
Submitted:
03 October 2025
Posted:
06 October 2025
You are already at the latest version
Abstract
Type 2 diabetes mellitus (T2DM) is characterized by uncontrolled increase in blood glucose levels, insulin resistance and metabolic inflammation. Vascular complication in T2DM have the inflammatory nature. Drugs with different mechanisms of action have been developed and used to treat T2DM, initially aimed to control blood glucose levels. Among them sodium-glucose cotransporter 2 inhibitors (SGLT2-i) were developed as specific inhibitors of glucose reabsorption in the kidneys, but along with lowering blood glucose levels, they demonstrated multiple (including non-glycemic) positive effects in the treatment of T2DM related to their beneficial effects on the immune system. SGLT2 inhibitors can reduce the risk of diabetic cardiomyopathy (DCM) and chronic kidney disease (CKD) development in patients with and without diabetes. SGLT2-is improve cardio-renal complications through a number of signalling pathways, including those dependent on the involvement of non-coding RNAs (ncRNAs) and their targets. The best-studied classes of ncRNAs are microRNAs, which are short (are less than 200 bases) RNAs (miRNAs), long non-coding RNAs (lncRNAs) (are more than 200 bases), and circular RNAs (circRNAs). The regulatory effect of ncRNAs has broad physiological significance, and changes in the ncRNAs expression are associated with the pathogenesis of different diseases including T2DM. RNA-seq allowed to construct networks of interactions of lncRNA/circRNA-miRNA-mRNA called competitive endogenous RNA (ceRNA) networks, to identify clinically significant molecular markers, to improve mechanistic understanding of pathogenesis, and to contribute to the development of new diagnostics and therapies. Our review summarizes the role of non-coding RNA in the action of SGLT2 inhibitors in cardio-renal complications in T2DM. We focus on methods of detection, genetics and mechanism of action of non-coding RNA. Specific attention is given to the role of non-coding RNAs in the inflammatory reactions of innate immune cells in relation to the SGLT2 inhibitors.
Keywords:
type 2 diabetes mellitus
; SGLT2 inhibitors
; chronic inflammation
; diabetic cardiovascular complications
; diabetic nephropathy
; non-coding RNAs (ncRNAs)
; signaling pathways
; cardiomyocytes
; cardiac fibroblasts
; endothelial cells
; innate immune cells
; macrophages
; podocytes
; tubular epithelial cells
; mesangial renal cells
; ncRNA genetic variants
1. Introduction
Diabetes mellitus is a group of metabolic diseases characterized by an uncontrolled increase in blood glucose levels. Type 2 diabetes mellitus (T2DM) accounts for 90-95% of all cases of diabetes mellitus [1]. T2DM is characterized by insulin resistance in cells of various tissues, as well as dysfunction of the beta cells of the pancreas islets [2]. Type 2 diabetes is one of the leading causes of death and severe complications, such as blindness, renal and/or heart failure, strokes, and lower limb amputations [1,3,4]. Drugs with different mechanisms of action have been developed and used to treat T2DM, which are initially aimed at controlling blood glucose levels. Metformin is currently used as first-line treatment, which leads to the desired glucose-lowering effect for many patients with T2DM. However, in 30% of cases of patients with T2DM, metformin is ineffective, so other therapeutic agents have been being developed and are used.
Currently, among the many drugs used to treat T2DM, sodium-glucose cotransporter 2 inhibitors (SGLT2i) attract the significant attention. These drugs were developed as specific inhibitors of glucose reabsorption in the kidneys, but along with lowering blood glucose levels, they have shown multiple (including non-glycemic) positive effects in the treatment of T2DM. In particular, it turned out that they downregulate the inflammatory processes and, as a result, reduce the risk of developing micro- and macrovascular complications of diabetes [5,6]. Due to the pleiotropic therapeutic effect of these drugs, they are used to treat many diseases that occur as a complication of T2DM and not only [7,8]. Clinical studies have shown that SGLT2 inhibitors can reduce the risk of diabetic cardiomyopathy (DCM) and chronic kidney disease (CKD) development in patients with and without diabetes [9,10,11]. Studies of the molecular mechanisms have shown that the SGLT2 inhibitors improve cardio-renal complications through a number of signaling pathways, including those dependent on the involvement of non-coding RNAs (ncRNAs) and their targets.
Increasingly functionally studied in healthy state and different pathologies, ncRNAs are molecules that do not have a traditional protein-coding function. The best-studied classes of ncRNAs that can influence gene expression are microRNAs, which are short (less than 200 bases) RNAs (miRNAs), long non-coding RNAs (lncRNAs) (more than 200 bases), and circular RNAs [12,13,14]. MicroRNAs are single-stranded RNA molecules approximately 22 nucleotides long, and the generally accepted mechanism of their action is to bind with the corresponding sites in the 3'-untranslated region (3'-UTR) of messenger RNA (mRNA), which leads to mRNA degradation and translational arrest [15]. LncRNAs are involved in the regulation of gene expression through multiple mechanisms, both at the transcriptional level and at the level of RNA splicing, turnover, and translation. In addition, a number of lncRNAs have been shown to regulate DNA replication or repair [13,14]. Circular RNAs have a closed circular structure formed as one of the splicing products (mainly non-canonical) during the formation of mature mRNA [13]. Circular RNAs, as well as linear lncRNAs, demonstrate potential to modulate gene expression at the transcriptional and post-transcriptional levels [16]. Although the spatial structures of lncRNAs and circRNAs are different, both can act as competitors of mRNAs for binding to miRNAs, allowing them to influence the regulatory functions of miRNAs at the translational level [17,18].
The regulatory effect of ncRNAs has broad physiological significance. These molecules are involved in the processes of cell growth and differentiation, organ and tissue development, apoptosis, inflammation, glucose metabolism, intracellular signaling, etc. [13]. It has also been shown that changes in the expression of non-coding RNAs are associated with the pathogenesis of such significant diseases as cancer, cardiovascular diseases and diseases associated with metabolic disorders, including type 2 diabetes mellitus [18,19,20,21,22,23]. Whole transcriptome sequencing (RNA-seq) provides information on all transcriptional products (lncRNAs, circRNAs, microRNAs, and mRNAs) in selected cells and tissues under conditions of interest [24]. RNA-seq results are successfully used to identify non-coding RNAs and construct networks of interactions of lncRNA/circRNA-miRNA-mRNA (competitive endogenous RNA (ceRNA) networks) associated with the development of T2DM and response to drug therapy, as well as to identify clinically significant molecular markers, which allows deeper insight into the pathogenesis molecular mechanisms and contributes to the development of new approaches to their diagnosis and treatment [16,22,25,26,27,28,29].
In this review, we highlight the current state of candidate and high-throughput studies, genetic and functional research on ncRNAs regulatory role in the T2DM cardio/renal complications development, as well as their involvement in the SGLT2 inhibitors action that prevent T2DM – induced complications on the heart and kidneys.
2. Regulatory non-coding RNAs in T2DM
2.1. Identification of T2DM miRNA Biomarkers
To date, a number of studies have shown the involvement of ncRNAs in the development of T2DM and its complications. Most of studies focused on the individual candidate microRNAs and lncRNAs. As a result, dozens of ncRNAs were identified with a decreased or increased levels in various tissues and cell types both in model animals and in human (liver, kidney and muscle biopsies, pancreatic islets, whole blood, PBMC, CD14+ monocytes) as reviewed elsewhere [30,31,32,33,34,35,36]. Changes in the concentrations of ncRNAs in the blood plasma or serum in T2DM have been identified as well [37,38,39,40,41]. New biomarkers with high precision and specificity are urgently needed to identify high-risk individuals for early and personalized therapy due to the limitations of current detection techniques [40]. Differentially expressed ncRNA circulating in the blood plasma/serum are considered as biomarkers for the prognosis of T2DM and its complications, especially in the early clinical stage, as well as potential targets for therapy [31,38,40,41].
Large-scale analysis by microarray techniques with RT- PCR validation was used for the massive detection of previously unknown ncRNAs associated with T2DM and its complications [39,42,43,44,45,46,47]. Further bioinformatics analysis was applied to identify genes associated with differentially expressed microRNAs and lncRNAs, as well as signaling pathways associated with these genes, potentially involved in the pathogenesis of T2DM. Whole-transcriptome studies (RNA-seq) opened new horizons in the study of the role of microRNAs, lncRNAs, and circular RNAs in the development of T2DM. This approach allows simultaneous identification of the expression patterns of lncRNAs and circular RNAs, microRNAs, and mRNAs, and identification of networks of regulatory interactions of competing endogenous RNAs (ceRNAs) that are crucial for the development of the pathology [22,35,48,49,50,51,52]. The accumulated data indicate that all types of endogenous RNA transcripts are in dynamic interaction, competing for binding with a limited set of microRNAs, forming a ceRNA network [53]. This research field is highly dynamic and rapidly accumulating data bring new knowledge about the role of microRNA, lncRNA and circular RNA in the pathogenesis of T2DM.
2.2. Circulating miRNA as Biomarkers of T2DM
Diabetes is characterized by insufficient function of pancreatic beta cells leading to the relative or absolute insulin deficiency as well as to the reduced sensitivity of peripheral target organs and tissues to insulin that together result in disturbances in glucose, lipid, and protein metabolism [34]. Yang et al summarised miRNAs that are involved in the beta cell proliferation and function, including the insulin synthesis and secretion as follows: miR-375, miR-30a, miR-34a, miR-7a, miR-486-5p miR-17-92 cluster, miR-130a-3p, miR-130b-3p, and miR-152-3p [34]. Insulin resistance (IR), also referred as an impaired insulin sensitivity, was found to be associated with a dysregulation of miR-543 miR-592 miR-15b in the liver, miR-29 family, miR-103 and miR-107 in the adipose tissue.
In a high-fat diet-induced diabetes mouse model, whole-transcriptome analysis of pancreatic islets RNA expression identified novel regulatory axes associated with impaired insulin secretion (miR-6948-5p/Cacna1c, miR-6964-3p/Cacna1b, miR-3572-5p/Hk2, miR-3572-5p/Cckar, and miR-677-5p/Camk2d) and increased pancreatic beta-islet mass (miR-216a-3p/FKBP5, miR-670-3p/Foxo3, miR-677-5p/RIPK1, miR-802-3p/Smad2 и ENSMUST00000176781/Caspase9) [35]. The development of T2DM in this model was associated with regulatory interactions of ceRNA, which was centered on the differentially expressed microRNA 12_4382, miR-1188-5p, miR-216a-3p, miR-3572-5p, miR-670-3p, miR-677-5p, and miR-6964-3p, 15 associated lncRNAs and 38 mRNAs [35].
Recently accumulated results allowed to propose circulating microRNA as potential biomarkers for the pre-diabetic and early diabetes states [41,54,55], T2DM [25,35,55,56,57], as well as biomarkers that discriminate between five previously defined subtypes that characterize the differences in pathophysiological processes in T2DM: severe insulin-resistant diabetes (SIRD), severe insulin-deficient diabetes (SIDD), mild age-related diabetes (MARD), mild obesity-related diabetes (MOD), and mild early-onset diabetes (MEOD) [58] . Sulaiman et al used sequencing techniques to analyze the profile of circulating microRNAs in patients with T2DM, which made it possible to associate each subtype of T2DM with a specific microRNA profile that differs from the profile of healthy individuals. In particular, three increased circulating miRNAs were found to be unique for the SIRD, one for the MARD, nine for the MOD и eighteen for the MEOD. Among miRNAs which levels were decreased, eleven were specific for the SIRD, nine for the SIDD, two for the MARD and one for the MEOD [58].
According to Mahjoob study T2DM development was associated by increased expression of the circulating miR-9, miR-30a-5p, miR-150, miR-9, miR-29a, miR-30d, miR-34a, miR-124a, miR-146a, miR-27a, и miR-320a; along with the decrease of the circulating miR-126, miR-21, miR-103, miR-28-3p, miR-15a, miR-145, miR-375, miR-223 expression [57]. Grieco et al conducted a meta-analysis of the literature, which resulted in the identification of ten circulating microRNAs that can be considered as candidate markers of T2DM (miR-21-5p, miR-15a-5p, miR-24-3p, miR-30d-5p, miR-30c-5p, miR-34a-5p, miR-126-3p, miR-146a-5p, miR-148a-3p, и miR-223-3p) [56]. miR-375 miR-126, miR-7a, and let-7 are the most often reported in T2DM patients that highlights their potential as early T2DM detection markers [41].
Song et al. using RNA-seq found a number of differentially expressed lncRNAs, miRNAs, and mRNAs in the plasma of newly diagnosed T2DM patients [25]. Analysis using GO, PPI network, and lncRNA-miRNA-mRNA network revealed that the differentially expressed mRNAs of SLC25A4, PLCB1, AGTR2, PRKN, and SCD5 genes might be under the control of miR-199b-5p, miR-202-5p, miR-548o-3p, and miR-1255b-5p [25]. The discovered miRNAs potentially associated with T2DM pathogenesis identified in large-scale studies represent potential markers and targets for therapy, which requires further experimental verification.
2.3. LncRNA and circRNA as Biomarkers of T2DM
LncRNAs and circular RNAs have been intensively studied during the last several years in the context of T2DM pathogenesis. LncMALAT1, lncRNA steroid receptor RNA activator (SRA), lncRNA βFaar are involved in the beta cell proliferation and function, while lncRNA MEG3, lncRNA HOTAIR are involved in the insulin resistance [34].
To identify new lncRNAs as potential molecular markers of T2DM Lin et al. used bioinformatics analysis of available transcriptome data and constructed a T2DM-associated ceRNA network consisting of 98 genes, 86 microRNAs and 167 lncRNAs. The highest degree nodes were VEGFA/ESR1, hsa-mir-21 and lncRNA MIR22HG, respectively [43]. Next, a ceRNA subnetwork centered on mTOR was generated, since the mTOR signaling pathway plays a central role in energy metabolism, which consisted out of the mTOR gene, three microRNAs and 15 lncRNAs. The authors showed experimentally that lncRNA-NEAT1 can control the expression of the gene encoding the mTOR signaling protein mlST8 by competing with the corresponding mRNA for binding to miR-181b [43].
Ma Q et al used microarray techniques for comparative analysis lncRNA and mRNA expression in PBMC from patients and healthy donors [45]. In this study, 68 differentially expressed lncRNAs and 74 differentially expressed mRNAs were identified in PBMCs from patients with T2D, their function being in the multiple biological processes associated with diabetes. Results of the study revealed that XR_108954.2 and its target gene E2F2 function in the glucose and glycolipid metabolic pathways during T2DM progression and have potential diagnostic value in T2DM [45]. Ali et al aimed to more accurately determine those at risk of developing prediabetes and progressing to T2DM by combining several preselected circulating RNA biomarkers [54]. They used bioinformatics analysis to select candidate mRNA genes which are related to T2DM, insulin resistance signaling, STimulator of Interferon Gene (STING), and NLR signaling pathways from public microarray databases that allowed to retrieve a set TMEM173 and CHUK mRNAs. Further, the mRNA - miRNA - lncRNA regulatory panel was constructed and the potential biomarkers expression levels were estimated in the blood serum from T2DM, pre-DM and healthy donors [54]. This study identified novel STING/NOD/IR RNA panel (TMEM173, CHUK mRNAs, miR-611 and miR -1976 and lncRNA RP4-605O3.4) as pre-DM and T2DM associated biomarker panel [54].
Su et al performed a systematic review and meta-analysis of human case–control or cohort studies on differential lncRNA expression in T2DM that allowed to conclude that lncRNAs may be promising diagnostic markers for prediabetes, T2DM and its complications [55]. Authors also aimed to validate lncRNA biomarkers for T2DM that are prevalent in animals and humans by using Venn diagram statistics. According to results EPB41L4A-AS1, MALAT1, TUG1, DRAIR, MIAT, Kcnq1ot1 were lncRNAs validated in animals and humans, whereas MALAT1 and NEAT1 were validated in diabetic cells, tissues, and blood as valid diagnostic markers for T2DM [55].
CircRNA CDR1as has been reported to modulate insulin production and transcription in islet cells by inhibiting miR-7 through regulating endogenous target genes Pax6 and Myrip [59]. Circ-Tulp4, circANKRD36, circRNA_008565 are the examples of circRNAs important for the beta cells function and insulin secretion. Insulin sensitivity was found to be regulated in the peripheral tissues by circRNA_0054633, circH19, circRNA_000203 [34].
3. Regulatory Non-Coding RNAs in Diabetic Cardiovascular Complications
3.1. Cardiovascular Complications in T2DM
Emerging studies have shown that risk of adverse cardiovascular events increases several times for T2DM patients compared with clinically healthy individuals [36,52,60]. T2DM can lead to the development of diabetic cardiomyopathy (DCM), heart failure (HF) and myocardial infarction (MI) [36,52]. Epidemiological studies have confirmed the correlation between T2DM markers such as elevated serum glucose, glycated hemoglobin levels, impaired glucose tolerance and increased risk of heart diseases [61,62]. Continuous cascade reactions induced by hyperglycemia and insulin resistance lead to chronic inflammation in T2DM, where innate immunity has a central role [63,64,65,66]. The inflammatory factors produced by monocytes and macrophages in metabolic conditions induce pathological activation of endothelial cells, as well as apoptosis and pyroptosis of cardiomyocytes, that aggravate the progression of DCM [67,68,69,70,71]. DCM can be clinically manifested as a restrictive phenotype which is eventually evolving into HF with preserved ejection fraction (HFpEF) and dilated phenotype which is evolving into HF with reduced ejection fraction (HFrEF) [72]. DCM is a unique cardiomyopathy regarding to the molecular mechanism of its development related to the metabolic dysregulation in the heart [36]. DCM is characterized by changes in the myocardial tissue induced by impaired insulin signalling via dysregulated adenosine monophosphate-activated protein kinase (AMPK), protein kinase C (PKC), and mitogen-activated protein kinase (MAPK) signalling leading to fibrosis by increased collagen deposition and the changed protein structure in extracellular matrix. Fibrotic process is also enhanced by EMT under hyperglycemic conditions, as well as abnormal lipid metabolism with increased fatty acid uptake leads to the development of lipotoxicity, endothelial dysfunction and atherosclerosis.
3.2. MiRNAs in Diabetic Cardiovascular Complications
Increasing number of studies confirms the role of miRNAs in the regulation of pathophysiological alterations associated with diabetic heart injury [31,36,60,73]. Clinical studies highlight the significance of certain miRNAs as potential diagnostic biomarkers for T2DM-associated cardiovascular complications. Recent review by Kura et al summarizes the differentially expressed miRNAs in DCM that are associated with oxidative stress (miRNA-21, miRNA-141, miRNA-1, miRNA-200c, miRNA-503, miRNA-22, miRNA-133a/b, miRNA-144, miRNA-499), inflammation (miRNA-21, miRNA-92a, miRNA-204, miRNA-24, miRNA-146a), cardiac hypertrophy (miRNA-195, miRNA-208a, miRNA-451, miRNA-133a, miRNA-150, miRNA-30c), myocardial fibrosis (miRNA-21, miRNA-122-5p, miRNA-150-5p, miRNA-9, miRNA-503, miRNA-142, miRNA-133a, miRNA-700, miRNA-495), apoptosis (miRNA-1, miRNA-34a, miRNA-195, miRNA-208, miRNA-320, miRNA-21, miRNA-181a, miRNA-30c), mitochondrial dysfunction and metabolic disturbances (let-7, miRNA-141, miRNA-223, miRNA-320, miRNA-30c) [36].
A number of miRNAs have been identified as highly expressed in cardiomyocytes, playing significant roles in the healthy state of the heart and in cardiac pathology, among them miRNA-1, miRNA-16, miRNA-27b, miRNA-30d, miRNA-126, miRNA-133, miRNA-143, miRNA-208, and the let-7 family [36,74,75,76] (Figure 1). For example, abundantly expressed in normal cardiac tissue miR-133a was significantly downregulated in DCM mice. An in vitro study showed that miR-133a attenuated cardiomyocyte hypertrophy induced by hypeglyceamia [77]. MiRNAs such as miR-186-5p, miR-22, and miR-29a are shown to suppress hyperglycemia-induced apoptosis in cardiomyocytes in DCM, thereby potentially offering therapeutic targets to protect cardiomyocytes [78]. In contrast, miRNAs like miR-34a, miR-483, and miR-207 promote apoptosis and exacerbate the loss of cardiomyocytes [78]. Notably, miR-144 exhibits a dual role by targeting different mRNAs, thereby both promoting and suppressing apoptosis depending on the specific context within the hyperglycemic environments [78].
Endothelial cells in the myocardium modulating vascular tone and structure. Certain endothelial-enriched miRNAs have been found to have effects on the cardiac function and remodeling, such as miR-126, miR-92a, miR-155, miR-222, miR-221 [31,36]. EMT plays significant role in the DCM pathogenesis. A number of microRNAs were found to be upregulated in cardiac endothelial cells in DCM as miR-21, miR-92a miR-200c, miR-195-5p miRNA-204, while miR-18a-5p miR-142-3p, miR-222, miR146a, miRNA-24, miR-9, miR-126 were downregulated [31,36,75,79,80] (Figure 1). Interesting circulating prognostic marker is miR-92a, which increases several years before the development of coronary artery diseases in T2DM [57]. Cardiac fibroblasts are highly abundant cells in the myocardium, and are essential for maintaining the heart’s extracellular matrix. In response to stress or injury, they rapidly proliferate and produce excess extracellular matrix proteins, leading to cardiac fibrosis. A number of differentially expressed miRNAs have been shown to be associated with pathological processes in cardiac fibroblasts, among them upregulated miR-1, miR-21, miR-24, miR-122, miR-150-5p and downregulated miR-135b, miRNA-141, miR-495 [31,36,75,80,81] (Figure 1). The involvement of miR-141 in the development of diabetic cardiomyopathy was shown in the work of Che H, et al., who found a decrease in miR-141 levels in STZ-induced mouse T2DM model and in the cultured mouse neonatal cardiac fibroblasts under the influence of high glucose levels [82]. The authors found in primary cardiac fibroblasts under HG condition an increased level of lncRNA MALAT1, which is a sponge for miR-141. The work demonstrated the central role of miR-141 by reducing the expression of NLRP3 and TGF-β1 target genes in the anti-fibrotic effect of the hormone melatonin, since suppression of miR-141 expression abolished this effect in cultured cardiac fibroblasts [82].
It is of note, that within 5 years of diabetic cardiomyopathy onset, increasing cardiac hypertrophy assessed using cardiac magnetic resonance (CMR) was independent of glycemic control and paralleled by the up-regulation of the circulating miR122-5p targeting the extracellular matrix MMP2 gene expression [81]. Jakubik et al summarized published data and generated the network of ncRNAs and their target genes involved in pathophysiology of DCM [31]. Cardiac fibrosis, hypertrophy, oxidative stress, inflammation, apoptosis, autophagy, and pyroptosis were presented. miR-146a and miR-195 appeared to be the most promising miRNAs as regulators in DCM, since they can target at least six different genes and can regulate different biological processes involved in DCM [31].
As a result of high-throughput miRNA studies, the list of potential circulating miRNA markers for DCM has been being increased. Bielska A et al. conducted a miRNA expression profiling study and validated the diagnostic significance of miR-615-3p, miR-3147, miR-1224- 5p, miR-5196- 3p, miR-6732- 3p and miR-548b-3p for DCM, all tested miRNAs showed high diagnostic value. Notably, the miRNAs tested under the study were more effective than the non-specific inflammatory parameters like chemokines CXCL12 and macrophage migration inhibitory factor (MIF) [47]. A unique profile of plasma-derived miRNAs in T2DM patients with CAD was revealed in the recent study of Szydełko et al [39]. Authors used expression profiling of 2,578 miRNAs by microarray techniques that allowed to develop a panel of hsa-miR-4505, hsa-miR-4743-5p and hsa-miR-4750-3p. Authors postulate, that this panel could serve as a novel non-invasive biomarker for T2DM-CAD patients compared with T2DM subjects and controls [39]. For coronary artery diseases (CAD) that develop against the background of T2DM, a panel of circulating microRNAs has been identified, the differential expression level of which may be diagnostically significant [57]. The level of some miRNAs increases in coronary artery disease, for example, miR-1, miR-132, miR-133 and miR-373-3-p [57]. In a recent study, Mansuori et al showed elevated plasma miR-1 and miR-133a levels in 3 groups of patients with myocardial infarction (MI): MI patients, MI patients with pre-diabetes (metformin non-users), and MI patients with diabetes (metformin users) compared with healthy donors [83]. Also, miR-1 and miR-133a levels were lower in metformin-user patients than in non-user patients, indicating a potential mechanism for cardiac protection by the treatment [83].
Recently Yin published a systematic review of the contribution of circulating miRNAs as biomarkers of diabetic complications, based on analysis of the selected 71 studies screened and 75 miRNAs validated [40]. Using KEGG pathway enrichment analysis they demonstrated that in DCM, miR-130, miR-21, and miR-199 are implicated in a considerable number of pathways, among them fatty acid synthesis and metabolism pathway was significantly enriched [40].
3.3. LncRNA and circRNA in Cardiovascular Complications
Recently Accumulating data demonstrate that lncRNAs may participate in the development of cardiac hypertrophy and heart failure in DCM through the regulation of oxidative stress, inflammation, cardiac hypertrophy, myocardial fibrosis, apoptosis, mitochondrial dysfunction and metabolic disturbances [29,62,78,84,85]. LncRNAs associated with diabetic cardiomyopathy according to the rodent DCM models and cultured cells studies inlcude DCRF, DACH1, TINCR, Kcnq1ot1, ZNF593-AS, MALAT1, Airn ZFAS1, H19, GAS5, HOTAIR, Crnde, NEAT1, MIAT, NORAD, AK081284, ANRIL, NONRATT007560.2, MEG3, PVT1 (as reviewed by [62]).
Meng et al. found that lncRNA TINCR was significantly up-regulated in an STZ-induced DCM rat model, promoted cardiomyocyte pyroptosis, and aggravated cardiac dysfunction [86]. They further found that TINCR interacted with NLRP3 and stabilized NLRP3 mRNA, thereby accelerating the initiation and progression of DCM. NON-RATT007560.2 [87], HOTAIR [88] and ANRIL [89] are associated with cardiac remodeling in DCM through the regulation of cardiomyocyte apoptosis and oxidative stress [85]. Another lncRNA, AK081284, was proven to be associated with cardiac interstitial fibrosis via TGF-β1. Zhang et al. discovered that AK081284 mediated the effect of IL-17 on interstitial fibrosis in the diabetic heart [90]. In addition, the lncRNA Crnde, which was primarily increased in CFs with TGF-β stimulation, was found to exert a protective effect against cardiac fibrosis [91].
The lncRNA-miRNA-mRNA axis, which is involved in the pathophysiological processes of cardiomyocytes plays a significant role in instigating and advancing the development of DCM [78]. Wu et al emphasized the advancements related to the lncRNA-miRNA-mRNA axis concerning cardiac apoptosis in the context of DCM progression. They reviewed the data demonstrating the involvement of MALAT1/ miR-181a-5p/ miR-22 axis, GAS5/ miR-138/ miR-320/ miR-126a/b-5p axis, H19/ miR-675 axis, KCNQ1OT1/ miR-181a-5p axis, MIAT/ miR-22-3p axis and AK139328/miR-204-3p axis [78]. Each lncRNA is involved in a number of lncRNA-miRNA-mRNA axes taking part in diabetic heart disease progression. As an example, lncRNA KCNQ1OT1 expression was found to be increased in T2DM patients, high glucose-induced cardiomyocyte model, and diabetic mouse models. KCNQ1OT1 knockdown suppresses pyroptosis in high-glucose-induced AC16 cells and primary cardiomyocytes by affecting miR-214-3p and Caspase-1 levels, and also restores cytoskeletal structure and reduces calcium overload in cultivated cardiomyocytes improving cardiac structure and function [92]. According to Zhao and colleagues KCNQ1OT1 and its target gene miR-181a-5p control cardiomyocyte apoptosis in DCM by influencing the regulation of PDCD4 gene [93]. Recently Leng reported that KCNQ1OT1/circ_0020316-miR-92a-2-5p-MAPK3 regulatory networks were involved in vascular injury in T2DM [94].
High-throughput techniques application allows to discover novel ncRNAs which were not known before to be associated with diabetes pathogenesis. Microarray data from Gene Expression Omnibus (GEO) (GSE26887 dataset) were used to construct a ceRNA network associated with diabetic cardiomyopathy [44]. The results revealed that the lncRNA XIST and its two interacting miR-424-5p, miR-497-5p were involved in the pathogenesis of cardiomyopathic complications in T2DM patients [44]. Recently, high-fat diet (HFD) mice model was used to address the mechanisms that links HFD-induced cardiac injury and myocardial infarction [52]. High-throughput sequencing identified differentially expressed mRNAs and long non-coding RNAs in diabetic mice with MI [52]. Heart tissue lncRNA and mRNA expression profile was obtained that allowed to construct lncRNA-gene network in HFD-treated MI group. Three core lncRNAs AC165273.2, 2310039LI5Rik, and AC160401.1 were identified in the network map and selected for further analysis. Analysis of the shared intersections among these core lncRNAs identified 8 target genes, among them Rapgef5 and Ing1 were involved in proliferation and apoptosis of cardiomyocytes induced by HG/HL with hypoxia/reoxygenation [52].
Recent studies have confirmed that differentially expressed circRNAs have a vital role in diabetic complications [84,95]. Li X et al aimed to identify the expression characteristics of circRNAs in the peripheral blood of CAD patients and T2DM patients [96]. Using microarray analysis of circRNA in RNA samples from whole peripheral blood, followed by verification study on the independent cohort, they identified hsa-circRNA11783-2 as significantly related to CAD and T2DM [96]. Several circular RNAs have been implicated in DCM progression, including circ_000203, circRNA_010567, circHIPK3, CACR, and circCDR1as, all of which were upregulated in DCM development [84,95,97]. CircRNA_010567, circHIPK3, and circRNA_000203 are upregulated in the heart tissues of diabetic mice, and are related to the myocardial fibrosis pathogenesis [98,99]. Hsa_circ_0076631, called caspase-1-associated circRNA (CACR) as well, has elevated expression in the hyperglyceamia-exposed cardiomyocytes and in the patients with diabetes [100]. CircCDR1as was upregulated in DCM hearts of STZ-induced diabetic mice, promoting cardiomyocyte apoptosis [84,95]. Recently Yuan Q have discovered that circRNA mm9_circ_008009 is downregulated in mouse hearts with DCM and in cardiomyocytes treated with the advanced glycation end products (AGEs). They named it as well as its conserved human circular RNA hsa_circ_0131202 as DICAR (the diabetes-induced circulation associated circular RNA). DICAR was able to efficiently inhibit the HG-induced pyroptosis in the primary mouse cardiomyocytes and in the HL-1 and HCM cardiomyocyte cell lines via the interaction with valosin-containing protein (VCP) and blocking of the Med12 protein degradation [101].
Accumulating data support the potential of several ncRNAs as valuable markers of T2DM associated cardiac disorders. Dysregulated ncRNAs have been proved to be involved in the DCM pathogenesis, when studied in patients, as well as in animal models in vivo and cell cultures in vitro. However, the reviewed studies demonstrate sometimes contradictory results due to different human heart pathologies and corresponding animal models under investigation. It is obvious, that further collaborative laboratory and clinical studies are urgently needed to enforce progression in the field.
4. Regulatory Non-Coding RNAs in Diabetic Nephropathy
4.1. Renal Complications in T2DM
Accumulating Normal renal function is the result of cooperative work of a number of highly specialized epithelial, endothelial, immune and interstitial cell types and subtypes [102]. Diabetic nephropathy (DN) is a severe microvascular complication of diabetes, characterized by renal interstitial inflammation and fibrosis. It is developed due to the hyperglycemic (HG) conditions in T2DM, that promotes the formation of glycosylation end products that activate signaling pathways including protein kinase C (PKC), transforming growth factor-β (TGF-β), mitogen-activated protein kinase (MAPK), stress-activated protein kinase/jun N-terminal kinase (SAPK/JNK), Janus kinase/signal transducer and activator of transcription (JAK/STAT3), nuclear factor-κB (NF-κB), PI3K/AKT, Nrf2/ARE, and AMPK [103,104]. These pathological events in the renal cells result in the downstream effects including impaired antioxidant defense (via inhibition of Nrf2), enhanced inflammation (through NF-κB and JAK/STAT3), fibrotic signaling (via TGF-β1 and MAPK components p38, JNK, and ERK), and inhibition of autophagy (through suppressed AMPK and altered mTOR signaling) [103,104].
HG conditions activate renin-angiotensin-aldosterone system (RAAS) that accelerates renal damage by increasing calcium influx into podocytes, stimulating proinflammatory cytokines, matrix metalloproteinase-9 (MMP-9), transforming growth factor-β (TGF-β), and activating macrophages. This also enhances aldosterone secretion, upregulating profibrotic factors and promoting kidney fibrosis [104,105]. DN manifest histologically as thickening of the glomerular and thylakoid membranes, podocyte loss, leading to glomerulosclerosis, renal interstitial inflammation, fibrosis, and progressive renal function decline [103].
4.2. MiRNAs in Diabetic Nephropathy
In both human clinical and animal experimental studies, the altered expression of regulatory RNAs such as miRNAs, lncRNAs, and circRNAs were reported in DN in clinical samples including serum, plasma, whole blood, and tissues [40,106,107,108,109]. Abdelmaksoud et al comprehensively reviewed the role of miRNAs in DN, specifically focusing on their impact on key signaling pathways implicated in DN progression [106]. They summarized the results coming from the primary and cultured renal cells, experiments in animal DN models, as well as clinical trials, indicating that miRNAs regulate several essential signaling pathways including Notch, JAK/STAT, Wnt/β-Catenin, PTEN/PI3K/AKT/mTORC1, and TGF-β-mediated pathways. MiR-145–5p, miR-30 family (miR-30a/b/c/d/e), miR-146a, downregulated the Notch signaling pathway in human podocytes and tubular epithelial cells, thus preventing fibrosis, podocyte damage and inflammation in DN [106,110,111,112]. In contrast, miR-135a activates the Notch pathway in renal cells therefore inhibiting miR-135a causes diabetic rats to have less kidney fibrosis [113]. MiR-150, miR-214, miR-155, miR-33-5p upregulate JAK/ STAT pathway and were shown to be increased in DN, thus inducing its progression [114,115,116]. MiR-20a, miR-30c-5p expression levels were found to be decreased in the renal tissues in of patients with diabetic kidney disease, therefore their overexpression was proposed to provide a potential therapeutic effect in DN by JAK/ STAT suppression [117,118]. The aberrant activation of Wnt/β-catenin pathway results in the podocyte and mesangial renal cells damage, leading to the enhancement of diabetic kidney disease. MiR-27a, miR-21, miR-671–5p, miR-135a/b, miR-221, miR-17–5p, miR-466o–3p, miR-499–5p were shown to play roles in DN models in vivo and in vitro by regulation of Wnt/β-catenin pathway [119,120,121,122]. MiR-214, miR-26a, miR-22, miR-141–3p, miR-21, miR-188–5p, miR-142–5p, miR-155–5p, and miR-181b-5p have been found to be upregulated in different models of DN, inducing PI3K/ PTEN/ AKT/ mTOR signaling pathway activation predominantly by suppressing of PTEN expression [42,106,123,124,125]. TGF-β pathway is strongly linked to DN as far as its activation is associated with renal hypertrophy, inflammation, fibrosis and glomerular epithelial cell injury. Aberrant expression of miR-21, miR-155, miR-146a, miR-92b-3p, miR-27a, miR-135a-5p, miR-192, miR-200a, miR-10a/b, miR-7a-5p, miR-26a lead to renal complications by abnormal regulation of the TGF-β pathway in DN [106,126,127,128,129,130]. MiRNA expression changes in renal cells are summarized in Figure 1.
NcRNA-mediated autophagy is involved in various pathological processes of DN, such as cell apoptosis, inflammation, mitochondrial dysfunction, renal injury and fibrosis. miRNA and lncRNA affect autophagy during DN progression [107]. MiR-217, miR-155, miR-34a, miR-150-5p, miR-1187, miR-7002-5p, miR-218, miR-379, miR-214, miR-22, miR-192, miR-32, miR-543 are listed as autophagy- inhibiting miRNAs in podocytes and tubular epithelial cells, leading to the renal damage in DN [107]. MiR-451 was reported to be downregulated in HG-treated HK-2 cells and db/db mice, contributing to renal injury and fibrosis, as well as increased expression of miR-451 was shown to alleviate these effects [131].
Biomarkers of the early DN state and predictors of the diabetic kidney disease development risk are of importance for the personalized T2DM treatment. Bijkerk R et al. were the first to conduct pre-screening of circulating miRNA profiles in three healthy donors and eight DN patients, followed by a larger study confirming that DN patients had lower circulating levels of 11 miRNAs (miR-25, miR -27a, miR -126, miR -130b, miR -132, miR -152, miR -181a, miR -320, miR -326, miR -340, and miR -660) compared to healthy controls [132].
Recently Yin has published a systematic overview to collect and summarize the current trials of T2DM patient blood miRNAs as promising diagnostic biomarkers for diabetic complications. Authors reviewed a total of 24 differentially expressed miRNAs selected from the literature screened for DN, as follows: hsa-miR-9-3p, miR-15a-3p, miR-16-1-3p, miR-21-3p, miR-25-3p, miR-27a-3p, miR-29c-3p, miR-31-3p, miR-126-3p, miR-130-3p, miR-132-3p, miR-152-3p, miR-192-3p, miR-200a-3p, miR-214-3p, miR-223-3p, miR-320a, miR-326, miR-340-3p, miR-377-3p, miR-378a-3p, miR-451a, miR-574-3p, miR-660-3p [40]. MiR-126 and miR-192 occurred three and four times, respectively, both of these two miRNAs were significantly downregulated in DN patients [40]. According to KEGG pathway enrichment analysis the top three significantly enriched miRNAs in DN were miR-27, miR-25, and miR-29. Notably, T2DM patients with renal and cardiovascular complications have differentially expressed common miRNAs such as miR-29 and miR-126 [40]. Mechanistically, the involvement of miR-29 in the development of glomerular fibrosis may be related to TGF-β/SMAD signaling pathway regulation [133]. MiR-126 can suppress inflammation and ROS production in human umbilical vein endothelial cells (HUVECs) treated by high glucose by modulating the HMGB1 expression [134,135].
Exosomes are cell-generated small extracellular vesicles that carry a number of active biomolecules including microRNAs, which can influence various pathological processes associated with kidney diseases. Plasma exosomes do not normally pass through the glomerular filtration barrier therefore the exosomes present in the urine tend to originate mainly from cells of the urogenital system, reflecting the state of the kidneys [104,136]. There is a large number of discovered exosomal biomarkers that correlate with various stages of DN progression. Urinary exosomal miR-133b-3p, miR-342-3p, and miR-30a-5p were found to be elevated in diabetic patients with normoalbuminuria and increase significantly with the progression to micro- or macro-albuminuria in patients with T2DM, potentially serving as early indicators of DN [104,136]. Exosomal miR-21-5p and miR-23b-3p in the urine were elevated in patients with T2DM and were linked to altered renal function, renal sclerosis and fibrosis [104], and miR-451 was correlated with renal failure [104,136].
4.3. LncRNA and circRNA in Diabetic Nephropathy
Emerging studies discovered lncRNAs involved in DN pathogenesis, however the functional roles of lncRNAs are still under investigation. There is an increasing body of evidence indicating that lncRNAs can act as competing endogenous RNAs (ceRNAs), and play a critical role in the molecular mechanisms of DN [137]. Functional studies demonstrated, that NEAT1/miR-93-5p/CXCL8 and LINC00960/miR-1237-3p/MMP-2 might be potential RNA regulatory pathways to regulate the disease progression of DN [138,139]. Li et al. demonstrated that lncRNA Tug1/PGC1a had renoprotective effect via regulating mitochondrial remodeling and urea cycle metabolites in diabetic mice [140]. LncRNA Erbb4-IR was reported to promote renal fibrosis via inhibiting miR-29b in db/db mice DN model [141]. LncRNA KCNQ1OT1 affects cell proliferation, apoptosis and fibrosis through regulating miR-18b-5p/SORBS2 axis and NF-ĸB pathway in DN model of HG-treated human glomerular mesangial cells (HGMCs) and human renal glomerular endothelial cells (HRGECs) [142]. LncRNA SNHG14 level was elevated in the serum of DKD patients and in HG-induced HK-2 cells [143]. Silencing of lncRNA SNHG14 alleviated renal tubular injury via the miR-483-5p/HDAC4 axis in HG-induced HK-2 cells [143]. Studies in the STZ-induced rat DN model and HG-treated HK-2 and 293T showed that lncRNA MALAT1 promotes renal fibrosis and injury in DN [144].
Cao et al used GEO base microarray datasets to identify the differentially expressed genes (DEGs) between DN samples and control human tubulointerstitial tissues [46]. Enrichment analyses, construction of Protein-protein interaction (PPI) network, and visualization of the co-expressed network between mRNAs and microRNAs (miRNAs) were performed. Finally, they identified CD53, CSF2RB, and LAPTM5 as hub genes of tubulointerstitial lesions in DN. Authors speculated that these genes may be closely related to the pathogenesis of DN and the predicted RNA regulatory pathway of NEAT1/XIST-hsa-miR-155-5p/hsa-miR-486-5p-CSF2RB presents a biomarker axis to the occurrence and development of DN [46]. MiR-155 Increased levels of miR-155 were detected in the human renal glomerular endothelial cells in DN, leading to the expression reduction of the target gene ETS-1, as well as its targets VCAM-1, MCP-1 and caspase-3, inducing an inflammatory response and apoptosis [145]. HG-induced mouse podocytes also showed the involvement of miR-155, the increased expression of which leads to the suppression of the target gene BDNF expression, causing oxidative stress and inflammation in mouse models of DN [146].
Study by Guo M intended to construct a ceRNA network in human renal tissue via bioinformatics analysis to determine the potential molecular mechanisms of DN pathogenesis [147]. Based on the analysis of the ceRNA network, five differentially expressed lncRNAs (DE lncRNAs) (SNHG6, KCNMB2-AS1, LINC00520, DANCR, and PCAT6), five DE miRNAs (miR-130b-5p, miR-326, miR-374a-3p, miR-577, and miR-944), and five DE mRNAs (PTPRC, CD53, IRF8, IL10RA, and LAPTM5) were demonstrated to be related to the pathogenesis of DN [147]. Their findings declared a novel underlying molecular mechanism of SNHG6/miR-944, LINC00520/miR-577, PCAT6/miR-326, DANCR/miR-577, and KCNMB2-AS1/miR-374a-3p signaling axis in promoting DN progression [147]. Recently the clinical value and function of DANCR in DN were further explored by Kuang et al [148]. Expression levels of DANCR in the serum of patients with DN or HG-treated human proximal tubular epithelial cells (HK2) were analyzed, demonstrating that abnormal upregulation of DANCR expression level in DN can induce renal tubular injury via the miR-214-5p/KLF5 axis [148]. In the recent study, Hao J et al established DN models (the STZ-treated mice and HG stimulated HK-2 cells) in order to evaluate the xanthohumol treatment molecular mechanism [149]. LncRNA SNHG10 was downregulated in the renal tissues of DN mice and HG induced HK-2 cells, and xanthohumol inhibited the progression of DN by regulating SNHG10/miR-378b [149].
Significant component of DN pathogenesis is hyperglycemia-induced ROS generation leading to the functional disturbance of different types of renal cells, among them renal proximal tubule epithelial cells, podocytes and mesangial cells being highly important [150]. The involvement of multiple lncRNAs aberrant expression in DN development have been demonstrated for these cell types. Recently Li L et al reviewed the studies of lncRNAs that modulate processes such as epithelial–mesenchymal transition (EMT), fibrosis, proliferation, and cell death, ultimately contributing to the pathological manifestations of DN [150]. Aberrant regulation of NEAT1/miR-27b-3p/ZEB1, Dlx6-os1/EZH2/SOX6 pathways in mesangial cells promotes interstitial fibrosis in DN. Expression changes of CASC2/miR135a-5p/TIMP3, CDKN2B/miR15b-5p/WNT2B, HOTAIR/miR147a/WNT2B, NEAT1/miR-423-5p/GLIPR2 pathways lead to enhanced proliferation of mesangial cells and result in mesangial hypertrophy. Podocyte damage and renal function decrease were associated with MIAT/miR-130b/SOX4, Dlx6-AS1/miR346/GSK-3β, Glis2/miR-328/Sirt1[150] . Tubular epithelial cell injury and death were induced by ANRIL/miR-497/TXNIP, MALAT/miR-23c/ELAVL1, NEAT1/miR-34c/NRLP3, CDKN2B-AS1/miR98-5p/NOTCH2, and ZFAS1/miR-525/SGK1 signaling pathways [150].
According to a systematic review and meta-analysis of human case–control or cohort studies on differential lncRNA expression, 18 lncRNAs validated both in animals and humans in DN were as follows: CYP4B1-PS1–001, SOX2OT, HOXB3OS, NEAT1, GAS5, XIST, Lnc-ISG20, CASC2, PVT1, OIP5–AS1, H19, MEG3, NONHSAG053901, TUG1, CES1P1, MIAT, ANRIL, MALAT1 [151]. Four lncRNAs from the list - NEAT1, CASC2, MALAT1, and ANRIL, have been validated in diabetic cells, tissues, and blood [151].
Recent review by Yu S et al emphasizes the modulation of lncRNA, regulating autophagy during DN progression [107]. LncRNAs AK044604, SPAG5-AS1, SNHG17, NEAT1, Linc279227 were reported to be upregulated in DN mouse models and HG-treated podocytes, thus promoting podocyte apoptosis and kidney injury. In contrast, Gm5524/ Gm15645, SOX2OT, Hoxb3os, XIST enhance the viability of podocytes and reduce kidney fibrosis [107].
A number of studies indicate, that circRNAs take part in DN development and can serve as biomarkers for monitoring DN [97,104,152]. Although most circRNAs have negative effects on DN, certain circRNAs have protective effects against DN. Decreased CircRNA_010383 increases proteinuria and renal fibrosis, while elevated Circ_0125310 and Circ_DLGAP4 promote mesangial cell proliferation and fibrosis. CircHOMER1 increases oxidative stress, inflammation, and ECM deposition [104,152]. Evidence for the roles of circRNAs in DN mainly comes from mesangial cells (MCs), tubular epithelial cells (TECs) and podocytes [95]. For example, investigation by Wang et al 2021 in a mouse DN model and HG-exposed mesangial cells (MCs), have demonstrated that circ_0037128/miR-17-3p/AKT3 axis significantly suppressed cell proliferation and fibrosis in DN pathogenesis [153]. The results published by Hu et al suggested that the abnormal expression of CircRNA_15698/miR-185/TGF-β1 axis in HG-exposed MCs has a role in diabetic renal fibrosis [154]. Mou et al confirmed 18 upregulated circRNAs and 22 downregulated circRNAs in the DN kidney from db/db mice using circRNA-seq [49]. Furthermore, circ_0000491 levels were significantly augmented in DN mice and HG-induced mouse MCs. In addition, circ-0000491 sponged miR-101b and activated TGFβR1, leading to ECM accumulation [49]. Upregulation of circ_0000491 in SV40-MES13 cells by HG was found to stimulate the expression of Hmgb1, which is a target of miR-455-3p. Knockdown of Circ_0000491 suppressed HG-induced apoptosis, inflammation, oxidative stress, and fibrosis by regulating the miR-455-3p/Hmgb1 axis [155]. Accumulating data demonstrate that circRNAs are associated with tubular epithelial cell damage in DN. Wen et al explored the circRNA expression profiles and found that circular RNA actin related protein 2 (circACTR2) was upregulated in HG-exposed HK-2 cells and mediated inflammation and pyroptosis [156]. Knockdown of circACTR2 prevented HG-induced pyroptosis, inflammation and fibrosis of TECs, suggesting that circACTR2 has a vital role in the pathogenesis of DN [156]. Circular RNA of eukaryotic translation initiation factor 4 gamma 2 (circEIF4G2) was elevated in db/db mice and in the HG-induced NRK-52E cells [157]. By contrast, the downregulation of circEIF4G2 mitigated renal fibrosis in DN by sponging miR-218 [157]. Recent evidence also suggested that circRNAs are involved in the in the injury and apoptosis of podocytes in DN. Yao et al found that circ_0000285 was increased in kidney tissues of mouse models of DN and podocytes exposed to HG, leading to inflammation and podocyte injury through sponging miR-654-3p/mitogen-activated protein kinase 6 (MAPK6) [158].
5. Inflammation and Innate Immune Cells in T2DM: The Role of Non-Coding RNAs
A T2DM is characterized by the development of chronic low-grade inflammation in the cardiovascular system, kidneys, adipose tissue and other organs, in which monocyte/macrophages play the leading role [70,159]. In T2DM, monocytes/macrophages have increased ability to adhere to the vessel wall, to migration through the endothelial layer, and to differentiate towards pro-inflammatory M1 macrophages, contributing to the development of macro- and microvascular complications [160,161,162]. Monocytes and macrophages control innate immune responses not only to pathogens, but also to the metabolic waste components using their broad arsenal of scavenger receptors [163,164,165]. They secrete inflammatory cytokines and reactive oxygen species, exacerbating endothelial dysfunction in vascular complications of diabetes, such as cardiomyopathy [166,167], nephropathy [161,168], and retinopathy [162,169]. Hyperglycaemia is known to stimulate inflammatory reactions in monocytes/macrophages, and even to have at least middle-term pro-inflammatory memory linked to the increased expression of S100A9 and S100A10 that have damaging effects on endothelial cells [69,70,170,171]. Key signaling pathways regulating macrophage polarization include NF-κB, JAK/STAT, Notch, and TGF-β/SMAD [166]. However, there are still significant gaps in the understanding the molecular basis of proinflammatory programming of monocytes and macrophages in T2DM, making further investigation urgently needed [167]. Single-cell sequencing (scRNA) of the total pool of peripheral blood mononuclear cells (PBMCs) of three patients with T2DM and three healthy donors provided new insight into immune status and potential immune mechanisms involved in the pathogenesis of T2DM. As a result, a transcriptional map of immune cells was created, 119 DEGs in T cells and 178 DEGs in monocytes circulating in the blood of patients with T2DM were identified [172].
5.1. MiRNAs in Monocytes in T2DM
The study by Wang SS et al explored the connections between T2DM and insulin resistance with expression of stress-related miR-18a and miR-34c using PBMCs of T2DM patients, subjects with impaired fasting glucose (IFG) and healthy individuals [173]. The increased levels of miR-18a and decreased levels of miR-34c in PBMCs were associated with risk of T2DM and IFG. Expression levels of miR-18a and miR-34c were found to significantly correlated chronic-stress associated factors, as cortisol, corticotropin- releasing factor (CRF) and IL-6 levels in the blood [173]. MiR-18a and miR-34c in PBMCs may be important marker of stress reaction and may play a role in vulnerability to T2DM as well as IR [173].
Due to their phenotypic plasticity, macrophages play a role in both inflammation and resolution during cardiac and renal injury in T2DM. Studies demonstrated miRNAs expression changes in the context of monocyte differentiation and pro-inflammatory M1 macrophage activation in T2DM - associated chronic inflammation (Figure 1). MiRNAs were found to be uniquely regulated in human macrophages polarized toward M1 and M2 [174]. MiR-125a-3p, -193b, miR-27a-3p, miR-155-3p, and miR-29b-3p exhibited increased expression during macrophage activation, both in M1 and M2 phenotype, while miR-26a shows decreased expression. MiR-222-3p expression is uniquely upregulated in M2 macrophages but downregulated in M1 ones [175]. Distinct miRNA expression profiles were found in mice for M1 versus M2 macrophages, with miR-181a, miR-155-5p, miR-204-5p, and miR-451 being highly upregulated and miR-125-5p, miR-146a, miR-143-3p, and miR-145-5p significantly downregulated [176]. In mouse models of inflammation-associated T2DM complications miR-155 was shown to be upregulated in M1 polarized macrophages [31]. The aggravation of diabetic cardiomyopathy caused by estrogen deficiency was prevented by treatment with antagomiR-155, which induced M2 macrophages infiltration and ameliorated the structure and function of the heart. It is suggested that miR-155 inhibition therapy could serve as a promising approach to improve cardiac function in DCM [177]. Under HG conditions, the expression of miR-126 in macrophages was downregulated, and overexpression of miR-126 diminished efferocytosis, phagocytic clearance of the apoptotic cells, impairment in macrophages of DCM patients [31].
Increased expression of miR-99a in murine bone marrow-derived macrophages suppresses TNFα production, preventing M1 polarization, and reduces the level of proinflammatory markers in T2DM [178]. The therapeutic potential of miR-99a in db/db mice was evident due to an improved glucose tolerance and insulin sensitivity after miR-99a mimics were administered [178]. Conversely, increased expression of miR-130b, miR-330-5p and miR-495 enhances M1 polarization of peritoneal/adipose-tissue derived macrophages, which has been shown in murine models of high-fat diet induced T2DM [179,180,181]. Aggravated endothelial injury and impaired endothelial repair capacity was found to be associated with overexpression of miR-483-3p in the M2-type macrophages and in the endothelial cells of T2DM patients [182]. Overexpression of miR-483-3p increased endothelial and macrophage apoptosis and impaired reendothelialization in vitro [182]. Significant upregulation of miR-471-3p expression was detected in the diabetic mouse bone marrow-derived macrophages (db/db) and advanced glycation end products (AGE)-treated RAW264.7 cells [183]. Meanwhile, inhibition of miR-471-3p reduced proinflammatory macrophage polarization. Bioinformatic analysis identified SIRT1 (coding NAD-dependent deacetylase sirtuin-1) as a target of miR-471-3p, which was then confirmed in a luciferase assay [183]. Recently a therapeutic role was demonstrated for miR-369-3p in halting diabetes-associated atherosclerosis by regulating GPR91 and macrophage succinate metabolism [184]. In the study by Rawal et al miR-369-3p was found reduced in peripheral blood mononuclear cells from diabetic patients with coronary artery disease (CAD) and in aortic intimal lesions from Ldlr-/- mice on a high-fat sucrose-containing diet. In vitro, oxLDL treatment reduced miR-369-3p expression in mouse bone marrow-derived macrophages (BMDMs) [184].
As mentioned above miR-21 dysregulation was reported in T2DM-associated heart failure with preserved ejection fraction (HFpEF) [185]. Recent study investigated the effects of miR-21-3p on macrophage polarization and mitophagy in the chronic heart failure model of the isoproterenol (ISO)-induced myocardial structural disruption and fibrosis in rats. The ISO-challenged rats displayed increased left ventricular internal diameter systolic (LVIDs), decreased left ventricular ejection fraction (LVEF), which are traditional clinical indicators of HF, as well as elevated serum levels of creatine kinase-myocardial band, a marker of myocardial necrosis. In this model, heart failure was exacerbated by miR-21-3p [186]. MiR-21-3p accelerated M1 macrophage polarization in vivo in chronic heart failure model and in the co-culture system of macrophages and rat cardiomyocytes H9c2 cell line; furthermore miR-21-3p mimics promoted M1 polarization, which exacerbated H9c2 cell damage [186].
5.2. LncRNAs and circRNAs in Monocytes in T2DM
There is also multiple evidence for the role of lncRNAs and circRNAs in diabetes-related inflammatory processes [97]. For example, Reddy et al. showed that the long-ncRNA DRAIR plays an important role in the reducing diabetes-related inflammation [187]. DRAIR suppression leaded to a decrease in the expression of anti-inflammatory genes in the THP1 monocyte-like cells, increased monocyte adhesion to the vessel wall and the expression of pro-inflammatory genes such as IL-1β [187]. In contrast, to DRAIR, lncRNA transcribed from the opposite strand of the dynamin 3 gene (Dnm3os) increased inflammatory responses that contribute to complications in T2DM. Dnm3os expression is significantly upregulated in CD14+ monocytes from different murine diabetes models, leading to the increased expression of proinflammatory genes in macrophages and increased phagocytic activity [188].
Transcriptome analysis of monocytes isolated out of peripheral blood of patients with T2DM and control healthy volunteers revealed two significantly elevated lncRNAs (ENSG00000287255 and ENSG00000289424) and one significantly decreased lncRNA (ENSG00000276603) in patients versus controls [51]. The most likely associated with T2DM were lncRNAs ENSG00000225822 (UBXN7-AS1), ENSG0000028654, ENSG00000261326 (LINC01355), ENSG00000260135 (MMP2-AS1), ENSG00000262097 and ENSG00000241560 (ZBTB20-AS1) [51]. Bioinformatic analysis of differentially expressed genes revealed the networks associated with cell mobility, growth and development [51]. Yang et al. obtained RNA-seq data for PBMCs from three T2DM patients and three healthy individuals, which were used to construct a ceRNA network, demonstrating that lncRNAs and circRNAs play a regulatory role by interacting with multiple miRNAs, and, thus, co-regulating more mRNAs [22]. The authors also showed the important role of differentially expressed lncRNAs and circRNAs in T2DM patients in glucose metabolism, mTOR signaling pathway, lysosomal pathway, and apoptotic pathway [22]. In the study by Ma Q et al., 68 differentially expressed lncRNAs and 74 differentially expressed mRNAs were found using microarray analysis of the PBMC transcriptome of five T2DM patients and paired controls. Searching for lncRNA-mRNA interaction pairs revealed that the target gene of lncRNA XR_108954.2 encodes the transcription factor E2F2, which is involved in the control of the cell cycle, as well as glucose and lipid metabolism [45].
A comparative study of PBMC using RNA-seq revealed 220 circRNAs to be differentially expressed between patients with T2DM and healthy individuals [48]. circANKRD36 levels were upregulated in peripheral blood leukocytes and correlated with chronic inflammation in T2DM [48]. Another study analyzed the circRNA expression by RNA-seq in the polarized human THP-1 macrophage-like cells incubated in M1 (interferon-γ + LPS) or M2 (interleukin-4) conditions [50]. 71 up-regulated circRNAs and 69 down-regulated circRNAs from 9720 detected circRNAs were found in the M1-type THP-1 cells. CircRNF19B expression was significantly up-regulated in M1 conditions [50]. Bone marrow (BM) derived murine macrophages polarized to a pro-inflammatory phenotype (INFγ+TNFα) or anti-inflammatory phenotype (IL-10, IL-4, and TGF-β) were studied to determine whether circRNAs modulate monocyte/macrophage biology and function [189]. Authors used circRNA microarray analyses to assess transcriptome changes, which allowed to choose circ-Cdr1as as playing a key role in regulating anti-inflammatory phenotype of macrophages. Overexpression of circ-Cdr1as increased transcription of anti-inflammatory markers and percentage of CD206+ cells in naïve and pro-inflammatory macrophages in vitro [189].
To summarize, accumulated published data indicate that dysregulated miRNA, lncRNA, and circRNA are extensively involved in the pathogenesis of T2DM and diabetes–induced cardio/renal complications and can act as key nodes in the disease regulatory network. NcRNAs associated with T2DM demonstrating a potential as effective biomarkers have been found in animal models as well as in human studies. Unique as well as common biomarkers for patients and challnaged rodents were found. Notably, despite intensive research in the field, there are still gaps in the understanding of the complex integrating molecular mechanisms of dozens ncRNAs involvement in the T2DM initiation and its major complications development. Therefore, further investigations are urgently needed based on the high throughput RNA sequencing techniques, refined computational analysis, accumulated data integration and construction of T2DM molecular networks, including external inducing signals, internal multiple signaling pathways and target processes under the ncRNA regulation. Developing of the strategy for integrative data analysis is necessary to evaluate potential ncRNA-based biomarkers for clinical practice, to provide novel targets for disease treatment and new drug development.
6. Involvement of ncRNAs in the SGLT2i Effect on Cardiac and Renal Complications of T2DM
6.1. SGLT2i Effect on the Cardiovascular System State and Function
Canagliflozin, dapagliflosin, empagliflosin, ertugliflosin, and sotagliflosin are the five therapeutic agents of a class of SGLT2 inhibitors that have been approved to date by the FDA; they are now being considered as a second-line therapy for the management of T2DM [3,8,190,191]. SGLT2i improve cardiovascular complications in T2DM through a combination of hemodynamic, metabolic, anti-inflammatory, and vascular effects on the myocardium [52]. These drugs reduce inflammation, oxidative stress, apoptosis, lipotoxicity, enhance endothelial function, and promote a shift towards more efficient energy utilization. Studies have explored various cardiac cell types, including cardiomyocytes, endothelial cells, fibroblasts, and smooth muscle cells, as potential targets for the observed cardiovascular benefits of SGLT2 inhibitors [191]. Empagliflozin reduced cardiac fibrosis through modulation of human myofibroblast function, and suppression of the expression of several pro-fibrotic factors, including collagen type I α1, actin alpha 2 (smooth muscle), connective tissue growth factor, fibronectin 1, and matrix metalloproteinase 2 [8,192]. Dapagliflozin inhibited JNK, p38, and FoxO1 signaling pathways in association with the reduction of myocardial interstitial and perivascular fibrosis. The down-regulation of mitogen-activated protein kinase/activator protein-1 signaling by dapagliflozin has been found to be Na+/H+ exchanger 1 (NHE1)-dependent [193]. Dapagliflozin protected against myocardial fibrosis by inhibiting resident fibroblast activation, collagen secretion through suppression of EMT and AMP-activated protein kinase α-mediated inhibition of TGF-β/Smad signaling [194]. Numerous studies evidenced that empagliflosin directly inhibit Na+/H+ exchanger 1 (NHE1) activity in cardiac cells, including cardiomyocytes, endothelial cells, and cardiofibroblasts [195]. A recent study emphasized that dapagliflozin and/or liraglutide attenuated cardiac tissue injury via reducing key elements of oxidative stress, inflammation and apoptosis in diabetes-induced cardiomyopathy rats [196]. Empagliflozin partially exerted anti-oxidative stress and anti-apoptotic effects on cardiomyocytes under high glucose conditions by activating AMPK/PGC-1α and suppressing other RhoA/ Rho-associated protein kinase (ROCK) pathway in diabetes-induced cardiomyopathy mice [197]. In addition to inflammation and oxidative stress, lipotoxicity also plays a critical role in diabetic HFpEF. Sun et al. showed that canagliflozin can attenuate lipotoxicity and inflammatory injury in cardiomyocytes, and can protect diabetic mouse hearts via inhibiting the mTOR/hypoxia-inducible factor-1α (HIF-1α) pathway [198].
SGLT2is improved cardiac energetics and enhanced mitochondrial function by inhibiting cardiac glucose uptake and glycolysis, increasing cardiac glucose oxidation, ketone metabolism and fatty acids metabolism [190,199]. Canagliflozin ameliorated myocardial remodeling in HFpEF rats by optimizing cardiac energy metabolism, enhancing mitochondrial function, and consequently reducing myocardial hypertrophy and fibrosis, with simultaneous improvement of diastolic function [200].
6.2. Effect of microRNA on the Cardiovascular System Condition and Functions Under the Treatment with SGLT2-i
Protective effects of SGLT2 inhibitors action in various states and conditions, including cancer, metabolic diseases and others, by affecting ncRNAs expression were demonstrated in cultured cells, animal models and patient studies for a number of SGLT2 inhibitors [7]. The involvement of non-coding RNAs in the SGLT2 inhibitors-induced cardioprotective effects in T2DM was demonstrated for empagliflozin and dapagliflozin (Figure 1; Table 1).
6.2.1. MiR-21 and miR-92 in the Effect of SGLT2-i
miR-21 plays a pivotal role in cardiac function regulation therefore it has been frequently investigated, taking into account its prospects in clinical therapy. This miRNA is significantly upregulated in conditions such as coronary arterial disease (CAD), cardiac fibrosis (CF) and heart failure (HF) in T2DM patients [201,202,203]. In contrast, according to Tao L the circulating miR-21 levels were significantly decreased in the diabetic cardiomyopathy patients compared to the non-DCM group [204]. The disparate findings on miR-21 in cardiomyopathy suggest that its pathological activity depends on the cell type and on the disease context. Based on the results of the bioinformatics study by Sessa et al, the role of miR-21 in cardiovascular diseases is related to its interaction with a number of hub genes: programmed cell death 4 (PDCD4), RAS guanyl-releasing protein 1 (RASGRP1), and B-cell translocation gene 2 (BTG2) (bioinformatics analysis of the experimental data from CVD patients) [205]. miR-21 is associated with elevated reactive oxygen species (ROS) levels and is involved in the endothelial dysfunction. It targets key genes involved in regulation of ROS production, such as Krev/Rap1 interaction trapped-1 (KRIT1) and superoxide dismutase 2 (SOD2), resulting in increased oxidative stress when overexpressed [206]. Experiments in the cellular and animal models, as well as in patients indicate that miR-21 facilitates the accumulation of superoxide in cells, particularly in the context of high glucose concentrations, which is particularly relevant in diabetic vascular complications [206].
Ridwan et al. in the Wistar rat model of STZ-induced hyperglycemia found a trend of miR-21 decreased expression in the heart under the influence of empagliflozin compared to metformin (Table 1). The authors associate this effect of SGLT2-i with the detected decrease in the TGF-β1 level, and the increased expression of the matrix metalloproteinase-2 (MMP-2) gene, which may reduce the risk of cardiac fibrosis occurrence during treatment with empagliflozin, but not metformin [207]. It should be noted that in an earlier study in a STZ /HFD-induced mouse model of diabetic cardiomyopathy, increased miR-21 expression was associated with promoted fibrosis, decreased autophagy, and attenuated the protective effects of the hypoglycemic drug vildalgliptin, a dipeptidyl peptidase 4 inhibitor, on cardiac function [208]. In this study, miR-21 was also upregulated in the rat cardiomyocyte cell line H9c2 under high glucose condition, which was downregulated by vildalgliptin. The authors showed that pathological changes in the heart under hyperglycemia occur as a result of autophagy reduction through the SPRY1/ERK/mTOR signaling pathway, the activity of which is suppressed by increased levels of miR-21, and normalized by the vildalgliptin therapy [208].
Mone et al. assessed the effect of empagliflozin on the expression of circulating microRNAs involved in the regulation of endothelial function in a group of age-related T2DM patients with heart failure with preserved ejection fraction (HFpEF) [209]. The authors analyzed by RT-PCR a panel of microRNAs previously reported to be involved in the regulation of endothelial dysfunction [209]. Compared with the control group, significantly increased levels of circulating miR-21 and miR-92 were found in HFpEF patients [209]. After 3 months of empagliflozin treatment circulating miR-21 and miR-92 levels in T2DM patients with HFpEF decreased, while they didn’t change in response to metformin or insulin [209]. As noted above, both microRNAs have been previously associated with inflammatory diseases, in particular, heart failure, and may have value as prognostic markers for cardiomyopathy in T2DM. MiR-92a has been found to inhibit heme oxygenase-1 (HO-1), an important enzyme responsible for the degradation of heme to form biliverdin and bilirubin, which have antioxidant properties [210]. When miR-92a was inhibited, HO-1 levels increased, thereby enhancing the antioxidant potential of human umbilical vein endothelial cells and db/db mouse aortas [210].
Notably, in the obese T2DM patients, abdominal fat excision reduced inflammation, normalized left ventricular diastolic function over 1 year of follow-up, these improvements were accompanied by decreased miR-21, miR-92 and increased miR-126 levels in the blood serum [211]. Previously, miR-126 was shown to be involved in the protection of the cardiovascular system by modulating angiogenesis, inflammation, and apoptosis in patients with T2DM, with a number of studies establishing a decrease in the level of this microRNA in the blood serum of patients with cardiomyopathy and T2DM [56,135]. It was shown that miR-126 is expressed at significantly lower levels in the heart of diabetic rats, which is associated with diastolic heart failure [212]. This microRNA is a potential target for the anti-inflammatory effect of SGLT2 inhibitor drugs investigation.
As mentioned above, increased miR-21 and decreased miR-126 were found to induce pro-inflammatory M1 macrophage phenotype in cardiac T2DM disorders, therefore one can propose macrophage pivotal role in the SGLT2i-induced patients health improvements [41,186]. This question has not been examined yet and needs further macrophage-targeted investigation.
6.2.2. MiR-30d in the Effect of SGLT2-i
In the work by Zhang, the therapeutic effect of SGLT2 inhibitors (not specified) on the development of diabetic cardiomyopathy was studied in a rat model on a high-fat diet and with streptozotocin (STZ)-induced diabetes [213]. The use of SGLT2 inhibitors for 10 weeks improved cardiac function as a result of enhancing autophagy by reducing the expression level of miR-30d, which was significantly increased during the development of DCM in this model [213]. In addition, in the case of miR-30d knockdown, cardiac function was also improved. The study showed that SGLT2 inhibitor improves the animal condition with diabetic cardiomyopathy by affecting the miR-30d/KLF9/VEGFA pathway, which regulates the expression of autophagy factors protein light chain 3 (LC3-II) and p62/SQSTM1 (p62/sequestosome 1) [213]. Earlier, Li X (2014) also found an increase in mir-30d levels, which contributed to cardiomyocyte pyroptosis in rats with streptozotocin (STZ)-induced diabetes, as well as in cardiomyocytes treated with a high level of glucose [214]. Pyroptosis is one of the types of programmed cell death, which is accompanied by an inflammatory response, playing a significant role in the pathogenesis of cardiomyopathy. Li X showed that the induction of cardiomyocyte pyroptosis under hyperglycemia under the influence of increased mir-30d expression occurs through an increase in the level of caspase-1 and proinflammatory cytokines IL-1β, IL-18 as a result of a decrease in the expression level of foxo3a and its direct target activity-regulated cytoskeleton-associated protein (ARC) [214].
El Khayari et al. reviewed the important role of miR-30d and other members of the miR-30 family (particularly miR-30a and miR-30e) in the development of T2DM complications on the heart and kidneys [190]. Recent review has summarized the data obtained in the cellular and animal models, as well as in patients, and described the mechanisms underlying the effect of SGLT2 inhibitors and miR-30 family members on endothelial cells and cardiomyocytes. The mechanistic elements targeted by SGLT2 inhibitors and miR-30 family members included transcription factor 21 (TCF21), Toll-like receptor 4 (TLR-4), AMP-activated protein kinase (AMPK) and angiopoietin 2, followed by suppression of nuclear factor-kB (NF-κB) signalling and decreased expression of proinflammatory cytokines and adhesion molecules [190]. Given the mutual influence of SGLT2 inhibitors and miR-30 on inflammation, angiogenesis and endothelial cell survival, critical role of interaction between different cell types, including immune cells seem highly likely. Thus, members of the miR-30 family may be involved in mediating the action of SGLT2 inhibitors in endothelial cells and cardiomyocytes [190]. However, this was not directly demonstrated, and such interaction in immune cells cannot be excluded.
6.2.3. MiR-181a and Empagliflozin
Empagliflozin can significantly mitigate myocardial fibrosis following MI, leading to marked enhancements in cardiac functionality and a decrease in the incidence of ventricular fibrillation, as it was shown in the rat model of myocardial infarction (MI) [215]. Empagliflozin had the potential to ameliorate MI – induced myocardial fibrosis by inhibiting the expression of miR-181a suppressing the TGF-β1/Smad3 signaling pathways in the myocardial tissue [215]. miR-181a was earlier associated with CAD development in T2DM both in patient studies, animal CAD models and HG-induced endothelial cells [181]. Moreover, study by Chen demonstrated that miR-181a upregulation leads to the downregulation of HMGB2 in the angiotensin II - treated primary cardiomyocytes (ex vivo model of cardiac hypertrophy) [216].
6.2.4. MiR-181a and Empagliflozin
Molecular mechanisms of empagliflozin effects were studied in the obese ZSF-1 leptin-receptor knockout rats (heart failure with preserved ejection fraction mode, HFpEF l) and in the obese ZSF-1 rats treated with SU5416 to stimulate resting pulmonary hypertension (obese+sugen, CpcPH model) [217]. In the heart failure with preserved ejection fraction (HFpEF), metabolic syndrome promoted pulmonary vascular dysfunction and exercise-induced pulmonary hypertension (EIPH) by enhancing ROS production and increasing miR-193b levels. This factors had suppressive effect on the nuclear factor Y α subunit (NFYA)-dependent soluble guanylate cyclase β1 subunit (sGCβ1) expression in the pulmonary arterial smooth muscle cells [217]. Application of empagliflozin in CpcPH rat model with combined precapillary and postcapillary pulmonary hypertension ameliorated metabolic syndrome and reduced mitochondrial reactive oxygen species generation, downregulates miR-193b, resulting in restoration of NFYA-sGCβ1-cGMP signaling and improvement of EIPH symptoms [217]. miR-193b was recently identified as a potential marker of T2DM in a large-scale analysis of 1934 miRNAs circulating in the blood plasma of newly diagnosed, drug-naive patients [218]. When miR-193b-3p levels were increased, triosephosphate isomerase (TPI1) levels were decreased in both plasma of T2DM patients and HepG2 hepatocellular carcinoma cells [218]. The multi-omics approach revealed that miR-193b-3p may affect glucose metabolism by directly targeting tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta (YWHAZ)/14-3-3ζ and upregulating the transcription factor forkhead box protein O1 (FOXO1), which is involved in the effects of the PI3K-AKT signaling pathway. Based on the results observed in cultured HepG2 cells derived from liver tissue, it can be speculated that the effects of miR-193b-3p on glucose metabolism in other tissues may be similar [218]. The involvement of differential expression of miR-193b-3p in the development of cardiovascular complications of T2DM and in the implementation of the protective effect of SGLT2 inhibitors seems promising for further investigation.
6.2.5. MiR 30e-5p and miR199a-3p in the Effect of Dapagliflozin
In the study of Solini et al, patients with T2DM and hypertension (blood pressure >130/80 mm Hg) were randomly divided into 2 groups receiving either dapagliflozin or hydrochlorothiazide (HCT) during 4 weeks, their background medication was unchanged during the study [219]. The authors assessed the effect of short-term treatment on endothelial function, systemic and renal vascular function using standard generally accepted instrumental approaches in association with changes in the expression of circulating microRNAs. Dapagliflozin, but not HCT, significantly altered the expression of two microRNAs previously associated with heart failure (miR-30e-5p and miR-199a-3p), suggesting the involvement of these microRNAs in the therapeutic effect of dapagliflozin [219]. As demonstrated earlier, miR-30e-5p, which is selectively up-regulated by dapagliflozin, inhibits myocardiocyte autophagy through modulation the angiotensin-converting enzyme 2 (ACE2) pathway in doxorubicin-induced heart cardiomyopathy [220]. Increased expression of miR-30e-5p was associated with antihypertrophic and antiapoptotic effects on angiotensin II (Ang-II)-stimulated hypertrophic cardiomyocytes [221]. In contrast, miRNA-199a is a prohypertrophic miRNA, and its overexpression in mouse cardiomyocytes induced hypertrophy and suppressed autophagy [222]. The reduced expression of miR-199a-3p induced by dapagliflozin might be also intriguing in mechanistic terms, as far as antagonizing miR-199-3p could derepress cardiac peroxisome proliferator-activated receptor delta (PPARδ) levels, thus restoring mitochondrial fatty acid oxidation and improving cardiac function in the failing heart [223].
6.2.6. Potential Role of miR-141 in the Effect of SGLT2-i
Li et al. demonstrated that SGLT2 mRNA is a direct target of miR-141, so miR-141 overexpression reduced the development of cardiac fibrosis in a rat myocardial infarction model by reducing the expression level of SGLT2 [224]. The study revealed an increase in the expression level of SGLT2 in association with a decrease in the expression of miR-141 during the development of fibrosis after the induction of myocardial infarction in rats via the permanent ligation of the left anterior descending branch of the coronary artery, and SGLT2 knockdown improved the condition of the heart muscle, as did an increase in miR-141 expression [224]. The results of this work open the question about possible synergistic effect of SGLT2 inhibitors and miR-141 in the anti-fibrotic effect in cardiomyopathy.
6.3. SGLT2i Effect on the Renal System
Protective effects on renal function, which is impaired during the development of T2DM, have been demonstrated in cultured cells and in vivo models for several SGLT2 inhibitors [3,7,225,226]. A literature review by Castoldi G, 2024 summarized the data demonstrating that SGLT2 inhibitors also effectively improve kidney state and function in various non-diabetic kidney disease models [227].
In the diabetic kidney disease models, SGLT2 inhibitors like dapagliflozin, empagliflozin and canagliflozin have shown significant anti-inflammatory, anti-oxidative, and anti-fibrotic properties. They reduced calcification in vascular smooth muscle cells, inhibited epithelial–mesenchymal transition, and delayed fibrosis by modulating pathways such as YAP/TAZ, TXNDC5, and NLRP3 [7,225,228,229]. Dapagliflozin, empagliflozin, and canagliflozin have demonstrated pleiotropic vascular protective effects beyond the glycemic control with benefits observed both in patients and the animal models. These agents mitigate mitochondrial ROS production, suppress NOX4 expression, activate the AMPK/SIRT1/PGC-1α axis, preserve mitochondrial membrane potential, and enhance eNOS phosphorylation. All these mechanisms contribute to the improvement of the endothelial function and reduction of arterial stiffness [226,230]. Furthermore, SGLT2 inhibitors impact kidney function by regulating cell migration, reducing oxidative stress, and mitigating the effects of high glucose levels on the proximal tubule cells, demonstrating high potential as therapeutic agents for diabetic nephropathy [225]. SGLT2 inhibitors have been shown to impact kidney proximal tubule cell migration by modulating various cellular pathways involved in cell motility [231]. Empagliflozin exerted the nephroprotective effects in high-glucose-induced cultured kidney proximal tubule cells in vitro [232]. Dapagliflozin may delay tubulointerstitial fibrosis in diabetic kidney disease, inhibiting YAP/TAZ activation in tubular epithelial cells [229]. Anti-fibrotic and anti-ferroptotic effects of dapagliflozin in DN mouse model have been associated with its interaction with the Nrf2 and TGF-β signaling pathways [228]. Recent whole-transcriptome study of the kidney tissue by Wu et al indicated that empagliflozin can improve the outcome of DN in the mouse T2DM model by inhibiting inflammation, apoptosis, senescence, and other related pathways [233]. The KEGG enrichment of upregulated genes in the DN and DNE groups was concentrated in the p53 pathway, which was related to apoptosis and senescence, and the aggregated peroxidase proliferators activate receptor-related (PPAR) pathway, which was the main inflammatory pathway [233].
6.4. Effect of microRNA and lncRNA on the Renal System Condition and Functions Under SGLT2-i Treatment
Protective effects on the impaired renal function in T2DM were demonstrated in cultured cells, animal models and patient studies for SGLT2 inhibitors canagliflozin, dapagliflozin, empagliflozin, irtugliflozin and sotagliflozin. The involvement of several non-coding RNAs in the SGLT2 inhibitors-induced nephroprotective effects was demonstrated for empagliflozin and dapagliflozin (Figure 1; Table 1).
6.4.1. MiR-21 in Effect of DN Therapy by Empagliflozin and Dapagliflozin in Different Models
Empagliflozin exerted a protective effect on human renal proximal tubular cells (HK-2) cultured in the presence of high glucose in vitro [232]. Empagliflozin attenuated HG-induced oxidative stress, matrix metalloproteinase (MMP2) activation and RECK suppression which led to stimulation of fibrosis in the kidneys of db/db mice by activating EMT and renal proximal tubular cells migration [232,234]. Das et al showed that in HG-treated HK-2 cells the advanced glycation end products (AGE) activated their receptor RAGE, causing the increased expression of TRAF3-interacting protein 2 (TRAF3IP2). Next, through the activation of NF-κB, p38MAPK led to an increase in the expression of miR-21 and suppression of the RECK expression, which is a direct target of miR-21 [232]. Thus, downregulation of miR-21 expression is one of the pathways mediating the therapeutic effect of empagliflozin on the renal proximal tubular cells in vitro. Recently Al-Tantawy SM et al studied antidiabetic effect of the treatment by dapagliflosin in combination with gold nanoparticles (AuNPs) in the streptozotocin-induced rat model of diabetic nephropathy [235]. The treatment for 7 weeks significantly increased catalase activity, serum total antioxidant capacity, along with a substantial decline in malondialdehyde indicating the alleviation of DN [235]. Autophagy activation, inhibition of apoptosis, and renal fibrosis was detected in renal tissue which could be due to their inhibitory impact on miR-192 and miR-21 renal expression [235].
Protective effect of dapagliflozin was examined in the gentamicin-induced renal injury in Wistar rats, and improvement in kidney function and structure as found to be associated with an increase of miR-21 and a decrease in miR-181a levels in the kidney tissue [236]. Dapagliflozin administration decreased markers of kidney injury, such as increased creatinine, blood urea, and urine protein. The authors suggest that miR-21 and miR-181a expression changes in the kidney tissue by dapagliflozin lead to a decrease in renal tubular cell apoptosis [236]. Thus, SGLT2 inhibitors normalize abnormal miRNA expression in renal tissue in different pathological states: both chronic metabolic stress- associated with increased miR-21 levels, and acute drug-induced injury associated with decreased miR-21 levels.
6.4.2. MiR-21 in Effect of DN Therapy by Empagliflozin and Dapagliflozin in Different Models
Since ncRNAs appear to be promising predictive biomarkers for therapy efficacy, there is great interest in identifying novel differentially expressed ncRNAs using large-scale analysis. To assess the pharmacological effects of empagliflozin therapy on the kidneys, Shepard used a mouse model of early T2DM associated with obesity and insulin resistance [237]. The TallyHo/Jng mouse line, derived from Swiss-Webster, is a polygenic model of progressive insulin resistance leading to the development of T2DM on a high-fat diet [237]. The kidney function was assessed using a number of criteria, including urine microRNA expression profiling using large-scale microarray analysis, and changes of several identified microRNAs were verified by PCR analysis [237] (Shepard BD, 2024). Empagliflozin treatment resulted in improved renal function parameters and changes in urinary exosomal microRNA profiles, including increased levels of miR 27a-3p, 190a-5p, and 196b-5p. Network analysis identified “cancer pathways” and “FOXO signaling” as the main pathways regulated by the drug [237]. An earlier study by An Y et al. found that increased levels of another member of the miR-196 family (miR-196a) in the urine of patients with diabetic nephropathy was a prognostic marker for kidney disease progression [238]. Four-week treatment with dapagliflozin of patients with hypertension and T2DM demonstrated a beneficial effect on the ability of kidneys to dilate blood vessels in association with significant up-regulation of circulating miR-30e-5p and downregulation of miR-199a-3p, which are involved in the pathophysiology of CV disease and heart failure [219]. Of note, the nephroprotective effect correlated with the pretreatment expression levels of circulating miR-27b and miR-200b [219].
6.4.3. LncRNA, circRNA and miRNA Identification and Study in DN Treatment with Dapagliflozin and Empagliflozin
The effect of dapagliflozin was studied using genome-wide sequencing and ceRNA network construction in a diabetic nephropathy (DN) model (db/db mice), which allowed comparative analysis of the expression of lncRNAs and mRNAs in the kidney tissue of healthy mice, db/db mice, and dapagliflozin-treated db/db mice [238]. Db/db mice were characterized with glucose intolerance, high urinary albumin/creatinine ratio and renal damage, which were found improved in the dapagliflozin group after 12-week treatment. Large-scale RNA sequencing identified 172 differentially expressed lncRNAs between DN and control mice, and 40 differentially expressed lncRNAs between DN without and with dapagliflozin. In these two pairwise comparisons, 7 common differentially expressed lncRNAs were found (NR_015554.2, XR_382492.3, XR_382493.3, XR_382494.3, XR_388840.1, XR_873495.2, and XR_876705.2).
The results of quantitative RT-PCR showed that dapagliflozin could reverse the expression changes of XR_382492.4, XR_873495.3, XR-388840.1, NR-015554.2, XR-382493.3, and XR-876705.2 associated with diabetic nephropathy [238]. The authors used the results of RNA sequencing and network pharmacology analysis of therapeutic pathways and targets to identify potential mechanisms of dapagliflozin effect in DN. As a result, the involvement of TGF-β signaling pathways, peroxisome proliferator-activated receptor (PPAR) signaling pathway, chemokine signaling pathway, and a number of other pathways that affect 11 key genes (SMAD9, PPARG, CD36, CYP4A12A, CYP4A12B, CASP3, H2-DMB2, MAPK1, MAPK3, C3, IL-10) in the action of dapagliflozin was revealed [238]. It should be noted that the comprehensive analysis of the results revealed for the first time the involvement of the SMAD9, CYP4A12a, CYP4A12b and H2-DMB2 genes in DN, while these genes were previously unknown potential targets for dapagliflozin in DN [238].
Recently Wu T. et al. conducted a comprehensive study of the mouse transcriptome in a model of induced diabetic nephropathy in mice treated with streptozocin to determine the dynamics of various types of ncRNAs and construct a ceRNA network of the response to empagliflozin [233]. They experimentally identified differentially expressed mRNAs in kidney tissue as a result of two pairwise comparisons: between the negative control and diabetic nephropathy, and between DN and DN with empagliflozin administration. A total of 232 genes were found downregulated and 282 genes upregulated for both comparisons. The authors focused on genes encoding proteins involved in metabolism, for which protein-protein interaction networks were constructed, while all data on lncRNAs and miRNAs that could potentially interact with them were obtained from the corresponding databases [233]. Construction of the ceRNA network allowed to identify the β-1,4-mannosyl glycoprotein 4-β-n acetylglucosamine transferase (Mgat3), ST8α-n-acetylceramide α-2,8-sialic transferase 1 (St8sia1), and glucosamine-6-phosphate deaminase 1(Gnpda1) genes as central ones. The role of the associated key microRNAs miR-466i-3p, miR466f-3p, miR-709, miR-207, and miR-297b-3p in DN and in the empagliflozin effects was not recognized, and requires further investigation [233].
Song et al. studied the mechanisms of dapagliflozin action on cultured human renal proximal tubular epithelial cells HK-2 under HG conditions [239]. The results obtained using RNA microarray hybridization were used to construct ceRNA networks and functional analysis, which revealed the involvement of multiple circRNAs, their target miRNAs, and proteins in the action of dapagliflozin on renal proximal tubular cells. The experimental validation identified the hsa_circRNA_012448-hsa-miR-29b-2-5p-GSK3β pathway involved in oxidative stress response as an important pathway mediating the action of dapagliflozin in the treatment of diabetic kidney disease [239]. This finding is in accordance with the data from Wang et al, which demonstrated that miR-29 exhibited an anti-fibrotic effect in the renal tissue [240].
6.5. Effect of SGLT Inhibitors on Immune Cells in T2DM: Involvement of ncRNAs
One of the key mechanisms of SGLT inhibitors that underlay their cardioprotective and renoprotective effects in patients with T2DM is reduction of chronic low-grade inflammation [225,240]. Recently Shen et al performed high-throughput analysis and demonstrated that pathways related to immune response and metabolism pathways were critically involved in diabetic nephropathy progression and reversed by dapagliflozin [241] (Table 1). ATAC-seq, RNA-seq, and weighted gene correlation network analysis has been performed to assess the chromatin accessibility in combination with mRNA expression in the comparative study of kidney tissue from mouse diabetic nephropathy model: db/m mice (Controls), db/db mice (DN), and db/db mice treated with dapagliflozin (DN treatment group) [241]. Combined analysis of the data identified some candidate genes (including UDP glucuronosyltransferase 1 family, Dock2, Tbc1d10c) and transcriptional regulators (KLF6 and GFI1) that might be associated with DN and dapagliflozin restoration [241]. However, specificity of the action of these genes in the distinct cell types was not addressed. In particular, it would be of interest to examine the mechanistic role of KLF6 in kidney macrophages. There are data that KLF6 is involved in the macrophage polarisation and, non-coding RNA regulate KL6 pathways in macrophages. For examples. miR-21-5p carried by adMSC-derived apoptotic bodies was shown to KLF6 leading to M2 polarization in skin wound healing [242]. In liver fibrosis induced by carbon tetrachloride in mice, miR-148a regulated intrahepatic macrophage functions through KLF6/STAT3 signalling [243]. Another study used the mode for the sterile kidney inflammation kidney based on the ischemia/reperfusion-induced acute kidney injury (IRI-AKI). In this model, formononetin (a natural, O-methylated isoflavone of a plant origin) significantly improved renal function and reduced inflammation in IRI-AKI, and the authors attributed these effects to the inhibition of KLF6/STAT3-mediated macrophage pro-inflammatory activation [244].
Intensive investigation of molecular mechanisms of SGLT inhibitors action on macrophages, key innate immune cells, suggests that anti-inflammatory effect of inhibitors relies on the attenuation of inflammasome activity and inhibition of TLR4/NF-κB [225,240]. The anti-inflammatory effect is also related to the changes in the activity of other signaling pathways (AMPK, PI3K/Akt, ERK 1/2-MAPK, and JAKs/STAT) in macrophages [225]. Recent study investigated the inhibitory effect of dapagliflozin on macrophage M1 polarization and apoptosis in the murine model of streptozotocin- induced diabetic nephropathy [245]. Experiments ex vivo demonstrated, that dapagliflozin attenuated M1 macrophage polarization and mitigated apoptosis in the abdominal macrophages of diabetic mice, by the suppressing the phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) signaling pathway (PI3K, AKT, phosphorylated protein kinase B) [245]. However, the role of ncRNAs in SGLT inhibitors effects on the innate immune cells remains poorly investigated, and only isolated reports are available.
Abdollahi et al. studied the effect of dapagliflozin on human macrophages differentiated out of peripheral blood mononuclear cells and activated by LPS [246]. Addition of dapagliflozin reduced the M1/M2 ratio and LPS-induced TLR-4 overexpression in macrophages under high and low glucose conditions, and also suppressed the high glucose-induced activation of the NF-κB signaling pathway. Dapagliflozin treatment abolished the LPS-induced change in the expression of two microRNAs, miR-146a (anti-inflammatory) and miR-155 (pro-inflammatory), both are known to play a role in the regulation of inflammation through activation of the NF-κB signaling pathway [246]. Earlier in a model of STZ-induced DN in wild-type mice miR-146a expression increased in both peritoneal and intrarenal macrophages, playing anti-inflammatory role [247]. Also, miR-146a deficiency during STZ-induced diabetic nephropathy in miR-146a−/− mice led to the increased expression of M1 activation markers and to the suppression of M2 markers in renal macrophages. Mechanistically, miR-146a suppresses expression of its target genes Irak1 and Traf6 in renal macrophages. IRAK1 and TRAF6 are known promote NFκB-mediated upregulation of proinflammatory cytokines, such as IL-1β, IL-18, and TNF-α, inhibiting inflammatory response [247]. miRNA-155 is also involved in innate immune response, playing a pro-inflammatory role [80]. The overexpression of miR-155 promoted the activation of the ox-LDL-induced NLRP3 inflammasome in PMA-preprocessed THP-1 macrophages, thus aggravating the carotid atherosclerotic lesion in ApoE-/- mice [248]. Repressing expression of miR-155 while promoting that of miR-146a, dapagliflozin can effectively modulate inflammatory responses preceding T2DM-induced vascular complications development.
7. Genetic Variants in Non-Coding RNA in T2DM
T2DM and diabetic complications are multifactorial diseases with a strong genetic component [249]. To date, genome-wide association studies (GWAS) have identified around 700 variants (mainly SNPs) associated with predisposition to these diseases [250]. Information about SNPs associated with ncRNAs regulatory action in the development of T2DM also comes from sources independent of GWAS. Summarized, the largest part of already discovered SNPs affects mRNA binding sites interacting with microRNAs due to complementarity, and less part pertains to substitutions in the seed sequence of the microRNAs [251].
7.1. T2DM-Associated Variants in miRNAs
In a study by D. Jahantigh and colleagues, an association was found between the C rs4705342 and C rs4705343 alleles, localized in the promoter region of the MIR143/145 microRNA gene cluster [252], with an increased and decreased risk of developing T2DM, respectively (Table 2).
Moreover, individuals with the rs4705342-CC genotype have significantly higher levels of low-density lipoprotein cholesterol, fasting blood glucose, glycated hemoglobin, and miR-143 expression than carriers of the rs4705342-TT genotype [253]. It has previously been shown that obesity-induced increased expression of miR-143 impairs the ability of insulin to activate the phosphatidylinositol 3-kinase (PI3K)/Akt pathway in the liver and muscle tissue, causing insulin resistance and impaired glucose metabolism [254]. In addition, miR143 is involved in the process of autophagy inhibition, which can also lead to insulin resistance [255]. In turn, miR145 may play a protective role in the pathogenesis of T2DM due to decrease of chronic inflammation. For example, in the study by He M, miR145 expression in PBMC of T2DM patients was significantly lower than in the control group [256]. Inhibition of miR145 by intravenous administration of an antisense oligonucleotide to C57BL/6 J mice stimulated macrophage infiltration into the liver and their polarization into a proinflammatory phenotype. Restoration of miR145 expression resulted in decreased weight in mice, decreased fasting blood glucose levels, improved glucose tolerance, and reduced macrophage infiltration into the liver [256]. Thus, rs4705342 and rs4705343 are promising candidates for further study of their role in regulating miR-143/miR-145 expression and mechanisms of T2DM development.
In the work of Sun et al., an association was established between rs8089787 and rs10877887, located in the promoter regions of miR-133a-1 and let-7f, respectively, and impaired diastolic cardiac function in T2DM [257]. It has been demonstrated that let-7f mimetics can suppress the expression level of IGF1R (insulin-like growth factor 1 receptor), whereas let-7f inhibitors promote the increase in the expression level of IGF1R [258]. In addition, let-7f expression levels have been shown to be decreased in individuals with T2DM compared to healthy controls [259]. In turn, the expression level of miR-133a is higher in individuals with T2DM compared to healthy individuals. Moreover, a positive correlation was observed between the expression level of miR-133a and the levels of triglycerides, total cholesterol, fasting blood glucose and glycated hemoglobin [260]. An association with an increased risk of T2DM has also been established with alleles of non-coding variants: rs1292037-C and rs13137-T, located in the downstream region of miR-21 (-281 bp and -404 bp from the transcription start site, respectively) [261], and rs13283671-C in the downstream region of miR-31 (-444 bp from the transcription start site) [262]. The rs1076064-G allele in the upstream region of miR-378a (+222 bp from the transcription start site) has a protective role in T2DM [262].
Some studies demonstrate discordant results. For example, study of Khan MS analyzed volunteers from Pakistan, and found association of the rs11614913 and rs2910164 polymorphisms located in the stem-loop miR-196a-2 (MIR196A2) and seed sequence miR-146a (MIR146A) with T2DM [263]. It was shown that the C allele of rs11614913 and G allele of rs2910164 are associated with an increased risk of developing T2DM [263]. In contrast, other studies reported an association of the T allele of rs11614913 with an increased risk of developing cardiovascular diseases in patients with T2DM [264], and the G allele of rs2910164 with a reduced risk of developing T2DM [265]. Genetic variants that affect miRNA processing are also thought to contribute to the development of T2DM. For example, rs72631823, located in the terminal loop of pre-miR-34a, has been shown to affect the expression of the mature form of miR-34a [266]. miR-34a is involved in maintaining the functioning of pancreatic β-cells, and disruption of its activity leads to the development of endothelial dysfunction, oxidative stress, and inflammation and predict new-onset diabetes in coronavirus disease 2019 (COVID-19) patients [267,268]. Transfection of plasmid constructs containing the pre-miR-34a sequence with the G or A-rs72631823 allele into INS-1 (rat insulinoma) and MIN6 (mouse insulinoma) cell lines showed increased expression of miR-34a with the A allele in all cell lines compared to the G allele. This is due to the fact that the terminal loop of pre-miR-34a containing the A allele of rs72631823 is in a more relaxed form than the G allele, which leads to more efficient processing of pre-miR-34a by Drosha and Dicer ribonucleases [266].
In a study by Y. Yang et al., it was shown that the mutant allele rs895819-C, located in the terminal loop of pre-miR-27a, leads to a decrease in the expression of both pre-miR-27a itself and its mature form in neural progenitor cells (NPCs) compared to the wild-type allele rs895819-T [269]. It is known that the C allele is associated with a reduced risk of developing T2DM [270] and coronary heart disease [271], which is one of the complications of diabetes. Given the involvement of miR-27a in the development of obesity, insulin resistance and inflammation [272,273,274], it can be assumed that the protective role of rs895819-C in the pathogenesis of T2DM is carried out by suppressing the activity of miR-27a. It was also found that carriers of the G allele and the GG genotype of the rs3746444 polymorphism, located in the stem loop miR-499 precursor, the mature form of which is involved in the PI3K/AKT signaling pathway and glycogen synthesis, have an increased risk of developing T2DM both without and with polyneuropathy [275].
In the Han Chinese population, a 3′ UTR variant in the TBX5 gene, increased the risk for congenital heart disease such as atrial and ventricular septal defects by nearly two-fold [276]. The mutant allele has increased binding affinity to miR-9 and miR-30a which reduced TBX5 expression [276]. Circulating levels of miR-146a were significantly elevated in the cohort of 30 healthy controls compared with 30 T2DM patients [277]. Rs2910164-C allele was associated with the reduced expression levels of the miR-146a but not its mRNAs targets and cytokine levels in diabetic patients [277].
Novel genetic predictors of T2DM development have been increasingly discovered both by high-throughput and targeted studies. Yang et al conducted a case-control study to evaluate the associations of TGFBRAP1 (TGF-β1 receptor-associated protein 1) and TGFBR2 (TGF-β1 receptor 2) SNPs with T2DM risk [278]. Aimed to study the genetic effects of SNPs on diabetes-related miRNA expression they screened T2DM-related miRNA in the patient blood plasma by microarray and 15 differential expressed miRNAs were further validated in 75 T2DM and 75 normal glucose tolerance (NGT). Their findings suggest that genetic polymorphisms of TGFBRAP1, especially rs224179, may contribute to the genetic susceptibility of T2DM by mediating diabetes-related miR-30b-5p and miR-93-5p expression [278].
7.2. T2DM-Associated Variants in lncRNAs
Data on T2DM-associated variants in lncRNAs are still limited. Based on a number of GWAS studies in various ethnic groups, a link between rs10811661 and the risk of developing T2DM was established [279] and confirmed [280] (Table 2). rs10811661 is located 13 kb downstream of the lncRNA ANRIL transcription start site in a locus that includes the MTAP, CDKN2A, CDKN2B and ANRIL genes [281]. ANRIL is known to be a negative transcriptional regulator of CDKN2A and CDKN2B, which encode inhibitors of cyclin-dependent kinases p14/p16 and p15, respectively, which are responsible for pancreatic β-cell proliferation and regeneration [282]. Mechanistically ANRIL suppresses the transcriptional activity of these genes by recruiting the polycomb repressor complexes PRC2 and PRC1, which add the repressing histone modifications H3K27me3 and H2AK119ub [283]. Considering the availability of information on the association of rs10811661 with impaired insulin secretion and changes in ANRIL expression [281], it can be assumed that this polymorphism plays an important role in the pathogenesis of T2DM. An association with T2DM has also been shown for rs10757278, located in the ANRIL intron [284]. In this case-control study conducted on volunteers from China, individuals with the GG genotype were found to have a higher risk of developing T2DM. In addition, individuals with T2DM who had the GG genotype had lower HbA1C and fasting glucose levels, as well as serum ANRIL expression levels [284].
A study by Gonzalez-Moro et al showed that rs2076380, located in a DNA region corresponding to lncRNA TGM2, is involved in T2DM pathogenesis through allele-specific downregulation of TGM2 gene expression in human beta cells, which impairs glucose-stimulated insulin release. The TGM2 risk allele A, associated with T2DM, has been shown to disrupt the secondary structure of this lncRNA, affecting its stability and, consequently, TGM2 expression in pancreatic beta cells [285]. G/G genotype of lncRNA MEG3 rs7158663 single nucleotide polymorphism is associated with protective effect against late-stage diabetic retinopathy in T2DM Patients [286]. An association has also been shown between rs12427129 and rs1899663 in lncRNA HOTAIR and the development of diabetic retinopathy [287], the rs2151280-G allele in CDKN2B-AS1 and an increased risk of proliferative diabetic retinopathy, decreased glomerular filtration rate and high-density lipoprotein levels [288], rs55829688 in lncRNA GAS5 and diabetic kidney disease [289], rs2891168 in lncRNA CDKN2B-AS1 and cardiovascular complications in diabetes [290], rs1333049 in CDKN2B-AS1 and carotid atherosclerosis [291] and HbA1c levels [292].
Pharmacotherapy in T2D is routinely selected based on the patient comorbidities, medication side effects, cost, and availability, without consideration of the underlying pathophysiology driving disease phenotype [293]. Pharmacogenetics is a promising precision medicine approach to tailor treatment choices by a patient’s underlying genetic background [294]. Using genetics to guide medication selection might decrease the risk for comorbidities, or to suggest lower medication doses, reducing side effects [295]. Specific loci have been linked to the response to metformin [296,297], sulfonylureas [298], and GLP-1 receptor agonists [299,300,301].
Examples of SNPs in ncRNAs associated with the response to drug therapy for different diseases have been published [302,303,304]. However, the data about SNPs in ncRNAs association with the response to antidiabetic drugs are very limited. The rs1333049-C allele, located in the CDKN2B-AS1 intron, was found to be associated with poor response to metformin (absolute HbA1c level changed by less than 0.5% from baseline after a three-month course of treatment) in Filipinos with T2DM [305]. Chen Z.R. et al, showed that the G allele of rs60432575 located in the MIR4532 gene promotes efficient binding of hsa-miR-4532 to the 3'-untranslated region of KCNJ11 mRNA, decreasing the expression of KCNJ11 and the Kir6.2 protein encoded by KCNJ11 in HEK293 cells, as well as insulin secretion after stimulation of MIN6 cells with three sulfonylurea drugs (gliclazide, glibenclamide and glimepiride) [306]. In contrast, disruption of hsa-miR-4532 binding to KCNJ11 by the A allele did not affect the expression of KCNJ11, Kir6.2 and insulin secretion [306]. Thus, further study of the role of rs60432575 in the response to sulfonylurea drugs is of great interest. According to our knowledge, there are no publications on the studies of SNPs in ncRNAs associated with the response to SGLTis.
7.3. Future Perspectives in T2DM Pharmacogenetics
As far as the genetic background of T2DM and its complications has been increasingly supported, it is reasonable to expect the genetic component involved in the response to SGLT2 inhibitors. Identification of the susceptible loci associated with T2DM complications risk such as renal and cardiac comorbidities may support early identification of T2DM patients with an increased risk, while identification of SGLT2 inhibitors targets may support targeted therapeutic interventions through the analysis of a panel of genetic markers [307]. Li et al. used canonical correlation analysis based on multivariate regression analysis on the summary statistics data from two large independent meta-analyses of GWAS [308]. This led to the identification of 4624 SNPs and 1745 genes associated with chronic kidney disease (CKD) and heart failure (HF) [308]. Validation of these genes against the transcriptome-wide gene expression data for CKD and HF was followed by gene enrichment and KEGG pathway analyses to explore their potential functional significance. This study identified 169 putative pleiotropic genes significantly associated with at least one of the two conditions. Among these genes, 21 were predicted as potential therapeutic targets of SGLT2 inhibitors in both CKD and HF [308]. It is of interest to investigate whether these genes function in the circulating monocytes or in the resident macrophages.
Transcriptome sequencing (RNA-seq) and the search for allele-asymmetric events (ASE) is currently an accepted approach to identifying SNPs with regulatory function [309,310]. Along with other approaches based on the principle of searching for allele-asymmetric events, this method allows to evaluate the functionality of the examined SNP. Moreover, it allows to reduce the number of the individuals included into the study up to a single individual since allele-asymmetric events are studied for each individual on an identical genetic background and under identical external and internal environmental conditions [309,311]. This fact makes ASE-based approach rather attractive compared with GWAS methodology, which is based on the genetic analysis of hundreds and thousands of patients and controls, in most cases of pharmaceutical studies being unavailable. Therefore, RNA-seq-based ASE search seems very promising for identifying regulatory SNPs associated with different responses to therapeutic agents. For example, a large-scale search of allele-asymmetric events for 50 different substances (hormones, nutrients, several drugs, and environmental pollutants) in transcriptome data was conducted using primary cultures of 5 cell types (LCLs, PBMCs, HUVECs, SMCs, and melanocytes), each represented by samples from three individuals [312]. The authors carried out analysis of the RNA-seq data which allowed to identify 253 SNPs, the misbalance in the representation of alleles of which in the transcriptomes either newly appeared or significantly increased in response to treatment with a particular drug. Using the same approach, 561 ASEs were identified that responded to the treatment of CD4+ lymphocytes (from 24 genotyped individuals) with immobilized antibodies to CD3/CD28 [313]. We believe that the application of this approach can significantly extend the possibilities of searching for potentially regulatory SNPs in ncRNAs and their mRNA targets, including those involved in the mechanisms of response to antidiabetic drugs. Thus, we made bioinformatic analysis of the data from Moyerbrailean et al., which demonstrated that 58 of the 253 SNPs found by the authors represent variations in the microRNA binding sites in the 3'UTR of mRNAs, which, according to calculations, significantly affect the efficiency of their binding. In particular, several discovered SNPs were potentially involved in the individual response to dexamethasone, vitamin D and selenium – containing pharmaceuticals [304].
8. Conclusion
The search for differentially expressed non-coding RNAs, as well as genetic variants affecting their expression during the development of T2DM and the associated cardiac / renal complications, seems to be of current interest. Investigations are necessary to develop the most thorough view of the regulatory relationships of ncRNAs, mRNAs including external and internal stimuli affecting them, which normally ensure the working state of catabolic processes needed for a healthy energy supply. The studies under the review have been aimed to clarify the molecular mechanisms involving ncRNAs that lead to the development of T2DM and associated complications. The expression of ncRNAs is tissue-specific, in this regard, various pathways with their engagement have been identified for the heart / kidney specific cell types. Since chronic inflammation is one of the important processes leading to pathological changes in T2DM, ncRNAs expression changes during the transformation of circulating monocytes and resident macrophages in the heart and kidneys attract significant attention.
Approaches to the detection and functional annotation of T2DM pathogenesis-associated ncRNAs are rapidly developing from targeted study of individual molecules to large-scale detection in whole-transcriptome data with subsequent bioinformatic analysis and construction of competing RNA networks. Data obtained on differentially expressed ncRNAs coming from clinical studies are juxtaposed with the results of animal models and cell culture experiments and further verified on independent patient cohorts to evaluate the clinical significance. The results obtained are extremely valuable, as they can be applied in clinical practice in the near future. Based on differentially expressed ncRNAs in the blood plasma or urine, low-invasive biomarkers for early diagnosis and prognosis of T2DM and related cardiac / renal complications can be developed. In addition, ncRNAs being involved into the coordination of practically all intracellular events are the creative candidates for the novel treatment and prevention strategies for T2DM. The development of ncRNA-based therapy includes two main directions, one of which will use ncRNAs as therapeutic tools to affect specific genes, and the other considers ncRNAs as targets for therapeutic intervention [26,38,137]. The development of ncRNA-based therapeutic approaches for T2DM is emerging, and drug prototypes have not yet reached clinical trials, although laboratory research indicate they are promising [314]. There are drugs in clinical trials for metabolic disease [27]. MiR-34 and miR-16 mimics (MRX34 and MesomiR-1, respectively), have entered clinical trials to assess their potential in cancer therapy, notably these miRs have been shown to be involved in the diabetic cardiomyopathy pathogenesis [60]. Matboli et al used CRISPR/CAS9 to knock out lncRNA-RP11-773H22.4 in peripheral blood mononuclear cells (PBMCs) of T2DM patients, and the expression of insulin resistance-related genes was significantly altered [315].
A topical issue is to clarify the mechanisms of therapeutic action of modern antidiabetic drugs associated with altered expression profile of ncRNA in heart, kidney and immune cells. Particular attention is drawn to SGLT2 inhibitors aimed initially at reducing blood glucose levels by inhibiting the SGLT2 protein glucose-transporting activity, which is involved in the glucose reabsorbtion in the kidney. Understanding the SGLT2-i treatment mechanisms, which along with normalizing glucose levels lead to a decrease in chronic inflammation and a reduction in the severity of T2DM complications, is necessary for the development of ncRNAs-based therapeutic tools, which can help to significantly improve the treatment specificity and reduce unwanted side effects. Identification of genetic variants in ncRNAs and their targets that are associated with T2DM and its complications development, as well as with therapeutic effect of SGLT2i, are needed to expand our understanding of the treatment molecular mechanisms and to make a new contribution to the development of accurate predictive algorithms for personalized therapy of T2DM patients.
Author Contributions
Conceptualization, E.R. and J.K.; writing—original draft preparation, E.R., I.D., and J.K.; writing—review and editing J.K. and T.M.; visualization, I.D. and E.S.; supervision, J.K.; project administration T.M.; funding acquisition, T.M. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the Russian Science Foundation (grant no. 23-15-00113).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
No.
Conflicts of Interest
The authors declare no conflicts of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Yamada, T.; Wakabayashi, M.; Bhalla, A.; Chopra, N.; Miyashita, H.; Mikami, T.; Ueyama, H.; Fujisaki, T.; Saigusa, Y.; Yamaji, T.; et al. Cardiovascular and renal outcomes with SGLT-2 inhibitors versus GLP-1 receptor agonists in patients with type 2 diabetes mellitus and chronic kidney disease: a systematic review and network meta-analysis. Cardiovasc. Diabetol. 2021, 20, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.C.; Andreozzi, F.; Sesti, G. Pharmacogenetics of type 2 diabetes mellitus, the route toward tailored medicine. Diabetes/Metabolism Res. Rev. 2018, 35, e3109. [Google Scholar] [CrossRef] [PubMed]
- Zelniker, T.A.; Braunwald, E. Mechanisms of Cardiorenal Effects of Sodium-Glucose Cotransporter 2 Inhibitors. JACC 2020, 75, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Wang, Z.; Okoth, K.; Toulis, K.A.; Denniston, A.K.; Singh, B.M.; Crowe, F.L.; Sainsbury, C.; Wang, J.; Nirantharakumar, K. Associations of antidiabetic drugs with diabetic retinopathy in people with type 2 diabetes: an umbrella review and meta-analysis. Front. Endocrinol. 2024, 14, 1303238. [Google Scholar] [CrossRef]
- Jarosz-Popek, J.; Eyileten, C.; Gager, G.M.; Nowak, A.; Szwed, P.; Wicik, Z.; Palatini, J.; von Lewinski, D.; Sourij, H.; Siller-Matula, J.M.; et al. The interaction between non-coding RNAs and SGLT2: A review. Int. J. Cardiol. 2023, 398, 131419. [Google Scholar] [CrossRef]
- Rolski, F.; Mączewski, M. Cardiac Fibrosis: Mechanistic Discoveries Linked to SGLT2 Inhibitors. Pharmaceuticals 2025, 18, 313. [Google Scholar] [CrossRef]
- O’hara, D.V.; Lam, C.S.P.; McMurray, J.J.V.; Yi, T.W.; Hocking, S.; Dawson, J.; Raichand, S.; Januszewski, A.S.; Jardine, M.J. Applications of SGLT2 inhibitors beyond glycaemic control. Nat. Rev. Nephrol. 2024, 20, 513–529. [Google Scholar] [CrossRef]
- Heerspink, H.J.L.; Jongs, N.; Chertow, G.M.; Langkilde, A.M.; McMurray, J.J.V.; Correa-Rotter, R.; Rossing, P.; Sjöström, C.D.; Stefansson, B.V.; Toto, R.D.; et al. Effect of dapagliflozin on the rate of decline in kidney function in patients with chronic kidney disease with and without type 2 diabetes: a prespecified analysis from the DAPA-CKD trial. Lancet Diabetes Endocrinol. 2021, 9, 743–754. [Google Scholar] [CrossRef]
- Anker, S.D.; Butler, J.; Filippatos, G.; Ferreira, J.P.; Bocchi, E.; Böhm, M.; Brunner–La Rocca, H.-P.; Choi, D.-J.; Chopra, V.; Chuquiure-Valenzuela, E.; et al. Empagliflozin in Heart Failure with a Preserved Ejection Fraction. N. Engl. J. Med. 2021, 385, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Huang, X.; Chen, J.; Lin, W.; Tian, J. Roles of non-coding RNA in diabetic cardiomyopathy. Cardiovasc. Diabetol. 2024, 23, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- O'BRien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef]
- Kundu, I.; Varshney, S.; Karnati, S.; Naidu, S. The multifaceted roles of circular RNAs in cancer hallmarks: From mechanisms to clinical implications. Mol. Ther. - Nucleic Acids 2024, 35, 102286. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Khorkova, O.; Hsiao, J.; Wahlestedt, C. Basic biology and therapeutic implications of lncRNA. Adv. Drug Deliv. Rev. 2015, 87, 15–24. [Google Scholar] [CrossRef]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging role of non-coding RNA in health and disease. Metab. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef]
- Beňačka, R.; Szabóová, D.; Guľašová, Z.; Hertelyová, Z.; Radoňak, J. Non-Coding RNAs in Human Cancer and Other Diseases: Overview of the Diagnostic Potential. Int. J. Mol. Sci. 2023, 24, 16213. [Google Scholar] [CrossRef]
- Dehghanian, F.; Azhir, Z.; Khalilian, S.; Grüning, B. Non-coding RNAs underlying the pathophysiological links between type 2 diabetes and pancreatic cancer: A systematic review. J. Diabetes Investig. 2021, 13, 405–428. [Google Scholar] [CrossRef]
- Yang, F.; Chen, Y.; Xue, Z.; Lv, Y.; Shen, L.; Li, K.; Zheng, P.; Pan, P.; Feng, T.; Jin, L.; et al. High-Throughput Sequencing and Exploration of the lncRNA-circRNA-miRNA-mRNA Network in Type 2 Diabetes Mellitus. BioMed Res. Int. 2020, 2020, 8162524. [Google Scholar] [CrossRef] [PubMed]
- Kudriashov, V.; Sufianov, A.; Mashkin, A.; Beilerli, A.; Ilyasova, T.; Liang, Y.; Lyulin, S.; Beylerli, O. The role of long non-coding RNAs in carbohydrate and fat metabolism in the liver. Non-coding RNA Res. 2023, 8, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Kutikhin, A.G.; Sinitsky, M.Y.; Yuzhalin, A.E.; Velikanova, E.A. Whole-Transcriptome Sequencing: A Powerful Tool for Vascular Tissue Engineering and Endothelial Mechanobiology. BioTech 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Nie, L.; Wang, M.; Liao, W.; Huan, C.; Jia, Z.; Wei, D.; Liu, P.; Fan, K.; Mao, Z.; et al. Differential Expression of lncRNA-miRNA-mRNA and Their Related Functional Networks in New-Onset Type 2 Diabetes Mellitus among Chinese Rural Adults. Genes 2022, 13, 2073. [Google Scholar] [CrossRef]
- Grillone, K.; Caridà, G.; Luciano, F.; Cordua, A.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. A systematic review of non-coding RNA therapeutics in early clinical trials: a new perspective against cancer. J. Transl. Med. 2024, 22, 1–13. [Google Scholar] [CrossRef]
- Li, S.; Xiong, F.; Zhang, S.; Liu, J.; Gao, G.; Xie, J.; Wang, Y. Oligonucleotide therapies for nonalcoholic steatohepatitis. Mol. Ther. - Nucleic Acids 2024, 35, 102184. [Google Scholar] [CrossRef]
- Jing, F.; Shi, Y.; Jiang, D.; Li, X.; Sun, J.; Zhang, X.; Guo, Q. Deciphering the role of non-coding RNAs involved in sorafenib resistance. Heliyon 2024, 10, e29374. [Google Scholar] [CrossRef]
- Spinetti, G.; Mutoli, M.; Greco, S.; Riccio, F.; Ben-Aicha, S.; Kenneweg, F.; Jusic, A.; de Gonzalo-Calvo, D.; Nossent, A.Y.; Novella, S.; et al. Cardiovascular complications of diabetes: role of non-coding RNAs in the crosstalk between immune and cardiovascular systems. Cardiovasc. Diabetol. 2023, 22, 1–28. [Google Scholar] [CrossRef]
- Dieter, C.; Lemos, N.E.; Corrêa, N.R.d.F.; Assmann, T.S.; Crispim, D. The Impact of lncRNAs in Diabetes Mellitus: A Systematic Review and In Silico Analyses. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef]
- Jakubik, D.; Fitas, A.; Eyileten, C.; Jarosz-Popek, J.; Nowak, A.; Czajka, P.; Wicik, Z.; Sourij, H.; Siller-Matula, J.M.; De Rosa, S.; et al. MicroRNAs and long non-coding RNAs in the pathophysiological processes of diabetic cardiomyopathy: Emerging biomarkers and potential therapeutics. Cardiovasc. Diabetol. 2021, 20, 55. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, D.; Jiang, Z.; Gao, Y.; Sun, L.; Li, R.; Chen, M.; Lin, C.; Liu, D. Non-coding RNAs in diabetes mellitus and diabetic cardiovascular disease. Front. Endocrinol. 2022, 13, 961802. [Google Scholar] [CrossRef] [PubMed]
- Taheri, M.; Eghtedarian, R.; Ghafouri-Fard, S.; Omrani, M.D. Non-coding RNAs and type 2 diabetes mellitus. Arch. Physiol. Biochem. 2020, 129, 526–535. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, H. Emerging Roles of ncRNAs in Type 2 Diabetes Mellitus: From Mechanisms to Drug Discovery. Biomolecules 2024, 14, 1364. [Google Scholar] [CrossRef]
- Ma, J.; Gao, R.; Xie, Q.; Pan, X.; Tong, N. Whole transcriptome sequencing analyses of islets reveal ncRNA regulatory networks underlying impaired insulin secretion and increased β-cell mass in high fat diet-induced diabetes mellitus. PLOS ONE 2024, 19, e0300965. [Google Scholar] [CrossRef]
- Kura, B.; Kindernay, L.; Singla, D.; Dulova, U.; Bartekova, M. Mechanistic insight into the role of cardiac-enriched microRNAs in diabetic heart injury. Am. J. Physiol. Circ. Physiol. 2025, 328, H865–H884. [Google Scholar] [CrossRef]
- Mo, F.; Lv, B.; Zhao, D.; Xi, Z.; Qian, Y.; Ge, D.; Yang, N.; Zhang, D.; Jiang, G.; Gao, S. Small RNA Sequencing Analysis of STZ-Injured Pancreas Reveals Novel MicroRNA and Transfer RNA-Derived RNA with Biomarker Potential for Diabetes Mellitus. Int. J. Mol. Sci. 2023, 24, 10323. [Google Scholar] [CrossRef]
- Macvanin, M.T.; Gluvic, Z.; Bajic, V.; Isenovic, E.R. Novel insights regarding the role of noncoding RNAs in diabetes. World J. Diabetes 2023, 14, 958–976. [Google Scholar] [CrossRef]
- Szydełko, J.; Czop, M.; Petniak, A.; Lenart-Lipińska, M.; Kocki, J.; Zapolski, T.; Matyjaszek-Matuszek, B. Identification of plasma miR-4505, miR-4743-5p and miR-4750-3p as novel diagnostic biomarkers for coronary artery disease in patients with type 2 diabetes mellitus: a case-control study. Cardiovasc. Diabetol. 2024, 23, 1–23. [Google Scholar] [CrossRef]
- Yin, R.; Zhang, Y.; Fang, X.; Zhang, Y.; Miao, R.; Yao, Y.; Guan, H.; Tian, J. Discovering diabetes complications-related microRNAs: meta-analyses and pathway modeling approach. BMC Med Genom. 2025, 18, 1–15. [Google Scholar] [CrossRef]
- Ahmed, S.; Adnan, H.; Khawaja, M.A.; Butler, A.E. Novel Micro-Ribonucleic Acid Biomarkers for Early Detection of Type 2 Diabetes Mellitus and Associated Complications—A Literature Review. Int. J. Mol. Sci. 2025, 26, 753. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Wang, S.-S.; Han, Z.; Han, F.; Chang, Y.-P.; Yang, Y.; Xue, M.; Sun, B.; Chen, L.-M. Triptolide Restores Autophagy to Alleviate Diabetic Renal Fibrosis through the miR-141-3p/PTEN/Akt/mTOR Pathway. Mol. Ther. - Nucleic Acids 2017, 9, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Li, X.; Zhan, X.; Sun, L.; Gao, J.; Cao, Y.; Qiu, H. Construction of competitive endogenous RNA network reveals regulatory role of long non-coding RNAs in type 2 diabetes mellitus. J. Cell. Mol. Med. 2017, 21, 3204–3213. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Ma, Y.; Wu, S.; Zhuang, Y.; Liu, X.; Lv, L.; Zhang, G. Construction and analysis of a lncRNA-miRNA-mRNA network based on competitive endogenous RNA reveals functional lncRNAs in diabetic cardiomyopathy. Mol. Med. Rep. 2019, 20, 1393–1403. [Google Scholar] [CrossRef]
- Ma, Q.; Wang, L.; Wang, Z.; Su, Y.; Hou, Q.; Xu, Q.; Cai, R.; Wang, T.; Gong, X.; Yi, Q. Long non-coding RNA screening and identification of potential biomarkers for type 2 diabetes. J. Clin. Lab. Anal. 2022, 36, e24280. [Google Scholar] [CrossRef]
- Cao, H.; Rao, X.; Jia, J.; Yan, T.; Li, D. Identification of tubulointerstitial genes and ceRNA networks involved in diabetic nephropathy via integrated bioinformatics approaches. Hereditas 2022, 159, 1–12. [Google Scholar] [CrossRef]
- Bielska, A.; Niemira, M.; Bauer, W.; Sidorkiewicz, I.; Szałkowska, A.; Skwarska, A.; Raczkowska, J.; Ostrowski, D.; Gugała, K.; Dobrzycki, S.; et al. Serum miRNA Profile in Diabetic Patients With Ischemic Heart Disease as a Promising Non-Invasive Biomarker. Front. Endocrinol. 2022, 13, 888948. [Google Scholar] [CrossRef]
- Fang, Y.; Wang, X.; Li, W.; Han, J.; Jin, J.; Su, F.; Zhang, J.; Huang, W.; Xiao, F.; Pan, Q.; et al. Screening of circular RNAs and validation of circANKRD36 associated with inflammation in patients with type 2 diabetes mellitus. Int. J. Mol. Med. 2018, 42, 1865–1874. [Google Scholar] [CrossRef]
- Mou, X.; Chenv, J.W.; Zhou, D.Y.; Liu, K.; Chen, L.J.; Zhou, D.; Hu, Y. A novel identified circular RNA, circ_0000491, aggravates the extracellular matrix of diabetic nephropathy glomerular mesangial cells through suppressing miR-101b by targeting TGFβRI. Mol. Med. Rep. 2020, 22, 3785–3794. [Google Scholar] [CrossRef]
- Zhou, R.-M.; Shi, Z.-H.; Shan, K.; Zhang, S.-J.; Zhang, Y.-H.; Liang, Y.; Yan, B.; Zhao, C. Comparative Analysis of Differentially Expressed Circular RNAs in Polarized Macrophages. Front. Genet. 2022, 13, 823517. [Google Scholar] [CrossRef]
- Halabi, N.; Thomas, B.; Chidiac, O.; Robay, A.; AbiNahed, J.; Jayyousi, A.; Al Suwaidi, J.; Bradic, M.; Khalil, C.A. Dysregulation of long non-coding RNA gene expression pathways in monocytes of type 2 diabetes patients with cardiovascular disease. Cardiovasc. Diabetol. 2024, 23, 1–23. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, S.; Xia, Y.; Li, H.; Liu, D.; Du, Y. Identification of novel target genes in exaggerated cardiac remodeling following myocardial infarction in diabetes. Front. Endocrinol. 2025, 16, 1536639. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.S.; Kamel, M.M.; Agwa, S.H.A.; Hakeem, M.S.A.; El Meteini, M.S.; Matboli, M. Analysis of mRNA-miRNA-lncRNA differential expression in prediabetes/type 2 diabetes mellitus patients as potential players in insulin resistance. Front. Endocrinol. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Huang, H.; Lai, J.; Lin, S.; Huang, Y. Long noncoding RNAs as potential diagnostic biomarkers for diabetes mellitus and complications: A systematic review and meta-analysis. J. Diabetes 2023, 16. [Google Scholar] [CrossRef] [PubMed]
- Grieco, G.E.; Besharat, Z.M.; Licata, G.; Fignani, D.; Brusco, N.; Nigi, L.; Formichi, C.; Po, A.; Sabato, C.; Dardano, A.; et al. Circulating microRNAs as clinically useful biomarkers for Type 2 Diabetes Mellitus: miRNomics from bench to bedside. Transl. Res. 2022, 247, 137–157. [Google Scholar] [CrossRef]
- Mahjoob, G.; Ahmadi, Y.; Rajani, H.F.; Khanbabaei, N.; Abolhasani, S. Circulating microRNAs as predictive biomarkers of coronary artery diseases in type 2 diabetes patients. J. Clin. Lab. Anal. 2022, 36, e24380. [Google Scholar] [CrossRef]
- Sulaiman, F.; Khyriem, C.; Dsouza, S.; Abdul, F.; Alkhnbashi, O.; Faraji, H.; Farooqi, M.; Al Awadi, F.; Hassanein, M.; Ahmed, F.; et al. Characterizing Circulating microRNA Signatures of Type 2 Diabetes Subtypes. Int. J. Mol. Sci. 2025, 26, 637. [Google Scholar] [CrossRef]
- Xu, H.; Guo, S.; Li, W.; Yu, P. The circular RNA Cdr1as, via miR-7 and its targets, regulates insulin transcription and secretion in islet cells. Sci. Rep. 2015, 5, srep12453. [Google Scholar] [CrossRef]
- Macvanin, M.T.; Gluvic, Z.; Radovanovic, J.; Essack, M.; Gao, X.; Isenovic, E.R. Diabetic cardiomyopathy: The role of microRNAs and long non-coding RNAs. Front. Endocrinol. 2023, 14, 1124613. [Google Scholar] [CrossRef]
- Dillmann, W.H. Diabetic Cardiomyopathy. Circ. Res. 2019, 124, 1160–1162. [Google Scholar] [CrossRef]
- Ma, X.; Mei, S.; Wuyun, Q.; Zhou, L.; Sun, D.; Yan, J. Epigenetics in diabetic cardiomyopathy. Clin. Epigenetics 2024, 16, 1–17. [Google Scholar] [CrossRef]
- Edgar, L.; Akbar, N.; Braithwaite, A.T.; Krausgruber, T.; Gallart-Ayala, H.; Bailey, J.; Corbin, A.L.; Khoyratty, T.E.; Chai, J.T.; Alkhalil, M.; et al. Hyperglycemia Induces Trained Immunity in Macrophages and Their Precursors and Promotes Atherosclerosis. Circulation 2021, 144, 961–982. [Google Scholar] [CrossRef]
- Riksen, N.P.; Bekkering, S.; Mulder, W.J.M.; Netea, M.G. Trained immunity in atherosclerotic cardiovascular disease. Nat. Rev. Cardiol. 2023, 20, 799–811. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Geng, K.; Wang, H.-Y.; Wan, S.-R.; Ma, X.-M.; Long, Y.; Xu, Y.; Jiang, Z.-Z. Hyperglycemia-induced STING signaling activation leads to aortic endothelial injury in diabetes. Cell Commun. Signal. 2023, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Vinci, M.C.; Costantino, S.; Damiano, G.; Rurali, E.; Rinaldi, R.; Vigorelli, V.; Sforza, A.; Carulli, E.; Pirola, S.; Mastroiacovo, G.; et al. Persistent epigenetic signals propel a senescence-associated secretory phenotype and trained innate immunity in CD34+ hematopoietic stem cells from diabetic patients. Cardiovasc. Diabetol. 2024, 23, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive oxygen species (ROS) in macrophage activation and function in diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef]
- Shariatzadeh, M.; Payán-Gómez, C.; Kzhyshkowska, J.; Dik, W.A.; Leenen, P.J.M. Polarized Macrophages Show Diverse Pro-Angiogenic Characteristics Under Normo- and Hyperglycemic Conditions. Int. J. Mol. Sci. 2025, 26, 4846. [Google Scholar] [CrossRef]
- Badillo-Garcia, L.E.; Liu, Q.; Ziebner, K.; Balduff, M.; Sevastyanova, T.; Schmuttermaier, C.; Klüter, H.; Harmsen, M.; Kzhyshkowska, J. Hyperglycemia amplifies TLR-mediated inflammatory response of M(IL4) macrophages to dyslipidemic ligands. J. Leukoc. Biol. 2024, 116, 197–204. [Google Scholar] [CrossRef]
- Mossel, D.M.; Moganti, K.; Riabov, V.; Weiss, C.; Kopf, S.; Cordero, J.; Dobreva, G.; Rots, M.G.; Klüter, H.; Harmsen, M.C.; et al. Epigenetic Regulation of S100A9 and S100A12 Expression in Monocyte-Macrophage System in Hyperglycemic Conditions. Front. Immunol. 2020, 11, 1071. [Google Scholar] [CrossRef]
- Chee, Y.J.; Dalan, R.; Cheung, C. The Interplay Between Immunity, Inflammation and Endothelial Dysfunction. Int. J. Mol. Sci. 2025, 26, 1708. [Google Scholar] [CrossRef]
- Seferović, P.M.; Paulus, W.J. Clinical diabetic cardiomyopathy: a two-faced disease with restrictive and dilated phenotypes. Eur. Hear. J. 2015, 36, 1718–1727. [Google Scholar] [CrossRef]
- Srivastava, S.P.; Goodwin, J.E.; Tripathi, P.; Kanasaki, K.; Koya, D. Interactions among Long Non-Coding RNAs and microRNAs Influence Disease Phenotype in Diabetes and Diabetic Kidney Disease. Int. J. Mol. Sci. 2021, 22, 6027. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, H.; Zhu, X.; Jiang, F.; Yu, Q.; Zhang, J.; Ji, Y. miR-208 inhibits myocardial tissues apoptosis in mice with acute myocardial infarction by targeting inhibition of PDCD4. J. Biochem. Mol. Toxicol. 2022, 36, e23202. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Cardiac-specific miRNA in cardiogenesis, heart function, and cardiac pathology (with focus on myocardial infarction). J. Mol. Cell. Cardiol. 2016, 94, 107–121. [Google Scholar] [CrossRef]
- Li, J.; Ren, Y.; Shi, E.; Tan, Z.; Xiong, J.; Yan, L.; Jiang, X. Inhibition of the Let-7 Family MicroRNAs Induces Cardioprotection Against Ischemia-Reperfusion Injury in Diabetic Rats. Ann. Thorac. Surg. 2016, 102, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Chen, S.; George, B.; Feng, Q.; Chakrabarti, S. miR133a regulates cardiomyocyte hypertrophy in diabetes. Diabetes/Metabolism Res. Rev. 2009, 26, 40–49. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Y.; Liu, C. The interregulatory circuit between non-coding RNA and apoptotic signaling in diabetic cardiomyopathy. Non-coding RNA Res. 2024, 9, 1080–1097. [Google Scholar] [CrossRef]
- Li, J.; Yang, C.; Wang, Y. miR-126 overexpression attenuates oxygen-glucose deprivation/reperfusion injury by inhibiting oxidative stress and inflammatory response via the activation of SIRT1/Nrf2 signaling pathway in human umbilical vein endothelial cells. Mol. Med. Rep. 2020, 23, 1–1. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Gambardella, J.; Sardu, C.; Lombardi, A.; Santulli, G. Functional Role of miR-155 in the Pathogenesis of Diabetes Mellitus and Its Complications. Non-Coding RNA 2021, 7, 39. [Google Scholar] [CrossRef]
- Pofi, R.; Giannetta, E.; Galea, N.; Francone, M.; Campolo, F.; Barbagallo, F.; Gianfrilli, D.; Venneri, M.A.; Filardi, T.; Cristini, C.; et al. Diabetic Cardiomiopathy Progression is Triggered by miR122-5p and Involves Extracellular Matrix: A 5-Year Prospective Study. JACC Cardiovasc. Imaging 2021, 14, 1130–1142. [Google Scholar] [CrossRef]
- Che, H.; Wang, Y.; Li, H.; Li, Y.; Sahil, A.; Lv, J.; Liu, Y.; Yang, Z.; Dong, R.; Xue, H.; et al. Melatonin alleviates cardiac fibrosis via inhibiting lncRNA MALAT1/miR-141-mediated NLRP3 inflammasome and TGF-β1/Smads signaling in diabetic cardiomyopathy. FASEB J. 2020, 34, 5282–5298. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, F.; Mohammadzad, M.S. Bioinformatics analyses of potential microRNAs and their target genes in myocardial infarction patients with diabetes. Diabetes Vasc. Dis. Res. 2025, 22. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Pang, H.; Xie, Z.; Huang, G.; Zhou, Z. Circular RNAs in diabetes mellitus and its complications. Front. Endocrinol. 2022, 13, 885650. [Google Scholar] [CrossRef]
- Galis, P.; Bartosova, L.; Farkasova, V.; Bartekova, M.; Ferenczyova, K.; Rajtik, T. Update on clinical and experimental management of diabetic cardiomyopathy: addressing current and future therapy. Front. Endocrinol. 2024, 15, 1451100. [Google Scholar] [CrossRef]
- Meng, L.; Lin, H.; Huang, X.; Weng, J.; Peng, F.; Wu, S. METTL14 suppresses pyroptosis and diabetic cardiomyopathy by downregulating TINCR lncRNA. Cell Death Dis. 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Yu, M.; Shan, X.; Liu, Y.; Zhu, J.; Cao, Q.; Yang, F.; Liu, Y.; Wang, G.; Zhao, X. RNA-Seq analysis and functional characterization revealed lncRNA NONRATT007560.2 regulated cardiomyocytes oxidative stress and apoptosis induced by high glucose. J. Cell. Biochem. 2019, 120, 18278–18287. [Google Scholar] [CrossRef]
- Gao, L.; Wang, X.; Guo, S.; Xiao, L.; Liang, C.; Wang, Z.; Li, Y.; Liu, Y.; Yao, R.; Liu, Y.; et al. LncRNA HOTAIR functions as a competing endogenous RNA to upregulate SIRT1 by sponging miR-34a in diabetic cardiomyopathy. J. Cell. Physiol. 2018, 234, 4944–4958. [Google Scholar] [CrossRef]
- Dai, W.; Lee, D. Interfering with long chain noncoding RNA ANRIL expression reduces heart failure in rats with diabetes by inhibiting myocardial oxidative stress. J. Cell. Biochem. 2019, 120, 18446–18456. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.-Y.; Li, T.-T.; Wang, J.; Jiang, Y.; Zhao, Y.; Jin, X.-X.; Xue, G.-L.; Yang, Y.; Zhang, X.-F.; et al. Ablation of interleukin-17 alleviated cardiac interstitial fibrosis and improved cardiac function via inhibiting long non-coding RNA-AK081284 in diabetic mice. J. Mol. Cell. Cardiol. 2018, 115, 64–72. [Google Scholar] [CrossRef]
- Zheng, D.; Zhang, Y.; Hu, Y.; Guan, J.; Xu, L.; Xiao, W.; Zhong, Q.; Ren, C.; Lu, J.; Liang, J.; et al. Long noncoding RNA Crnde attenuates cardiac fibrosis via Smad3-Crnde negative feedback in diabetic cardiomyopathy. FEBS J. 2019, 286, 1645–1655. [Google Scholar] [CrossRef]
- Yang, F.; Qin, Y.; Wang, Y.; Li, A.; Lv, J.; Sun, X.; Che, H.; Han, T.; Meng, S.; Bai, Y.; et al. LncRNA KCNQ1OT1 Mediates Pyroptosis in Diabetic Cardiomyopathy. Cell. Physiol. Biochem. 2018, 50, 1230–1244. [Google Scholar] [CrossRef]
- Zhao, S.-F.; Ye, Y.-X.; Xu, J.-D.; He, Y.; Zhang, D.-W.; Xia, Z.-Y.; Wang, S. Long non-coding RNA KCNQ1OT1 increases the expression of PDCD4 by targeting miR-181a-5p, contributing to cardiomyocyte apoptosis in diabetic cardiomyopathy. Acta Diabetol. 2021, 58, 1251–1267. [Google Scholar] [CrossRef] [PubMed]
- Leng, Y.; Wang, M.-Z.; Xie, K.-L.; Cai, Y. Identification of Potentially Functional Circular RNA/Long Noncoding RNA-MicroRNA-mRNA Regulatory Networks Associated with Vascular Injury in Type 2 Diabetes Mellitus by Integrated Microarray Analysis. J. Diabetes Res. 2023, 2023, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Duan, J.; Zhou, H. Perspectives of circular RNAs in diabetic complications from biological markers to potential therapeutic targets (Review). Mol. Med. Rep. 2023, 28, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, Z.; Jian, D.; Li, W.; Tang, H.; Li, M. Hsa-circRNA11783-2 in peripheral blood is correlated with coronary artery disease and type 2 diabetes mellitus. Diabetes Vasc. Dis. Res. 2017, 14, 510–515. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Yang, Y.; Liu, Z.; Sun, S.; Li, R.; Zhu, H.; Li, T.; Zheng, J.; Li, J.; et al. Circular RNAs in human diseases. Medcomm 2024, 5, e699. [Google Scholar] [CrossRef]
- Tang, C.-M.; Zhang, M.; Huang, L.; Hu, Z.-Q.; Zhu, J.-N.; Xiao, Z.; Zhang, Z.; Lin, Q.-X.; Zheng, X.-L.; -Yang, M.; et al. CircRNA_000203 enhances the expression of fibrosis-associated genes by derepressing targets of miR-26b-5p, Col1a2 and CTGF, in cardiac fibroblasts. Sci. Rep. 2017, 7, 40342. [Google Scholar] [CrossRef]
- Zhou, B.; Yu, J.-W. A novel identified circular RNA, circRNA_010567, promotes myocardial fibrosis via suppressing miR-141 by targeting TGF-β1. Biochem. Biophys. Res. Commun. 2017, 487, 769–775. [Google Scholar] [CrossRef]
- Yang, F.; Li, A.; Qin, Y.; Che, H.; Wang, Y.; Lv, J.; Li, Y.; Li, H.; Yue, E.; Ding, X.; et al. A Novel Circular RNA Mediates Pyroptosis of Diabetic Cardiomyopathy by Functioning as a Competing Endogenous RNA. Mol. Ther. - Nucleic Acids 2019, 17, 636–643. [Google Scholar] [CrossRef]
- Yuan, Q.; Sun, Y.; Yang, F.; Yan, D.; Shen, M.; Jin, Z.; Zhan, L.; Liu, G.; Yang, L.; Zhou, Q.; et al. CircRNA DICAR as a novel endogenous regulator for diabetic cardiomyopathy and diabetic pyroptosis of cardiomyocytes. Signal Transduct. Target. Ther. 2023, 8, 1–12. [Google Scholar] [CrossRef]
- Balzer, M.S.; Rohacs, T.; Susztak, K. How Many Cell Types Are in the Kidney and What Do They Do? Annu. Rev. Physiol. 2021, 84, 507–531. [Google Scholar] [CrossRef]
- Wang, B.; Xiong, Y.; Deng, X.; Wang, Y.; Gong, S.; Yang, S.; Yang, B.; Yang, Y.; Leng, Y.; Li, W.; et al. The role of intercellular communication in diabetic nephropathy. Front. Immunol. 2024, 15, 1423784. [Google Scholar] [CrossRef] [PubMed]
- Joumaa, J.P.; Raffoul, A.; Sarkis, C.; Chatrieh, E.; Zaidan, S.; Attieh, P.; Harb, F.; Azar, S.; Ghadieh, H.E. Mechanisms, Biomarkers, and Treatment Approaches for Diabetic Kidney Disease: Current Insights and Future Perspectives. J. Clin. Med. 2025, 14, 727. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, C. Recent Advances in the Management of Diabetic Kidney Disease: Slowing Progression. Int. J. Mol. Sci. 2024, 25, 3086. [Google Scholar] [CrossRef] [PubMed]
- Abdelmaksoud, N.M.; Al-Noshokaty, T.M.; Abdelhamid, R.; Abdellatif, N.; Mansour, A.; Mohamed, R.; Mohamed, A.H.; Khalil, N.A.E.; Abdelhamid, S.S.; Mohsen, A.; et al. Deciphering the role of MicroRNAs in diabetic nephropathy: Regulatory mechanisms and molecular insights. Pathol. - Res. Pr. 2024, 256, 155237. [Google Scholar] [CrossRef]
- Yu, S.; Li, Y.; Lu, X.; Han, Z.; Li, C.; Yuan, X.; Guo, D. The regulatory role of miRNA and lncRNA on autophagy in diabetic nephropathy. Cell. Signal. 2024, 118, 111144. [Google Scholar] [CrossRef]
- Lv, J.; Wu, Y.; Mai, Y.; Bu, S. Noncoding RNAs in Diabetic Nephropathy: Pathogenesis, Biomarkers, and Therapy. J. Diabetes Res. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Wada, J. Noncoding RNAs and diabetic kidney disease. J. Diabetes Investig. 2024, 16, 8–9. [Google Scholar] [CrossRef]
- Ishii, H.; Kaneko, S.; Yanai, K.; Aomatsu, A.; Hirai, K.; Ookawara, S.; Ishibashi, K.; Morishita, Y. MicroRNAs in Podocyte Injury in Diabetic Nephropathy. Front. Genet. 2020, 11, 993. [Google Scholar] [CrossRef]
- Wei, B.; Liu, Y.; Guan, H. MicroRNA-145-5p attenuates high glucose-induced apoptosis by targeting the Notch signaling pathway in podocytes. Exp. Ther. Med. 2020, 19, 1915–1924. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, S.; Jia, Q.; Zhang, A.; Li, Y.; Zhu, Y.; Lv, S.; Zhang, J. The microRNA in ventricular remodeling: the miR-30 family. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Zhang, L.-Y.; Wang, Y.; Yang, Y.-R.; Shao, J.-J.; Liang, B. MiR-135a regulates renal fibrosis in rats with diabetic kidney disease through the Notch pathway. 2020, 24, 1979–1987. [CrossRef]
- Luan, J.; Fu, J.; Wang, D.; Jiao, C.; Cui, X.; Chen, C.; Liu, D.; Zhang, Y.; Wang, Y.; Yuen, P.S.; et al. miR-150-Based RNA Interference Attenuates Tubulointerstitial Fibrosis through the SOCS1/JAK/STAT Pathway In Vivo and In Vitro. Mol. Ther. - Nucleic Acids 2020, 22, 871–884. [Google Scholar] [CrossRef]
- Tsai, Y.-C.; Kuo, P.-L.; Hung, W.-W.; Wu, L.-Y.; Wu, P.-H.; Chang, W.-A.; Kuo, M.-C.; Hsu, Y.-L. Angpt2 Induces Mesangial Cell Apoptosis through the MicroRNA-33-5p-SOCS5 Loop in Diabetic Nephropathy. Mol. Ther. - Nucleic Acids 2018, 13, 543–555. [Google Scholar] [CrossRef]
- Xi, W.; Zhao, X.; Wu, M.; Fu, X.; Jia, W.; Lu, M.; Li, H. MiR-214 promotes renal fibrosis in diabetic nephropathy via targeting SOCS1. Trop. J. Pharm. Res. 2021, 18, 1009–1015. [Google Scholar] [CrossRef]
- Wang, S.-Z.; Zhang, Y.-L.; Shi, H.-B. Potential Repressive Impact of microRNA-20a on Renal Tubular Damage in Diabetic Kidney Disease by Targeting C-X-C Motif Chemokine Ligand 6. Arch. Med Res. 2021, 52, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.-H.; Wu, H.; Wang, X.; Ji, L.-L.; Chen, C. MiR-30c-5p inhibits high glucose-induced EMT and renal fibrogenesis by down-regulation of JAK1 in diabetic nephropathy. 2020, 24, 1338–1349. [CrossRef]
- Wang, H.; Zhang, R.; Wu, X.; Chen, Y.; Ji, W.; Wang, J.; Zhang, Y.; Xia, Y.; Tang, Y.; Yuan, J. The Wnt Signaling Pathway in Diabetic Nephropathy. Front. Cell Dev. Biol. 2022, 9, 701547. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wan, J.; Hou, X.; Geng, J.; Li, X.; Bai, X. MicroRNA-27a promotes podocyte injury via PPARγ-mediated β-catenin activation in diabetic nephropathy. Cell Death Dis. 2017, 8, e2658–e2658. [Google Scholar] [CrossRef]
- Wang, X.; Gao, Y.; Tian, N.; Zou, D.; Shi, Y.; Zhang, N. Astragaloside IV improves renal function and fibrosis via inhibition of miR-21-induced podocyte dedifferentiation and mesangial cell activation in diabetic mice. Drug Des. Dev. Ther. 2018, ume 12, 2431–2442. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, S.; Sun, X.; Lou, Y.; Bao, J.; Yu, J. Hyperoside ameliorates diabetic nephropathy induced by STZ via targeting the miR-499–5p/APC axis. J. Pharmacol. Sci. 2021, 146, 10–20. [Google Scholar] [CrossRef]
- Bera, A.; Das, F.; Ghosh-Choudhury, N.; Mariappan, M.M.; Kasinath, B.S.; Choudhury, G.G. Reciprocal regulation of miR-214 and PTEN by high glucose regulates renal glomerular mesangial and proximal tubular epithelial cell hypertrophy and matrix expansion. Am. J. Physiol. Physiol. 2017, 313, C430–C447. [Google Scholar] [CrossRef]
- El-Said, Y.A.M.; Sallam, N.A.A.; Ain-Shoka, A.A.-M.; Abdel-Latif, H.A.-T. Geraniol ameliorates diabetic nephropathy via interference with miRNA-21/PTEN/Akt/mTORC1 pathway in rats. Naunyn-Schmiedeberg's Arch. Pharmacol. 2020, 393, 2325–2337. [Google Scholar] [CrossRef]
- Khokhar, M.; Purohit, P.; Tomo, S.; Agarwal, R.G.; Gadwal, A.; Bajpai, N.K.; Bohra, G.K.; Shukla, R.K. PTEN, MMP2, and NF-κB and Regulating MicroRNA-181 Aggravate Insulin Resistance and Progression of Diabetic Nephropathy: A Case-Control Study. Kidney Dial. 2023, 3, 121–138. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Yan, R.; Liang, L.; Zhou, X.; Liu, H.; Zhang, X.; Mao, Y.; Peng, W.; Xiao, Y.; et al. BMP-7 inhibits renal fibrosis in diabetic nephropathy via miR-21 downregulation. Life Sci. 2019, 238, 116957. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Jung, E.-H.; Kim, G.-Y.; Kim, B.-C.; Lim, J.H.; Woo, C.-H. Itch E3 Ubiquitin Ligase Positively Regulates TGF-β Signaling to EMT via Smad7 Ubiquitination. Mol. Cells 2015, 38, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Jiang, M. MicroRNA-27a Adjusts Diabetic Nephropathy Patients and Inhibits TGF-β/Smad Signaling Pathway. Int. J. Clin. Exp 2017, 10, 14266–14274. [Google Scholar]
- Wang, L.; Wang, H.-L.; Liu, T.-T.; Lan, H.-Y. TGF-Beta as a Master Regulator of Diabetic Nephropathy. Int. J. Mol. Sci. 2021, 22, 7881. [Google Scholar] [CrossRef]
- Du, Y.; Yang, Y.; Tang, G.; Jia, J.; Zhu, N.; Yuan, W. Butyrate alleviates diabetic kidney disease by mediating the miR-7a-5p/P311/TGF-β1 pathway. FASEB J. 2020, 34, 10462–10475. [Google Scholar] [CrossRef]
- Fluitt, M.B.; Shivapurkar, N.; Kumari, M.; Singh, S.; Li, L.; Tiwari, S.; Ecelbarger, C.M. Systemic inhibition of miR-451 increases fibrotic signaling and diminishes autophagic response to exacerbate renal damage in Tallyho/Jng mice. Am. J. Physiol. Physiol. 2020, 319, F476–F486. [Google Scholar] [CrossRef]
- Bijkerk, R.; Duijs, J.M.G.J.; Khairoun, M.; ter Horst, C.J.H.; van der Pol, P.; Mallat, M.J.; Rotmans, J.I.; de Vries, A.P.J.; de Koning, E.J.; de Fijter, J.W.; et al. Circulating MicroRNAs Associate With Diabetic Nephropathy and Systemic Microvascular Damage and Normalize After Simultaneous Pancreas–Kidney Transplantation. Am. J. Transplant. 2015, 15, 1081–1090. [Google Scholar] [CrossRef]
- Qin, W.; Chung, A.C.; Huang, X.R.; Meng, X.-M.; Hui, D.S.; Yu, C.-M.; Sung, J.J.; Lan, H.Y. TGF-β/Smad3 Signaling Promotes Renal Fibrosis by Inhibiting miR-29. J. Am. Soc. Nephrol. 2011, 22, 1462–1474. [Google Scholar] [CrossRef]
- Tang, S.-T.; Wang, F.; Shao, M.; Wang, Y.; Zhu, H.-Q. MicroRNA-126 suppresses inflammation in endothelial cells under hyperglycemic condition by targeting HMGB1. Vasc. Pharmacol. 2017, 88, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, H.; Wang, Y.; Xuan, J.; Gao, X.; Ding, H.; Ma, C.; Chen, Y.; Yang, Y. Roles of physical exercise-induced MiR-126 in cardiovascular health of type 2 diabetes. Diabetol. Metab. Syndr. 2022, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Zeng, M.; Yang, Y.; Liang, Y.; Fu, P.; Dong, Z. Exosomes in Diabetic Kidney Disease. Kidney Dis. 2023, 9, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zhou, M.; Zhang, J.; Huang, X.; Jiang, D.; Liu, J.; Zhang, Q.; Chen, D.; Hu, Q. Mechanism of LncRNA-MiRNA in Renal Intrinsic Cells of Diabetic Kidney Disease and Potential Therapeutic Direction. DNA Cell Biol. 2025, 44, 304–324. [Google Scholar] [CrossRef]
- Li, G.; Zhang, J.; Liu, D.; Wei, Q.; Wang, H.; Lv, Y.; Ye, Z.; Liu, G.; Li, L. Identification of Hub Genes and Potential ceRNA Networks of Diabetic Nephropathy by Weighted Gene Co-Expression Network Analysis. Front. Genet. 2021, 12. [Google Scholar] [CrossRef]
- Yu, Y.; Jia, Y.-Y.; Wang, M.; Mu, L.; Li, H.-J. PTGER3 and MMP-2 play potential roles in diabetic nephropathy via competing endogenous RNA mechanisms. BMC Nephrol. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Li, L.; Long, J.; Mise, K.; Galvan, D.L.; Overbeek, P.A.; Tan, L.; Kumar, S.V.; Chan, W.K.; Lorenzi, P.L.; Chang, B.H.; et al. PGC1α is required for the renoprotective effect of lncRNA Tug1 in vivo and links Tug1 with urea cycle metabolites. Cell Rep. 2021, 36, 109510–109510. [Google Scholar] [CrossRef]
- Sun, S.F.; Tang, P.M.; Feng, M.; Xiao, J.; Huang, X.R.; Li, P.; Ma, R.C.; Lan, H.Y. Novel lncRNA Erbb4-IR Promotes Diabetic Kidney Injury in db/db Mice by Targeting miR-29b. Diabetes 2017, 67, 731–744. [Google Scholar] [CrossRef]
- Jie, R.; Zhu, P.; Zhong, J.; Zhang, Y.; Wu, H. LncRNA KCNQ1OT1 affects cell proliferation, apoptosis and fibrosis through regulating miR-18b-5p/SORBS2 axis and NF-ĸB pathway in diabetic nephropathy. Diabetol. Metab. Syndr. 2020, 12, 1–11. [Google Scholar] [CrossRef]
- Huang, Q.; Qiu, T.; Chen, H.; Tian, T.; Wang, D.; Lu, C. Silencing LncRNA SNHG14 alleviates renal tubular injury via the miR-483-5p/HDAC4 axis in diabetic kidney disease. Hormones 2024, 24, 123–135. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, G.; Ge, Z. lncRNA MALAT1 Promotes Renal Fibrosis in Diabetic Nephropathy by Targeting the miR-2355-3p/IL6ST Axis. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- He, K.; Chen, Z.; Zhao, J.; He, Y.; Deng, R.; Fan, X.; Wang, J.; Zhou, X. The role of microRNA-155 in glomerular endothelial cell injury induced by high glucose. Mol. Biol. Rep. 2022, 49, 2915–2924. [Google Scholar] [CrossRef]
- Gao, J.; Liang, Z.; Zhao, F.; Liu, X.; Ma, N. Triptolide inhibits oxidative stress and inflammation via the microRNA-155-5p/brain-derived neurotrophic factor to reduce podocyte injury in mice with diabetic nephropathy. Bioengineered 2022, 13, 12275–12288. [Google Scholar] [CrossRef]
- Guo, M.; Dai, Y.; Jiang, L.; Gao, J. Bioinformatics Analysis of the Mechanisms of Diabetic Nephropathy via Novel Biomarkers and Competing Endogenous RNA Network. Front. Endocrinol. 2022, 13, 934022. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.; Yang, J.; Sun, M.; Rui, T.; Yang, Z.; Shi, M. Depression of LncRNA DANCR alleviates tubular injury in diabetic nephropathy by regulating KLF5 through sponge miR-214-5p. BMC Nephrol. 2024, 25, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Li, H.; Yu, W. Xanthohumol ameliorates diabetic kidney disease through suppression of renal fibrosis by regulating SNHG10/miR-378b. Front. Pharmacol. 2025, 16, 1532517. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, Y.-Q.; Yang, J.-E. Stress-Related LncRNAs and Their Roles in Diabetes and Diabetic Complications. Int. J. Mol. Sci. 2025, 26, 2194. [Google Scholar] [CrossRef]
- Su, X.; Huang, H.; Lai, J.; Lin, S.; Huang, Y. Long noncoding RNAs as potential diagnostic biomarkers for diabetes mellitus and complications: A systematic review and meta-analysis. J. Diabetes 2023, 16. [Google Scholar] [CrossRef]
- Shu, H.; Zhang, Z.; Liu, J.; Chen, P.; Yang, C.; Wu, Y.; Wu, D.; Cao, Y.; Chu, Y.; Li, L. Circular RNAs: An emerging precise weapon for diabetic nephropathy diagnosis and therapy. Biomed. Pharmacother. 2023, 168, 115818. [Google Scholar] [CrossRef]
- Wang, Q.; Cang, Z.; Shen, L.; Peng, W.; Xi, L.; Jiang, X.; Ge, X.; Xu, B.; Huang, S. circ_0037128/miR-17-3p/AKT3 axis promotes the development of diabetic nephropathy. Gene 2021, 765, 145076. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Han, Q.; Zhao, L.; Wang, L. Circular RNA circRNA_15698 aggravates the extracellular matrix of diabetic nephropathy mesangial cells via miR-185/TGF-β1. J. Cell. Physiol. 2018, 234, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, S.; Li, W.; Zhao, M.; Li, K. Circ_0000491 Promotes Apoptosis, Inflammation, Oxidative Stress, and Fibrosis in High Glucose-Induced Mesangial Cells by Regulating miR-455-3p/Hmgb1 Axis. Nephron 2021, 146, 72–83. [Google Scholar] [CrossRef]
- Wen, S.; Li, S.; Li, L.; Fan, Q. circACTR2: A Novel Mechanism Regulating High Glucose-Induced Fibrosis in Renal Tubular Cells via Pyroptosis. Biol. Pharm. Bull. 2020, 43, 558–564. [Google Scholar] [CrossRef]
- Xu, B.; Wang, Q.; Li, W.; Xia, L.; Ge, X.; Shen, L.; Cang, Z.; Peng, W.; Shao, K.; Huang, S. Circular RNA circEIF4G2 aggravates renal fibrosis in diabetic nephropathy by sponging miR-218. J. Cell. Mol. Med. 2020, 26, 1799–1805. [Google Scholar] [CrossRef]
- Yao, T.; Zha, D.; Hu, C.; Wu, X. Circ_0000285 promotes podocyte injury through sponging miR-654-3p and activating MAPK6 in diabetic nephropathy. Gene 2020, 747, 144661. [Google Scholar] [CrossRef]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.-A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. Rev. 2019, 14, 50–59. [Google Scholar] [CrossRef]
- Kanter, J.E.; Hsu, C.-C.; Bornfeldt, K.E. Monocytes and Macrophages as Protagonists in Vascular Complications of Diabetes. Front. Cardiovasc. Med. 2020, 7, 10. [Google Scholar] [CrossRef]
- Hou, G.; Dong, Y.; Jiang, Y.; Zhao, W.; Zhou, L.; Cao, S.; Li, W. Immune inflammation and metabolic interactions in the pathogenesis of diabetic nephropathy. Front. Endocrinol. 2025, 16, 1602594. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, X.; Li, Z.; Wang, Q. Immune cell contribution to vascular complications in diabetes. Front. Endocrinol. 2025, 16, 1549945. [Google Scholar] [CrossRef]
- Kzhyshkowska, J.; Neyen, C.; Gordon, S. Role of macrophage scavenger receptors in atherosclerosis. Immunobiology 2012, 217, 492–502. [Google Scholar] [CrossRef]
- Kzhyshkowska, J.; Gratchev, A.; Brundiers, H.; Mamidi, S.; Krusell, L.; Goerdt, S. Phosphatidylinositide 3-kinase activity is required for stabilin-1-mediated endosomal transport of acLDL. Immunobiology 2005, 210, 161–173. [Google Scholar] [CrossRef]
- Kazakova, E.; Iamshchikov, P.; Larionova, I.; Kzhyshkowska, J. Macrophage scavenger receptors: Tumor support and tumor inhibition. Front. Oncol. 2023, 12, 1096897. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Dai, H.; Zhou, Q.; Meng, X. The immunology of diabetic cardiomyopathy. Front. Endocrinol. 2025, 16, 1542208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhu, X.; Chen, Y.; Wen, Z.; Shi, P.; Ni, Q. The role and therapeutic potential of macrophages in the pathogenesis of diabetic cardiomyopathy. Front. Immunol. 2024, 15, 1393392. [Google Scholar] [CrossRef] [PubMed]
- Torres, Á.; Muñoz, K.; Nahuelpán, Y.; Saez, A.-P.R.; Mendoza, P.; Jara, C.; Cappelli, C.; Suarez, R.; Oyarzún, C.; Quezada, C.; et al. Intraglomerular Monocyte/Macrophage Infiltration and Macrophage–Myofibroblast Transition during Diabetic Nephropathy Is Regulated by the A2B Adenosine Receptor. Cells 2020, 9, 1051. [Google Scholar] [CrossRef]
- Saadane, A.; Veenstra, A.A.; Minns, M.S.; Tang, J.; Du, Y.; Elghazali, F.A.; Lessieur, E.M.; Pearlman, E.; Kern, T.S. CCR2-positive monocytes contribute to the pathogenesis of early diabetic retinopathy in mice. Diabetologia 2023, 66, 590–602. [Google Scholar] [CrossRef]
- Torres-Castro, I.; Arroyo-Camarena, Ú.D.; Martínez-Reyes, C.P.; Gómez-Arauz, A.Y.; Duẽnas-Andrade, Y.; Hernández-Ruiz, J.; Béjar, Y.L.; Zaga-Clavellina, V.; Morales-Montor, J.; Terrazas, L.I.; et al. Corrigendum to ‘Human monocytes and macrophages undergo M1-type inflammatory polarization in response to high levels of glucose’ [Immunol. Lett. 176 (2016) 81–89]. Immunol. Lett. 2017, 192, 106. [Google Scholar] [CrossRef]
- Moganti, K.; Li, F.; Schmuttermaier, C.; Riemann, S.; Klüter, H.; Gratchev, A.; Harmsen, M.C.; Kzhyshkowska, J. Hyperglycemia induces mixed M1/M2 cytokine profile in primary human monocyte-derived macrophages. Immunobiology 2017, 222, 952–959. [Google Scholar] [CrossRef]
- Zhao, J.; Fang, Z. Single-cell RNA sequencing reveals the dysfunctional characteristics of PBMCs in patients with type 2 diabetes mellitus. Front. Immunol. 2025, 15, 1501660. [Google Scholar] [CrossRef]
- Wang, S.; Li, Y.; Liang, Y.; Dong, J.; He, Y.; Zhang, L.; Yan, Y. Expression of miR-18a and miR-34c in circulating monocytes associated with vulnerability to type 2 diabetes mellitus and insulin resistance. J. Cell. Mol. Med. 2017, 21, 3372–3380. [Google Scholar] [CrossRef]
- Wassaifi, S.; Kaeffer, B.; Zarrouk, S. Cellular Phenotypic Transformation During Atherosclerosis: The Potential Role of miRNAs as Biomarkers. Int. J. Mol. Sci. 2025, 26, 2083. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.W.; Dickson, A.M.; Clay, G.; McCaffrey, A.P.; Wilson, M.E. Identifying Functional MicroRNAs in Macrophages with Polarized Phenotypes. J. Biol. Chem. 2012, 287, 21816–21825. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Zhong, M.; Suo, Q.; Lv, K. Expression profiles of miRNAs in polarized macrophages. Int. J. Mol. Med. 2013, 31, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Chen, H.; Wei, M.; Chen, X.; Zhang, Y.; Cao, L.; Yuan, P.; Wang, F.; Yang, G.; Ma, J. Gold nanoparticle-based miR155 antagonist macrophage delivery restores the cardiac function in ovariectomized diabetic mouse model. Int. J. Nanomed. 2017, 12, 4963–4979. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.; Reddy, S.S.; Maurya, M.; Maurya, P.; Barthwal, M.K. MicroRNA-99a mimics inhibit M1 macrophage phenotype and adipose tissue inflammation by targeting TNFα. Cell. Mol. Immunol. 2019, 16, 495–507. [Google Scholar] [CrossRef]
- Sun, J.; Huang, Q.; Li, S.; Meng, F.; Li, X.; Gong, X. miR-330-5p/Tim-3 axis regulates macrophage M2 polarization and insulin resistance in diabetes mice. Mol. Immunol. 2018, 95, 107–113. [Google Scholar] [CrossRef]
- Hu, F.; Tong, J.; Deng, B.; Zheng, J.; Lu, C. MiR-495 regulates macrophage M1/M2 polarization and insulin resistance in high-fat diet-fed mice via targeting FTO. Pfl?gers Arch. Eur. J. Physiol. 2019, 471, 1529–1537. [Google Scholar] [CrossRef]
- Szydełko, J.; Matyjaszek-Matuszek, B. MicroRNAs as Biomarkers for Coronary Artery Disease Related to Type 2 Diabetes Mellitus—From Pathogenesis to Potential Clinical Application. Int. J. Mol. Sci. 2022, 24, 616. [Google Scholar] [CrossRef]
- Kuschnerus, K.; Straessler, E.T.; Müller, M.F.; Lüscher, T.F.; Landmesser, U.; Kränkel, N. Increased Expression of miR-483-3p Impairs the Vascular Response to Injury in Type 2 Diabetes. Diabetes 2018, 68, 349–360. [Google Scholar] [CrossRef]
- Liu, G.; Yan, D.; Yang, L.; Sun, Y.; Zhan, L.; Lu, L.; Jin, Z.; Zhang, C.; Long, P.; Chen, J.; et al. The effect of miR-471-3p on macrophage polarization in the development of diabetic cardiomyopathy. Life Sci. 2021, 268, 118989. [Google Scholar] [CrossRef]
- Rawal, S.; Randhawa, V.; Rizvi, S.H.M.; Sachan, M.; Wara, A.K.; Pérez-Cremades, D.; Weisbrod, R.M.; Hamburg, N.M.; Feinberg, M.W. miR-369-3p ameliorates diabetes-associated atherosclerosis by regulating macrophage succinate-GPR91 signalling. Cardiovasc. Res. 2024, 120, 1693–1712. [Google Scholar] [CrossRef]
- Mone, P.; Varzideh, F.; Jankauskas, S.S.; Pansini, A.; Lombardi, A.; Frullone, S.; Santulli, G. SGLT2 Inhibition via Empagliflozin Improves Endothelial Function and Reduces Mitochondrial Oxidative Stress: Insights From Frail Hypertensive and Diabetic Patients. Hypertension 2022, 79, 1633–1643. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, Y.; Cai, Z.; Ferrari, M.W.; Li, C.; Zhang, T.; Lyu, G.; Wang, Z. MiR-21-3p inhibitor exerts myocardial protective effects by altering macrophage polarization state and reducing excessive mitophagy. Commun. Biol. 2024, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.A.; Amaram, V.; Das, S.; Tanwar, V.S.; Ganguly, R.; Wang, M.; Lanting, L.; Zhang, L.; Abdollahi, M.; Chen, Z.; et al. LncRNA DRAIR is downregulated in diabetic monocytes and modulates inflammatory phenotype via epigenetic mechanisms. J. Clin. Investig. 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Reddy, M.A.; Senapati, P.; Stapleton, K.; Lanting, L.; Wang, M.; Amaram, V.; Ganguly, R.; Zhang, L.; Devaraj, S.; et al. Diabetes Mellitus–Induced Long Noncoding RNA Dnm3os Regulates Macrophage Functions and Inflammation via Nuclear Mechanisms. Arter. Thromb. Vasc. Biol. 2018, 38, 1806–1820. [Google Scholar] [CrossRef]
- Gonzalez, C.; Cimini, M.; Cheng, Z.; Benedict, C.; Wang, C.; Trungcao, M.; Mallaredy, V.; Rajan, S.; Garikipati, V.N.S.; Kishore, R. Role of circular RNA cdr1as in modulation of macrophage phenotype. Life Sci. 2022, 309, 121003–121003. [Google Scholar] [CrossRef]
- El Khayari, A.; Hakam, S.M.; Malka, G.; Rochette, L.; El Fatimy, R. New insights into the cardio-renal benefits of SGLT2 inhibitors and the coordinated role of miR-30 family. Genes Dis. 2024, 11, 101174. [Google Scholar] [CrossRef]
- Cinti, F.; Laborante, R.; Cappannoli, L.; Morciano, C.; Gugliandolo, S.; Pontecorvi, A.; Burzotta, F.; Donniacuo, M.; Cappetta, D.; Patti, G.; et al. The effects of SGLT2i on cardiac metabolism in patients with HFpEF: Fact or fiction? Cardiovasc. Diabetol. 2025, 24, 1–13. [Google Scholar] [CrossRef]
- Kang, S.; Verma, S.; Hassanabad, A.F.; Teng, G.; Belke, D.D.; Dundas, J.A.; Guzzardi, D.G.; Svystonyuk, D.A.; Pattar, S.S.; Park, D.S.; et al. Direct Effects of Empagliflozin on Extracellular Matrix Remodelling in Human Cardiac Myofibroblasts: Novel Translational Clues to Explain EMPA-REG OUTCOME Results. Can. J. Cardiol. 2020, 36, 543–553. [Google Scholar] [CrossRef]
- Lin, K.; Yang, N.; Luo, W.; Qian, J.-F.; Zhu, W.-W.; Ye, S.-J.; Yuan, C.-X.; Xu, D.-Y.; Liang, G.; Huang, W.-J.; et al. Direct cardio-protection of Dapagliflozin against obesity-related cardiomyopathy via NHE1/MAPK signaling. Acta Pharmacol. Sin. 2022, 43, 2624–2635. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, M.; Suo, M.; Liu, D.; Wang, X.; Liu, M.; Pan, J.; Jin, T.; An, F. Dapagliflozin alleviates cardiac fibrosis through suppressing EndMT and fibroblast activation via AMPKα/TGF-β/Smad signalling in type 2 diabetic rats. J. Cell. Mol. Med. 2021, 25, 7642–7659. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Xu, Y.; Wang, D.; Chen, F.; Tu, Z.; Qian, J.; Xu, S.; Xu, Y.; Hwa, J.; Li, J.; et al. Cardioprotective mechanism of SGLT2 inhibitor against myocardial infarction is through reduction of autosis. Protein Cell 2021, 13, 336–359. [Google Scholar] [CrossRef] [PubMed]
- El-Shafey, M.; El-Agawy, M.S.E.-D.; Eldosoky, M.; Ebrahim, H.A.; Elsherbini, D.M.A.; El-Sherbiny, M.; Asseri, S.M.; Elsherbiny, N.M. Role of Dapagliflozin and Liraglutide on Diabetes-Induced Cardiomyopathy in Rats: Implication of Oxidative Stress, Inflammation, and Apoptosis. Front. Endocrinol. 2022, 13, 862394. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhu, Q.-X.; Li, G.-Z.; Wang, T.; Zhou, H. Empagliflozin ameliorates diabetic cardiomyopathy probably via activating AMPK/PGC-1α and inhibiting the RhoA/ROCK pathway. World J. Diabetes 2023, 14, 1862–1876. [Google Scholar] [CrossRef]
- Sun, P.; Wang, Y.; Ding, Y.; Luo, J.; Zhong, J.; Xu, N.; Zhang, Y.; Xie, W. Canagliflozin attenuates lipotoxicity in cardiomyocytes and protects diabetic mouse hearts by inhibiting the mTOR/HIF-1α pathway. iScience 2021, 24, 102521. [Google Scholar] [CrossRef]
- Santos-Gallego, C.G.; Requena-Ibanez, J.A.; Antonio, R.S.; Ishikawa, K.; Watanabe, S.; Picatoste, B.; Flores, E.; Garcia-Ropero, A.; Sanz, J.; Hajjar, R.J.; et al. Empagliflozin Ameliorates Adverse Left Ventricular Remodeling in Nondiabetic Heart Failure by Enhancing Myocardial Energetics. JACC 2019, 73, 1931–1944. [Google Scholar] [CrossRef]
- Zhang, G.; Zuo, Q.; Ma, S.; He, L.; Wang, Z.; Zhai, J.; Zhang, T.; Wang, Y.; Guo, Y. Metabolomic Analysis of the Effects of Canagliflozin on HFpEF Rats and Its Underlying Mechanism. Endocrine, Metab. Immune Disord. - Drug Targets 2025, 25, 1–16. [Google Scholar] [CrossRef]
- Al-Hayali, M.A.; Sozer, V.; Durmus, S.; Erdenen, F.; Altunoglu, E.; Gelisgen, R.; Atukeren, P.; Atak, P.G.; Uzun, H. Clinical Value of Circulating Microribonucleic Acids miR-1 and miR-21 in Evaluating the Diagnosis of Acute Heart Failure in Asymptomatic Type 2 Diabetic Patients. Biomolecules 2019, 9, 193. [Google Scholar] [CrossRef]
- Marketou, M.; Kontaraki, J.; Zacharis, E.; Maragkoudakis, S.; Fragkiadakis, K.; Kampanieris, E.; Plevritaki, A.; Savva, E.; Malikides, O.; Chlouverakis, G.; et al. Peripheral Blood MicroRNA-21 as a Predictive Biomarker for Heart Failure With Preserved Ejection Fraction in Old Hypertensives. Am. J. Hypertens. 2023, 37, 298–305. [Google Scholar] [CrossRef]
- Abolhasani, S.; Ahmadi, Y.; Fattahi, D.; Rostami, Y.; Chollou, K.M. microRNA-Mediated Regulation of Oxidative Stress in Cardiovascular Diseases. J. Clin. Lab. Anal. 2025, 39, e70017. [Google Scholar] [CrossRef]
- Tao, L.; Huang, X.; Xu, M.; Qin, Z.; Zhang, F.; Hua, F.; Jiang, X.; Wang, Y. Value of circulating miRNA-21 in the diagnosis of subclinical diabetic cardiomyopathy. Mol. Cell. Endocrinol. 2020, 518, 110944. [Google Scholar] [CrossRef]
- Sessa, F.; Salerno, M.; Esposito, M.; Cocimano, G.; Pisanelli, D.; Malik, A.; Khan, A.A.; Pomara, C. New Insight into Mechanisms of Cardiovascular Diseases: An Integrative Analysis Approach to Identify TheranoMiRNAs. Int. J. Mol. Sci. 2023, 24, 6781. [Google Scholar] [CrossRef]
- Rezazadeh-Gavgani, E.; Oladghaffari, M.; Bahramian, S.; Majidazar, R.; Dolati, S. MicroRNA-21: A critical underestimated molecule in diabetic retinopathy. Gene 2023, 859, 147212. [Google Scholar] [CrossRef] [PubMed]
- Ridwan, M.; Dimiati, H.; Syukri, M.; Lesmana, R. Effects of SGLT2-inhibitor on The Expression of MicroRNA-21, Transforming Growth Factor-β1, and Matrix Metalloproteinase-2 in The Process of Cardiac Fibrosis in Hyperglycemic Model Rats. Indones. Biomed. J. 2024, 16, 48–55. [Google Scholar] [CrossRef]
- Li, X.; Meng, C.; Han, F.; Yang, J.; Wang, J.; Zhu, Y.; Cui, X.; Zuo, M.; Xu&Ast, J.; Chang, B.; Xu; Ast, J. ; Chang, B. Vildagliptin Attenuates Myocardial Dysfunction and Restores Autophagy via miR-21/SPRY1/ERK in Diabetic Mice Heart. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Mone, P.; Lombardi, A.; Kansakar, U.; Varzideh, F.; Jankauskas, S.S.; Pansini, A.; Marzocco, S.; De Gennaro, S.; Famiglietti, M.; Macina, G.; et al. Empagliflozin Improves the MicroRNA Signature of Endothelial Dysfunction in Patients with Heart Failure with Preserved Ejection Fraction and Diabetes. J. Pharmacol. Exp. Ther. 2022, 384, 116–122. [Google Scholar] [CrossRef]
- Gou, L.; Zhao, L.; Song, W.; Wang, L.; Liu, J.; Zhang, H.; Huang, Y.; Lau, C.W.; Yao, X.; Tian, X.Y.; et al. Inhibition of miR-92a Suppresses Oxidative Stress and Improves Endothelial Function by Upregulating Heme Oxygenase-1 in db/db Mice. Antioxidants Redox Signal. 2018, 28, 358–370. [Google Scholar] [CrossRef]
- Sardu, C.; D'ONofrio, N.; Trotta, M.C.; Balestrieri, M.L.; Nicoletti, G.F.; D'AMico, G.; Fumagalli, C.; Contaldi, C.; Pacileo, G.; Scisciola, L.; et al. Could Ghrelin Expression Regulate Diastolic Cardiac Function in Type 2 Diabetic Obese Patients? Diabetes/Metabolism Res. Rev. 2025, 41, e70049. [Google Scholar] [CrossRef]
- Huang, J.-P.; Chang, C.-C.; Kuo, C.-Y.; Huang, K.-J.; Sokal, E.M.; Chen, K.-H.; Hung, L.-M. Exosomal microRNAs miR-30d-5p and miR-126a-5p Are Associated with Heart Failure with Preserved Ejection Fraction in STZ-Induced Type 1 Diabetic Rats. Int. J. Mol. Sci. 2022, 23, 7514. [Google Scholar] [CrossRef]
- Zhang, W.-Y.; Wang, J.; Li, A.-Z. A study of the effects of SGLT-2 inhibitors on diabetic cardiomyopathy through miR-30d/KLF9/VEGFA pathway. 2020, 24, 6346–6359.
- Li, X.; Du, N.; Zhang, Q.; Li, J.; Chen, X.; Liu, X.; Hu, Y.; Qin, W.; Shen, N.; Xu, C.; et al. MicroRNA-30d regulates cardiomyocyte pyroptosis by directly targeting foxo3a in diabetic cardiomyopathy. Cell Death Dis. 2014, 5, e1479–e1479. [Google Scholar] [CrossRef]
- Shen, J.; Huang, M.; Ye, Z.; Jing, Y.; Yuan, Z.; Xiang, Z.; Ding, M.; Li, T.; Ye, Q. Empagliflozin attenuates ventricular fibrillation postmyocardial infarction associated with reduced transforming growth factor- β 1/Smad3 signaling and miR-181a expression. J. Int. Med Res. 2025, 53. [Google Scholar] [CrossRef]
- Chen, F.; Li, W.; Zhang, D.; Fu, Y.; Yuan, W.; Luo, G.; Liu, F.; Luo, J. MALAT1 regulates hypertrophy of cardiomyocytes by modulating the miR-181a/HMGB2 pathway. Eur. J. Histochem. 2022, 66. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Wang, L.; Espinosa-Diez, C.; Wang, B.; Hahn, S.A.; Noda, K.; Rochon, E.R.; Dent, M.R.; Levine, A.R.; Baust, J.J.; et al. Metabolic Syndrome Mediates ROS-miR-193b-NFYA–Dependent Downregulation of Soluble Guanylate Cyclase and Contributes to Exercise-Induced Pulmonary Hypertension in Heart Failure With Preserved Ejection Fraction. Circulation 2021, 144, 615–637. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhao, M.; Li, Z.; Nie, H.; He, J.; Chen, Z.; Yuan, J.; Guo, H.; Zhang, X.; Yang, H.; et al. Plasma miR-193b-3p Is Elevated in Type 2 Diabetes and Could Impair Glucose Metabolism. Front. Endocrinol. 2022, 13, 814347. [Google Scholar] [CrossRef] [PubMed]
- Solini, A.; Seghieri, M.; Giannini, L.; Biancalana, E.; Parolini, F.; Rossi, C.; Dardano, A.; Taddei, S.; Ghiadoni, L.; Bruno, R.M. The Effects of Dapagliflozin on Systemic and Renal Vascular Function Display an Epigenetic Signature. J. Clin. Endocrinol. Metab. 2019, 104, 4253–4263. [Google Scholar] [CrossRef]
- Lai, L.; Chen, J.; Wang, N.; Zhu, G.; Duan, X.; Ling, F. MiRNA-30e mediated cardioprotection of ACE2 in rats with Doxorubicin-induced heart failure through inhibiting cardiomyocytes autophagy. Life Sci. 2017, 169, 69–75. [Google Scholar] [CrossRef]
- Wang, W.; Wu, C.; Ren, L.; Bao, Y.; Han, Y.; Li, C.; Li, Y. MiR-30e-5p is sponged by Kcnq1ot1 and represses Angiotensin II-induced hypertrophic phenotypes in cardiomyocytes by targeting ADAMExp. Cell Res. 2020, 394, 112140. [Google Scholar] [CrossRef]
- Li, Z.; Song, Y.; Liu, L.; Hou, N.; An, X.; Zhan, D.; Li, Y.; Zhou, L.; Li, P.; Yu, L.; et al. miR-199a impairs autophagy and induces cardiac hypertrophy through mTOR activation. Cell Death Differ. 2015, 24, 1205–1213. [Google Scholar] [CrossRef]
- el Azzouzi, H.; Leptidis, S.; Dirkx, E.; Hoeks, J.; van Bree, B.; Brand, K.; McClellan, E.A.; Poels, E.; Sluimer, J.C.; Hoogenhof, M.M.v.D.; et al. The Hypoxia-Inducible MicroRNA Cluster miR-199a∼214 Targets Myocardial PPARδ and Impairs Mitochondrial Fatty Acid Oxidation. Cell Metab. 2013, 18, 341–354. [Google Scholar] [CrossRef]
- Li, G.; Zhao, C.; Fang, S. SGLT2 promotes cardiac fibrosis following myocardial infarction and is regulated by miR-141. Exp. Ther. Med. 2021, 22, 1–9. [Google Scholar] [CrossRef]
- Rykova, E.Y.; Klimontov, V.V.; Shmakova, E.; Korbut, A.I.; Merkulova, T.I.; Kzhyshkowska, J. Anti-Inflammatory Effects of SGLT2 Inhibitors: Focus on Macrophages. Int. J. Mol. Sci. 2025, 26, 1670. [Google Scholar] [CrossRef]
- Roy, B. Pathophysiological Mechanisms of Diabetes-Induced Macrovascular and Microvascular Complications: The Role of Oxidative Stress. Med Sci. 2025, 13, 87. [Google Scholar] [CrossRef]
- Castoldi, G.; Carletti, R.; Barzaghi, F.; Meani, M.; Zatti, G.; Perseghin, G.; Di Gioia, C.R.T.; Zerbini, G. Sodium Glucose Cotransporter-2 Inhibitors in Non-Diabetic Kidney Disease: Evidence in Experimental Models. Pharmaceuticals 2024, 17, 362. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, L.; Dai, Y.; Lian, Y.; Song, H.; Dai, X.; Su, R.; Yin, J.; Gu, R. Dapagliflozin inhibits ferroptosis and ameliorates renal fibrosis in diabetic C57BL/6J mice. Sci. Rep. 2025, 15, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Chen, Y.; Li, N.; Yang, X.; Zhou, L.; Li, H.; Wang, T.; Xie, M.; Liu, H. Dapagliflozin delays renal fibrosis in diabetic kidney disease by inhibiting YAP/TAZ activation. Life Sci. 2023, 322, 121671. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tai, S.; Zhang, N.; Fu, L.; Wang, Y. Dapagliflozin prevents oxidative stress-induced endothelial dysfunction via sirtuin 1 activation. Biomed. Pharmacother. 2023, 165, 115213. [Google Scholar] [CrossRef] [PubMed]
- Panchapakesan, U.; Pegg, K.; Gross, S.; Komala, M.G.; Mudaliar, H.; Forbes, J.; Pollock, C.; Mather, A. Effects of SGLT2 Inhibition in Human Kidney Proximal Tubular Cells—Renoprotection in Diabetic Nephropathy? PLOS ONE 2013, 8, e54442. [Google Scholar] [CrossRef]
- Das, N.A.; Carpenter, A.J.; Belenchia, A.; Aroor, A.R.; Noda, M.; Siebenlist, U.; Chandrasekar, B.; DeMarco, V.G. Empagliflozin reduces high glucose-induced oxidative stress and miR-21-dependent TRAF3IP2 induction and RECK suppression, and inhibits human renal proximal tubular epithelial cell migration and epithelial-to-mesenchymal transition. Cell. Signal. 2020, 68, 109506–109506. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, Z.; Huang, H.; Wu, X. RNA-Seq Analysis of ceRNA-Related Networks in the Regulatory Metabolic Pathway of Mice with Diabetic Nephropathy Subjected to Empagliflozin Intervention. Arch. Espanoles De Urol. 2023, 76, 680–689. [Google Scholar] [CrossRef]
- Aroor, A.R.; Das, N.A.; Carpenter, A.J.; Habibi, J.; Jia, G.; Ramirez-Perez, F.I.; Martinez-Lemus, L.; Manrique-Acevedo, C.M.; Hayden, M.R.; Duta, C.; et al. Glycemic control by the SGLT2 inhibitor empagliflozin decreases aortic stiffness, renal resistivity index and kidney injury. Cardiovasc. Diabetol. 2018, 17, 1–14. [Google Scholar] [CrossRef]
- Al-Tantawy, S.M.; Eraky, S.M.; Eissa, L.A. Promising renoprotective effect of gold nanoparticles and dapagliflozin in diabetic nephropathy via targeting miR-192 and miR-21. J. Biochem. Mol. Toxicol. 2023, 37, e23430. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, D.I.; Khairy, E.; Saad, S.S.; Habib, E.K.; Hamouda, M.A. Potential protective effects of Dapagliflozin in gentamicin induced nephrotoxicity rat model via modulation of apoptosis associated miRNAs. Gene 2019, 707, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Shepard, B.D.; Chau, J.; Kurtz, R.; Rosenberg, A.Z.; Sarder, P.; Border, S.P.; Ginley, B.; Rodriguez, O.; Albanese, C.; Knoer, G.; et al. Nascent shifts in renal cellular metabolism, structure, and function due to chronic empagliflozin in prediabetic mice. Am. J. Physiol. Physiol. 2024, 326, C1272–C1290. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Xie, T.; Liu, T.; Chen, Z.; Yu, L.; Zhang, C.; Luo, J.; Chen, L.; Zhao, X.; Xiao, Y. An integrated RNA sequencing and network pharmacology approach reveals the molecular mechanism of dapagliflozin in the treatment of diabetic nephropathy. Front. Endocrinol. 2022, 13, 967822. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Guo, F.; Liu, Y.; Huang, F.; Fan, X.; Zhao, L.; Qin, G. Identification of circular RNAs and functional competing endogenous RNA networks in human proximal tubular epithelial cells treated with sodium-glucose cotransporter 2 inhibitor dapagliflozin in diabetic kidney disease. Bioengineered 2022, 13, 3911–3929. [Google Scholar] [CrossRef]
- Mashayekhi, M.; Safa, B.I.; Gonzalez, M.S.; Kim, S.F.; Echouffo-Tcheugui, J.B. Systemic and organ-specific anti-inflammatory effects of sodium-glucose cotransporter-2 inhibitors. Trends Endocrinol. Metab. 2024, 35, 425–438. [Google Scholar] [CrossRef]
- Shen, J.; Ying, L.; Wu, J.; Fang, Y.; Zhou, W.; Qi, C.; Gu, L.; Mou, S.; Yan, Y.; Tian, M.; et al. Integrative ATAC-seq and RNA-seq analysis associated with diabetic nephropathy and identification of novel targets for treatment by dapagliflozin. Cell Biochem. Funct. 2024, 42, e3943. [Google Scholar] [CrossRef]
- Li, J.; Wei, C.; Yang, Y.; Gao, Z.; Guo, Z.; Qi, F. Apoptotic bodies extracted from adipose mesenchymal stem cells carry microRNA-21–5p to induce M2 polarization of macrophages and augment skin wound healing by targeting KLFBurns 2022, 48, 1893–1908. [CrossRef]
- Tian, S.; Zhou, X.; Zhang, M.; Cui, L.; Li, B.; Liu, Y.; Su, R.; Sun, K.; Hu, Y.; Yang, F.; et al. Mesenchymal stem cell-derived exosomes protect against liver fibrosis via delivering miR-148a to target KLF6/STAT3 pathway in macrophages. Stem Cell Res. Ther. 2022, 13, 1–20. [Google Scholar] [CrossRef]
- Zhang, N.-X.; Guan, C.; Li, C.-Y.; Xu, L.-Y.; Xin, Y.-L.; Song, Z.; Li, T.-Y.; Yang, C.-Y.; Zhao, L.; Che, L.; et al. Formononetin Alleviates Ischemic Acute Kidney Injury by Regulating Macrophage Polarization through KLF6/STAT3 Pathway. Am. J. Chin. Med. 2024, 52, 1487–1505. [Google Scholar] [CrossRef]
- Xiong, S.-X.; Huang, L.-J.; Liu, H.-S.; Zhang, X.-X.; Li, M.; Cui, Y.-B.; Shao, C.; Hu, X.-L. Dapagliflozin exerts anti-apoptotic effects by mitigating macrophage polarization via modulation of the phosphoinositide 3-kinase/protein kinase B signaling pathway. World J. Diabetes 2025, 16, 97287. [Google Scholar] [CrossRef]
- Abdollahi, E.; Keyhanfar, F.; Delbandi, A.-A.; Falak, R.; Hajimiresmaiel, S.J.; Shafiei, M. Dapagliflozin exerts anti-inflammatory effects via inhibition of LPS-induced TLR-4 overexpression and NF-κB activation in human endothelial cells and differentiated macrophages. Eur. J. Pharmacol. 2022, 918, 174715. [Google Scholar] [CrossRef]
- Bhatt, K.; Lanting, L.L.; Jia, Y.; Yadav, S.; Reddy, M.A.; Magilnick, N.; Boldin, M.; Natarajan, R. Anti-Inflammatory Role of MicroRNA-146a in the Pathogenesis of Diabetic Nephropathy. J. Am. Soc. Nephrol. 2016, 27, 2277–2288. [Google Scholar] [CrossRef]
- Yin, R.; Zhu, X.; Wang, J.; Yang, S.; Ma, A.; Xiao, Q.; Song, J.; Pan, X. MicroRNA-155 promotes the ox-LDL-induced activation of NLRP3 inflammasomes via the ERK1/2 pathway in THP-1 macrophages and aggravates atherosclerosis in ApoE−/− mice. Ann. Palliat. Med. 2019, 8, 676–689. [Google Scholar] [CrossRef]
- Cole, J.B.; Florez, J.C. Genetics of diabetes mellitus and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Imamura, M.; Maeda, S. Perspectives on genetic studies of type 2 diabetes from the genome-wide association studies era to precision medicine. J. Diabetes Investig. 2024, 15, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Chhichholiya, Y.; Suryan, A.K.; Suman, P.; Munshi&Ast, A.; Singh, S. SNPs in miRNAs and Target Sequences: Role in Cancer and Diabetes. Front. Genet. 2021, 12. [CrossRef]
- Jahantigh, D.; Sargazi, F.M.; Sargazi, S.; Saravani, R.; Zidanloo, S.G.; Nia, M.H.; Piri, M. Relationship between Functional miR-143/145 Cluster Variants and Susceptibility to Type 2 Diabetes Mellitus: A Preliminary Case-Control Study and Bioinformatics Analyses. Endocr. Res. 2021, 46, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Duan, Y.; Wang, J.; Liu, Y. A functional polymorphism of microRNA-143 is associated with the risk of type 2 diabetes mellitus in the northern Chinese Han population. Front. Endocrinol. 2022, 13, 994953. [Google Scholar] [CrossRef]
- Blumensatt, M.; Wronkowitz, N.; Wiza, C.; Cramer, A.; Mueller, H.; Rabelink, M.J.; Hoeben, R.C.; Eckel, J.; Sell, H.; Ouwens, D.M. Adipocyte-derived factors impair insulin signaling in differentiated human vascular smooth muscle cells via the upregulation of miR-143. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2014, 1842, 275–283. [Google Scholar] [CrossRef]
- Wei, J.; Ma, Z.; Li, Y.; Zhao, B.; Wang, D.; Jin, Y.; Jin, Y. miR-143 inhibits cell proliferation by targeting autophagy-related 2B in non-small cell lung cancer H1299 cells. Mol. Med. Rep. 2014, 11, 571–576. [Google Scholar] [CrossRef]
- He, M.; Wu, N.; Leong, M.C.; Zhang, W.; Ye, Z.; Li, R.; Huang, J.; Zhang, Z.; Li, L.; Yao, X.; et al. miR-145 improves metabolic inflammatory disease through multiple pathways. J. Mol. Cell Biol. 2019, 12, 152–162. [Google Scholar] [CrossRef]
- Sun, D.; Zhu, Z.; Zhang, Y.; Bai, R.; Zhu, F.; Shan, Z.; Ma, C.; Yang, J. Relation of genetic polymorphisms in microRNAs with diastolic and systolic function in type 2 diabetes mellitus. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 2877–2882. [Google Scholar] [CrossRef]
- Hu, W.; Li, T.; Hu, R.; Wu, L.; Li, M.; Meng, X. MicroRNAlet-7aandlet-7fas novel regulatory factors of the sika deer (Cervus nippon)IGF-1Rgene. Growth Factors 2013, 32, 27–33. [Google Scholar] [CrossRef]
- Santovito, D.; De Nardis, V.; Marcantonio, P.; Mandolini, C.; Paganelli, C.; Vitale, E.; Buttitta, F.; Bucci, M.; Mezzetti, A.; Consoli, A.; et al. Plasma Exosome MicroRNA Profiling Unravels a New Potential Modulator of Adiponectin Pathway in Diabetes: Effect of Glycemic Control. J. Clin. Endocrinol. Metab. 2014, 99, E1681–E1685. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, H.; Karimi, J.; Khodadadi, I.; Tavilani, H. Correlation between miR-103 and miR-133a Expression and the Circulating ANGPTL8 in Type 2 Diabetic Patients and Healthy Control Subjects. Clin. Lab. 2019, 65, 2149–2155. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, Y.; Huang, J.; Liu, Y.; Zhu, Y. Association of genetic variants in lncRNA GAS5/miR-21/mTOR axis with risk and prognosis of coronary artery disease among a Chinese population. J. Clin. Lab. Anal. 2020, 34, e23430. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yang, M.; Yang, Y.; Xu, F.; Wang, F.; Jiao, M.; Tao, W.; Li, Y. Association of MiRNA Polymorphisms Involved in the PI3K/ATK/GSK3β Pathway with T2DM in a Chinese Population. Pharmacogenomics Pers. Med. 2025, ume 18, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Rahman, B.; Haq, T.U.; Jalil, F.; Khan, B.M.; Maodaa, S.N.; Al-Farraj, S.A.; El-Serehy, H.A.; Shah, A.A. Deciphering the Variants Located in the MIR196A2, MIR146A, and MIR423 with Type-2 Diabetes Mellitus in Pakistani Population. Genes 2021, 12, 664. [Google Scholar] [CrossRef]
- Buraczynska, M.; Zukowski, P.; Wacinski, P.; Ksiazek, K.; Zaluska, W. Polymorphism in microRNA-196a2 contributes to the risk of cardiovascular disease in type 2 diabetes patients. J. Diabetes its Complicat. 2014, 28, 617–620. [Google Scholar] [CrossRef]
- Deng, Z.; Gao, W.; Luo, W.; Ai, L.; Hu, M. Significant Association between Microrna Gene Polymorphisms and Type 2 Diabetes Mellitus Susceptibility in Asian Population: A Meta-Analysis. Iran. J. Public Heal. 2020, 49, 830–836. [Google Scholar] [CrossRef]
- Locke, J.M.; Allen, H.L.; Harries, L.W. A rare SNP in pre-miR-34a is associated with increased levels of miR-34a in pancreatic beta cells. Acta Diabetol. 2013, 51, 325–329. [Google Scholar] [CrossRef]
- Mone, P.; de Donato, A.; Varzideh, F.; Kansakar, U.; Jankauskas, S.S.; Pansini, A.; Santulli, G. Functional role of miR-34a in diabetes and frailty. Front. Aging 2022, 3, 949924. [Google Scholar] [CrossRef]
- Mone, P.; Jankauskas, S.S.; Manzi, M.V.; Gambardella, J.; Coppola, A.; Kansakar, U.; Izzo, R.; Fiorentino, G.; Lombardi, A.; Varzideh, F.; et al. Endothelial Extracellular Vesicles Enriched in microRNA-34a Predict New-Onset Diabetes in Coronavirus Disease 2019 (COVID-19) Patients: Novel Insights for Long COVID Metabolic Sequelae. J. Pharmacol. Exp. Ther. 2024, 389, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, W.; Ning, M.; Zhou, X.; Wan, X.; Mi, Q.; Yang, X.; Zhang, D.; Zhang, Y.; Jiang, B.; et al. A functional SNP rs895819 on pre-miR-27a is associated with bipolar disorder by targeting NCAM1. Commun. Biol. 2022, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liu, B.; Jiang, Q.; Zhang, J.; Xin, S.; Xu, K. The association of two common polymorphisms in miRNAs with diabetes mellitus. Medicine 2019, 98, e17414. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.; Elfaki, I.; Javid, J.; Barnawi, J.; Altayar, M.A.; Albalawi, S.O.; Jalal, M.M.; Tayeb, F.J.; Yousif, A.; Ullah, M.F.; et al. Genetic Determinants of Cardiovascular Disease: The Endothelial Nitric Oxide Synthase 3 (eNOS3), Krüppel-Like Factor-14 (KLF-14), Methylenetetrahydrofolate Reductase (MTHFR), MiRNAs27a and Their Association with the Predisposition and Susceptibility to Coronary Artery Disease. Life 2022, 12, 1905. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, O.-K. Potential Roles of Adipocyte Extracellular Vesicle–Derived miRNAs in Obesity-Mediated Insulin Resistance. Adv. Nutr. Int. Rev. J. 2020, 12, 566–574. [Google Scholar] [CrossRef]
- El-Jawad, A.M.A.; Ibrahim, I.H.; Zaki, M.E.; Elias, T.R.; Rasheed, W.I.; Amr, K.S. The potential role of miR-27a and miR-320a in metabolic syndrome in obese Egyptian females. J. Genet. Eng. Biotechnol. 2022, 20, 75–10. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, Y.; Liu, Y.; Zhu, D.; Yu, J.; Li, G.; Sun, Z.; Wang, W.; Jiang, H.; Hong, Z. MiR-27a promotes insulin resistance and mediates glucose metabolism by targeting PPAR-γ-mediated PI3K/AKT signaling. Aging 2019, 11, 7510–7524. [Google Scholar] [CrossRef]
- Burada, E.; Roșu, M.-M.; Sandu, R.E.; Burada, F.; Cucu, M.G.; Streață, I.; Petre-Mandache, B.; Popescu-Hobeanu, G.; Cara, M.-L.; Țucă, A.-M.; et al. miR-499a rs3746444 A>G Polymorphism Is Correlated with Type 2 Diabetes Mellitus and Diabetic Polyneuropathy in a Romanian Cohort: A Preliminary Study. Genes 2023, 14, 1543. [Google Scholar] [CrossRef]
- Wang, F.; Liu, D.; Zhang, R.-R.; Yu, L.-W.; Zhao, J.-Y.; Yang, X.-Y.; Jiang, S.-S.; Ma, D.; Qiao, B.; Zhang, F.; et al. A TBX5 3′UTR variant increases the risk of congenital heart disease in the Han Chinese population. Cell Discov. 2017, 3, 17026–17026. [Google Scholar] [CrossRef]
- Alipoor, B.; Ghaedi, H.; Meshkani, R.; Omrani, M.D.; Sharifi, Z.; Golmohammadi, T. The rs2910164 variant is associated with reduced miR-146a expression but not cytokine levels in patients with type 2 diabetes. J. Endocrinol. Investig. 2017, 41, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Chen, X.; Yang, M.; Zhao, X.; Chen, Y.; Zhao, H.; Liu, C.; Shen, C. The variant at TGFBRAP1 is significantly associated with type 2 diabetes mellitus and affects diabetes-related miRNA expression. J. Cell. Mol. Med. 2018, 23, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Voight, B.F.; Lyssenko, V.; Burtt, N.P.; de Bakker, P.I.W.; Chen, H.; Roix, J.J.; Kathiresan, S.; Hirschhorn, J.N.; Daly, M.J.; et al. Genome-Wide Association Analysis Identifies Loci for Type 2 Diabetes and Triglyceride Levels. Science 2007, 316, 1331–1336. [Google Scholar] [CrossRef]
- Chauhan, G.; Spurgeon, C.J.; Tabassum, R.; Bhaskar, S.; Kulkarni, S.R.; Mahajan, A.; Chavali, S.; Kumar, M.K.; Prakash, S.; Dwivedi, O.P.; et al. Impact of Common Variants of PPARG, KCNJ11, TCF7L2, SLC30A8, HHEX, CDKN2A, IGF2BP2, and CDKAL1 on the Risk of Type 2 Diabetes in 5,164 Indians. Diabetes 2010, 59, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Sharma, R.B.; Ly, S.; Stamateris, R.E.; Jesdale, W.M.; Alonso, L.C. CDKN2A/B T2D Genome-Wide Association Study Risk SNPs Impact Locus Gene Expression and Proliferation in Human Islets. Diabetes 2018, 67, 872–884. [Google Scholar] [CrossRef]
- Dechamethakun, S.; Muramatsu, M. Long noncoding RNA variations in cardiometabolic diseases. J. Hum. Genet. 2016, 62, 97–104. [Google Scholar] [CrossRef]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.-M. Molecular Interplay of the Noncoding RNA ANRIL and Methylated Histone H3 Lysine 27 by Polycomb CBX7 in Transcriptional Silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef]
- Li, X.; Yang, A.; Liu, X.; Zhang, R.; Zhou, H.; Xu, S. Genetic association of long non-coding RNA ANRIL polymorphism with the risk of type 2 diabetes mellitus in the Chinese Han population. Diabetol. Metab. Syndr. 2025, 17, 1–11. [Google Scholar] [CrossRef]
- González-Moro, I.; Rojas-Márquez, H.; Sebastian-Delacruz, M.; Mentxaka-Salgado, J.; Olazagoitia-Garmendia, A.; Mendoza, L.M.; Lluch, A.; Fantuzzi, F.; Lambert, C.; Blanco, J.A.; et al. A long non-coding RNA that harbors a SNP associated with type 2 diabetes regulates the expression of TGM2 gene in pancreatic beta cells. Front. Endocrinol. 2023, 14, 1101934. [Google Scholar] [CrossRef]
- Brondani, L.d.A.; Dandolini, I.; Girardi, E.; Canani, L.H.; Crispim, D.; Dieter, C. Association between the G/G genotype of the lncRNA MEG3 rs7158663 polymorphism and proliferative diabetic retinopathy. Arq. Bras. de Endocrinol. Metabol. 2024, 68, e240024. [Google Scholar] [CrossRef]
- Chuang, C.-C.; Wang, K.; Yang, Y.-S.; Kornelius, E.; Tang, C.-H.; Lee, C.-Y.; Chien, H.-W.; Yang, S.-F. Association of Long Noncoding RNA HOTAIR Polymorphism and the Clinical Manifestations of Diabetic Retinopathy. Int. J. Environ. Res. Public Heal. 2022, 19, 14592. [Google Scholar] [CrossRef]
- Yao, Y.-P.; Chien, H.-W.; Wang, K.; Yang, Y.-S.; Su, S.-C.; Chang, L.-C.; Lin, H.-Y.; Yang, S.-F. Genetic association of diabetic retinopathy with long noncoding RNA CDKN2B-AS1 gene polymorphism. Eur. J. Ophthalmol. 2024, 35, 637–644. [Google Scholar] [CrossRef]
- Yang, P.-J.; Ting, K.-H.; Tsai, P.-Y.; Su, S.-C.; Yang, S.-F. Association of long noncoding RNA GAS5 gene polymorphism with progression of diabetic kidney disease. Int. J. Med Sci. 2024, 21, 2201–2207. [Google Scholar] [CrossRef]
- Yari, A.; Karam, Z.M.; Meybodi, S.M.E.; Sargazi, M.L.; Saeidi, K. CDKN2B-AS (rs2891168), SOD2 (rs4880), and PON1 (rs662) polymorphisms and susceptibility to coronary artery disease and type 2 diabetes mellitus in Iranian patients: A case-control study. Heal. Sci. Rep. 2023, 6, e1717. [Google Scholar] [CrossRef] [PubMed]
- Letonja, J.; Petrovič, D.; Petrovič, D. CDKN2B-AS1 polymorphism rs1333049 is associated with advanced carotid artery atherosclerosis in a Slovenian population. Biomol. Biomed. 2024, 25, 1362–1368. [Google Scholar] [CrossRef]
- Jiménez-Osorio, A.S.; Musalem-Younes, C.; Cárdenas-Hernández, H.; Solares-Tlapechco, J.; Costa-Urrutia, P.; Medina-Contreras, O.; Granados, J.; López-Saucedo, C.; Estrada-Garcia, T.; Rodríguez-Arellano, M.E. Common Polymorphisms Linked to Obesity and Cardiovascular Disease in Europeans and Asians are Associated with Type 2 Diabetes in Mexican Mestizos. Medicina 2019, 55, 40. [Google Scholar] [CrossRef] [PubMed]
- Florez, J.C.; Pearson, E.R. A roadmap to achieve pharmacological precision medicine in diabetes. Diabetologia 2022, 65, 1830–1838. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Haffner, S.M.; Heise, M.A.; Herman, W.H.; Holman, R.R.; Jones, N.P.; Kravitz, B.G.; Lachin, J.M.; O'Neill, M.C.; Zinman, B.; et al. Glycemic Durability of Rosiglitazone, Metformin, or Glyburide Monotherapy. New Engl. J. Med. 2006, 355, 2427–2443. [Google Scholar] [CrossRef]
- Kreienkamp, R.J.; Voight, B.F.; Gloyn, A.L.; Udler, M.S. Genetics of Type 2 Diabetes; 2023.
- MetGen Investigators; Zhou, K.; DPP Investigators; ACCORD Investigators; Yee, S.W.; Seiser, E.L.; van Leeuwen, N.; Tavendale, R.; Bennett, A.J.; Groves, C.J.; et al. Variation in the glucose transporter gene SLC2A2 is associated with glycemic response to metformin. Nat. Genet. 2016, 48, 1055–1059. [CrossRef]
- Li, J.H.; Perry, J.A.; Jablonski, K.A.; Srinivasan, S.; Chen, L.; Todd, J.N.; Harden, M.; Mercader, J.M.; Pan, Q.; Dawed, A.Y.; et al. Identification of Genetic Variation Influencing Metformin Response in a Multiancestry Genome-Wide Association Study in the Diabetes Prevention Program (DPP). Diabetes 2023, 72, 1161–1172. [Google Scholar] [CrossRef]
- Dawed, A.Y.; Yee, S.W.; Zhou, K.; van Leeuwen, N.; Zhang, Y.; Siddiqui, M.K.; Etheridge, A.; Innocenti, F.; Xu, F.; Li, J.H.; et al. Genome-Wide Meta-analysis Identifies Genetic Variants Associated With Glycemic Response to Sulfonylureas. Diabetes Care 2021, 44, 2673–2682. [Google Scholar] [CrossRef] [PubMed]
- Umapathysivam, M.M.; Araldi, E.; Hastoy, B.; Dawed, A.Y.; Vatandaslar, H.; Sengupta, S.; Kaufmann, A.; Thomsen, S.; Hartmann, B.; Jonsson, A.E.; et al. Type 2 Diabetes Risk Alleles in Peptidyl-Glycine Alpha-Amidating Monooxygenase Influence GLP-1 Levels and Response to GLP-1 Receptor Agonists 2023.
- Geng, Z.; Li, Q.; Huang, R.; Wang, J.; Weng, J.; Zhou, K.; Liang, H.; Wang, Y. KCNQ1 Variant rs163184 is a Potential Biomarker of Glycemic Response to Exenatide. Pharmacogenomics 2022, 23, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, K.; Liu, H.; Cao, R. GLP1R Variant is Associated with Response to Exenatide in Overweight Chinese Type 2 Diabetes Patients. Pharmacogenomics 2019, 20, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Zhao, L.; Zhang, X.; Li, H.; Yin, Z. Effect of rs67085638 in long non-coding RNA (CCAT1) on colon cancer chemoresistance to paclitaxel through modulating the microRNA-24-3p and FSCN1. J. Cell. Mol. Med. 2021, 25, 3744–3753. [Google Scholar] [CrossRef]
- Wen, Q.-W.; Chen, C.-J.; Zhang, H.-Q.; Yu, H.; Zeng, Z.; Jin, T.; Wang, T.-H.; Qin, W.; Huang, H.; Wu, X.-D.; et al. Associations Between microRNA-related Genetic Polymorphisms and Clinical Response to Methotrexate in Chinese Rheumatoid Arthritis Patients. Curr. Pharm. Des. 2022, 28, 3167–3173. [Google Scholar] [CrossRef]
- Rykova, E.; Ershov, N.; Damarov, I.; Merkulova, T. SNPs in 3′UTR miRNA Target Sequences Associated with Individual Drug Susceptibility. Int. J. Mol. Sci. 2022, 23, 13725. [Google Scholar] [CrossRef]
- Paz-Pacheco, E.; Nevado, J.B.; la Paz, E.M.C.C.-D.; Jasul, G.V.; Aman, A.Y.C.L.; A Ribaya, E.L.; Francisco, M.D.G.; Guanzon, M.L.V.V.; Uyking-Naranjo, M.L.; Añonuevo-Cruz, M.C.S.; et al. Variants of SLC2A10 may be Linked to Poor Response to Metformin. J. Endocr. Soc. 2022, 6, bvac092. [Google Scholar] [CrossRef]
- Chen, Z.-R.; He, F.-Z.; Liu, M.-Z.; Hu, J.-L.; Xu, H.; Zhou, H.-H.; Zhang, W. MIR4532 gene variant rs60432575 influences the expression of KCNJ11 and the sulfonylureas-stimulated insulin secretion. Endocrine 2018, 63, 489–496. [Google Scholar] [CrossRef]
- Klen, J.; Dolžan, V. SGLT2 Inhibitors in the Treatment of Diabetic Kidney Disease: More than Just Glucose Regulation. Pharmaceutics 2023, 15, 1995. [Google Scholar] [CrossRef]
- Li, H.; Mai, Z.; Yu, S.; Wang, B.; Lai, W.; Chen, G.; Zhou, C.; Liu, J.; Yang, Y.; Chen, S.; et al. Exploring the Pleiotropic Genes and Therapeutic Targets Associated with Heart Failure and Chronic Kidney Disease by Integrating metaCCA and SGLT2 Inhibitors’ Target Prediction. BioMed Res. Int. 2021, 2021. [Google Scholar] [CrossRef]
- Degtyareva, A.O.; Antontseva, E.V.; Merkulova, T.I. Regulatory SNPs: Altered Transcription Factor Binding Sites Implicated in Complex Traits and Diseases. Int. J. Mol. Sci. 2021, 22, 6454. [Google Scholar] [CrossRef]
- Aguet, F.; Barbeira, A.N.; Bonazzola, R.; Jo, B.; Kasela, S.; Liang, Y.; Parsana, P.; Aguet, F.; Battle, A.; Brown, A.; et al. The GTEx Consortium The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science 2020, 369, 1318–1330. [Google Scholar] [CrossRef]
- Harvey, C.T.; Moyerbrailean, G.A.; Davis, G.O.; Wen, X.; Luca, F.; Pique-Regi, R. QuASAR: quantitative allele-specific analysis of reads. Bioinformatics 2014, 31, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Moyerbrailean, G.A.; Richards, A.L.; Kurtz, D.; Kalita, C.A.; Davis, G.O.; Harvey, C.T.; Alazizi, A.; Watza, D.; Sorokin, Y.; Hauff, N.; et al. High-throughput allele-specific expression across 250 environmental conditions. Genome Res. 2016, 26, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Arcelus, M.; Baglaenko, Y.; Arora, J.; Hannes, S.; Luo, Y.; Amariuta, T.; Teslovich, N.; Rao, D.A.; Ermann, J.; Jonsson, A.H.; et al. Allele-specific expression changes dynamically during T cell activation in HLA and other autoimmune loci. Nat. Genet. 2020, 52, 247–253. [Google Scholar] [CrossRef]
- Wei, H.-H.; Zheng, L.; Wang, Z. mRNA therapeutics: New vaccination and beyond. Fundam. Res. 2023, 3, 749–759. [Google Scholar] [CrossRef]
- Matboli, M.; Kamel, M.M.; Essawy, N.; Bekhit, M.M.; Abdulrahman, B.; Mohamed, G.F.; Eissa, S. Identification of Novel Insulin Resistance Related ceRNA Network in T2DM and Its Potential Editing by CRISPR/Cas9. Int. J. Mol. Sci. 2021, 22, 8129. [Google Scholar] [CrossRef]
- Scott, L.J.; Mohlke, K.L.; Bonnycastle, L.L.; Willer, C.J.; Li, Y.; Duren, W.L.; Erdos, M.R.; Stringham, H.M.; Chines, P.S.; Jackson, A.U.; et al. A Genome-Wide Association Study of Type 2 Diabetes in Finns Detects Multiple Susceptibility Variants. Science 2007, 316, 1341–1345. [Google Scholar] [CrossRef]
- Cook, J.P.; Morris, A.P. Multi-ethnic genome-wide association study identifies novel locus for type 2 diabetes susceptibility. Eur. J. Hum. Genet. 2016, 24, 1175–1180. [Google Scholar] [CrossRef]
- Grarup, N.; Rose, C.S.; Andersson, E.A.; Andersen, G.; Nielsen, A.L.; Albrechtsen, A.; Clausen, J.O.; Rasmussen, S.S.; Jørgensen, T.; Sandbæk, A.; et al. Studies of Association of Variants Near the HHEX, CDKN2A/B, and IGF2BP2 Genes With Type 2 Diabetes and Impaired Insulin Release in 10,705 Danish Subjects. Diabetes 2007, 56, 3105–3111. [Google Scholar] [CrossRef]
- Hribal, M.L.; Presta, I.; Procopio, T.; Marini, M.A.; Stančáková, A.; Kuusisto, J.; Andreozzi, F.; Hammarstedt, A.; Jansson, P.-A.; Grarup, N.; et al. Glucose tolerance, insulin sensitivity and insulin release in European non-diabetic carriers of a polymorphism upstream of CDKN2A and CDKN2B. Diabetologia 2011, 54, 795–802. [Google Scholar] [CrossRef]
- Hart, L.M. ‘.; Simonis-Bik, A.M.; Nijpels, G.; van Haeften, T.W.; Schäfer, S.A.; Houwing-Duistermaat, J.J.; Boomsma, D.I.; Groenewoud, M.J.; Reiling, E.; van Hove, E.C.; et al. Combined Risk Allele Score of Eight Type 2 Diabetes Genes Is Associated With Reduced First-Phase Glucose-Stimulated Insulin Secretion During Hyperglycemic Clamps. Diabetes 2009, 59, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Haq, T.U.; Zahoor, A.; Ali, Y.; Chen, Y.; Jalil, F.; Shah, A.A. Genetic Variants of MIR27A, MIR196A2 May Impact the Risk for the Onset of Coronary Artery Disease in the Pakistani Population. Genes 2022, 13, 747. [Google Scholar] [CrossRef] [PubMed]
- Vinelli-Arzubiaga, D.; Campos, C.E.S.; Laso-Salazar, M.C.; Abarca-Barriga, H. Polymorphic variants and risk of diabetic peripheral neuropathy in patients with type 2 diabetes mellitus: systematic review and meta-analysis. BMC Endocr. Disord. 2025, 25, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, J.; Tao, W.; Yang, M.; Wang, X.; Lu, T.; Li, C.; Yang, Y.; Yao, Y. The Single Nucleotide Polymorphisms (rs1292037 and rs13137) in miR-21 Were Associated with T2DM in a Chinese Population. Diabetes, Metab. Syndr. Obesity: Targets Ther. 2022, ume 15, 189–198. [CrossRef]
- Agiannitopoulos, K.; Samara, P.; Papadopoulou, M.; Efthymiadou, A.; Papadopoulou, E.; Tsaousis, G.N.; Mertzanos, G.; Babalis, D.; Lamnissou, K. miRNA polymorphisms and risk of premature coronary artery disease. Hell. J. Cardiol. 2021, 62, 278–284. [Google Scholar] [CrossRef]
- Zal, N.; Safi, S.; Ahmadieh, H.; Fekri, S.; Najafi, S.; Forouhari, A.; Moghaddasi, A.; Hejazi, M.; Kheiri, B.; Eslami, M.; et al. Assessment of MIR200B Polymorphisms Association with Sight-Threatening Diabetic Retinopathy. J. Curr. Ophthalmol. 2023, 35, 355–361. [Google Scholar] [CrossRef]
- Jianbing, H.; Xiaotian, L.; Jie, T.; Xueying, C.; Honge, J.; Bo, Z.; Lirong, H.; Lei, Z. The Effect of Allograft Inflammatory Factor-1 on Inflammation, Oxidative Stress, and Autophagy via miR-34a/ATG4B Pathway in Diabetic Kidney Disease. Oxidative Med. Cell. Longev. 2022, 2022, 1668000. [Google Scholar] [CrossRef]
- Zhuang, X.; Sun, Z.; Du, H.; Zhou, T.; Zou, J.; Fu, W. Metformin inhibits high glucose-induced apoptosis of renal podocyte through regulating miR-34a/SIRT1 axis. Immunity, Inflamm. Dis. 2024, 12, e1053. [Google Scholar] [CrossRef]
- Cardenas-Gonzalez, M.; Srivastava, A.; Pavkovic, M.; Bijol, V.; Rennke, H.G.; E Stillman, I.; Zhang, X.; Parikh, S.; Rovin, B.H.; Afkarian, M.; et al. Identification, Confirmation, and Replication of Novel Urinary MicroRNA Biomarkers in Lupus Nephritis and Diabetic Nephropathy. Clin. Chem. 2017, 63, 1515–1526. [Google Scholar] [CrossRef]
Figure 1.
Proposed mechanism of SGLT2 inhibitors via microRNA modulation in diabetic complications. The diagram illustrates the key cellular players and proposed miRNA networks in diabetic cardiomyopathy (left) and diabetic nephropathy (right). In both organs, specific cell types—including cardiomyocytes, endothelial cells, fibroblasts, podocytes, tubular epithelial cells, and mesangial cells—exhibit altered miRNA expression profiles under diabetic conditions, driving pathological processes such as fibrosis, hypertrophy, and inflammation. Infiltrating macrophages contribute to these pathways in both compartments. The model further highlights the emerging role of SGLT2 inhibitors (Empagliflozin and Dapagliflozin) in expression regulation of a subset of these dysregulated miRNAs (indicated by arrows), thereby mitigating cellular dysfunction and interrupting the progression of both complications.
Figure 1.
Proposed mechanism of SGLT2 inhibitors via microRNA modulation in diabetic complications. The diagram illustrates the key cellular players and proposed miRNA networks in diabetic cardiomyopathy (left) and diabetic nephropathy (right). In both organs, specific cell types—including cardiomyocytes, endothelial cells, fibroblasts, podocytes, tubular epithelial cells, and mesangial cells—exhibit altered miRNA expression profiles under diabetic conditions, driving pathological processes such as fibrosis, hypertrophy, and inflammation. Infiltrating macrophages contribute to these pathways in both compartments. The model further highlights the emerging role of SGLT2 inhibitors (Empagliflozin and Dapagliflozin) in expression regulation of a subset of these dysregulated miRNAs (indicated by arrows), thereby mitigating cellular dysfunction and interrupting the progression of both complications.
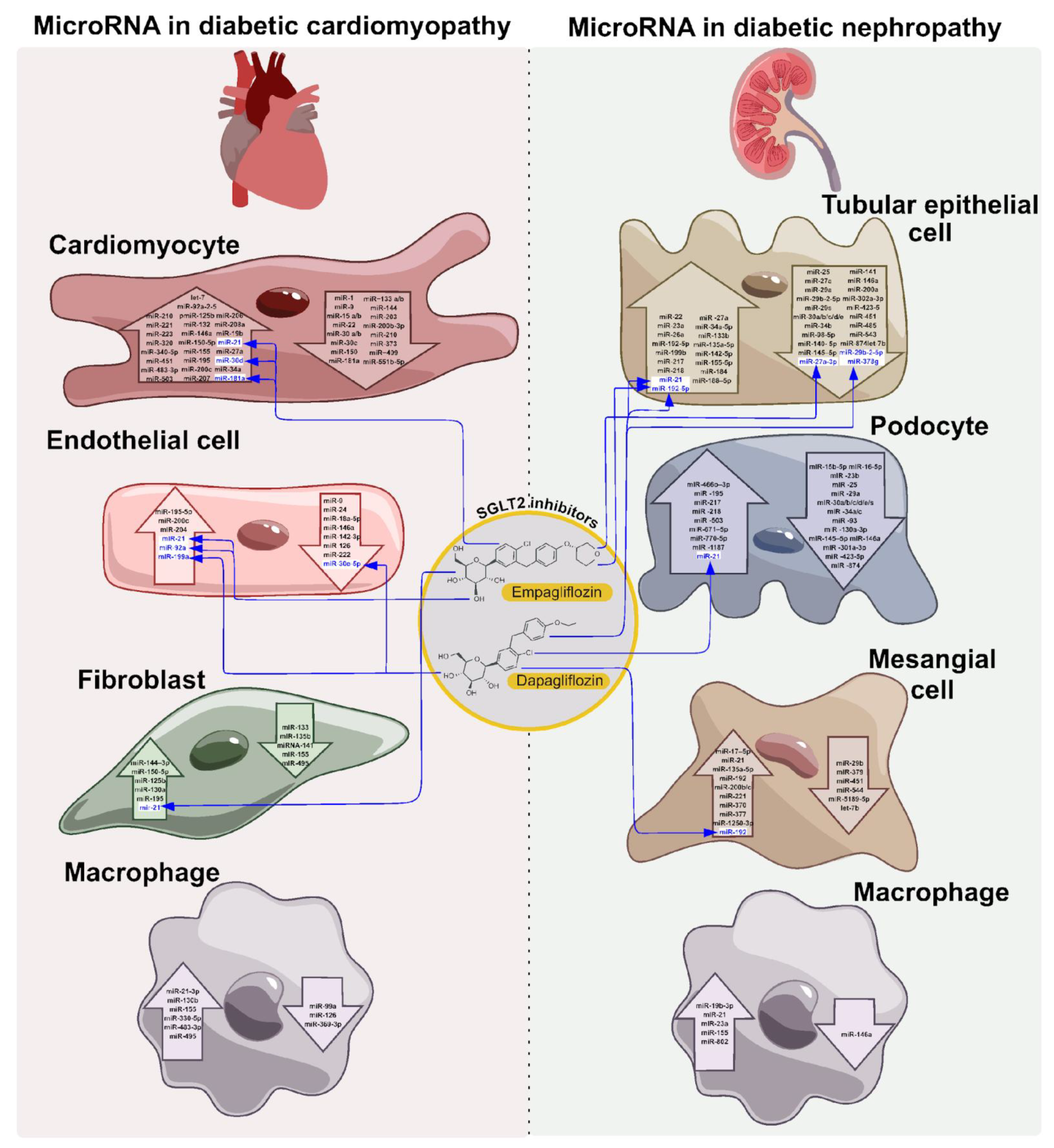

Table 1.
NcRNAs as modulators of therapeutic effects of SGLT inhibitors: focus on kidney and heart.
| ncRNA | Effect of specific inhibitor | Mouse/rat model, patients’ cohort | Kidney/ Heart | Major effect in cells or in circulation | Refs |
| miR-21 | empagliflozin treatment leads to decreased TGF-β1 level/ increased expression of the matrix metalloproteinase-2 (MMP-2) in heart tissue | rat model of STZ-induced hyperglycemia | Heart | empagliflozin administration for seven weeks decreases (compared to metformin) miR-21 levels in heart tissue | [207] |
| miR-21miR-92 | empagliflozin improves endothelial function by reducing mitochondrial calcium overload and the generation of reactive oxygen species | age-related T2DM patients with heart failure with preserved ejection fraction (HFpEF) | Heart HFpEF | empagliflozin (not metformin) decreases biomarkers of HF: circulating miR-21 and miR-92 levels in T2DM patients with HFpEF | [185,209] |
| miR-30d | SGLT2 inhibitor (not specified) improved cardiac function due to enhancing autophagy by reducing the expression level of miR-30d | rat model of diabetic cardiomyopathy on a high-fat diet and with (STZ)-induced diabetes | Heart DCM | SGLT2i improves DCM by affecting the miR-30d/KLF9/VEGFA pathway, which regulates the expression of autophagy factors protein light chain 3 (LC3-II) and p62/SQSTM1 (p62/sequestosome 1) | [213] |
| miR-193b | empagliflozin ameliorates metabolic syndrome and reduces mitochondrial reactive oxygen species generation | rat model of combined precapillary and postcapillary pulmonary hypertension (CpcPH): obese ZSF-1 rats treated with SU5416 to stimulate resting pulmonary hypertension | Heart, HFpEF | empagliflozin downregulates miR-193b, resulting in restoration of NFYA-sGCβ1-cGMP signaling in pulmonary arterial smooth muscle cells and improvement of EIPH symptoms | [217] |
| miR-30e-5p miR-199a-3p | short-term treatment (4 weeks) by dapagliflozin improves endothelial function, systemic vascular function | patients with T2DM and hypertension (blood pressure >130/80 mm Hg) | Heart TD2M hypertension | dapagliflozin increases circulating levels of antihypertrophic miR-30e-5p, decrease of circulating levels of prohypertrophic miR-199a-3p, which are involved in the pathophysiology of CV disease and heart failure | [219] |
| miR-27b and miR-200b | dapagliflozin demonstrates beneficial effect on the kidneys' ability to dilate blood vessels | patients with T2DM and hypertension (blood pressure >130/80 mm Hg) | Kidney renal vascular function | dapagliflozin treatment demonstrates the nephroprotective effect which correlates with the pretreatment levels of circulating miR-27b and miR-200b | [219] |
| miR-21 | empagliflozin treatment ameliorated HG-induced proinflammatory effects, and inhibited epithelial-to-mesenchymal transition (EMT) and HK-2 cell migration. | high glucose (HG) treated human kidney proximal tubule cells (HK-2) | Kidney | empagliflozin ameliorates AGE - induced RECK expression suppression via oxidative stress/TRAF3IP2/NF-κB and p38 MAPK/miR-21 pathways in HK-2 cells | [232] |
| miR-21miR-181a | dapagliflozin administration decreases markers of kidney injury, such as increased creatinine, blood urea, and urine protein | gentamicin-induced renal injury in Wistar rats | Kidney | dapagliflozin improves kidney function and structure in association with an increase of miR-21 and a decrease in miR-181a expression in kidney tissue | [236] |
| miR 27a-3p, 190a-5p, 196b-5p | empagliflozin treatment results in improved renal function parameters | murine model of early T2DM associated with obesity and insulin resistance on a high-fat diet | Kidney | empagliflozin changes urinary exosomal microRNA profiles, including increased levels of miR 27a-3p, 190a-5p, and 196b-5p. | [237] |
| circRNA_012448miR-29b-2-5p | the target genes of dapagliflozin-related circRNAs primarily focused on gene expression, glucose metabolic process, extracellular exosome, epigenetics, glyoxylate and dicarboxylate metabolism, amino acid metabolism, and lysosome pathways | HG-treated cultured human renal proximal tubular epithelial cells (HK-2) | Kidney |
dapagliflozin treatment of HG-treated HK-2 cells induced changes of multiple circRNAsexpression including the hsa_circRNA_012448-hsa-miR-29b-2-5p-GSK3β pathway |
[239] |
| lncRNA XR_382492.4, XR_873495.3, XR-388840.1, NR-015554.2, XR-382493.3, and XR-876705.2 | dapagliflozin improves glucose intolerance, high urinary albumin/creatinine ratio and renal damage | db/db mice model of T2DM nephropathy, RNA-seq study | Kidney DN | dapagliflozin treatment reversed the expression changes of XR_382492.4, XR_873495.3, XR-388840.1, NR-015554.2, XR-382493.3, and XR-876705.2 associated with DN in kidney tissue |
[238] |
| miR-466i-3p, miR466f-3p, miR-709, miR-207, and miR-297b-3p | the analysis of DEGs indicated that empagliflozin may inhibit the progression of diabetic nephropathy by inhibiting inflammation, apoptosis, and senescence | Murine streptozocin (STZ) - treated model of induced DN | Kidney DN | empagli-flozin treatment inhibits diabetic nephropa-thy ac-cording to the data of RNA-seq of kidney tis-sue from DN mice, empagli-flozin-treated DN mice and negative control | [233] |
Table 2.
MiRNA Genetics in T2DM.
|
SNPs/chromosome/ involvement in SGLTi effect |
SNP/gene and its associationwith diabetes complications | Description of the effect | Refs |
| rs2076380G>A Chromosome: 20:38165127 (GRCh38) 20:36793529 (GRCh37) Location: 20:38165127 Cytogenetic region:20q11.23 SGLT2-i – |
intron TGM2/ lncRNA TGM2 Mapped gene(s): TGM2 intron_variant lncRNA TGM2 is associated with T2DM No data in Diabetic Nephropathy, and Cardiomyopathy |
The AA genotype is associated with an increased risk of developing T2DM; risk allele A disrupts the secondary structure of this lncRNA, affecting its stability and the expression of TGM2 in pancreatic beta cells. Diminished LncTGM2 in human beta cells impairs glucose-stimulated insulin release. |
[285] |
| rs7158663A>G Chromosome: 14:100853087 (GRCh38) 14:101319424 (GRCh37) SGLT2-i – |
Exon MEG3/lnc MEG3 lncRNA MEG3 is associated with Diabetic Nephropathy No data in Diabetic Cardiomyopathy |
The GG genotype is associated with a protective effect against late-stage diabetic retinopathy in patients with type 2 diabetes. | [286] |
| rs12427129 C>T, Chromosome: 12:53973906 (GRCh38) 12:54367690 (GRCh37) rs1899663G>T Chromosome: 12:53967210 (GRCh38) 12:54360994 (GRCh37) SGLT2-i – |
intronHOTAIR/lnc HOTAIR Gene: HOXC11 HOTAIR lncRNA HOTAIR is associated with Diabetic Nephropathy, and Diabetic Cardiomyopathy |
The rs12427129-T allele is associated with an increased risk of diabetic retinopathy The rs1899663-TT genotype is associated with an increased risk of diabetic retinopathy |
[287] |
| rs1333049G>A,C Chromosome: 9:22125504 (GRCh38) 9:22125503 (GRCh37) SGLT2-i – Metformin + |
found at the intron of cyclin dependent kinase inhibitor 2B antisense RNA 1 (CDKN2B-AS1) lncRNA CDKN2B-AS1 is associated with Diabetic Nephropathy no data in Diabetic Cardiomyopathy |
rs1333049 have shown significant association with poor metformin response rs1333049 is found to be significantly associated with HbA1c level in Mexican nonobese T2DM patients rs1333049 (SNP) in CDKN2B-AS1, located in the 9p21 region.is associated with advanced carotid artery atherosclerosis |
[291,292,305] |
| rs2151280G>A Chromosome: 9:22034720 (GRCh38) 9:22034719 (GRCh37) SGLT2-i – |
intron CDKN2B-AS1/ lnc CDKN2B-AS1 Gene: CDKN2B-AS1 lncRNA CDKN2B-AS1 is associated with Diabetic Nephropathy |
The G allele is associated with an increased risk of proliferative diabetic retinopathy, decreased glomerular filtration rate and high-density lipoprotein levels | [288] |
| rs2891168A>G Chromosome: 9:22098620 (GRCh38) 9:22098619 (GRCh37) SGLT2-i – |
Gene: CDKN2B-AS1 lncRNA CDKN2B-AS1 is associated with Diabetic Nephropathy no data in Diabetic Cardiomyopathy |
susceptibility to coronary artery disease and type 2 diabetes mellitus | [290] |
| rs55829688 Chromosome: 1:173868168 (GRCh38) 1:173837306 (GRCh37) SGLT2-i – |
Gene: GAS5 lncRNA GAS5 is associated with Diabetic Nephropathy and Cardiomyopathy |
association of GAS5 gene polymorphisms with the progression of DKD. Carriers of at least one minor allele (C) of rs55829688 (TC and CC) more frequently suffer from advanced DKD than do those homozygotes for the major allele (TT). | [289] |
| rs55829688T>C Chromosome: 1:173868168 (GRCh38) 1:173837306 (GRCh37) SGLT2-i – |
intronGAS5/lnc GAS5 Gene: GAS5 lncRNA GAS5 is associated with Diabetic Nephropathy and Cardiomyopathy |
The C allele is associated with an increased risk of developing diabetic kidney disease, a decrease in the glomerular filtration rate | [289] |
| rs10811661T>C Chromosome: 9:22134095 (GRCh38) 9:22134094 (GRCh37) SGLT2-i – |
+13 kbfrom TSS ANRIL/lnc ANRIL lncRNA ANRIL is associated with Diabetic Nephropathy and Cardiomyopathy |
The T allele is associated with an increased risk of T2DM | [279,316,317] |
| The T allele is associated with decreased insulin secretion | [318,319,320] | ||
| The T allele is associated with increased expression of ANRIL in the islets of Langerhans | [281] | ||
| rs895819 A>G Chromosome: 19:13836478 (GRCh38) 19:13947292 (GRCh37) SGLT2-i + in Diabetic Nephropathy |
miRNAs27a Gene: MIR23AHG MIR23A MIR24-2 MIR27A miR-27a-3pand miR-27b are associated with T2DM, Coronary Artery Disease and Diabetic Nephropathy |
miR-27ars895819-GG genotype protects from coronary artery disease CAD risk subgroup analysis demonstrated that rs895819 C allele conveyed a significant protective effect against T2DM development in Caucasians |
[270,271,321] |
| rs3746444 Chromosome: 20:34990448 (GRCh38) 20:33578251 (GRCh37) SGLT2-i -- |
miR-499a Gene: MYH7B, MIR499A, MIR499B miR-499a is associated with T2DM and Diabetic Cardiomyopathy |
miR-499a rs3746444 A>G Polymorphism Is Correlated with T2DM and Diabetic Polyneuropathy with carriers of the GG genotype and the G allele being at an increased risk in the Romanian population | [275,322] |
| rs8089787T>A,C,G Chromosome: 18:21826640 (GRCh38) 18:19406601 (GRCh37) SGLT2-i – rs10877887T>C Chromosome: 12:62603400 (GRCh38) 12:62997180 (GRCh37) SGLT2-i – |
miR-133a-1 Gene: MIB1 MIR133A1 MIR133A1HG miR-133a-1 is associated with myocardial fibrosis MiRNA-let-7f Gene: LINC01465 MIRLET7I let-7f is associated with mitochondrial dysfunction and metabolic disturbances in T2DM |
miR-133a-1-rs8089787 and let-7f-rs10877887 were associated with impaired cardiac diastolic function in T2DM | [257] |
| rs10757278 A>G Chromosome: 9:22124478 (GRCh38) 9:22124477 (GRCh37) SGLT2-i – |
intron ANRIL/lncRNA ANRIL lncRNA ANRIL is associated with Diabetic Nephropathy and Diabetic Cardiomyopathy |
The relative fast plasma glucose (FPG) and HbA1c levels were relatively lower in individuals with the AA genotype and higher in those with the GG genotype | [284] |
| rs1292037T>C Chromosome: 17:59841547 (GRCh38) 17:57918908 (GRCh37) SGLT2-i + |
miR-21 Gene: VMP1 MIR21 miR-21 is associated with susceptibility to Coronary Heart Disease, Diabetic Cardiomyopathy andDiabetic Nephropathy |
rs1292037C allele and C/C genotype in miR-21 were strongly associated with elevated susceptibility to coronary heart disease (CHD) in a Chinese Han population | [261] |
| rs1292037T>C (see above) rs13137A>G,T Chromosome: 17:59841670 (GRCh38) 17:57919031 (GRCh37) miR-21 SGLT2-i + rs78312845A>C,G 17:7018112 (GRCh38) 17:6921431 (GRCh37) SGLT2-i – |
miR-21 Gene: VMP1 MIR21 miR-21 miR-21 is associated with susceptibility to Diabetic Cardiomyopathy and Diabetic Nephropathy miR-195 Gene: MIR195 miR-195 is associated with apoptosis in Diabetic Cardiomyopathy |
the C allele of rs1292037 in miR-21 could increase the risk of T2DM the T allele of rs13137 in miR-21 could be a risk factor for T2DM rs78312845 in miR-195 contributed to the level of fasting plasma glucose (FPG) and HbA1C in nondiabetic group in the Han Chinese population |
[323] |
| rs1076064A>G Chromosome: 5:149732603 (GRCh38) 5:149112166 (GRCh37 SGLT2-i – rs13283671C>A,T Chromosome: 9:21511741 (GRCh38) 9:21511740 (GRCh37) SGLT2-i – |
miR-378a Gene: MIR378A miR-378a-3pis associated with Diabetic Nephropathy miR-31 Gene: MIR31 MIR31HG C allele could increase the risk of developing T2DM miR-31-3p is associated with Diabetic Nephropathy |
The results showed that miR-378a rs1076064 G allele could be a protective factor in T2DM, whereas the miR-31 rs13283671 C allele could increase the risk of T2DM | [262] |
| rs11614913C>T Chromosome: 12:53991815 (GRCh38) 12:54385599 (GRCh37) SGLT2-i + |
exonMIR196A2/ stem-loop miR-196a-2 Gene: MIR196A2 miR 196b-5p is associated with Diabetic Nephropatahy |
The C allele is associated with an increased risk of T2DM | [263] |
| The T allele is associated with an increased risk of cardiovascular diseases in T2DM miR196a2 C>T (rs11614913) and miR499 A>G (rs3746444) were found to be strongly associated with increased risk for CAD |
[264,324] |
||
| rs72563729 G>A Chromosome: 1:1167183 (GRCh38) 1:1102563 (GRCh37) SGLT2-i + |
Gene: MIR200B miR 200b is associated with Diabetic Nephropathy miRNA-200c is associated with Diabetic cardiomyopathy |
allelic association of MIR200B variations with sight-threatening Diabetic Retinopathy | [325] |
| rs2910164C>G Chromosome: 5:160485411 (GRCh38) 5:159912418 (GRCh37) SGLT2-i – rs531564G>C Chromosome: 8:9903189 (GRCh38) 8:9760699 (GRCh37) SGLT2-i – |
exon MIR146A/ seed sequence miR-146a Gene: MIR146A MIR3142HG miRNA-146a is associated with inflammation in Diabetic Cardiomyopathy Gene: MIR124-1HG MIR124-1 miR-124a is a T2DM circulating marker |
The G allele is associated with an increased risk of T2DM | [263] |
| miR-146a rs2910164 (G allele) and miR-124a rs531564 (C allele) might function as protective factors in T2DM in Asian population | [265] | ||
| rs4705342T>C Chromosome: 5:149428408 (GRCh38) 5:148807971 (GRCh37) rs4705343T>C Chromosome: 5:149428518 (GRCh38) 5:148808081 (GRCh37) SGLT2-i – |
promoter region of a gene cluster MIR143/145/ miR143/145 Gene: MIR143 CARMN upstream_transcript_variant |
The C-rs4705342 and C-rs4705343 alleles are associated with an increased and decreased risk of developing T2DM, respectively The CC genotype of rs4705342 might be a risk factor in T2DM by increasing the expression of miRNA-143 in the northern Chinese Han population |
[252,253] |
| rs72631823 G>A Chromosome: 1:9151723 (GRCh38) 1:9211782 (GRCh37) SGLT2-i – Metformin+ |
exon MIR34A / terminal loop pre-miR-34a Gene: MIR34A MIR34AHG miR-34a is associated with Diabetic Kidney Disease |
Allele A promotes the formation of a relaxed form of the pre-miR-34a terminal loop, which facilitates its processing and, leads to an increase in miR-34a expression in INS-1 and MIN6 cells. The Effect of Allograft Inflammatory Factor-1 on Inflammation, Oxidative Stress, and Autophagy via miR-34a/ATG4B Pathway in Diabetic Kidney Disease |
[266,326,327] |
| rs60432575G>A Chromosome: 20:57895403 (GRCh38) 20:56470459 (GRCh37) SGLT2-i – Sulfonylurea drug+ |
MIR4532/ hsa-miR-4532 urinary microRNA biomarkers in lupus nephritis and Diabetic Nephropathy |
The G allele promotes efficient binding of miR-4532 to the 3'-untranslated region of KCNJ11 mRNA, resulting in decreased expression of KCNJ11 and Kir6.2 in HEK293 cells, as well as insulin secretion after stimulation of MIN6 cells with sulfonylurea drugs | [306,328] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



