Submitted:
26 September 2025
Posted:
30 September 2025
You are already at the latest version
Abstract
Female infertility, affecting millions worldwide, involves complex molecular mechanisms such as chronic inflammation, impaired cellular death, and protein regulation. This study explores how the inflammatory protein IL-6, autophagy marker LC3, ubiquitination processes, and small RNA molecules of miR-146a-5p, miR-9-5p, and miR-9-3p contribute to the control of ovarian function and female infertility. Two expression profile datasets (GSE199225 and GSE146856) were screened and downloaded from GEO. DEGs were screened using the GEO2R and ggVennDiagram tools. The three miRNAs were retrieved from datasets using the multiMiR tool, and IL6-targeted genes were retrieved from MSigDB. IL6, miRNAs interactions were constructed. Further, LC3, Ubiquitination, and their cross-relation with the DEGs associated miRNAs were demonstrated. Meanwhile, GO/KEGG pathway enrichment analyses, and molecular network interaction analysis were performed. Lastly, immunohistochemistry and quantitative PCR (qPCR) were used to confirm the expression of IL6, LC3, and miRNA in ovarian endometrial tissues compared to control. Results showed that IL-6 drives inflammation in conditions of PCOS and ovarian endometriosis, which then disrupt ovulation and embryo implantation. The miR-146a-5p reduced inflammation by targeting gene TRAF6, while miR-9-5p regulated protein degradation via SQSTM1. In agreement with the bioinformatic approach, experimental analysis revealed a reduced IL6 protein expression in ovarian endometriosis tissues while mRNA IL6 level was increased, suggesting the presence of post-transcriptional regulatory mechanisms that act to limit excessive inflammation, probably through miRNAs. Indeed, miR-146a-5 levels, which plays a role in immune modulation and inflammatory signaling were significantly upregulated. Interestingly, an induction of autophagic mechanism revealed by elevated LC3 was also observed. Aligned with these experimental data, bioinformatic analysis showed that autophagy genes LC3, ATG5 and ubiquitination processes were tightly linked to ovarian health, with disruptions accelerating follicle loss and oxidative damage. In conclusion, results showed that IL-6, miRNAs and autophagy work together to control inflammation and cellular repair in ovarian disorders. This study opens new avenues for targeted treatments to improve fertility outcomes by connecting molecular networks to clinical insights.
Keywords:
Femle fertility
; Ovary
; Interleukin-6
; LC3
; Ubiquitination
; miRNAs
1. Introduction
Female infertility affects approximately 10–15% of couples globally, with polycystic ovary syndrome (PCOS), endometriosis, and idiopathic ovarian dysfunction representing the leading causes [1]. These conditions are driven by molecular dysregulation, including chronic inflammation, oxidative stress, and defective autophagy, which impairs folliculogenesis, oocyte quality, and endometrial receptivity [2]. Among inflammatory mediators, interleukin-6 (IL-6) is a well-established cornerstone of the chronic low-grade inflammation characteristic of PCOS. Elevated serum and follicular fluid levels of IL-6 are consistently reported in women with PCOS and are strongly correlated with insulin resistance, hyperandrogenism, and dysfunctional ovulation [3-6]. In endometriosis, IL-6 drives a pro-inflammatory microenvironment that promotes lesion survival and impairs endometrial receptivity [5].
MicroRNAs (miRNAs), small non-coding RNAs, are emerging as master regulators of reproductive health [7-9]. MicroRNAs (miRNAs) have emerged as critical post-translational regulators of the pathways disrupted in PCOS and endometriosis. Specifically, miR-146a-5p is recognized for its role in dampening innate immune response. In PCOS, miR-146a-5p is dysregulated and has been shown to target key genes like TRAF6 and IRAK1 in granulosa cells, directly linking it to the inflammatory signaling prevalent in the syndrome [10-12]. Similarly, the miR-9 family (miR-9-3p/5p) is involved in granulosa cell apoptosis and follicular atresia. Studies indicate that miR-9 influences aromatase expression and estrogen production, processes fundamental to follicular development that are impaired in PCOS [13-15]. Despite their significance, the interplay between miRNAs, IL-6, and downstream pathways of autophagy and ubiquitination remains underexplored.
Autophagy, a cellular clearance mechanism marked by the conversion of LC3-I to LC3-II, is crucial for maintaining oocyte quality and granulosa cell function. Growing evidence indicates that autophagy is impaired in PCOS ovaries, contributing to oxidative stress and the accumulation of damaged organelles, which in turn exacerbates metabolic and reproductive dysfunction [16-18]. The ubiquitination system works in concert with autophagy to regulate protein turnover, and its dysregulation is increasingly implicated in ovarian aging and PCOS pathogenesis [19-21]. Dysregulation of these processes is linked to oxidative damage, mitochondrial dysfunction, and apoptosis of the hallmarks of ovarian aging and implantation failure [22].
Current research on female infertility primarily examines isolated pathways, neglecting the integrative roles of IL-6, miRNAs, autophagy, and ubiquitination. A systems-level approach is needed to unravel their crosstalk and identify therapeutic targets. This study hypothesizes that IL-6-driven inflammation, miRNA dysregulation, and impaired autophagy-ubiquitination networks synergistically contribute to female infertility. By leveraging bioinformatics analyses of gene expression datasets GSE199225 [23] and GSE146856 [24], we aim to map these interactions. These datasets were selected for their focus on PCOS, utilization of high-resolution RNA-seq technology, and well-matched control groups, providing a robust foundation for our analysis. The results obtained by bioinformatic tools were confirmed by experimental analysis using immunohistochemistry and quantitative PCR (qPCR) techniques.
2. Materials and Methods
2.1. Data Processing
The gene expression profiles related to female infertility were retrieved and downloaded from the Gene Expression Omnibus (GEO) public repository of the National Center for Biotechnology Information (NCBI). The retrieved Boolean queries were including keywords "Interleukin-6" OR "IL-6", "LC3" OR "MAP1LC3", "Ubiquitination" OR "Proteasome pathway", "miR-146a-5p" OR "miR-9-1", and conditions related to female infertility was "Endometriosis" OR "PCOS" OR "Ovarian Dysfunction". To ensure high-quality data, datasets were prioritized, retrieved, and downloaded using Expression Profiling by Array or RNA-seq technologies, which provide comprehensive annotations.
GSE199225 [23] is an RNA-seq expression profile based on GPL24676 Illumina NovaSeq 6000 (Homo sapiens) and contains sample of six healthy lean control women, all aged between 18-40 years and PCOS. GSE146856 [24] is an expression profile based datasets based on GPL20795 HiSeq X Ten (Homo sapiens) provided by International Peace Maternity and Child Health Hospital, Shanghai, China and it contains sample of control tissue and PCOS. The samples of the two expression profile datasets were classified and analyzed based on the PCOS condition.
IL6 signaling related genes were extracted from the Molecular Signature Database (MSigDB) by Human collection Biocarta, Apoptosis, and IL6 pathway gene sets category [25]. microRNAs (miRs) are fundamental regulators of protein-coding genes [26]. The miR-146a-5p is a small, non-coding RNA molecule involved in the post-transcriptional regulation of gene expression [27]. miR-9 is a microRNA (miRNA) that plays a crucial role in brain development and function. miR-9-5p and miR-9-3p are two single-stranded microRNAs (miR) derived from the same RNA duplex [26]. The three miRs targeted protein coding genes were derived from miRRecords, miRbase, and TarBase databases by using multiMiR R library [28]. The workflow of this study is illustrated in Figure 1.
2.2. Analysis of Microarray Datasets
Based on the PCOS role in Female Infertility, in the subsequent analysis, samples of each dataset were defined into two groups “Control” and “PCOS”. Raw expression data and metadata extracted by using GEO2R [29, 30]. Normalization and log2 conversion were carried out for each dataset to filter out the DEGs of the two datasets, and the DEGs are displayed as volcano plots. The filtering conditions were as follows: |log2-fold change| ≥ 1 and adjusted P-value (adj. P) < 0.05. The ggVennDiagram R package was used to compare and analyze the intersection of genes [31]. Based on the intersection results, the final DEGs were obtained and integrated with the results for further analysis.
2.3. Identification of miRNA-Target Interactions
To identify high-confidence genes targeted by the three miRNAs miR-146a-5p, miR-9-5p and miR-9-3p, we utilized the multiMiR R package [28], which provides a comprehensive compilation of miRNA-target interactions from multiple validated and predicted datasets. To ensure high reliability, the analysis was restricted to experimentally validated targets from curated sources within multiMiR, including miRTarBase, TarBase, and miRRecords. The multimir function was used with the parameters table = 'validated' and predicted.site = 'conserved' to retrieve a list of target genes for each miRNA supported by experimental evidence. The resulting gene symbols for each miRNA were then used for subsequent analyses. ENTREZID IDs were mapped for these target gene lists using org.Hs.eg.db database to facilitate KEGG pathway enrichment analysis using the clusterProfiler package [32] with the following parameters: organism (hsa), adjusted p-value cutoff (pvalueCutoff = 0.05), and Benjamini-Hochberg correction (pAdjustMethod = "BH"). Enriched pathways were filtered for relevance to IL6, autophagy, and ubiquitination, and results were visualized using dot plots with significance thresholds (qvalueCutoff = 0.05).
2.4. Analysis of IL6 and miRNA Interactions
A total of 648 genes were retrieved. 566 IL6 unique genes were found after the deletion of duplicated genes. DEGs with female infertility were first cross-referenced with IL6-related genes obtained from the MSigDB database to identify overlapping genes potentially regulated by IL6 signaling. The overlapped IL6 genes interacted with the three miRNAs targeted genes. To examine the interactions more broadly, a relationship network model was constructed between the DEGs associated with IL6 genes and the targeted miRNA genes using Cytoscape [33]. Enrichment analysis of the key genes from the network was performed for the interacted genes using Enrichr [34]. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway [35] and GO enrichment analyses were used to identify the significant pathways.
2.5. Analysis of miRNAs Interactions with LC3 and Ubiquitination
Autophagy-related genes were curated based on their roles in LC3-mediated autophagy and ubiquitination, based on a literature study. Overlapping targets of miR-146a-5p, miR-9-3p, and miR-9-5p were identified using the multiMiR package [28]. Gene symbols were mapped to Entrez IDs via the org.Hs.eg.db database, and GO Biological Process (BP) enrichment was performed with clusterProfiler using Benjamini-Hochberg correction where pAdjustMethod = "BH" and ont = "BP" [32]. Significant terms were visualized via dot plots with a pvalueCutoff = 0.05.
2.6. Network Construction of the LC3, Ubiquitination and miRNAs
To visualize miRNA-gene interactions and their role in autophagy and ubiquitination, a regulatory network was constructed by using Cytoscape [33]. Signaling pathways of upstream regulators of MTOR, TRAF6, and NF-κB were added to the network to show the indirect effect of the genes. The signaling-related genes were downloaded from STRING-DB [36]. While STRING-DB identified broader interactions within the NF-κB pathway, it focused on TRAF6 → NFKB1 → ULK1 due to their direct roles in autophagy regulation and relevance to female infertility [37, 38]. In the style panel, miRNA nodes with red triangles, orange diamond shapes, and dashed line edges were added to the signaling regulators, and all the gene targets were added in a blue circle with solid line edges.
2.7. Pathway Analysis of the LC3-Ubiquitination and miRNAs
The protein-coding genes from the autophagy, ubiquitination and miRNAs network were uploaded to the Enrichr [34]. The organism selected is Human. The KEGG Pathways, Reactome Pathways, and GO Biological Processes databases were selected to analyze the pathways. The database generated pathways along with p-values and adjusted p-values. The top pathways were sorted based on the p-values and adjusted p-values.
2.8. qRT-PCR
A group of women with ovarian endometriosis was confirmed histologically on formalin-fixed paraffin-embedded (FFPE) tissues. FFPE tissue samples were acquired by using a rotary microtome, tissue sections, enabling the visualization of a single cell layer. Genomic DNA and RNA were extracted from patients with endometriosis and controls using a commercial kit (AllPrep DNA/RNA FFPE kit, Qiagen, Germany). All clinical data were used anonymously to protect patient privacy. The quantity and quality of DNA and RNA were measured using a Nanodrop ND-2000C spectrophotometer (Thermo Scientific, Wilmington, DE, USA). The extracted nucleotides were kept at −20°C until further analysis. Following the manufacturer’s protocol, total RNA (2000 ng/μL) was transcribed into cDNA using a high-capacity cDNA Reverse Transcription Kit (Applied Biosystems 4368814, Carlsbad, CA). Optimal amplification was achieved using qRT-PCR. Primers were designed manually to generate amplicons of ≤120 bp. Each primer was at least 18 nucleotides in length with an appropriate GC content to minimize the formation of secondary structures. To avoid primer dimer formation, both forward and reverse primers were designed with minimal 3′ end complementarity and equal melting temperatures (Tm). Primer specificity was confirmed through BLAST analysis. The oligonucleotide primers (Table 1) were synthesized by Macrogen, Inc. (Seoul, South Korea) and diluted in DNase/RNase-free water to a final working concentration of 10 pmol. qRT-PCR reactions were prepared with Universal SYBR Green Supermix (1725120, Bio-Rad, Hercules, CA, USA). Each well of the 96-well plate contained 10 μL of reaction mix, composed of 4 μL SYBR Green Supermix, 1 μL ROX, 2 μL cDNA, 1 μL of each primer, and 2 μL nuclease-free water. Amplification was performed on a QuantStudio 7 Flex Real-Time PCR System (Applied Biosystems, Hercules, CA, USA). All reactions were run in triplicate. GAPDH served as the reference gene for normalization. Data analysis was performed using SPSS software (version 20; SPSS Inc., Chicago, IL, USA).
2.9. Immunohistochemical Staining (IHC)
Sections obtained from FFPE tissue samples and mounted on coated slides were deparaffinized and immersed in xylene twice for 15 min each, rehydrated using a graded alcohol series, and washed in distilled water. The slides were heated in a citric acid buffer for 5 min in a microwave for antigen retrieval. The sections were then rinsed in distilled water to suppress endogenous peroxidase activity and subsequently treated for 5 minutes with a 0.3% hydrogen peroxide solution. Afterward, 0.01 M phosphate-buffered saline PBS (pH 7.2) was used to rinse the sections. The sections were then incubated with a blocking solution for an hour to minimize non-specific background staining. Next, the primary antibodies (anti-IL6 (ABCAM, ab9324), anti-LC3 (ABCAM, ab229327) and anti-actin (ABCAM, ab1801)), were applied to the samples at 4°C in a wet chamber overnight. After washing with PBS, the slides were incubated with the corresponding secondary antibody (ABCAM, ab6728), for one hour at room temperature. Haematoxylin was used as a counterstain to provide the stained antigen with a contrasting background. The slides were dehydrated, and xylene was used to clear the sections after passing through a graded alcohol series.
2.10. Statistical Analysis
GraphPad Prism software (GraphPad Software version 10.2, San Diego, CA, USA) was used to compare gene and protein expression levels between the control and patient groups. P-values less than 0.05 were considered statistically significant.
2.11. Ethical Approval
This study was approved by the Ethical Committee and Institutional Review Board of the College of Medicine Research Center at King Saud University Medical City (reference number E-22-6750).
3. Results
3.1. Identification of DEGs
The correlation of all DEGs from the two expression profile array datasets showed in the volcano plots (Figure 2). After screening and comprehensive analysis with the ggVennDiagram, 8 upregulated and downregulated genes were identified in the control group (Figure 3A,B). Similarly, 8 upregulated and downregulated genes were identified in the PCOS group also (Figure 3C,D). PSG4, CFI, CHI3L1, BDKRB2, GUCY1A2, SFRP4, IL33, PADI2 upregulated genes in control group but downregulated in PCOS group were shared in both datasets and PITX1, ISG15, NEURL1, OAS2, PRSS3, PI16, L3MBTL4, MX2 downregulated genes in control group but upregulated in PCOS group were shared in both datasets. Finally, a total of 2312 DEGs were screened (|log2-fold change| ≥ 1 and adjusted P-value (adj. P) < 0.05) after deletion of duplicated genes, where 1148 genes were upregulated and 1164 were downregulated expression genes.
3.2. miRs Gene Target Pathway Analysis
KEGG enrichment analysis of the combined target genes of miR-146a-5p, miR-9-3p, and miR-9-5p targets revealed significant associations with pathways central to inflammation, autophagy, and ubiquitination (p.adjust ≤ 5e-06). The MAPK signaling pathway (p.adjust = 1e-06), linked to IL6-driven inflammation and follicular dysfunction, was the most enriched, followed by Ubiquitin-mediated proteolysis (p.adjust = 2e-06), implicating miRNA regulation of protein degradation in ovarian aging (Figure 4). Pathways such as FoxO signaling (p.adjust = 4e-06) and Protein processing in endoplasmic reticulum (p.adjust = 3e-06) highlighted autophagy (LC3-associated) roles in endometrial stress responses. Notably, Cellular senescence (p.adjust = 5e-06) and p53 signaling (p.adjust = 5e-06) tied miR-9-3p/5p to oxidative stress and apoptosis in ovarian follicles. IL6-associated pathways (e.g., Focal adhesion, Cytoskeleton regulation) were enriched (GeneRatio = 0.03–0.05), with miR-146a-5p targeting genes in Endocytosis and Proteoglycans in cancer, suggested roles in immune dysregulation and implantation failure.
3.3. Identification of IL-6-miRNA Interactions
A total of 566 IL6 genes and 2312 DEGs were intersected and 91 common genes were intersected (Figure 5A). The Venn diagram demonstrated that the 91 IL6 genes matched with the three miRNAs to demonstrate the relationship between IL6 and the miRNAs (Figure 5). No miRNAs were exclusive to IL6 alone (Figure 5B). That indicates that IL6 interacts with these miRNAs in combination with other pathways. The largest overlap occurring between mir-146a-5p and IL6 is 28 genes CFH, CASP7, CCND2, ICAM1, CXCL8, BAK1, TNFRSF21, HIF1A, PIM1, BNIP3L, C3, C4BPB, DUSP5, F10, HAS2, LRP1, PDGFB, PDGFRB, TIMP2, IER3, EHD1, PDE4B, TNFRSF1B, DGKG, BST2, KCNIP2, TXNIP, MCL1, that suggests strong co-regulation between this miRNA and IL6. A core set of 2 genes, HAS2 and PDGFRB, was shared among IL6, mir-146a-5p, mir-9-3p, and mir-9-5p, highlighting their central role in IL6-associated processes like inflammation [39]. Smaller overlaps included 11 genes CCND2, ICAM1, CXCL8, HIF1A, PIM1, DUSP5, HAS2, LRP1, PDGFRB, TIMP2 and EHD1 shared by IL6, mir-146a-5p, and mir-9-5p. 5 genes PDGFRB, SLC7A2, EREG, HAS2, ITGB8 shared by IL6, mir-9-3p, and mir-9-5p. A core set of 7 genes CASP7, BAK1, TNFRSF21, BNIP3L, HAS2, PDGFRB, MCL1 shared by IL6 mir-146a-5p, and mir-9-3p. These results emphasize that IL6 collaborates with mir-146a-5p, mir-9-3p, and mir-9-5p through shared regulatory networks, rather than acting independently, to influence cellular responses [40, 41]. To visualize the functional relationships, a network model was constructed focusing on the 91 genes that were common to both the IL-6 signaling pathway and the infertility-associated DEGs (Figure 6). IL6-interacting genes include ICAM1, which is related to inflammation [42], TNFRSF21 related to apoptosis [43], and CXCL8 related to immune signaling [44], which are co-regulated by the miRNAs. mir-146a-5p and mir-9-5p strongly overlap with IL6-associated genes like ICAM1 and TNFRSF21, suggesting their role in modulating IL6-driven pathways. mir-9-3p partially overlaps CXCL8 and DUSP5, linking it to chemokine signaling and stress responses [44, 45]. The network reveals shared targets between IL6 and these miRNAs, indicating coordinated regulation of inflammation and immune processes. mir-146a-5p and mir-9-5p directly targeted IL6-regulated genes, while mir-9-3p co-regulated CXCL6 and DUSP5.
Several key genes in the network model (Figure 6) are associated with female infertility. HAS2 has a critical role for cumulus cell expansion during ovulation [46]. HBEGF gene is essential for embryo implantation and endometrial receptivity [47]. The ICAM1 is linked to endometriosis-associated inflammation and infertility [42, 48]. HIF1A influences placental development and abortion [49]. CASP7 regulates apoptosis in ovarian follicles, affecting ovarian reserve [50]. Also, CXCL8 modulates inflammatory processes during ovulation [51].
3.4. Enrichment Analysis of Key IL6 Genes
To identify pathways most critical to the IL-6-miRNA crosstalk, we focused enrichment analysis on a subset of 9 key IL-6 associated genes (ICAM1, TNFRSF21, CXCL8, DUSP5, HAS2, HBEGF, HIF1A, CASP7, IL6). These genes were selected based on their central position in the interaction network with the three miRNAs (Figure 6) and their established roles in reproductive processes. The p < 0.05 was considered as the cutoff criterion for significant enrichment. The genes revealed significant associations with inflammatory, immune, and developmental pathways critical to female infertility (Figure 7). Key enriched terms included "Cellular Response to Cytokine Stimulus" (GO:0071345) (Figure 7A), "TNF Signaling Pathway" (KEGG, Figure 7E), and "IL6/JAK-STAT3 Signaling" (MSigDB, Figure 7D), highlighting roles in inflammation, endometrial dysfunction, and folliculogenesis. Genes such as ICAM1, HAS2, and HBEGF were linked to extracellular matrix remodeling and embryo implantation, while HIF1A and CASP7 were tied to hypoxia and ovarian follicle apoptosis. The MSigDB pathway analysis confirmed IL6 as a central hub coordinating these processes.
3.5. Identification of LC3 and Ubiquitination Interaction with miRNAs
The curated autophagy related genes (Table-1) were used to run the GO enrichment analysis that revealed significant associations between miRNA targets and autophagy-related processes where p.adjust ≤ 6e-06 (Figure 8). The top enriched terms are “mitophagy of mitochondrion” with a p.adjust value of 2e-06, “macroautophagy” with a p.adjust value of 4e-06), and autophagosome assembly with a p.adjust value of 6e-06), with a high gene ratio 0.6–1.0. Key genes MAP1LC3A and LC3B, which are associated with “autophagosome formation”, SQSTM1 and p62 are “ubiquitin-binding” genes, and PARK2 is associated with “mitophagy”, were prominent in these pathways. Additionally, “response to starvation” and “cellular response to nutrient levels” implicated miRNAs in stress-induced autophagy, which is critical for ovarian follicle survival and oocyte quality [52].
3.6. Network Analysis of the LC3-Ubiquitination and miRNAs
Cytoscape network analysis revealed the miR-146a-5p, miR-9-3p, and miR-9-5p as central regulators of upstream autophagy and inflammatory signaling (Figure 9). The network highlights cross-regulation between autophagy (ATG5, MAP1LC3B) and inflammatory pathways (TRAF6, NFKB1), with miRNAs acting as molecular bridges. miR-146a-5p suppresses TRAF6 and NFKB1, dampening IL6-driven inflammation while indirectly promoting autophagy via reduced MTOR activation [60-62]. miR-9-3p targets MTOR by directly stimulating autophagy initiation through ULK1 and BECN1 [63]. Also, miR-9-5p regulation of SQSTM1/p62, linking ubiquitination to autophagosome formation via MAP1LC3B (LC3B) [64].
3.7. GO Analysis of the LC3-Ubiquitination and miRNAs
GO enrichment analysis of upstream signaling networks revealed significant associations between miR-146a-5p, miR-9-3p, miR-9-5p, and pathways critical to autophagy, ubiquitination, and inflammation (Figure 10). Strong enrichment of macroautophagy (GO:0016236) (Figure 10E), autophagosome assembly (GO:0000045) (Figure 10A), and mitophagy (GO:0000423) (Fig 10A), highlighting miRNA regulation of mitochondrial quality control and autophagic flux in the biological process. In the cellular component pathway (Figure 10B), enrichment of autophagosome membrane (GO:0009421) and TORC1 complexes (GO:0031931), linking miRNAs to nutrient-sensing pathways (e.g., MTOR) in ovarian follicle survival. Also, ubiquitin ligase binding (GO:0031625) (Figure 10C) and protein kinase binding (GO:0019901) (Figure 10C), implicating miRNAs in ubiquitination and stress signaling, are revealed in the molecular functions. Moreover, in the KEGG and RCTM pathways (Figure 10D-E), autophagy, PINK1-PARKIN-mediated mitophagy, and NF-κB signaling dominated, with PD-1 checkpoint pathways and the significant pathways and their overlap genes (Table 2) suggesting immune-infertility crosstalk.
3.8. Interaction of LC3, IL6, and Ubiquitination Genes with Key Biological Processes
The clustergram (Figure 11) highlights significant associations between IL6-miRNA network genes and autophagy-related biological processes with log10(Adj.Pval) ≥ 0.2–0.8. MTOR exhibited the strongest link to negative regulation of programmed cell death (GO:0043069, log10(Adj.Pval) = 0.8), followed by NFKB1 and TRAF6 with cellular response to starvation (GO:0009267, log10(Adj.Pval) = 0.6) [65]. Autophagy genes MAP1LC3B, BECN1, and ATG5 clustered tightly with macroautophagy (GO:0016236) and autophagosome assembly (GO:0000045), underscoring their roles in ovarian follicle survival. SQSTM1/p62 is strongly linked to mitophagy (GO:0000423), emphasizing its role in mitochondrial quality control [66].
3.9. Experimental Analysis
We performed analyses of the mRNA and protein expression of IL-6 expression in ovarian endometriosis. Protein expression analysis using immunohistochemistry (IHC) revealed a paradoxical downregulation of IL-6 protein levels in endometriotic lesions relative to normal (Figure 14A-G)., while a significant increase in IL-6 mRNA revealed by quantitative real-time PCR (qRT-PCR) analysis was observed (Figure 14H). The observed discrepancy between IL-6 mRNA and protein expression indicated the possibility of post-transcriptional regulatory mechanisms, including miRNAs. Our findings indicated a notable upregulation to miR-146a-5p (Figure 14I)., which plays a key role in immune modulation and inflammatory signaling, highlighting the significance of immune dysregulation in disease progression. Interestingly, protein expression analysis using IHC consistently showed a notable increase in LC3 protein levels in ovarian endometriotic lesions compared to control (Figure 14J-P), in accordance with the qRT-PCR result which demonstrated a significant increase in LC3 mRNA levels (Figure 14 Q).
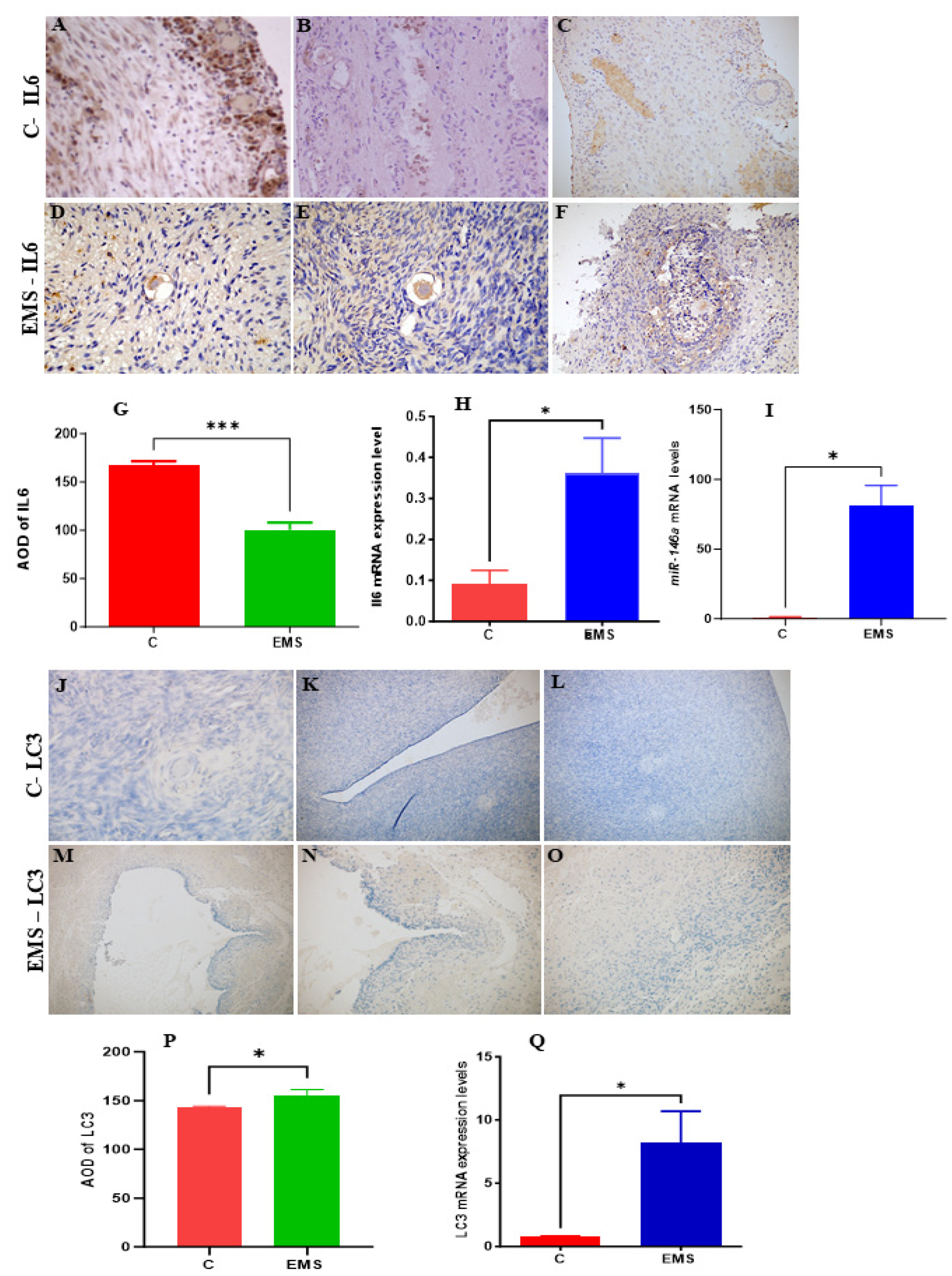
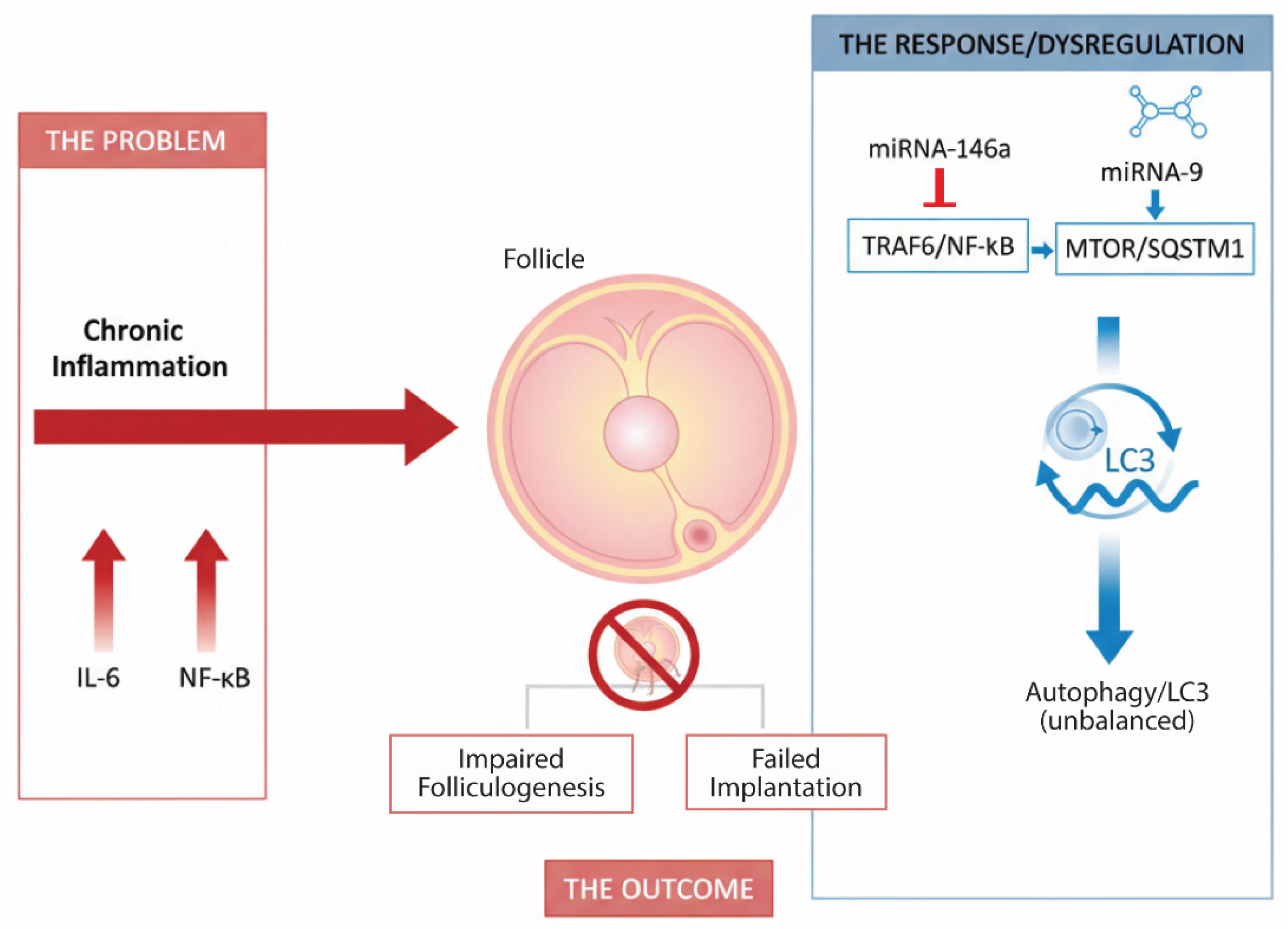
Figure 14.
(A–G): Representative immunohistochemical staining analysis of IL-6 expression in endometrial tissues from control and endometriosis (EMS) groups. Immunohistochemistry showing IL-6 localization predominantly in the cytoplasm and nuclei of glandular and stromal cells. EMS tissues display less strong IL-6 staining (D-F) compared to control samples (A-C), indicating reduced protein expression in the patient group. (H): qRT-PCR analysis demonstrates a significant increase of IL-6 mRNA levels in EMS tissues relative to controls (p < 0.05). Figure 2. (I–O): Representative immunohistochemical staining of LC3 expression in endometrial tissues from control and endometriosis (EMS) groups. Immunohistochemistry showing weak LC3 staining in the control group (I–K) compared to EMS tissues (L-N), with faint cytoplasmic localization in glandular and stromal cells. (M): qRT-PCR analysis demonstrates a significant upregulation of lc3 mRNA levels in EMS tissues compared to controls samples (p < 0.05).
Figure 14.
(A–G): Representative immunohistochemical staining analysis of IL-6 expression in endometrial tissues from control and endometriosis (EMS) groups. Immunohistochemistry showing IL-6 localization predominantly in the cytoplasm and nuclei of glandular and stromal cells. EMS tissues display less strong IL-6 staining (D-F) compared to control samples (A-C), indicating reduced protein expression in the patient group. (H): qRT-PCR analysis demonstrates a significant increase of IL-6 mRNA levels in EMS tissues relative to controls (p < 0.05). Figure 2. (I–O): Representative immunohistochemical staining of LC3 expression in endometrial tissues from control and endometriosis (EMS) groups. Immunohistochemistry showing weak LC3 staining in the control group (I–K) compared to EMS tissues (L-N), with faint cytoplasmic localization in glandular and stromal cells. (M): qRT-PCR analysis demonstrates a significant upregulation of lc3 mRNA levels in EMS tissues compared to controls samples (p < 0.05).
Figure 15.
A proposed model for the crosstalk between IL-6, miRNAs, autophagy, and ubiquitination in female infertility. Chronic inflammation driven by IL-6 disrupts ovarian function. In response, miRNAs like miR-146a-5p and miR-9 are dysregulated, by creating a feedback loop that attempts to control inflammation and manage cellular stress through autophagy (LC3) and ubiquitination. The breakdown of this regulatory network leads to impaired folliculogenesis and failed implantation.
Figure 15.
A proposed model for the crosstalk between IL-6, miRNAs, autophagy, and ubiquitination in female infertility. Chronic inflammation driven by IL-6 disrupts ovarian function. In response, miRNAs like miR-146a-5p and miR-9 are dysregulated, by creating a feedback loop that attempts to control inflammation and manage cellular stress through autophagy (LC3) and ubiquitination. The breakdown of this regulatory network leads to impaired folliculogenesis and failed implantation.
4. Discussion
Female infertility is a major global health issue, with PCOS and ovarian endometriosis being among the most prevalent endocrine disorders associated with it [1]. Both endometriosis and PCOS impact up to 10% of women, severely affecting their health, fertility, and overall quality of life [67]. PCOS is characterized by hormonal imbalances, chronic inflammation, and metabolic dysregulation, which collectively impair ovulation, endometrial receptivity, and follicular development [4]. In this study, we employed a comprehensive bioinformatics approach and experimental analysis to unravel the molecular crosstalk between interleukin-6 (IL6), autophagy, ubiquitination, and three microRNAs, miR-146a-5p, miR-9-3p, and miR-9-5p in the context of female infertility. By analyzing gene expression profiles from PCOS and control groups from GSE199225 and GSE146856 datasets, we identified 2,312 differentially expressed genes (DEGs), including 1,148 upregulated and 1,164 downregulated genes. These DEGs were intricately linked to IL6 signaling, autophagy markers (LC3), ubiquitination pathways, and miRNA interactions, revealing a complex regulatory network that underscores the multifactorial nature of infertility [2].
MicroRNAs, small non-coding RNAs that have fine-tuned gene expression, emerged as pivotal regulators in this study. Using the multiMiR database [28], we identified 12,094 miRNA-target interactions, with miR-146a-5p, miR-9-3p, and miR-9-5p targeting 5,505, 1,359, and 5,230 genes, respectively. The KEGG enrichment analysis [35] highlighted their roles in IL6-driven inflammatory cascades, autophagy, and ubiquitin-mediated proteolysis. The MAPK signaling pathway, central to IL6-mediated inflammation that was strongly enriched with miR-146a-5p by suppressing MAPK inhibitors (DUSP5), thereby amplifying inflammatory signals that disrupt folliculogenesis and endometrial function [68]. Concurrently, miR-9-5p regulated ubiquitin ligases (FBXW7) to stabilize LC3-associated autophagic proteins, which is a process critical for clearing damaged cellular components in aging ovarian follicles [19, 69-71]. These findings position miRNAs as molecular bridges connecting inflammation, autophagy, and protein degradation in reproductive pathophysiology.
Network modeling further elucidated the collaborative roles of IL6 and miRNAs in female infertility. IL6, a pro-inflammatory cytokine that has been reported to be highly expressed in ovarian endometriosis [72], dominated pathways of TNF signaling and NF-κB activation, which are hyperactive in endometriosis and implantation failure [73]. Notably, miR-146a-5p suppressed ICAM1, a gene overexpressed in endometriosis that promotes leukocyte infiltration and tissue damage (Figure 6) [74]. This suggests a feedback loop where IL6-induced inflammation upregulates ICAM1, while miR-146a-5p acts as a compensatory brake, a mechanism likely disrupted in infertility. In accordance with the network modeling findings, experimental analysis using qRT-PCR analysis revealed a significant upregulation of IL-6 mRNA levels. However, protein expression analysis using IHC showed a paradoxical downregulation of IL-6 protein levels. This inconsistency—elevated mRNA levels but reduced protein expression—may contribute to the persistence of ovarian endometriotic lesions by mitigating excessive inflammation through the suppression of IL-6 protein levels. One potential explanation for this regulation involves miRNAs, which are critical in post-transcriptional gene regulation by inhibiting the translation of target mRNAs [75]. Specifically, numerous studies have demonstrated altered miRNA expression in the ectopic and eutopic endometrial tissues [76-78]. Our findings showed that miR-146a-5p, which plays a key role in regulating inflammation, immune responses, and autophagy [79], was significantly upregulated in ovarian endometrial tissues compared to controls. Consistent with previous research [80], miR-146a-5p suppresses the production of pro-inflammatory cytokines like IL-6 through post-transcriptional mechanisms. By binding to the 3′ untranslated region of IL-6 mRNA, miR-146a-5p can inhibit translation or promote mRNA degradation, resulting in decreased IL-6 protein expression.
Similarly, miR-9-5p regulated CASP7, an apoptosis executor, linking IL6-driven oxidative stress to accelerated follicular atresia [81]. Extracellular matrix genes HAS2 and HBEGF, essential for ovulation and embryo implantation, were modulated by IL6 and miRNAs. miR-9-3p indirectly influenced HBEGF-mediated embryo attachment by targeting DUSP5, a phosphatase that regulates endometrial signaling [13]. These interactions mirror clinical observations in PCOS, where IL6 overexpression correlates with defective follicle rupture and implantation failure [4].
GO and KEGG enrichment analyses emphasized the centrality of autophagy and ubiquitination in maintaining ovarian and endometrial homeostasis (Figure 8, Figure 10 and Figure 11). Autophagy-related terms "mitophagy" (GO:0000423) and "autophagosome assembly" (GO:0000045) were enriched with genes MAP1LC3B and SQSTM1/p62, which interact with miR-9-3p and miR-9-5p to maintain mitochondrial and protein quality [22]. Dysregulation of these genes, as seen in endometriosis or PCOS, could impair autophagic flux, exacerbating oxidative damage and fibrosis [16]. The NF-κB signaling pathway further connected IL6 to chronic inflammation, while the Autophagy-Animal pathway highlighted LC3-mediated processes critical for ovarian reserve [19]. Cluster analysis (Figure 11) revealed tight associations between IL6-associated genes (MTOR, NFκB1) and autophagy effectors BECN1 and ATG5, demonstrating their convergence on pathways governing follicular atresia and nutrient sensing (Figure 13). Furthermore, MTOR, a master regulator of autophagy (Figure 12), suppressed apoptosis under metabolic stress but inhibited autophagy via ULK1 repression [82]. miR-9-3p’s targeting of MTOR could restore this balance, enhancing oocyte survival in PCOS [13]. Aligned with the bioinformatic analysis, our experimental findings demonstrated a significant upregulation of LC3 mRNA and protein levels in ovarian endometriotic tissues, as confirmed through qRT-PCR, and IHC. These results align with previous studies that creported increased expression of autophagic markers, such as Beclin-1 and LC3-II, in ovarian endometriomas compared to eutopic and normal endometria, suggesting heightened autophagic activity in endometrial tissues [83, 84]. This elevated autophagic activity likely enables ectopic endometrial cells to survive under adverse conditions, such as oxidative stress and inflammation, by providing a mechanism for adaptation to hypoxic and inflammatory environments. These findings are consistent with earlier research showing that enhanced autophagy supports the proliferation and survival of endometrial cells [85]. Therefore, targeting autophagy-related pathways could represent a promising therapeutic strategy to disrupt the survival mechanisms of endometriotic cells.
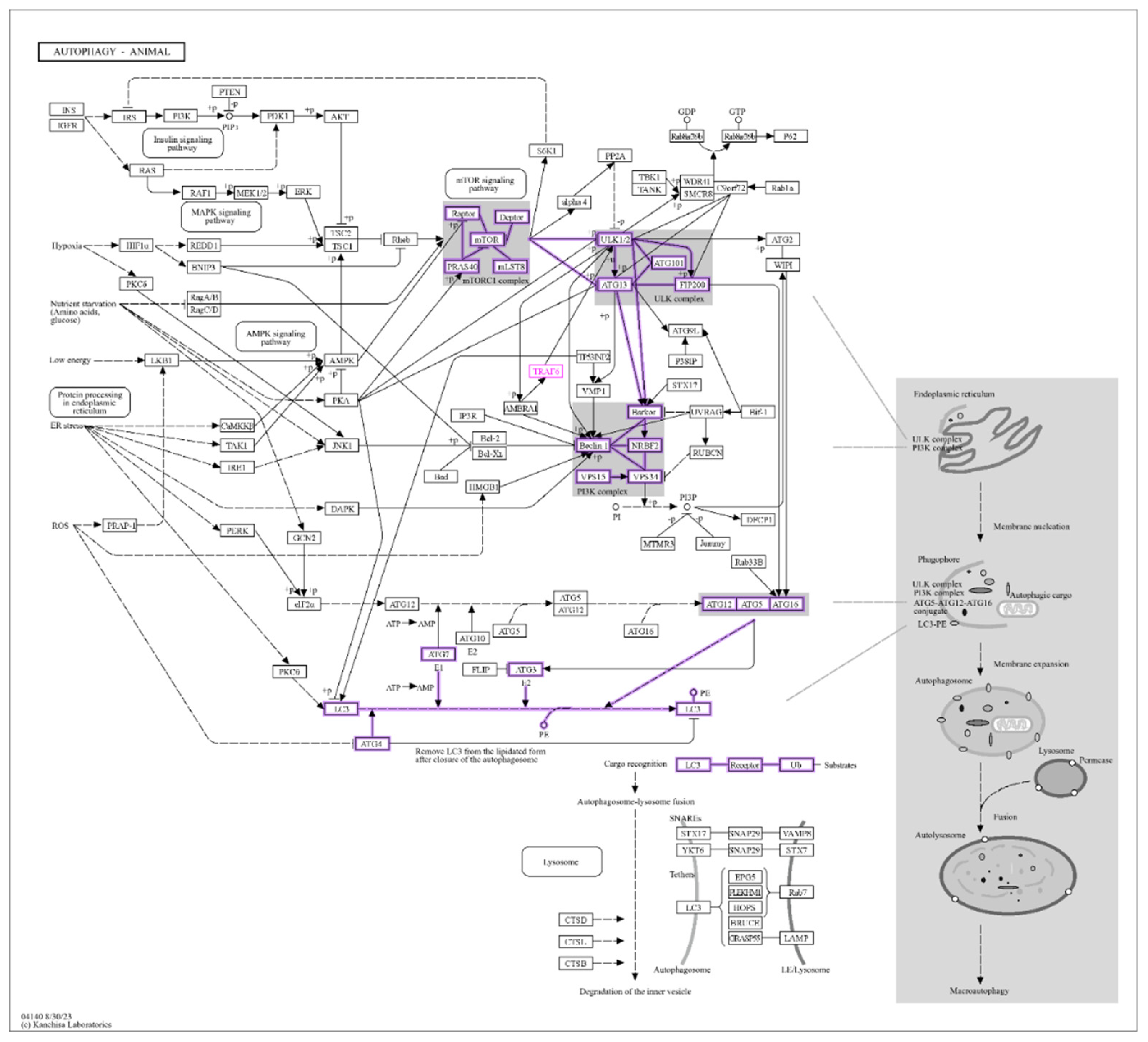
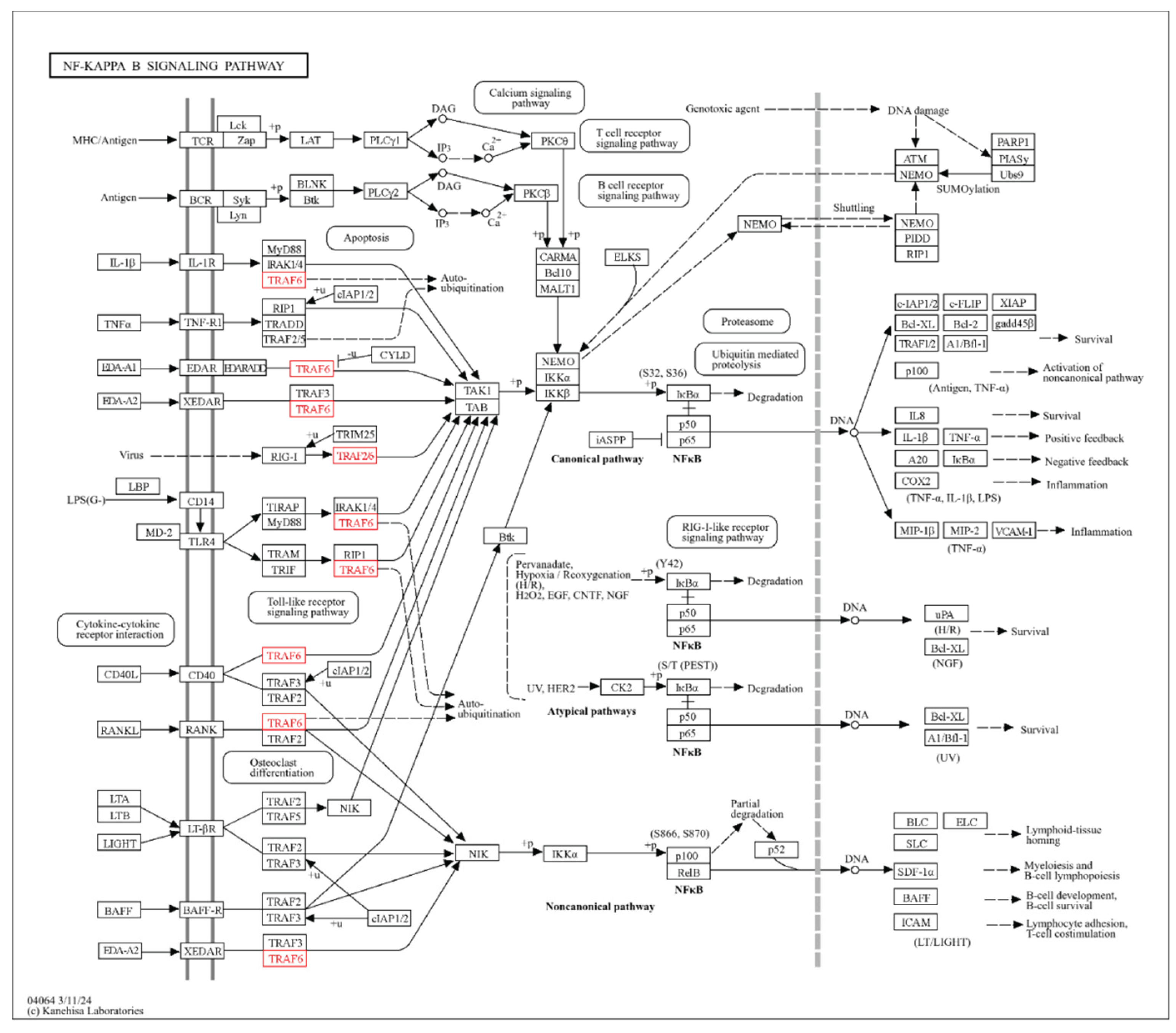
Figure 12.
KEGG Autophagy pathway (nt06532). The core molecular machinery of macroautophagy, including the mTOR signaling pathway, which inhibits ULK1 and the LC3 conjugation system. Genes highlighted in purple (ULK1/2, Beclin 1, LC3, ATG5, ATG7) are key targets of miR-9-3p and miR-9-5p identified in this study, demonstrating their role in regulating autophagic flux.
Figure 12.
KEGG Autophagy pathway (nt06532). The core molecular machinery of macroautophagy, including the mTOR signaling pathway, which inhibits ULK1 and the LC3 conjugation system. Genes highlighted in purple (ULK1/2, Beclin 1, LC3, ATG5, ATG7) are key targets of miR-9-3p and miR-9-5p identified in this study, demonstrating their role in regulating autophagic flux.
Figure 13.
NF-κB signaling pathway (nt06516). It depicts the canonical and non-canonical NF-κB activation pathways. Genes highlighted in red (TRAF6) represent key nodes targeted by miR-146a-5p, indicating their role in suppressing inflammatory signaling downstream of IL-6 and other cytokines.
Figure 13.
NF-κB signaling pathway (nt06516). It depicts the canonical and non-canonical NF-κB activation pathways. Genes highlighted in red (TRAF6) represent key nodes targeted by miR-146a-5p, indicating their role in suppressing inflammatory signaling downstream of IL-6 and other cytokines.
Based on our bioinformatic and experimental findings, we propose a working model for the pathophysiology of female infertility centered on the crosstalk between IL-6, miRNAs, autophagy, and ubiquitination. In this model, chronic inflammation, driven by elevated IL-6, is the initial insult that disrupts the follicular environment and endometrial receptivity. Dysregulated miRNAs act as critical molecular tuners where miR-146a-5p is upregulated as a compensatory brake to suppress IL-6/TRAF6/NF-κB signaling and mitigate inflammation, while the miR-9 family members regulate autophagic (via MTOR & SQSTM1) and apoptotic pathways to manage cellular stress. The interplay between these pathways determines cellular fate. The miR-mediated response influences the autophagy-ubiquitination system (evidenced by LC3 elevation), which is essential for clearing inflammation-induced damage. The paradoxical findings, such as increased IL-6 mRNA but decreased protein, alongside elevated miR-146a-5p and LC3, epitomize this dynamic, failed repair process. The breakdown of this feedback loop results in a detrimental cycle of oxidative stress, impaired oocyte quality, and failed implantation, characterizing conditions like PCOS and endometriosis.
All together, these finding offers critical insights and a collective impact on reproductive health. IL-6, a central driver of chronic inflammation, disrupts ovarian folliculogenesis, endometrial receptivity, and follicular survival, particularly in conditions of endometriosis and PCOS. miRNAs act as molecular tuners, modulating IL-6 signaling to balance inflammation, autophagy, and protein degradation. miR-146a-5p suppresses inflammatory mediators (TRAF6/NFκB), while miR-9-5p regulates ubiquitination (via SQSTM1) to maintain cellular quality. Autophagy, marked by LC3-mediated processes, and ubiquitination emerge as essential mechanisms for mitochondrial health, protein clearance, and oocyte survival processes impaired in infertility. Neutralizing IL-6 (with tocilizumab) may mitigate inflammation-driven endometrial damage and ovarian dysfunction. miR-146a-5p mimics may suppress inflammation and enhance autophagy, while miR-9-3p/5p mimics may restore autophagy-ubiquitination balance to improve oocyte quality. Also, activating LC3-mediated pathways (via rapamycin analogs) may rescue follicular atresia and oxidative stress in aging ovaries.
5. Conclusions and Perspectives
This study identifies novel regulatory axes with translational potential. The dual role of miR-146a-5p in suppressing inflammation (via TRAF6/NFκB) and promoting autophagy offers a unique therapeutic avenue for endometriosis [10]. Similarly, miR-9-3p mimics could enhance autophagic clearance in ovarian follicles, while miR-9-5p modulation might stabilize SQSTM1/p62-LC3B interactions to mitigate protein aggregation in aging ovaries [22]. IL6 inhibitors (tocilizumab) combined with miRNA-based therapies could address multifactorial infertility by simultaneously dampening inflammation and enhancing autophagy [86]. While our bioinformatics approach provides robust insights, which have been in part validated by some experimental studies, more experimentation is also required. For instance, luciferase assays could confirm miRNA-gene targeting [87], and in vitro models of ovarian/endometrial cells could clarify tissue-specific effects. Additionally, pathways like "Shigellosis" (Figure 10D) and "SARS-CoV-1 infection," (Figure 10E) though statistically enriched, lack clear biological relevance to infertility and warrant cautious interpretation [88]. Future studies may prioritize clinical cohorts to validate these networks and explore immune-autophagy crosstalk suggested by terms like "PD-1 checkpoint pathway."
Author Contributions
Conceptualization, A.H. Harrath; Investigation, S. Nehdi; Methodology, S. Nehdi, M. Arafah and A.H. Harrath; Supervision, M. Arafah and A.H. Harrath; Writing – original draft, S. Nahdi and A.H. Harrath; Writing – review & editing, S. Nahdi, M. Arafa and A.H. Harrath.
Funding
This work was funded by Ongoing Research Funding Program (ORF-2025-17) at King Saud University, Riyadh.
Institutional Review Board Statement
Not applicable.
Data availability statement
The data that support the findings of this study are available from the corresponding author (Abdel Halim Harrath) upon reasonable request.
Acknowledgments
The authors extend their appreciation to Ongoing Research Funding Program (ORF-2025-17) at King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carson, S.A. and A.N. Kallen, Diagnosis and Management of Infertility: A Review. JAMA, 2021. 326(1): p. 65-76.
- Agarwal, A., S. Gupta, and R. Sharma, Oxidative stress and its implications in female infertility - a clinician's perspective. Reprod Biomed Online, 2005. 11(5): p. 641-50. [CrossRef]
- Li, L., et al., GnRH agonist treatment regulates IL-6 and IL-11 expression in endometrial stromal cells for patients with HRT regiment in frozen embryo transfer cycles. Reprod Biol, 2022. 22(2): p. 100608. [CrossRef]
- Rudnicka, E., et al., Chronic Low Grade Inflammation in Pathogenesis of PCOS. Int J Mol Sci, 2021. 22(7). [CrossRef]
- Tan, B. and J. Wang, Role of IL-6 in Physiology and Pathology of the Ovary. CEOG, 2024. 51(9). [CrossRef]
- Banerjee, S., L.G. Cooney, and A.K. Stanic, Immune Dysfunction in Polycystic Ovary Syndrome. Immunohorizons, 2023. 7(5): p. 323-332. [CrossRef]
- He, C., et al., Roles of noncoding RNA in reproduction. Frontiers in genetics, 2021. 12: p. 777510.
- Fabová, Z., et al., Does the miR-105–1-Kisspeptin Axis Promote Ovarian Cell Functions? Reproductive Sciences, 2024. 31(8): p. 2293-2308. [CrossRef]
- Sirotkin, A.V., et al., MicroRNAs control transcription factor NF-kB (p65) expression in human ovarian cells. Functional & Integrative Genomics, 2015. 15(3): p. 271-275. [CrossRef]
- Carletti, M.Z., S.D. Fiedler, and L.K. Christenson, MicroRNA 21 blocks apoptosis in mouse periovulatory granulosa cells. Biol Reprod, 2010. 83(2): p. 286-95. [CrossRef]
- Ashrafnezhad, Z., et al., Evaluating the Differential Expression of miR-146a, miR-222, and miR-9 in Matched Serum and Follicular Fluid of Polycystic Ovary Syndrome Patients: Profiling and Predictive Value. Int J Mol Cell Med, 2022. 11(4): p. 320-333. [CrossRef]
- Heidarzadehpilehrood, R. and M. Pirhoushiaran, Biomarker potential of competing endogenous RNA networks in Polycystic Ovary Syndrome (PCOS). Noncoding RNA Res, 2024. 9(2): p. 624-640. [CrossRef]
- Hawkins, S.M., et al., Functional microRNA involved in endometriosis. Mol Endocrinol, 2011. 25(5): p. 821-32. [CrossRef]
- Faraji, M., et al., Elevated expression of microRNA-155, microRNA-383, and microRNA-9 in Iranian patients with polycystic ovary syndrome. Biochem Biophys Rep, 2025. 42: p. 101997. [CrossRef]
- Kong, F., C. Du, and Y. Wang, MicroRNA-9 affects isolated ovarian granulosa cells proliferation and apoptosis via targeting vitamin D receptor. Mol Cell Endocrinol, 2019. 486: p. 18-24. [CrossRef]
- Choi, A.M.K., S.W. Ryter, and B. Levine, Autophagy in Human Health and Disease. New England Journal of Medicine, 2013. 368(7): p. 651-662.
- Kumariya, S., et al., Autophagy in ovary and polycystic ovary syndrome: role, dispute and future perspective. Autophagy, 2021. 17(10): p. 2706-2733. [CrossRef]
- Ji, R., et al., BOP1 contributes to the activation of autophagy in polycystic ovary syndrome via nucleolar stress response. Cell Mol Life Sci, 2024. 81(1): p. 101. [CrossRef]
- Mizushima, N. and M. Komatsu, Autophagy: renovation of cells and tissues. Cell, 2011. 147(4): p. 728-41. [CrossRef]
- Tsakiri, E.N. and I.P. Trougakos, The amazing ubiquitin-proteasome system: structural components and implication in aging. Int Rev Cell Mol Biol, 2015. 314: p. 171-237.
- Sun, L., et al., The E3 Ubiquitin Ligase SYVN1 Plays an Antiapoptotic Role in Polycystic Ovary Syndrome by Regulating Mitochondrial Fission. Oxid Med Cell Longev, 2022. 2022: p. 3639302. [CrossRef]
- Levine, B. and G. Kroemer, Biological Functions of Autophagy Genes: A Disease Perspective. Cell, 2019. 176(1-2): p. 11-42. [CrossRef]
- McIlvenna, L.C., et al., Transforming growth factor beta1 impairs the transcriptomic response to contraction in myotubes from women with polycystic ovary syndrome. J Physiol, 2022. 600(14): p. 3313-3330. [CrossRef]
- Liu, X., et al., Novel PGK1 determines SKP2-dependent AR stability and reprograms granular cell glucose metabolism facilitating ovulation dysfunction. EBioMedicine, 2020. 61: p. 103058.
- Subramanian, A., et al., Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A, 2005. 102(43): p. 15545-50. [CrossRef]
- Kim, C.K., et al., Differential Stability of miR-9-5p and miR-9-3p in the Brain Is Determined by Their Unique Cis- and Trans-Acting Elements. eNeuro, 2020. 7(3).
- Iacona, J.R. and C.S. Lutz, miR-146a-5p: Expression, regulation, and functions in cancer. Wiley Interdiscip Rev RNA, 2019. 10(4): p. e1533.
- Ru, Y., et al., The multiMiR R package and database: integration of microRNA-target interactions along with their disease and drug associations. Nucleic Acids Res, 2014. 42(17): p. e133. [CrossRef]
- Barrett, T., et al., NCBI GEO: archive for functional genomics data sets--update. Nucleic Acids Res, 2013. 41(Database issue): p. D991-5. [CrossRef]
- Edgar, R., M. Domrachev, and A.E. Lash, Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res, 2002. 30(1): p. 207-10. [CrossRef]
- Dusa, C.-H.G.a.A., ggVennDiagram: A 'ggplot2' Implement of Venn Diagram. 2024.
- Yu, G., et al., clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS, 2012. 16(5): p. 284-7. [CrossRef]
- Shannon, P., et al., Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res, 2003. 13(11): p. 2498-504. [CrossRef]
- Kuleshov, M.V., et al., Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res, 2016. 44(W1): p. W90-7. [CrossRef]
- Kanehisa, M. and S. Goto, KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res, 2000. 28(1): p. 27-30. [CrossRef]
- Szklarczyk, D., et al., The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res, 2023. 51(D1): p. D638-D646. [CrossRef]
- Ham, J., W. Lim, and G. Song, Ethalfluralin impairs implantation by aggravation of mitochondrial viability and function during early pregnancy. Environ Pollut, 2022. 307: p. 119495. [CrossRef]
- Tuncdemir, M., et al., NFKB1 rs28362491 and pre-miRNA-146a rs2910164 SNPs on E-Cadherin expression in case of idiopathic oligospermia: A case-control study. Int J Reprod Biomed, 2018. 16(4): p. 247-254. [CrossRef]
- Virakul, S., et al., Basic FGF and PDGF-BB synergistically stimulate hyaluronan and IL-6 production by orbital fibroblasts. Mol Cell Endocrinol, 2016. 433: p. 94-104. [CrossRef]
- Li, W. and Y. Zheng, MicroRNAs in Extracellular Vesicles of Alzheimer's Disease. Cells, 2023. 12(10). [CrossRef]
- Bros, M., et al., Differentially Tolerized Mouse Antigen Presenting Cells Share a Common miRNA Signature Including Enhanced mmu-miR-223-3p Expression Which Is Sufficient to Imprint a Protolerogenic State. Front Pharmacol, 2018. 9: p. 915. [CrossRef]
- Bonacini, M., et al., miR-146a and miR-146b regulate the expression of ICAM-1 in giant cell arteritis. J Autoimmun, 2024. 144: p. 103186. [CrossRef]
- Pakdaman Kolour, S.S., et al., Extracecellulr vesicles (EVs) microRNAs (miRNAs) derived from mesenchymal stem cells (MSCs) in osteoarthritis (OA); detailed role in pathogenesis and possible therapeutics. Heliyon, 2025. 11(3): p. e42258. [CrossRef]
- Jia, W., et al., Immune-related gene methylation prognostic instrument for stratification and targeted treatment of ovarian cancer patients toward advanced 3PM approach. EPMA Journal, 2024. 15(2): p. 375-404. [CrossRef]
- Kraczkowska, W. and P.P. Jagodzinski, The Long Non-Coding RNA Landscape of Atherosclerotic Plaques. Mol Diagn Ther, 2019. 23(6): p. 735-749. [CrossRef]
- Faizal, A.M., et al., Unravelling the role of HAS2, GREM1, and PTGS2 gene expression in cumulus cells: implications for human oocyte development competency - a systematic review and integrated bioinformatic analysis. Front Endocrinol (Lausanne), 2024. 15: p. 1274376. [CrossRef]
- Broi, M.G.D., et al., Expression of PGR, HBEGF, ITGAV, ITGB3 and SPP1 genes in eutopic endometrium of infertile women with endometriosis during the implantation window: a pilot study. JBRA Assist Reprod, 2017. 21(3): p. 196-202.
- Liao, C., et al., Effect of Chitosan-assisted Combination of Laparoscope and Hysteroscope on the Levels of IFN-gamma and ICAM-1 in Treatment of Infertility Caused by Obstruction of Fallopian Tubes. Cell Mol Biol (Noisy-le-grand), 2023. 69(4): p. 101-104. [CrossRef]
- Padmanabhan, R.A., et al., Autoimmune regulator (AIRE): Takes a hypoxia-inducing factor 1A (HIF1A) route to regulate FOXP3 expression in PCOS. Am J Reprod Immunol, 2023. 89(2): p. e13637. [CrossRef]
- Samare-Najaf, M., et al., The constructive and destructive impact of autophagy on both genders' reproducibility, a comprehensive review. Autophagy, 2023. 19(12): p. 3033-3061. [CrossRef]
- Jenabi, M., et al., Evaluation of expression CXCL8 chemokine and its relationship with oocyte maturation and embryo quality in the intracytoplasmic sperm injection method. Mol Biol Rep, 2022. 49(9): p. 8413-8427. [CrossRef]
- Peters, A.E., et al., Autophagy in Female Fertility: A Role in Oxidative Stress and Aging. Antioxid Redox Signal, 2020. 32(8): p. 550-568. [CrossRef]
- Rahman, M.A., et al., An Update on the Study of the Molecular Mechanisms Involved in Autophagy during Bacterial Pathogenesis. Biomedicines, 2024. 12(8).
- Rahman, M.A., et al., Molecular Insights into the Multifunctional Role of Natural Compounds: Autophagy Modulation and Cancer Prevention. Biomedicines, 2020. 8(11).
- Popli, P. and R. Kommagani, Autophagy is required for stem-cell-mediated endometrial programming and the establishment of pregnancy. Autophagy, 2024. 20(4): p. 970-972. [CrossRef]
- Rahman, M.A., et al., Therapeutic Aspects and Molecular Targets of Autophagy to Control Pancreatic Cancer Management. Biomedicines, 2022. 10(6).
- Rahman, M.A., et al., Autophagy Modulation in Aggresome Formation: Emerging Implications and Treatments of Alzheimer's Disease. Biomedicines, 2022. 10(5).
- Harrath, A.H., et al., Autophagy and Female Fertility: Mechanisms, Clinical Implications, and Emerging Therapies. Cells, 2024. 13(16). [CrossRef]
- Cheng, D., et al., The Roles of Autophagy in the Genesis and Development of Polycystic Ovary Syndrome. Reprod Sci, 2023. 30(10): p. 2920-2931. [CrossRef]
- Liu, X., et al., miR-146a-5p Plays an Oncogenic Role in NSCLC via Suppression of TRAF6. Front Cell Dev Biol, 2020. 8: p. 847. [CrossRef]
- Vergani, E., et al., miR-146a-5p impairs melanoma resistance to kinase inhibitors by targeting COX2 and regulating NFkB-mediated inflammatory mediators. Cell Commun Signal, 2020. 18(1): p. 156.
- Chen, Z., Q. Gu, and R. Chen, miR-146a-5p regulates autophagy and NLRP3 inflammasome activation in epithelial barrier damage in the in vitro cell model of ulcerative colitis through the RNF8/Notch1/mTORC1 pathway. Immunobiology, 2023. 228(4): p. 152386. [CrossRef]
- Gozuacik, D., et al., Autophagy-Regulating microRNAs and Cancer. Front Oncol, 2017. 7: p. 65.
- Guan, X., et al., Mechanistic Insights into Selective Autophagy Subtypes in Alzheimer's Disease. Int J Mol Sci, 2022. 23(7). [CrossRef]
- Deng, T., et al., TRAF6 autophagic degradation by avibirnavirus VP3 inhibits antiviral innate immunity via blocking NFKB/NF-kappaB activation. Autophagy, 2022. 18(12): p. 2781-2798. [CrossRef]
- Yamada, T., et al., SQSTM1/p62 promotes mitochondrial ubiquitination independently of PINK1 and PRKN/parkin in mitophagy. Autophagy, 2019. 15(11): p. 2012-2018. [CrossRef]
- Dinsdale, N.L. and B.J. Crespi, Endometriosis and polycystic ovary syndrome are diametric disorders. Evolutionary applications, 2021. 14(7): p. 1693-1715. [CrossRef]
- Taganov, K.D., et al., NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci U S A, 2006. 103(33): p. 12481-6. [CrossRef]
- Harrath, A.H., et al., Autophagy and female fertility: mechanisms, clinical implications, and emerging therapies. Cells, 2024. 13(16): p. 1354. [CrossRef]
- Harrath, A.H., et al., Benzene exposure causes structural and functional damage in rat ovaries: occurrence of apoptosis and autophagy. Environmental Science and Pollution Research, 2022. 29(50): p. 76275-76285. [CrossRef]
- Jalouli, M., et al., Allethrin promotes apoptosis and autophagy associated with the oxidative stress-related PI3K/AKT/mTOR signaling pathway in developing rat ovaries. International Journal of Molecular Sciences, 2022. 23(12): p. 6397. [CrossRef]
- Oală, I.E., et al., Endometriosis and the role of pro-inflammatory and anti-inflammatory cytokines in pathophysiology: a narrative review of the literature. Diagnostics, 2024. 14(3): p. 312. [CrossRef]
- Hannan, N.J., J. Evans, and L.A. Salamonsen, Alternate roles for immune regulators: establishing endometrial receptivity for implantation. Expert Rev Clin Immunol, 2011. 7(6): p. 789-802. [CrossRef]
- Kuessel, L., et al., Soluble VCAM-1/soluble ICAM-1 ratio is a promising biomarker for diagnosing endometriosis. Hum Reprod, 2017. 32(4): p. 770-779. [CrossRef]
- Lee, R.C., R.L. Feinbaum, and V. Ambros, The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. cell, 1993. 75(5): p. 843-854. [CrossRef]
- Toloubeydokhti, T., et al., The expression and ovarian steroid regulation of endometrial micro-RNAs. Reproductive sciences, 2008. 15: p. 993-1001. [CrossRef]
- Ohlsson Teague, E.M.C., et al., MicroRNA-regulated pathways associated with endometriosis. Molecular endocrinology, 2009. 23(2): p. 265-275. [CrossRef]
- Filigheddu, N., et al., Differential expression of microRNAs between eutopic and ectopic endometrium in ovarian endometriosis. BioMed Research International, 2010. 2010(1): p. 369549. [CrossRef]
- Zhou, Z., et al., Long noncoding RNA SNHG16 targets miR-146a-5p/CCL5 to regulate LPS-induced WI-38 cell apoptosis and inflammation in acute pneumonia. Life sciences, 2019. 228: p. 189-197. [CrossRef]
- Yan, F., et al., MicroRNA miR-146a-5p inhibits the inflammatory response and injury of airway epithelial cells via targeting TNF receptor-associated factor 6. Bioengineered, 2021. 12(1): p. 1916-1926. [CrossRef]
- Yu, F., et al., Epigenetically-Regulated MicroRNA-9-5p Suppresses the Activation of Hepatic Stellate Cells via TGFBR1 and TGFBR2. Cell Physiol Biochem, 2017. 43(6): p. 2242-2252. [CrossRef]
- Morita, Y., et al., Caspase-2 deficiency prevents programmed germ cell death resulting from cytokine insufficiency but not meiotic defects caused by loss of ataxia telangiectasia-mutated (Atm) gene function. Cell Death Differ, 2001. 8(6): p. 614-20. [CrossRef]
- Zheng, J., et al., Decreased expression of HOXA10 may activate the autophagic process in ovarian endometriosis. Reproductive Sciences, 2018. 25(9): p. 1446-1454. [CrossRef]
- Allavena, G., et al., Autophagy is upregulated in ovarian endometriosis: a possible interplay with p53 and heme oxygenase-1. Fertility and sterility, 2015. 103(5): p. 1244-1251. e1. [CrossRef]
- Yang, H.-L., et al., Autophagy in endometriosis. American Journal of Translational Research, 2017. 9(11): p. 4707.
- Genovese, M.C., et al., Interleukin-6 receptor inhibition with tocilizumab reduces disease activity in rheumatoid arthritis with inadequate response to disease-modifying antirheumatic drugs: the tocilizumab in combination with traditional disease-modifying antirheumatic drug therapy study. Arthritis Rheum, 2008. 58(10): p. 2968-80. [CrossRef]
- Rottiers, V. and A.M. Naar, MicroRNAs in metabolism and metabolic disorders. Nat Rev Mol Cell Biol, 2012. 13(4): p. 239-50.
- Kanehisa, M., et al., KEGG: integrating viruses and cellular organisms. Nucleic Acids Res, 2021. 49(D1): p. D545-D551. [CrossRef]
Figure 1.
Flowchart of the integrated analysis study.
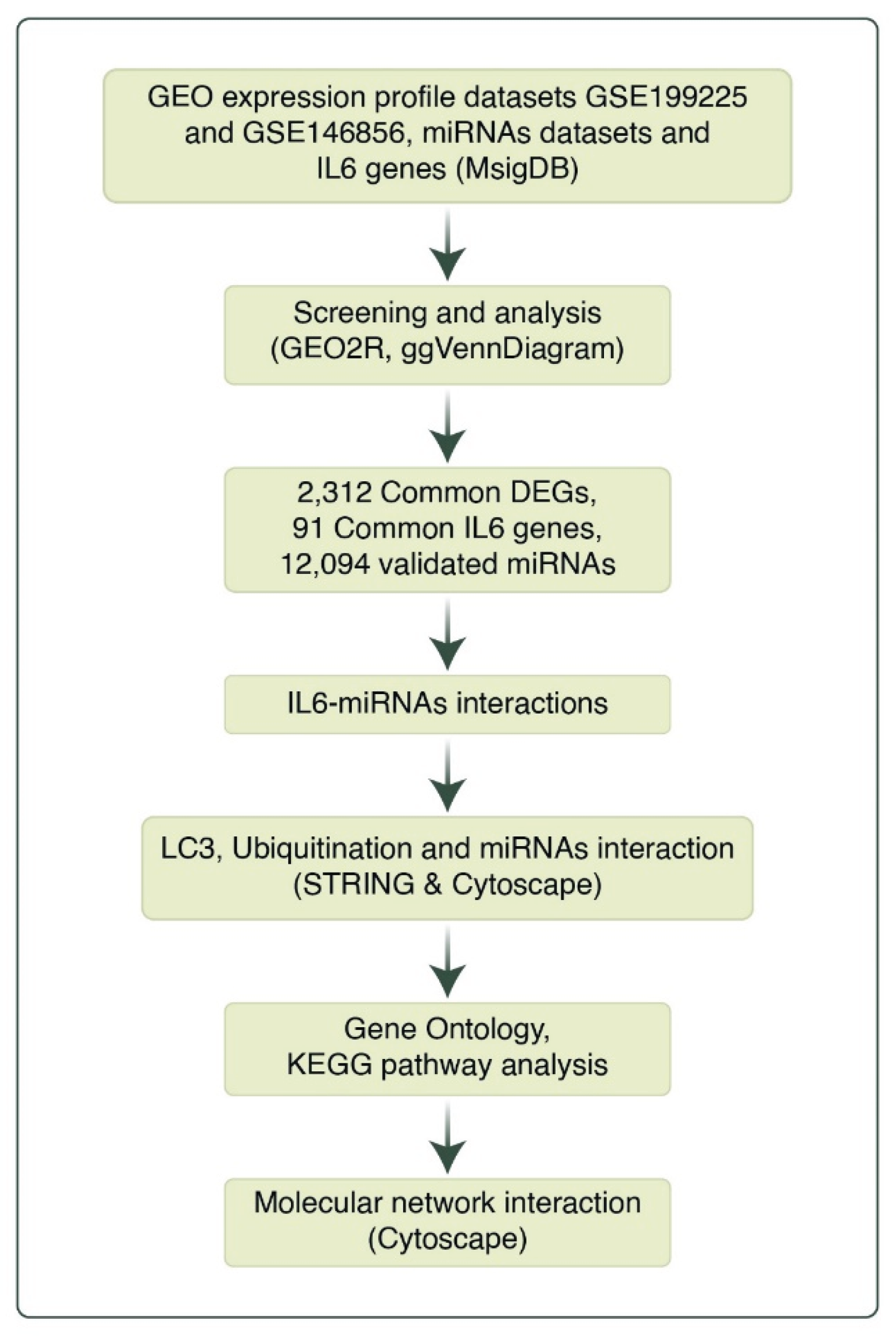
Figure 2.
Volcano Plot of Differentially Expressed Genes in GSE199225 and GSE146856: Identification of Up-regulated and Down-regulated Genes.
Figure 2.
Volcano Plot of Differentially Expressed Genes in GSE199225 and GSE146856: Identification of Up-regulated and Down-regulated Genes.
Figure 3.
DEGs screening via intersection analysis. (A) upregulated genes and (B) downregulated genes in the control group. (C) upregulated genes and (D) downregulated genes in the PCOS group.
Figure 3.
DEGs screening via intersection analysis. (A) upregulated genes and (B) downregulated genes in the control group. (C) upregulated genes and (D) downregulated genes in the PCOS group.
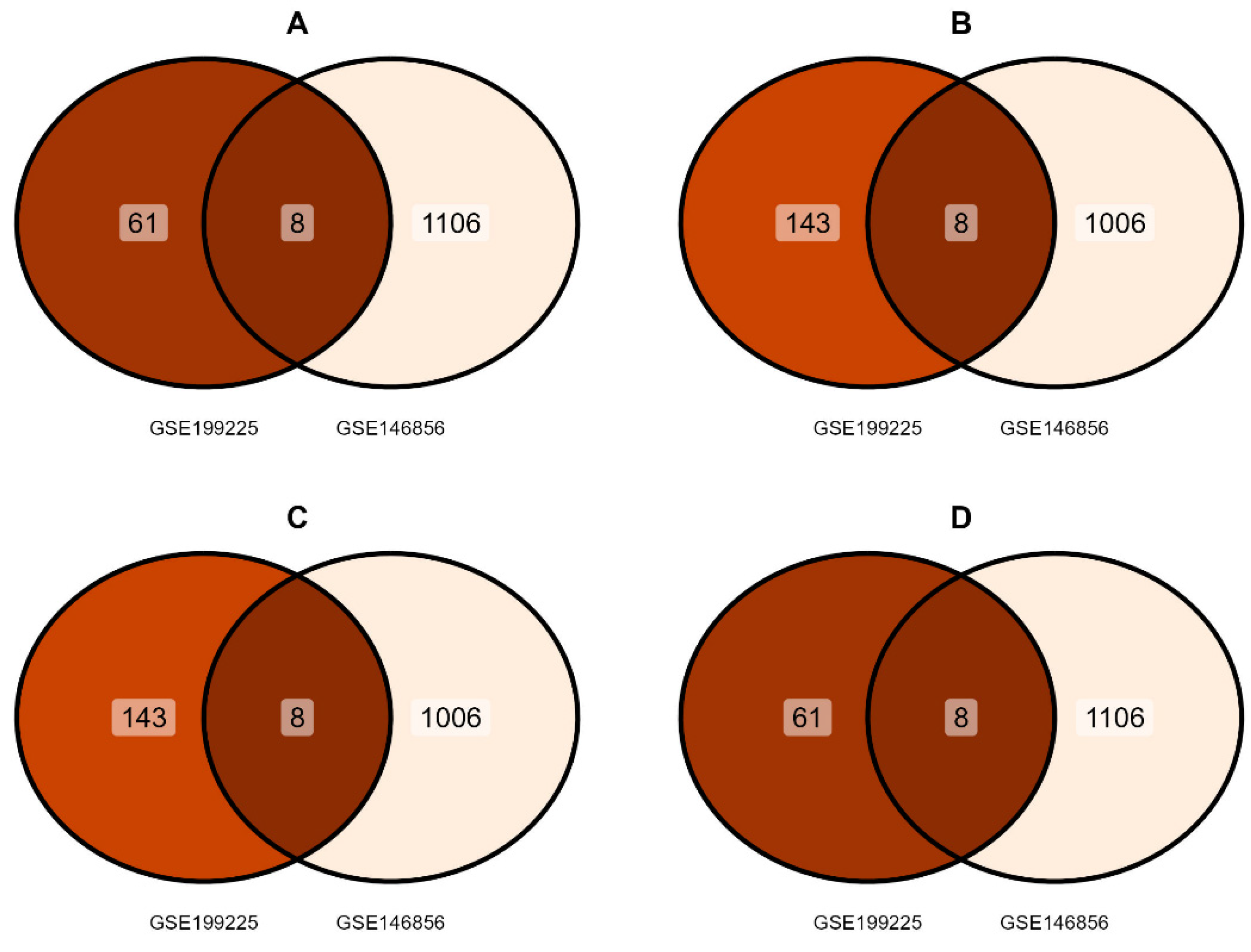
Figure 4.
KEGG pathway enrichment analysis of the predicted target genes for miR-146a-5p, miR-9-3p, and miR-9-5p. The analysis reveals significant associations with pathways related to inflammation, autophagy, and ubiquitination.
Figure 4.
KEGG pathway enrichment analysis of the predicted target genes for miR-146a-5p, miR-9-3p, and miR-9-5p. The analysis reveals significant associations with pathways related to inflammation, autophagy, and ubiquitination.
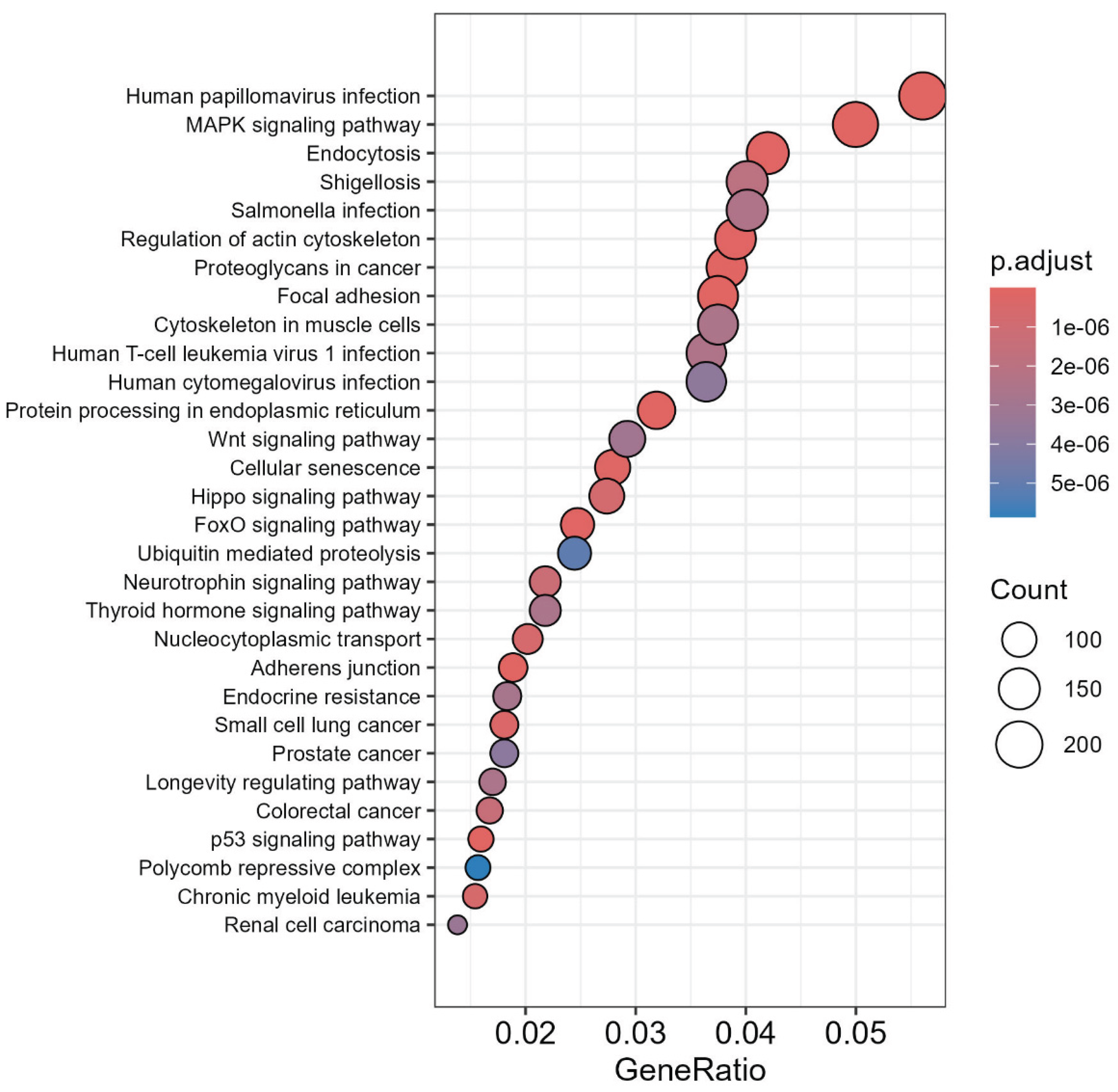
Figure 5.
Interaction Between DEGs, miRNAs, and IL6 Signaling. (A) IL6 targeted genes in the DEGs. (B) DEGs overlapped IL6 genes in the miRNAs.
Figure 5.
Interaction Between DEGs, miRNAs, and IL6 Signaling. (A) IL6 targeted genes in the DEGs. (B) DEGs overlapped IL6 genes in the miRNAs.
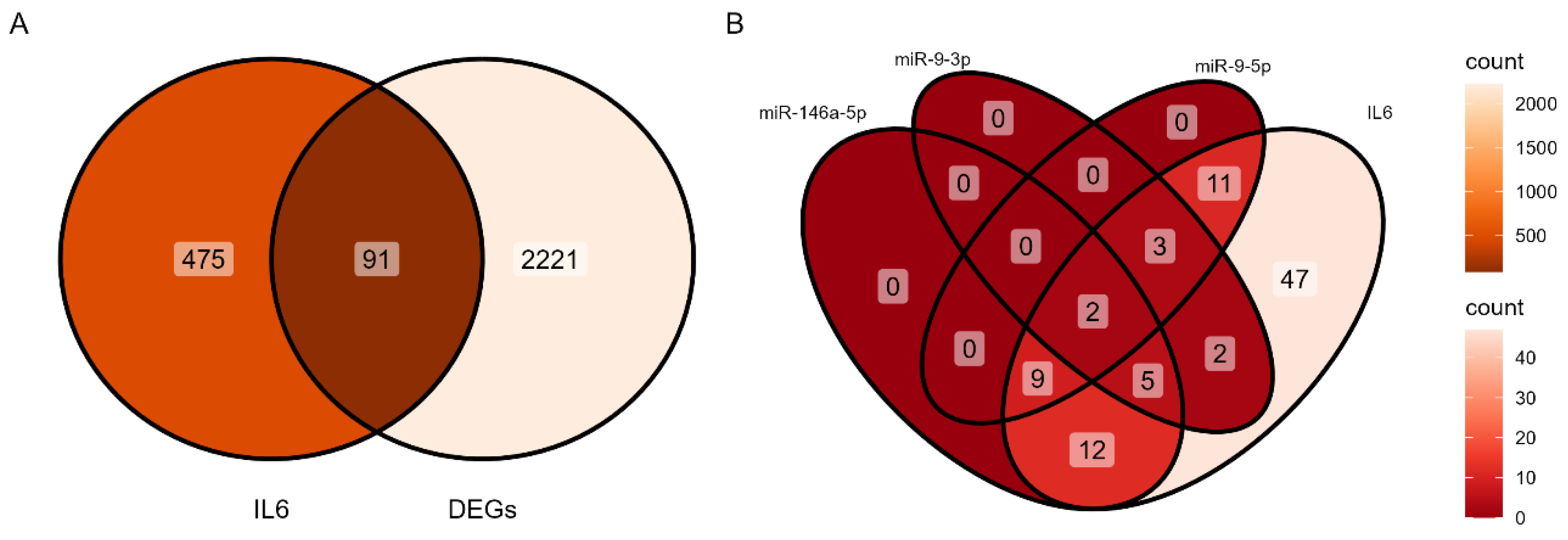
Figure 6.
Regulatory network of miRNAs and their target genes within the IL-6 signaling pathway that are dysregulated in female infertility. The model shows how miR-146a-5p, miR-9-3p, and miR-9-5p potentially regulate the shared IL-6/DEG targets.
Figure 6.
Regulatory network of miRNAs and their target genes within the IL-6 signaling pathway that are dysregulated in female infertility. The model shows how miR-146a-5p, miR-9-3p, and miR-9-5p potentially regulate the shared IL-6/DEG targets.
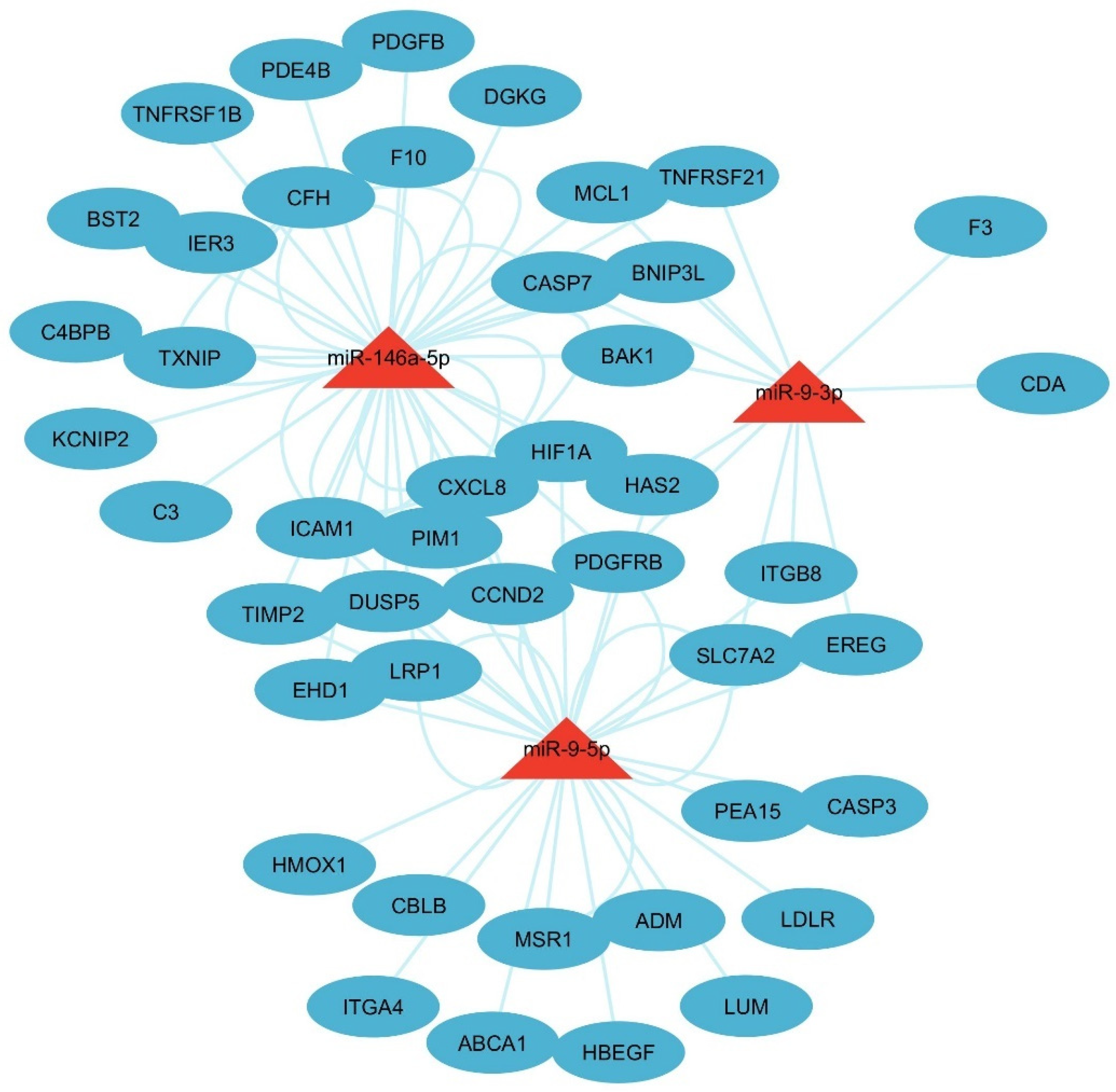
Figure 7.
Key IL6 genes associated with apoptosis, female infertility and the miRs targeted genes enrichment analysis. (A) Biological process. (B) Cellular components. (C) Molecular functions. (D) MSigDB Hallmark pathways. (E) KEGG pathways. (F) Reactome pathways. Pathways were considered significant at p < 0.05 based on Enrichr's statistical output.
Figure 7.
Key IL6 genes associated with apoptosis, female infertility and the miRs targeted genes enrichment analysis. (A) Biological process. (B) Cellular components. (C) Molecular functions. (D) MSigDB Hallmark pathways. (E) KEGG pathways. (F) Reactome pathways. Pathways were considered significant at p < 0.05 based on Enrichr's statistical output.
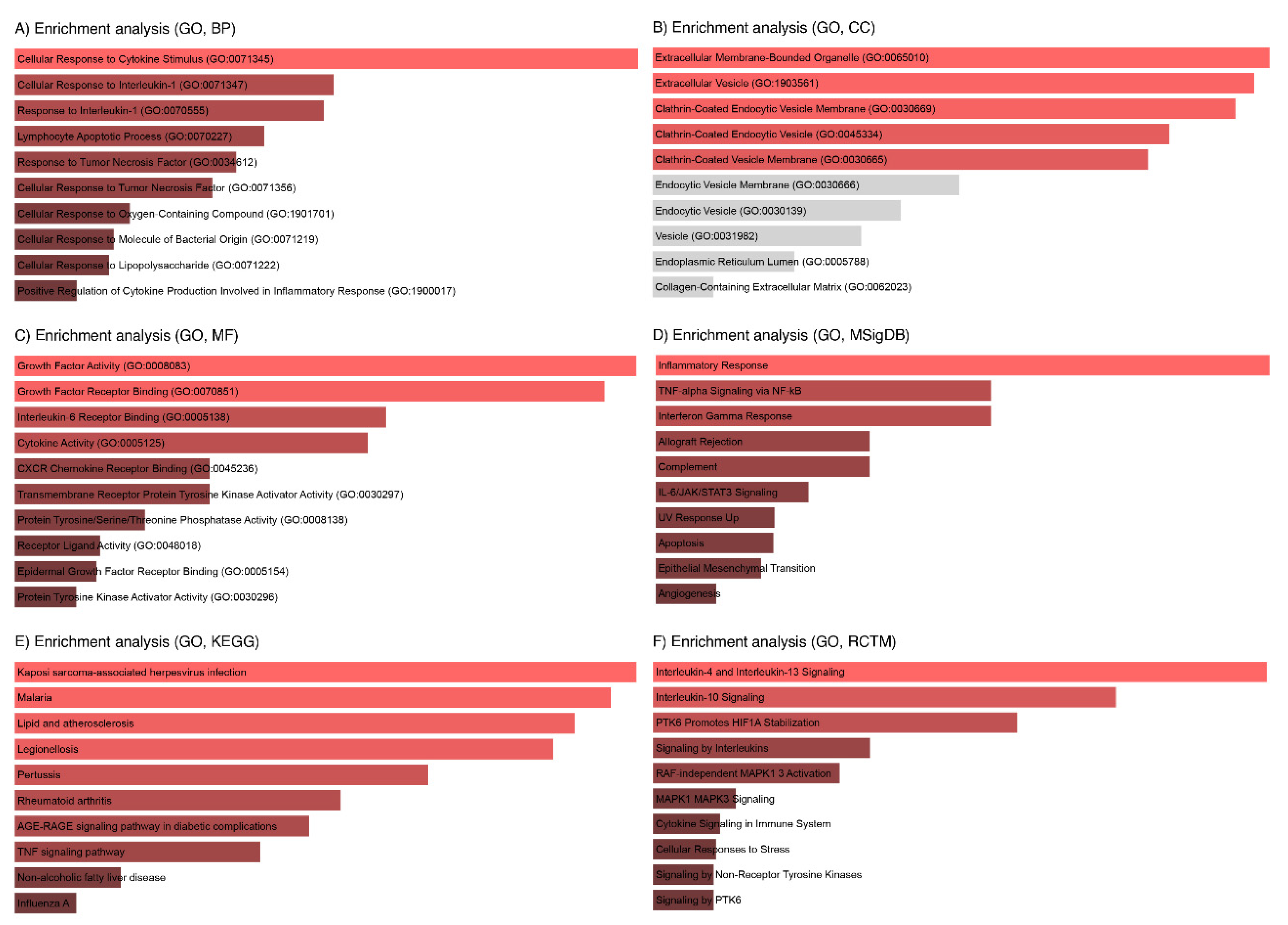
Figure 8.
Gene Ontology (GO) enrichment of the key autophagy genes.
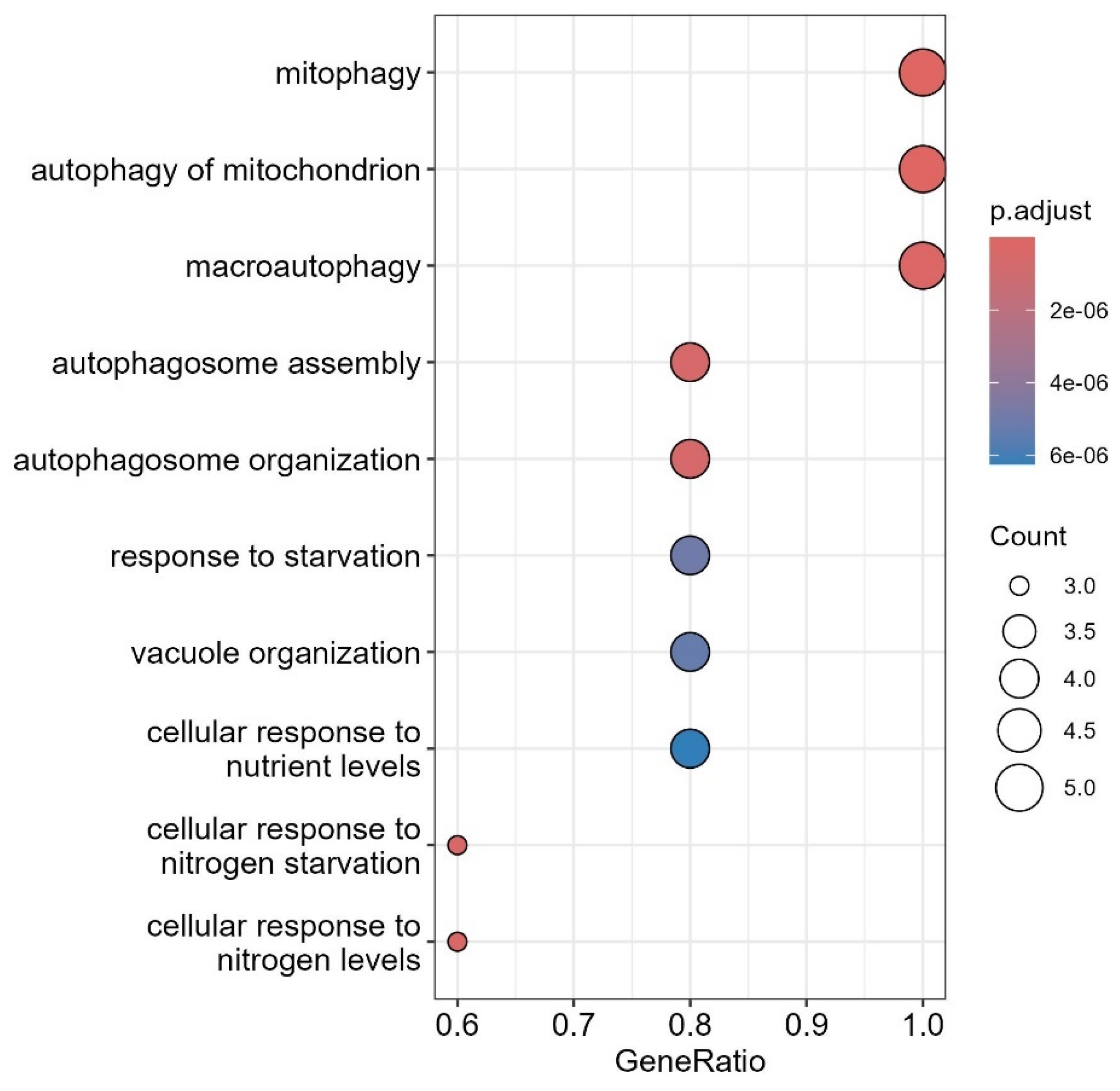
Figure 9.
miRNAs interactions with autophagy, ubiquitination and upstream signaling regulators.
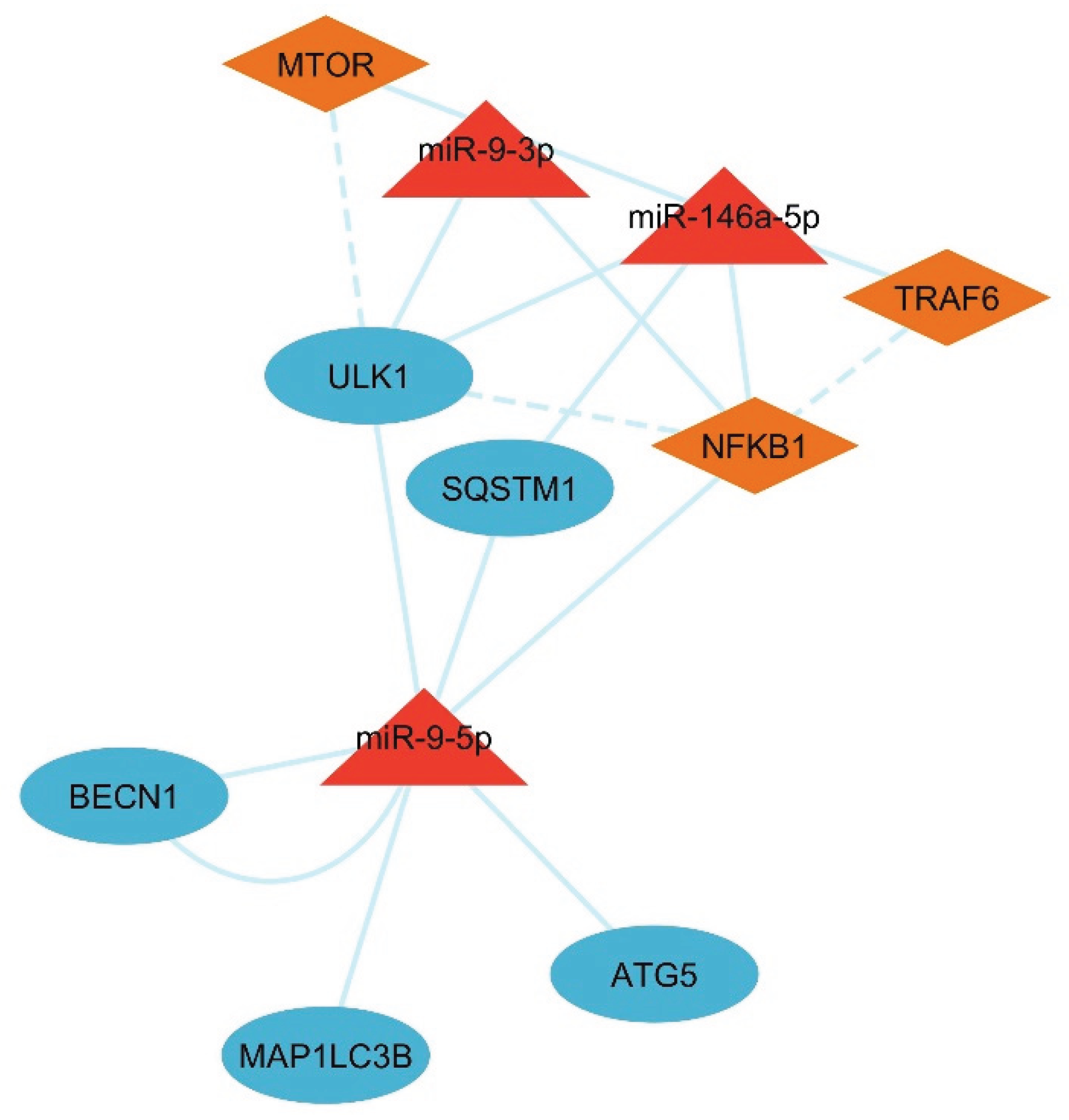
Figure 10.
LC3-Ubiquitination and miRNAs targeted genes enrichment analysis. (A) Biological process. (B) Cellular components. (C) Molecular functions. (D) KEGG pathways. (E) Reactome pathways. Enriched terms shown met statistical significance thresholds (p < 0.05) as calculated by Enrichr.
Figure 10.
LC3-Ubiquitination and miRNAs targeted genes enrichment analysis. (A) Biological process. (B) Cellular components. (C) Molecular functions. (D) KEGG pathways. (E) Reactome pathways. Enriched terms shown met statistical significance thresholds (p < 0.05) as calculated by Enrichr.
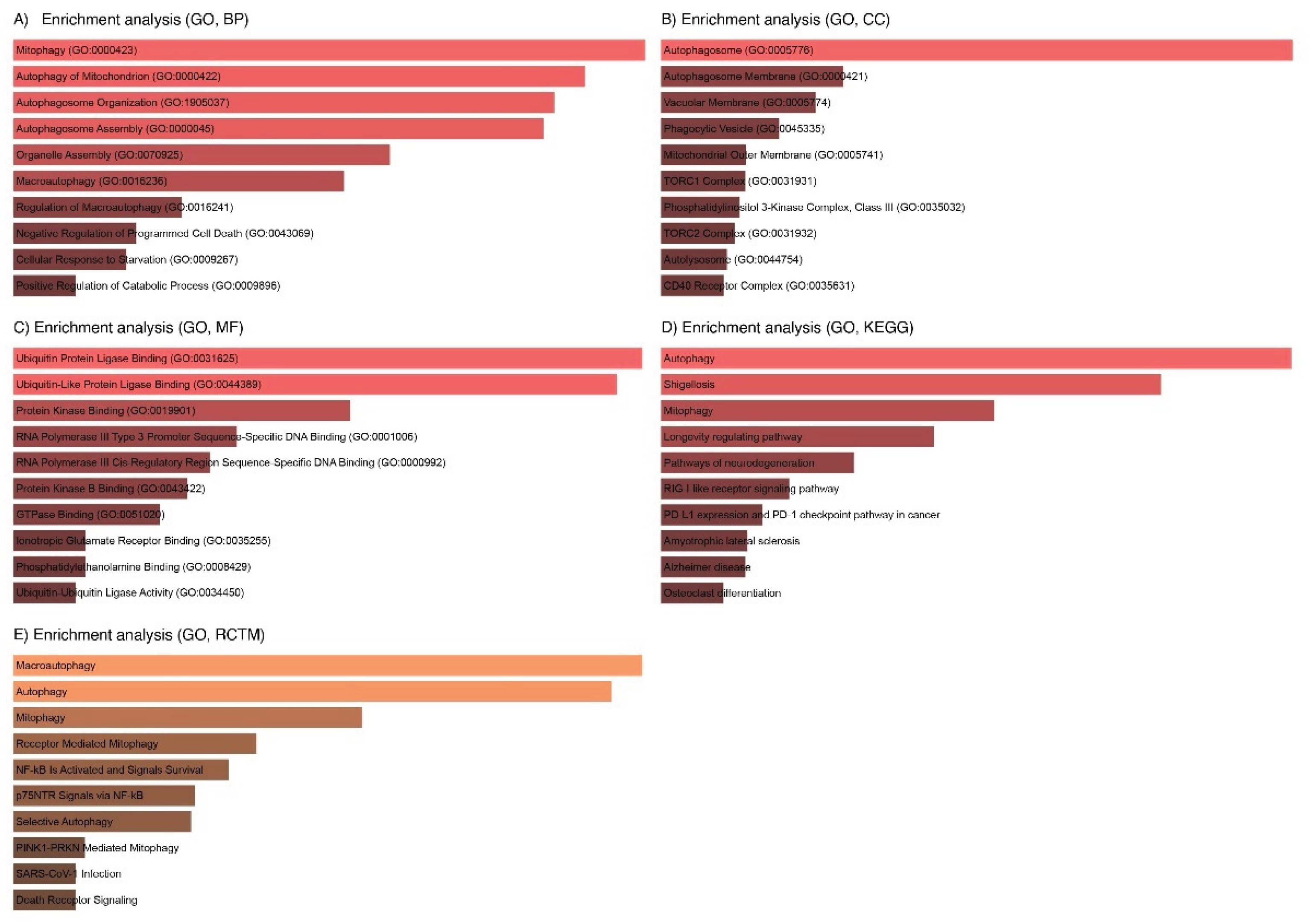
Figure 11.
Clustered Associations of Autophagy, Inflammation, and Ubiquitination Genes with Key Biological Processes.
Figure 11.
Clustered Associations of Autophagy, Inflammation, and Ubiquitination Genes with Key Biological Processes.
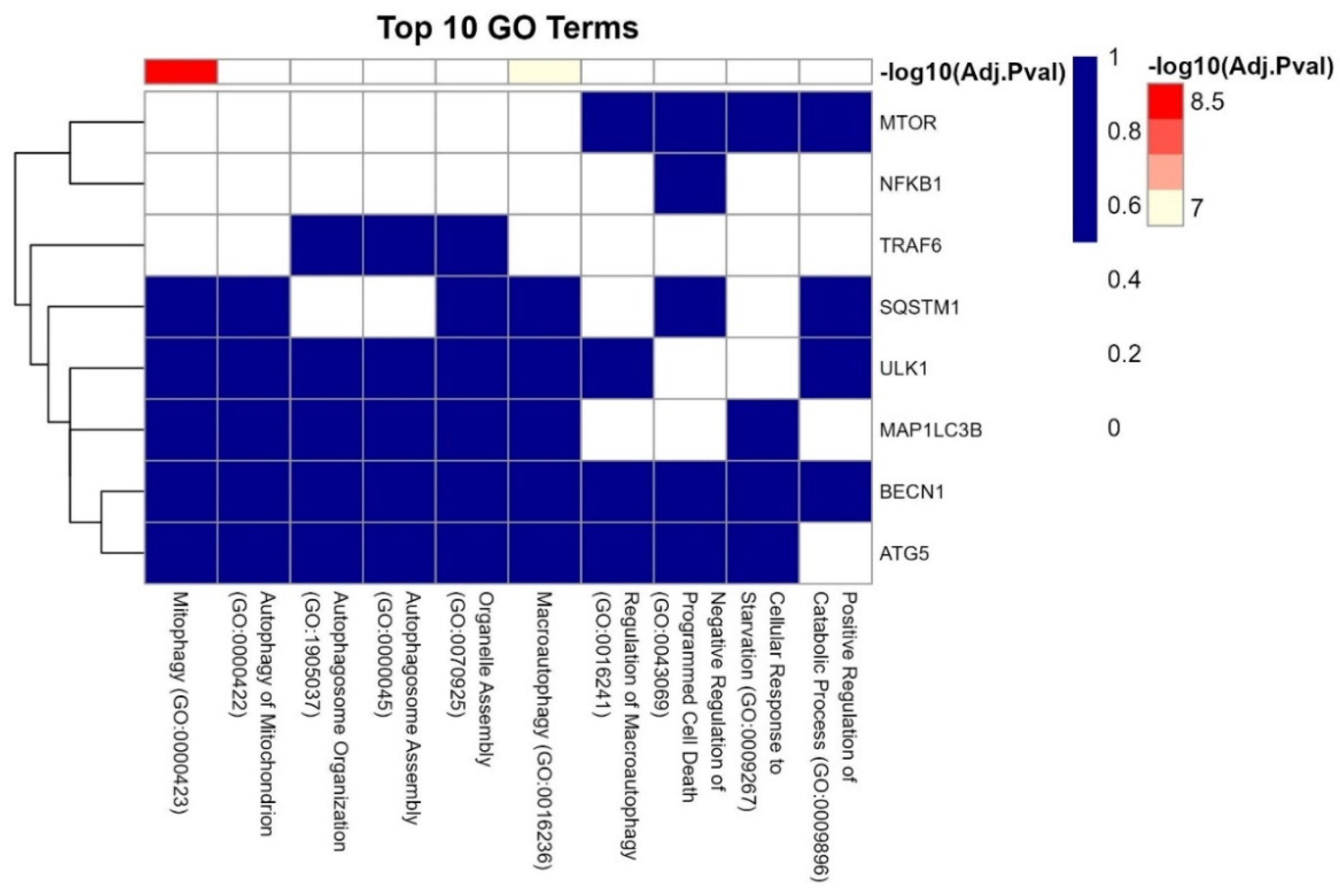

Table 1.
Key Autophagy/ubiquitination Genes and Their Roles in Female Infertility.
| Gene | Functional Role | Associated Reproductive Process | Reference |
| MAP1LC3B | Autophagosome membrane formation | Impaired oocyte maturation, embryo quality | [53] |
| ATG5 | Autophagosome elongation (ATG5-ATG12 complex) | Follicular atresia, granulosa cell apoptosis | [54] |
| BECN1 | Autophagy initiation (PI3K complex) | Dysregulated autophagy in endometrial cells | [55, 56] |
| SQSTM1 | Selective autophagy of ubiquitinated proteins | Protein aggregate clearance in ovarian cells | [53, 57] |
| ULK1 | Master kinase regulating autophagy initiation | Stress response in cumulus cells | [58, 59] |
Table 2.
Top 10 significant p-values and q-values for the protein coding genes in KEGG/Reactome Pathways 2024.
Table 2.
Top 10 significant p-values and q-values for the protein coding genes in KEGG/Reactome Pathways 2024.
| Term | p-Value | q-Value | Overlap Genes |
| Macroautophagy | 3.325892e-12 | 5.372732e-10 | BECN1, MAP1LC3B, ULK1, SQSTM1, MTOR, ATG5 |
| Autophagy | 6.105378e-12 | 5.372732e-10 | BECN1, MAP1LC3B, ULK1, SQSTM1, MTOR, ATG5 |
| Mitophagy | 8.588874e-10 | 5.038806e-08 | MAP1LC3B, ULK1, SQSTM1, ATG5 |
| Receptor-Mediated Mitophagy | 6.919482e-09 | 3.044572e-07 | MAP1LC3B, ULK1, ATG5 |
| NF-kB Is Activated and Signals Survival | 1.198939e-08 | 4.220264e-07 | TRAF6, SQSTM1, NFKB1 |
| p75NTR Signals via NF-kB | 2.346278e-08 | 6.359648e-07 | TRAF6, SQSTM1, NFKB1 |
| Selective Autophagy | 2.529405e-08 | 6.359648e-07 | MAP1LC3B, ULK1, SQSTM1, ATG5 |
| PINK1-PRKN Mediated Mitophagy | 2.071973e-07 | 4.393258e-06 | MAP1LC3B, SQSTM1, ATG5 |
| SARS-CoV-1 Infection | 2.496169e-07 | 4.393258e-06 | BECN1, MAP1LC3B, TRAF6, NFKB1 |
| Death Receptor Signaling | 2.496169e-07 | 4.393258e-06 | TRAF6, ULK1, SQSTM1, NFKB1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



