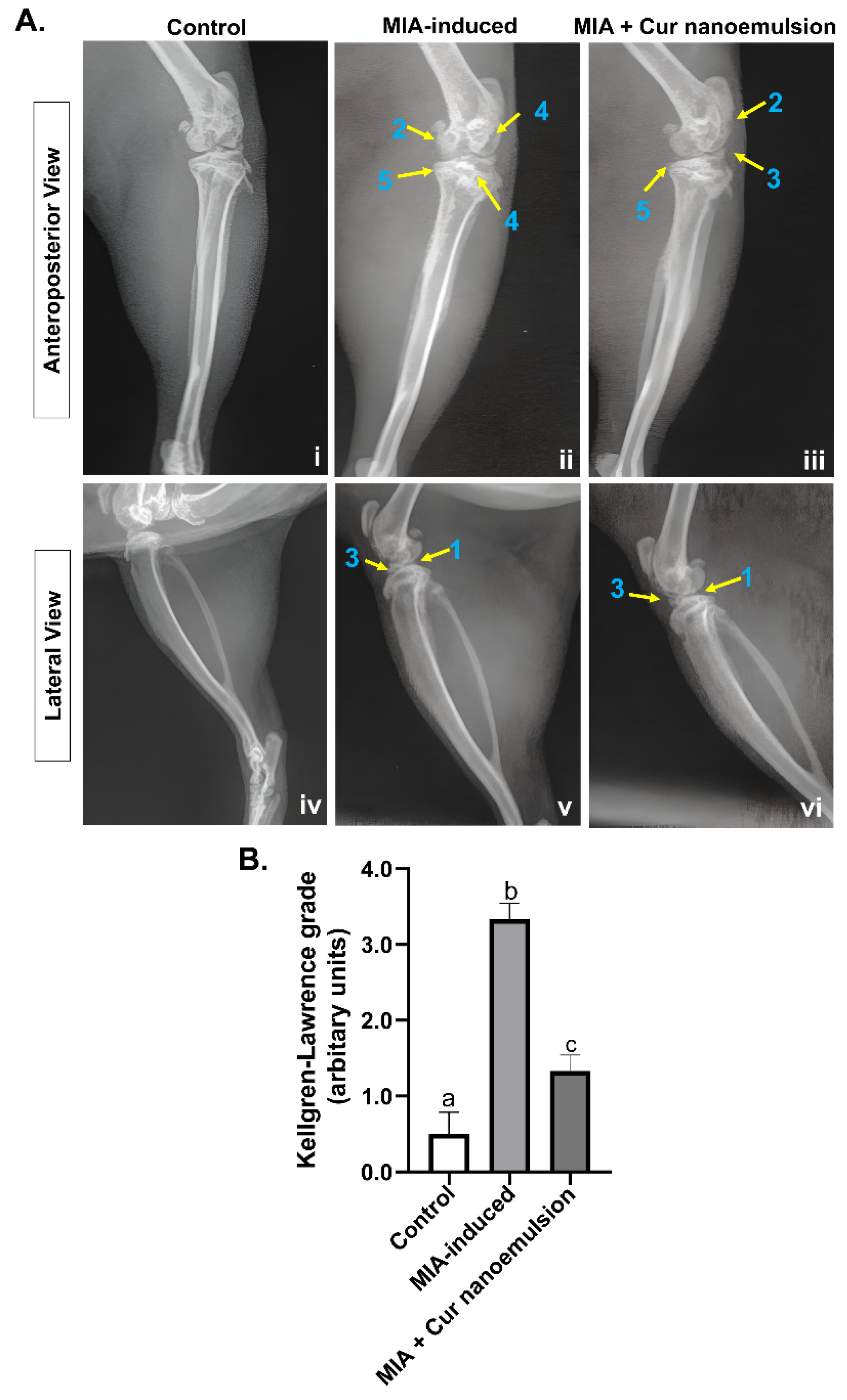
Figure 3.
Histopathological assessment using different staining methods and quantitative scoring of the knee joint in MIA-induced osteoarthritis rats treated with curcumin nanoemulsion. (A) Histology sections of the femorotibial joint in rats stained with (i-iii) Haematoxylin-Eosin, (iv-vi) Safranin O/Fast Green, (vii-ix) Toluidine Blue/Fast Green for control, MIA-induced rats treated with or without nanoemulsion. Control rats (i, iv, vii) exhibited a smooth cartilage surface and an intact tidemark, with normal cellularity and staining. MIA-induced rats (ii, v, viii) showed distinct cartilage erosion (1), deep fibrillation (2), loss of tidemark (3), depleted staining of matrix proteoglycans (4), matrix atrophy (5), and hypocellularity (6). Curcumin nanoemulsion (iii, vi, ix) increased matrix proteoglycans staining (4) displayed mild matrix atrophy (5), reduced fibrillation (2), reduced hypocellularity (6) and restored tidemark integrity (3). (B) Quantification of the osteoarthritic knee joints using modified Mankin scoring. Data were analyzed using one-way ANOVA with Tukey’s multiple comparisons and represented as mean ± SEM (n = 3), with unlike letters denoting significance at p < 0.05 compared to the control.
Figure 3.
Histopathological assessment using different staining methods and quantitative scoring of the knee joint in MIA-induced osteoarthritis rats treated with curcumin nanoemulsion. (A) Histology sections of the femorotibial joint in rats stained with (i-iii) Haematoxylin-Eosin, (iv-vi) Safranin O/Fast Green, (vii-ix) Toluidine Blue/Fast Green for control, MIA-induced rats treated with or without nanoemulsion. Control rats (i, iv, vii) exhibited a smooth cartilage surface and an intact tidemark, with normal cellularity and staining. MIA-induced rats (ii, v, viii) showed distinct cartilage erosion (1), deep fibrillation (2), loss of tidemark (3), depleted staining of matrix proteoglycans (4), matrix atrophy (5), and hypocellularity (6). Curcumin nanoemulsion (iii, vi, ix) increased matrix proteoglycans staining (4) displayed mild matrix atrophy (5), reduced fibrillation (2), reduced hypocellularity (6) and restored tidemark integrity (3). (B) Quantification of the osteoarthritic knee joints using modified Mankin scoring. Data were analyzed using one-way ANOVA with Tukey’s multiple comparisons and represented as mean ± SEM (n = 3), with unlike letters denoting significance at p < 0.05 compared to the control.
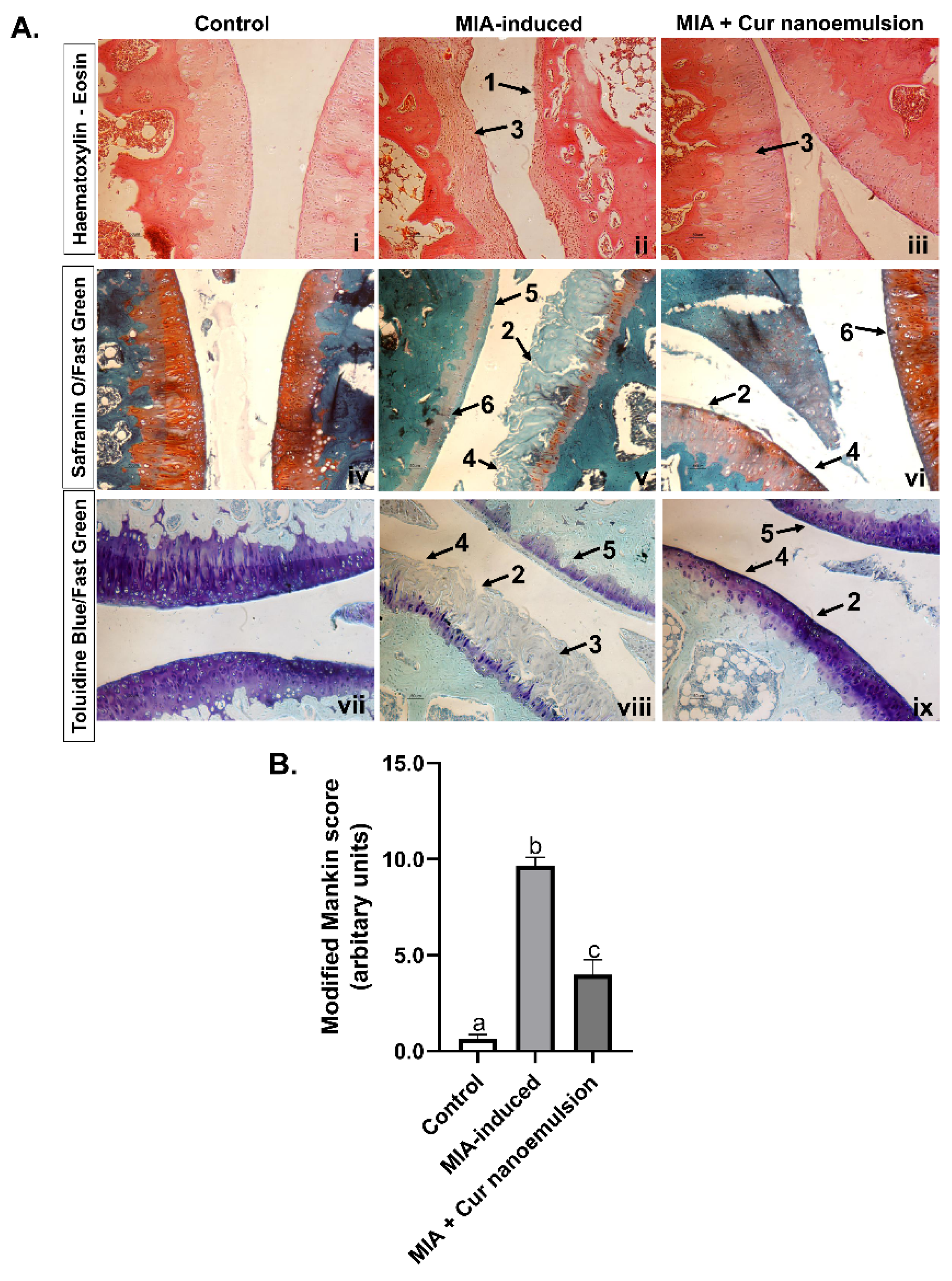
Figure 4.
Effect of curcumin nanoemulsion on synovial inflammation and oxidative stress in MIA-induced OA rats. (A) Representative histology sections of the synovial membrane in the knee joint of rats stained with Haematoxylin-Eosin (40x). Black arrows highlight the synovial lining, and the pointed triangle indicates infiltration of inflammatory cells. (B) Quantification of synovial inflammation. (C) Systemic oxidative stress measured by TBARS assay. Data were analyzed using one-way ANOVA with Tukey’s multiple comparisons and represented as Mean ± SEM (n = 3/group), with letters denoting significance at p < 0.05 compared to the control.
Figure 4.
Effect of curcumin nanoemulsion on synovial inflammation and oxidative stress in MIA-induced OA rats. (A) Representative histology sections of the synovial membrane in the knee joint of rats stained with Haematoxylin-Eosin (40x). Black arrows highlight the synovial lining, and the pointed triangle indicates infiltration of inflammatory cells. (B) Quantification of synovial inflammation. (C) Systemic oxidative stress measured by TBARS assay. Data were analyzed using one-way ANOVA with Tukey’s multiple comparisons and represented as Mean ± SEM (n = 3/group), with letters denoting significance at p < 0.05 compared to the control.
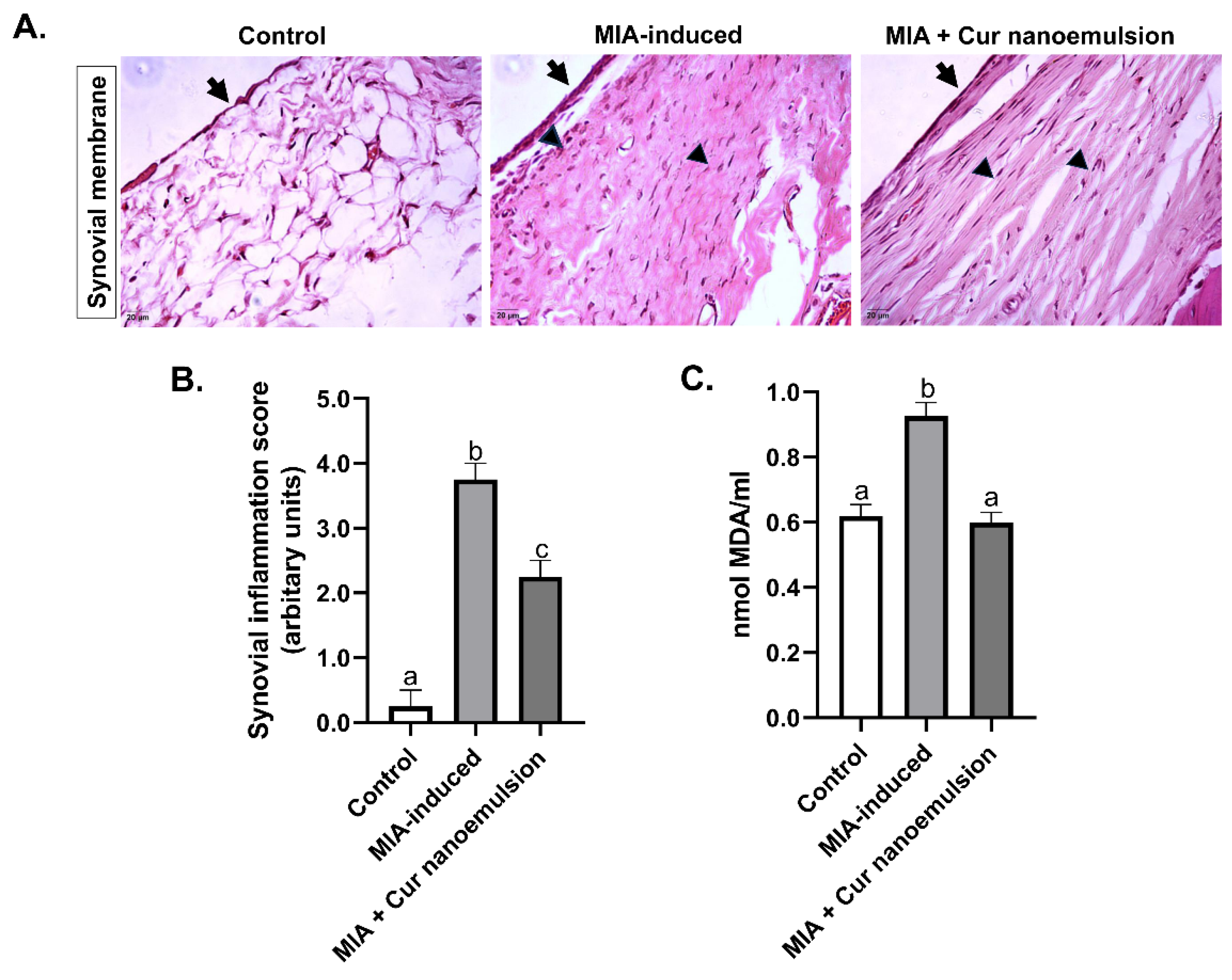
Figure 5.
Effect of curcumin nanoemulsion on cell viability and chemotactic migration of macrophages towards IL-1beta-induced chondrocytes. (A,B) Cell viability by CHON-001 and RAW 264.7 cells, respectively, exposed to different concentrations of curcumin and nanoemulsion (0.1-100μM) for 24 hours. Cell viability was evaluated using the MTT assay and calculated by comparing optical density values between the control and treatment groups after normalizing them with blank values, and expressed as a percentage of the control. Each value is represented as mean ± SEM (n=8), (C) Transwell co-culture model, (D) Representative images of calcein AM (green) stained macrophages on the underside of 8μm insert after 24hours of co-culture. Chondrocytes were cultured in the lower chamber and exposed to stimulus, while macrophages were incubated in the insert with plain assay media. Images (magnification, 10x) represent macrophage migration towards (i) unstimulated chondrocytes, (ii) IL1β-induced chondrocytes, (iii) IL1β-induced chondrocytes treated with curcumin (10μM), and (iv) IL1β-induced chondrocytes treated with nanoemulsion (10μM); (C) Quantification of migrated cells using mean fluorescence intensity per unit area. Data were analyzed using one-way ANOVA with Tukey’s multiple comparisons, and p < 0.05 was considered statistically significant compared to the control. Letters denote significant differences.
Figure 5.
Effect of curcumin nanoemulsion on cell viability and chemotactic migration of macrophages towards IL-1beta-induced chondrocytes. (A,B) Cell viability by CHON-001 and RAW 264.7 cells, respectively, exposed to different concentrations of curcumin and nanoemulsion (0.1-100μM) for 24 hours. Cell viability was evaluated using the MTT assay and calculated by comparing optical density values between the control and treatment groups after normalizing them with blank values, and expressed as a percentage of the control. Each value is represented as mean ± SEM (n=8), (C) Transwell co-culture model, (D) Representative images of calcein AM (green) stained macrophages on the underside of 8μm insert after 24hours of co-culture. Chondrocytes were cultured in the lower chamber and exposed to stimulus, while macrophages were incubated in the insert with plain assay media. Images (magnification, 10x) represent macrophage migration towards (i) unstimulated chondrocytes, (ii) IL1β-induced chondrocytes, (iii) IL1β-induced chondrocytes treated with curcumin (10μM), and (iv) IL1β-induced chondrocytes treated with nanoemulsion (10μM); (C) Quantification of migrated cells using mean fluorescence intensity per unit area. Data were analyzed using one-way ANOVA with Tukey’s multiple comparisons, and p < 0.05 was considered statistically significant compared to the control. Letters denote significant differences.
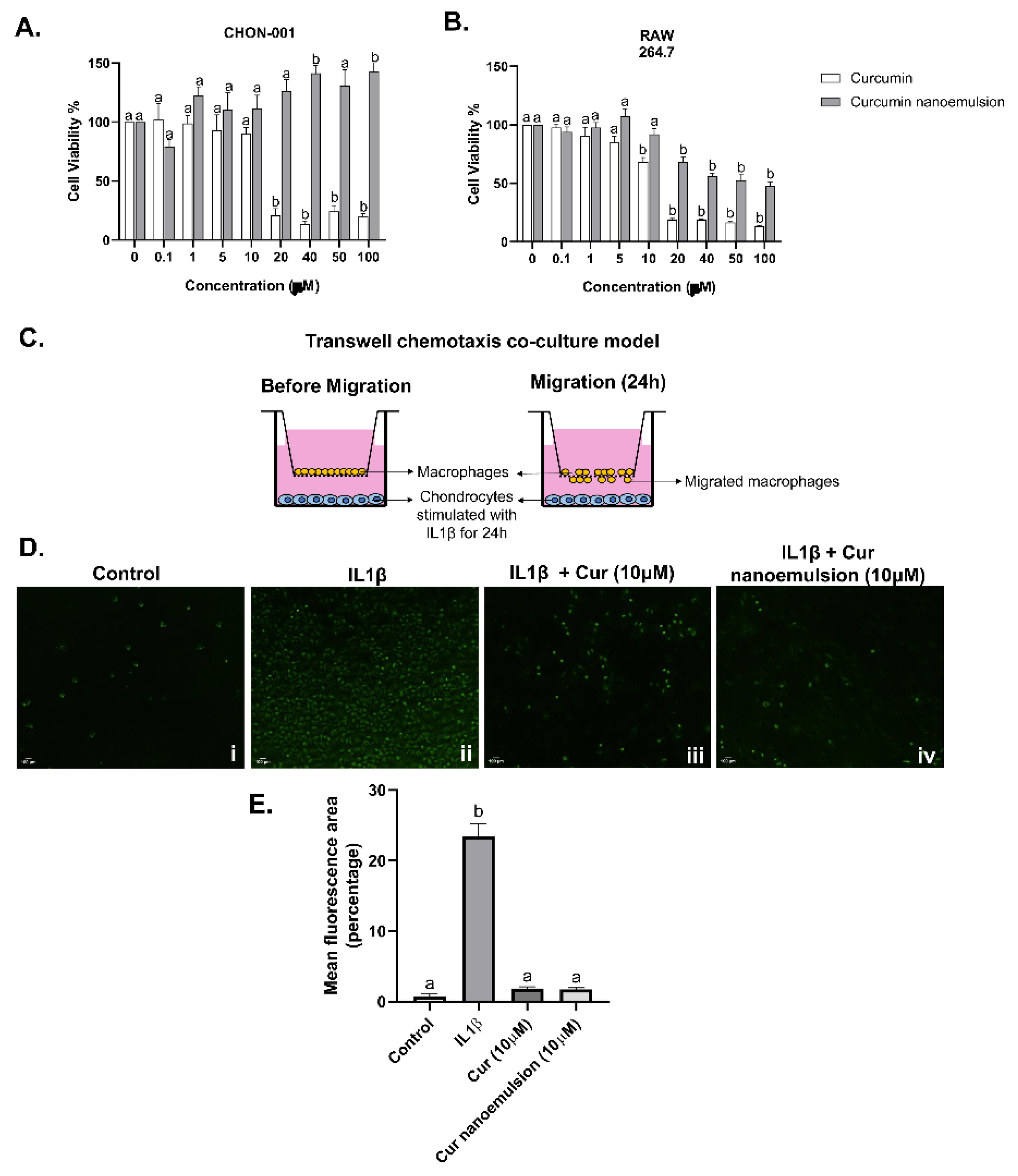
Figure 6.
Expression of inflammatory mediator NF-κB in RAW 264.7 macrophages following IL1β stimulation in the presence of curcumin/nanocurcumin (5-10μM). (A) Immunoblot of NF-κB and (B) its relative protein expression normalized with β-actin (mean ± SEM), (C) Nuclear and cytosolic expression of NF-κB in RAW 264.7 macrophages using immunocytochemistry after treating with IL1β and curcumin nanoemulsion (10μM). Cells were stained with Alexa Fluor 594-tagged NF-κB (red) and DAPI (blue) to stain nuclei with original magnification of 40x. Images were captured using a confocal laser scanning microscope with an oil immersion objective (TCS SP5, Leica Microsystems, Germany). (D) Corrected total cell fluorescence (CTCF) was calculated, analysed statistically and expressed in arbitrary units [CTCF = integrated density – (area of selected cell × mean fluorescence of background readings)]. Data were analyzed using one-way ANOVA with post-hoc Tukey’s multiple comparison test, with p < 0.05 considered significant, and results depicted using different letters.
Figure 6.
Expression of inflammatory mediator NF-κB in RAW 264.7 macrophages following IL1β stimulation in the presence of curcumin/nanocurcumin (5-10μM). (A) Immunoblot of NF-κB and (B) its relative protein expression normalized with β-actin (mean ± SEM), (C) Nuclear and cytosolic expression of NF-κB in RAW 264.7 macrophages using immunocytochemistry after treating with IL1β and curcumin nanoemulsion (10μM). Cells were stained with Alexa Fluor 594-tagged NF-κB (red) and DAPI (blue) to stain nuclei with original magnification of 40x. Images were captured using a confocal laser scanning microscope with an oil immersion objective (TCS SP5, Leica Microsystems, Germany). (D) Corrected total cell fluorescence (CTCF) was calculated, analysed statistically and expressed in arbitrary units [CTCF = integrated density – (area of selected cell × mean fluorescence of background readings)]. Data were analyzed using one-way ANOVA with post-hoc Tukey’s multiple comparison test, with p < 0.05 considered significant, and results depicted using different letters.
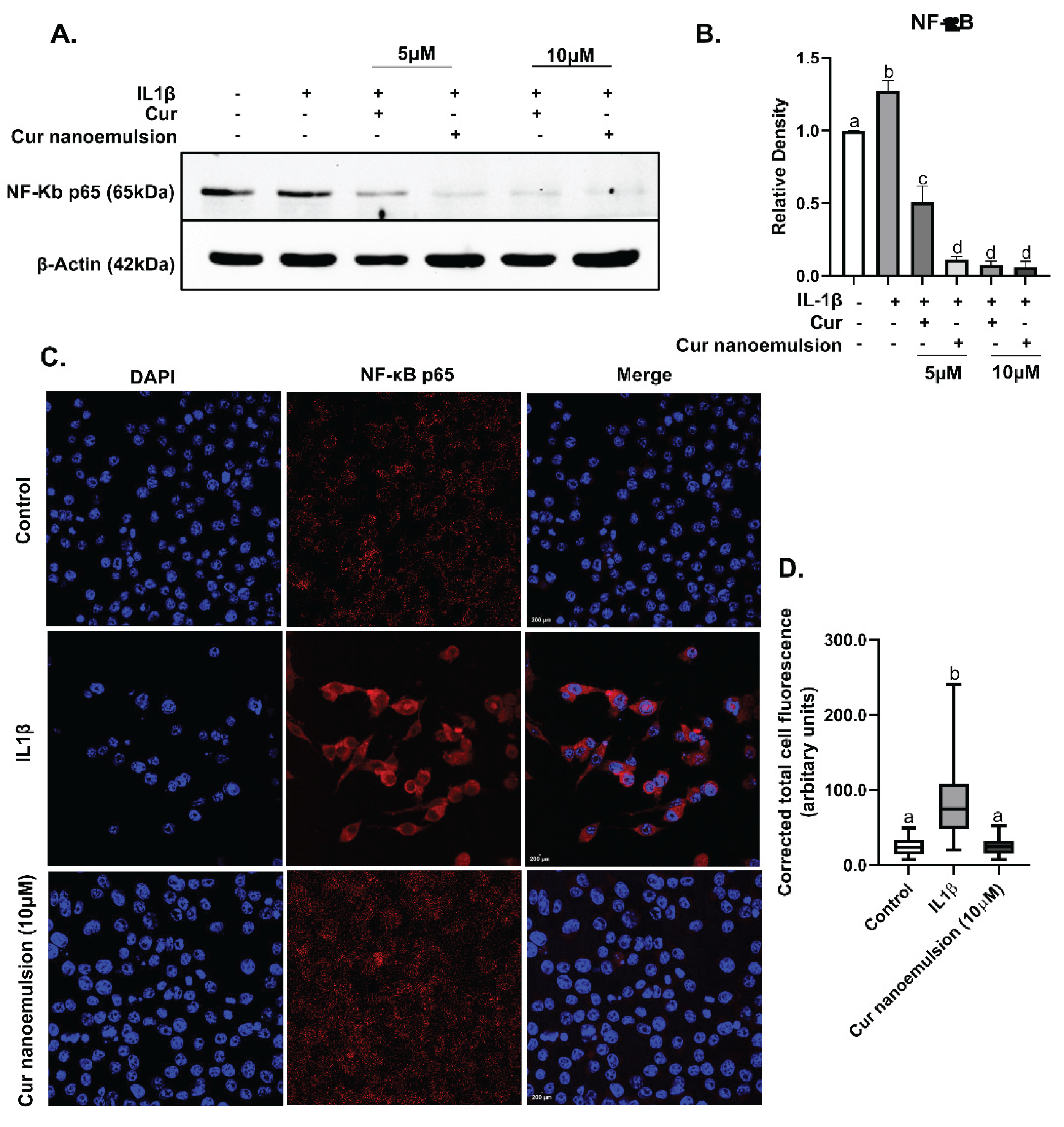

Table 1.
Relative mRNA expression1 of pro-inflammatory and matrix regulatory genes in chondrocytes after treatment with curcumin nanoemulsion2.
Table 1.
Relative mRNA expression1 of pro-inflammatory and matrix regulatory genes in chondrocytes after treatment with curcumin nanoemulsion2.
| Type of Gene | Gene Symbol | Control (Unstimulated) | IL1β (10ng/mL) | IL1β + Cur (10μM) | IL1β + Cur- Nanoemulsion (10μM) |
| Inflammatory modulators | |||||
| NFκB | 1.01 ± 0.04a | 4.41 ± 0.67b | 2.37 ± 0.05c | 2.47 ± 0.07c | |
| NFκB-IB | 1.01 ± 0.06a | 2.28 ± 0.32b | 3.14 ± 0.33b | 3.58 ± 0.22bc | |
| TNFα | 1.01 ± 0.04a | 1.65 ± 0.11b | 0.90 ± 0.03a | 0.97 ± 0.09a | |
| Matrix regulators | |||||
| TGFβ1 | 1.01 ± 0.02a | 0.57 ± 0.03b | 1.35 ± 0.06a | 1.24 ± 0.17a | |
| BMP2 | 1.01 ± 0.05a | 8.61 ± 2.29b | 12.78 ± 1.07b | 13.64 ± 1.31b | |
| SMAD1 | 1.01 ± 0.04a | 0.95 ± 0.05a | 2.20 ± 0.07b | 1.40 ± 0.08c | |
| SMAD5 | 1.01 ± 0.02a | 0.70 ± 0.22a | 0.52 ± 0.02a | 0.32 ± 0.02ab | |
| SPARC | 1.01 ± 0.02a | 0.39 ± 0.13b | 0.13 ± 0.01b | 0.18 ± 0.01b | |
| MMP2 | 1.01 ± 0.01a | 1.57 ± 0.10b | 1.56 ± 0.07b | 1.44 ± 0.18b | |
| MMP9 | 1.01 ± 0.02a | 1.94 ± 0.49b | 0.93 ± 0.16a | 1.94 ± 0.13b | |
| MMP14 | 1.01 ± 0.02a | 0.84 ± 0.23a | 1.52 ± 0.09b | 1.47 ± 0.04ab | |
| TIMP1 | 1.01 ± 0.03a | 1.98 ± 0.32b | 2.08 ± 0.18b | 2.53 ± 0.16b | |
1 The level of mRNA expression of genes was quantified after normalizing with endogenous control, TBP and calculated according to the 2-ΔΔCt method. Data are expressed as mean of relative mRNA ± SEM, n=3. *p < 0.05 vs. control. 2 mRNA expression was measured in IL1β stimulated CHON-001 cells after incubating with curcumin (10µM) or curcumin nanoemulsion (10µM) 24h.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



