Submitted:
03 October 2025
Posted:
07 October 2025
You are already at the latest version
Abstract
Cervical cancer remains a major global burden largely driven by persistent infection with high-risk human papillomavirus (HPV). Biological diversity across HPV phylogenetic clades (α-7 and α-9) may shape tumor programs and outcomes. To accelerate therapeutic options, we implemented an expression based, clade informed drug repurposing pipeline using OCTAD. We built disease signatures for HPV-α7 and HPV-α9 tumors, ranked compounds whose drug perturbation profiles inversely correlated with those signatures, and restricted analyses to FDA-approved agents. The screen yielded 41 and 52 candidates for α7 and α9, respectively, enriched for mechanistically coherent classes, including histone-deacetylase inhibitors, estrogen-pathway modulators, and HMG-CoA-reductase inhibitors. Notably, stronger signature inversion was associated with greater in-vitro sensitivity in relevant cell lines. These results support the feasibility of clade informed, transcriptome guided repurposing and nominate a concise, testable set of therapies for HPV-driven cervical cancer. While prospective validation in appropriate experimental models is essential, our findings highlight both shared and lineage-specific vulnerabilities that could guide rational combinations and biomarker driven studies.
Keywords:
cervical cancer
; HPV
; drug repositioning
; transcriptome signatures
; OCTAD
; precision oncology
1. Introduction
Cervical cancer (CC) is a major public health concern and ranks fourth worldwide for both incidence and mortality among women [1]. A substantial share of the burden is concentrated in low and middle income countries, where access to timely diagnosis and effective treatment remains limited and coverage of prophylactic vaccination programs is insufficient [2]. As a consequence, nearly 482,000 deaths are projected by 2040, representing a 38.2% increase compared with 2022 [1]. These figures underscore the urgent need to advance effective and accessible therapeutic strategies to improve survival and quality of life.
Etiologically, CC is attributable almost entirely to persistent infection with high-risk human papillomavirus (HPV) [3], whose oncogenes function as key carcinogenic drivers [4,5] Besides, during tumor progression, HPV helps sustain malignancy and may promote invasiveness [6,7]. Interestingly, high-risk genotypes display marked biological diversity that shapes molecular programs and clinical outcomes [8]. This diversity is evident at the level of HPV phylogenetic clades, among which the most clinically relevant are α-9, which includes HPV16, HPV31, HPV33, HPV35, HPV52, HPV58, and HPV67, and α-7, which includes HPV18, HPV39, HPV45, HPV59, HPV68, HPV70, HPV85, and HPV97 [9] In keeping with these groupings, tumors associated with presence of HPV genotypes belonging to α-7 clade, are typically less differentiated, more likely to harbor integrated viral transcripts, and more aggressive, patterns that correlate with poorer prognosis and shorter recurrence-free survival. By contrast, tumors linked to α-9 HPV clade, generally retain epithelial differentiation, show higher keratin expression, and predominantly exhibit episomal viral transcripts with lower levels of genomic integration, features that together are associated with comparatively better outcomes [9,10]. Despite these differences, it remains uncertain whether HPV genotype modulates therapeutic response, because the available evidence is inconsistent [11,12,13]. Consequently, genotype is not incorporated into routine clinical decision making, and management of advanced CC continues to rely on surgery, radiotherapy, and chemotherapy with cisplatin as the mainstay despite substantial toxicity and limited specificity [14,15,16,17]. Although targeted agents directed at VEGF, EGFR, or PD-L1 have shown promise, their clinical use remains limited and overall prognosis is still poor [18,19].
Within this context, the search for new therapeutic strategies is constrained by the high cost, long timelines, and high attrition of de novo drug development. Only about one in 5,000 to 10,000 anticancer candidates ultimately secures FDA approval, and each new molecular entity typically requires more than US$2 billion and approximately 13 years from discovery to market [20]. As a practical alternative, drug repurposing identifies new indications for already approved agents and thereby offers a faster, less expensive, lower-risk, and operationally efficient pathway to effective treatments compared to novo discovery [21]. In oncology, multiple successes have been reported for drugs originating both within and outside cancer therapeutics [22]; however, progress in cervical cancer has been comparatively slow, in large part due to tumor heterogeneity linked to its pathogenesis [19]. Nevertheless, several candidates could plausibly potentiate chemoradiotherapy or act synergistically with immunotherapy [23,24]. What remains limited, however, is preclinical and clinical evidence, including systematic assessment of adverse effects, which underscores the need for rigorous translational evaluation [24]
Building on this landscape, drug repositioning is becoming increasingly systematic, adopting a precision framework that integrates multi-omic and pharmacological resources with computational analytics to align disease mechanisms with existing agents, thereby advancing system level target discovery and therapeutic design [25,26,27]. A central pillar of this framework is expression based connectivity mapping, which compares disease transcriptional profiles with drug-perturbation signatures and prioritizes compounds whose profiles are inversely correlated with the disease state; this approach is grounded in the original Connectivity Map and its next-generation LINCS compendium that together provide millions of drug–gene expression relationships for systematic inference [28,29]. Within oncology, methodologic syntheses and case studies show how primary in silico integration, secondary prioritization, and tertiary experimental validation can be assembled into an efficient pipeline for precision repurposing across tumor contexts [21,30,31]. Empirically, expression-guided repurposing has produced prospective validations, including identification of pyrvinium pamoate for hepatocellular carcinoma with in vitro activity across multiple lines and tumor growth inhibition in xenografts [32], confirmation of auranofin and ganetespib activity in Ewing sarcoma with improved survival in vivo when combined[33], and discovery of tricyclic antidepressants with preclinical efficacy in small-cell lung cancer models[34] continued methodological refinements have further demonstrated expression-guided identification of homoharringtonine as a liver cancer candidate with supporting experiment[35].
Within this precision space, the Open Cancer TherApeutic Discovery platform operationalizes transcriptome-based connectivity mapping at the patient-group level by constructing disease signatures, matching them against large drug-perturbation repositories, and summarizing anticorrelation scores with built in validation steps; protocol-level demonstrations and related studies illustrate feasibility and reproducibility across indications[36]. Accordingly, by leveraging clade-specific HPV transcriptional programs and applying OCTAD-style screening, the present study provides a concise and prioritized set of plausible pharmacological candidates for cervical cancer that warrant systematic preclinical testing and careful translational evaluation.
2. Results
2.1. Differential Expression Analysis Across HPV Clades Supports Both Shared and Clade-Specific Transcriptional Programs
Building on previous genomic studies that have reported that high risk HPV phylogenetic clades differ in their molecular landscapes and clinical behavior [9,10,37], within we divided our dataset of primary cervical carcinomas into samples α-A7 and α-A9, according with of HPV genotype present in tumors [9,38]. We then performed differential gene expression analysis for tumors versus controls using edgeR, DESeq2, and limma-voom (Figure 1A). Most genes showed consistent expression patterns across methods, and we defined a consensus signature as the intersection of the three analytical outputs. Based on this definition, we identified 1,455 underexpressed and 548 overexpressed genes in α-A7 tumors, while α-A9 tumors displayed 2,037 underexpressed and 883 overexpressed genes (Figure 1B). Interestingly, we observed overlap between the two clades, with 53.5% of overexpressed and 60.6% of underexpressed genes shared.
To further explore the biological implications of these signatures, we performed functional enrichment analysis (Figure 1C). As expected, several pathways were shared between both clades, which is consistent with the fact that all samples correspond to primary tumors with comparable clinical stage distributions. These common processes primarily involved cell cycle regulation, chromosome segregation, extracellular matrix organization, and immune-related functions. Beyond these similarities, each clade displayed distinct enrichment patterns. α-A7 tumors displayed upregulation of cytoskeletal programs, including microtubule binding and annealing activity. In contrast, α-A9 tumors exhibited upregulation of interferon-mediated signaling and DNA helicase activity, alongside downregulation of histone H2AXY142 kinase activity and intracellularly calcium-gated channel activity. Overall, these results support the idea that, while α-A7 and α-A9 share a broad transcriptional program, each clade may exhibit distinct functional features consistent with their phylogenetic origin [10,37,39,40]. Such divergence has been suggested to contribute to differences in tumor biology and could potentially influence variability in treatment response [11,12,13].
(A) Volcano plots show significantly overexpressed and underexpressed genes in tumors with presence of α-A7 and α-A9 HPV clades, computed with edgeR (red), DESeq2 (blue), and limma-voom (green). Numbers of differentially expressed genes (DEGs) are indicated for each method. (B) Venn diagrams depicting the overlap of overexpressed and underexpressed DEGs across methods within each clade. (C) Gene Ontology (GO) enrichment analysis of consensus signatures. Selected biological processes (BP), cellular components (CC), and molecular functions (MF) are displayed for over- and underexpressed genes in each clade. Dot size reflects the number of genes per category, and color represents enrichment significance (–log10 adjusted p-value).
2.2. Drug Candidates for Repositioning in HPV Clade-Specific Cervical Cancer
Reversal-based transcriptomic approaches have been extensively employed to identify compounds capable of counteracting disease-associated gene expression programs [32,41]. Following this rationale, we calculated sRGES values for drugs in HPV-driven cervical cancer, thereby generating a comprehensive landscape of potential repositioning opportunities across both α-A7 and α-A9 clades. For the α-A7 signature, 5,569 compounds were evaluated, of which 931 corresponded to FDA-approved drugs. In the case of α-A9, we obtained 4,014 compounds that were tested, including 693 FDA-approved.
To further contextualize these results, we performed enrichment analysis of the FDA-approved drugs (Supplementary Figure S1). Using MeSH categories (Medical Subject Headings), we observed significant enrichment for pharmacological classes such as histone deacetylase inhibitors, selective estrogen receptor modulators, estrogen antagonists, and HMG-CoA reductase inhibitors, reflecting both well-established anticancer agents and broader therapeutic categories. In parallel, ChEMBL target-based annotations highlighted recurrent involvement of molecular families including HDAC isoforms, ABC transporters, and nuclear hormone receptors. Notably, these enrichments were supported by multiple independent drugs converging on the same categories or targets, thereby reinforcing the biological plausibility and therapeutic relevance of the predicted candidates.
We next focused on the most significant candidates, defined as drugs with sRGES ≤ −0.20, and we identified 63 compounds that represent the strongest predicted reversal of the clade-specific transcriptional signatures (Figure 3). Of these, 31 were shared across α-A7 and α-A9; however, their sRGES values differed between clades, indicating variation in the predicted magnitude of reversal by context. By contrast, the remaining 32 appeared in only one clade, thereby delineating clade-restricted sets. Within the FDA-approved subset, 10 agents met the threshold for α-A7 and 21 for α-A9. Taken together, these results reveal a landscape that includes broadly acting options as well as group-specific candidates, supporting study designs that test pan-group repositioning alongside strategies tailored to α-A7 or α-A9 tumors.
Heatmap of FDA approved drug candidates with significant reversal potential (sRGES ≤ −0.20) across α-A7 and α-A9 cervical cancer signatures. Rows correspond to clades and columns to individual drugs, ordered by consensus sRGES. The color scale represents sRGES (darker blue = stronger predicted reversal). The annotation bar at the top indicates compounds common to both clades (green), unique to α-A7 (orange), or unique to α-A9 (purple).
2.3. Validation of Predicted Drug Response
Following the OCTAD workflow, we evaluated whether more negative sRGES scores translate into better pharmacologic response in the most relevant cell lines (Figure 4). Interestingly, we observed consistent and directionally concordant associations for both clades. In HPV-A7, sRGES showed a positive correlation with the logarithm of the median half maximal inhibitory concentration (log10[median IC50]) and a negative correlation with the area under the drug response curve (AUC). Here, IC50 represents the concentration of a drug required to reduce cell viability by 50%, with lower values indicating greater potency. On the other hand, the recomputed AUC, as defined in PharmacoGx, reflects the integral of the drug response across doses, calculated as the area of (1–viability); in this framework, higher AUC values indicate stronger overall sensitivity (Supplementary Figure S2). Thus, our results show that stronger transcriptomic reversal, captured by more negative sRGES values, is associated simultaneously with lower IC50 and higher AUC. The same trend was observed in HPV-A9, supporting that negative sRGES values consistently track with increased pharmacologic sensitivity across clades. Together, the concordance between sRGES and drug response strengthens the evidence that our top candidates provide a robust foundation for experimental validation and clade-aware drug repositioning.
(A) HPV-A7; (B) HPV-A9. Scatterplots show the relationship between sRGES (x-axis) and log10(median IC50) (left) or AUC (recomputed) (right) after collapsing to a single point per drug (median across tested lines). Each point is a candidate predicted drug; the black line is a least-squares fit with 95% confidence band. The in-panel label reports the Pearson correlation (r) and p-value computed on the collapsed data. Points are colored by whether the drug was Strongly predicted (sRGES ≤ −0.20) and shaped by FDA status (triangle = FDA-launched; circle = other).
3. Discussion
Cervical cancer is a malignancy shaped by both intrinsic host variables and HPV features [42]. The association between high-risk HPV and cervical cancer has been studied extensively, from the initial viral infection of host cells to long term in the maintenance of malignancy [7]. Increasing evidence indicates that phylogenetic clades of high-risk HPV contribute distinctive biological features to tumors [10,37]. Our analysis of clade-stratified differential expression signatures shows that both α-A7 and α-A9 tumors converge on a broad malignant core, highlighting that fundamental pathways of tumorigenesis remain conserved regardless of viral lineage [43,44]. However, we also identified that tumors associated with high risk HPV clades differ in their transcriptional expression. These differences that we observed are expected, because several multi-omics studies have described how HPV-clade specifically drives distinct regulatory programs through epigenetic mechanisms like variation in DNA methylation, as well as histone marks [10]. Such patterns are consistent with the known biology of HPV persistence and integration: HPV18 (α-A7) is almost invariably integrated into host DNA, whereas HPV16 (α-A9) shows a comparatively lower rate of integration, with a fraction of viral genomes persisting episomally [37,45]. Also breakpoints tend to cluster at common fragile sites and active regulatory regions, occasionally creating domains similar to super enhancers that increase the expression of nearby oncogenes and remodel transcriptional programs [46,47]. Building on this mechanistic context, large multi-cohort analyses have defined intrinsic CC subtypes [48,49]. And also it could be a systematic stratification for precision therapy, however deriving these subtypes typically requires multi-omic profiling and sophisticated classifiers that limit immediate clinical use. By contrast, HPV genotyping is already embedded in diagnostic workflows and biologically informative [14], and it may serve as a practical axis for precision treatment decisions in future studies.
By constructing clade resolved signatures and using the OCTAD framework, we derived a sRGES score that identified a concise set of FDA approved agents with strong predicted activity. The enrichment of pharmacologic classes with a priori relevance most notably the . Histone deacetylases (HDACs) inhibitors such as panobinostat, vorinostat and belinostat appeared prominently across clades, which is mechanistically coherent in HPV-driven disease. In CC context, HDAC modulates chromatin structure and transcription, also their overactivity contributes to silencing tumor suppressors expression and can contribute with viral oncoprotein functions [50]. Accordingly, HDAC blockade can restore proapoptotic and cell cycle restraining programs, and has demonstrated antiproliferative activity in CC models [51,52], with combination strategies under early clinical investigation [53]. In this light, our results reinforce that epigenetic targeting is relevant across both α-7 and α-9 tumors.
We also identified hormone-modulating agents, selective estrogen receptor modulators (SERMs) and antagonists, such as fulvestrant, raloxifene and toremifene. These align with strong evidence that estrogen signaling drives HPV related CC.In HPV transgenic mouse models, both the pure ER antagonist fulvestrant and the SERM raloxifene produced dramatic tumor regressions, with one-month treatment clearing established cervical cancers and dysplasias [54]. In vitro, raloxifene blocked estradiol induced proliferation and E6/E7 transcription in cervical cells [55]. These data imply that repurposing antiestrogens for HPV positive CC is biologically plausible.
On the metabolic front, HMG-CoA reductase inhibitors such as simvastatin and pitavastatin also emerged as promising candidates. These lipophilic statins disrupt the mevalonate pathway, reducing prenylation of small GTPases and thereby attenuating oncogenic RAS–MAPK and PI3K–AKT signaling [56]. In CC models, simvastatin has been shown to suppress proliferation, induce apoptosis, and enhance the effects of cytotoxic chemotherapy through broad inhibition of prenylation dependent pathways [57]. Complementarily, pitavastatin induces oxidative stress–mediated DNA damage that activates caspase-9 and caspase-3, leading to PARP cleavage and apoptosis [58,59]. Together, these mechanistic insights and our findings highlight metabolic rewiring as a shared vulnerability in cervical tumors, supporting systematic testing of statins in combination with established agents.
Interestingly, several antimicrobial and antiparasitics agents emerged as notable candidates, Among anthelmintics, pyrvinium showed some of the strongest predicted activity, acting as a Wnt/β-catenin inhibitor that induces apoptosis and potentiates cisplatin in cervical models [60], and is already under early clinical evaluation in pancreatic cancer [61]. In the same class, closantel demonstrated anti-angiogenic effects and suppression of tumor growth in zebrafish xenografts implanted with CC cells [62]. Moving to antibiotic derivatives, monensin, also targeting Wnt/β-catenin signaling, reduced tumor growth in vivo with low toxicity [63], while nifuroxazide, a nitrofuran compound, inhibited STAT3 activity in cervical cells, promoting apoptosis and limiting metastatic signaling [64]. By contrast, ivermectin, despite wide interest, only achieved antiproliferative effects in cervical models at supraphysiologic concentrations, limiting its translational feasibility [65]. Collectively, these findings indicate that while antiparasitic and antimicrobial compounds are attractive for their cost and oral availability, their promise in cervical cancer requires cautious interpretation and rigorous clade-aware preclinical validation before advancing toward clinical application.
To complement repositioning candidates, several agents recovered by our screen overlap with conventional chemotherapies. DNA-targeting drugs such as methotrexate, 6-mercaptopurine, teniposide, etoposide, and SN-38 (the active metabolite of irinotecan) were recovered, reflecting their broad cytotoxicity rather than novel repositioning. Indeed, irinotecan has shown activity in cervical cancer when combined with cisplatin, with Phase II studies reporting overall response rates close to 60% in advanced or recurrent disease [66,67]. Etoposide in combination with cisplatin has similarly provided moderate but consistent benefit in recurrent settings [68]. Importantly, our analysis also retrieved bortezomib, whose rationale is especially relevant in HPV-driven cancers: by inhibiting the proteasome, bortezomib stabilizes p53 and PDZ-domain tumor suppressors such as hDlg and hScrib, counteracting key oncogenic effects of HPV oncoproteins. In preclinical HeLa xenografts models, bortezomib combined with cisplatin produced synergistic tumor regression [69].
Our screen further suggests palbociclib, an inhibitor of CDK4/6, due to its capacity to reduce proliferation and trigger apoptosis via oxidative stress in CC models [70]. Combined with SHetA2, it synergistically suppresses phosphorylated Rb and tumor growth in SiHa xenografts[71]. In parallel, the mTOR inhibitor temsirolimus achieved mostly disease stabilization but few objective responses in a phase II trial of recurrent or metastatic cervical carcinoma, reflecting modest single-agent efficacy and highlighting the need for biomarker-guided or combinatorial approaches [72].
Moreover, phenothiazines (antipsychotic agents), impact multiple signaling axes, such as calmodulin, PI3K/AKT/mTOR and autophagy. In cervical models, chlorpromazine induces time dependent cytomorphologic disruption and detachment in human CC cells [73]. Overall, phenothiazines remain mechanistically interesting but dose intensity and central nervous system adverse effects raise translational constraints.
Taken together, these additional drug classes expand the therapeutic landscape beyond our primary leads. Their inclusion highlights that vulnerabilities in cervical cancer span epigenetic, metabolic, hormonal, signaling, proteostatic, and immunomodulatory axes. Integration into clade-aware combination regimens for example, HDAC inhibitors with statins, ER antagonists with proteasome inhibitors, or CDK4/6 blockade with mTOR/STAT3 inhibition, could enhance tumor control by exploiting complementary mechanisms. Importantly, the majority of the predicted candidates converge on pathways known to be hijacked by HPV oncogenes, including cell-cycle dysregulation, epigenetic silencing, estrogen receptor signaling, Wnt/β-catenin activity, autophagy, and proteasome-mediated degradation [44]. Several compounds have direct preclinical or clinical support in cervical systems, while others such as antiparasitics and antimicrobials remain attractive but demand more rigorous validation. Overall, our results provide a coherent set of repositioning candidates supported by varying degrees of prior anticancer evidence, but best viewed as hypothesis generating rather than definitive treatment rules.
Signature-based repurposing pipelines have repeatedly shown that transcriptional inversion predicts antitumor activity across malignancies, progressing in some cases from in silico prediction to bench validation and early clinical exploration [29,32]. In cervical cancer, most prior efforts have generated large drug catalogs without stratifying by viral genotype. By contrast, our approach deploys an OCTAD workflow tuned to HPV clades, yielding a concise and biologically plausible list of FDA-approved agents that align with clade-specific transcriptional signatures and are reinforced by cell-line sensitivity correlations. Nevertheless, several limitations temper interpretation: bulk RNA-seq signatures conflate epithelial and microenvironmental signals; HPV genotype, histology, and integration status are partially collinear; and reversal scoring favors pleiotropic transcriptional modulators. Future studies incorporating single-cell analyses, carefully matched experimental systems, and orthogonal validation strategies will be required to disentangle these axes and refine therapeutic hypotheses.
4. Materials and Methods
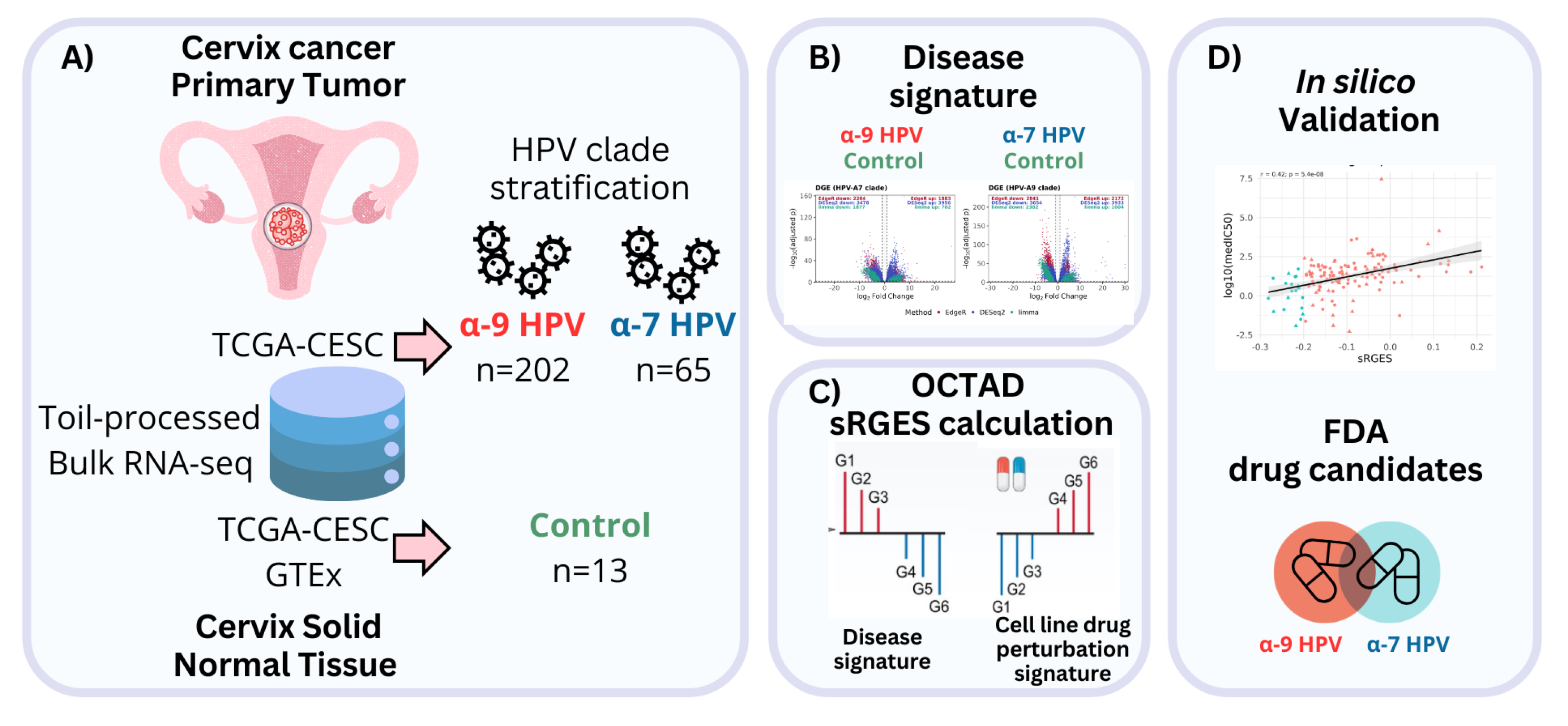
A) Bulk RNA-seq data uniformly processed by Toil were obtained from TCGA-CESC tumors and GTEx/TCGA normal cervix samples. HPV genotypes were assigned with HPV-EM, and only mono-infected cases were retained. Tumor samples were stratified into α-A9 (n = 202) and α-A7 (n = 65) clades, with controls from normal cervix tissues (n = 13). B) Differential expression analysis was performed separately for each clade versus controls using edgeR, DESeq2, and limma-voom with RUVSeq correction. Consensus DEG signatures were functionally interpreted by GO over-representation analysis. C) Drug predictions were derived through OCTAD by calculating summarized Reversal Gene Expression Scores (sRGES), integrating tumor–cell line similarity and LINCS L1000 perturbation profiles, and filtered for FDA-approved compounds with sRGES ≤ −0.20. D) Finally, predictions were validated in silico against pharmacogenomic sensitivity data (AUC and IC₅₀) from cancer relevant cell lines.
4.1. Data and Cohort Definition
All analyses were conducted in R (v4.5.1) using octad (v1.9.0) and octad.db (v1.10.0) packages [36,74]. RNA-seq expression profiles were obtained from the OCTAD platform, which integrates uniformly Toil-processed RNA-seq data (HDF5 format) from TCGA and GTEx and the associated sample metadata (dataset EH7274), allowing for reliable cross-cohort comparisons (Figure 5). From the TCGA-CESC project, we selected 267 cervical carcinoma samples. HPV genotypes were assigned according to the annotations reported with the HPV-EM algorithm [38], and only mono-infected cases with high-risk types were considered. Therefore, samples were stratified into two phylogenetic clades: α-A7 (n = 66; HPV 18, 39, 45, 59) and α-A9 (n = 202; HPV 16, 31, 33, 35, 52, 58) [9]. The control group consisted of 13 normal cervical tissue samples, including 10 from GTEx and 3 adjacent normals from TCGA.
4.2. Differential Expression Analysis
Differential gene expression between tumors and controls was assessed separately for each clade group (α-A7 and α-A9) using three methods: edgeR, DESeq2, and limma-voom. In order to mitigate unwanted variation, RUVSeq was applied with k = 1 for edgeR and DESeq2, following OCTAD’s default pipeline. Genes were considered significantly differentially expressed when |log₂FC| ≥ 1 and FDR ≤ 0.01, based on the adjusted p-values returned by each method.
4.3. Over-Representation Analysis
To explore the functional relevance of the consensus DEG signatures, we performed Over-Representation Analysis (ORA) using the clusterProfiler (v4.16.0) R package [75] . For each clade (α-A7 and α-A9), up- and downregulated genes were analyzed independently across the three gene ontology (GO) domains: biological process (BP), cellular component (CC), and molecular function (MF). ORA was conducted to capture all significant categories (p ≤ 0.05, q ≤ 0.20, BH-adjusted), and subsequently refined by collapsing redundant terms based on semantic similarity (cutoff = 0.25), yielding a simplified set of representative GO categories for each clade and regulation group.
4.4. sRGES Calculation
Candidate drugs were prioritized using summarized Reversal Gene Expression Scores (sRGES) implemented in the OCTAD framework [1]. This metric integrates differential expression (DE) signatures with cohort–cell line similarity weights to estimate the ability of a compound to reverse disease associated transcriptional profiles. Similarity between tumor cohorts (α-A7 and α-A9) and cancer cell lines was first assessed with the function computeCellLine using the octad.small reference set; only lines with a median correlation (medcor) > 0.30 were retained. Clade-specific DE signatures were then analyzed with the function runRGES, which assigns each compound an individual RGES defined as the difference between enrichment scores of up- and downregulated disease genes within LINCS L1000 perturbation profiles. The resulting scores were adjusted by the retained cell-line weights and summarized into an sRGES per drug [32].
Within each group of samples, only drugs consistently identified across the three DE methods were considered with a consensus sRGES calculated as the arithmetic mean of the method-specific values. Analyses were then restricted to FDA-approved compounds, and the final lists were filtered to retain only candidates with consensus sRGES ≤ −0.20. All selected cell lines and clade-specific predictions were archived for reproducibility (Supplementary Table S1).
4.5. Drug Set Enrichment Analysis
To further contextualize the drug predictions, we carried out enrichment analysis of the consensus FDA-approved sRGES lists using octadDrugEnrichment function. Two complementary classifications were examined: (i) MeSH (Medical Subject Headings), which organizes compounds by pharmacological action or therapeutic application and (ii) ChEMBL targets, which group drugs according to annotated protein targets or families. Enrichment significance was evaluated at FDR ≤ 0.01 and for each enriched set, we additionally reported the number of predicted compounds contributing to the signal.
4.6. Validation of Predicted Drug Response
To evaluate the robustness of the predictions, we validated the consensus sRGES values against pharmacogenomic response data from cancer cell lines. For each group of samples (clade α-A7 and α-A9), we applied the OCTAD function topLineEval, which compares drug-specific sRGES scores with experimental sensitivity profiles available for the corresponding cell lines. The function returns two complementary measures for all overlapping compounds: (i) the area under the drug response curve (AUC), as recomputed in PharmacoGx from raw viability data and defined as the integral of (1–viability) across the tested dose range (with higher AUC values indicating greater sensitivity), and (ii) the median inhibitory concentration (IC₅₀), reported after log₁₀ transformation, which represents the concentration required to reduce viability by 50% (lower IC₅₀ reflecting higher potency). In addition, the function calculates Pearson’s correlation coefficients (r) together with two-sided P-values, thereby providing a direct statistical assessment of the association between predicted reversal scores and observed drug sensitivities.
5. Conclusions
In summary, leveraging CC HPV clade resolved transcriptional programs with the OCTAD platform yielded prioritized sets of 41 and 52 FDA approved candidates with strong predicted activity for HPV-α7 and HPV-α9 tumors, respectively, enriched for mechanistically coherent drug classes and supported by correlations with experimental sensitivity. These findings argue that both shared and clade-specific vulnerabilities exist in HPV-driven cervical cancer and that expression-guided repurposing can surface actionable therapeutic hypotheses. While confirmation in appropriate experimental systems is indispensable, this work ultimately provides a repertoire of plausible repositioning agents and should be regarded primarily as a hypothesis generating resource for future preclinical and translational research.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org.
Author Contributions
Conceptualization—J.R.-H., G.d.A.-J. and E.H.-L.; methodology—J.R.-H., G.d.A.-J. and E.H.-L.; validation—J.R.-H. and E.H.-L.; formal analysis—J.R.-H.; investigation—J.R.-H., and E.H.-L.; resources—E.H.-L.; data curation—J.R.-H.; writing—original draft preparation—J.R.-H.; writing—review and editing—J.R.-H., G.d.A.-J. and E.H.-L. All authors have read and agreed to the published version of the manuscript.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. J.R.-H. is a doctoral student from Programa de Maestria y Doctorado en Ciencias Bioquimicas (PMDCB), Universidad Nacional Autónoma de México (UNAM), and received fellowship CVU 1100839 from SECIHTI.
Informed Consent Statement
Informed consent was received from all participants or their representatives.
Data Availability Statement
All code involved in this work is publicly available at https://github.com/JoelRuiz26/CESC_Network/tree/bfbec389f3e206aac35260cd7a1548334ae27b7a/6_OCTAD
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- International Agency for Research on Cancer Global Cancer Observatory: Cancer Today. Available online: https://gco.iarc.who.int/today/ (accessed on 24 September 2025).
- Brisson, M.; Kim, J.J.; Canfell, K.; Drolet, M.; Gingras, G.; Burger, E.A.; Martin, D.; Simms, K.T.; Bénard, É.; Boily, M.-C.; et al. Impact of HPV Vaccination and Cervical Screening on Cervical Cancer Elimination: A Comparative Modelling Analysis in 78 Low-Income and Lower-Middle-Income Countries. Lancet Lond. Engl. 2020, 395, 575–590. [Google Scholar] [CrossRef]
- Chan, C.K.; Aimagambetova, G.; Ukybassova, T.; Kongrtay, K.; Azizan, A. Human Papillomavirus Infection and Cervical Cancer: Epidemiology, Screening, and Vaccination-Review of Current Perspectives. J. Oncol. 2019, 2019, 3257939. [Google Scholar] [CrossRef]
- Vallejo-Ruiz, V.; Gutiérrez-Xicotencatl, L.; Medina-Contreras, O.; Lizano, M. Molecular Aspects of Cervical Cancer: A Pathogenesis Update. Front. Oncol. 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Castro-Oropeza, R.; Piña-Sánchez, P. Epigenetic and Transcriptomic Regulation Landscape in HPV+ Cancers: Biological and Clinical Implications. Front. Genet. 2022, 13, 886613. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhou, J.; Wang, F.; Shi, H.; Li, Y.; Li, B. HPV-16 E6/E7 Promotes Cell Migration and Invasion in Cervical Cancer via Regulating Cadherin Switch in Vitro and in Vivo. Arch. Gynecol. Obstet. 2015, 292, 1345–1354. [Google Scholar] [CrossRef] [PubMed]
- Wilting, S.M.; Steenbergen, R.D.M. Molecular Events Leading to HPV-Induced High Grade Neoplasia. Papillomavirus Res. 2016, 2, 85–88. [Google Scholar] [CrossRef]
- Muñoz-Bello, J.O.; Carrillo-García, A.; Lizano, M. Epidemiology and Molecular Biology of HPV Variants in Cervical Cancer: The State of the Art in Mexico. Int. J. Mol. Sci. 2022, 23, 8566. [Google Scholar] [CrossRef]
- Rader, J.S.; Tsaih, S.; Fullin, D.; Murray, M.W.; Iden, M.; Zimmermann, M.T.; Flister, M.J. Genetic Variations in Human Papillomavirus and Cervical Cancer Outcomes. Int. J. Cancer 2019, 144, 2206–2214. [Google Scholar] [CrossRef]
- Gagliardi, A.; Porter, V.L.; Zong, Z.; Bowlby, R.; Titmuss, E.; Namirembe, C.; Griner, N.B.; Petrello, H.; Bowen, J.; Chan, S.K.; et al. Analysis of Ugandan Cervical Carcinomas Identifies Human Papillomavirus Clade–Specific Epigenome and Transcriptome Landscapes. Nat. Genet. 2020, 52, 800–810. [Google Scholar] [CrossRef]
- Nobeyama, H.; Sumi, T.; Misugi, F.; Okamoto, E.; Hattori, K.; Matsumoto, Y.; Yasui, T.; Honda, K.-I.; Iwai, K.; Ishiko, O. Association of HPV Infection with Prognosis after Neoadjuvant Chemotherapy in Advanced Uterine Cervical Cancer. Int. J. Mol. Med. 2004, 14, 101–105. [Google Scholar] [CrossRef]
- Ferdousi, J.; Nagai, Y.; Asato, T.; Hirakawa, M.; Inamine, M.; Kudaka, W.; Kariya, K.-I.; Aoki, Y. Impact of Human Papillomavirus Genotype on Response to Treatment and Survival in Patients Receiving Radiotherapy for Squamous Cell Carcinoma of the Cervix. Exp. Ther. Med. 2010, 1, 525–530. [Google Scholar] [CrossRef]
- Wang, C.-C.; Lai, C.-H.; Huang, H.-J.; Chao, A.; Chang, C.-J.; Chang, T.-C.; Chou, H.-H.; Hong, J.-H. Clinical Effect of Human Papillomavirus Genotypes in Patients with Cervical Cancer Undergoing Primary Radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, 1111–1120. [Google Scholar] [CrossRef]
- Comprehensive Cervical Cancer Control: A Guide to Essential Practice; World Health Organization, World Health Organization, Eds. ; Second edition.; World Health Organization: Geneva, 2014; ISBN 978-92-4-154895-3. [Google Scholar]
- Li, H.; Wu, X.; Cheng, X. Advances in Diagnosis and Treatment of Metastatic Cervical Cancer. J. Gynecol. Oncol. 2016, 27, e43. [Google Scholar] [CrossRef]
- Fernandes, A.; Viveros-Carreño, D.; Hoegl, J.; Ávila, M.; Pareja, R. Human Papillomavirus-Independent Cervical Cancer. Int. J. Gynecol. Cancer 2022, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, C.A.; Khan, S.F.; Schäfer, G.; Mbatani, N.; Adams, T.; Moodley, J.; Prince, S. Cervical Cancer Therapies: Current Challenges and Future Perspectives. Tumour Virus Res. 2022, 13, 200238. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Luo, H.; Zhang, W.; Shen, Z.; Hu, X.; Zhu, X. Molecular Mechanisms of Cisplatin Resistance in Cervical Cancer. Drug Des. Devel. Ther. 2016, 10, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, M.; Li, X.; Yin, S.; Wang, B. An Overview of Novel Agents for Cervical Cancer Treatment by Inducing Apoptosis: Emerging Drugs Ongoing Clinical Trials and Preclinical Studies. Front. Med. 2021, 8. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, L.; Xie, N.; Nice, E.C.; Zhang, T.; Cui, Y.; Huang, C. Overcoming Cancer Therapeutic Bottleneck by Drug Repurposing. Signal Transduct. Target. Ther. 2020, 5, 113. [Google Scholar] [CrossRef]
- Langedijk, J.; Mantel-Teeuwisse, A.K.; Slijkerman, D.S.; Schutjens, M.-H.D.B. Drug Repositioning and Repurposing: Terminology and Definitions in Literature. Drug Discov. Today 2015, 20, 1027–1034. [Google Scholar] [CrossRef]
- Weth, F.R.; Hoggarth, G.B.; Weth, A.F.; Paterson, E.; White, M.P.J.; Tan, S.T.; Peng, L.; Gray, C. Unlocking Hidden Potential: Advancements, Approaches, and Obstacles in Repurposing Drugs for Cancer Therapy. Br. J. Cancer 2024, 130, 703–715. [Google Scholar] [CrossRef]
- Park, S.-H.; Kim, M.; Lee, S.; Jung, W.; Kim, B. Therapeutic Potential of Natural Products in Treatment of Cervical Cancer: A Review. Nutrients 2021, 13, 154. [Google Scholar] [CrossRef]
- Capistrano I, R.; Paul, S.; Boere, I.; Pantziarka, P.; Chopra, S.; Nout, R.A.; Bouche, G. Drug Repurposing as a Potential Source of Innovative Therapies in Cervical Cancer. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2022, 32, 1377–1386. [Google Scholar] [CrossRef] [PubMed]
- de Anda-Jáuregui, G.; Guo, K.; McGregor, B.A.; Hur, J. Exploration of the Anti-Inflammatory Drug Space Through Network Pharmacology: Applications for Drug Repurposing. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- de Anda-Jáuregui, G.; Guo, K.; Hur, J. Network-Based Assessment of Adverse Drug Reaction Risk in Polypharmacy Using High-Throughput Screening Data. Int. J. Mol. Sci. 2019, 20, 386. [Google Scholar] [CrossRef] [PubMed]
- Coria-Rodríguez, H.; Ochoa, S.; de Anda-Jáuregui, G.; Hernández-Lemus, E. Drug Repurposing for Basal Breast Cancer Subpopulations Using Modular Network Signatures. Comput. Biol. Chem. 2023, 105, 107902. [Google Scholar] [CrossRef]
- Lamb, J.; Crawford, E.D.; Peck, D.; Modell, J.W.; Blat, I.C.; Wrobel, M.J.; Lerner, J.; Brunet, J.-P.; Subramanian, A.; Ross, K.N.; et al. The Connectivity Map: Using Gene-Expression Signatures to Connect Small Molecules, Genes, and Disease. Science 2006, 313, 1929–1935. [Google Scholar] [CrossRef]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e17. [Google Scholar] [CrossRef]
- Mejía-Pedroza, R.A.; Espinal-Enríquez, J.; Hernández-Lemus, E. Pathway-Based Drug Repositioning for Breast Cancer Molecular Subtypes. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Hernández-Lemus, E.; Martínez-García, M. Pathway-Based Drug-Repurposing Schemes in Cancer: The Role of Translational Bioinformatics. Front. Oncol. 2021, 10. [Google Scholar] [CrossRef]
- Chen, B.; Ma, L.; Paik, H.; Sirota, M.; Wei, W.; Chua, M.-S.; So, S.; Butte, A.J. Reversal of Cancer Gene Expression Correlates with Drug Efficacy and Reveals Therapeutic Targets. Nat. Commun. 2017, 8, 16022. [Google Scholar] [CrossRef]
- Pessetto, Z.Y.; Chen, B.; Alturkmani, H.; Hyter, S.; Flynn, C.A.; Baltezor, M.; Ma, Y.; Rosenthal, H.G.; Neville, K.A.; Weir, S.J.; et al. In Silico and in Vitro Drug Screening Identifies New Therapeutic Approaches for Ewing Sarcoma. Oncotarget 2016, 8, 4079–4095. [Google Scholar] [CrossRef]
- Jahchan, N.S.; Dudley, J.T.; Mazur, P.K.; Flores, N.; Yang, D.; Palmerton, A.; Zmoos, A.-F.; Vaka, D.; Tran, K.Q.T.; Zhou, M.; et al. A Drug Repositioning Approach Identifies Tricyclic Antidepressants as Inhibitors of Small Cell Lung Cancer and Other Neuroendocrine Tumors. Cancer Discov. 2013, 3, 1364–1377. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, H.; Chen, M.; Wang, S.; Qian, R.; Zhang, L.; Huang, X.; Wang, J.; Liu, Z.; Qin, W.; et al. A Survey of Optimal Strategy for Signature-Based Drug Repositioning and an Application to Liver Cancer. eLife 2022, 11, e71880. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Glicksberg, B.S.; Newbury, P.; Chekalin, E.; Xing, J.; Liu, K.; Wen, A.; Chow, C.; Chen, B. OCTAD: An Open Workspace for Virtually Screening Therapeutics Targeting Precise Cancer Patient Groups Using Gene Expression Features. Nat. Protoc. 2021, 16, 728–753. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.D.; Chen, Z.; Saller, C.; Tarvin, K.; Carvalho, A.L.; Scapulatempo-Neto, C.; Silveira, H.C.; Fregnani, J.H.; Creighton, C.J.; Anderson, M.L.; et al. Integrated Genomic and Molecular Characterization of Cervical Cancer. Nature 2017, 543, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Inkman, M.J.; Jayachandran, K.; Ellis, T.M.; Ruiz, F.; McLellan, M.D.; Miller, C.A.; Wu, Y.; Ojesina, A.I.; Schwarz, J.K.; Zhang, J. HPV-EM: An Accurate HPV Detection and Genotyping EM Algorithm. Sci. Rep. 2020, 10, 14340. [Google Scholar] [CrossRef]
- Lim, J.; Straub, E.; Stubenrauch, F.; Iftner, T.; Schindler, M.; Simon, C. An Enhanced Triple Fluorescence Flow-Cytometry-Based Assay Shows Differential Activation of the Notch Signaling Pathway by Human Papillomavirus E6 Proteins. Sci. Rep. 2022, 12, 3000. [Google Scholar] [CrossRef]
- Yu, L.; Majerciak, V.; Zheng, Z.-M. HPV16 and HPV18 Genome Structure, Expression, and Post-Transcriptional Regulation. Int. J. Mol. Sci. 2022, 23, 4943. [Google Scholar] [CrossRef]
- Zhao, G.; Newbury, P.; Ishi, Y.; Chekalin, E.; Zeng, B.; Glicksberg, B.S.; Wen, A.; Paithankar, S.; Sasaki, T.; Suri, A.; et al. Reversal of Cancer Gene Expression Identifies Repurposed Drugs for Diffuse Intrinsic Pontine Glioma. Acta Neuropathol. Commun. 2022, 10, 150. [Google Scholar] [CrossRef]
- Alrefai, E.A.; Alhejaili, R.T.; Haddad, S.A. Human Papillomavirus and Its Association With Cervical Cancer: A Review. Cureus 2024, 16, e57432. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qiu, K.; Ren, J.; Zhao, Y.; Cheng, P. Roles of Human Papillomavirus in Cancers: Oncogenic Mechanisms and Clinical Use. Signal Transduct. Target. Ther. 2025, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Porter, V.L.; Ng, M.; O’Neill, K.; MacLennan, S.; Corbett, R.D.; Culibrk, L.; Hamadeh, Z.; Iden, M.; Schmidt, R.; Tsaih, S.-W.; et al. Rearrangements of Viral and Human Genomes at Human Papillomavirus Integration Events and Their Allele-Specific Impacts on Cancer Genome Regulation. Genome Res. 2025, 35, 653–670. [Google Scholar] [CrossRef] [PubMed]
- McBride, A.A.; Warburton, A. The Role of Integration in Oncogenic Progression of HPV-Associated Cancers. PLoS Pathog. 2017, 13, e1006211. [Google Scholar] [CrossRef]
- Løvestad, A.H.; Repesa, A.; Costanzi, J.-M.; Lagström, S.; Christiansen, I.K.; Rounge, T.B.; Ambur, O.H. Differences in Integration Frequencies and APOBEC3 Profiles of Five High-Risk HPV Types Adheres to Phylogeny. Tumour Virus Res. 2022, 14, 200247. [Google Scholar] [CrossRef]
- Chakravarthy, A.; Reddin, I.; Henderson, S.; Dong, C.; Kirkwood, N.; Jeyakumar, M.; Rodriguez, D.R.; Martinez, N.G.; McDermott, J.; Su, X.; et al. Integrated Analysis of Cervical Squamous Cell Carcinoma Cohorts from Three Continents Reveals Conserved Subtypes of Prognostic Significance. Nat. Commun. 2022, 13, 5818. [Google Scholar] [CrossRef]
- Zhu, X.; Li, S.; Luo, J.; Ying, X.; Li, Z.; Wang, Y.; Zhang, M.; Zhang, T.; Jiang, P.; Wang, X. Subtyping of Human Papillomavirus-Positive Cervical Cancers Based on the Expression Profiles of 50 Genes. Front. Immunol. 2022, 13, 801639. [Google Scholar] [CrossRef]
- Li, X.; Zhou, M.; Yu, J.; Yu, S.; Ruan, Z. Histone Modifications in Cervical Cancer: Epigenetic Mechanisms, Functions and Clinical Implications (Review). Oncol. Rep. 2025, 54, 131. [Google Scholar] [CrossRef]
- Dejligbjerg, M.; Grauslund, M.; Litman, T.; Collins, L.; Qian, X.; Jeffers, M.; Lichenstein, H.; Jensen, P.B.; Sehested, M. Differential Effects of Class I Isoform Histone Deacetylase Depletion and Enzymatic Inhibition by Belinostat or Valproic Acid in HeLa Cells. Mol. Cancer 2008, 7, 70. [Google Scholar] [CrossRef]
- Banerjee, N.S.; Moore, D.W.; Broker, T.R.; Chow, L.T. Vorinostat, a Pan-HDAC Inhibitor, Abrogates Productive HPV-18 DNA Amplification. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, E11138–E11147. [Google Scholar] [CrossRef]
- Borcoman, E.; Cabarrou, B.; Francisco, M.; Bigot, F.; Ghiringhelli, F.; Vansteene, D.; Legrand, F.; Halladjian, M.; Dupain, C.; Le Saux, O.; et al. Efficacy of Pembrolizumab and Vorinostat Combination in Patients with Recurrent and/or Metastatic Squamous Cell Carcinomas: A Phase 2 Basket Trial. Nat. Cancer 2025, 6, 1370–1383. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.-H.; Lambert, P.F. Prevention and Treatment of Cervical Cancer in Mice Using Estrogen Receptor Antagonists. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 19467–19472. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.-Q.; Wang, X.-H.; Tang, L.-P.; Chen, X.-W.; Lou, G. Raloxifene Suppress Proliferation-Promoting Function of Estrogen in CaSKi Cervical Cells. Int. J. Clin. Exp. Med. 2015, 8, 5571–5575. [Google Scholar] [PubMed]
- Buccioli, G.; Testa, C.; Jacchetti, E.; Pinoli, P.; Carelli, S.; Ceri, S.; Raimondi, M.T. The Molecular Basis of the Anticancer Effect of Statins. Sci. Rep. 2024, 14, 20298. [Google Scholar] [CrossRef]
- Pan, Q.; Xu, J.; Ma, L. Simvastatin Enhances Chemotherapy in Cervical Cancer via Inhibition of Multiple Prenylation-Dependent GTPases-Regulated Pathways. Fundam. Clin. Pharmacol. 2020, 34, 32–40. [Google Scholar] [CrossRef]
- Hacıseyitoğlu, A.Ö.; Doğan, T.Ç.; Dilsiz, S.A.; Canpınar, H.; Eken, A.; Bucurgat, Ü.Ü. Pitavastatin Induces Caspase-Mediated Apoptotic Death through Oxidative Stress and DNA Damage in Combined with Cisplatin in Human Cervical Cancer Cell Line. J. Appl. Toxicol. JAT 2024, 44, 623–640. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Wu, J.-X.; Yang, S.-F.; Wu, Y.-C.; Hsiao, Y.-H. Molecular Mechanisms Underlying the Anticancer Properties of Pitavastatin against Cervical Cancer Cells. Int. J. Mol. Sci. 2024, 25, 7915. [Google Scholar] [CrossRef]
- Xu, H.; Wang, Z.; Xu, L.; Mo, G.; Duan, G.; Wang, Y.; Sun, Z.; Chen, H. Targeting the eIF4E/β-Catenin Axis Sensitizes Cervical Carcinoma Squamous Cells to Chemotherapy. Am. J. Transl. Res. 2017, 9, 1203–1212. [Google Scholar]
- Ponzini, F.M.; Schultz, C.W.; Leiby, B.E.; Cannaday, S.; Yeo, T.; Posey, J.; Bowne, W.B.; Yeo, C.; Brody, J.R.; Lavu, H.; et al. Repurposing the FDA-Approved Anthelmintic Pyrvinium Pamoate for Pancreatic Cancer Treatment: Study Protocol for a Phase I Clinical Trial in Early-Stage Pancreatic Ductal Adenocarcinoma. BMJ Open 2023, 13, e073839. [Google Scholar] [CrossRef]
- Zhu, X.-Y.; Xia, B.; Liu, H.-C.; Xu, Y.-Q.; Huang, C.-J.; Gao, J.-M.; Dong, Q.-X.; Li, C.-Q. Closantel Suppresses Angiogenesis and Cancer Growth in Zebrafish Models. Assay Drug Dev. Technol. 2016, 14, 282–290. [Google Scholar] [CrossRef]
- Fu, B.; Fang, L.; Wang, R.; Zhang, X. Inhibition of Wnt/β-Catenin Signaling by Monensin in Cervical Cancer. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2024, 28, 21–30. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, Y. Long Non-Coding RNA MEG3 Inhibits Cervical Cancer Cell Growth by Promoting Degradation of P-STAT3 Protein via Ubiquitination. Cancer Cell Int. 2019, 19, 175. [Google Scholar] [CrossRef] [PubMed]
- Qabbus, M.B.; Hunt, K.S.; Dynka, J.; Woodworth, C.D.; Sur, S.; Samways, D.S.K. Ivermectin-Induced Cell Death of Cervical Cancer Cells in Vitro a Consequence of Precipitate Formation in Culture Media. Toxicol. Appl. Pharmacol. 2022, 449, 116073. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Yakushiji, M.; Noda, K.; Ikeda, M.; Kudoh, R.; Yajima, A.; Tomoda, Y.; Terashima, Y.; Takeuchi, S.; Hiura, M.; et al. Phase II Study of Irinotecan and Cisplatin as First-Line Chemotherapy in Advanced or Recurrent Cervical Cancer. Oncology 2000, 58, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Mabuchi, S.; Yokoi, E.; Shimura, K.; Komura, N.; Matsumoto, Y.; Sawada, K.; Isobe, A.; Tsutsui, T.; Kitada, F.; Kimura, T. A Phase II Study of Irinotecan Combined with S-1 in Patients with Advanced or Recurrent Cervical Cancer Previously Treated with Platinum Based Chemotherapy. Int. J. Gynecol. Cancer 2019, 29, 474–479. [Google Scholar] [CrossRef]
- Al-Saleh, E.; Hoskins, P.J.; Pike, J.A.; Swenerton, K.D. Cisplatin/Etoposide Chemotherapy for Recurrent or Primarily Advanced Cervical Carcinoma. Gynecol. Oncol. 1997, 64, 468–472. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Nakagawa, S.; Wada-Hiraike, O.; Seiki, T.; Tanikawa, M.; Hiraike, H.; Sone, K.; Nagasaka, K.; Oda, K.; Kawana, K.; et al. Sequential Effects of the Proteasome Inhibitor Bortezomib and Chemotherapeutic Agents in Uterine Cervical Cancer Cell Lines. Oncol. Rep. 2013, 29, 51–57. [Google Scholar] [CrossRef]
- Dam, P.V.; Wouters, A.; Dirix, L.; Rolfo, C. Abstract 2317: Palbociclib Monotherapy Exhibits Potent Activity in Cervical Cancer Cell Lines. Cancer Res. 2018, 78, 2317. [Google Scholar] [CrossRef]
- Wang, M.; Liu, J.; Liao, X.; Yi, Y.; Xue, Y.; Yang, L.; Cheng, H.; Liu, P. The SGK3-Catalase Antioxidant Signaling Axis Drives Cervical Cancer Growth and Therapy Resistance. Redox Biol. 2023, 67, 102931. [Google Scholar] [CrossRef]
- Tinker, A.V.; Ellard, S.; Welch, S.; Moens, F.; Allo, G.; Tsao, M.S.; Squire, J.; Tu, D.; Eisenhauer, E.A.; MacKay, H. Phase II Study of Temsirolimus (CCI-779) in Women with Recurrent, Unresectable, Locally Advanced or Metastatic Carcinoma of the Cervix. A Trial of the NCIC Clinical Trials Group (NCIC CTG IND 199). Gynecol. Oncol. 2013, 130, 269–274. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Zhang, B. Atomic Force Microscopy Study on Chlorpromazine-Induced Morphological Changes of Living HeLa Cells In Vitro. Scanning 2009, 31, 259–265. [Google Scholar] [CrossRef]
- Chekalin, E.; Paithankar, S.; Zeng, B.; Chen, B. Octad.Db: Open Cancer TherApeutic Discovery (OCTAD) Database.
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
Figure 1.
Differential expression signatures in HPV-associated cervical cancer by clade.
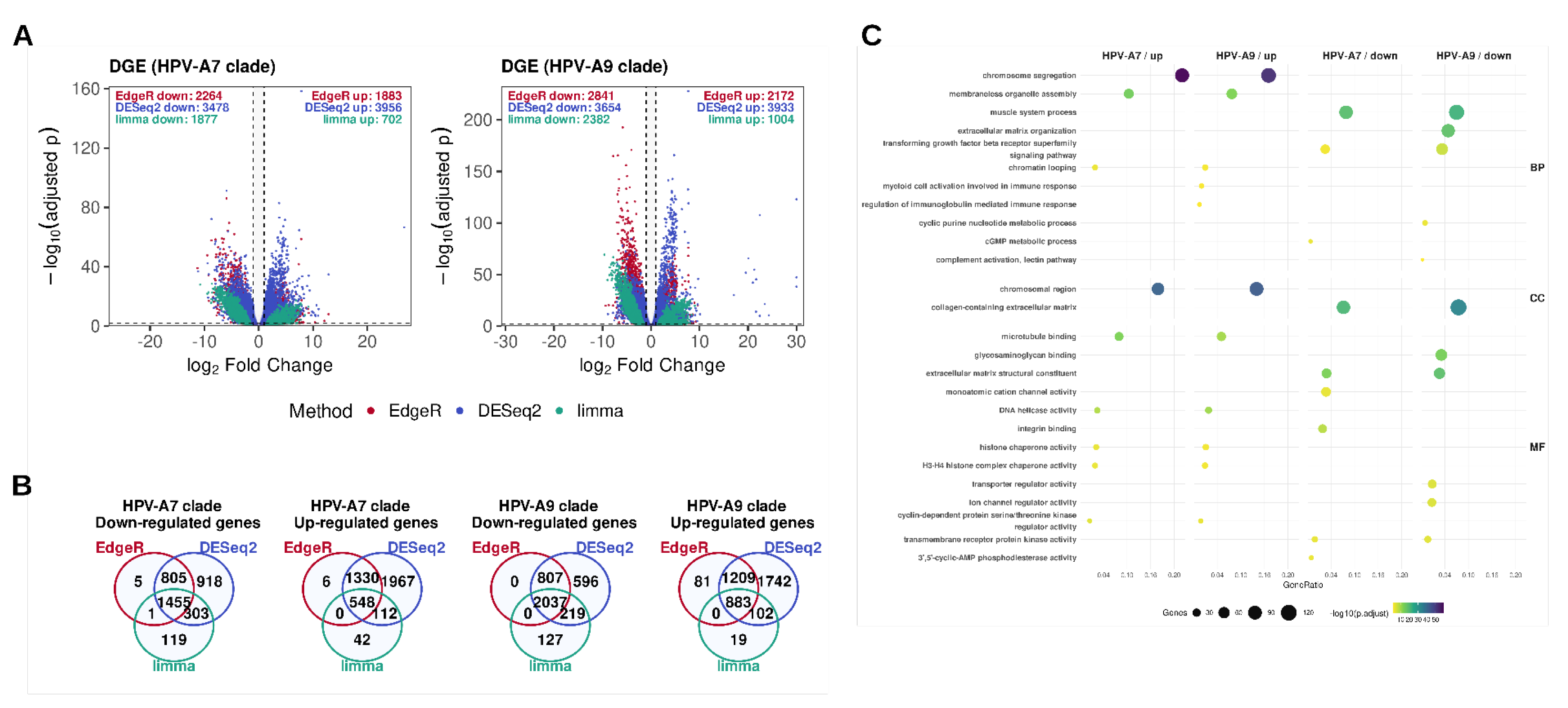
Figure 3.
Drug candidates for repositioning in α-A7 and α-A9 cervical carcinomas.
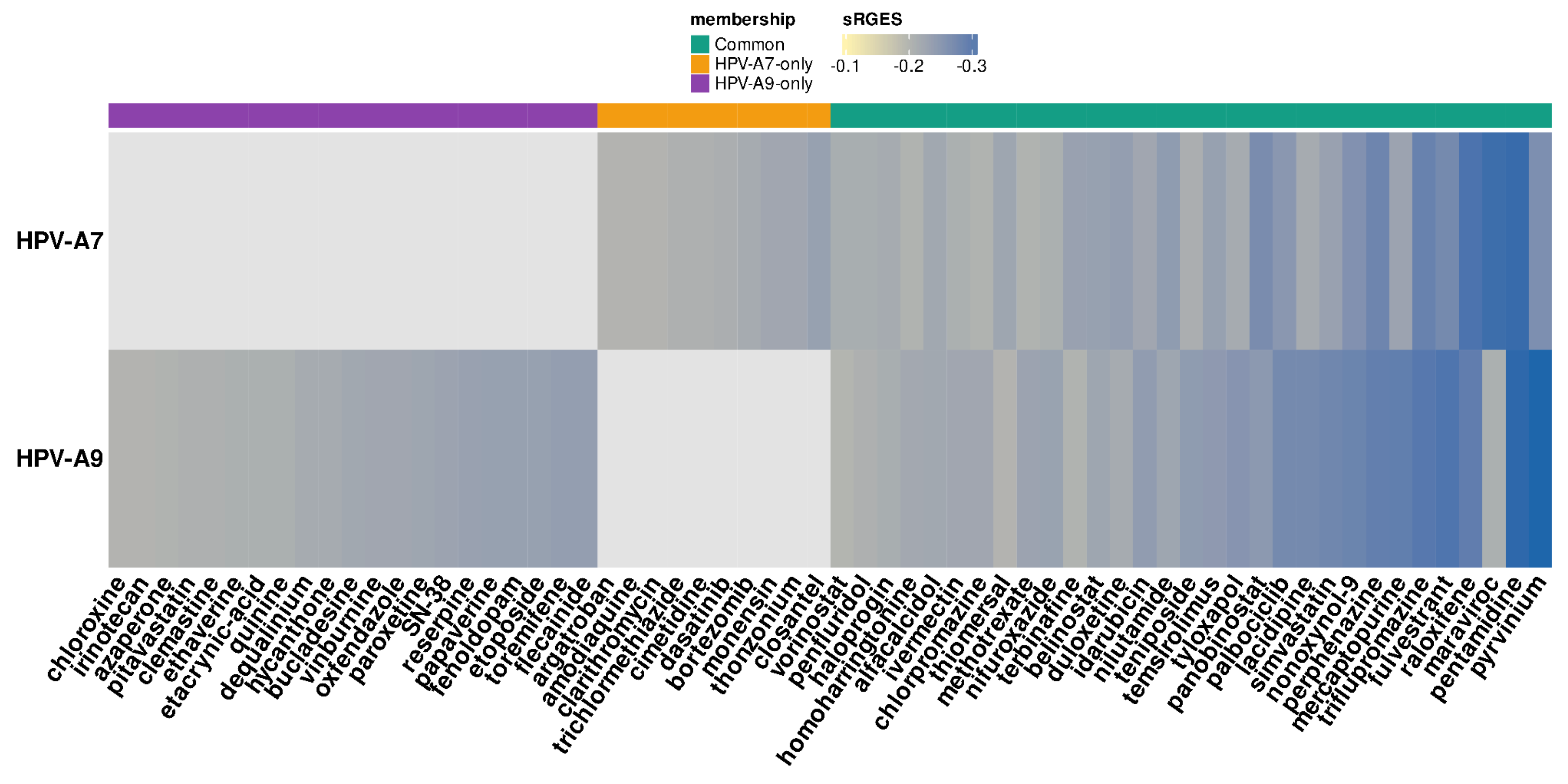
Figure 4.
Validation of sRGES with pharmacologic response.
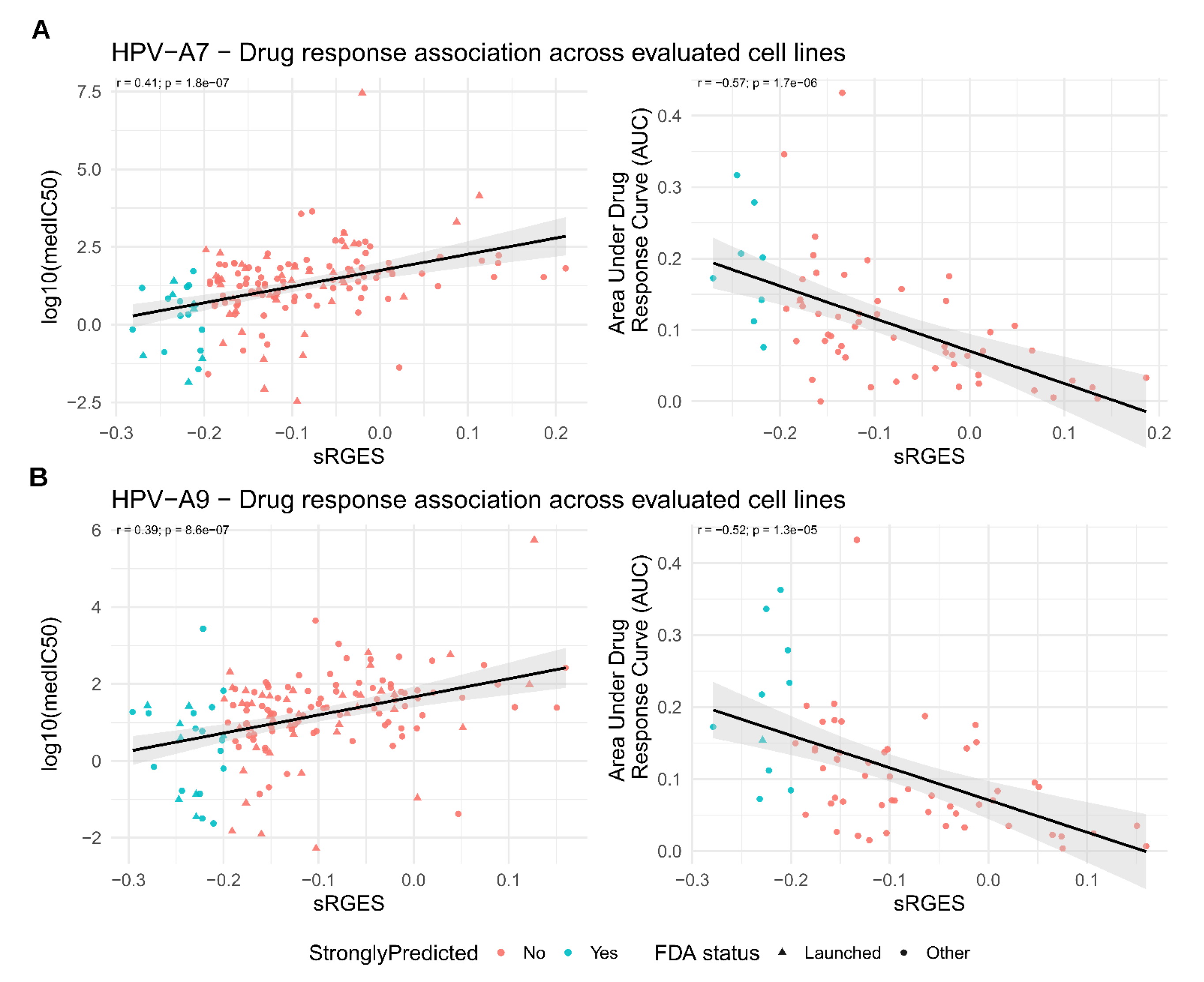
Figure 5.
Graphical overview of the study design and analytical workflow.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



