Submitted:
26 September 2025
Posted:
29 September 2025
You are already at the latest version
Abstract
Objectives: This review provides a comprehensive overview of the complex and dynamic bacterial composition of the human gastrointestinal (GI) microbiota and explores its integral role in the microbiome-gut-brain axis. Methods: It discusses the physiological and molecular pathways through which the gut microbiota communicates with the central nervous system, highlighting key barriers that can impede effective signaling along this axis. The review also delves into the influence of the microbiota on brain health, including cognitive function, mood regulation, and neuroinflammation, and considers how disruptions in this system—known as dysbiosis—can contribute to metabolic and neurological dysfunction. Results: A central focus of the article is the role of the commonly prescribed antidiabetic drug Metformin, not only in regulating glucose metabolism but also in its emerging potential to beneficially modulate the gut microbiome. By doing so, Metformin may indirectly support improved brain health. Conclusions: Ultimately, the article seeks to inform both healthcare practitioners and patients about the promising therapeutic implications of microbiome-targeted strategies, particularly the use of Metformin, in promoting cognitive and neurological well-being.
Keywords:
brain health
; gut-brain axis
; metformin
; metabolic dysfunction
; microbiota
; probiotics
1. Gut Microbiota
This term refers to the various microorganisms residing in the human intestine, whose collective genomes and metabolites form the microbiome [1,2]. Dominant phyla include Bacteroidetes and Firmicutes, which produce short-chain fatty acids (SCFAs) critical for gut integrity and host metabolism [3,4]. These microbes assist in digesting nutrients, synthesizing vitamins, and regulating immune responses and metabolic pathways. They also protect against pathogens by maintaining epithelial barriers and producing antimicrobial compounds [5]. Disruptions in microbial composition (i.e., dysbiosis) contributed to various diseases, prompting interest in probiotics for therapeutic use..
2. Gut-Brain Axis (GBA)
The gut microbiota plays a critical role in the GBA. Microbe-derived signals reach the central nervous system (CNS) either directly through systemic circulation or indirectly via receptors on enteroendocrine cells (EECs), enterochromaffin cells (ECCs) in the gut, or the mucosal immune system [6], which in turn trigger a CNS response through the autonomic nervous system (ANS), and the hypothalamic-pituitary-adrenal (HPA) axis [7]. Strong evidence of microbiota and brain linkages in clinical practice is that gut dysbiosis has been causally associated with neurological illnesses such as autism and anxiety-depressive behaviors, as well as functional gastrointestinal disorders, including irritable bowel syndrome [8,9]. Numerous channels of communication, including immunological, endocrine, neuronal, and metabolic pathways, have been identified along the GBA [10,11,12,13].
2.1. Microbiota and Neurotransmitters Synthesis
Various neurotransmitters, such as acetylcholine, noradrenaline, serotonin, and dopamine are synthesized by numerous bacterial species residing in the gastrointestinal tract [14,15,16,17], contribute to the intricate communication network between the gut and the brain [18]. Gamma-aminobutyric acid (GABA) is synthesized by Bifidobacterium and Lactobacillus species; Noradrenaline is synthesized by the species Bacillus, Escherichia, and Saccharomyces; serotonin by Streptococcus, Escherichia, Candida, and Enterococcus species, while acetylcholine is synthesized by Lactobacillus and dopamine by Bacillus [15]. These microbiota-generated neurotransmitters cross the intestinal mucosal barrier; however, their effects on brain function are thought to be mediated indirectly via interactions with the enteric nervous system (ENS) [19,20].
2.2. Microbiota and Enteroendocrine Signaling
Bacterial metabolites interact with the gut epithelium, prompting EECs to create active amines through the intracellular decarboxylation of active amine precursors. These are subsequently deposited in secretory vesicles [21]. EECs are considered crucial sensors of gut microbiota and/or microbial metabolites, playing a vital role in maintaining mucosal immunity and gut barrier function, visceral hyperalgesia, and gastrointestinal (GI) motility in health and disease [13,22,23,24].
The brain controls the functions of intestine functional effector cells, including enteric neurons, interstitial cells of Cajal, immune cells, epithelial cells, smooth muscle cells, and enterochromaffin cells, through a combination of neurological and hormonal channels of communication. On the other hand, the gut microbiota, which is involved in the gut-brain reciprocal connections, also influences these same cells. It’s currently emerging that a microbiome GBA exists [7].
2.3. Microbiota Metabolites
The gut microbiota produces a remarkably diverse array of metabolites derived from the anaerobic fermentation of dietary components and endogenous chemicals produced by both the host and microbes [33]. Many of these resulting compounds hinder the growth of their rivals, thus maintaining the diversity of commensal species and eliminating pathogenic bacteria [26]. Furthermore, these gut microbiota metabolites have roles in a variety of key physiological processes, such as host energy metabolism and immunology as well as other unknown activities, composing the human metabolome [34].
Colonic bacteria possess carbohydrate-active enzymes, enabling them to break down complex carbohydrates and produce metabolites such as SCFAs [35]. Propionate, butyrate, and acetate are the three most common SCFAs in the GI tract, with a 1:1:3 ratio [36]. These SCFAs are quickly absorbed by epithelial cells in the GI tract, where they regulate cellular processes such as gene expression, chemotaxis, differentiation, proliferation, and cell death [37].
3. Barriers to Microbiota-Gut-Brain (MGB) Signaling
The intestinal barrier and the BBB are the two primary obstacles to MGB signaling. Gut flora, inflammatory signals, and stress influence the permeability of both dynamic barriers. In a healthy condition, these tight barriers inhibit microbiome-related immunological signals reaching the brain [38,39].
3.1. Intestinal Barrier
The intestinal barrier comprises multiple layers. The outer layer is composed of mucus, commensal gut flora, and defense proteins, including secretory immunoglobulin A (sIgA) and antimicrobial proteins (AMPs). The intermediate layer consists of intestinal epithelial cells (IECs), whereas the inner layer is made up of innate and adaptive immune cells [40]. The mucus layer is divided into two layers: a loose outside layer (lumen-facing) and an interior layer linked to the epithelial tissues. Commensal microorganisms mostly occupy the outer layer, both because it is their preferred habitat for biofilm formation and because it provides microbes with nutrients (glycans) in the absence of dietary fiber [41]. Dietary fiber deficiency, which is characteristic of the traditional American diet, causes thinning of the outer layer, resulting in greater permeability of microorganisms through the outer layer [42]. The inner layer is normally devoid of bacteria and protects the epithelial cells from the microbiota through innate immune mechanisms (antimicrobial peptides), adaptive immune mechanisms (secretory immunoglobulin A), and physical separation [43]. Fortunately, diet- or chronic stress-induced shrinkage of the outer layer increases the likelihood that commensal bacteria cell wall components interact with TLRs on dendritic cell extensions, resulting in cytokine release from these cells and stimulation of other cells in the gut-associated immune system [44]. Cytokine production may also loosen the tight connections between epithelial cells, permitting microorganisms to pass across the intestinal barrier through microfold cells, which in turn allows more local immune cell activation and dissemination all through the systemic circulation, a condition known as metabolic endotoxemia [45]. Similarly, SCFAs generated by specific gut microorganisms play an important role in preserving intestinal structural integrity by keeping cell junctions tight and limiting the activation of gut-associated immune cells [46].
3.2. Blood–Brain Barrier (BBB)
The BBB controls the transport of substances from the circulatory system to the cerebrospinal fluid [47]. The BBB is composed of capillary endothelial cells with tight junctions, astrocytes, and pericytes. Tight junction proteins, which limit the paracellular transport of water-soluble compounds from the blood to the brain [48], are mostly composed of transmembrane proteins such as claudins, tricellulin, and occluding [49].
The gut microbiota, which modulates the intestinal barrier, may affect BBB permeability. The gut microbiome boosts the synthesis of tight junction proteins like Claudin-5 and occludin, which reduces BBB permeability [50]. The Colonization of the gut with SCFA-producing bacteria also reduces the permeability of the BBB, indicating that SCFAs play a crucial role in the development and maintenance of the BBB [51] - Figure 1.
4. Role of the Gut Microbiota in Health
With its extensive genetic makeup and metabolic diversity, the gut microbiota offers several beneficial characteristics to the host. Among these bacteria’s most significant functions are their assistance in preserving the mucosal barrier’s integrity, their provision of nutrients including vitamins, and their defense against infections. Furthermore, for the immune system to operate properly, commensal microbiota and the mucosal immune system must interact [52].
The alterations in gut bacterial composition and disruption of gut homeostasis, which have been linked to the etiology of gut-brain diseases, are frequently brought on by dietary patterns, antibiotic use, and viral and bacterial infections [53].
SCFAs are signaling molecules produced only by gut microorganisms during the fermentation of dietary fiber, since humans lack the enzymes necessary to digest fiber [54]. Once absorbed by colonic epithelial cells, SCFAs activate free fatty acid 2 (FFA2), FFA3, GPR109a, and Olfr78 receptors, and act on a variety of targets. SCFAs have been linked to various physiological processes, including neuroplasticity, gene expression, dietary intake, and immune system modulation [55,56].
The GI microbiota is also important for the de novo production of critical vitamins that the host can’t generate [57]. Lactic acid bacteria are essential for the creation of vitamin B12, which cannot be generated by animals, plants, or fungus [58]. Bifidobacteria are the primary providers of folate, a vitamin involved in essential host metabolic processes such as DNA synthesis and repair [59]. Vitamin K, riboflavin, biotin, nicotinic acid, pantothenic acid, pyridoxine, and thiamine are other vitamins that human gut bacteria have been demonstrated to manufacture [60]).
5. Role of GBA in Health
Bacterial colonization of the gut is essential for the formation and maturity of both the ENS and the CNS. The lack of microbial colonization leads to changes in gene expression and neurotransmitter turnover in both the CNS and ENS. Additionally, it results in altered gut sensory-motor functions, resulting in delayed gastric emptying and intestinal transit [7].
Microbiota regulates the HPA axis set point, which generates glucocorticoids such as cortisol in humans and corticosterone in rodents, which modulates stress reactivity and anxiety-like behavior [63]. Several mental disorders and behavioral changes have been linked to alterations in the HPA axis [64,65].
Cortisol can influence the MGB axis via several routes. Cortisol receptors are present on various gut cells, such as epithelial cells, immune cells, and EECs, suggesting a direct impact of cortisol on gut function [7,66,67]. By modifying gut transit time, intestinal permeability, and nutritional availability, cortisol can impact the gut microbiota, which in turn affects its diversity and composition. Central effects of cortisol occur by binding to glucocorticoid receptors in the prefrontal cortex, amygdala, and hippocampus. Additionally, there is proof that microorganisms in the stomach can trigger stress circuits in the CNS and ENS by means of the vagus nerve and sensory neurons, respectively.
Microbiota also regulate serotonergic pathways in the limbic system [7,68,69]. It has a strong influence on the EECs, which synthesize and secrete around 90% of serotonin. Furthermore, the gut microbiome has been discovered to influence hippocampal serotonin levels, possibly by affecting the peripheral availability of tryptophan [70]. Gut-derived serotonin cannot penetrate the BBB, but serotonin derivatives (N-acetyl serotonin and melatonin) and their precursor (5-hydroxytryptophan) can, and can affect the CNS [71].
Stress, anxiety, and violence are all triggered by serotonin (5-HT). Research has shown that disorders related to gut microbes and serotonergic system have a major influence on the etiology of neuropsychiatric and neurological illnesses [72]. They are linked to numerous CNS diseases, such as Alzheimer’s disease [73], Parkinson’s disease [74], and amyotrophic lateral sclerosis [75]. As a result, gut microbe disorders are thought to be a major cause of dementia.
6. Dysbiosis of the Gut Microbiome Contributes to Metabolic Dysfunction
Gut microbiota is a major environmental factor in the regulation of human metabolism, leading to the occurrence and development of chronic illnesses, such as obesity, diabetes, and atherosclerosis [76]. Reported causes include increases in systemic LPS, changes in bile acid metabolism [77], changes in SCFAs synthesis [37,78,79], changes in gut hormone secretion [80,81,82] and changes in circulating branched chain amino acids [83,84,85].
6.1. Lipopolysaccharide (LPS)
LPS, or endotoxin, is a bacterial cell wall component primarily found in Gram-negative bacteria that triggers an inflammatory response by activating TLR-4 and transforming growth factor (TGF)-mediated pathways [86,87]. Increases in systemic LPS or lipoprotein binding protein have been linked to low-grade, chronic inflammation in obesity [88], metabolic syndrome [89], and type 2 diabetes [90].
Several potential routes by which the gut microbiota may influence circulating LPS levels. These include: 1) Changes in the balance and types of intestinal bacteria can affect LPS bioavailability. For example, in diabetes, dysbiosis is characterized by a reduction in butyrate-producing, LPS-lacking, Gram-positive Clostridial species, and an increase in LPS-containing, Gram-negative opportunistic pathogens, including certain Bacteroidetes and Proteobacteria species [91,92,93]. 2) Increased intestinal permeability, often referred to as “leaky gut,” can enable LPS to move through intercellular pathways. The gut microbiome plays a key role in regulating gut permeability by supporting the health of intestinal cells, maintaining their tight junctions, and preserving a protective mucous layer. This regulation is partly achieved by supplying nutrients, such as SCFAs, to the epithelial cells [94,95,96].
Probiotics, such as Streptococcus thermophilus and Lactobacillus acidophilus, have been shown to prevent TNF-α and Interferon Gamma (IF)-induced increase in human intestinal epithelial cells’ permeability in vitro. This highlights the important role certain bacteria play in maintaining a healthy intestinal barrier [97,98].
6.2. Bile Acids
The gut microbiome plays a vital role in bile acid metabolism. Bile acids are initially synthesized from cholesterol in the liver and conjugated with glycine or taurine to form bile salts. These salts are secreted into the small intestine, where roughly 95% are reabsorbed in the ileum and transported back to the liver via enterohepatic circulation. However, around 400–600 mg of bile salts reach the large intestine, where anaerobic bacteria convert them into secondary bile acids, such as deoxycholic acid and lithocholic acid. These secondary bile acids can exert widespread effects, including on the brain [99,100].
Secondary bile acids activate the farnesoid X receptor (FXR) in the ileum [101,102], which stimulates the production of fibroblast growth factor 19 (FGF19). FGF19 then enters the bloodstream, crosses the BBB, and activates the arcuate nucleus of the hypothalamus [103]. This hypothalamic activation enhances glucose metabolism regulation and reduces HPA axis activity. The antidiabetic effects of vertical sleeve gastrectomy, a type of bariatric surgery, have been shown to rely on FXR signaling [104]. Similarly, an intestinal FXR agonist has been demonstrated to improve insulin sensitivity [105]. In the pancreas, FXR activation influences insulin transport and secretion [106] and may also provide protection to islets against lipotoxicity [107].
L cells in the ileum express Takeda G protein-coupled receptor 5 (TGR5), which is activated by secondary bile acids. These secondary bile acids are exclusively produced by intestinal bacteria and their levels are influenced by the composition of the gut microbiota [108]. Activation of TGR5 enhances glucose homeostasis by stimulating L cells to release glucagon-like peptide-1 (GLP-1). This increase in GLP-1 regulates ingestive behavior and food intake [109].
6.3. Short Chain Fatty Acid (SCFAs)
Nondigestible carbohydrates are fermented by bacteria in the colon to produce SCFAs, with butyrate, acetate, and propionate being the main products. Dietary fiber content, microbiota, and SCFAs interact because diets high in oligosaccharides change the makeup of microorganisms, produce more SCFAs, and lower the pH of the luminal fluid [55,56].
Preclinical and clinical research have demonstrated that the synthesis of SCFAs induces the ileum’s L cells to release the satiety hormone GLP-1, resulting in behavioral changes and altered satiety perceptions [110]. Additionally, SCFAs influence how 5-HT, or serotonin, is synthesized in ECCs [111]. By stimulating AMP kinase and free fatty acid receptors 2 and 3 (FFAR2 and 3), sometimes referred to as G-protein coupled receptors 43 and 41 [112], SCFAs behave as signaling molecules. SCFAs was shown to prevent de novo development of non-alcoholic fatty liver disease by stimulating fatty acid oxidation [113].
6.4. Gut Hormone Secretion
The release of gut hormones like GLP-1 and peptide YY (PYY), which regulate energy balance and glucose metabolism, is linked to SCFAs [80]. In response to dietary intake, proglucagon undergoes tissue-specific processing to produce GLP-1, which enhances insulin secretion from pancreatic β-cells. Both GLP-1 and PYY act in the hypothalamus to suppress food intake [116]. These hormones are also thought to contribute to the metabolic benefits observed after gastric bypass surgery, with GLP-1 playing a role in Metformin’s glucose-lowering effect [117,118].
PYY, like GLP-1, is synthesized by L-cells in the ileum and colon and regulates satiety by activating Agouti-related peptide (AgRP) neurons in the hypothalamus and Y2 receptors on neuropeptide Y (NPY). This suppresses appetite by disinhibiting the satiety-inducing proopiomelanocortin/alpha-melanocyte stimulating hormone (POMC/α-MSH) pathway [119]. The gut microbiota’s influence on PYY secretion is significant for understanding obesity and metabolic diseases [120]. Additionally, secondary bile acids stimulate PYY secretion via pathways like those for GLP-1 [121].
6.5. Microbial Synthesis of Amino Acids
The human microbes take part in the synthesis of amino acids and serum amino acid levels [122]. Bacteria may serve as a source of branched-chain amino acids (BCAAs), as these are more abundant in bacterial cells compared to eukaryotic ones. Notably, bacteria can synthesize all 20 amino acids required for protein production. Various lines of evidence suggest that gut microbiota influence both the composition and levels of amino acids absorbed by the host. Circulating amino acids play a role in maintaining glucose homeostasis by promoting the release of insulin and glucagon. BCAAs, in particular, appear to have a distinct role in glucose regulation, which may be linked to an increased risk of diabetes. A prospective, nested case-control study found that plasma concentrations of five branched-chain and aromatic amino acids (isoleucine, leucine, valine, tyrosine, and phenylalanine) predicted the development of diabetes, independent of traditional risk factors [123].
7. The Role of Metformin in Modulating the Gut Microbiome
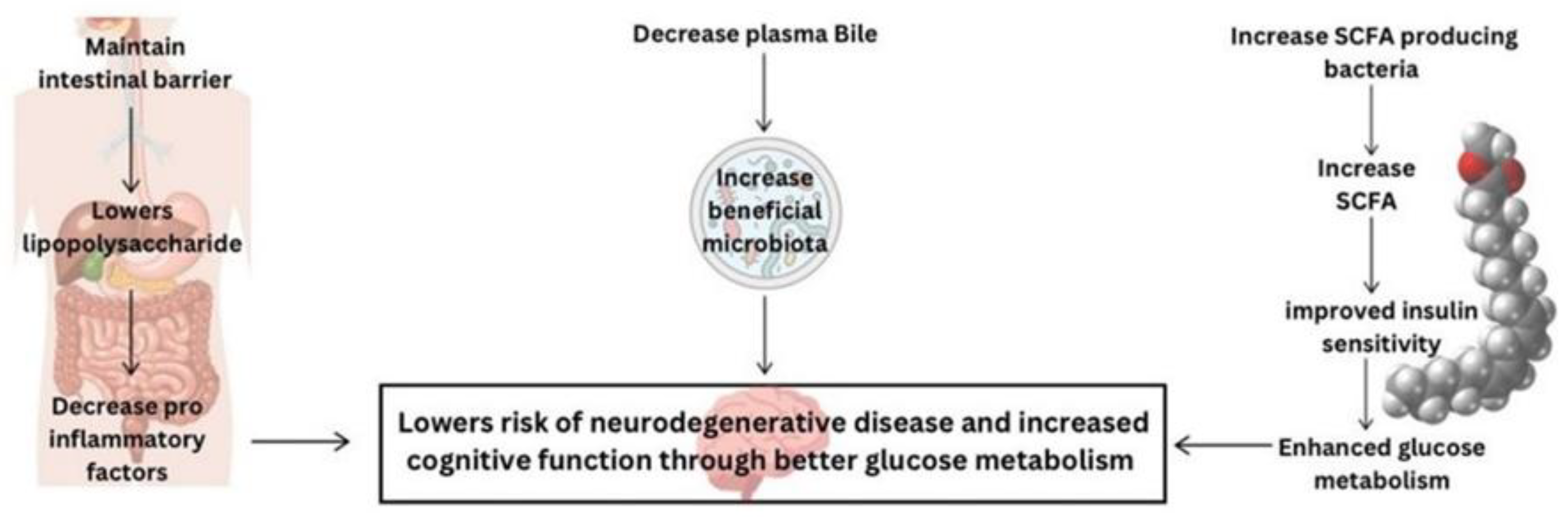
Metformin, a common medication for managing type 2 diabetes, has shown potential in altering gut microbiota, which plays a crucial role in glucose metabolism and overall metabolic health [124]. Dysbiosis, characterized by a reduced diversity of beneficial bacteria (e.g., Bifidobacterium spp. and Akkermansia muciniphila) and an increase in harmful bacteria (e.g., Firmicutes and Proteobacteria), has been linked to impaired glucose metabolism and insulin resistance [125] – Figure 2.
7.1. Metformin Mechanism of Action
Metformin primarily acts through the activation of AMP-activated kinase (AMPK), which influences various cellular processes [126]. It lowers glucose production in the liver and improves insulin sensitivity [127]. Additionally, Metformin exhibits strong anti-inflammatory and neuroprotective properties, potentially mediated by its effects on gut microbiota [128,129].
One of Metformin’s key actions is maintaining intestinal barrier integrity, which reduces serum LPS levels and enhances glucose metabolism by preventing the migration of pro-inflammatory factors. The drug also promotes SCFAs production, improving insulin sensitivity by modulating substrate metabolism in peripheral tissues. This mechanism entails an increase in the population of SCFAs-producing bacteria, reinforcing Metformin’s role in regulating glucose levels. Additionally, Metformin regulates bile acid levels, contributing to improved glucose metabolism. Studies have shown that it elevates plasma bile acid while altering gut microbiota to enhance metabolic outcomes. Its effect on gut microbiota composition, particularly through the promotion of beneficial bacteria such as Akkermansia muciniphila, plays a significant role in glucose homeostasis and overall metabolic health [4,130]. Metformin may influence glucose transfer from the intestinal lumen to the bloodstream and enhance glucose sensing in the gut. This highlights an additional mechanism by which Metformin contributes to its glucose-lowering effects.
Metformin’s modulation of gut microbiota may have implications for brain health, potentially addressing cognitive disorders [128]. Through the GBA, Metformin’s influence on gut microbiota opens possibilities for microbiome-targeted therapies that could offer cognitive benefits. These findings underline the importance of exploring Metformin’s broader effects beyond its traditional role in metabolic regulation.
7.2. Effects of Metformin on Cognition
Metformin has been associated with considerably reduced risk of dementia and neurodegenerative disorders, as well as enhancements in three cognitive domains: memory, semantic memory, and executive function [131,132]. Such neuroprotective benefits may occur via improving neuronal AMPK-induced energy homeostasis.
Metformin treatment was also shown to effectively counteract amyloid-beta-induced effects on human neural stem cells (hNSCs) by suppressing caspase activity and reducing cytosolic cytochrome c. Furthermore, co-treatment with Metformin played a key role in restoring mitochondrial structure in affected stem cells, bringing it closer to normal morphology [2]. Metformin-induced AMPK activation protected the stem cells against cytotoxicity caused by advanced glycation end products [133].
Metformin has been shown to reduce Alzheimer’s disease-associated neuropathological alterations in differentiated mouse neuroblastoma cell lines, such as Neuro-2a [134]. It also inhibited the phosphorylation of tau protein in cultured neurons and in mice brains. Metformin was additionally shown to prevent apoptotic cell death in primary cortical neurons [135], and restore the decreased cell proliferation and neuroblast differentiation caused by type 2 diabetes in the dentate gyrus of the rat hippocampus [136].
In humans, Metformin was associated with a reduction in the incidence rate of dementia compared to those treated with sulfonylureas and thiazolidinediones. Combination therapy with Metformin and thiazolidinedione lowered the risk of all-cause dementia. Combination therapy with Metformin and sulfonylureas protected against all types of dementia over 2-year time [137].
Given that mitochondria-related metabolic disturbances and impaired molecular pathways play a role in the pathogenesis of multiple neurological degenerative diseases and manifest differently at different stages of disease progression, it is fundamental to precisely measure the altered mitochondria-related molecular pathways, which have great potential to serve as biomarkers to guide metformin-effective treatment for targeted therapy at the specific stage of disease [138].
8. Conclusions
The importance of microbiota in health cannot be overstated. Emerging research has highlighted the crucial role that the diverse community of microorganisms residing in our bodies plays in various physiological processes. These microbial populations play a crucial role in digestion, immune function, and the synthesis of essential nutrients.
Furthermore, microbiota influences the GBA, affecting mental health and cognitive function. Disruptions in microbial balance have been linked to a range of health issues, including gastrointestinal disorders, metabolic conditions, and even neurodegenerative diseases.
Metformin impacts gut microbiota composition, improving glucose regulation and potentially enhancing brain health. Since mitochondrial dysfunction contributes to various neurodegenerative diseases, precise measurements of altered mitochondrial pathways may help identify biomarkers to guide stage-specific metformin therapy. However, more research is needed to fully understand how Metformin influences gut microbiota and its role in managing type 2 diabetes and cognitive function. Future studies should investigate the microbial changes associated with therapeutic benefits, paving the way for new treatments for metabolic and cognitive disorders.
Author Contributions
“Conceptualization, A.N.S. and V.F.F.; writing—original draft preparation, A.N.S., V.F.F., F.A.I., and H.A.E; writing—review and editing, H.S., and H.S.S..
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest
Abbreviations
The following abbreviations are used in this manuscript:
AgRP
Agouti-related peptide
AMPs
Antimicrobial proteins
ANS
Autonomic nervous system
BBB
Blood-brain barrier
CNS
Central Nervous System
ECCs
Enterochromaffin cells
EECs
Enteroendocrine cells
ENS
Enteric nervous system
FFA2
Free fatty acid 2
FGF19
Fibroblast growth factor 19
FXR
Farnesoid X receptor
GABA
Gamma-aminobutyric acid
GALT
Gut-associated lymphoid tissues
GBA
Gut-Brain Axis
GI
Gastrointestinal
GLP-1
Glucagon-like peptide-1
hNSCs
Human neural stem cells
HPA
Hypothalamic-pituitary-adrenal
IECs
Intestinal epithelial cells
IFᵞ
Interferon Gamma
IL-10
Interleukin 10
LPS
Lipopolysaccharide
NPY
Neuropeptide Y
PG
Peptidoglycan
POMC/α-MSH
proopiomelanocortin/alpha-melanocyte-stimulating
hormone
PYY
Peptide YY
SCFAs
Short-chain fatty acids
sIgA
Secretory immunoglobulin A
TGF
Transforming growth factor
TGR5
Takeda G protein-coupled receptor 5
TLRs
Toll-like receptors
References
- de Vos WM, Tilg H, Van Hul M, Cani PD: Gut microbiome and health: mechanistic insights. Gut 2022, 71, 1020–1032. [CrossRef]
- Chiang MC, Cheng YC, Chen SJ, Yen CH, Huang RN: Metformin activation of AMPK-dependent pathways is neuroprotective in human neural stem cells against Amyloid-beta-induced mitochondrial dysfunction. Exp. Cell Res. 2016, 347, 322–331. [CrossRef]
- Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, Fernandes GR, Tap J, Bruls T, Batto JM et al: Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [CrossRef] [PubMed]
- Wang X, Huang H, Zhu Y, Li S, Zhang P, Jiang J, Xi C, Wu L, Gao X, Fu Y et al: Metformin acts on the gut-brain axis to ameliorate antipsychotic-induced metabolic dysfunction. Biosci. Trends 2021, 15, 321–329. [CrossRef] [PubMed]
- Khosravi F, Amiri Z, Masouleh NA, Kashfi P, Panjizadeh F, Hajilo Z, Shanayii S, Khodakarim S, Rahnama L: Shoulder pain prevalence and risk factors in middle-aged women: A cross-sectional study. Journal of bodywork and movement therapies 2019, 23, 752–757. [CrossRef] [PubMed]
- Muller PA, Schneeberger M, Matheis F, Wang P, Kerner Z, Ilanges A, Pellegrino K, Del Marmol J, Castro TBR, Furuichi M et al: Microbiota modulate sympathetic neurons via a gut-brain circuit. Nature 2020, 583, 441–446.
- Carabotti M, Scirocco, A., Maselli, M.A. and Severi, C.: The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Annals of Gastroenterology 2015, 28(2):203.
- Mitrea L, Nemes SA, Szabo K, Teleky BE, Vodnar DC: Guts Imbalance Imbalances the Brain: A Review of Gut Microbiota Association With Neurological and Psychiatric Disorders. Front Med (Lausanne), 2022, 9:813204.
- Dong TS, Mayer E: Advances in Brain-Gut-Microbiome Interactions: A Comprehensive Update on Signaling Mechanisms, Disorders, and Therapeutic Implications. Cell Mol. Gastroenterol. Hepatol. 2024, 18, 1–13. [CrossRef]
- Tsigos C, Chrousos GP: Hypothalamic-pituitary-adrenal axis, neuroendocrine factors and stress. J. Psychosom. Res. 2002, 53, 865–871. [CrossRef]
- Foster JA MNK: Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [CrossRef]
- Mayer EA, Savidge T, Shulman RJ: Brain-gut microbiome interactions and functional bowel disorders. Gastroenterology 2014, 146, 1500–1512. [CrossRef]
- Martin CR, Osadchiy V, Kalani A, Mayer EA: The Brain-Gut-Microbiome Axis. Cell Mol. Gastroenterol. Hepatol. 2018, 6, 133–148. [CrossRef] [PubMed]
- Holzer P, Farzi A: Neuropeptides and the microbiota-gut-brain axis. Advances in experimental medicine and biology 2014, 817:195-219.
- Lyte M: Microbial endocrinology in the microbiome-gut-brain axis: how bacterial production and utilization of neurochemicals influence behavior. PLoS Pathog. 2013, 9, e1003726.
- Obrenovich M, Sankar Chittoor Mana, T., Rai, H., Shola, D., Sass, C., McCloskey, B. and Levison, B.S.: Recent findings within the microbiota–gut–brain–endocrine metabolic interactome. Pathology and Laboratory Medicine International.
- Ignatow G: The microbiome-gut-brain and social behavior. Journal for the Theory of Social. Behaviour 2022, 52, 164–182. [CrossRef]
- Sorboni SG, Moghaddam HS, Jafarzadeh-Esfehani R, Soleimanpour S: A Comprehensive Review on the Role of the Gut Microbiome in Human Neurological Disorders. Clin. Microbiol. Rev. 2022, 35, e0033820.
- Cryan JF, O’Riordan KJ, Sandhu K, Peterson V, Dinan TG: The gut microbiome in neurological disorders. Lancet Neurol. 2020, 19, 179–194. [CrossRef]
- Chen Y, Xu J, Chen Y: Regulation of Neurotransmitters by the Gut Microbiota and Effects on Cognition in Neurological Disorders. Nutrients 2021, 13.
- Boyd CA: Amine uptake and peptide hormone secretion: APUD cells in a new landscape. J Physiol, 2001, 531(Pt 3):581.
- Worthington JJ, Reimann F, Gribble FM: Enteroendocrine cells-sensory sentinels of the intestinal environment and orchestrators of mucosal immunity. Mucosal Immunol. 2018, 11, 3–20. [CrossRef]
- Levy M, Thaiss CA, Elinav E: Metabolites: messengers between the microbiota and the immune system. Genes. Dev. 2016, 30, 1589–1597. [CrossRef]
- Yu Y, Yang W, Li Y, Cong Y: Enteroendocrine Cells: Sensing Gut Microbiota and Regulating Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2020, 26, 11–20. [CrossRef]
- Morbe UM, Jorgensen PB, Fenton TM, von Burg N, Riis LB, Spencer J, Agace WW: Human gut-associated lymphoid tissues (GALT); diversity, structure, and function. Mucosal Immunol. 2021, 14, 793–802. [CrossRef]
- Man AWC, Zhou Y, Xia N, Li H: Involvement of Gut Microbiota, Microbial Metabolites and Interaction with Polyphenol in Host Immunometabolism. Nutrients 2020, 12.
- Duan T, Du Y, Xing C, Wang HY, Wang RF: Toll-Like Receptor Signaling and Its Role in Cell-Mediated Immunity. Front Immunol, 2022, 13:812774.
- Yiu JH, Dorweiler B, Woo CW: Interaction between gut microbiota and toll-like receptor: from immunity to metabolism. J Mol Med (Berl) 2017, 95, 13–20. [CrossRef]
- Carlson NG WW, Chen J, Bacchi A, Rogers SW, Gahring LC: Inflammatory cytokines IL-1α, IL-1β, IL-6, and TNF-α impart neuroprotection to an excitotoxin through distinct pathways. J. Immunol. 1999, 163, 3963–3968. [CrossRef]
- Schwarz MJ: Cytokines, neurophysiology, neuropsychology, and psychiatric symptoms. Dialogues Clin. Neurosci. 2003, 5, 139–153. [CrossRef] [PubMed]
- Walker AW, Ince J, Duncan SH, Webster LM, Holtrop G, Ze X, Brown D, Stares MD, Scott P, Bergerat A et al: Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [CrossRef] [PubMed]
- Vrieze A, Van Nood E, Holleman F, Salojarvi J, Kootte RS, Bartelsman JF, Dallinga-Thie GM, Ackermans MT, Serlie MJ, Oozeer R et al: Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology.
- Rooks MG, Garrett WS: Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [CrossRef] [PubMed]
- Liu J, Tan Y, Cheng H, Zhang D, Feng W, Peng C: Functions of Gut Microbiota Metabolites, Current Status and Future Perspectives. Aging Dis. 2022, 13, 1106–1126. [CrossRef]
- Thursby E, Juge N: Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [CrossRef]
- Silva YP, Bernardi A, Frozza RL: The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol (Lausanne) 2020, 11:25.
- Kim CH: Complex regulatory effects of gut microbial short-chain fatty acids on immune tolerance and autoimmunity. Cell Mol. Immunol. 2023, 20, 341–350. [CrossRef]
- Daneman R, Rescigno M: The gut immune barrier and the blood-brain barrier: are they so different? Immunity 2009, 31, 722–735. [CrossRef]
- Scalise AA, Kakogiannos N, Zanardi F, Iannelli F, Giannotta M: The blood-brain and gut-vascular barriers: from the perspective of claudins. Tissue Barriers 2021, 9, 1926190. [CrossRef]
- Vancamelbeke M, Vermeire S: The intestinal barrier: a fundamental role in health and disease. Expert. Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [CrossRef]
- Johansson ME, Phillipson M, Petersson J, Velcich A, Holm L, Hansson GC: The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. U S A 2008, 105, 15064–15069. [CrossRef]
- Pelaseyed T, Bergstrom JH, Gustafsson JK, Ermund A, Birchenough GM, Schutte A, van der Post S, Svensson F, Rodriguez-Pineiro AM, Nystrom EE et al: The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 2014, 260, 8–20.
- Herath M, Hosie S, Bornstein JC, Franks AE, Hill-Yardin EL: The Role of the Gastrointestinal Mucus System in Intestinal Homeostasis: Implications for Neurological Disorders. Front Cell Infect Microbiol 2020, 10:248.
- Lian P, Braber S, Garssen J, Wichers HJ, Folkerts G, Fink-Gremmels J, Varasteh S: Beyond Heat Stress: Intestinal Integrity Disruption and Mechanism-Based Intervention Strategies. Nutrients 2020, 12.
- Tsiaoussis GI, Assimakopoulos SF, Tsamandas AC, Triantos CK, Thomopoulos KC: Intestinal barrier dysfunction in cirrhosis: Current concepts in pathophysiology and clinical implications. World J. Hepatol. 2015, 7, 2058–2068. [CrossRef] [PubMed]
- Shin Y, Han S, Kwon J, Ju S, Choi TG, Kang I, Kim SS: Roles of Short-Chain Fatty Acids in Inflammatory Bowel Disease. Nutrients 2023, 15.
- Schreiner TG, Romanescu C, Popescu BO: The Blood-Brain Barrier-A Key Player in Multiple Sclerosis Disease Mechanisms. Biomolecules 2022, 12.
- Hawkins BT, Davis TP: The blood-brain barrier/neurovascular unit in health and disease. Pharmacol. Rev. 2005, 57, 173–185. [CrossRef]
- Tscheik C, Blasig IE, Winkler L: Trends in drug delivery through tissue barriers containing tight junctions. Tissue Barriers 2013, 1, e24565. [CrossRef]
- Fock E, Parnova R: Mechanisms of Blood-Brain Barrier Protection by Microbiota-Derived Short-Chain Fatty Acids. Cells 2023, 12.
- Fusco W, Lorenzo MB, Cintoni M, Porcari S, Rinninella E, Kaitsas F, Lener E, Mele MC, Gasbarrini A, Collado MC et al: Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15.
- Bested AC, Logan AC, Selhub EM: Intestinal microbiota, probiotics and mental health: from Metchnikoff to modern advances: Part II - contemporary contextual research. Gut Pathog. 2013, 5, 3. [CrossRef]
- Rogers GB, Keating DJ, Young RL, Wong ML, Licinio J, Wesselingh S: From gut dysbiosis to altered brain function and mental illness: mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [CrossRef]
- Roy CC, Kien CL, Bouthillier L, Levy E: Short-chain fatty acids: ready for prime time? Clin. Pract. 2006, 21, 351–366.
- den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud DJ, Bakker BM: The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [CrossRef] [PubMed]
- Nogal A, Valdes, A.M. and Menni, C.: The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut microbes 2021, 13(1):1-24.
- LeBlanc JG MC, de Giori GS, Sesma F, van Sinderen D, Ventura M.: Bacteria as vitamin suppliers to their host: a gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24(2):160-168.
- Watanabe F, Bito T: Vitamin B(12) sources and microbial interaction. Exp Biol Med (Maywood) 2018, 243, 148–158. [CrossRef] [PubMed]
- Pompei A, Cordisco L, Amaretti A, Zanoni S, Matteuzzi D, Rossi M: Folate production by bifidobacteria as a potential probiotic property. Appl. Environ. Microbiol. 2007, 73, 179–185. [CrossRef]
- Hill MJ: Intestinal flora and endogenous vitamin synthesis. Eur, S: Prev 1997, 6 Suppl 1, 1997.
- Ivanov, II, Honda K: Intestinal commensal microbes as immune modulators. Cell Host Microbe 2012, 12, 496–508. [CrossRef]
- Zheng A, Fowler JR: The Effectiveness of Preoperative Antibiotic Prophylaxis in Ulnar Nerve Release at the Cubital Tunnel. Hand (N Y) 2024, 19(2):224-227.
- Huo R, Zeng B, Zeng L, Cheng K, Li B, Luo Y, Wang H, Zhou C, Fang L, Li W et al: Microbiota Modulate Anxiety-Like Behavior and Endocrine Abnormalities in Hypothalamic-Pituitary-Adrenal Axis. Front Cell Infect Microbiol 2017, 7:489.
- Schatzberg AF, Keller J, Tennakoon L, Lembke A, Williams G, Kraemer FB, Sarginson JE, Lazzeroni LC, Murphy GM: HPA axis genetic variation, cortisol and psychosis in major depression. Mol. Psychiatry 2014, 19, 220–227. [CrossRef]
- Fries GR, Vasconcelos-Moreno MP, Gubert C, dos Santos BT, Sartori J, Eisele B, Ferrari P, Fijtman A, Ruegg J, Gassen NC et al: Hypothalamic-pituitary-adrenal axis dysfunction and illness progression in bipolar disorder. Int J Neuropsychopharmacol 2014, 18.
- Tetel MJ, de Vries GJ, Melcangi RC, Panzica G, O’Mahony SM: Steroids, stress and the gut microbiome-brain axis. J Neuroendocrinol 2018, 30.
- Mayer EA: The neurobiology of stress and gastrointestinal disease. Gut 2000, 47, 861–869. [CrossRef]
- Lyte M, Li W, Opitz N, Gaykema RP, Goehler LE: Induction of anxiety-like behavior in mice during the initial stages of infection with the agent of murine colonic hyperplasia Citrobacter rodentium. Physiol. Behav. 2006, 89, 350–357. [CrossRef] [PubMed]
- Geng ZH, Zhu Y, Li QL, Zhao C, Zhou PH: Enteric Nervous System: The Bridge Between the Gut Microbiota and Neurological Disorders. Front Aging Neurosci, 8104.
- Appleton J: The Gut-Brain Axis: Influence of Microbiota on Mood and Mental Health. Integr Med (Encinitas) 2018, 17, 28–32.
- Maffei ME: 5-Hydroxytryptophan (5-HTP): Natural Occurrence, Analysis, Biosynthesis, Biotechnology, Physiology and Toxicology. Int J Mol Sci 2020, 22.
- Dorszewska J F-WJ, Kowalska M, Stanski M, Kowalewska A, Kozubski W: Serotonin in Neurological Diseases. In Serotonin - A Chemical Messenger Between All Types of Living Cells. 2017.
- Wu SC, Cao, Z.S., Chang, K.M. and Juang, J.L.: Intestinal microbial dysbiosis aggravates the progression of Alzheimer’s disease in Drosophila. Nature communications 2017, 8(1):24.
- Scheperjans F, Aho V, Pereira PA, Koskinen K, Paulin L, Pekkonen E, Haapaniemi E, Kaakkola S, Eerola-Rautio J, Pohja M et al: Gut microbiota are related to Parkinson’s disease and clinical phenotype. Mov. Disord. 2015, 30, 350–358.
- Zhai CD, Zheng JJ, An BC, Huang HF, Tan ZC: Intestinal microbiota composition in patients with amyotrophic lateral sclerosis: establishment of bacterial and archaeal communities analyses. Chin Med J (Engl) 2019, 132, 1815–1822. [CrossRef]
- Brial F, Le Lay A, Dumas ME, Gauguier D: Implication of gut microbiota metabolites in cardiovascular and metabolic diseases. Cell Mol. Life Sci. 2018, 75, 3977–3990. [CrossRef]
- Ethanic M, Stanimirov B, Pavlovic N, Golocorbin-Kon S, Al-Salami H, Stankov K, Mikov M: Pharmacological Applications of Bile Acids and Their Derivatives in the Treatment of Metabolic Syndrome. Front Pharmacol, 1382.
- Jia X, Xu W, Zhang L, Li X, Wang R, Wu S: Impact of Gut Microbiota and Microbiota-Related Metabolites on Hyperlipidemia. Front Cell Infect Microbiol, 2021, 11:634780.
- Facchin S, Bertin L, Bonazzi E, Lorenzon G, De Barba C, Barberio B, Zingone F, Maniero D, Scarpa M, Ruffolo C et al: Short-Chain Fatty Acids and Human Health: From Metabolic Pathways to Current Therapeutic Implications. Life (Basel) 2024, 14.
- Utzschneider KM, Kratz M, Damman CJ, Hullar M: Mechanisms Linking the Gut Microbiome and Glucose Metabolism. J. Clin. Endocrinol. Metab. 2016, 101, 1445–1454. [CrossRef]
- Fukui H, Xu X, Miwa H: Role of Gut Microbiota-Gut Hormone Axis in the Pathophysiology of Functional Gastrointestinal Disorders. J. Neurogastroenterol. Motil. 2018, 24, 367–386. [CrossRef] [PubMed]
- Leeuwendaal NK, Cryan JF, Schellekens H: Gut peptides and the microbiome: focus on ghrelin. Curr. Opin. Endocrinol. Diabetes Obes. 2021, 28, 243–252. [CrossRef] [PubMed]
- Tajiri K, Shimizu Y: Branched-chain amino acids in liver diseases. Transl, 4: Hepatol 2018, 3, 2018.
- Zhou M, Shao J, Wu CY, Shu L, Dong W, Liu Y, Chen M, Wynn RM, Wang J, Wang J et al: Targeting BCAA Catabolism to Treat Obesity-Associated Insulin Resistance. Diabetes 2019, 68, 1730–1746. [CrossRef] [PubMed]
- Agus A, Clement K, Sokol H: Gut microbiota-derived metabolites as central regulators in metabolic disorders. Gut 2021, 70, 1174–1182. [CrossRef]
- Meseguer V, Alpizar YA, Luis E, Tajada S, Denlinger B, Fajardo O, Manenschijn JA, Fernandez-Pena C, Talavera A, Kichko T et al: TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nat Commun 2014, 5:3125.
- Mazgaeen L, Gurung P: Recent Advances in Lipopolysaccharide Recognition Systems. Int J Mol Sci 2020, 21.
- Pavlov VA: The evolving obesity challenge: targeting the vagus nerve and the inflammatory reflex in the response. Pharmacol, 1: 222, 1077.
- Fedulovs A, Pahirko L, Jekabsons K, Kunrade L, Valeinis J, Riekstina U, Pirags V, Sokolovska J: Association of Endotoxemia with Low-Grade Inflammation, Metabolic Syndrome and Distinct Response to Lipopolysaccharide in Type 1 Diabetes. Biomedicines 2023, 11.
- Metz CN, Brines M, Xue X, Chatterjee PK, Adelson RP, Roth J, Tracey KJ, Gregersen PK, Pavlov VA: Increased plasma lipopolysaccharide-binding protein and altered inflammatory mediators reveal a pro-inflammatory state in overweight women. BMC Womens Health 2025, 25, 57.
- Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F, Liang S, Zhang W, Guan Y, Shen D et al: A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [CrossRef]
- Salguero MV, Al-Obaide MAI, Singh R, Siepmann T, Vasylyeva TL: Dysbiosis of Gram-negative gut microbiota and the associated serum lipopolysaccharide exacerbates inflammation in type 2 diabetic patients with chronic kidney disease. Exp. Ther. Med. 2019, 18, 3461–3469.
- Meng L, Song Z, Liu A, Dahmen U, Yang X, Fang H: Effects of Lipopolysaccharide-Binding Protein (LBP) Single Nucleotide Polymorphism (SNP) in Infections, Inflammatory Diseases, Metabolic Disorders and Cancers. Front Immunol, 6818.
- Di Tommaso N, Gasbarrini A, Ponziani FR: Intestinal Barrier in Human Health and Disease. Int J Environ Res Public Health 2021, 18.
- Di Vincenzo F, Del Gaudio A, Petito V, Lopetuso LR, Scaldaferri F: Gut microbiota, intestinal permeability, and systemic inflammation: a narrative review. Intern. Emerg. Med. 2024, 19, 275–293. [CrossRef]
- Poto R, Fusco W, Rinninella E, Cintoni M, Kaitsas F, Raoul P, Caruso C, Mele MC, Varricchi G, Gasbarrini A et al: The Role of Gut Microbiota and Leaky Gut in the Pathogenesis of Food Allergy. Nutrients 2023, 16.
- Resta-Lenert S, Barrett KE: Probiotics and commensals reverse TNF-alpha- and IFN-gamma-induced dysfunction in human intestinal epithelial cells. Gastroenterology 2006, 130, 731–746. [CrossRef] [PubMed]
- Virk MS, Virk MA, He Y, Tufail T, Gul M, Qayum A, Rehman A, Rashid A, Ekumah JN, Han X et al: The Anti-Inflammatory and Curative Exponent of Probiotics: A Comprehensive and Authentic Ingredient for the Sustained Functioning of Major Human Organs. Nutrients 2024, 16.
- Ridlon JM, Harris SC, Bhowmik S, Kang DJ, Hylemon PB: Consequences of bile salt biotransformations by intestinal bacteria. Gut Microbes 2016, 7, 22–39. [CrossRef] [PubMed]
- Urdaneta V, Casadesus J: Interactions between Bacteria and Bile Salts in the Gastrointestinal and Hepatobiliary Tracts. Front Med (Lausanne) 2017, 4:163.
- Stofan M, Guo GL: Bile Acids and FXR: Novel Targets for Liver Diseases. Front Med (Lausanne) 2020, 7:544.
- Chiang JYL, Ferrell JM: Discovery of farnesoid X receptor and its role in bile acid metabolism. Mol Cell Endocrinol, 2022, 548:111618.
- Wean JB, Smith BN: Fibroblast Growth Factor 19 Increases the Excitability of Pre-Motor Glutamatergic Dorsal Vagal Complex Neurons From Hyperglycemic Mice. Front Endocrinol (Lausanne), 2021, 12, 765359.
- Li K, Zou J, Li S, Guo J, Shi W, Wang B, Han X, Zhang H, Zhang P, Miao Z et al: Farnesoid X receptor contributes to body weight-independent improvements in glycemic control after Roux-en-Y gastric bypass surgery in diet-induced obese mice. Molecular metabolism 2020, 37:100980.
- Boland BB, Mumphrey MB, Hao Z, Townsend RL, Gill B, Oldham S, Will S, Morrison CD, Yu S, Munzberg H et al: Combined loss of GLP-1R and Y2R does not alter progression of high-fat diet-induced obesity or response to RYGB surgery in mice. Molecular metabolism 2019, 25:64-72.
- Fleishman JS, Kumar S: Bile acid metabolism and signaling in health and disease: molecular mechanisms and therapeutic targets. Signal Transduct. Target. Ther. 2024, 9, 97. [CrossRef]
- Ruze R, Song J, Yin X, Chen Y, Xu R, Wang C, Zhao Y: Mechanisms of obesity- and diabetes mellitus-related pancreatic carcinogenesis: a comprehensive and systematic review. Signal Transduct. Target. Ther. 2023, 8, 139. [CrossRef]
- Masse KE, Lu VB: Short-chain fatty acids, secondary bile acids and indoles: gut microbial metabolites with effects on enteroendocrine cell function and their potential as therapies for metabolic disease. Front Endocrinol (Lausanne), 1169.
- Lun W, Yan Q, Guo X, Zhou M, Bai Y, He J, Cao H, Che Q, Guo J, Su Z: Mechanism of action of the bile acid receptor TGR5 in obesity. Acta Pharm. Sin. B 2024, 14, 468–491. [CrossRef]
- Tolhurst G, Heffron H, Lam YS, Parker HE, Habib AM, Diakogiannaki E, Cameron J, Grosse J, Reimann F, Gribble FM: Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [CrossRef]
- Margolis KG, Cryan JF, Mayer EA: The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [CrossRef]
- May KS, den Hartigh LJ: Modulation of Adipocyte Metabolism by Microbial Short-Chain Fatty Acids. Nutrients 2021, 13.
- Mishra SP, Karunakar P, Taraphder S, Yadav H: Free Fatty Acid Receptors 2 and 3 as Microbial Metabolite Sensors to Shape Host Health: Pharmacophysiological View. Biomedicines 2020, 8.
- Arora T, Tremaroli V: Therapeutic Potential of Butyrate for Treatment of Type 2 Diabetes. Front Endocrinol (Lausanne), 2021, 12:761834.
- Mayorga-Ramos A, Barba-Ostria C, Simancas-Racines D, Guaman LP: Protective role of butyrate in obesity and diabetes: New insights. Front Nutr, 2022, 9:1067647.
- Abou-Samra M, Venema K, Ayoub Moubareck C, Karavetian M: The Association of Peptide Hormones with Glycemia, Dyslipidemia, and Obesity in Lebanese Individuals. Metabolites 2022, 12.
- Bahne E, Sun EWL, Young RL, Hansen M, Sonne DP, Hansen JS, Rohde U, Liou AP, Jackson ML, de Fontgalland D et al: Metformin-induced glucagon-like peptide-1 secretion contributes to the actions of Metformin in type 2 diabetes. JCI Insight 2018, 3.
- Hansen LS, Gasbjerg LS, Bronden A, Dalsgaard NB, Bahne E, Stensen S, Hellmann PH, Rehfeld JF, Hartmann B, Wewer Albrechtsen NJ et al: The role of glucagon-like peptide 1 in the postprandial effects of Metformin in type 2 diabetes: a randomized crossover trial. Eur. J. Endocrinol. 2024, 191, 192–203.
- Loh K, Herzog H, Shi YC: Regulation of energy homeostasis by the NPY system. Trends Endocrinol. Metab. 2015, 26, 125–135. [CrossRef] [PubMed]
- Larraufie P, Martin-Gallausiaux C, Lapaque N, Dore J, Gribble FM, Reimann F, Blottiere HM: SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 74.
- Kuhre RE, Wewer Albrechtsen NJ, Larsen O, Jepsen SL, Balk-Moller E, Andersen DB, Deacon CF, Schoonjans K, Reimann F, Gribble FM et al: Bile acids are important direct and indirect regulators of the secretion of appetite- and metabolism-regulating hormones from the gut and pancreas. Molecular metabolism 2018, 11:84-95.
- Metges CC: Contribution of microbial amino acids to amino acid homeostasis of the host. J Nutr 2000, 130, 1857S–1864S. [CrossRef]
- Wang TJ, Larson MG, Vasan RS, Cheng S, Rhee EP, McCabe E, Lewis GD, Fox CS, Jacques PF, Fernandez C et al: Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [CrossRef]
- Yang G, Wei J, Liu P, Zhang Q, Tian Y, Hou G, Meng L, Xin Y, Jiang X: Role of the gut microbiota in type 2 diabetes and related diseases. Metabolism, 2021, 117:154712.
- Crudele L, Gadaleta RM, Cariello M, Moschetta A: Gut microbiota in the pathogenesis and therapeutic approaches of diabetes. EBioMedicine, 2023, 97:104821.
- Zhou G, Myers R, Li Y, Chen Y, Shen X, Fenyk-Melody J, Wu M, Ventre J, Doebber T, Fujii N et al: Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Invest. 2001, 108, 1167–1174. [CrossRef]
- Giannarelli R, Aragona M, Coppelli A, Del Prato S: Reducing insulin resistance with Metformin: the evidence today. Diabetes Metab, 2003, 29(4 Pt 2):6S28-35.
- Cao G, Gong T, Du Y, Wang Y, Ge T, Liu J: Mechanism of metformin regulation in central nervous system: Progression and future perspectives. Biomed Pharmacother, 2022, 156:113686.
- Lin H, Ao H, Guo G, Liu M: The Role and Mechanism of Metformin in Inflammatory Diseases. J Inflamm Res, 2023, 16:5545-5564.
- Zhang Q, Hu N: Effects of Metformin on the Gut Microbiota in Obesity and Type 2 Diabetes Mellitus. Diabetes Metab Syndr Obes, 2020, 13:5003-5014.
- He R, Zheng R, Li J, Cao Q, Hou T, Zhao Z, Xu M, Chen Y, Lu J, Wang T et al: Individual and Combined Associations of Glucose Metabolic Components With Cognitive Function Modified by Obesity. Front Endocrinol (Lausanne) 2021, 12:769120.
- Rosell-Diaz M, Fernandez-Real JM: Metformin, Cognitive Function, and Changes in the Gut Microbiome. Endocr. Rev. 2024, 45, 210–226. [CrossRef] [PubMed]
- Chung MM, Chen YL, Pei D, Cheng YC, Sun B, Nicol CJ, Yen CH, Chen HM, Liang YJ, Chiang MC: The neuroprotective role of Metformin in advanced glycation end product treated human neural stem cells is AMPK-dependent. Biochim. Biophys. Acta 2015, 1852, 720–731. [CrossRef] [PubMed]
- Gupta A, Bisht B, Dey CS: Peripheral insulin-sensitizer drug metformin ameliorates neuronal insulin resistance and Alzheimer’s-like changes. Neuropharmacology 2011, 60, 910–920. [CrossRef] [PubMed]
- El-Mir MY, Detaille D, G RV, Delgado-Esteban M, Guigas B, Attia S, Fontaine E, Almeida A, Leverve X: Neuroprotective role of antidiabetic drug metformin against apoptotic cell death in primary cortical neurons. J. Mol. Neurosci. 2008, 34, 77–87. [CrossRef]
- Hwang IK, Kim IY, Joo EJ, Shin JH, Choi JW, Won MH, Yoon YS, Seong JK: Metformin normalizes type 2 diabetes-induced decrease in cell proliferation and neuroblast differentiation in the rat dentate gyrus. Neurochem. Res. 2010, 35, 645–650. [CrossRef]
- Tang X, Brinton RD, Chen Z, Farland LV, Klimentidis Y, Migrino R, Reaven P, Rodgers K, Zhou JJ: Use of oral diabetes medications and the risk of incident dementia in US veterans aged >/=60 years with type 2 diabetes. BMJ Open Diabetes Res Care 2022, 10.
- Loan A, Syal C, Lui M, He L, Wang J: Promising use of Metformin in treating neurological disorders: biomarker-guided therapies. Neural Regen. Res. 2024, 19, 1045–1055. [CrossRef]
Figure 1.
This diagram shows the effect of the gut microbiome on the blood-brain barrier. BBB: Blood-Brain Barrier - SCFA: Short-Chain Fatty Acid.
Figure 1.
This diagram shows the effect of the gut microbiome on the blood-brain barrier. BBB: Blood-Brain Barrier - SCFA: Short-Chain Fatty Acid.
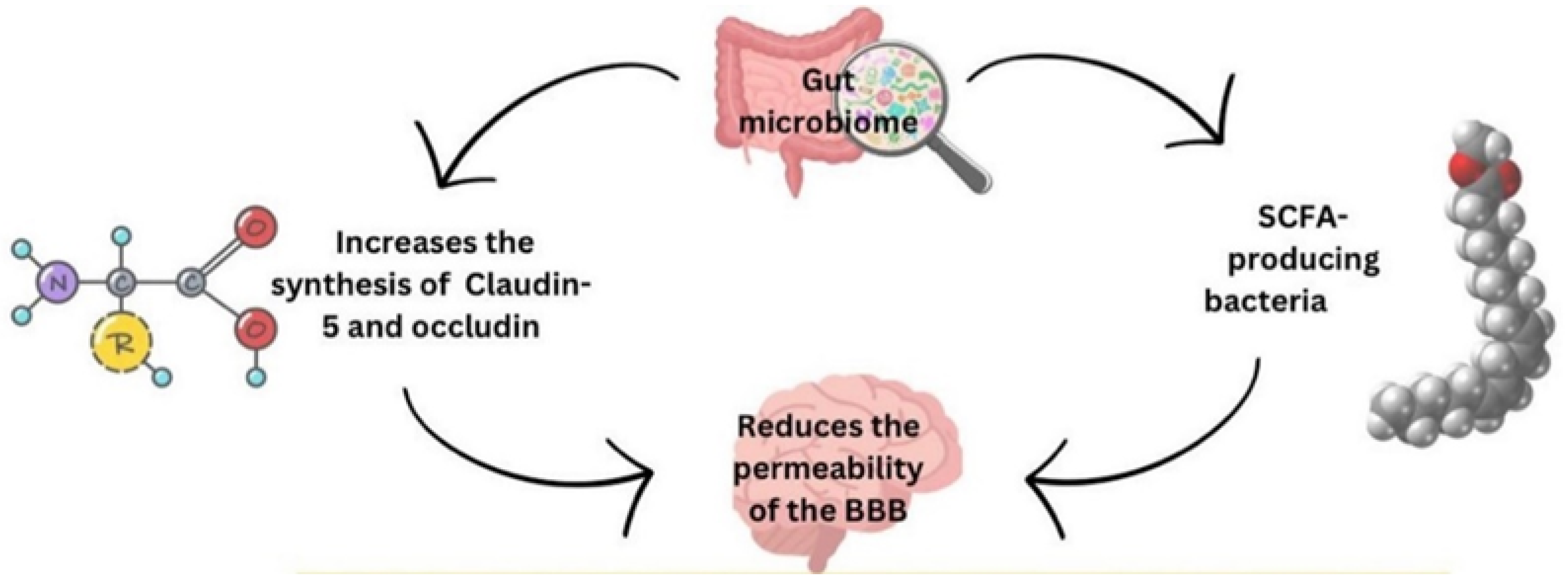
Figure 2.
This diagram shows the role of Metformin in modulating the gut microbiome to improve human brain health. SCFA: Short-Chain Fatty Acid.
Figure 2.
This diagram shows the role of Metformin in modulating the gut microbiome to improve human brain health. SCFA: Short-Chain Fatty Acid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



