
Submitted:
01 October 2025
Posted:
02 October 2025
You are already at the latest version
Abstract
Phytochemicals are bioactive plant-derived compounds with significant antioxidant, anti-inflammatory, and epigenetic modulatory properties, offering promising avenues for the prevention and management of chronic diseases. Chronic conditions such as cancer, cardiovascular, neurodegenerative, metabolic, and autoimmune disorders often involve oxidative stress, inflammation, and epigenetic dysregulation. Evidence from in vitro and in vivo studies indicates that phytochemicals, including flavonoids, carotenoids, phenolic acids, organosulfur compounds, and terpenes, can attenuate oxidative damage by enhancing endogenous antioxidant defenses, modulate inflammatory signaling pathways such as NF-κB and Nrf2, and influence gene expression through the regulation of DNA methylation, histone modifications, and non-coding RNAs. Specific compounds—such as hesperidin, genistein, phloretin, lycopene, caffeic acid, p-coumaric acid, anacardic acid, allyl mercaptan, silybin, artemisinin, and geraniol—demonstrate multi-targeted actions, reversing aberrant epigenetic marks and restoring cellular homeostasis. By simultaneously acting on redox balance, inflammatory mediators, and epigenetic regulators, these natural compounds provide a mechanistic basis for chemoprevention and adjuvant therapy. However, translation into clinical applications requires rigorous trials to assess bioavailability, synergistic effects, and safety in humans. Understanding the molecular mechanisms of phytochemical action may advance novel, integrative strategies for chronic disease prevention and health promotion.
Keywords:
phytochemicals
; anti-oxidant activity
; epigenetic mechanisms
; anti-inflammatory activity
; chemoprevention
1. Introduction
Phytochemicals are plant secondary metabolites, produced to act as defense mole-cules against external pathogens and stress induced by environmental factors [1]. They have a great chemical diversity ranging from low molecular-weight compounds, with single aromatic-ring structure, as phenolic acids to large, complex molecules as proanthocyanidins and ellagitannins [2,3]. The natural occurring compounds possess antiviral, antifungal, and antibiotic properties, along with photoprotective function and participate in plant metabolism and survival process under biotic and abiotic stress [4].
For centuries, plant-derived treatments were used to heal and alleviate acute and chronic conditions including diabetes, depression, hypertension, hypercholesterolemia, insomnia and inflammation [5,6]. According to WHO, the primary health care of almost 70-80% of world population still depends on traditional medicines based mostly on medicinal plant extracts [7].
In the past decades scientific data illustrated that natural bioactive compounds found in fruit, vegetables and medicinal plants are biocompatible with cellular and molecular target sites and often induce less toxicity to normal cells, than chemical synthetic drugs [8,9]. Several preclinical studies have reported that many phytochemicals with anti-inflammatory, anti-oxidant and anti-proliferative properties can prevent cancer initiation and development by triggering apoptosis and activating antioxidant enzymes [10,11]. Moreover, the phytochemicals can impact cellular and molecular processes by acting as substrates, cofactors or inhibitors in biochemical and enzymatic reactions. They also function as ligands, either activating or blocking cell surface or intracellular receptors, scavengers of reactive oxygen species or other toxic chemicals, and enhancers of nutrient absorption and stability [11,12,13,14]. However, the chemopreventive potential of phytochemicals, despite promising preclinical research, has not been successfully translated into clinical practice.
Nowadays, there is mounting evidence indicating that many chronic conditions such as cancer, metabolic, neurodegenerative and autoimmune disorders are triggered by epi-genetic dysregulations [15]. Epigenetic alterations can occur due to exposure at various environmental factors, such as air pollution, chemical pollutants, electromagnetic radiation and life style (e.g. alcohol consumption, poor nutrition, smoking and lack of physical activity) [16]. Generally, epigenetic refers to heritable changes in gene expression that do not involve alteration to the DNA sequence itself [17]. These changes are primally mediated through specific mechanisms: DNA methylation (typically associated with gene silencing); post-translational modification of histone proteins (such as acetylation, methylation, phosphorylation, etc), non-coding RNAs, including microRNAs and long non-coding RNAs, which regulate gene expression post-transcriptionally [18,19].
Bioactive food components can trigger protective epigenetic changes throughout life, with early nutrition playing a particularly crucial role. These epigenetic modifications, influenced by diet and other environmental factors, can affect gene expression and potentially impact health outcomes, including the risk of chronic diseases, across a lifespan [20].
Early identification of epigenome abnormalities that may onset various pathologies and the possibility of eradicating them by using natural bioactive compounds from diet represent a new approach in prevention. The capacity of phytochemicals to modulate the epigenetic mechanisms, by reversing the aberrations related with onset and development of diverse diseases is intensively studied. All these factors can impact the epigenome and consequently might influence gene expression and biological processes
Therefore, a deeper understanding of the molecular mechanisms through which plant-derived compounds exert antioxidant and anti-inflammatory effects is needed to clarify their impact on human health. Further research into these mechanisms is crucial for developing new therapeutic strategies and promoting overall well-being.
This review aims to introduce the role of phytochemicals as natural bioactive compounds capable of modulating key molecular and epigenetic mechanism involved in the prevention and management chronic diseases. It highlights their antioxidant, anti-inflammatory, and gene-regulatory functions, emphasizing the potential of plant-derived compounds to influence human health by reversing or preventing pathological processes triggered by environmental and lifestyle-related factors. Furthermore, this review seeks to present current scientific evidence on chemopreventive properties of phytochemicals and to underscore the need for further research to translate these findings into effective clinical application.
2. Epigenetic Dysregulations in Chronic Diseases
In the past decades, many scientific reports have highlighted the essential role of the altered expression and function of epigenetic-regulatory proteins in the onset and development of various chronic diseases [21].
Epigenetic mechanisms are the main regulators of gene expression through chemical modifications of DNA, histones and RNA. For example, the transformation of a condensed chromatin region (heterochromatin), which is transcriptionally inactive into euchromatin as transcriptionally active structure, is achieved through the addition of methyl or acetyl groups to histone proteins. The addition of these chemical radicals along with gene’s promoter demethylation can loosen the chromatin structure, making DNA more accessible to the transcriptional machinery [22,23].
Enzymes that regulate epigenetic modifications are categorized into “writers,” “erasers,” “readers,” and “remodelers” based on their functions [24]. Writers are adding the specific chemical groups on DNA or RNA bases or histone’s amino acids, whereas erasers are acting in reverse by re-moving these modifications. For instance, DNA methyltransferase (DNMT) catalyzes the addition of methyl groups to form 5-methylcytosine (m5C) in gene’s promoter DNA sequence [25], whereas the ten-eleven translocation (TET) enzymes initiate DNA demethylation process by converting m5C into other derivatives, such as 5-hydroxymethylcytosine (5hmC), [26]. Typically, transcriptionally active genes exhibit a lower methylation pattern, whereas inactive genes or with a lower expression level are heavily methylated. Readers are proteins that recognize and bind to these chemical modifications, such as the methyl-CpG-binding domain (MBD) which is able to recognize 5mC [27]. The reader proteins are dynamically influencing chromatin status and by recruiting or collaborating with other protein complexes are regulating gene expression [28].
Remodelers such as SWI/SNF family, which possess DNA-stimulated ATPase activity are involved in nucleosome disruption with a crucial role in chromatin accessibility. They achieve this by disrupting nucleosomes and either moving or removing them from key regulatory regions like enhancers and promoters, thus influencing gene expression [29]. Furthermore, there are unique classes of epigenetic regulators, represented by non-coding RNAs (ncRNAs) which directly bind to various genomic regions or specific RNA sequences to modulate gene expression [30].
The long non-coding RNAs (lncRNAs) have a variety of functions and importantly they can interact with proteins, DNA sequences, RNAs regulating directly or indirectly the whole epigenetic machinery, by affecting gene expression and chromatin organization [31,32].
When epigenetic regulators, either epigenetic enzymes or ncRNAs expression, be-come imbalanced—by being excessively active or insufficiently functional—they disrupt cellular homeostasis and could induce the development of diseases [33]. Consequently, by investigating these individual epigenetic alterations such as DNA methylation, histone modifications and ncRNAs dysfunctional expression, represents a promising opportunity for chronic dis-ease therapy, fueling the creation of targeted treatments [34,35]. As an example, the abnormal epigenetic mechanisms are present at various stages of cancer development, including initiation, progression, invasion, migration, and chemotherapy resistance. DNA methylation patterns are changes with hypermethylation of specific tumor suppressors and hypomethylation of oncogenes promoters, which drives tumor development [36]. The silencing of tumor suppressor and DNA repair genes leads to the disruption of normal cell proliferation and differentiation and fostering the malignant phenotype of tumor cells [37]. Moreover, DNA methylation loss in oncogene promoter regions, and extensive demethylation of DNA repetitive sequences, compromises the chromatin stability, facilitating tumor development. [38].
For metabolic disorders the physiological conditions, the pre-existence of genetic mutations and also the exposure to environmental factors, such as dietary patterns can significantly influence the epigenome [39]. Several studies have shown that in mice feed with high-fat diet the hypermethylation of the promoters of genes like Rac family small guanosine triphosphate hydrolase (GTPase), PPARα and others is linked with retinal damage and microangiopathy in diabetic retinopathy models [40,41].
Diet-induced epigenetic modifications can affect future generations, heightening their susceptibility to metabolic diseases, including diabetes, non-alcoholic fatty liver disease and others. Furthermore, diet-related epigenetic changes have been observed impacting the function of epigenetic regulators enzymes along with their cofactors, such as TET enzymes and α-ketoglutarate (α-KG) derived from the tricarboxylic acid (TCA) cycle, in metabolic disorders [21,42]. This disruption further impairs epigenetic regulation and accelerates disease progression.
Thus, the reversible characteristics of epigenetic modifications that regulate the activation or inhibition of enzymes may serve as promising targets and valuable tools for understanding cellular and biological processes.
2.1. Oxidative Stress Impacting Epigenetic Mechanisms
The most studied property of plant-derived natural compounds is their antioxidant capacity. The scavenging of reactive oxygen species (ROS) is thought to be an effective measure to depress the level of oxidative stress in cells.
ROS are represented by free radicals such as hydrogen peroxide, hydroxyl radical (OH−), peroxy radical (ROO−), superoxide anion radical (O2−), and singlet oxygen, among others, which are necessarily produced in the human body as a result of normal physio-logical occurring processes, and also by exposure to environmental stressors. Oxidative stress emerges when the balance between the levels of ROS exceeds and cellular antioxidant defense system is overwhelmed [43].
Oxidative stress is a key modulator of the cellular environment, often leading to alterations in gene expression via epigenetic modifications. The imbalance between pro-oxidant species and antioxidant defenses can lead to changes in DNA methylation patterns, histone modifications, and non-coding RNA expression, which collectively impact chromatin structure and gene transcription. For instance, oxidative stress induces the formation of 8-oxo-2'-deoxyguanosine (8-oxo-dG), a DNA lesion that recruits repair machinery but also affects the local methylation status, potentially silencing tumor suppressor genes or activating oncogenes [44]. Such oxidative DNA damage has been linked to disrupted DNMT activity and subsequent global DNA hypo-methylation [45].
Multiple lines of scientific evidence have linked oxidative stress process to pathophysiology associated with various diseases including cancer [46,47]. The ROS can effectively induce DNA damage by nucleotide alterations, DNA strand breakage and mutations within genome of normal cells and thus promote further disease related transformation. Moreover, ROS are able to activate certain transcription factors including NF-κB, cyclooxygenases (COX), p53, HIF-1α, PPAR-γ, β-catenin/Wnt, and Nrf2, that leads to the expression of over 500 different genes, including growth factors, pro and anti-inflammatory cytokines, cell cycle regulatory molecules. [48,49]. Moreover, Nrf2, a crucial transcription factor that acts as a master regulator of cellular defense mechanisms against oxidative stress, electrophiles, and other harmful substances, is capable to influence epigenetic mechanisms by modulating histone acetylation and deacetylation [50]. This activity reshapes the cell's epigenetic landscape, impacting gene expression in response to oxidative stress [51].
Besides, the ROS can influence the DNA methylation pattern by directly oxidizing the catalytic site of DNMTs or by reducing the availability of the DNMT cofactor S-adenosyl methionine, SAM [52]. Cellular ROS levels themselves have been suggested to be the subject of epigenetic modulation. Thereby, ROS-generating systems of mitochondria and, including different subunits of the NOX complex, as well as antioxidant enzymes such as SODs and catalase, have been reported to be epigenetically regulated by different mechanisms [53]. ROS can modulate the activity of other epigenetic regulators such as histone acetyltransferases (HATs) and histone deacetylases (HDACs), influencing the histone acetylation epigenetic mechanism and thus inducing dysregulated gene expression in cancer and cardiovascular disorders [52].
Phytochemicals are highly bioactive molecules, yet a particular class, namely phenolics (phenolic acids, flavonoids), are the most abundant and exert the highest anti-oxidant activity in cellular environment [53,54].
Different mechanisms underlie the antioxidant capacity of phytochemicals, including the free-radical scavenging, hydrogen atom donation, ferric or cupric ions chelation, single oxygen quenching, and acting as a substrate for superoxide and hydroxyl radicals (Figure 1) [11,55]. The antioxidant activity of phytochemicals relies also on their capacity to inhibit essential enzymes responsible to the endogenous production of reactive oxygen species (ROS) such COXs, cytochrome P450 (CP450), lipoxygenase (LO), NADPH oxidase (NOX), nitric oxide synthases (NOSs) and xanthine oxidase (XO) [56]. The production of these enzymes is increased during inflammatory conditions; thus, inhibiting them can prevent ROS over-production, thus mitigating the associated cellular damage and inflammatory [57]. Since mediators of inflammation play key role in various pathologies, like cancer, metabolic and neurodegenerative disorders, the natural bioactive compounds might exert simultaneously anti-oxidant and anti-inflammatory activities [58].
In the view of recent scientific reports, the main molecular mechanism of phytochemicals acting to reduce cellular oxidative stress is by activating the Nrf2 pathway, which boosts antioxidant defense systems and diminishes inflammation [59,60]. Phytochemicals activate the Nrf2 pathway to combat oxidative stress primarily by modifying Keap1, the central negative regulator of Nrf2, or by promoting phosphorylation of Nrf2. Under normal conditions, Nrf2 is bound to Keap1 in the cytoplasm, which leads to its ubiquitination and degradation [61]. When phytochemicals induce oxidative or electrophilic stress, they cause conformational changes in Keap1 or directly modify it, leading to the release of Nrf2. Freed Nrf2 then translocate to the nucleus, where it forms heterodimers with Maf proteins and binds to antioxidant response elements (AREs) in the DNA. This binding initiates the transcription of various antioxidant and detoxification genes, such as heme oxygenase-1 (HO-1), glutathione-S-transferase (GST), and NAD(P)H quinone dehydrogenase 1 (NQO1), which enhance cellular antioxidant defenses and reduce oxidative damage [62,63].
At the same time, the phytochemicals influence epigenetic processes acting as epigenetic modulators primarily inhibiting the key epigenetic enzymes, including DNMTs, HDACs, HATs, BETs [64].
Moreover, the phytochemicals can directly donate the methyl group and act as co-substrates in DNA, RNA or histone methylation process or indirectly by affecting the methyl pool [25].
The combined action, as anti-oxidants, anti-inflammatory molecules and epigenetic modulators the phytochemicals represent valuable candidates for disease prevention and treatment through coordinated regulation of redox balance, inflammation process and epigenetic mechanisms(Figure 1).
2.2. Epigenetic Dysregulations in Inflammation Process
Inflammation is a highly dynamic and tightly regulated biological response, essential for defending the body against infections, injuries, and other harmful stimuli [65]. However, when the mechanism of controlling inflammation becomes dysregulated, the results can be chronic inflammatory states that contribute to wide range of diseases, from autoimmune disorders to cancer and cardiovascular pathologies [66]. Epigenetic mechanisms are increasingly recognized for their essential role in inflammation. They act as modulators of normal inflammatory responses by fine-tunning the protein expression and also as drivers of pathological inflammation when disrupted [67].
Epigenetic dysregulation is increasingly recognized as a critical factor in modulating inflammation and immune responses. DNA methylation plays a dual role, either suppressing or activating key pro-inflammatory genes. Aberrant activity of DNA methyltransferases (DNMTs) can lead to inappropriate methylation patterns that sustain chronic inflammation by silencing anti-inflammatory genes. For example, DNMT3B has been shown to hypermethylate promoter regions of tumor suppressor genes, thereby contributing to sustained inflammatory signaling [45].
During an inflammatory response, immune cells such as macrophages, dendritic cells, and lymphocytes undergo rapid changes in gene expression [68]. This transcriptional reprogramming is orchestrated in part by epigenetic modification, which enable these cells to quickly activate or repress large sets of genes in response to signals like cytokines, pathogen-associated molecular patterns (PAMPs), or damage- associated molecule patterns (DAMPs) [69,70,71]. Epigenetic control not only dictates which genes are expressed, but also how quickly and intensely cells react to danger signals [72].
Multiple epigenetic modifications may occur in a large number of diseases connected or related to inflammatory processes. For instance, the most investigated class of diseases, cancers, are often associated with epigenetic deregulation, that can occur at tumoral stroma or in immune cells. Epigenetics includes a series of reversible modifications that do not include gene mutations, which are strongly modulated by internal environment, mainly that is located close to the tumoral processes. In this way the pro-inflammatory surrounding the tumor can be responsible for the induction of epigenetic modifications [73,74].
Yang [75] analyzed the implications of epigenetic modifications – DNA methylation, chromatin remodeling, regulation of non-coding RNAs, histone changes – during the progression from chronic inflammation to colorectal cancer. The study also outlined the accelerating effects of such changes on the activation of cancer signaling pathways. Among these, of notable impact appear to be NF-kB, and STAT3 pathways. A similar analysis was provided by Pandareesh at all., for the case of prostate cancer, with an almost identical distribution and interaction between epigenetic factors, tumor environment and pro-inflammatory environment. [76].
When epigenetic controls mechanism malfunction, the balance between pro-inflammatory and anti-inflammatory pathways can be disrupted [71,77]. For instance: aberrant DNA methylation patterns may silence anti-inflammatory genes (e.g. IL-10) or abnormally activate pro-inflammatory ones, such as IL-6 or TNF-α [78,79,80]. Moreover, histone modification can alter chromatin accessibility, maintain certain genes in a persistently active state, even in the absence inflammatory stimuli, for example, increased histone acetylation of NF-kB target gene promotors has been linked to sustained inflammation in rheumatoid arthritis and inflammatory bowel disease [81,82,83]. Notable, non-coding RNAs, such as miR-155 or miR-21 are often overexpressed in chronic inflammation and can modulate immune cell differentiation, cytokine production, and apoptosis resistance [84,85]. The eraser proteins HDACs inhibition leads to increased histone acetylation, opening chromatin and facilitating the expression of anti-proliferative and regulatory genes like p21 [86,87]. The p21 gene plays a multifaceted role in cellular processes. It acts as a cyclin-dependent kinase (CDK) inhibitor, primarily inhibiting CDK2 and CDK1, which are crucial for cell cycle progression [88]. This inhibition leads to cell cycle arrest, particularly at the G1/S and G2/M transitions, preventing uncontrolled cell division and promoting genomic stability after DNA damage. Beyond cell cycle regulation, p21 also influences the cellular environment through the secretion of bioactive molecules, contributing to immunomodulatory functions and potentially aiding in the resolution of inflammation [89].
Histone modifications further contribute to inflammation-driven gene expression. Pro-inflammatory cytokines like TNF-α and IL-6 are regulated by dynamic histone acetylation and methylation at their promoter regions, with histone acetyltransferases (HATs) enhancing expression and histone deacetylases (HDACs) serving as brakes on transcription. Natural polyphenols such as resveratrol and quercetin have been shown to modulate these enzymes, reducing pro-inflammatory histone marks while enhancing the expression of anti-inflammatory genes [90].
Non-coding RNAs, especially microRNAs (miRNAs), play a pivotal role in fine-tuning inflammation. For instance, miR-146a suppresses NF-κB signaling, a master regulator of inflammation. Under oxidative stress conditions, miR-146a expression can be epigenetically downregulated, leading to sustained inflammation [44]. This highlights a complex interplay between ROS, epigenetic mechanisms, and inflammation.
Interestingly, dietary polyphenols may exert protective effects on inflammation also, through modulation of Nrf2, that coordinates antioxidant responses and modulates inflammatory pathways. By activating the Nrf2-ARE pathway, compounds like curcumin, sulforaphane, and EGCG (epigallocatechin gallate) can upregulate antioxidant genes while simultaneously downregulating pro-inflammatory mediators [51]. This dual action highlights their potential as epigenetic modulators in chronic inflammatory diseases.
Furthermore, recent evidence from a clinical study suggests that polyphenol-rich diets can alter epigenetic aging markers in immune cells, indirectly influencing inflammation status [91]. Such interventions might slow down immunosenescence and mitigate inflammation-associated disorders.
Notably, such epigenetic changes can affect not only immune cells but stromal and epithelial cells within the inflamed microenvironment, contributing to persistent tissue remodeling and fibrosis [74].
Another field of major interest is coronary artery disease, where, on one hand, disease progression, in which epigenetic changes – such as histone methylation, phosphorylation and acetylation, pay key roles. Besides these, the lncRNAs, miRNAs and circRNAs are also involved in epigenetic processes, and help the regulation of gene expression in response to environmental factors and lifestyle. Such modifications in de-regulation of genes regulating vascular function, inflammation, lipid metabolism or oxidative stress [92,93].
Collectively, this epigenetic modification can act as a form of inflammatory memory, locking cells into a pro-inflammatory state even after the initial trigger has passed [69]. This phenomenon is especially evident in chronic diseases like atherosclerosis, rheumatoid arthritis, and asthma, where immune cells show persistent activation profiles [94].
A defending feature of epigenetic regulation is its plasticity of epigenetic marks that are not static and can be influenced by environmental exposures [95]. Factors such as smoking, diet, psychological stress, pollution, infections, and gut microbiota derived metabolites can all modify the epigenetic landscape [96,97].
For instance, short-chain fatty acids like butyrate, produced by commensal bacteria, can inhibit HDACs and thereby affect gene expression in immune cells [98]. Similarly oxidative stress resulting from environmental pollutants can trigger histone modification that amplify inflammatory gene transcriptions [99,100]. These influences suggest that chronic exposure to harmful environmental factors may not only trigger inflammation but may epigenetically fix it into the biology of the tissues [101,102].
Even subtle lifestyle factors, such as circadian rhythm disruption or chronic low-grade stress, may modulate the expression of inflammatory genes through epigenetic routes [103].These influences suggest that chronic exposure to harmful environmental factors may not only trigger inflammation but may epigenetically fix it into the biology of the tissue [104].
Of interest is also the fact that natural compounds, for instance naringenin and naringin (pharmacologically active in anti-cancer and anti-cardiovascular diseases, displaying anti-oxidant and anti-inflammatory activities), are also involved in epigenetic mechanisms, thus broadening the mechanisms in supporting restoration of health [105,106].
Overall, the integration of dietary components, oxidative stress, and epigenetic alterations forms a complex network that modulates inflammation. Targeting these pathways through nutraceuticals offers a promising avenue for managing chronic inflammation-related diseases.
The reversibility of epigenetic marks makes them attractive targets for novel therapeutic strategies. Drugs that inhibit DNMT1 or HDACs are already being explored in cancer therapy but they show promise in treating chronic inflammatory diseases as well. By reprogramming immune cells epigenetically either to reduce their inflammatory potential or restore tolerance such intervention could offer more precise and durable solution than broad spectrum immunosuppressants [107]. For example, HDAC inhibitors may promote the expression of p21 and other regulatory genes, helping to dampen persistent inflammation. Similarly, DNMT inhibitors could potentially reactivate silenced anti-inflammatory genes or detoxifying enzymes like GSTP1, restoring homeostasis in chronically inflamed tissues [108].
3. Phytochemicals as Antioxidant, Anti-Inflammatory Agents with Epigenetic Modulator Capacities
Chronic inflammation and oxidative stress are fundamental contributors to the pathogens of numerous non-communicable diseases, including cardiovascular disorders, cancer, neurodegenerative conditions, diabetes, and autoimmune syndromes. Oxidative stress arises from an imbalance between reactive oxygen species and the antioxidant defense system, which may induce also the inflammation by elevating the levels of pro-inflammatory mediators and cytokines. These two pathological states are tightly interconnected and often exacerbate one another, creating a self-perpetuating cycle of tissue damage and immune dysregulation [109,110].
In recent years, phytochemicals found in fruit, vegetables, herbs, and medicinal plants have gained significant attention for their ability to counteract oxidative and inflammation damage. These compounds include flavonoids, phenolic acids, terpenoids, alkaloids, and organosulfur compounds, many of which possess strong radical-scavenging capabilities and modulate inflammatory signaling pathways such as NF-kB, Nrf2, PI3K/Akt, and MAPKs (Table 1) [111,112] .
Unlike conventional pharmaceuticals, phytochemicals often have multi-target potential, by modulating cellular processes simultaneously, with lower toxicity profiles. For instance, flavonoids enhance antioxidant enzymes activities, such as SOD, CAT, GSH, while simultaneously inhibiting pro-inflammatory mediators like TNF-α, IL-6 and COX-2 through suppression of transcription factors and kinases [113,114,115]. These mechanisms not only alleviate oxidative stress but also reduce systemic and local inflammation, contributing to disease prevention and adjunctive therapy. Notably, flavonoids represent a structurally diverse class of compounds-including flavonols, flavones, flavanones and anthocyanidins each with distinct bioactivities and tissue selectivity [116]. This diversity allows them to modulate also a broad range of molecular targets implicated in inflammation, oxidative damage, and metabolic imbalance [117]. The phytochemicals multi-target capacity supports their potential in disease prevention and adjunctive therapy.
In Table 1 are presented the phytochemicals with in vitro demonstrated anti-oxidant, anti-inflammatory and potential epigenetic modulator capacities.

Table 1.
Example of phytochemicals with antioxidant, anti-inflammatory activities and epigenetic modulator capacity, investigated in vitro.
Table 1.
Example of phytochemicals with antioxidant, anti-inflammatory activities and epigenetic modulator capacity, investigated in vitro.
| Class of compound | Phytochemicals | Type of study | Antioxidant | Anti-inflammatory | Epigenetic modulator | References |
|---|---|---|---|---|---|---|
| Flavonoids | Hesperidin | MCF-7 ( 50 µM) |
Increases activities of SOD, CAT, GSH; reduces lipid peroxidation; Activates Nrf2 |
Reduces expressions of TNF-α, IL-6 ex-pression | Inhibits DNMT1; hypo-methylation of p16 promoter | [118,119,120] |
| Phloretin | RAW 264.7 macrophages (25 µM); A549 cells |
Inhibits ROS production and increases GSH activity | Suppresses NF-κB activation and reduces iNOS and COX-2 expression | Inhibits HDAC; increases histone acetylation, p21 expression | [121,122,123] | |
| Genistein | MCF-7, PC3, HL-60 cells | Increases antioxidant enzymes, reduces lipid peroxidation | Reduces pro-inflammatory cytokines (IL-6, IL-1β) expression via NF-κB | Inhibits DNMT1, demethylates and reactivates tumor suppressor genes | [124,125,126,127] | |
| Phenolic acids | Caffeic acid | Caco-2, HeLa cells (20 µM) |
Neutralizes free radicals and boosts antioxidant enzymes | Inhibits pro-inflammatory cytokines and NF-κB activation | Inhibits DNMT; DNA hypomethylation; gene reactivation | [128,129,130] |
| Coumaric acid | LPS-stimulated RAW 264.7 cells; MDA-MB-231 cells (30 µM) |
Free radical scavenger and increases SOD activity | Reduces NO production and COX-2 expression | Inhibits HDAC; increases histone acetylation | [131,132,133] | |
| Terpenoids | Lycopene | HepG2, PC3 cells (10 µM) |
Reduces ROS and increases antioxidant enzyme activity | Inhibits COX-2 expression and lowers prostaglandin E2 levels | ||
| Silibinin | HepG2, DU145 cells (50 µM) | Increases SOD and CAT activity; reduces lipid peroxidation | Inhibits pro-inflammatory cytokines and NF-κB activation | Inhibits DNMT and HDAC; hypomethylation and histone acetylation | [134,135,136] | |
| Sesquit-erpenoids | Artemisinin | HeLa cells (50 µM) |
Reduces oxidative stress by lowering ROS and activating Nrf2 | Inhibits IL-1β, TNF-α, and COX-2 expression | — | [137] |
| Mono-terpenoids | Geraniol | RAW 264.7 macrophages (50 µM) | Reduces ROS production and increases GSH activity | Suppresses NO, IL-6, and TNF-α production by inhibiting NF-κB | — | [138] |
| Organo-sulfur com-pounds | Allyl mercaptan | HUVEC, HT-29 cells (15 µM) | Enhances antioxidant enzyme activity and reduces ROS | Inhibits adhesion molecules and reduces vascular inflammation | Inhibits HDAC; increases histone acetylation; gene reactivation | [139,140,141,142] |
Hesperidin, a bioactive flavonoid predominantly found in citrus such as orange lemons has demonstrated significant antioxidant and anti-inflammatory properties, particularly in the context of breast cancer [143,144]. In vitro studies on MCF-7 breast cancer cells treated with 50 µM hesperidin revealed an increase in antioxidant enzymes such as SOD, CAT, and glutathione, alongside a reduction in lipid peroxidation [145]. These effects are attributed to the activation of the Nrf2 signaling pathway, which plays crucial role in cellular defense against oxidative stress. Furthermore, hesperidin significantly decreased the expression of pro-inflammatory cytokines such as TNF-alpha and IL-6 by inhibiting pathway, thereby modulating inflammation within the tumor microenvironment [146,147]. These findings underscore hesperidin’s potential as therapeutic agent managing oxidative stress and inflammation in breast cancer models [148]. Beyond this antioxidant and anti-inflamatory activities hesperidin also exerts epigenetic modulatory effects. Studies have demonstrate that hesperidin can reduce global DNA methylation levels by inhibiting DNA methyltransferases (DNMTs) particularly in cancer models [149]. In vitro and in vivo models of cancers have shown that hesperidin downregulates hypermethylation of repetitive DNA elements, such as KINE-1 and ALU sequences, and modulates microRNA expression, including the upregulation of tumor-suppressive miR-34a and downregulation of oncogenic miR-221 [150]. These epigenetic modifications contribute to the reactivation of tumor suppressor genes and the inhibition of oncogenic pathways, further supporting hesperidin”s role in cancer and chemoprevention. These findings suggest that hesperidin supports human health through several interconnected mechanisms. By enhancing antioxidant defenses, reducing harmful inflammation, and influencing epigenetic processes that control gene activity, hesperidin shows great promise as a natural compound that could help prevent or support the treatment of cancer in an integrative approach.
Genistein is a naturally occurring isoflavone, it is predominantly found in leguminous plants, especially soybeans (Glycine max), where it exists mainly in its glycosylated from, genistin [151]. Genistein exhibits a spectrum of biological activities beyond its well know estrogenic and anti-cancer effects, including moderate antioxidant and anti-inflammatory actions, as well as epigenetic mechanisms regulator [152,153]. This compound exerts moderate but significant antioxidant effects, primarily by reducing ROS [57] and enhances the activities of antioxidant enzymes such as SOD), catalase CAT and glutathione peroxidase, largely via NRf2 signaling pathway [154]. These effects stabilize redox balance and protect cellular components from oxidative damage especially under inflammation or tumorigenic conditions. In addition to its redox balancing effects, genistein demonstrated anti-inflammatory activity by downregulation the Nf-kB signaling pathway and suppressing the expression of pro inflammatory cytokines [155] such TNF- alpha, Il-6, and Il-1ß [156].
Genistein also inhibits the activity of enzymes like COX-2 and iNOS, further reducing the inflammatory response [157]. This is particularly relevant in chronic diseases where low-grade inflammation plays a pathogenic role in atherosclerosis and arthritis. One of genistein most compelling biological features is its to ability to modulate epigenetic mechanisms by inhibiting DNMTs, especially DNMT1, leading to reactivation of silenced tumor suppressor genes such as p16, RASSF1A and BRCA1 [158,159]. Recent findings also suggest that genistein can modulate the activity sirtuins and potentially interact with TET-enzymes, impacting DNA demethylation processes and metabolic-epigenetic crosstalk [160].
Phloretin, a dihydrochalcone flavonoid predominantly found in apples, has demonstrated notable antioxidant and anti-inflammatory properties (10.3390/molecules24020278). In vitro studies utilizing RAQ 264.7 macrophages treated with 25 µM phloretin have shown a significant reduction in reactive oxygen species (ROS) production and an enhancement of intracellular GSH levels, indicating its potential in mitigating oxidative stress [161].
Furthermore, phloretin has been observed to suppress the activation of nuclear factor kappa B (NF-κB), a pivotal transcription factor in the inflammatory response. This suppression leads to the downregulation of pro-inflammatory enzymes such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2). Additionally, phloretin modulates the mitogen-activated protein kinase (MAPK) pathways, including extracellular signal-regulated kinase (ERK), c-Jun-n-TERMINAL KINASE (JNK), and p38 MAPK [162]. These findings are supported by Leet et al, which demonstrated that phloretin attenuates lipopolysaccharide-induced acute lung injury in mice via blockade of the NF-kB and MAPK pathways [163]. These pathways are integral to the production of pro-inflammatory mediators [161]. The combined modulation of these signaling pathways by phloretin suggests its potential application in managing chronic inflammatory conditions and cardiovascular diseases [164,165].
Figure 1.
Role of phytochemicals in cell survival and metabolism. Phytochemicals act as homeostatic factors for cells: They intervene directly on ROS by modulation of SOD, CAT and GSH activity. At the same time, they promote recruitment of repair mechanisms to counteract the DNA damage done by ROS, thus promoting survival of the cells. Phytochemicals counteract the epigenetic effects of ROS by directly modulating DNA methylation and histone acetylation. Alongside these cell survival effects, phytochemicals interact with some extracellular proliferation and inflammation signals, by inhibiting their internal pathways, thus altering gene expression of NO, Cox2, IL1B, TNF-α and their subsequent pro-proliferative effects. .
Figure 1.
Role of phytochemicals in cell survival and metabolism. Phytochemicals act as homeostatic factors for cells: They intervene directly on ROS by modulation of SOD, CAT and GSH activity. At the same time, they promote recruitment of repair mechanisms to counteract the DNA damage done by ROS, thus promoting survival of the cells. Phytochemicals counteract the epigenetic effects of ROS by directly modulating DNA methylation and histone acetylation. Alongside these cell survival effects, phytochemicals interact with some extracellular proliferation and inflammation signals, by inhibiting their internal pathways, thus altering gene expression of NO, Cox2, IL1B, TNF-α and their subsequent pro-proliferative effects. .
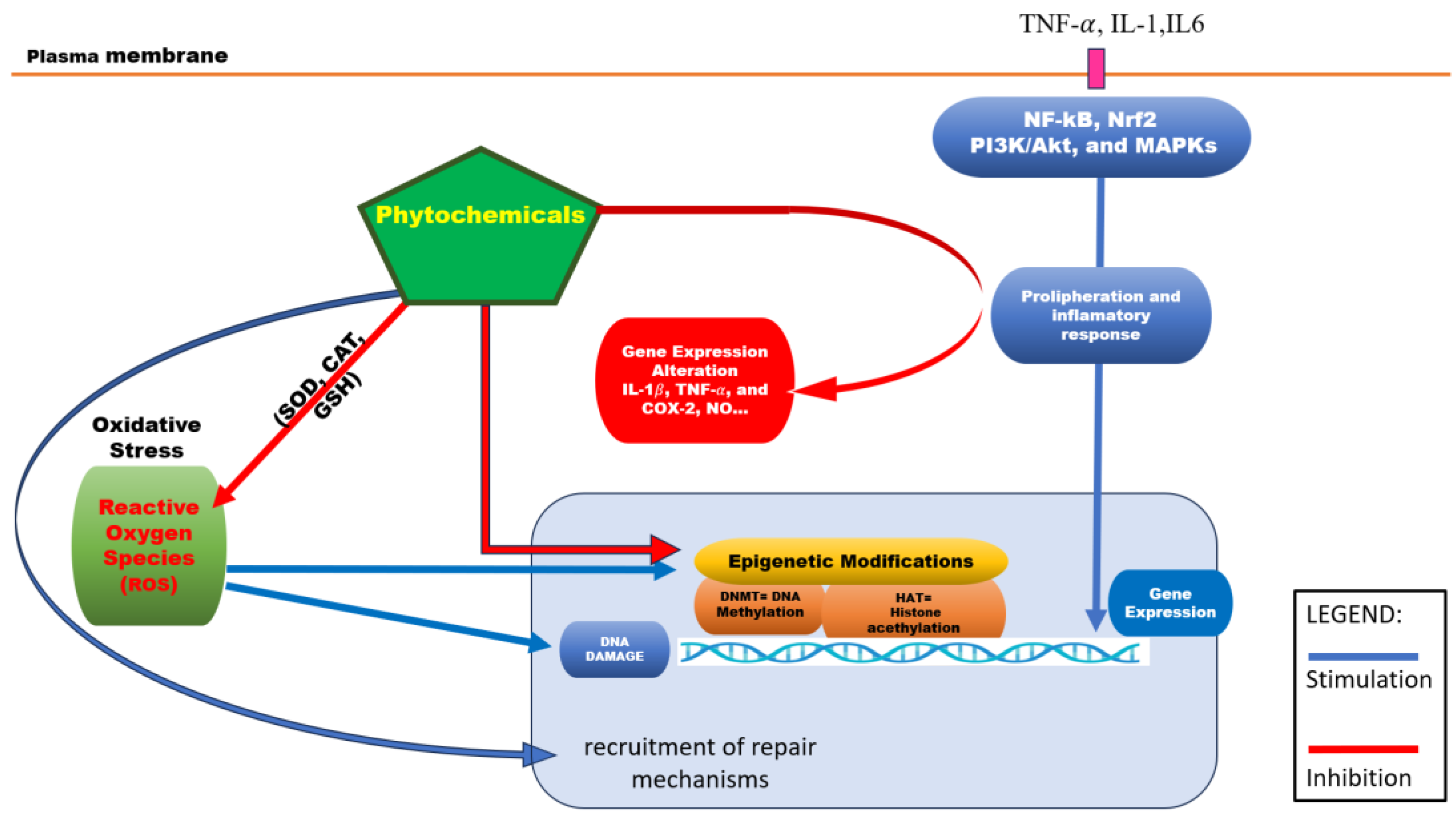
In addition to these well-established antioxidant and anti-inflammatory actions, emerging evidence highlights phloretin’s potential role as an epigenetic modulator. According to recent findings, phloretin can inhibit DNA methyltransferases (DNMTs), enzymes responsible for adding methyl groups to DNA and often involved the silencing of tumor suppressor genes [45] . By modulating DNA methylation patterns, phloretin may help restore normal gene expression profiles that are disrupted in various chronic diseases, including cancer. Although further research is needed to clarify its impact on histone modification and microRNA regulation, these initial insights suggest that phloretin’s health benefits extended beyond redox and inflammatory pathways to include epigenetic regulation of gene expression. Altogether, these multiple molecular action-combining antioxidant, anti-inflammatory, and epigenetic modulatory effects-position phloretin as a promising candidate for managing chronic inflammatory diseases, metabolic disorders and cancer prevention.
Lycopene a lipophilic carotenoid found in tomatoes and watermelon acts, as a potent scavenger of ROS in HepG2 cells [166]. At a concentration 10 µM lycopene reduced ROS levels and increased antioxidant enzyme activity [167]. Additionally, it inhibited COX-2 expression and decreased prostaglandin E2 synthesis, indicating an anti-inflammatory effect. Lycopene also modulates transcription factors such as AP-1 and STAT3, suggesting its potential for liver cancer prevention and hepatic inflammation reduction [168]. In addition to these well-established effects, lycopene also exerts epigenetic modulatory activity. According to recent findings, lycopene can influence DNA methylation patterns by inhibiting DNA methyltransferases (DNMTs). This inhibition may contribute to the reactivation of silenced tumor suppressor genes and the correction of aberrant gene expression associated with carcinogenesis. While further research is needed to fully elucidate its effects on histone modification and non-coding RNAs, these early insights suggest that lycopene contributes to regulation of gene activity through epigenetic pathways, adding a new layer to its chemopreventive potential[169]. Overall, these findings suggest that lycopene supports liver health through several complementary actions. By reducing oxidative stress, calming inflammation, and influencing gene activity through epigenetic mechanism, lycopene shows promise cells and reduce the risk of liver cancer. [45]
Caffeic acid, a hydroxycinnamic acid prevalent in coffee, fruits, and vegetables, has demonstrated significant antioxidant and anti-inflammatory properties, particularly within intestinal epithelial cells [169,170]. In vitro studies utilizing Caco-2 cells treated with 20 µM caffeic acid have shown a notable reduction of ROS production and an enhancement of endogenous antioxidant enzyme activities, including SOD, CAT, and GSH [171,172]. These effects are primarily mediated through the activation of Nrf2 signaling pathway, which plays a crucial role in cellular defense against oxidative stress [173,174].
Moreover, caffeic acid has been observed to inhibit the activation of nuclear factor kappa B NF-kB, a key transcription factor involved in the inflammatory response [175]. This inhibition leads to a decrease in the section of pro-inflammatory cytokines, such as interleukin-8 (IL-8), thereby mitigating inflammation within the intestinal environment [176]. Additionally, caffeic acid modulates the phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) and mitogen-activated protein kinase (MAPK) signaling pathways, including extracellular signal-regulated kinases (ERK), c-Jun N-terminal kinase (JNK), and p38 MAPK [177]. These pathways are integral to the regulation of cellular responses to oxidative stress and inflammation [178].
Collectively, these findings suggest that caffeic acid exerts protective effects against oxidative stress and inflammatory conditions in intestinal epithelial cells, highlighting its potential therapeutic applications in managing intestinal inflammation and related disorders [179].
These antioxidant and anti-inflammatory mechanism, caffeic acid has also been identified as a modulator of epigenetic processes.
Recent research highlights its capacity to influence DNA methylation and histone modifications, two key mechanism controlling gene expression without altering the underlying DNA sequence [180] Specifically, caffeic acid has been shown to inhibit the activity of DNA methyltransferases (DNMTs), potentially reversing anormal gene silencing associated with chronic diseases such as cancer and inflammatory disorders. Moreover, caffeic acid may impact histone acetylation and methylation states, further contributing to the regulation of genes involved in oxidative stress responses, inflammation and cellular homeostasis.
These findings show that caffeic acid protects intestinal cells not only by reducing oxidative stress and inflammation but also by helping restore healthy gene activity, making it a promising natural compound for managing intestinal disorders.
p-Coumaric acid, a naturally occurring hydroxycinnamic acid found in foods like red wine and balsamic vinegar, exhibits significant antioxidant and anti-inflammatory properties [181,182]. In vitro studies using lipopolysaccharide (LPS)-simulated RAW 264.7 macrophages have demonstrated that treatment with p-coumaric acid at concentration ranging from 10 to 100 μg/mL effectively reduces the production of nitric oxide (NO) and suppresses the expression of pro-inflammatory mediators such as inducible nitric oxide synthase (iNOS) and cyclooxygenase 2 (COX-2) at both mRNA and protein levels [131,183].
The anti-inflammatory effects of p-coumaric acid primarily mediated through the inhibition of the nuclear factor kappa B (NF-kB) signaling pathway. Specifically, p-coumaric prevents the phosphorylation and subsequent degradation of IκBα, an inhibitor of NF-kB, thereby blocking the translocation of the NF-kB p65 subunit into the nucleus and reducing the transcription of pro-inflammatory genes [131].
Additionally, p-coumaric acid modules the mitogen-activated protein kinase (MAPK) pathways by inhibiting the phosphorylation of extracellular signal-regulated kinase (ERK1/2) and c-Jun N-terminal kinase (JNK), which are crucial in the expression of inflammatory mediators [184].
Beyond its effects on macrophages, p-coumaric acid has been shown to exert protective effects against atherosclerosis. In studies involving THP-1 macrophages treated with oxidized low-density lipoprotein (ox-LDL) and LPS, p-coumaric acid at 20 μM significantly inhibited lipid accumulation and foam cell formation. This effect is attributed to the upregulation of cholesterol efflux-related genes, such as ATP binding cassette transporter A1 (ABCA1), liver X receptor alpha (LXRα), and peroxisome proliferator-activated receptor gamma (PPARγ), and the downregulation of lipid uptake related genes, including lecithin-like oxidized low-density lipoprotein receptor-1 (LOX-1), cluster of differentiation 36 (CD36), and scavenger receptor class A1 (SR-A1). Furthermore, p-coumaric acid suppressed the expression of NF-kB, COX-2, tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), indicating its potential role in mitigating inflammatory response associated with atherosclerosis. These findings suggest that p-coumaric acid hold promise as a therapeutic agent in managing oxidative stress-related inflammatory diseases, such as atherosclerosis, by modulating key. Inflammatory signaling pathways and promoting cholesterol homeostasis [185].
Anacardic acid, a phenolic lipid predominantly found in cashew nuts (Anacardium occidentale) and mango seeds, has gained attention for its potent neuroprotective properties, particularly in the context of neurodegenerative diseases such as Parkinson’s and Alzheimer’s [186].
In a study by Augusto et al (2020), purified anacardic acid (AAs) were administered at a dose of 50mg/kg in a rotenone induced mouse model Parkinson’s disease. The treatment led to a significant reduction in lipid peroxidation and NO levels, alongside an increase in the glutathione (GSH)/oxidized glutathione (GSSH) ratio in both the substantia nigra and striatum regions of the brain. These changes indicate a restoration of redox balance and mitigation of oxidative stress-induced neuronal damage [187].
Furthermore, the study observed that AAs attenuated the activation of nuclear kappa B (NF-kB) and reduced the expression of pro-inflammatory cytokines such as interleukin-1β (IL-1β). This anti-inflammatory effect was complemented by the downregulation of matrix metalloproteine-9 (MMP-9) and glial fibrillary acid protein (GFAP), markers associated with neuroinflammation and astrogliosis.
In addition to its neuroprotective and anti-inflammatory effects, anacardic acid also shows potential as a natural modulator of gene activity. Recent studies suggest that it can block the activity of histone acetyltransferases (HATs) particularly p300/CBP, enzymes that normally loosen chromatin structure to activate gene expression [188]. By limiting histone acetylation, anacardic acid may help turn down the expression of pro-inflammatory genes and other harmful pathways involved in neurodegenerative diseases. This ability to influence gene regulation adds another important layer to its protective action, complementing its antioxidant and anti-inflammatory effects. Together, these findings highlight anacardic acid as a promising natural compound that works on multiple levels to protect neurons and slow the progression of neurodegeneration. Allyl mercaptan, a sulfur containing compound derived from garlic (Allium sativum), has been shown to enhance the activity of key antioxidant enzymes such as superoxide dismutase (SOD) and catalase (CAT), while also reducing intracellular reactive oxygen species (ROS) levels in HUVEC cells treated at a concentration of 15μM.(https://doi.org/10.1016/j.jff.2023.105657
[189]. This antioxidant enhancement contributes to the perseveration of redox homeostasis in vascular endothelial cells [190].
In addition to its antioxidant effects, allyl mercaptan also inhibits the expression of adhesion molecules, such as ICAM1 and VCAM-1, and reduces vascular inflammation by preventing NF-kB nuclear translocation, a key event in endothelial activation. Although the precise molecular pathway remains under investigators, some studies have demonstrated that offer garlic-derived compounds like diallyl disulfide and diallyl trisulfide may act. Synergistically or more potently through NF-k-B inhibition [191].
These findings support the therapeutic potential of allyl mercaptan as a candidate for endothelial protection and cardiovascular disease prevention, particularly in the context oxidative stress and inflammation [192].
Moreover, beyond its antioxidant and anti-inflammatory action, alyl mercaptan has been reported to exert epigenetic regulatory effects. According to findings from carcinogenesis ([141], allyl mercaptan can inhibit DNA methyltransferase DNMT activity, leading to hypomethylation of promoter regions in key genes involved in cell regulation, such as p21 WAF1. This epigenetic modulation results in the reactivation of tumor suppressor pathways and contributes to the control of cellular proliferation and apoptosis. Therefore, the protective role of allyl mercaptan in in vascular endothelial cells may not only involve redox homeostasis and inflammation suppression but also the regulation of gene expression through epigenetic modifications, highlighting its complex mechanism of action in maintaining cardiovascular health.
Silybin, a flavonolignan extracted from Silybum marianum (milk thistle), is a widely recognized for its hepatoprotective, antioxidant, anti-inflammatory effects. In vitro studies on HepG2 cells have shown that silybin increase the activity of endogenous antioxidant enzymes, including SOD and CAT, while significantly decreased lipid peroxidation, thereby alleviating oxidative damage [193,194]. The compound also inhibits the activation of NF-Kb, the key transcription factor regulating inflammatory cytokines [195]. As a result, treatment with silybin leads to reduced expression of TNF-α, IL-6, and iNOS, contributing to a pronounced anti-inflammatory effect. Moreover, it modulates JAK/STAT and TGF-βsignaling pathways, both of which play central roles in liver fibrosis and hepatocarcinogenesis [196].
Beyond its cellular effects, silibinin is under clinical investigation for its antifibrotic and chemoprotective potential, particularly in the context of chronic liver diseases such as non-alcoholic fatty liver disease (NAFLD) and hepatoprotective carcinoma (HCC) [197]. Its multifunctional properties-spanning antioxidation, inflammation suppression, and fibrotic pathway interference-highlight its promise as a therapeutic agent in hepatic disorders [198].
In addition to these biological effects, silybin has also been shown to exert epigenetic regulatory activities. According to research [199] silybin modulates the activity of DNA methyltransferases (DNMTs) and histone deacetylases (HDACs), leading to changes in DNA methylation and histone acetylation status in liver cells. These epigenetic modification results in the reactivation of tumor suppressor genes and suppression of oncogenic pathways, contributing to its chemopreventive potential in hepatocarcinogenesis. Therefore, the therapeutic role of silybin in liver diseases may also involve the regulation of gene expression at epigenetic level, providing a broader mechanistic basis for its hepatoprotective and anticancer effects.
Artemisin, a sesquiterpene lactone extracted from Artemisia annua, has shown strong antioxidant, anti-inflammatory, and anticancer properties [200,201]. In HeLa cells treated with 50μM artemisinin, studies have demonstrated a significant reduction in ROS and enhanced expression of antioxidant enzymes such as a heme oxygenase-1 (HO-1) via activation of the Nrf2 signaling pathway [202,203].
In addition to its antioxidant effects, artemisin effectively inhibits NF-kB activation, resulting in the downregulation of pro-inflammatory cytokines such as IL-1β, TNF-α, and COX-2 [204,205]. These molecular effects contribute to the compound’s ability to suppress the inflammatory tumor microenvironment and modulate immune responses.
Artemisinin also exerts pro-apoptotic activity, inducing mitochondrial membrane potential loss, caspase activation, and cell cycle arrest in various cancer cell lines, including breast, cervical, and colorectal cancers [206]. Moreover, its antiangiogenic properties, achieved through inhibition of VEGF-mediated endothelial cell migration and proliferation, make it a valuable candidate in anti-tumor therapy, particularly for limiting tumor neovascularization [207].
Due to this multimodal activity profile-antioxidant, anti-inflammatory, apoptotic, and anti-angiogenic, artemisin is increasingly regarded as a promising adjunct in oncological treatment and is currently under investigation in both preclinical and early-phase clinical studies [208].
Geraniol, a monoterpene found in rose and citronella essential oils, reduces ROS levels and increases GSH activity in RAW 264.7 macrophages (50 μM). By inhibiting NF-kB, it decreases NO, IL-6, and TNF-α [209]. Geraniol also modulates P13K/Akt and MAPK pathways, with potential application in managing chronic inflammatory and metabolic disorders [210],[202]
These effects, recent research has uncovered an emerging role for geraniol in epigenetic regulation. A 2025 study demonstrate that geraniol can influence gene expression by modulating DNA methylation and histone acetylation levels, processes essential for controlling how genes are turned on or off [211]. Specifically, geraniol was found to inhibit the expression of DNA methyltransferases (DNMTs) and histone deacetylases (HDACs), enzymes that play key roles in maintaining aberrant gene silencing in inflammation and metabolic dysfunction. Through this action, geraniol may help restore normal gene expression patterns, further supporting its protective effects at the molecular level. This evidence suggests that geraniol supports health by reducing oxidative stress and inflammation while also helping to restore healthy gene activity, making it a promising natural option for preventing chronic diseases.
Various classes of bioactive compounds including flavonoids (hesperidin, phloretin, silybin), carotenoids (lycopene), phenolic acids (caffeic acid, p-coumaric acid, anacardic acid), sulfur-containing molecules (allyl mercaptan), terpenes (geraniol), and sesquiterpene lactones (artemisinin)-have demonstrated strong antioxidant and anti-inflammatory activities in multiple in vitro studies,
Most of these phytochemicals enhance the activity of endogenous antioxidant enzymes such as SOD, CAT, and GSH, while reducing lipid peroxidation and ROS accumulation. These mechanisms contribute to cellular protection against oxidative damage.
Additionally, several compounds, such as hesperidin, lycopene, artemisn, and silybin, demonstrate antiproliferative effects through inhibition of NF-kB signaling and suppression of pro-inflammatory mediators like TNF-α, IL-6, COX-2, and iNOS. These bioactive compounds support potential roles as chemopreventive and therapeutic agents in inflammation-associated disease, including cancer, cardiovascular, and neurodegenerative disorders.
Moreover, several compounds, such as hesperidin, lycopene, artemisinin, and silibinin, demonstrate antiproliferative effects through inhibition of NF-κB signaling and suppression of pro-inflammatory mediators like TNF-α, IL-6, COX-2, and iNOS. These bioactivities support their potential roles as chemopreventive and therapeutic agents in inflammation-associated diseases, including cancer, cardiovascular, and neurodegenerative disorders.
Interestingly, some phytochemicals are able to regulate epigenetically the levels of NRF2, which acts as a master regulator of antioxidant response. As such the coupling NRF2 to the antioxidant response element sequences in promoters of genes results in mechanisms of antioxidant detoxification. Yates et al Phytochemicals documented to modulate NRF2 signaling act by reversing hypermethylated states in the CpG islands of NFE2L2 or Nfe2l2, via the inhibition of DNA methyltransferases (DNMTs) and histone deacetylases (HDACs), through the induction of ten-eleven translocation (TET) enzymes, or by inducing miRNA to target the 3′-UTR of the corresponding mRNA transcripts. To date, fewer than twenty phytochemicals have been reported as NRF2 epigenetic modifiers, including cur-cumin, sulforaphane, resveratrol, reserpine, and ursolic acid. (Bhattacharjee S, Dashwood RH. Epigenetic Regulation of NRF2/KEAP1 by [212] Bhattachaarjee et al. provided a broader view of the action of phytochemicals on a broad array of disorders, including cancer and chronic diseases. A more focused insight is provided by Nurkolis et al (Journal of Agriculture and Food Research, Volume 21, 2025, [213], who focused on natural compound acting as epigenetic regulators in diabetes mellitus. As major actors were revealed curcumin, resveratrol, sulforaphane, genistein and quercetin.
Naturally occurring phytochemicals such as those mentioned above, along with other well studied compound like resveratrol [214], quercetin [215], apigenin [216] are commonly used in herbal extracts and show potential as complementary therapies in managing inflammatory diseases, cancer, and other chronic condition.
When these bioactive compounds are consumed from natural sources, they typically present alongside other related phytochemicals. This combination allows them to act on multiple regulatory pathways at the same time [217] . Additionally, these natural compounds can be combined with synthetic medicines to create more complex therapeutic strategies that target several disease mechanism simultaneously[218,219].
4. Conclusions and Future Perspective
The preventive potential of phytochemicals is a rapidly growing area of research, with in vitro and in vivo studies demonstrating their ability to target multiple cellular and molecular pathways simultaneously, leading to enhanced biological effects. These natural bioactive compounds are increasingly recognized for their roles in promoting health and preventing chronic disease. The majority of chronic diseases are characterized by early epigenetic dysregulations which might disturb the normal patterns of gene expression. The intrinsic factors like aging and genetic background alongside with extrinsic environmental factors (e.g. diet, pollution, radiation, stress) can significantly influence epigenetic mechanisms, ultimately impacting human’s health.
The reversibility of epigenetic modifications and the ability to reset cellular homeo-stasis through compounds with epigenetic modulator capacity have opened new avenues for prevention interventions, particularly in aging, cancer, cardiovascular and metabolic disorders. Targeting these reversible changes with dietary bioactive compounds offers a promising approach to prevent diseases where early epigenetic abnormalities play a key role.
This review emphasizes that certain phytochemicals, including flavonoids, that possess both antioxidant and anti-inflammatory properties, and additionally, they can directly or indirectly inhibit the aberrant activities of epigenetic regulators. The underlying molecular mechanisms of phytochemicals multifaceted actions is incompletely unveiled, but certainly their capability to act through various epigenetic pathways, including DNA methylation, histone modifications and chromatin remodeling are potentially impacting a range of cellular processes.
The translation of preclinical data to preventive medicine is a challenging process requiring rigorous clinical trials, acknowledging individual variability in the gut microbiota and its impact on phytochemical metabolism. Long-term trials which should consider the synergistic effects of dietary phytochemicals, are also needed to evaluate overall efficacy and safety in humans, as the diet provides a combination of phytochemicals with specific cellular and organ-level actions.
Author Contributions
Conceptualization, D.C., M.C.R., M.I.,.; methodology, R.A., S.P., C.T., M.L.P., ; software, D.C., M.I.; validation, M.C.R, R.A., S.P., formal analysis, R.A. S.P., C.T., M.L.P.,; investigation, D.C, M.C.R., M.I., S.P.; resources, D.C., M.C.R., M.I.,R.A., S.P.,C.T., M.L.P.,; data curation, R.A., C.T., M.L.P.; writing—original draft preparation, D.C., M.C.R, M.I.; writing—review and editing, R.A., C.T., M.L.P.; visualization, S.P., C.T., M.L.P.; supervision, R.A., C.T., M.L.P.; project administration, C.T., M.L.P.; funding acquisition, D.C., M.C.R., M.I., R.A., S.P., C.T., M.L.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors have reviewed and edited the output and take full responsibility for the content of this publication.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Anjali; Kumar, S.; Korra, T.; Thakur, R.; Arutselvan, R.; Kashyap, A.S.; Nehela, Y.; Chaplygin, V.; Minkina, T.; Keswani, C. Role of Plant Secondary Metabolites in Defence and Transcriptional Regulation in Response to Biotic Stress. Plant Stress 2023, 8, 100154. [CrossRef]
- Full Text.
- Rabizadeh, F.; Mirian, M.S.; Doosti, R.; Kiani-Anbouhi, R.; Eftekhari, E. Phytochemical Classification of Medicinal Plants Used in the Treatment of Kidney Disease Based on Traditional Persian Medicine. Evidence-Based Complementary and Alternative Medicine 2022, 2022, 1–13. [CrossRef]
- Veiga, M.; Costa, E.M.; Silva, S.; Pintado, M. Impact of Plant Extracts upon Human Health: A Review. Critical Reviews in Food Science and Nutrition 2020, 60, 873–886. [CrossRef]
- Panossian, A.G.; Efferth, T.; Shikov, A.N.; Pozharitskaya, O.N.; Kuchta, K.; Mukherjee, P.K.; Banerjee, S.; Heinrich, M.; Wu, W.; Guo, D.; et al. Evolution of the Adaptogenic Concept from Traditional Use to Medical Systems: Pharmacology of Stress- and Aging-related Diseases. Medicinal Research Reviews 2021, 41, 630–703. [CrossRef]
- Chihomvu, P.; Ganesan, A.; Gibbons, S.; Woollard, K.; Hayes, M.A. Phytochemicals in Drug Discovery—A Confluence of Tradition and Innovation. IJMS 2024, 25, 8792. [CrossRef]
- Medicinal Plants and Their Traditional Uses in Different Locations. In Phytomedicine; Elsevier, 2021; pp. 207–223 ISBN 978-0-12-824109-7.
- Dar, R.A.; Shahnawaz, M.; Ahanger, M.A.; Majid, I.U. Exploring the Diverse Bioactive Compounds from Medicinal Plants: A Review. J Phytopharmacol 2023, 12, 189–195. [CrossRef]
- Howes, M.R.; Perry, N.S.L.; Vásquez-Londoño, C.; Perry, E.K. Role of Phytochemicals as Nutraceuticals for Cognitive Functions Affected in Ageing. British J Pharmacology 2020, 177, 1294–1315. [CrossRef]
- George, B.P.; Chandran, R.; Abrahamse, H. Role of Phytochemicals in Cancer Chemoprevention: Insights. Antioxidants 2021, 10, 1455. [CrossRef]
- Albulescu, L.; Suciu, A.; Neagu, M.; Tanase, C.; Pop, S. Differential Biological Effects of Trifolium Pratense Extracts—In Vitro Studies on Breast Cancer Models. Antioxidants 2024, 13, 1435. [CrossRef]
- Liu, R.H. Potential Synergy of Phytochemicals in Cancer Prevention: Mechanism of Action. The Journal of Nutrition 2004, 134, 3479S-3485S. [CrossRef]
- Ionescu, V.S.; Popa, A.; Alexandru, A.; Manole, E.; Neagu, M.; Pop, S. Dietary Phytoestrogens and Their Metabolites as Epigenetic Modulators with Impact on Human Health. Antioxidants 2021, 10, 1893. [CrossRef]
- Alarabei, A.A.; Abd Aziz, N.A.L.; Ab Razak, N.I.; Abas, R.; Bahari, H.; Abdullah, M.A.; Hussain, M.K.; Abdul Majid, A.M.S.; Basir, R. Immunomodulating Phytochemicals: An Insight into Their Potential Use in Cytokine Storm Situations. Adv Pharm Bull 2023. [CrossRef]
- Samanta, S.; Rajasingh, S.; Cao, T.; Dawn, B.; Rajasingh, J. Epigenetic Dysfunctional Diseases and Therapy for Infection and Inflammation. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2017, 1863, 518–528. [CrossRef]
- Tiffon, C. The Impact of Nutrition and Environmental Epigenetics on Human Health and Disease. IJMS 2018, 19, 3425. [CrossRef]
- Gibney, E.R.; Nolan, C.M. Epigenetics and Gene Expression. Heredity 2010, 105, 4–13. [CrossRef]
- Bure, I.V.; Nemtsova, M.V.; Kuznetsova, E.B. Histone Modifications and Non-Coding RNAs: Mutual Epigenetic Regulation and Role in Pathogenesis. IJMS 2022, 23, 5801. [CrossRef]
- The Role of DNA Methylation and Histone Modifications in Transcriptional Regulation in Humans. In Subcellular Biochemistry; Springer Netherlands: Dordrecht, 2013; pp. 289–317 ISBN 978-94-007-4524-7.
- Lorenzo, P.M.; Izquierdo, A.G.; Rodriguez-Carnero, G.; Fernández-Pombo, A.; Iglesias, A.; Carreira, M.C.; Tejera, C.; Bellido, D.; Martinez-Olmos, M.A.; Leis, R.; et al. Epigenetic Effects of Healthy Foods and Lifestyle Habits from the Southern European Atlantic Diet Pattern: A Narrative Review. Advances in Nutrition 2022, 13, 1725–1747. [CrossRef]
- Dai, W.; Qiao, X.; Fang, Y.; Guo, R.; Bai, P.; Liu, S.; Li, T.; Jiang, Y.; Wei, S.; Na, Z.; et al. Epigenetics-Targeted Drugs: Current Paradigms and Future Challenges. Sig Transduct Target Ther 2024, 9. [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res 2011, 21, 381–395. [CrossRef]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic Modifications: Basic Mechanisms and Role in Cardiovascular Disease. Circulation 2011, 123, 2145–2156. [CrossRef]
- Gillette, T.G.; Hill, J.A. Readers, Writers, and Erasers: Chromatin as the Whiteboard of Heart Disease. Circulation Research 2015, 116, 1245–1253. [CrossRef]
- Pop, S.; Enciu, A.M.; Tarcomnicu, I.; Gille, E.; Tanase, C. Phytochemicals in Cancer Prevention: Modulating Epigenetic Alterations of DNA Methylation. Phytochem Rev 2019, 18, 1005–1024. [CrossRef]
- Zhang, X.; Zhang, Y.; Wang, C.; Wang, X. TET (Ten-Eleven Translocation) Family Proteins: Structure, Biological Functions and Applications. Sig Transduct Target Ther 2023, 8. [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacol 2013, 38, 23–38. [CrossRef]
- Mellor, J. Dynamic Nucleosomes and Gene Transcription. Trends in Genetics 2006, 22, 320–329. [CrossRef]
- Becker, P.B.; Workman, J.L. Nucleosome Remodeling and Epigenetics. Cold Spring Harbor Perspectives in Biology 2013, 5, a017905–a017905. [CrossRef]
- Kaikkonen, M.U.; Lam, M.T.Y.; Glass, C.K. Non-Coding RNAs as Regulators of Gene Expression and Epigenetics. Cardiovascular Research 2011, 90, 430–440. [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat Rev Mol Cell Biol 2021, 22, 96–118. [CrossRef]
- Pop, S.; Enciu, A.; Necula, L.G.; Tanase, C. Long Non-coding RNAs in Brain Tumours: Focus on Recent Epigenetic Findings in Glioma. J Cellular Molecular Medi 2018, 22, 4597–4610. [CrossRef]
- Wang, K.; Liu, H.; Hu, Q.; Wang, L.; Liu, J.; Zheng, Z.; Zhang, W.; Ren, J.; Zhu, F.; Liu, G.-H. Epigenetic Regulation of Aging: Implications for Interventions of Aging and Diseases. Sig Transduct Target Ther 2022, 7. [CrossRef]
- Copeland, R.A.; Olhava, E.J.; Scott, M.P. Targeting Epigenetic Enzymes for Drug Discovery. Current Opinion in Chemical Biology 2010, 14, 505–510. [CrossRef]
- Hsieh, H.-H.; Kuo, M.-Z.; Chen, I.-A.; Lin, C.-J.; Hsu, V.; HuangFu, W.-C.; Wu, T.-Y. Epigenetic Modifications as Novel Therapeutic Strategies of Cancer Chemoprevention by Phytochemicals. Pharm Res 2025, 42, 69–78. [CrossRef]
- Casalino, L.; Verde, P. Multifaceted Roles of DNA Methylation in Neoplastic Transformation, from Tumor Suppressors to EMT and Metastasis. Genes 2020, 11, 922. [CrossRef]
- Lee, A.V.; Nestler, K.A.; Chiappinelli, K.B. Therapeutic Targeting of DNA Methylation Alterations in Cancer. Pharmacology & Therapeutics 2024, 258, 108640. [CrossRef]
- Brückmann, N.H.; Pedersen, C.B.; Ditzel, H.J.; Gjerstorff, M.F. Epigenetic Reprogramming of Pericentromeric Satellite DNA in Premalignant and Malignant Lesions. Molecular Cancer Research 2018, 16, 417–427. [CrossRef]
- Sales, V.M.; Ferguson-Smith, A.C.; Patti, M.-E. Epigenetic Mechanisms of Transmission of Metabolic Disease across Generations. Cell Metabolism 2017, 25, 559–571. [CrossRef]
- Zhu, Y.; Wang, X.; Zhou, X.; Ding, L.; Liu, D.; Xu, H. DNMT1-Mediated PPARα Methylation Aggravates Damage of Retinal Tissues in Diabetic Retinopathy Mice. Biol Res 2021, 54. [CrossRef]
- Keleher, M.R.; Zaidi, R.; Hicks, L.; Shah, S.; Xing, X.; Li, D.; Wang, T.; Cheverud, J.M. A High-Fat Diet Alters Genome-Wide DNA Methylation and Gene Expression in SM/J Mice. BMC Genomics 2018, 19. [CrossRef]
- Paul, B.; Barnes, S.; Demark-Wahnefried, W.; Morrow, C.; Salvador, C.; Skibola, C.; Tollefsbol, T.O. Influences of Diet and the Gut Microbiome on Epigenetic Modulation in Cancer and Other Diseases. Clin Epigenet 2015, 7. [CrossRef]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive Oxygen Species, Toxicity, Oxidative Stress, and Antioxidants: Chronic Diseases and Aging. Arch Toxicol 2023, 97, 2499–2574. [CrossRef]
- Khan, M.I.; Rath, S.; Adhami, V.M.; Mukhtar, H. Targeting Epigenome with Dietary Nutrients in Cancer: Current Advances and Future Challenges. Pharmacological Research 2018, 129, 375–387. [CrossRef]
- Aanniz, T.; Bouyahya, A.; Balahbib, A.; El Kadri, K.; Khalid, A.; Makeen, H.A.; Alhazmi, H.A.; El Omari, N.; Zaid, Y.; Wong, R.S.-Y.; et al. Natural Bioactive Compounds Targeting DNA Methyltransferase Enzymes in Cancer: Mechanisms Insights and Efficiencies. Chemico-Biological Interactions 2024, 392, 110907. [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Medicine and Cellular Longevity 2017, 2017. [CrossRef]
- Zheng, X.; Sawalha, A.H. The Role of Oxidative Stress in Epigenetic Changes Underlying Autoimmunity. Antioxidants & Redox Signaling 2022, 36, 423–440. [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free Radical Biology and Medicine 2010, 49, 1603–1616. [CrossRef]
- The Double-Edged Sword Role of ROS in Cancer. In Handbook of Oxidative Stress in Cancer: Mechanistic Aspects; Springer Nature Singapore: Singapore, 2022; pp. 1103–1119 ISBN 978-981-15-9410-6.
- Silva-Llanes, I.; Shin, C.H.; Jiménez-Villegas, J.; Gorospe, M.; Lastres-Becker, I. The Transcription Factor NRF2 Has Epigenetic Regulatory Functions Modulating HDACs, DNMTs, and miRNA Biogenesis. Antioxidants 2023, 12, 641. [CrossRef]
- He, W.-J.; Lv, C.-H.; Chen, Z.; Shi, M.; Zeng, C.-X.; Hou, D.-X.; Qin, S. The Regulatory Effect of Phytochemicals on Chronic Diseases by Targeting Nrf2-ARE Signaling Pathway. Antioxidants 2023, 12, 236. [CrossRef]
- Kietzmann, T.; Petry, A.; Shvetsova, A.; Gerhold, J.M.; Görlach, A. The Epigenetic Landscape Related to Reactive Oxygen Species Formation in the Cardiovascular System. British J Pharmacology 2017, 174, 1533–1554. [CrossRef]
- Madugundu, G.S.; Cadet, J.; Wagner, J.R. Hydroxyl-Radical-Induced Oxidation of 5-Methylcytosine in Isolated and Cellular DNA. Nucleic Acids Research 2014, 42, 7450–7460. [CrossRef]
- Roy, A.; Khan, A.; Ahmad, I.; Alghamdi, S.; Rajab, B.S.; Babalghith, A.O.; Alshahrani, M.Y.; Islam, S.; Islam, Md.R. Flavonoids a Bioactive Compound from Medicinal Plants and Its Therapeutic Applications. BioMed Research International 2022, 2022. [CrossRef]
- Lü, J.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and Molecular Mechanisms of Antioxidants: Experimental Approaches and Model Systems. J Cellular Molecular Medi 2010, 14, 840–860. [CrossRef]
- Andrés, C.M.C.; Pérez De La Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Antioxidant Metabolism Pathways in Vitamins, Polyphenols, and Selenium: Parallels and Divergences. IJMS 2024, 25, 2600. [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxidants & Redox Signaling 2014, 20, 1126–1167. [CrossRef]
- Mucha, P.; Skoczyńska, A.; Małecka, M.; Hikisz, P.; Budzisz, E. Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes. Molecules 2021, 26, 4886. [CrossRef]
- Ngo, V.; Duennwald, M.L. Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants 2022, 11, 2345. [CrossRef]
- Qin, S.; Chen, Z.; Wen, Y.; Yi, Y.; Lv, C.; Zeng, C.; Chen, L.; Shi, M. Phytochemical Activators of Nrf2: A Review of Therapeutic Strategy in Diabetes. ABBS 2022. [CrossRef]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Molecular and Cellular Biology 2020, 40. [CrossRef]
- Jung, K.-A.; Kwak, M.-K. The Nrf2 System as a Potential Target for the Development of Indirect Antioxidants. Molecules 2010, 15, 7266–7291. [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. IJMS 2020, 21, 4777. [CrossRef]
- Carlos-Reyes, Á.; López-González, J.S.; Meneses-Flores, M.; Gallardo-Rincón, D.; Ruíz-García, E.; Marchat, L.A.; Astudillo-de La Vega, H.; Hernández De La Cruz, O.N.; López-Camarillo, C. Dietary Compounds as Epigenetic Modulating Agents in Cancer. Front. Genet. 2019, 10. [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [CrossRef]
- Yasmeen, F.; Pirzada, R.H.; Ahmad, B.; Choi, B.; Choi, S. Understanding Autoimmunity: Mechanisms, Predisposing Factors, and Cytokine Therapies. IJMS 2024, 25, 7666. [CrossRef]
- Zhang, S.; Meng, Y.; Zhou, L.; Qiu, L.; Wang, H.; Su, D.; Zhang, B.; Chan, K.; Han, J. Targeting Epigenetic Regulators for Inflammation: Mechanisms and Intervention Therapy. MedComm 2022, 3. [CrossRef]
- Cronkite, D.A.; Strutt, T.M. The Regulation of Inflammation by Innate and Adaptive Lymphocytes. Journal of Immunology Research 2018, 2018, 1–14. [CrossRef]
- Zhu, X.; Chen, Z.; Shen, W.; Huang, G.; Sedivy, J.M.; Wang, H.; Ju, Z. Inflammation, Epigenetics, and Metabolism Converge to Cell Senescence and Ageing: The Regulation and Intervention. Sig Transduct Target Ther 2021, 6. [CrossRef]
- Wu, D.; Shi, Y.; Zhang, H.; Miao, C. Epigenetic Mechanisms of Immune Remodeling in Sepsis: Targeting Histone Modification. Cell Death Dis 2023, 14. [CrossRef]
- Kaszycki, J.; Kim, M. Epigenetic Regulation of Transcription Factors Involved in NLRP3 Inflammasome and NF-kB Signaling Pathways. Front. Immunol. 2025, 16. [CrossRef]
- Jurkowska, R.Z. Role of Epigenetic Mechanisms in the Pathogenesis of Chronic Respiratory Diseases and Response to Inhaled Exposures: From Basic Concepts to Clinical Applications. Pharmacology & Therapeutics 2024, 264, 108732. [CrossRef]
- Vezzani, B.; Carinci, M.; Previati, M.; Giacovazzi, S.; Della Sala, M.; Gafà, R.; Lanza, G.; Wieckowski, M.R.; Pinton, P.; Giorgi, C. Epigenetic Regulation: A Link between Inflammation and Carcinogenesis. Cancers (Basel) 2022, 14, 1221. [CrossRef]
- Tan, S.Y.X.; Zhang, J.; Tee, W.-W. Epigenetic Regulation of Inflammatory Signaling and Inflammation-Induced Cancer. Front Cell Dev Biol 2022, 10, 931493. [CrossRef]
- Yang, Z.-H.; Dang, Y.-Q.; Ji, G. Role of Epigenetics in Transformation of Inflammation into Colorectal Cancer. World J Gastroenterol 2019, 25, 2863–2877. [CrossRef]
- Pandareesh, M.D.; Kameshwar, V.H.; Byrappa, K. Prostate Carcinogenesis: Insights in Relation to Epigenetics and Inflammation. Endocr Metab Immune Disord Drug Targets 2021, 21, 253–267. [CrossRef]
- Rostami, A.; White, K.; Rostami, K. Pro and Anti-Inflammatory Diets as Strong Epigenetic Factors in Inflammatory Bowel Disease. World J Gastroenterol 2024, 30, 3284–3289. [CrossRef]
- Das, D.; Karthik, N.; Taneja, R. Crosstalk Between Inflammatory Signaling and Methylation in Cancer. Front. Cell Dev. Biol. 2021, 9. [CrossRef]
- Bagni, G.; Biancalana, E.; Chiara, E.; Costanzo, I.; Malandrino, D.; Lastraioli, E.; Palmerini, M.; Silvestri, E.; Urban, M.L.; Emmi, G. Epigenetics in Autoimmune Diseases: Unraveling the Hidden Regulators of Immune Dysregulation. Autoimmunity Reviews 2025, 24, 103784. [CrossRef]
- Liu, L.; Ni, S.; Zhang, L.; Chen, Y.; Xie, M.; Huang, X. Molecular Insights and Clinical Implications of DNA Methylation in Sepsis-Associated Acute Kidney Injury: A Narrative Review. BMC Nephrol 2025, 26. [CrossRef]
- Bacher, S.; Meier-Soelch, J.; Kracht, M.; Schmitz, M.L. Regulation of Transcription Factor NF-κB in Its Natural Habitat: The Nucleus. Cells 2021, 10, 753. [CrossRef]
- Grabiec, A.M.; Tak, P.P.; Reedquist, K.A. Targeting Histone Deacetylase Activity in Rheumatoid Arthritis and Asthma as Prototypes of Inflammatory Disease: Should We Keep Our HATs On? Arthritis Res Ther 2008, 10, 226. [CrossRef]
- Li, C.; Gu, S.; Zhang, Y.; Zhang, Z.; Wang, J.; Gao, T.; Zhong, K.; Shan, K.; Ye, G.; Ke, Y.; et al. Histone Deacetylase in Inflammatory Bowel Disease: Novel Insights. Therap Adv Gastroenterol 2025, 18. [CrossRef]
- Raisch, J. Role of microRNAs in the Immune System, Inflammation and Cancer. WJG 2013, 19, 2985. [CrossRef]
- Alivernini, S.; Gremese, E.; McSharry, C.; Tolusso, B.; Ferraccioli, G.; McInnes, I.B.; Kurowska-Stolarska, M. MicroRNA-155—at the Critical Interface of Innate and Adaptive Immunity in Arthritis. Front. Immunol. 2018, 8. [CrossRef]
- Slaughter, M.J.; Shanle, E.K.; Khan, A.; Chua, K.F.; Hong, T.; Boxer, L.D.; Allis, C.D.; Josefowicz, S.Z.; Garcia, B.A.; Rothbart, S.B.; et al. HDAC Inhibition Results in Widespread Alteration of the Histone Acetylation Landscape and BRD4 Targeting to Gene Bodies. Cell Reports 2021, 34, 108638. [CrossRef]
- Phimmachanh, M.; Han, J.Z.R.; O’Donnell, Y.E.I.; Latham, S.L.; Croucher, D.R. Histone Deacetylases and Histone Deacetylase Inhibitors in Neuroblastoma. Front. Cell Dev. Biol. 2020, 8. [CrossRef]
- Al Bitar, S.; Gali-Muhtasib, H. The Role of the Cyclin Dependent Kinase Inhibitor P21cip1/Waf1 in Targeting Cancer: Molecular Mechanisms and Novel Therapeutics. Cancers 2019, 11, 1475. [CrossRef]
- Sturmlechner, I.; Zhang, C.; Sine, C.C.; Van Deursen, E.-J.; Jeganathan, K.B.; Hamada, N.; Grasic, J.; Friedman, D.; Stutchman, J.T.; Can, I.; et al. P21 Produces a Bioactive Secretome That Places Stressed Cells under Immunosurveillance. Science 2021, 374. [CrossRef]
- Arora, I.; Sharma, M.; Sun, L.Y.; Tollefsbol, T.O. The Epigenetic Link between Polyphenols, Aging and Age-Related Diseases. Genes 2020, 11, 1094. [CrossRef]
- Perlmutter, A.; Bland, J.S.; Chandra, A.; Malani, S.S.; Smith, R.; Mendez, T.L.; Dwaraka, V.B. The Impact of a Polyphenol-Rich Supplement on Epigenetic and Cellular Markers of Immune Age: A Pilot Clinical Study. Front. Nutr. 2024, 11. [CrossRef]
- Bergonzini, M.; Loreni, F.; Lio, A.; Russo, M.; Saitto, G.; Cammardella, A.; Irace, F.; Tramontin, C.; Chello, M.; Lusini, M.; et al. Panoramic on Epigenetics in Coronary Artery Disease and the Approach of Personalized Medicine. Biomedicines 2023, 11, 2864. [CrossRef]
- Dong, Z.; Hou, L.; Luo, W.; Pan, L.-H.; Li, X.; Tan, H.-P.; Wu, R.-D.; Lu, H.; Yao, K.; Mu, M.-D.; et al. Myocardial Infarction Drives Trained Immunity of Monocytes, Accelerating Atherosclerosis. Eur Heart J 2024, 45, 669–684. [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic Inflammation in Ageing, Cardiovascular Disease, and Frailty. Nat Rev Cardiol 2018, 15, 505–522. [CrossRef]
- Marsit, C.J. Influence of Environmental Exposure on Human Epigenetic Regulation. Journal of Experimental Biology 2015, 218, 71–79. [CrossRef]
- Begum, N.; Mandhare, A.; Tryphena, K.P.; Srivastava, S.; Shaikh, M.F.; Singh, S.B.; Khatri, D.K. Epigenetics in Depression and Gut-Brain Axis: A Molecular Crosstalk. Front. Aging Neurosci. 2022, 14. [CrossRef]
- Zhang, Q.; Liu, Y.; Li, Y.; Bai, G.; Pang, J.; Wu, M.; Li, J.; Zhao, X.; Xia, Y. Implications of Gut Microbiota-Mediated Epigenetic Modifications in Intestinal Diseases. Gut Microbes 2025, 17. [CrossRef]
- Liu, X.; Shao, J.; Liao, Y.-T.; Wang, L.-N.; Jia, Y.; Dong, P.; Liu, Z.; He, D.; Li, C.; Zhang, X. Regulation of Short-Chain Fatty Acids in the Immune System. Front. Immunol. 2023, 14. [CrossRef]
- Ji, H.; Khurana Hershey, G.K. Genetic and Epigenetic Influence on the Response to Environmental Particulate Matter. Journal of Allergy and Clinical Immunology 2012, 129, 33–41. [CrossRef]
- Rubio, K.; Hernández-Cruz, E.Y.; Rogel-Ayala, D.G.; Sarvari, P.; Isidoro, C.; Barreto, G.; Pedraza-Chaverri, J. Nutriepigenomics in Environmental-Associated Oxidative Stress. Antioxidants 2023, 12, 771. [CrossRef]
- Erratum. ILAR Journal 2017, 58, 413–413. [CrossRef]
- Breton, C.V.; Landon, R.; Kahn, L.G.; Enlow, M.B.; Peterson, A.K.; Bastain, T.; Braun, J.; Comstock, S.S.; Duarte, C.S.; Hipwell, A.; et al. Exploring the Evidence for Epigenetic Regulation of Environmental Influences on Child Health across Generations. Commun Biol 2021, 4. [CrossRef]
- Alegría-Torres, J.A.; Baccarelli, A.; Bollati, V. Epigenetics and Lifestyle. Epigenomics 2011, 3, 267–277. [CrossRef]
- Barouki, R.; Melén, E.; Herceg, Z.; Beckers, J.; Chen, J.; Karagas, M.; Puga, A.; Xia, Y.; Chadwick, L.; Yan, W.; et al. Epigenetics as a Mechanism Linking Developmental Exposures to Long-Term Toxicity. Environment International 2018, 114, 77–86. [CrossRef]
- Wu, X.; Wu, H.; Zhong, M.; Chen, Y.; Su, W.; Li, P. Epigenetic Regulation by Naringenin and Naringin: A Literature Review Focused on the Mechanisms Underlying Its Pharmacological Effects. Fitoterapia 2025, 181, 106353. [CrossRef]
- Wang, G.; Su, H.; Guo, Z.; Li, H.; Jiang, Z.; Cao, Y.; Li, C. Rubus Occidentalis and Its Bioactive Compounds against Cancer: From Molecular Mechanisms to Translational Advances. Phytomedicine 2024, 126, 155029. [CrossRef]
- Babar, Q.; Saeed, A.; Tabish, T.A.; Pricl, S.; Townley, H.; Thorat, N. Novel Epigenetic Therapeutic Strategies and Targets in Cancer. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2022, 1868, 166552. [CrossRef]
- Lagosz-Cwik, K.B.; Melnykova, M.; Nieboga, E.; Schuster, A.; Bysiek, A.; Dudek, S.; Lipska, W.; Kantorowicz, M.; Tyrakowski, M.; Darczuk, D.; et al. Mapping of DNA Methylation-Sensitive Cellular Processes in Gingival and Periodontal Ligament Fibroblasts in the Context of Periodontal Tissue Homeostasis. Front. Immunol. 2023, 14. [CrossRef]
- Nisar, A.; Jagtap, S.; Vyavahare, S.; Deshpande, M.; Harsulkar, A.; Ranjekar, P.; Prakash, O. Phytochemicals in the Treatment of Inflammation-Associated Diseases: The Journey from Preclinical Trials to Clinical Practice. Front. Pharmacol. 2023, 14. [CrossRef]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jönsson, L.S.; Kolb, H.; Lansink, M.; et al. Dietary Factors and Low-Grade Inflammation in Relation to Overweight and Obesity. Br J Nutr 2011, 106, S1–S78. [CrossRef]
- González-Gallego, J.; García-Mediavilla, M.V.; Sánchez-Campos, S.; Tuñón, M.J. Fruit Polyphenols, Immunity and Inflammation. Br J Nutr 2010, 104, S15–S27. [CrossRef]
- Hossain, Md.S.; Wazed, M.A.; Asha, S.; Amin, Md.R.; Shimul, I.M. Dietary Phytochemicals in Health and Disease: Mechanisms, Clinical Evidence, and Applications—A Comprehensive Review. Food Science & Nutrition 2025, 13. [CrossRef]
- Hasnat, H.; Shompa, S.A.; Islam, Md.M.; Alam, S.; Richi, F.T.; Emon, N.U.; Ashrafi, S.; Ahmed, N.U.; Chowdhury, Md.N.R.; Fatema, N.; et al. Flavonoids: A Treasure House of Prospective Pharmacological Potentials. Heliyon 2024, 10, e27533. [CrossRef]
- Fahmy, M.I.; Sadek, M.A.; Abdou, K.; El-Dessouki, A.M.; El-Shiekh, R.A.; Khalaf, S.S. Orientin: A Comprehensive Review of a Promising Bioactive Flavonoid. Inflammopharmacol 2025, 33, 1713–1728. [CrossRef]
- Caban, M.; Lewandowska, U. Polyphenols and the Potential Mechanisms of Their Therapeutic Benefits against Inflammatory Bowel Diseases. Journal of Functional Foods 2022, 95, 105181. [CrossRef]
- Liga, S.; Paul, C.; Péter, F. Flavonoids: Overview of Biosynthesis, Biological Activity, and Current Extraction Techniques. Plants 2023, 12, 2732. [CrossRef]
- Masenga, S.K.; Kabwe, L.S.; Chakulya, M.; Kirabo, A. Mechanisms of Oxidative Stress in Metabolic Syndrome. IJMS 2023, 24, 7898. [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; Gonzalez De Mejia, E. Phenolic Compounds from Coffee By-Products Modulate Adipogenesis-Related Inflammation, Mitochondrial Dysfunction, and Insulin Resistance in Adipocytes, via Insulin/PI3K/AKT Signaling Pathways. Food and Chemical Toxicology 2019, 132, 110672. [CrossRef]
- Li, Q.; Tanaka, Y.; Saitoh, Y.; Tanaka, H.; Miwa, N. Carcinostatic Effects of Platinum Nanocolloid Combined with Gamma Irradiation on Human Esophageal Squamous Cell Carcinoma. Life Sciences 2015, 127, 106–114. [CrossRef]
- Panieri, E.; Pinho, S.A.; Afonso, G.J.M.; Oliveira, P.J.; Cunha-Oliveira, T.; Saso, L. NRF2 and Mitochondrial Function in Cancer and Cancer Stem Cells. Cells 2022, 11, 2401. [CrossRef]
- Eckschlager, T.; Plch, J.; Stiborova, M.; Hrabeta, J. Histone Deacetylase Inhibitors as Anticancer Drugs. Int J Mol Sci 2017, 18, 1414. [CrossRef]
- Wu, B.; Tapadar, S.; Ruan, Z.; Sun, C.Q.; Arnold, R.S.; Johnston, A.; Olugbami, J.O.; Arunsi, U.; Gaul, D.A.; Petros, J.A.; et al. A Novel Liver Cancer-Selective Histone Deacetylase Inhibitor Is Effective Against Hepatocellular Carcinoma and Induces Durable Responses with Immunotherapy 2024.
- Liu, Z.; Huang, M.; Hong, Y.; Wang, S.; Xu, Y.; Zhong, C.; Zhang, J.; Zhuang, Z.; Shan, S.; Ren, T. Isovalerylspiramycin I Suppresses Non-Small Cell Lung Carcinoma Growth through ROS-Mediated Inhibition of PI3K/AKT Signaling Pathway. Int. J. Biol. Sci. 2022, 18, 3714–3730. [CrossRef]
- Record, I.R.; Dreosti, I.E.; McInerney, J.K. The Antioxidant Activity of Genistein in Vitro. The Journal of Nutritional Biochemistry 1995, 6, 481–485. [CrossRef]
- Kim, D.H.; Jung, W.-S.; Kim, M.E.; Lee, H.-W.; Youn, H.-Y.; Seon, J.K.; Lee, H.-N.; Lee, J.S. Genistein Inhibits Pro-Inflammatory Cytokines in Human Mast Cell Activation through the Inhibition of the ERK Pathway. International Journal of Molecular Medicine 2014, 34, 1669–1674. [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB Signaling in Inflammation. Sig Transduct Target Ther 2017, 2. [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10. [CrossRef]
- Zhao, Z.; Shin, H.S.; Satsu, H.; Totsuka, M.; Shimizu, M. 5-Caffeoylquinic Acid and Caffeic Acid Down-Regulate the Oxidative Stress- and TNF-α-Induced Secretion of Interleukin-8 from Caco-2 Cells. J. Agric. Food Chem. 2008, 56, 3863–3868. [CrossRef]
- Tyszka-Czochara, M.; Bukowska-Strakova, K.; Kocemba-Pilarczyk, K.A.; Majka, M. Caffeic Acid Targets AMPK Signaling and Regulates Tricarboxylic Acid Cycle Anaplerosis While Metformin Downregulates HIF-1α-Induced Glycolytic Enzymes in Human Cervical Squamous Cell Carcinoma Lines. Nutrients 2018, 10, 841. [CrossRef]
- Xia, L.; Zhu, G.; Peng, Q.; Li, X.; Zou, X.; Zhang, W.; Zhao, L.; Li, X.; Wu, P.; Luo, A.; et al. Natural Products Combating EGFR-TKIs Resistance in Cancer. European Journal of Medicinal Chemistry Reports 2025, 13, 100251. [CrossRef]
- Zhao, Y.; Liu, J. Anti-Inflammatory Effects of p-Coumaric Acid in LPS-Stimulated RAW264.7 Cells: Involvement of NF-κB and MAPKs Pathways. Med chem (Los Angeles) 2016, 06. [CrossRef]
- Wu, C.; Sun, C.; Han, X.; Ye, Y.; Qin, Y.; Liu, S. Sanyin Formula Enhances the Therapeutic Efficacy of Paclitaxel in Triple-Negative Breast Cancer Metastases through the JAK/STAT3 Pathway in Mice. Pharmaceuticals 2022, 16, 9. [CrossRef]
- Zhu, Y.; Yue, P.; Dickinson, C.F.; Yang, J.K.; Datanagan, K.; Zhai, N.; Zhang, Y.; Miklossy, G.; Lopez-Tapia, F.; Tius, M.A.; et al. Natural Product Preferentially Targets Redox and Metabolic Adaptations and Aberrantly Active STAT3 to Inhibit Breast Tumor Growth in Vivo. Cell Death Dis 2022, 13. [CrossRef]
- Bouyahya, A.; El Hachlafi, N.; Aanniz, T.; Bourais, I.; Mechchate, H.; Benali, T.; Shariati, M.A.; Burkov, P.; Lorenzo, J.M.; Wilairatana, P.; et al. Natural Bioactive Compounds Targeting Histone Deacetylases in Human Cancers: Recent Updates. Molecules 2022, 27, 2568. [CrossRef]
- Neelab; Zeb, A.; Jamil, M. Milk Thistle Protects against Non-Alcoholic Fatty Liver Disease Induced by Dietary Thermally Oxidized Tallow. Heliyon 2024, 10, e31445. [CrossRef]
- Surai, P. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [CrossRef]
- Sugihara, A.; Nguyen, L.C.; Shamim, H.M.; Iida, T.; Nakase, M.; Takegawa, K.; Senda, M.; Jida, S.; Ueno, M. Mutation in Fission Yeast Phosphatidylinositol 4-Kinase Pik1 Is Synthetically Lethal with Defect in Telomere Protection Protein Pot1. Biochemical and Biophysical Research Communications 2018, 496, 1284–1290. [CrossRef]
- Baş, H.; Kalender, Y.; Pandir, D.; Kalender, S. Effects of Lead Nitrate and Sodium Selenite on DNA Damage and Oxidative Stress in Diabetic and Non-Diabetic Rat Erythrocytes and Leucocytes. Environmental Toxicology and Pharmacology 2015, 39, 1019–1026. [CrossRef]
- Zhang, M.; Pan, H.; Xu, Y.; Wang, X.; Qiu, Z.; Jiang, L. Allicin Decreases Lipopolysaccharide-Induced Oxidative Stress and Inflammation in Human Umbilical Vein Endothelial Cells through Suppression of Mitochondrial Dysfunction and Activation of Nrf2. Cell Physiol Biochem 2017, 41, 2255–2267. [CrossRef]
- Meyer, K.; Ueberham, E.; Gebhardt, R. Influence of Organosulphur Compounds from Garlic on the Secretion of Matrix Metalloproteinases and Their Inhibitor TIMP-1 by Cultured HUVEC Cells. Cell Biol Toxicol 2004, 20, 253–260. [CrossRef]
- Nian, H.; Delage, B.; Pinto, J.T.; Dashwood, R.H. Allyl Mercaptan, a Garlic-Derived Organosulfur Compound, Inhibits Histone Deacetylase and Enhances Sp3 Binding on the P21WAF1 Promoter. Carcinogenesis 2008, 29, 1816–1824. [CrossRef]
- Astrain-Redin, N.; Sanmartin, C.; Sharma, A.K.; Plano, D. From Natural Sources to Synthetic Derivatives: The Allyl Motif as a Powerful Tool for Fragment-Based Design in Cancer Treatment. J. Med. Chem. 2023, 66, 3703–3731. [CrossRef]
- Addi, M.; Elbouzidi, A.; Abid, M.; Tungmunnithum, D.; Elamrani, A.; Hano, C. An Overview of Bioactive Flavonoids from Citrus Fruits. Applied Sciences 2021, 12, 29. [CrossRef]
- Chen, M.; Gu, H.; Ye, Y.; Lin, B.; Sun, L.; Deng, W.; Zhang, J.; Liu, J. Protective Effects of Hesperidin against Oxidative Stress of Tert-Butyl Hydroperoxide in Human Hepatocytes. Food and Chemical Toxicology 2010, 48, 2980–2987. [CrossRef]
- Rahmani, A.H.; Babiker, A.Y.; Anwar, S. Hesperidin, a Bioflavonoid in Cancer Therapy: A Review for a Mechanism of Action through the Modulation of Cell Signaling Pathways. Molecules 2023, 28, 5152. [CrossRef]
- Tejada, S.; Pinya, S.; Martorell, M.; Capó, X.; Tur, J.A.; Pons, A.; Sureda, A. Potential Anti-Inflammatory Effects of Hesperidin from the Genus Citrus. CMC 2019, 25, 4929–4945. [CrossRef]
- Li, C.; Schluesener, H. Health-Promoting Effects of the Citrus Flavanone Hesperidin. Critical Reviews in Food Science and Nutrition 2017, 57, 613–631. [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [CrossRef]
- Fernández-Bedmar, Z.; Anter, J.; Alonso-Moraga, A.; Martín De Las Mulas, J.; Millán-Ruiz, Y.; Guil-Luna, S. Demethylating and Anti-hepatocarcinogenic Potential of Hesperidin, a Natural Polyphenol of Citrus Juices. Molecular Carcinogenesis 2017, 56, 1653–1662. [CrossRef]
- Jiang, W.; Xia, T.; Liu, C.; Li, J.; Zhang, W.; Sun, C. Remodeling the Epigenetic Landscape of Cancer—Application Potential of Flavonoids in the Prevention and Treatment of Cancer. Front. Oncol. 2021, 11. [CrossRef]
- Dixon, R. Genistein. Phytochemistry 2002, 60, 205–211. [CrossRef]
- Setchell, K.D.R.; Cassidy, A. Dietary Isoflavones: Biological Effects and Relevance to Human Health. The Journal of Nutrition 1999, 129, 758S-767S. [CrossRef]
- Goh, Y.X.; Jalil, J.; Lam, K.W.; Husain, K.; Premakumar, C.M. Genistein: A Review on Its Anti-Inflammatory Properties. Front. Pharmacol. 2022, 13. [CrossRef]
- Valsecchi, A.E.; Franchi, S.; Panerai, A.E.; Rossi, A.; Sacerdote, P.; Colleoni, M. The Soy Isoflavone Genistein Reverses Oxidative and Inflammatory State, Neuropathic Pain, Neurotrophic and Vasculature Deficits in Diabetes Mouse Model. European Journal of Pharmacology 2011, 650, 694–702. [CrossRef]
- Zhou, X.; Yuan, L.; Zhao, X.; Hou, C.; Ma, W.; Yu, H.; Xiao, R. Genistein Antagonizes Inflammatory Damage Induced by β-Amyloid Peptide in Microglia through TLR4 and NF-κB. Nutrition 2014, 30, 90–95. [CrossRef]
- Smolińska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Węgrzyn, G.; Banecki, B.; Szczerkowska-Dobosz, A.; Purzycka-Bohdan, D.; Gabig-Cimińska, M. Molecular Action of Isoflavone Genistein in the Human Epithelial Cell Line HaCaT. PLoS ONE 2018, 13, e0192297. [CrossRef]
- Hämäläinen, M.; Nieminen, R.; Asmawi, M.; Vuorela, P.; Vapaatalo, H.; Moilanen, E. Effects of Flavonoids on Prostaglandin E2Production and on COX-2 and mPGES-1 Expressions in Activated Macrophages. Planta Med 2011, 77, 1504–1511. [CrossRef]
- Bilir, B.; Sharma, N.V.; Lee, J.; Hammarstrom, B.; Svindland, A.; Kucuk, O.; Moreno, C.S. Effects of Genistein Supplementation on Genome-Wide DNA Methylation and Gene Expression in Patients with Localized Prostate Cancer. International Journal of Oncology 2017, 51, 223–234. [CrossRef]
- Ko, K.-P.; Kim, S.-W.; Ma, S.H.; Park, B.; Ahn, Y.; Lee, J.W.; Lee, M.H.; Kang, E.; Kim, L.S.; Jung, Y.; et al. Dietary Intake and Breast Cancer among Carriers and Noncarriers of BRCA Mutations in the Korean Hereditary Breast Cancer Study. The American Journal of Clinical Nutrition 2013, 98, 1493–1501. [CrossRef]
- Lu, H.; Shi, J.-X.; Zhang, D.-M.; Wang, H.-D.; Hang, C.-H.; Chen, H.-L.; Yin, H.-X. Inhibition of Hemolysate-Induced iNOS and COX-2 Expression by Genistein through Suppression of NF-кB Activation in Primary Astrocytes. Journal of the Neurological Sciences 2009, 278, 91–95. [CrossRef]
- Habtemariam, S. The Molecular Pharmacology of Phloretin: Anti-Inflammatory Mechanisms of Action. Biomedicines 2023, 11, 143. [CrossRef]
- Hao, X.; Bai, Y.; Li, W.; Zhang, M.X. Phloretin Attenuates Inflammation Induced by Subarachnoid Hemorrhage through Regulation of the TLR2/MyD88/NF-kB Pathway. Sci Rep 2024, 14. [CrossRef]
- Lee, S.-W.; Lee, G.; Jo, J.-H.; Yang, Y.; Ahn, J.-H. Biosynthesis of Phloretin and Its C-Glycosides through Stepwise Culture of Escherichia Coli. Appl Biol Chem 2024, 67. [CrossRef]
- Chang, W.-T.; Huang, W.-C.; Liou, C.-J. Evaluation of the Anti-Inflammatory Effects of Phloretin and Phlorizin in Lipopolysaccharide-Stimulated Mouse Macrophages. Food Chemistry 2012, 134, 972–979. [CrossRef]
- Mansuri, M.L.; Parihar, P.; Solanki, I.; Parihar, M.S. Flavonoids in Modulation of Cell Survival Signalling Pathways. Genes Nutr 2014, 9. [CrossRef]
- Bin-Jumah, M.N.; Nadeem, M.S.; Gilani, S.J.; Mubeen, B.; Ullah, I.; Alzarea, S.I.; Ghoneim, M.M.; Alshehri, S.; Al-Abbasi, F.A.; Kazmi, I. Lycopene: A Natural Arsenal in the War against Oxidative Stress and Cardiovascular Diseases. Antioxidants 2022, 11, 232. [CrossRef]
- Imran, M.; Ghorat, F.; Ul-Haq, I.; Ur-Rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M.; et al. Lycopene as a Natural Antioxidant Used to Prevent Human Health Disorders. Antioxidants 2020, 9, 706. [CrossRef]
- Kulawik, A.; Cielecka-Piontek, J.; Czerny, B.; Kamiński, A.; Zalewski, P. The Relationship Between Lycopene and Metabolic Diseases. Nutrients 2024, 16, 3708. [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, Md.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12. [CrossRef]
- Birková, A. Caffeic Acid: A Brief Overview of Its Presence, Metabolism, and Bioactivity. Bioactive Compounds in Health and Disease 2020, 3, 74. [CrossRef]
- Pavlíková, N. Caffeic Acid and Diseases—Mechanisms of Action. IJMS 2022, 24, 588. [CrossRef]
- Ku, H.-C.; Lee, S.-Y.; Yang, K.-C.; Kuo, Y.-H.; Su, M.-J. Modification of Caffeic Acid with Pyrrolidine Enhances Antioxidant Ability by Activating AKT/HO-1 Pathway in Heart. PLoS ONE 2016, 11, e0148545. [CrossRef]
- Yao, J.; Liu, Y.; Lin, H.; Shao, C.; Jin, X.; Peng, T.; Liu, Y. Caffeic Acid Activates Nrf2 Enzymes, Providing Protection against Oxidative Damage Induced by Ionizing Radiation. Brain Research Bulletin 2025, 224, 111325. [CrossRef]
- Zielińska, D.; Zieliński, H.; Laparra-Llopis, J.M.; Szawara-Nowak, D.; Honke, J.; Giménez-Bastida, J.A. Caffeic Acid Modulates Processes Associated with Intestinal Inflammation. Nutrients 2021, 13, 554. [CrossRef]
- Yin, Y.; Wang, Z.; Hu, Y.; Wang, J.; Wang, Y.; Lu, Q. Caffeic Acid Hinders the Proliferation and Migration through Inhibition of IL-6 Mediated JAK-STAT-3 Signaling Axis in Human Prostate Cancer. OR 2024, 32, 1881–1890. [CrossRef]
- Shin, K.-M.; Kim, I.-T.; Park, Y.-M.; Ha, J.; Choi, J.-W.; Park, H.-J.; Lee, Y.S.; Lee, K.-T. Anti-Inflammatory Effect of Caffeic Acid Methyl Ester and Its Mode of Action through the Inhibition of Prostaglandin E2, Nitric Oxide and Tumor Necrosis Factor-α Production. Biochemical Pharmacology 2004, 68, 2327–2336. [CrossRef]
- Cortez, N.; Villegas, C.; Burgos, V.; Cabrera-Pardo, J.R.; Ortiz, L.; González-Chavarría, I.; Nchiozem-Ngnitedem, V.-A.; Paz, C. Adjuvant Properties of Caffeic Acid in Cancer Treatment. IJMS 2024, 25, 7631. [CrossRef]
- Park, J.-Y.; Yasir, M.; Lee, H.; Han, E.-T.; Han, J.-H.; Park, W.; Kwon, Y.-S.; Chun, W. Caffeic Acid Methyl Ester Inhibits LPS-induced Inflammatory Response through Nrf2 Activation and NF-κB Inhibition in Human Umbilical Vein Endothelial Cells. Exp Ther Med 2023, 26. [CrossRef]
- Wan, F.; Zhong, R.; Wang, M.; Zhou, Y.; Chen, Y.; Yi, B.; Hou, F.; Liu, L.; Zhao, Y.; Chen, L.; et al. Caffeic Acid Supplement Alleviates Colonic Inflammation and Oxidative Stress Potentially Through Improved Gut Microbiota Community in Mice. Front. Microbiol. 2021, 12. [CrossRef]
- Ding, Q.; Xu, Y.-M.; Lau, A.T.Y. The Epigenetic Effects of Coffee. Molecules 2023, 28, 1770. [CrossRef]
- Contardi, M.; Lenzuni, M.; Fiorentini, F.; Summa, M.; Bertorelli, R.; Suarato, G.; Athanassiou, A. Hydroxycinnamic Acids and Derivatives Formulations for Skin Damages and Disorders: A Review. Pharmaceutics 2021, 13, 999. [CrossRef]
- Xia, T.; Zhang, B.; Duan, W.; Zhang, J.; Wang, M. Nutrients and Bioactive Components from Vinegar: A Fermented and Functional Food. Journal of Functional Foods 2020, 64, 103681. [CrossRef]
- Jeong, Y.H.; Yang, H.J.; Li, W.; Oh, Y.-C.; Choi, J.-G. Immune-Enhancing Effects of Gwakhyangjeonggi-San in RAW 264.7 Macrophage Cells through the MAPK/NF-κB Signaling Pathways. IJMS 2024, 25, 9246. [CrossRef]
- Moustardas, P.; Aberdam, D.; Lagali, N. MAPK Pathways in Ocular Pathophysiology: Potential Therapeutic Drugs and Challenges. Cells 2023, 12, 617. [CrossRef]
- Moon, H.-R.; Yun, J.-M. P-Coumaric Acid Modulates Cholesterol Efflux and Lipid Accumulation and Inflammation in Foam Cells. Nutr Res Pract 2024, 18, 774. [CrossRef]
- Medeiros-Linard, C.F.B.; Andrade-da-Costa, B.L.D.S.; Augusto, R.L.; Sereniki, A.; Trevisan, M.T.S.; Perreira, R.D.C.R.; De Souza, F.T.C.; Braz, G.R.F.; Lagranha, C.J.; De Souza, I.A.; et al. Anacardic Acids from Cashew Nuts Prevent Behavioral Changes and Oxidative Stress Induced by Rotenone in a Rat Model of Parkinson’s Disease. Neurotox Res 2018, 34, 250–262. [CrossRef]
- Augusto, R.L.; Mendonça, I.P.; De Albuquerque Rego, G.N.; Pereira, D.D.; Da Penha Gonçalves, L.V.; Dos Santos, M.L.; De Souza, R.F.; Moreno, G.M.M.; Cardoso, P.R.G.; De Souza Andrade, D.; et al. Purified Anacardic Acids Exert Multiple Neuroprotective Effects in Pesticide Model of Parkinson’s Disease: In Vivo and in Silico Analysis. IUBMB Life 2020, 72, 1765–1779. [CrossRef]
- Tomasiak, P.; Janisiak, J.; Rogińska, D.; Perużyńska, M.; Machaliński, B.; Tarnowski, M. Garcinol and Anacardic Acid, Natural Inhibitors of Histone Acetyltransferases, Inhibit Rhabdomyosarcoma Growth and Proliferation. Molecules 2023, 28, 5292. [CrossRef]
- Rais, N.; Ved, A.; Ahmad, R.; Kumar, M.; Deepak Barbhai, M.; Radha; Chandran, D.; Dey, A.; Dhumal, S.; Senapathy, M.; et al. S-Allyl-L-Cysteine — A Garlic Bioactive: Physicochemical Nature, Mechanism, Pharmacokinetics, and Health Promoting Activities. Journal of Functional Foods 2023, 107, 105657. [CrossRef]
- Arreola, R.; Quintero-Fabián, S.; López-Roa, R.I.; Flores-Gutiérrez, E.O.; Reyes-Grajeda, J.P.; Carrera-Quintanar, L.; Ortuño-Sahagún, D. Immunomodulation and Anti-Inflammatory Effects of Garlic Compounds. Journal of Immunology Research 2015, 2015, 1–13. [CrossRef]
- Sasi, M.; Kumar, S.; Kumar, M.; Thapa, S.; Prajapati, U.; Tak, Y.; Changan, S.; Saurabh, V.; Kumari, S.; Kumar, A.; et al. Garlic (Allium Sativum L.) Bioactives and Its Role in Alleviating Oral Pathologies. Antioxidants 2021, 10, 1847. [CrossRef]
- Ansary, J.; Forbes-Hernández, T.Y.; Gil, E.; Cianciosi, D.; Zhang, J.; Elexpuru-Zabaleta, M.; Simal-Gandara, J.; Giampieri, F.; Battino, M. Potential Health Benefit of Garlic Based on Human Intervention Studies: A Brief Overview. Antioxidants 2020, 9, 619. [CrossRef]
- Wadhwa, K.; Pahwa, R.; Kumar, M.; Kumar, S.; Sharma, P.C.; Singh, G.; Verma, R.; Mittal, V.; Singh, I.; Kaushik, D.; et al. Mechanistic Insights into the Pharmacological Significance of Silymarin. Molecules 2022, 27, 5327. [CrossRef]
- Vargas-Mendoza, N. Hepatoprotective Effect of Silymarin. WJH 2014, 6, 144. [CrossRef]
- Lovelace, E.S.; Wagoner, J.; MacDonald, J.; Bammler, T.; Bruckner, J.; Brownell, J.; Beyer, R.P.; Zink, E.M.; Kim, Y.-M.; Kyle, J.E.; et al. Silymarin Suppresses Cellular Inflammation By Inducing Reparative Stress Signaling. J. Nat. Prod. 2015, 78, 1990–2000. [CrossRef]
- Chittezhath, M.; Deep, G.; Singh, R.P.; Agarwal, C.; Agarwal, R. Silibinin Inhibits Cytokine-Induced Signaling Cascades and down-Regulates Inducible Nitric Oxide Synthase in Human Lung Carcinoma A549 Cells. Molecular Cancer Therapeutics 2008, 7, 1817–1826. [CrossRef]
- Zhao, Y.; Zhou, Y.; Gong, T.; Liu, Z.; Yang, W.; Xiong, Y.; Xiao, D.; Cifuentes, A.; Ibáñez, E.; Lu, W. The Clinical Anti-Inflammatory Effects and Underlying Mechanisms of Silymarin. iScience 2024, 27, 111109. [CrossRef]
- Munteanu, C.; Schwartz, B. The Effect of Bioactive Aliment Compounds and Micronutrients on Non-Alcoholic Fatty Liver Disease. Antioxidants 2023, 12, 903. [CrossRef]
- Zadeh, M.M.; Motamed, N.; Ranji, N.; Majidi, M.; Falahi, F. Silibinin-Induced Apoptosis and Downregulation of MicroRNA-21 and MicroRNA-155 in MCF-7 Human Breast Cancer Cells. J Breast Cancer 2016, 19, 45. [CrossRef]
- Shoaib, M.; Shah, I.; Ali, N.; Adhikari, A.; Tahir, M.N.; Shah, S.W.A.; Ishtiaq, S.; Khan, J.; Khan, S.; Umer, M.N. Sesquiterpene Lactone! A Promising Antioxidant, Anticancer and Moderate Antinociceptive Agent from Artemisia Macrocephala Jacquem. BMC Complement Altern Med 2017, 17. [CrossRef]
- Taleghani, A.; Emami, S.A.; Tayarani-Najaran, Z. Artemisia: A Promising Plant for the Treatment of Cancer. Bioorganic & Medicinal Chemistry 2020, 28, 115180. [CrossRef]
- Zhang, Q.; Yi, H.; Yao, H.; Lu, L.; He, G.; Wu, M.; Zheng, C.; Li, Y.; Chen, S.; Li, L.; et al. Artemisinin Derivatives Inhibit Non-Small Cell Lung Cancer Cells Through Induction of ROS-Dependent Apoptosis/Ferroptosis. J. Cancer 2021, 12, 4075–4085. [CrossRef]
- Zhu, M.; Wang, Y.; Han, J.; Sun, Y.; Wang, S.; Yang, B.; Wang, Q.; Kuang, H. Artesunate Exerts Organ- and Tissue-Protective Effects by Regulating Oxidative Stress, Inflammation, Autophagy, Apoptosis, and Fibrosis: A Review of Evidence and Mechanisms. Antioxidants 2024, 13, 686. [CrossRef]
- Laurindo, L.F.; Santos, A.R.D.O.D.; Carvalho, A.C.A.D.; Bechara, M.D.; Guiguer, E.L.; Goulart, R.D.A.; Vargas Sinatora, R.; Araújo, A.C.; Barbalho, S.M. Phytochemicals and Regulation of NF-kB in Inflammatory Bowel Diseases: An Overview of In Vitro and In Vivo Effects. Metabolites 2023, 13, 96. [CrossRef]
- Efferth, T.; Oesch, F. The Immunosuppressive Activity of Artemisinin-type Drugs towards Inflammatory and Autoimmune Diseases. Medicinal Research Reviews 2021, 41, 3023–3061. [CrossRef]
- Ma, Z.; Woon, C.Y.-N.; Liu, C.-G.; Cheng, J.-T.; You, M.; Sethi, G.; Wong, A.L.-A.; Ho, P.C.-L.; Zhang, D.; Ong, P.; et al. Repurposing Artemisinin and Its Derivatives as Anticancer Drugs: A Chance or Challenge? Front. Pharmacol. 2021, 12. [CrossRef]
- Sagar, S.M.; Yance, D.; Wong, R.K. Natural Health Products That Inhibit Angiogenesis: A Potential Source for Investigational New Agents to Treat Cancer—Part 1. Current Oncology 2006, 13, 14–26. [CrossRef]
- Kazmi, S.T.B.; Fatima, H.; Naz, I.; Kanwal, N.; Haq, I. Pre-Clinical Studies Comparing the Anti-Inflammatory Potential of Artemisinic Compounds by Targeting NFκB/TNF-α/NLRP3 and Nrf2/TRX Pathways in Balb/C Mice. Front. Pharmacol. 2024, 15. [CrossRef]
- Silva, G.D.S.E.; Marques, J.N.D.J.; Linhares, E.P.M.; Bonora, C.M.; Costa, É.T.; Saraiva, M.F. Review of Anticancer Activity of Monoterpenoids: Geraniol, Nerol, Geranial and Neral. Chemico-Biological Interactions 2022, 362, 109994. [CrossRef]
- Ben Ammar, R.; Mohamed, M.E.; Alfwuaires, M.; Abdulaziz Alamer, S.; Bani Ismail, M.; Veeraraghavan, V.P.; Sekar, A.K.; Ksouri, R.; Rajendran, P. Anti-Inflammatory Activity of Geraniol Isolated from Lemon Grass on Ox-LDL-Stimulated Endothelial Cells by Upregulation of Heme Oxygenase-1 via PI3K/Akt and Nrf-2 Signaling Pathways. Nutrients 2022, 14, 4817. [CrossRef]
- Kim, H.; Li, S.; Nilkhet, S.; Baek, S.J. Anti-Cancer Activity of Rose-Geranium Essential Oil and Its Bioactive Compound Geraniol in Colorectal Cancer Cells. Appl Biol Chem 2025, 68. [CrossRef]
- Bhattacharjee, S.; Dashwood, R.H. Epigenetic Regulation of NRF2/KEAP1 by Phytochemicals. Antioxidants 2020, 9, 865. [CrossRef]
- Nurkolis, F.; Taslim, N.A.; Syahputra, R.A.; Annette d’Arqom; Tjandrawinata, R.R.; Purba, A.K.R.; Mustika, A. Food Phytochemicals as Epigenetic Modulators in Diabetes: A Systematic Review. Journal of Agriculture and Food Research 2025, 21, 101873. [CrossRef]
- Gowd, V.; Kanika; Jori, C.; Chaudhary, A.A.; Rudayni, H.A.; Rashid, S.; Khan, R. Resveratrol and Resveratrol Nano-Delivery Systems in the Treatment of Inflammatory Bowel Disease. The Journal of Nutritional Biochemistry 2022, 109, 109101. [CrossRef]
- Song, X.; Wang, Y.; Gao, L. Mechanism of Antioxidant Properties of Quercetin and Quercetin-DNA Complex. J Mol Model 2020, 26. [CrossRef]
- Olasehinde, T.A.; Olaokun, O.O. Apigenin and Inflammation in the Brain: Can Apigenin Inhibit Neuroinflammation in Preclinical Models? Inflammopharmacology 2024, 32, 3099–3108. [CrossRef]
- Pinto, C.; Cidade, H.; Pinto, M.; Tiritan, M.E. Chiral Flavonoids as Antitumor Agents. Pharmaceuticals (Basel) 2021, 14, 1267. [CrossRef]
- Ding, K.; Jiang, W.; Jia, H.; Lei, M. Synergistically Anti-Multiple Myeloma Effects: Flavonoid, Non-Flavonoid Polyphenols, and Bortezomib. Biomolecules 2022, 12, 1647. [CrossRef]
- Iordache, M.P.; Buliman, A.; Costea-Firan, C.; Gligore, T.C.I.; Cazacu, I.S.; Stoian, M.; Teoibaș-Şerban, D.; Blendea, C.-D.; Protosevici, M.G.-I.; Tanase, C.; et al. Immunological and Inflammatory Biomarkers in the Prognosis, Prevention, and Treatment of Ischemic Stroke: A Review of a Decade of Advancement. Int. J. Mol. Sci. 2025, 26, 7928. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



