Submitted:
07 October 2025
Posted:
08 October 2025
You are already at the latest version
Abstract
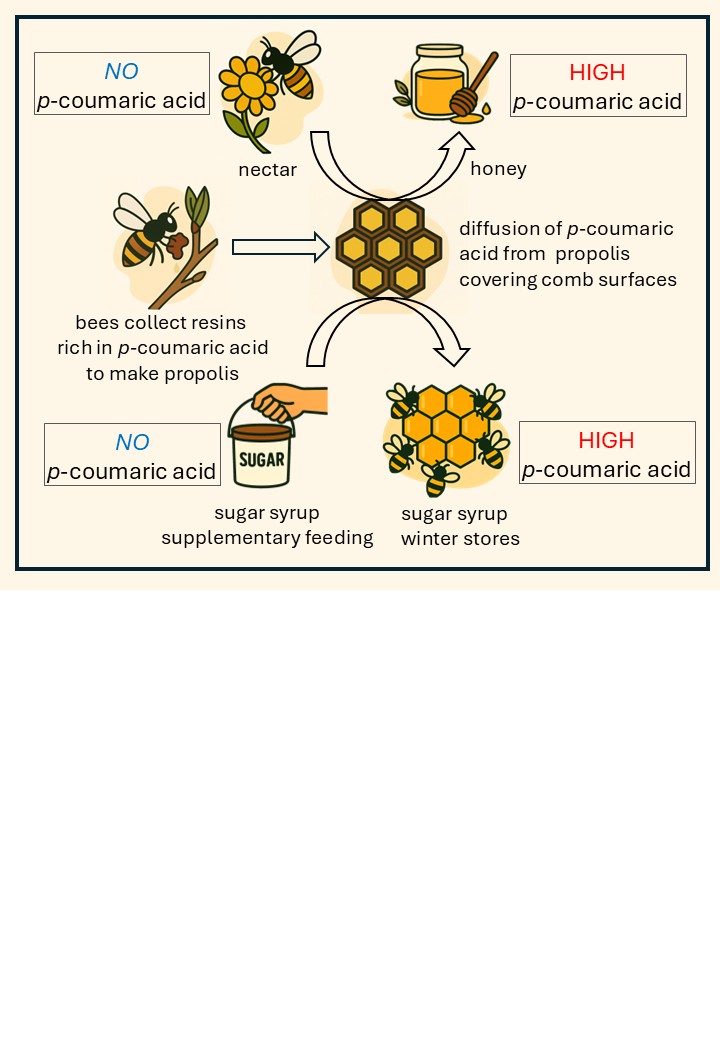
p-Coumaric acid is a natural phenolic compound supporting honey bee health by enhanc-27 ing detoxification, immunity and longevity. It also possesses antioxidant, anti-inflamma-28 tory and anticancer properties relevant to human health. While commonly detected in 29 honey and pollen, it is absent from nectar and sucrose-based supplements typically used 30 in beekeeping. Concerns have been raised whether supplemental saccharide feeding de-31 prives bees of this essential phytochemical. In this study, we quantified p-coumaric acid 32 in various bee-derived products and in supplementary sucrose syrup before and after 33 feeding to bees, using HPLC-HRMS analysis. Although fresh sucrose syrup contained 34 negligible amounts of p-coumaric acid, syrup stored in capped combs exhibited levels 35 comparable to or higher than those in honey. We identified propolis in combs as the pri-36 mary source of p-coumaric acid, diffusing into both honey and sucrose syrup during stor-37 age. Our findings demonstrate that supplemental feeding with sucrose syrup does not 38 diminish the bees' access to this key phytochemical and underscore the importance of 39 propolis-rich environment in bee health management.
Keywords:
honey bee
; phenolic compounds
; p-coumaric acid
; honey bee nutrition
; prop-41 olis
; honey
1. Introduction
p-Coumaric acid is a naturally occurring phenolic compound with a broad range of biological activities across taxa. In humans, it exhibits antioxidant, anti-inflammatory, antimicrobial and anticancer properties, contributing to the prevention of cardiovascular, neurodegenerative and oncological diseases [1,2,3]. Honey bee products (such as honey, bee bread or propolis), together with cereal bran and pigmented fruits (e.g., red grapes, cherries, blueberries), represent some of the richest reservoirs of this compound [4].
For honey bees (Apis mellifera), p-coumaric acid is essential to their health and colony sustainability. It supports detoxification pathways [5], enhances immune responses [6], extends lifespan [7], reduces pathogen load [8] and modulates gene expression linked to caste differentiation [9]. It is an intrinsic component of the honey bee diet, derived primarily from pollen and honey. The origin of p-coumaric acid in pollen is well established, as it is a major component of sporopollenin matrix of pollen walls [10]. However, the source of p-coumaric acid found in honey remains unclear, given its negligible concentration in nectar [11,12].
A widespread beekeeping practice involves the replacement of harvested honey with saccharide rich substitutes, most commonly sucrose syrup, fructose corn syrup or invert [13]. While studies have found no significant differences in overall colony winter survival between those fed honey and those fed sucrose syrup [14,15,16], bees overwintering on their own honey tend to exhibit enhanced physiological traits, including increased fat body reserves, increased lifespan and gene expression profiles associated with stress resilience [16,17]. Additionally, bees fed with honey show improved cognitive function and distinct gut microbiota profiles compared to those maintained on sucrose diet [18].
These findings suggest that supplemental saccharide feeds may lack nutritional components present in honey. The physiological disparities observed between bees fed honey and those fed artificial saccharide sources likely reflect the complex composition of honey that includes numerous bioactive compounds [19]. Notably, phenolic acids such as p-coumaric acid, caffeic acid, vanillic acid and chlorogenic acid may contribute to the health promoting effects of natural honey [20,21]. Consequently, concerns have been raised that sucrose syrup supplementation may deprive bees of these phytochemicals essential for coping with environmental and physiological stress [5,16].
In this study, we analyzed the p-coumaric acid content in honey and other honey bee products as well as in sucrose syrup before and after feeding to bees. Our goal was to assess whether winter feeding of sucrose syrup compromises p-coumaric acid availability in bee diets, potentially influencing colony health and survival.
2. Materials and Methods
2.1. Sources of Honey Bee Colonies and Honey Bee Products
Samples of honey, sucrose syrup stores, combs, queen cells, royal jelly, worker larval diet and propolis were taken from seven different apiaries in the region of South Bohemia, Czech Republic (Supplementary table ST1). The colonies of Apis mellifera carnica were managed according to standard practice, that included anti-varroa treatment (amitraz fumigation in autumn, oxalic acid drops in winter, formic acid in summer), honey harvest in June, and sucrose syrup feeding in August. The levels of Varroa destructor were regularly monitored by alcohol washes and did not exceed 2% levels throughout the season.
Honey samples were harvested in July 2021and July 2022 and honey bee colonies were then fed with sucrose syrup in August, according to a typical supplementary winter feeding protocol routinely used by the majority of bee keepers in the country. This comprised three doses of 5 kg granulated sucrose diluted with 3 liters of water, offered in one-week intervals (in total 15 kg sucrose for each colony was fed within two weeks). Empty combs originally built from wax foundations were inserted in the upper supper of the hive just before the supplementary feeding with sucrose syrup to avoid any honey deposition instead of the sucrose syrup stores. The capped combs full of sucrose syrup stores were marked and they were left in the colonies until springtime. The levels of p-coumaric acid in these capped sucrose syrup stores and in honey extracted from the same colonies were assessed in the spring 2022 (February, experiment in Figure 2A). In the second experiment (Figure 2B) the capped sucrose syrup store frames were analyzed two weeks after the supplementary feeding in August 2022 and stored outside the hive at room temperature in the laboratory, until next analysis in November 2022 and February 2023. For each analyzed comb, three sucrose stores samples originating from different parts of the comb were collected.
Royal jelly and worker larval diet samples were collected from queen and worker brood cells, respectively, using sterile spatulas immediately after larval removal. Corbicular pollen was obtained using pollen traps placed at hive entrances, while bee bread was harvested from comb cells following comb freezing to facilitate extraction. For the analysis of p-coumaric acid content in empty storage or brood combs, three samples were collected from different parts of each comb.
2.2. Analysis of p-Coumaric Acid Content in Honey, Sucrose Syrup Stores and Royal Jelly
Extraction of p-coumaric from these matrixes was based on [22]. 1 g of matrix was dissolved in 10 ml of water acidified with hydrochlorid acid to pH=2. After activating SPE (solid phase extraction) cartridge by methanol and washing with water acidified with HCl to pH=2, the dissolved sample was loaded. The cartridge was then washed with 6 ml of water acidified with HCl to pH=2. The p-coumaric acid was eluted by 6 ml of methanol. The eluent was concentrated under vacuum and subjected to HPLC-HRMS analysis. The concentration of p-coumaric acid was normalized to wet weight. Error bars in graphs represent standard deviations of biological triplicate measurements.
2.3. Analysis of p-Coumaric Acid in Empty Wax Combs, Queen Cells, Corbicular Pollen, Bee Bread and Propolis
Extraction of p-coumaric from these matrixes was performed according to a modified method of Mohdaly et al. [23]. Methanol extraction from these matrices proved more effective than water, with maximal compound recovery achieved after one to two weeks of extraction (supplementary figure S1). One gram of matrix was dispersed in 1 ml of methanol. After vortexing and ultrasounding the suspension, samples were macerated for two weeks with occasional shaking. After centrifugation, the supernatant was subjected to HPLC-HRMS. The concentration of p-coumaric acid was normalized to wet weight, error bars in graphs represent standard deviations of biological triplicate measurements.
To remove propolis rings from freshly built empty storage combs of different colours tweezers were used. For analysis of p-coumaric acid in queen cells, fifteen empty queen cells (within 24h after queen emergence) were collected from different hives, divided on light, medium dark and dark categories and each queen cell was analyzed separately for its content of p-coumaric acid.
2.4. The HPLC-HRMS Analyses
Samples for high performance liquid chromatography – high resolution mass spectrometry were analyzed on Dionex UltiMate 3000 UHPLC+ (Thermo Scientific, Sunnyvale, CA, USA) equipped with a diode-array detector. Separation of compounds was performed on reversed-phase C18 column (Phenomenex Kinetex, 150 × 4.6 mm, 2.6 µm, Torrance, CA, USA) using H2O (A) and Methanol (B) (both containing 0.1 % HCOOH) as a mobile phase at a flow rate of 0.6 ml min−1. The gradient was as follows: A/B 70/30 (0 min), 70/30 (in 1 min), 0/100 (in 20 min), 0/100 (in 30 min), and 70/30 (in 35 min). The mass spectrometer operated in negative ionisation with following settings: dry temperature 250°C; drying gas flow 10 L.m-1; nebulizer 3 bar; capillary voltage 3500 V; endplate offset 500 V. The spectra were collected in the range 100–1000 m/z with spectra rate 3 Hz. Sodium formate clusters were used as a calibrant at the beginning of each analysis.
2.5. Content of p-Coumaric Acid Calculation
The concentration of p-coumaric acid in the samples was determined using an external calibration curve constructed from a commercially available p-coumaric acid standard (Merck). Extracted ion chromatograms (EICs) corresponding to the protonated molecular ion [M+H]+ of p-coumaric acid (C9H8O3) were obtained for both standard solutions and sample extracts. In addition, due to in-source decarboxylation observed during ionization, a second EIC corresponding to the decarboxylated form (C8H8O) was evaluated. For quantification, sum of peak areas from both the sample EICs were compared to those of the standard, and only peaks with retention times matching those of the p-coumaric acid standard were considered.
2.6. Propolis Solubility in Sucrose Syrup and Honey
Propolis derived from hives was homogenized into powder using a blender. 150 mg of propolis was weighed into separate 2 ml Eppendorf tubes, mixed with 2.5 g honey or sucrose syrup (1:1 ratio sucrose to water) and incubated in a dark incubator at 35°C for 1 week to 3 months with occasional inverting of tubes. Honey with low basal level of p-coumaric acid was selected for the experiment. On the day of harvesting, samples with sucrose syrup were centrifuged at 10.000 g for 5 minutes and filtered through a 10 µm PSF membrane filter (Pall Corporation) to eliminate unsolubilized propolis particles. Samples containing honey were diluted with the same volume of water just before filtration and processed immediately the same way as samples with sucrose syrup. Content of p-coumaric acid was measured as described above and normalized to the weight of honey or sucrose syrup before filtration. Each point represents average values of biological triplicates.
2.7. Statistical Analysis
Each measurement was performed in biological triplicate. Statistical tests were performed by GraphPad Prism version 8.0.1 software (San Diego, California). Statistical significance: * p<0.05, ** p<0.01, *** p<0.001, **** p<0.0001.
3. Results
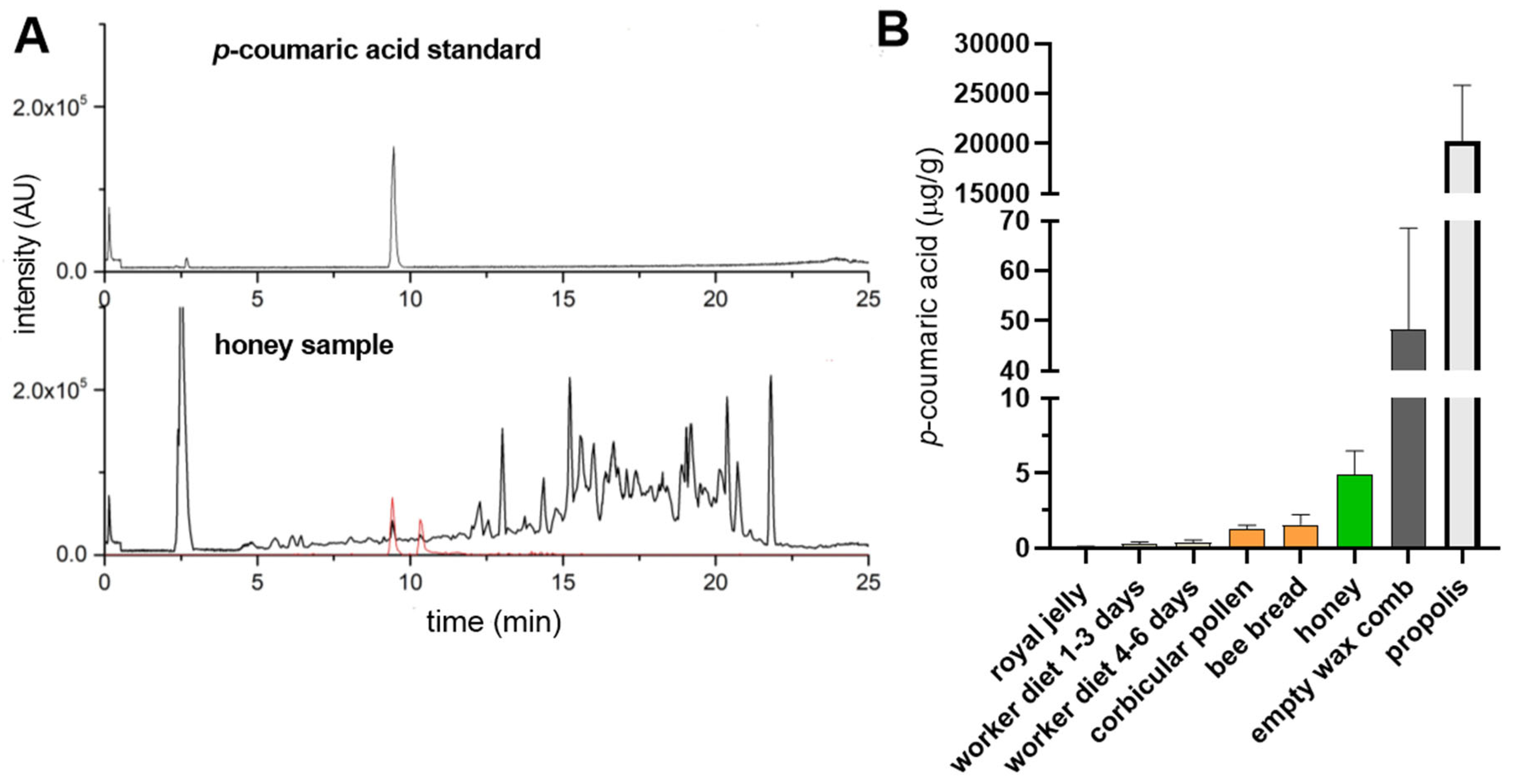
In order to evaluate the differences in the p-coumaric acid content in various bee products, we quantified its levels in royal jelly, worker diets for larvae of different ages, corbicular pollen, bee bread, honey, empty wax combs and propolis (Figure 1). As previously reported [9], p-coumaric acid was undetectable or present only at trace levels in royal jelly (0.05 μg/g on average) whereas it was detected in the worker larval diet, increasing with larval age (0.2 and 0.4 μg/g on average for larvae of 1-3 days and 4-6 days, respectively). Pollen and bee bread contained less than 2 mg/g of p-coumaric acid, while honey contained approximately 5 mg/g. Interestingly, empty wax combs contained on average ten times more p-coumaric acid than honey (48 mg/g). Propolis exhibited the highest concentrations, exceeding those in honey by over three orders of magnitude (20 mg/g).
To directly compare p-coumaric acid levels, we measured its concentration in honey and supplementary sucrose syrup before and after feeding to bees, using paired samples from the same colonies. In the first experiment, frames containing capped sucrose syrup stores were removed from colonies in the spring, following their storage in hives since the previous summer. As expected, freshly prepared sucrose syrup contained no detectable p-coumaric acid. However, capped sucrose stores recovered from the colonies contained measurable levels of p-coumaric acid, exceeding the concentrations found in honey samples collected from the same colonies prior to syrup feeding (Figure 2A).
To minimize the possibility that the observed increase resulted from mixing of sucrose syrup stores with residual honey by bees during winter, a second experiment was conducted. Capped sucrose syrup combs were removed from colonies shortly after feeding in summer, stored under controlled conditions in the laboratory and p-coumaric acid concentrations were monitored over time. The compound was detectable already after two weeks in storage and continued to accumulate throughout autumn and winter in most cases (Figure 2B). We observed that combs from apiaries 3 and 6 were dark, while the comb from apiary 7 was lighter.
Figure 1.
The content of p-coumaric acid in various honey bee products. (A) HPLC-HRMS chromatogram of p-coumaric acid standard solution (top) and honey (bottom, black line). Extracted ion chromatogram indicates ions contributing to p-coumaric acid signal (bottom, red line). (B) Apiary 1: Týn and Vltavou; apiary 2: Sedlce; apiary 3: Doubravice; apiary 4: České Budějovice 7; apiary 5: České Budějovice 31. Error bars represent standard deviations of biological triplicates.
Figure 1.
The content of p-coumaric acid in various honey bee products. (A) HPLC-HRMS chromatogram of p-coumaric acid standard solution (top) and honey (bottom, black line). Extracted ion chromatogram indicates ions contributing to p-coumaric acid signal (bottom, red line). (B) Apiary 1: Týn and Vltavou; apiary 2: Sedlce; apiary 3: Doubravice; apiary 4: České Budějovice 7; apiary 5: České Budějovice 31. Error bars represent standard deviations of biological triplicates.
Our results indicated that sucrose syrup acquired p-coumaric acid during storage in combs. Moreover, syrup stored in darker combs exhibited higher p-coumaric acid concentrations than syrup stored in light combs, suggesting a potential association between comb pigmentation and p-coumaric acid enrichment (Figure 2C). Since comb pigmentation is closely linked to propolis deposition, we hypothesize that propolis within the combs represents the primary source of p-coumaric acid, which diffuses into the bees’ saccharide stores.
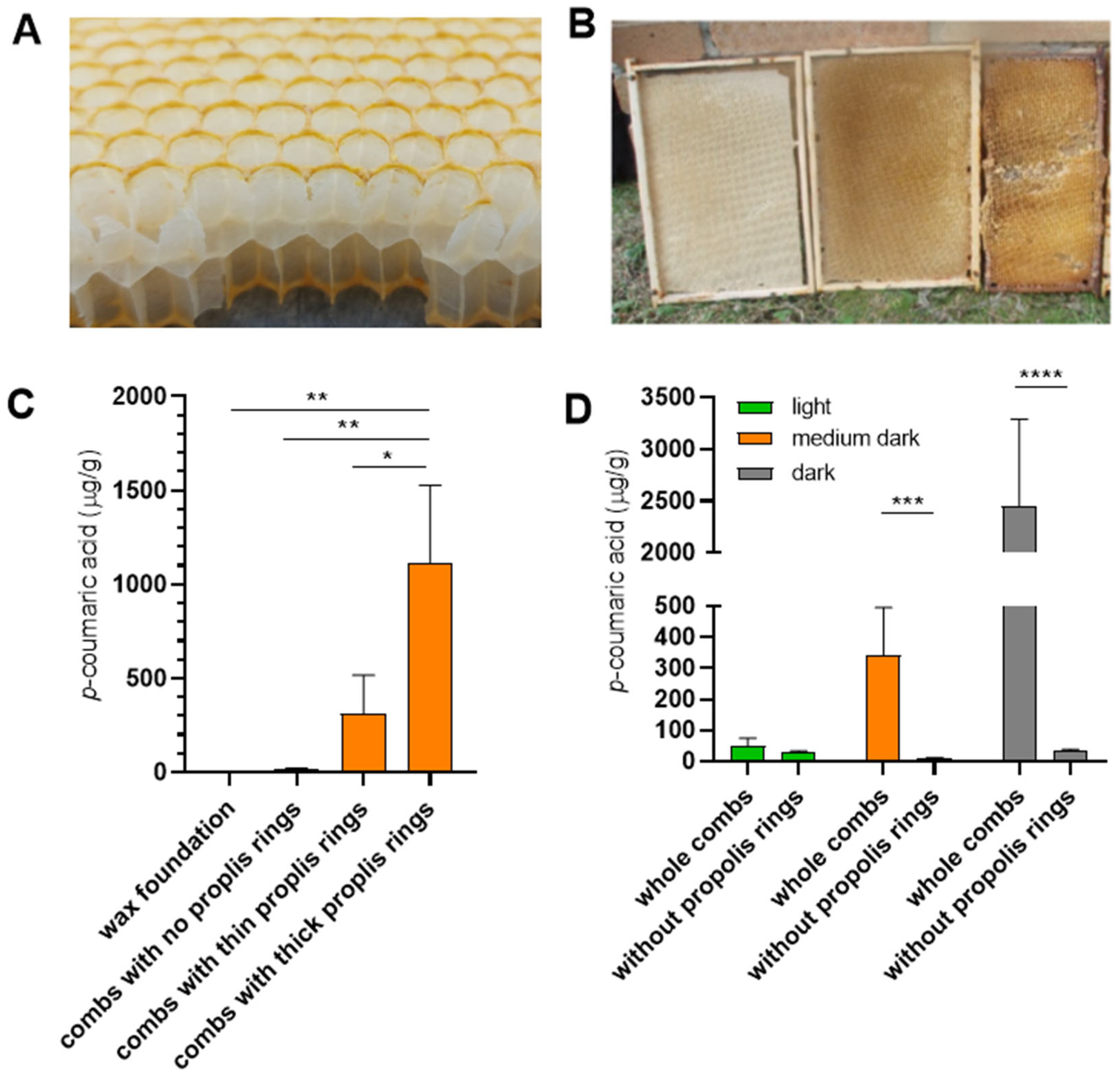
To further investigate this hypothesis, we examined the p-coumaric acid content of empty wax combs. Most of the wax combs dedicated for saccharide storage are marked by a propolis ring at the cell rims (Figure 3A and [24]), and the thickness of these rings correlates with the apparent darkness of the combs (Figure 3B). Empty combs with thick propolis rings contained significantly higher levels of p-coumaric acid than combs with thin or absent rings (Figure 3C). Mechanical removal of the propolis rings resulted in a marked reduction of p-coumaric acid content in the empty combs (Figure 3D). The data support the view that propolis incorporated in the comb structure may serve as a source of p-coumaric acid for saccharides stored by bees.
Finally, we directly tested whether p-coumaric acid from propolis could diffuse into saccharide stores in an in vitro experiment. Elevated levels of p-coumaric acid were detected in both honey and sucrose syrup already within one week of incubation with propolis and its concentration remained stable over extended period of three months (Figure 4A,B). The resulting p-coumaric acid concentrations reached 10-20 µg/g that is comparable to the concentrations detected in sucrose stores deposited by bees (compare with Figure 2).
Figure 2.
The content of p-coumaric acid in honey and sucrose syrup stores originating from the same colonies. (A) Honey was extracted in July and stored in glass jars. Sucrose syrup was fed to colonies in August. Empty combs were inserted in suppers just before feeding, freshly capped stores were labelled and left in the colonies until spring when the assays were performed. Significance according to paired two-tailed t-test. (B) In other colonies the freshly capped sucrose syrup stores were removed in summer, stored at room temperature in the laboratory and the levels of p-coumaric acid were analyzed 2 weeks after supplementary feeding in summer, then again in autumn and the spring. Apiary 3: Doubravice; apiary 6: České Budějovice 2; apiary 7: České Budějovice 2B. Significance according to two-way Anova with Sidak’s multiple comparisons test. (C) p-Coumaric acid levels were analyzed in honey extracted from storage combs of varying colours. Significance according to two-tailed t-test. Error bars represent standard deviations of biological triplicates, except in panel A where honey samples from each colonies were analyzed in single measurements.
Figure 2.
The content of p-coumaric acid in honey and sucrose syrup stores originating from the same colonies. (A) Honey was extracted in July and stored in glass jars. Sucrose syrup was fed to colonies in August. Empty combs were inserted in suppers just before feeding, freshly capped stores were labelled and left in the colonies until spring when the assays were performed. Significance according to paired two-tailed t-test. (B) In other colonies the freshly capped sucrose syrup stores were removed in summer, stored at room temperature in the laboratory and the levels of p-coumaric acid were analyzed 2 weeks after supplementary feeding in summer, then again in autumn and the spring. Apiary 3: Doubravice; apiary 6: České Budějovice 2; apiary 7: České Budějovice 2B. Significance according to two-way Anova with Sidak’s multiple comparisons test. (C) p-Coumaric acid levels were analyzed in honey extracted from storage combs of varying colours. Significance according to two-tailed t-test. Error bars represent standard deviations of biological triplicates, except in panel A where honey samples from each colonies were analyzed in single measurements.
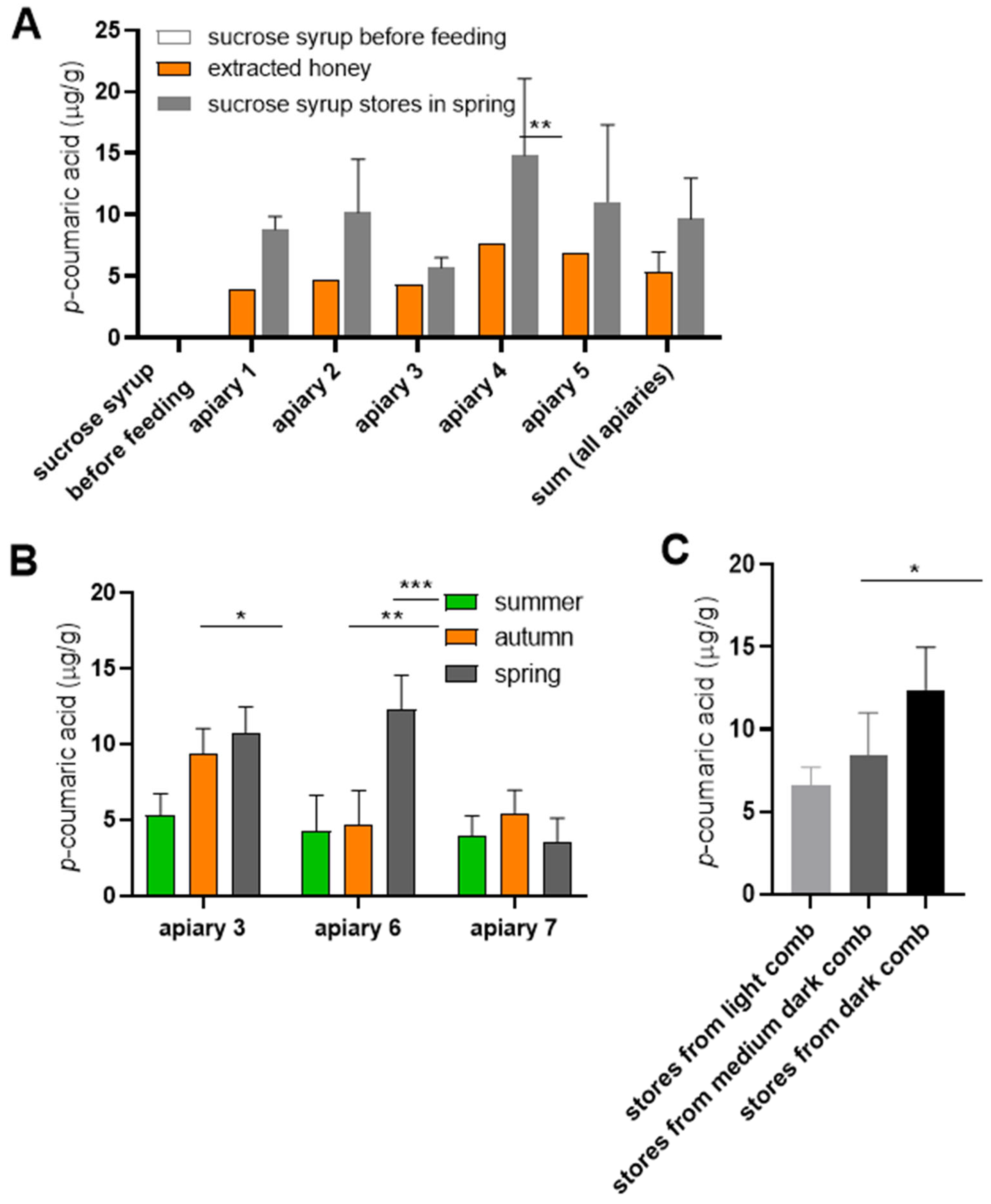
Figure 3.
The extraction of p-coumaric acid from empty storage combs differing in the presence of propolis rings and in their colour. (A) Cross section of a comb dedicated for storage of honey or sucrose syrup. The edges of wax cells are marked by a propolis ring. (B) These rings can be of various thickness, making the combs appear darker in colour. (C) p-coumaric acid levels in empty storage combs built without the use of wax foundations, differing in the intensities of their propolis rings. Significance according to one-way Anova and Tukey’s multiple comparisons test. (D) p-coumaric acid levels in combs depicted in (B) before and after the wax cell edges with propolis rings were removed. Significance according to one-way Anova and Tukey’s multiple comparisons test. Error bars represent standard deviations of biological triplicates.
Figure 3.
The extraction of p-coumaric acid from empty storage combs differing in the presence of propolis rings and in their colour. (A) Cross section of a comb dedicated for storage of honey or sucrose syrup. The edges of wax cells are marked by a propolis ring. (B) These rings can be of various thickness, making the combs appear darker in colour. (C) p-coumaric acid levels in empty storage combs built without the use of wax foundations, differing in the intensities of their propolis rings. Significance according to one-way Anova and Tukey’s multiple comparisons test. (D) p-coumaric acid levels in combs depicted in (B) before and after the wax cell edges with propolis rings were removed. Significance according to one-way Anova and Tukey’s multiple comparisons test. Error bars represent standard deviations of biological triplicates.
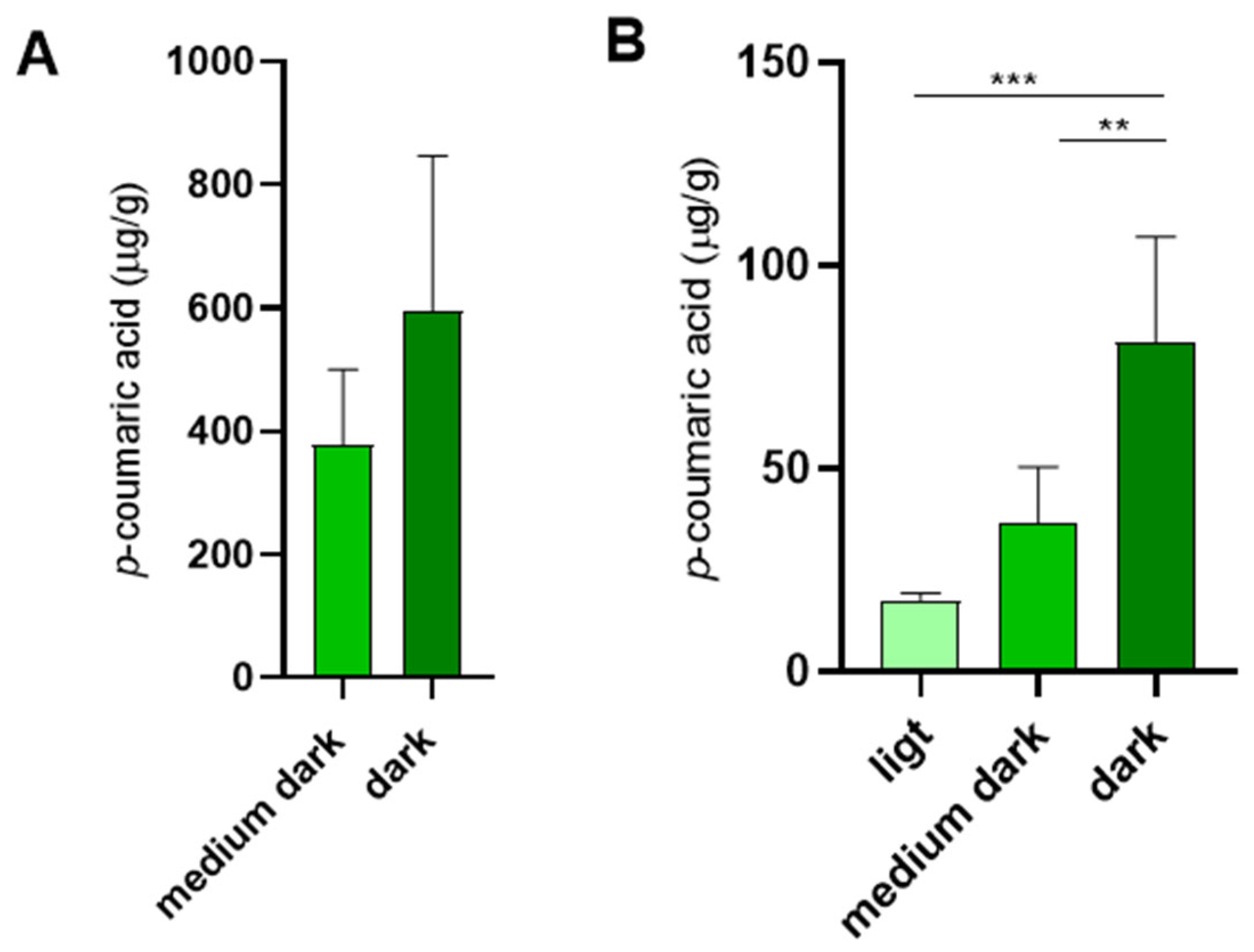
The use of propolis is not limited to combs designated for saccharide storage. Although brood combs lack the prominent propolis rings observed in storage combs, they are characterized by thin internal propolis linings along the cell walls [25,26]. We detected p-coumaric acid also in empty brood combs, in amounts comparable to those detected in medium dark empty storage combs (Figure 5A).
In addition to its other roles in promoting honey bee health, dietary p-coumaric acid has been shown to suppress ovary development in worker bees [9]. This may be one of the reasons why royal jelly fed to queens contains minimal levels of p-coumaric acid [27], as confirmed in our study (Figure 1B). We further examined whether p-coumaric acid is present in queen cells after the queen emergence. Indeed, p-coumaric acid was detected in empty queen cells, with its concentrations positively correlating with the intensity of queen cell pigmentation (Figure 5B).
Figure 4.
Diffusion of p-coumaric acid from propolis to honey and sucrose syrup. Propolis wax mixed with honey (A) or sucrose syrup (B) and incubated for indicated times in a dark incubator set at 35°C to mimic hive conditions. Significance according to one-way Anova and Tukey’s multiple comparisons test. Error bars represent standard deviations of biological triplicates.
Figure 4.
Diffusion of p-coumaric acid from propolis to honey and sucrose syrup. Propolis wax mixed with honey (A) or sucrose syrup (B) and incubated for indicated times in a dark incubator set at 35°C to mimic hive conditions. Significance according to one-way Anova and Tukey’s multiple comparisons test. Error bars represent standard deviations of biological triplicates.
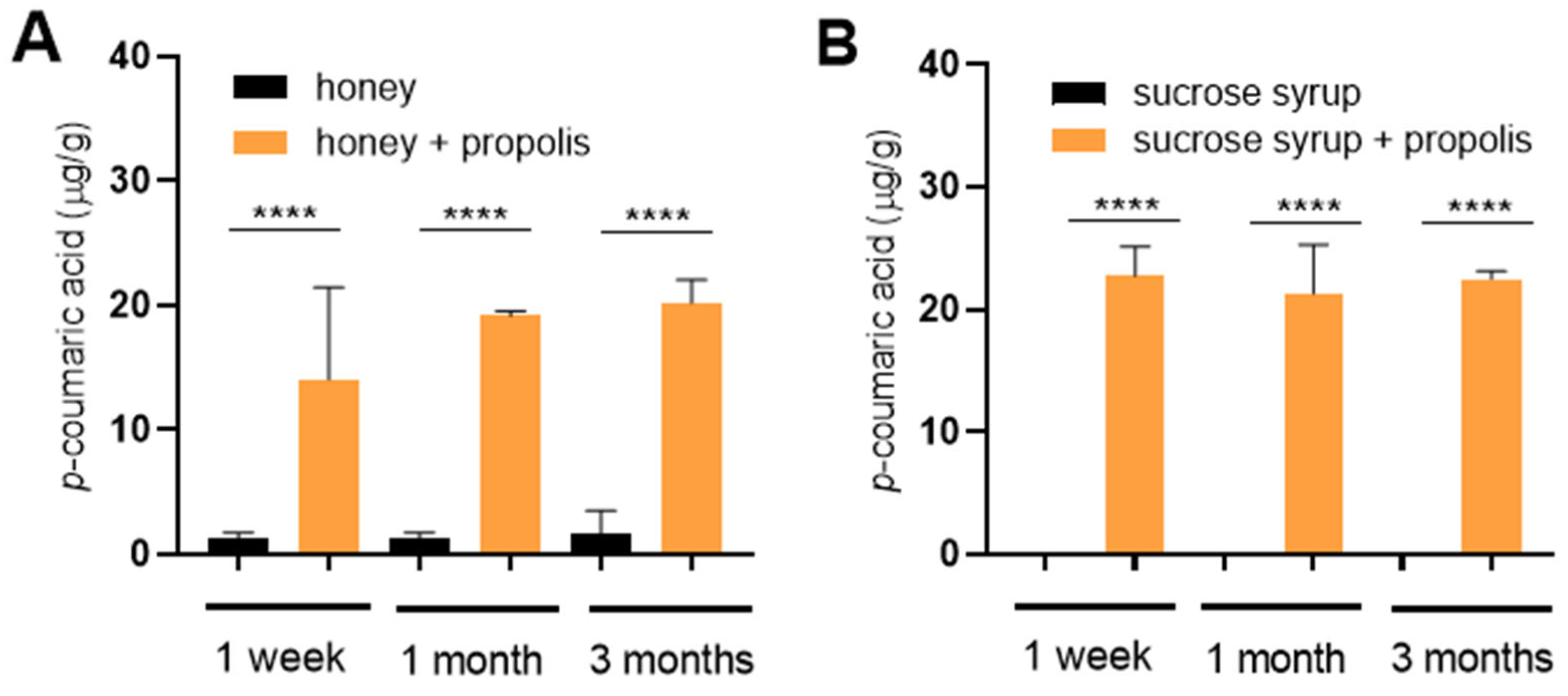
Figure 5.
The levels of p-coumaric acid in empty brood combs and empty queen cells. (A) p-coumaric acid levels in empty brood combs of various colours dedicated for rearing young worker bees. (B) p-coumaric acid levels in queen cells of various colours after queen emergence. Significance according to one-way Anova and Tukey’s multiple comparisons test. Error bars represent standard deviations of biological triplicates (A) and quintuplicates (B).
Figure 5.
The levels of p-coumaric acid in empty brood combs and empty queen cells. (A) p-coumaric acid levels in empty brood combs of various colours dedicated for rearing young worker bees. (B) p-coumaric acid levels in queen cells of various colours after queen emergence. Significance according to one-way Anova and Tukey’s multiple comparisons test. Error bars represent standard deviations of biological triplicates (A) and quintuplicates (B).
4. Discussion
p-Coumaric acid is a plant-derived metabolite that constitutes an important dietary component for honey bees through consumption of pollen and honey [28]. As bees lack the ability to synthesize p-coumaric acid, they rely entirely on its external sources. While concentrations vary depending on botanical origin, the levels of p-coumaric acid in pollen and honey generally fall within a similar range [29,30,31], a finding corroborated by our own data.
The richest source of p-coumaric acid and other beneficial polyphenols in the hive is propolis [32,33]. Derived from plant exudates such as tree buds, sap, and resins, propolis exhibits strong antimicrobial and antioxidant properties due to its high content of phenolic acids, flavonoids, terpenes, and other bioactive compounds [34]. While direct consumption of propolis by bees has not been documented in vivo, laboratory studies indicate that its ingestion can enhance immune responses and improve pathogen resistance [35]. Within the hive, bees apply propolis to comb surfaces and hive walls [24], presumably for antiseptic and structural purposes. Our data provide novel and unexpected evidence that bees utilize propolis as an indirect dietary source, whereby p-coumaric acid (and likely other polyphenols) diffuses from propolis-coated comb surfaces into the saccharide stores deposited within the comb cells.
Supplementary feeding of honey bees with sucrose syrup or other artificial saccharides has raised concerns about potential p-coumaric acid deficiencies that could negatively affect honey bee immunity and overwintering success [5,16]. Our findings contradict this hypothesis. We demonstrate that sucrose-based winter stores, after processing and storage by bees, contain p-coumaric acid in concentrations comparable to or even exceeding those in natural honey. These results indicate that the physiological and colony-level impairments observed under sucrose feeding regimes cannot be attributed to a deficiency of p-coumaric acid.
The mechanism underlying the enrichment of sucrose stores with p-coumaric acid may be explained by two non-mutually exclusive pathways. First, our model proposes that the p-coumaric acid originates from propolis that bees deposit on comb surfaces. Second, bees may actively contribute to the enrichment process during nectar or syrup processing by adding salivary gland secretions known to influence the chemical profile of stored honey [36]. Given that bees consume pollen during syrup processing, it is possible that p-coumaric acid from the ingested pollen is excreted into the processed syrup. However, as no direct evidence supporting this mechanism has been reported to date, our model appears more plausible.
Our data suggest that p-coumaric acid is not an inherent component of freshly secreted beeswax, as wax foundations did not contain it. Instead, the acid appears to accumulate in the saccharide stores over time, likely due to diffusion from propolis deposits in the combs. Despite its low water solubility, we show that p-coumaric acid can diffuse into aqueous sucrose solution or honey. This highlights the role of comb-associated propolis as a reservoir for phenolic compound enrichment of bee food stores.
Beyond its antimicrobial and antioxidant properties, p-coumaric acid is implicated in caste differentiation. As worker bee larvae consume pollen as part of their diet, they are exposed to this acid that has been shown to repress ovary development [9]. In contrast, queen-destined larvae are fed exclusively on royal jelly lacking p-coumaric acid [27]. Despite the importance of avoiding the exposure of p-coumaric acid to the developing queens, we detected this acid in queen cells. However, the continuous consumption of royal jelly by the developing larva likely limits its exposure to the p-coumaric acid, mitigating any adverse developmental effects. Similarly, while worker brood cells are coated with propolis, the primary source of p-coumaric acid for developing worker larvae likely remains dietary pollen [37].
In conclusion, our study provides new insights into the origin and dynamics of p-coumaric acid in the hive environment. We demonstrate that both natural and supplementary sucrose stores are enriched with this bioactive compound via comb-associated propolis. This challenges previous assumptions about nutrient deficiencies in artificial feeding regimes and highlights the multifaceted role of propolis as a nutritional source within the hive.
5. Conclusions
Our results indicate that propolis is the primary contributor of p-coumaric acid enrichment in honey bee saccharide stores. Passive diffusion from propolis rich combs shapes the phytochemical profile of both honey and supplemental feeds. Consequently, supplementary feeding deposited in combs are not devoid of p-coumaric acid but acquire levels comparable to those found in natural honey.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
VK: PU and AK designed the experiments, participated in their executions and in data interpretation, VK collected experimental samples, PU processed and measured all material, AK wrote the original draft of the manuscript, AK, PU and VK edited the manuscript and approved the final version.
Funding
The work was supported by the grant CZ.02.01.01/00/23_021/0009529, co-funded by the EU under the OP JAK programme, administered by the Ministry of Education, Youth and Sports of the Czech Republic.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Acknowledgments
We are grateful to Antonín Hosnedl, Aleš Křenek and Aleš Mikšátko for kindly providing samples of honey bee products.
Conflicts of Interest
The authors declare no conflicts of interests.
References
- TEHAMI, Wafâa, Abdelhafid NANI, Naim A. KHAN a Aziz HICHAMI. New Insights Into the Anticancer Effects of p -Coumaric Acid: Focus on Colorectal Cancer. Dose-Response [online]. 2023, 21(1) [cit. 2025-05-26]. ISSN 1559-3258. Dostupné z:. [CrossRef]
- ABAZARI, Mohammad Foad, Navid NASIRI, Shohreh Zare KARIZI, et al. An Updated Review of Various Medicinal Applications of p-Coumaric Acid: From Antioxidative and Anti-Inflammatory Properties to Effects on Cell Cycle and Proliferation. Mini-Reviews in Medicinal Chemistry [online]. 2021, 2021-11-11, 21(15), 2187-2201 [cit. 2025-05-26]. ISSN 13895575. Dostupné z:. [CrossRef]
- FALCÃO, Éverton Paredes, Danielle da Nóbrega ALVES, Susiany Pereira LOPES, Josy Goldoni LAZARINI, Pedro Luiz ROSALEN, Damião Pergentino DE SOUSA a Ricardo Dias DE CASTRO. Synthesis, antimicrobial activity, and toxicological evaluation of a p -coumaric acid derivative as a potential new antibacterial agent. Journal of Applied Microbiology [online]. 2025, 2025-04-01, 136(4) [cit. 2025-05-26]. ISSN 1365-2672. Dostupné z:. [CrossRef]
- SHARMA, Shashi, Anurag KHATKAR a Saloni KAKKAR. A Review: p-Coumaric Acid, A Medicinally Important Phenolic Acid Moiety. Indo Global Journal of Pharmaceutical Sciences [online]. Indo Global Journal of Pharmaceutical Sciences, 2021, 11(02), 120-129 [cit. 2025-07-29]. ISSN 2249-1023. Dostupné z:. [CrossRef]
- MAO, Wenfu, Mary A. SCHULER a May R. BERENBAUM. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proceedings of the National Academy of Sciences [online]. 2013, 2013-05-28, 110(22), 8842-8846 [cit. 2024-10-22]. ISSN 0027-8424. Dostupné z:. [CrossRef]
- MITTON, Giulia Angelica, Nicolás SZAWARSKI, Francesca Maria MITTON, Azucena IGLESIAS, Martín Javier EGUARAS, Sergio Roberto RUFFINENGO a Matías Daniel MAGGI. Impacts of dietary supplementation with p-coumaric acid and indole-3-acetic acid on survival and biochemical response of honey bees treated with tau-fluvalinate. Ecotoxicology and Environmental Safety [online]. 2020, 189 [cit. 2024-10-22]. ISSN 01476513. Dostupné z:. [CrossRef]
- LIAO, Ling-Hsiu, Wen-Yen WU a May BERENBAUM. Impacts of Dietary Phytochemicals in the Presence and Absence of Pesticides on Longevity of Honey Bees (Apis mellifera). Insects [online]. 2017, 8(1) [cit. 2024-10-22]. ISSN 2075-4450. Dostupné z:. [CrossRef]
- BERNKLAU, Elisa, Louis BJOSTAD, Alison HOGEBOOM, Ashley CARLISLE a Arathi H. S. Dietary Phytochemicals, Honey Bee Longevity and Pathogen Tolerance. Insects [online]. 2019, 10(1) [cit. 2024-10-22]. ISSN 2075-4450. Dostupné z:. [CrossRef]
- MAO, Wenfu, Mary A. SCHULER a May R. BERENBAUM. A dietary phytochemical alters caste-associated gene expression in honey bees. Science Advances [online]. 2015, 2015-08-07, 1(7) [cit. 2024-10-22]. ISSN 2375-2548. Dostupné z:. [CrossRef]
- KOSTIĆ, Aleksandar Ž., Yusuf Can GERCEK a Nesrin Ecem BAYRAM. Phenolic Acids in Pollen. In: ECEM BAYRAM, Nesrin, Aleksandar Ž. KOSTIC a Yusuf CAN GERCEK, ed. Pollen Chemistry & Biotechnology [online]. Cham: Springer International Publishing, 2023, s. 103-125 [cit. 2024-10-22]. ISBN 978-3-031-47562-7. Dostupné z:. [CrossRef]
- NEŠOVIĆ, Milica, Uroš GAŠIĆ, Tomislav TOSTI, et al. Polyphenol profile of buckwheat honey, nectar and pollen. Royal Society Open Science [online]. 2020, 7(12) [cit. 2024-11-08]. ISSN 2054-5703. Dostupné z:. [CrossRef]
- STEVENSON, Philip C., Susan W. NICOLSON, Geraldine A. WRIGHT a Jessamyn MANSON. Plant secondary metabolites in nectar: impacts on pollinators and ecological functions. Functional Ecology [online]. 2017, 31(1), 65-75 [cit. 2024-11-08]. ISSN 0269-8463. Dostupné z:. [CrossRef]
- PŘIDAL, Antonín, Jan MUSILA a Jiří SVOBODA. Condition and Honey Productivity of Honeybee Colonies Depending on Type of Supplemental Feed for Overwintering. Animals [online]. 2023, 13(3) [cit. 2024-10-08]. ISSN 2076-2615. Dostupné z:. [CrossRef]
- SEVERSON, D. W. a E. H. ERICKSON. Honey Bee (Hymenoptera: Apidae) Colony Performance in Relation to Supplemental Carbohydrates1. Journal of Economic Entomology [online]. 1984, 1984-12-01, 77(6), 1473-1478 [cit. 2024-10-08]. ISSN 1938-291X. Dostupné z:. [CrossRef]
- BRODSCHNEIDER, Robert, Rudolf MOOSBECKHOFER a Karl CRAILSHEIM. Surveys as a tool to record winter losses of honey bee colonies: a two year case study in Austria and South Tyrol. Journal of Apicultural Research [online]. 2015, 2015-04-02, 49(1), 23-30 [cit. 2024-10-08]. ISSN 0021-8839. Dostupné z:. [CrossRef]
- QUINLAN, Gabriela, Mehmet Ali DÖKE, Yarira ORTIZ-ALVARADO, Norma RODRIGUEZ-GOMEZ, Yilmaz Berk KORU, Robyn UNDERWOOD a Michael SIMONE-FINSTROM. Carbohydrate nutrition associated with health of overwintering honey bees. Journal of Insect Science [online]. 2023, 2023-11-01, 23(6) [cit. 2024-10-08]. ISSN 1536-2442. Dostupné z:. [CrossRef]
- HU, Yueyang, Fangming LU, Heyan YANG, Qizhong PAN, Xiaobo WU a Ramesh SAGILI. Effect of artificial sugar supplement on the lifespan and learning memory ability of honey bee (Apis cerana) worker bee offspring. Journal of Economic Entomology [online]. 2024, 2024-08-09 [cit. 2024-10-08]. ISSN 0022-0493. Dostupné z:. [CrossRef]
- WANG, Hongfang, Chunlei LIU, Zhenguo LIU, Ying WANG, Lanting MA a Baohua XU. The different dietary sugars modulate the composition of the gut microbiota in honeybee during overwintering. BMC Microbiology [online]. 2020, 20(1) [cit. 2024-10-08]. ISSN 1471-2180. Dostupné z:. [CrossRef]
- FERNANDES, Kenya E., Elizabeth A. FROST, Emily J. REMNANT, Kathleen R. SCHELL, Nural N. COKCETIN a Dee A. CARTER. The role of honey in the ecology of the hive: Nutrition, detoxification, longevity, and protection against hive pathogens. Frontiers in Nutrition [online]. 2022, 2022-7-25, 9 [cit. 2024-10-08]. ISSN 2296-861X. Dostupné z:. [CrossRef]
- SILICI, Sibel, Kemal SARIOGLU, Mahmut DOGAN a Kevser KARAMAN. HPLC-DAD Analysis to Identify the Phenolic Profile of Rhododendron Honeys Collected from Different Regions in Turkey. International Journal of Food Properties [online]. 2014, 2014-01-14, 17(5), 1126-1135 [cit. 2024-10-08]. ISSN 1094-2912. Dostupné z:. [CrossRef]
- BECERRIL-SÁNCHEZ, Ana L., Baciliza QUINTERO-SALAZAR, Octavio DUBLÁN-GARCÍA a Héctor B. ESCALONA-BUENDÍA. Phenolic Compounds in Honey and Their Relationship with Antioxidant Activity, Botanical Origin, and Color. Antioxidants [online]. 2021, 10(11) [cit. 2024-11-08]. ISSN 2076-3921. Dostupné z:. [CrossRef]
- PYRZYNSKA, Krystyna a Magdalena BIESAGA. Analysis of phenolic acids and flavonoids in honey. TrAC Trends in Analytical Chemistry [online]. 2009, 28(7), 893-902 [cit. 2024-11-05]. ISSN 01659936. Dostupné z:. [CrossRef]
- MOHDALY, Adel A.A., Awad A. MAHMOUD, Mohamed H.H. ROBY, Iryna SMETANSKA a Mohamed Fawzy RAMADAN. Phenolic Extract from Propolis and Bee Pollen: Composition, Antioxidant and Antibacterial Activities. Journal of Food Biochemistry [online]. 2015, 39(5), 538-547 [cit. 2024-11-05]. ISSN 01458884. Dostupné z:. [CrossRef]
- BORBA, Renata S., Karen K. KLYCZEK, Kim L. MOGEN a Marla SPIVAK. Seasonal benefits of a natural propolis envelope to honey bee immunity and colony health. Journal of Experimental Biology [online]. 2015, 2015-01-01 [cit. 2024-11-19]. ISSN 1477-9145. Dostupné z:. [CrossRef]
- MURRAY, Stephanie K., Colin M. KURKUL, Andrew J. MULARO, Vanessa L. HALE, Rachelle M. M. ADAMS a Reed M. JOHNSON. Antibacterial effects of propolis and brood comb extracts on the causative agent of European Foulbrood (Melissococcus plutonius) in honey bees (Apis mellifera). Journal of Apicultural Research [online]. 2023, 2023-08-08, 62(4), 813-822 [cit. 2024-11-19]. ISSN 0021-8839. Dostupné z:. [CrossRef]
- EVANS, Jay D. a Marla SPIVAK. Socialized medicine: Individual and communal disease barriers in honey bees. Journal of Invertebrate Pathology [online]. 2010, 103, S62-S72 [cit. 2024-11-19]. ISSN 00222011. Dostupné z:. [CrossRef]
- XUE, Xiaofeng, Liming WU a Kang WANG. Chemical Composition of Royal Jelly. In: ALVAREZ-SUAREZ, José M, ed. Bee Products - Chemical and Biological Properties [online]. Cham: Springer International Publishing, 2017, s. 181-190 [cit. 2024-11-19]. ISBN 978-3-319-59688-4. Dostupné z:. [CrossRef]
- DONG, Nai-Qian a Hong-Xuan LIN. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. Journal of Integrative Plant Biology [online]. 2021, 63(1), 180-209 [cit. 2024-11-08]. ISSN 1672-9072. Dostupné z:. [CrossRef]
- SPILIOTI, Eliana, Mari JAAKKOLA, Tiina TOLONEN, et al. Phenolic Acid Composition, Antiatherogenic and Anticancer Potential of Honeys Derived from Various Regions in Greece. PLoS ONE [online]. 2014, 2014-4-21, 9(4) [cit. 2024-11-19]. ISSN 1932-6203. Dostupné z:. [CrossRef]
- RAMANAUSKIENE, Kristina, Ada STELMAKIENE, Vitalis BRIEDIS, Liudas IVANAUSKAS a Valdas JAKŠTAS. The quantitative analysis of biologically active compounds in Lithuanian honey. Food Chemistry [online]. 2012, 132(3), 1544-1548 [cit. 2024-11-19]. ISSN 03088146. Dostupné z:. [CrossRef]
- NASCIMENTO, Kelly Souza do, José Augusto GASPAROTTO SATTLER, Luciene Fagundes LAUER MACEDO, et al. Phenolic compounds, antioxidant capacity and physicochemical properties of Brazilian Apis mellifera honeys. LWT [online]. 2018, 91, 85-94 [cit. 2024-11-19]. ISSN 00236438. Dostupné z:. [CrossRef]
- SLAVOV, Anton, Atanas TRIFONOV, Lyudmil PEYCHEV, Stela DIMITROVA, Stela PEYCHEVA, Velitchka GOTCHEVA a Angel ANGELOV. Biologically Active Compounds with Antitumor Activity in Propolis Extracts from Different Geographic Regions. Biotechnology & Biotechnological Equipment [online]. 2014, 2014-04-16, 27(4), 4010-4013 [cit. 2024-11-19]. ISSN 1310-2818. Dostupné z:. [CrossRef]
- VIVAR-QUINTANA, Ana María, María Inmaculada GONZÁLEZ-MARTÍN, Isabel REVILLA a Eddy Valentín BETANCES-SALCEDO. Determination and quantification of phenolic acids in raw propolis by reversed phase high performance liquid chromatography. Feasibility study for the use of near infrared spectroscopy. Journal of Apicultural Research [online]. 2018, 2018-10-20, 57(5), 648-656 [cit. 2024-11-19]. ISSN 0021-8839. Dostupné z:. [CrossRef]
- ANJUM, Syed Ishtiaq, Amjad ULLAH, Khalid Ali KHAN, et al. Composition and functional properties of propolis (bee glue): A review. Saudi Journal of Biological Sciences [online]. 2019, 26(7), 1695-1703 [cit. 2024-11-19]. ISSN 1319562X. Dostupné z:. [CrossRef]
- TURCATTO, Aline Patricia, Anete Pedro LOURENÇO a David DE JONG. Propolis consumption ramps up the immune response in honey bees infected with bacteria. Apidologie [online]. 2018, 49(3), 287-296 [cit. 2024-11-08]. ISSN 0044-8435. Dostupné z:. [CrossRef]
- AL-SHERIF, Aliaa A., Adel M. MAZEED, Mohamed A. EWIS, Emad A. NAFEA, El-Seid E. HAGAG a Ahmed A. KAMEL. Activity of salivary glands in secreting honey-elaborating enzymes in two subspecies of honeybee (Apis mellifera L). Physiological Entomology [online]. 2017, 42(4), 397-403 [cit. 2024-11-19]. ISSN 0307-6962. Dostupné z:. [CrossRef]
- WANG, Ying a Hongmei LI-BYARLAY. Physiological and Molecular Mechanisms of Nutrition in Honey Bees [online]. In: Elsevier, 2015, s. 25-58 [cit. 2024-11-19]. Advances in Insect Physiology. ISBN 9780128025864. Dostupné z:. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



