
Submitted:
26 September 2025
Posted:
28 September 2025
You are already at the latest version
Abstract
Pulses and pseudocereals are sustainable protein sources of bioactive peptides (BAPs) with potential antioxidant, antihypertensive, antidiabetic, antimicrobial, and immunomodulatory activities. BAPs are typically liberated during gastrointestinal digestion or through bio-based processes, among which enzymatic hydrolysis and microbial fermentation represent the most widely applied strategies. Enzymatic hydrolysis provides controlled and reproducible release of short peptide motifs; recent advances such as ultrasound- or high-pressure–assisted hydrolysis enhance yield and bioactivity. Fermentation exploits microbial proteolytic activity to generate complex peptide mixtures, while improving sensory quality, reducing antinutritional compounds, and responding to consumer demand for natural and “clean-label” products. In silico tools increasingly complement these approaches by accelerating peptide discovery, predicting interactions with molecular targets, and guiding process design. This review provides an updated overview of bio-based methods to produce BAPs from pulses and pseudocereals, emphasizing the comparative advantages of enzymatic and fermentation technologies and their integration with computational tools. Moreover, it examines regulatory frameworks in the European Union, the United States, Japan, and China, while discussing current challenges for industrial scale-up and application in functional foods and nutraceuticals. These combined strategies offer a promising pathway to unlock the health and sustainability potential of plant proteins.
Keywords:
bioactive peptides
; pulses
; pseudocereals
; bioactivity
; bio-based approaches
1. Introduction
1.1. Pulses and Pseudocereals
Different reviews have highlighted the numerous health benefits of constant consumption of vegetable proteins. The preference of vegetable protein sources is therefore necessary, not only to address environmental challenges, but also to assist in the management of the rising incidence of chronic metabolic diseases.
The principal vegetable protein sources are pulses of which peas, lentils, beans, chickpeas that contain 12-38 % high-quality protein. Furthermore, a high level of essential amino acids (EAAs), particularly lysine, is also present [1,2,3,4]. Pseudocereals, notably including quinoa, amaranth, buckwheat, and chia, have garnered interest for their high-quality protein content (up to 10-20 %) and potential health benefits. In contradistinction to conventional cereals, pseudocereals provide a more balanced amino acid profile and do not present gluten, rendering them valuable for plant-based diets and functional foods [5,6,7].
1.2. Structural and Physicochemical Characteristics of Bioactive Peptides
The high nutritional and functional value of pulse and pseudocereal proteins is also attributed to the presence of encrypted bioactive peptide (BAP) sequences. These sequences, inactive within the native protein structure, can be released during gastrointestinal digestion or through bio-based strategies such as enzymatic hydrolysis and microbial fermentation. In legumes, storage proteins, particularly globulins and albumins, as well as lectins and protease inhibitors, represent the primary reservoirs of BAPs. Similarly, in pseudocereals, 11S globulins and albumins play a central role as precursors of health-promoting peptides. These peptide fragments have garnered attention for their potential in modulating physiological functions and contributing to the development of functional foods and nutraceuticals [8,9,10,11].
Most BAPs consist of 2 to 18 amino acids long with a molecular weight < 3 kDa. Due to their length, the bioactivity is mainly attributed to their amino acid sequences despite their secondary or tertiary structures BAPs are rich in hydrophobic amino acids that can give a bitter taste, which is challenging for commercial application in food products, and EAAs that contribute to healthy value. Many functional peptides have sequences rich in proline (Pro), leucine (Leu), arginine (Arg) and lysine (Lys). Health effects are linked to particular motifs, for example the sequences VPP or IPP are linked to antihypertensive effects due to their angiotensin-converting enzyme (ACE) inhibition action, meanwhile sequences containing glutamine (Gln)- or cysteine (Cys)-rich domains often exhibit antioxidant properties [9,10,12,13,14].
A plethora of studies have been conducted with the objective of investigating the bioactivity potential of these peptides [15,16,17,18,19]. The primary field of research pertains to metabolic disorders, encompassing conditions such as diabetes, hypertension, and obesity. This focus is underpinned by the multifaceted properties of these peptides, which exhibit antioxidant, antihypertensive, antidiabetic, and anti-inflammatory effects, thereby highlighting their potential for applications in related diseases [15,20,21,22]. In addition, several studies have been conducted that have focused on the impact of these peptides on the gut microbiota indeed, these studies have described BAPs capacity to promote the growth of probiotic bacteria and to inhibit the growth of pathogenic bacteria in the gut [17,23].
The present review aims to summarize recent advances in the production of BAPs from pulses and pseudocereals, with particular emphasis on bio-based processing approaches such as enzymatic hydrolysis and microbial fermentation, bio-informatic prediction tools, bioactivity screening, and their potential applications in the development of functional foods and nutraceuticals, considering the current legislation in the European Union, United States, China and Japan.
2. Bio-Informatic Approaches for Bioactivity Prediction
In silico prediction methods have emerged as powerful tools for the identification and characterization of BAPs derived from pulses and pseudocereals. These computational approaches enable the efficient screening of extensive protein databases and the prediction of peptide sequences with potential physiological benefits. By simulating enzymatic hydrolysis and assessing structural and physicochemical parameters, in silico tools facilitate the preliminary evaluation of peptides for specific bioactivities such as antihypertensive, antioxidant, antidiabetic, and anticancer effects prior to experimental validation (Figure 1). This strategy significantly accelerates the discovery pipeline, reducing the time and resources required for laboratory testing [9,24,25].
2.1. In Silico Identification of BAPs
The identification of BAPs has been greatly enhanced by bioinformatics tools. Traditionally, peptide discovery was based on enzymatic hydrolysis and labor-intensive experimental workflows, which limited throughput and scalability. In silico approaches enable rapid screening of food protein sequences using curated databases (e.g., UniProt, NCBI), enzymatic cleavage simulation platforms (e.g., BIOPEP, PeptideCutter), and repositories of experimentally validated BAPs (e.g., AHTPDB, FeptideDB). These tools predict potential BAPs based on known enzymatic cleavage sites and reported bioactivities, facilitating the exploration of sequences with antioxidant, antihypertensive, antidiabetic, and anti-inflammatory properties. Further insight into peptide–target interactions is provided by molecular docking and molecular dynamics (MD) simulations, which estimate binding affinities and reveal structural stability in silico. Integrated strategies combining proteolytic simulation, docking, and MD are increasingly used to optimize candidate selection before experimental validation [24,26,27].
2.2. Activity Prediction and Peptide Optimization via QSAR Modeling
Quantitative structure–activity relationship (QSAR) models offer a complementary layer of prediction by correlating peptide structural features with biological activity. These computational models use descriptors such as amino acid composition, physicochemical properties, or 3D conformational features to predict outcomes like inhibitory concentration (IC₅₀), receptor binding potential, or binary classification of bioactivity. QSAR approaches can be regression-based, classification-based, or increasingly, deep learning–driven. Recent advances include the use of global descriptors (e.g., 3D-QSAR, CoMFA, CoMSIA) and protein language models (e.g., ProtTrans, ESM-2), which allow accurate feature extraction from variable-length peptides. Although dataset limitations remain a bottleneck, particularly the lack of high-quality negative samples and curated training data, QSAR models are now widely employed to screen large libraries of peptide candidates, prioritize sequences for synthesis, and reduce experimental costs. In BAP research, QSAR is especially valuable for predicting multi-bioactivity profiles and assessing properties such as allergenicity, organoleptic characteristics, absorption, distribution, metabolism, excretion, and toxicity (ADMET), paving the way for the rational design of functional peptides [24,25,28,29].
2.3. Structure-Based Approaches: Molecular Docking and Dynamics
While peptide identification and QSAR modeling offer sequence- and activity-level predictions, molecular docking and molecular dynamics (MD) simulations provide structure-based insights on how BAPs interact with their biological targets [27]. Docking simulations predict the preferred binding conformations and affinities of peptides within enzyme active sites or receptor pockets, commonly targeting ACE and dipeptidyl peptidase-IV (DPP-IV) due to their relevance in hypertension and diabetes management. These methods enable virtual screening of multiple peptide candidates derived from in silico hydrolysis or mass spectrometry workflows. However, scoring functions in docking software are limited in accuracy and may not fully capture the flexibility of short peptides [24,30].
To overcome these limitations, MD simulations are increasingly used to refine docking results by modeling peptide–receptor complexes under physiological conditions over time. MD allows evaluation of complex stability, conformational changes, and key interactions at the atomic level. Techniques such as root-mean-square deviation (RMSD) analysis, hydrogen bond profiling, and binding free energy estimations via molecular mechanics Poisson–Boltzmann surface area (MM-PBSA) or molecular mechanics generalized Born surface area (MM-GBSA) provide more reliable predictions of peptide efficacy. Although computationally intensive, these approaches offer critical mechanistic insights that help bridge the gap between in silico predictions and biological function, guiding the rational design and validation of functional food-derived peptides [24].
3. Bio-Based Approaches for the Production of BAPs
The production of BAPs from pulses and pseudocereals has been predominantly achieved through two bio-based strategies, namely enzymatic hydrolysis and microbial fermentation as shown in Figure 2 [31]. Enzymatic hydrolysis relies on the use of exogenous proteases to release specific peptide sequences under controlled conditions, whereas microbial fermentation exploits the proteolytic machinery of selected microorganisms to liberate peptides during growth.
These approaches are not mutually exclusive and, in fact, differ substantially in terms of efficiency, scalability, and cost. Enzymatic hydrolysis provides precise and reproducible control over proteolysis but is often limited by enzyme cost and the need for downstream processing. Conversely, fermentation is slower and less predictable, yet it is economically attractive, sustainable, and capable of improving both nutritional and sensorial attributes. The following sections synthesize the most recent evidence on enzymatic and fermentation-based production of BAPs from pulses and pseudocereals, with emphasis on bioactivity, structural characterization, and technological relevance.
3.1. Enzymatic Treatments
Enzymatic hydrolysis has been the method of choice for most mechanistic studies, as it enables the targeted release of short peptides under defined physicochemical conditions. Proteases such as Alcalase, trypsin, pepsin, papain, chymotrypsin, and Flavourzyme are widely employed, and the degree of hydrolysis can be modulated, mainly setting the time of hydrolysis and combining different enzymes, to generate peptides of desired molecular size and bioactivity [32,33]. A consistent finding across legume and pseudocereal proteins is that low-molecular-weight peptides (< 3 kDa) are enriched in hydrophobic, aromatic, or proline residues, features strongly associated with ACE and DPP-IV inhibition. As illustrated in Table 1, there is a wide range of applications of enzymatic hydrolysis on different pulses and pseudocereal matrices, with the subsequent production and bioactivity outcomes. This paragraph will provide a description of these applications.
Production of BAPs from chickpea has been highlighted by Mora-Melgem et al. (2023), who reported that hydrolysis with papain and trypsin generated 92 peptides with predicted DPP-IV inhibitory capacity, of which a substantial proportion also exhibited ACE inhibition. The dipeptide His–Phe was singled out as the most potent inhibitor, consistent with its ability to interact with key catalytic residues of DPP-IV [33]. Moreover Quintero-Soto et al. (2021) showed the antidiabetic properties of albumin and globulin fractions of chickpea subjected to Alcalase hydrolysis and chromatographic purification. They reported the yield of short sequences (FEI, FIE, and FEL) that inhibited DPP-IV with an IC₅₀ of 4.20 µg/mL, while FGKG preferentially targeted carbohydrate-digesting enzymes, reducing α-glucosidase and α-amylase activities by 56% and 54%, respectively [34].
Lentil proteins subjected to simulated gastrointestinal digestion released peptides such as KLRT, TLHGMV, and VNRLM, which displayed strong ACE-inhibitory activity (IC₅₀ = 0.0197 mg/mL), confirming the value of gastrointestinally stable lentil peptides as antihypertensive agents [35]. In addition, Garcia-Mora et al. (2015) reported that lentil proteins subjected to enzymatic hydrolysis with different proteases, in combination with high-pressure treatment, generated peptides with marked bioactivities. Among the tested conditions, Savinase-assisted hydrolysis at 300 MPa produced the most potent effects, yielding the highest ACE-inhibitory activity (69.5 %) together with a strong antioxidant capacity (403.86 μM TE/g). These findings indicate that pressure-assisted enzymatic hydrolysis can significantly enhance the release of multifunctional peptides from lentil proteins [36].
Bollati et al. (2022), showed that commercial pea protein contains peptides with dual ACE- and DPP-IV-inhibitory potential. In fact, the pea protein hydrolysate inhibited DPP-IV and ACE in vitro with IC₅₀ values of 1.33 and 0.61 mg/mL, respectively, while its <3 kDa fraction displayed even stronger activity (IC₅₀ = 1.0 mg/mL for DPP-IV and 0.43 mg/mL for ACE). Cell-based experiments confirmed these findings, as pea protein hydrolysate reduced the activity of both enzymes in Caco-2 cells in a dose-dependent manner without cytotoxic effects [37]. Ásledóttir et al. (2023) evaluated the gastrointestinal stability and bioactivity of pea and faba bean proteins digested with standardized human gastric and duodenal juices (INFOGEST). Peptidomic analysis predicted the release of 275 BAPs from pea and 268 from faba bean. From these, selected sequences such as DKPWWPK and NEPWWPK (pea), together with QQGPPPPPPPISL and ATPPPPPPPMSL (faba bean), were further characterized in cell assays. All tested peptides significantly reduced IL-8 secretion by up to ~40%, highlighting their immunomodulatory potential and suggesting that both pea and faba bean proteins represent promising dietary sources of immune-active peptides [38].
Quinoa proteins also yield multifunctional peptides. Hydrolysis with chymotrypsin produced peptides with inhibitory effects on cholesterol esterase and pancreatic lipase, with sequences such as QHPHGLGALCAAPPST and HVQGHPALPGVPAHW identified as active binders. These results suggest that quinoa-derived peptides may exert antihyperlipidemic effects by targeting lipid metabolism enzymes [32]. Additional studies demonstrated that simulated gastrointestinal digestion of quinoa proteins produced fractions with both ACE-inhibitory and antioxidant activity, underscoring the versatility of quinoa peptides [39]. The antidiabetic activity of quinoa proteins has been investigated by Abbasi et al. (2022), who reported that hydrolysates obtained through sequential treatment with Alcalase and trypsin exhibited inhibitory effects on carbohydrate-digesting enzymes. In particular, the fraction with molecular weight ≥3 kDa generated after 0.5 h of hydrolysis displayed the strongest activity, achieving 44.8% inhibition of α-glucosidase [40].
Ayala-Niño et al. (2019) demonstrated that sequential hydrolysis with Alcalase and Flavourzyme of amaranth yielded fractions displaying not only ACE-inhibitory activity (IC₅₀ = 0.134–0.808 mg/mL) but also thrombin inhibition and substantial antioxidant activity, thereby illustrating the multifunctionality of amaranth peptides in cardiovascular health [41].
Soy and lupin proteins are among the most extensively investigated, particularly in relation to metabolic health. Lammi et al. (2019) showed that soy protein hydrolysates below 3 kDa inhibited HMG-CoA reductase, upregulated LDL receptor expression, and inhibited DPP-IV activity in Caco-2 cells, with peptides such as IAVPTGVA, IAVPGEVA, and LPYP identified as active contributors [42]. Earlier work by Lammi et al. (2016) reported that IAVPTGVA (Soy1) and LTFPGSAED (Lup1) acted as potent DPP-IV inhibitors, with molecular docking analyses confirming their interaction with catalytic sites. Collectively, these results highlight the relevance of soy and lupin peptides in the modulation of glycemic control and lipid metabolism [43]. Moreover, the bioactivity of soybean protein hydrolysate has been investigated by Bollati et al. (2022), who demonstrated that peptidomic analysis revealed the presence of sequences with both ACE- and DPP-IV-inhibitory motifs. In vitro assays confirmed a strong multifunctional profile, with soybean hydrolysate inhibiting DPP-IV (IC₅₀ = 1.15 mg/mL) and ACE (IC₅₀ = 0.33 mg/mL). The <3 kDa peptide fraction further enhanced these effects (IC₅₀ = 0.82 and 0.40 mg/mL, respectively), highlighting the role of short peptides in bioactivity. Importantly, soybean hydrolysate also suppressed DPP-IV and ACE activity in Caco-2 cells without impairing cell viability, confirming its potential as a multifunctional anti-diabetic and antihypertensive ingredient [37]. Soy hydrolysate was also reported by Boschin et al., (2014) who showed that peptic hydrolysis of soybean proteins (pepsin digestion, 18 h, E:S = 1:100; <3 kDa fraction) generated peptide mixtures with strong ACE-inhibitory activity. The hydrolysate inhibited ACE in a dose-dependent manner, with an IC₅₀ of 0.224 mg/mL and reaching 88% inhibition at the highest concentration tested (0.983 mg/mL). Under the same experimental conditions pea proteins showed moderate ACE-inhibitory activity compared with soybean. The hydrolysate achieved an IC₅₀ of 0.595 mg/mL and a maximum inhibition of 71% at the highest concentration tested (0.861 mg/mL) [44]. Coscueta et al. (2019) showed that soybean proteins hydrolyzed with Corolase PP (1% E:S, 4 h, 50 °C) released BAPs with dual functionality. The resulting hydrolysates exhibited enhanced antioxidant activity (3.9 ± 0.1 μmol TE/mg) and a strong ACE-inhibitory effect (IC₅₀ = 52 μg/mL), confirming their potential as multifunctional ingredients for managing oxidative stress and hypertension [45]. Although enzymatic hydrolysis enables reproducible peptide production and in-depth characterization, its limitations include enzyme costs, potential formation of bitter hydrophobic peptides, and the need for inactivation and purification steps. Nonetheless, technological advances such as ultrasound- or high-pressure–assisted hydrolysis are increasingly being employed to improve peptide yield and bioactivity [46].

Table 1.
Bioactive peptides (BAPs) obtained through enzymatic hydrolysis of pulse and pseudocereal proteins, with details on production methods and reported bioactivities.
Table 1.
Bioactive peptides (BAPs) obtained through enzymatic hydrolysis of pulse and pseudocereal proteins, with details on production methods and reported bioactivities.
| Source | BAPs production | Bioactivity | Ref | |||||
|---|---|---|---|---|---|---|---|---|
| Treatements | Outcome | Type | Dosage | Control | Outcome | |||
| Adzuki bean | Flavourzyme | Adzuki F2 fraction | Antimicrobial | 2 mg/mL | Gentamicin, chloramphenicol | S. typhimurium inhibition (76%) | [47] | |
| Amaranth | Sequential Alcalase + Flavourzyme hydrolysis | Bioactive peptides identified in fraction “45” (e.g., HVQLGHY, SQIDTGS, NWACTL) | Antihypertensive/ Antithrombotic/ Antioxidant | 10 mg/mL amaranth protein hydrolysate | Hippuric acid; thrombin; unhydrolyzed amaranth proteins | Multifunctional: ACE inhibition (IC₅₀ = 0.134–0.808 mg/mL), thrombin inhibition (IC₅₀ = 0.155–0.167 mg/mL), ABTS antioxidant SC₅₀ = 0.992–6.931 mg/mL; | [41] | |
| Chickpea | In silico enzymatic hydrolysis (papain and trypsin) | Prediction of ninety-two peptides with potential DPP-IV and ACE inhibition activity | Antidiabeitc/ Antihypertensive | NA | Omarigliptin (PDB ID: 4PNZ) | His–Phe identified as most potent; in silico predicted DPP-IV and ACE-inhibitory | [33] | |
| Chickpea | Alcalase hydrolysis of albumin and globulin; chromatographic purification | Bioactive sequences identified (e.g., FEI, FIE, FEL and FGKG) | Antioxidant/ Antidiabeitc | 1 mg/mL for hydrolysate; 0.2 mg/mL for peptide fraction; 0.1 mg/mL for α-amylase inhibition assay | Acarbose (1 mmol/L); reaction buffer | High radical scavenging (ABTS and DPPH) ; FEI, FIE FEL had DPP-IV inhibition (IC₅₀ = 4.20 µg/mL) while FGKG showed α-glucosidase and α-amylase inhibition (56% and 54%) | [34] | |
| Cowpea seed | Enzymatic hydrolysis on the protein isolate with Alcalase, 1:200 (E:S), 4h, 55°C, pH 7.8; SEC purification | Cowpea seed protein hydrolysate (< 1 kDa) | Antimicrobial | 25–150 μg/mL | Ciprofloxacin | Broad-spectrum antimicrobial, membrane disruption confirmed | [48] | |
| Faba bean | Simulated digestion with human gastric & duodenal juices (INFOGEST) | 268 BAPs predicted | Anti-inflammatory | 0.1–1000 µM | IL-1β with positive, negative (Fresh serum-free growth medium), and IL-1Rα controls | Selected peptides (e.g., QQGPPPPPPPISL, ATPPPPPPPPMSL) reduced IL-8 up to ~40%, indicating immunomodulatory activity | [38] | |
| Black Jampa bean | Hydrolysis on protein isolate (5%, w/v) using pepsin (90 min, pH 2.5) followed by pancreatin (120 min, pH 7.5) at 1:20 w/w, E:S | Bean protein hydrolysate fractions (phaseolin-rich) | Antioxidant | 100 μg hydrolysates or 50 μg peptide fractions | Blank | 0.7–1.0 kDa peptides had highest Cu²⁺ chelation and moderate Fe²⁺ binding | [49] | |
| Lentil | In vitro simulated GI digestion of lentil flour; ion-exchange and gel filtration fractionation | Fraction 5 contained peptides KLRT, TLHGMV, VNRLM | Antihypertensive | 50 µL of the peptide sample | Hippuryl-His-Leu solutions (1.0, 2.0, 3.0, 4.0 and 10.0 mm) | ACE inhibition, IC₅₀ = 0.0197 mg/mL | [35] | |
| Lentil | Protamex, Savinase, Corolase and Alcalase hydrolysis assisted with high-pressure | The process increased the concentration of peptides under 3 kDa, particularly with pressurization at 300 MPa by all enzymes tested | Antihypertensive/Antioxidant | 0.5 mg/mL (for ACE inhibition assay) | Non-hydrolysed lentil proteins | Highest ACE-inhibitory (69.46%) and antioxidant activity (403.86 μM TE/g) through Savinase treatment at 300 MPa | [36] | |
| Lupin | Enzymatic in silico prediction/ peptide synthesis | LTFPGSAED (Lup1), IC₅₀=228 µM | Antidiabetic | Concentrations ranging from 10 to 1000 μM | Sitagliptin (0.1 μM) | DPP-IV inhibitory; docking confirmed binding; other lupin peptides inactive | [43] | |
| Lupin | Lupin protein isolate (10% w/v) hydrolysed with Alcalase 2.4L (15 min at pH 8, 50 °C, and E/S = 0.3 AU/g protein) | Lupin protein hydrolysate | Antidiabetic | 100 mg/kg (mice), 1 g/day 28 days (human) | Placebo | Inhibited DPP-IV, improved glucose control in mice and humans | [50] | |
| Lupin | Sequentially digested with pancreatin (pH 7.5) and pepsin (pH 2.0), in a ratio 1:20 w/w (E:S) at 37 °C for 1 h | Andean lupin γ-conglutin hydrolysate | Antidiabetic | 5 mg/mL | Untreated cells; metformin (1 mM) and insulin (100 nM) | Inhibited DPP-IV, ↑GLUT4 translocation, ↓gluconeogenesis | [51] | |
| Lupin | Alcalase hydrolysis for 15 min at pH 8, 50 °C, and E/S = 0.3 AU/g protein | Lupin protein hydrolysate | Anti-inflammatory | 0.1–0.5 mg/mL | LPS-treated co-culture, unstimulated cells | Blunted TNF-α, IL-1β, IL-6 upregulation, IL-1β below baseline | [52] | |
| Lupin | Alcalase hydrolysis on lupin protein isolate for 15 min | Lupin protein hydrolysate | Anti-inflammatory | 100 mg/kg oral prophylactic | EAE mice untreated | Reduced severity, better neurologic function | [53] | |
| Lupin | Alcalase 2.4L (E:S = 0.3 AU/g protein); pH 8.0 at 50°C for 15 min | Lupin protein hydrolysate | Prebiotic | 100 mg/kg/day | HFD mice untreated | Reduced obesity, improved metabolism, restored Akkermansia abundance | [54] | |
| Mung bean | Flavourzyme | Mung bean F4 fraction | Antimicrobial | 2 mg/mL | Gentamicin, chloramphenicol | S. aureus inhibition (71%) | [47] | |
| Pea | Industrial hydrolysis, ultrafiltration | < 3 kDa peptide fraction | Antidiabetic / Antihypertensive | DPP-IV inhibitory assay: in vitro = 0.01 to 2.0 mg/mL; cellular assay = 1.0, 2.5, and 5.0 mg/mL; ACE inhibitory activity: in vitro = 0.08, 0.17, 0.35, 0.7, and 1.0 mg/mL; cellular assay = 0.1 to 5.0 mg/mL | control (C) sample; growth medium and H2O | Inhibited DPP-IV (IC₅₀ = 1.33 mg/mL; < 3 kDa fraction IC₅₀ = 1.0 mg/mL) and ACE (IC₅₀ = 0.61 mg/mL; < 3 kDa fraction IC₅₀ = 0.43 mg/mL); active in Caco-2 cells | [37] | |
| Pea | Simulated digestion with human gastric & duodenal juices (INFOGEST) | 275 BAPs predicted | Anti-inflammatory | 0.1–1000 µM | IL-1β with positive, negative (Fresh serum-free growth medium), and IL-1Rα controls | Selected peptides (e.g., DKPWWPK, NEPWWPK) reduced IL-8 up to ~40%, indicating immunomodulatory activity | [38] | |
| Pea | Pepsin hydrolysis of total protein extract (18 h, E/S = 1:100), ultrafiltration <3 kDa | Peptide mixtures with ACE-inhibitory activity | Antihypertensive | 861 μg/ml | Inhibitor Blank (AIB): enzyme but no inhibitor; Reaction Blank (ARB): highest inhibitor concentration but no enzyme. | ACE inhibition: IC₅₀ = 0.595 mg/mL; maximum inhibition 71% at the highest concentration tested (0.861 mg/mL) | [44] | |
| Quinoa | Papain digestion of quinoa bran globulin powder | Quinoa bran globulin hydrolysate (SAPPP fraction) | Antihypertensive | IC₅₀ = 915 μM | ND | Stable ACE inhibition after digestion, pH fluctuations (2.0–10.0), pasteurization conditions, addition of ions | [55] | |
| Quinoa | Chymotrypsin hydrolysis (QPI, 2 h) | Peptides identified (e.g. QHPHGLGALCAAPPST) | Anti-hypercholesterolemic | 25 – 50 μL of sample | p-nitrophenyl butyrate | Highest inhibition of cholesterol esterase (IC₅₀=0.51 mg/mL) and pancreatic lipase (IC₅₀=0.78 mg/mL | [32] | |
| Quinoa | Simulated GI digestion of protein isolates | Peptide fractions < 5 kDa and > 5 kDa | Antioxidant | ND | Trolox (0.2–1.6 nmol); | Peptides with antioxidant and colon cancer cell viability inhibitory activity identified; hydrolysates showed strong radical-scavenging activity | [39] | |
| Quinoa | Alcalase and trypsin hydrolysis; MW cut-off = 3, 10 kDa | Different molecular weight peptide fractions | Antidiabetic | ND | pNPG substrates | Highest α-glucosidase inhibition (44.8%) obtained with 0.5 h hydrolysis time and 3 kDa ≥ MW. | [40] | |
| Quinoa | High pressure-assisted enzymatic hydrolysis with 0.14 AU of Alcalase/ g protein (1:20 w/v E:S) | Quinoa protein hydrolysate (< 3 kDa) | Antihypertensive | 0.1–0.5 mg/mL | Blank, non-hydrolyzed proteins, conventional hydrolysis | High-pressure hydrolysis improved ACE inhibition | [56] | |
| Quinoa | Simulated gastro-intestinal digestion, only pepsin and pancreatin | Quinoa albumin peptides (lunasin-rich) | Anti-inflammatory | 1 mg/mL | IL-1β-treated cells | Up to 74% reduction in NF-κB activity. In vitro digestion enhanced the effect | [57] | |
| Red quinoa | Hydrolysis with Alcalase 2.4 LFG, 2 h. | Red quinoa protein hydrolysate | Antioxidant/ Antihypertensive |
1000 mg/kg/day (8 weeks) |
Hypertensive rats untreated (water) or with vitamin C | Increased glutathione, decreased MDA, improved systemic antioxidant status, reduction systolic blood pressure | [58] | |
| Soy | Hydrolysis with pepsin and trypsin; ultrafiltration (<3 kDa) | Peptic (P) and tryptic (T) soybean hydrolysates | Antidiabetic | Range of 0.5–2.5 mg/mL | NA | HMG-CoA reductase inhibition (up to −77%), ↑LDL receptor expression and LDL uptake, DPP-IV inhibition (up to 43% in Caco-2); peptides characterized by LC-MS/MS | [42] | |
| Soy | Enzymatic in silico prediction/ peptide synthesis | Glycinin hydrolysis (Soy1 = IAVPTGVA), IC₅₀=106 µM | Antidiabetic | Concentrations ranging from 10 to 1000 μM | Sitagliptin (0.1 μM) | DPP-IV inhibitory peptide; docking confirmed binding | [43] | |
| Soy | Industrial hydrolysis, ultrafiltration | < 3 kDa peptide fraction | Antidiabetic/ Antihypertensive | DPP-IV inhibition: 0.01–2.0 mg/mL (in vitro), 1.0–5.0 mg/mL (cells); ACE inhibition: 0.08–1.0 mg/mL (in vitro), 0.1–5.0 mg/mL (cells). | Control (C) sample; growth medium and H2O | Inhibited DPP-IV (IC₅₀ = 1.15 mg/mL; < 3 kDa fraction IC₅₀ = 0.82 mg/mL) and ACE (IC₅₀ = 0.33 mg/mL; < 3 kDa fraction IC₅₀ = 0.40 mg/mL); active in Caco-2 cells | [37] | |
| Soy | Corolase PP hydrolysis (1% E:S, 4h, 50 °C) | Soy protein hydrolysate as biofunctional ingredient | Antihypertensive/ Antioxidant | 40 μL of sample | Trolox; ACE + Abz-Gly-Phe(NO2)-Pro | Improved antioxidant activity (3.9 ± 0.1 μmol TE/mg) and ACE inhibitory peptides (IC50 = 52 μg/mL) | [45] | |
| Soy | Pepsin hydrolysis of total protein extract (18 h, E/S = 1:100), ultrafiltration <3 kDa | Peptide mixtures with ACE-inhibitory activity | Antihypertensive | 983 μg/mL | Inhibitor Blank (AIB): enzyme but no inhibitor; Reaction Blank (ARB): highest inhibitor concentration but no enzyme. | ACE inhibition: IC₅₀ = 0.224 mg/mL; maximum inhibition 88% at the highest concentration tested (0.983 mg/mL) | [44] | |
| Soy | Simulation of Gastrointestinal Digestion | Germinated soybean peptides (5–10 kDa) | Antidiabetic | IC₅₀ 0.91 mg/mL | Diprotin A- | DPP-IV inhibition, active sequences from β-conglycinin, glycinin, P34 | [59] | |
| Soy | Proteinase PROTIN SD-NY10 (EC 3.4.24.28), 0.05% w/w, 50–55°C for 16 h | Soymilk hydrolysate tetrapeptide | Antihypertensive | 80 μg/kg/day (3 weeks) | Untreated SHR | Lowered BP, strong ACE inhibition | [60] | |
NA = information not available.
3.2. Production of BAPs Through Fermentation
Fermentation represents a complementary bioprocess that exploits the proteolytic capacity of microorganisms, particularly lactic acid bacteria, to release BAPs. In addition to peptide liberation, fermentation may confer benefits such as improved flavor, reduced antinutritional factors content, and the generation of additional bioactive metabolites. This makes fermentation highly attractive for the development of functional foods, even if it is generally less predictable than enzymatic hydrolysis [31,61]. As shown in Table 2, microbial fermentation has been explored in different pulse and pseudocereal matrices, resulting in diverse peptide production and bioactivities. In the following, we describe these applications more closely.
Chickpea proteins have been widely investigated as substrates for fermentation-driven bioactivity. Li and Wang (2021) showed that solid-state fermentation with Bacillus subtilis lwo promoted extensive proteolysis, generating low-molecular-weight peptides (<10 kDa, up to 25.8 mg/g after 12 h) and enhancing antioxidant capacity in a time-dependent manner [64]. Expanding on these findings, Chiacchio et al. (2025) demonstrated that lactic acid fermentation of chickpea puree with diverse LAB strains markedly improved the nutritional profile of the resulting flours, with increases in polyphenolic content and the release of BAPs including predicted DPP-IV and ACE inhibitors [65]. More recently, Xu et al. (2025) optimized fermentation with selenium-enriched Bacillus natto, achieving an ACE-inhibition rate of ~80.7% under specific conditions (2% inoculum, 19:1 liquid-to-solid ratio, 40 °C), highlighting the potential of Se-enrichment to combine nutritional fortification with antihypertensive functionality [66]. Collectively, these studies underscore the versatility of fermentation approaches to unlock antioxidant, antidiabetic, and antihypertensive peptides from chickpea proteins.
Studies on faba bean demonstrate the strong potential of fermentation. Jakubczyk et al. (2017) fermented faba bean flour with Lactiplantibacillus plantarum 299v at 30 °C for three days and identified multiple peptides in the <3 kDa fraction, which displayed potent ACE inhibition (IC₅₀ = 0.05 mg/mL). These sequences contained di- and tri-peptide motifs (e.g., GL, DA, MY) previously associated with antihypertensive activity. In peas, the same group showed that seven-day fermentation with L. plantarum 299v had limited effects on the intact protein, but subsequent gastrointestinal digestion released peptides that improved ACE-inhibitory activity (IC₅₀ = 0.19 mg/mL versus 0.37 mg/mL in the control), with KEDDEEEEQGEEE identified as an active sequence [35,67,68].
Tonini et al. (2024) explored the fermentation of red-lentil protein isolates with multiple LAB and yeasts, identifying strain-dependent differences in peptide release. The yeast Hanseniaspora uvarum SY1 produced the richest profile of BAPs, particularly those with antioxidant and ACE-inhibitory activities, underscoring the potential of non-conventional microbial starters for generating health-promoting peptides from lentils [69].
In lupin, Ayyash et al. (2019) reported that solid-state fermentation with L. plantarum K779 significantly improved both antioxidant and ACE-inhibitory activities compared to the unfermented controls, likely reflecting synergistic effects of liberated peptides and phytochemicals [70].
Fermentation also enhances the bioactivity of pseudocereals. Cruz-Casas et al. (2023) investigated the impact of fermentation with various lactic acid bacteria and Bacillus spp. on amaranth proteins, obtaining hydrolysates with bioactive properties. The extracts displayed strong antioxidant activity (9.18 μM TE/L), notable ACE-inhibitory effects (80.65%), and antimicrobial activity against foodborne pathogens, underscoring amaranth as a versatile pseudocereal for multifunctional peptide production through fermentation [62]. Morevoer, Rizzello et al. (2012) demonstrated that amaranth fermentation with Lactobacillus spp. induced the production of lunasin, a BAP with documented anticancer and cholesterol-lowering activity [63].
Solid-state fermentation with L. plantarum K779 on quinoa, applied in the study of Ayyash et al. (2019), showed significant improvement in ACE-inhibitory activities when compared to the unfermented control [70]. Li et al. (2022) reported that fermentation of quinoa flour with Lactobacillus paracasei CICC 20241 led to the release and identification of several antihypertensive peptides. Among them, NIFRPFAPEL and AALEAPRILNL were characterized as potent ACE inhibitors, with IC₅₀ values of 49.02 µM and 79.72 µM, respectively. These findings confirm that microbial fermentation can unlock quinoa-derived peptides with significant potential for blood pressure regulation [71]. Buckwheat fermentation has also been optimized: Wang and Ma (2024) reported that solid-state fermentation with L. plantarum under optimized conditions increased peptide content to 22.2 mg/mL, outperforming enzymatic methods in both yield and sensory improvement [72].
Fermentation thus offers a cost-effective and consumer-acceptable strategy for BAP production, with additional benefits and nutritional enhancement. Its limitations, however, include longer processing times, batch variability, and the need for rigorous microbial management.
Table 2.
Bioactive peptides (BAPs) obtained through microbial fermentation of pulse and pseudocereal proteins, with details on production methods and reported bioactivities.
Table 2.
Bioactive peptides (BAPs) obtained through microbial fermentation of pulse and pseudocereal proteins, with details on production methods and reported bioactivities.
| Source | BAPs production | Bioactivity | Ref | ||||
|---|---|---|---|---|---|---|---|
| Treatements | Outcome | Type | Dosage | Control | Outcome | ||
| Amaranth | Fermentation with various lactic acid bacteria and Bacillus spp | Multi potential dought | Antihypertensive/ Antimicrobial | 5-10 µL of sample for antioxidant and ACE-inhibtiory assays; 50 µL for antimicrobial activity assay |
Control time 0 min (unfermented amaranth dough) and 24 min (spontaneously fermented dough) | antioxidant (9.18 μM TE/L), ACE-inhibitory (80.65 %) and antimicrobial activities against pathogens | [62] |
| Amaranth seed protein hydrolysates | Fermentation with Enterococcus faecium vs. enzymatic hydrolysis | Protein hydrolysates contain novel peptides with antihypertensive activity | Antihypertensive | 0.00625 mg/mL | Blank | Enterococcus faecium hydrolysate strongest (79% inhibition) | [73] |
| Buckwheat | Solid-state fermentation with L. plantarum (12.87% inoculum, 60% moisture, 31.4 °C, 6 d) | Peptide content 22.18 mg/mL under optimal conditions; fermentation produced high peptide levels (and better flavor) | NA | NA | NA | NA | [72] |
| Chickpea | Fermentation of 20% chickpea puree with 14 LAB strains; 48 h; flours obtained by freeze-drying | Flour with enhanced bioactive peptides | Antidiabetic/ Antihypertensive | 200 µL of the sample extract | Trolox | Higher polyphenolic content; BAPs incl. DPP-IV/ACE-inhibitor candidates | [65] |
| Chickpea | Fermentation with selenium-enriched Bacillus natto | Under optimized condition (2% inoculum, 19:1 liquid-to-solid ratio (mL/g), and 40 °C) | Antihypertensive | 40 μL | HEPES (80 mmol/L) | ACE-inhibition rate ~80.7% | [66] |
| Chickpea | Solid-state fermentation with Bacillus subtilis lwo (SSF) | Peptides with MW < 10 kDa produced after 12h of fermentation (25.8 mg/g). | Antioxidant | 0.5-1 mL of extract | Control groups (blank or without extract/salicylic acid) | Antioxidant activity increases with fermentation time. | [64] |
| Chickpea | L. acidophilus fermentation of pretreated dried chickpeas | Fermented chickpea protein peptides | Antidiabetic | 5 mg/mL | Unfermented chickpea | L. acidophilus–fermented peptides suppressed α-glucosidase by >58% | [74] |
| Faba bean | Fermentation of faba bean flour with L. plantarum 299v (30 °C, 3 d) | 6 peptide sequences; most active fraction (3 kDa) contained di-/tripeptide motifs (e.g. GL, DA, MY) | Antihypertensive | NA | Control sample (without fermentation) | 3.5–7.0 kDa peptides with higher ACE inhibitory activity (IC50 of 0.28 mg/mL); | [67] |
| Lentil | Red-lentil protein isolate fermented with multiple LAB and yeasts | H. uvarum SY1 led to the highest abundance of BAPs | Antioxidant/ Antihypertensive | Different concentrations | Trolox; unfermented red lentil protein isolate | Antioxidant and ACE-inhibitory activities | [69] |
| Lupin | Solid-state fermentation with L. plantarum K779 (35 °C, 72 h) | Fermentation liberates peptides and phytochemicals | Antioxidant/ Antihypertensive | 20-200 µL | Non-inoculated samples | Enhanced antioxidant and ACE-inhibitory activity compared to raw | [70] |
| Pea | Fermentation of pea seeds with L. plantarum 299v (22 °C, 7 d) | LC fractions yielded peptide KEDDEEEEQGEEE | Antihypertensive | NA | Sample without fermentation process | After fermentation + simulated digestion, ACE inhibition IC₅₀ = 0.19 mg/mL (vs 0.37 control); | [68] |
| Quinoa | Solid-state fermentation with L. plantarum K779 (35 °C, 72 h) | Fermentation liberates peptides and phytochemicals | Antihypertensive | 20-200 µL of sample | non-inoculated samples | Enhanced antioxidant and ACE-inhibitory activity compared to raw | [70] |
| Quinoa | Solid fermentation of quinoa flour with Lactobacillus paracasei CICC 20241 | 5 potential ACE inhibitory peptides | Antihypertensive | 0.05 and 0.1 mg/mL | ACE without peptides | NIFRPFAPEL: IC50 = 49.02 µM; AALEAPRILNL IC50 = 79.72 µM) | [71] |
| Soy | Prozyme pretreated soy protein isolate fermented with Lactobacillus rhamnosus EBD1 (48 h at 37°C) | Fermented soy protein hydrolysate | Antihypertensive/ Prebiotic |
10-100 mg/kg hydrolysate per kg BW/day (6 weeks) | Spontaneously hypertensive rats (SHR) water or captopril | Rapid BP reduction, inhibited ACE, improved NO and SOD, remodelled gut microbiota (↓Firmicutes/Bacteroidetes ratio) | [75] |
| Soybean peptides | NA | Novel peptides identified | Prebiotic | NA | Undigested protein, MRS, FOS | Stimulated L. reuteri growth, unique fermentation profile | [76] |
NA = information not available.
3.3. Comparative Perspective
When considered together, enzymatic hydrolysis and microbial fermentation should not be seen as competing but rather as complementary approaches for the production of BAPs (Figure 3). From a practical standpoint, enzymatic hydrolysis provides high reproducibility and control over proteolysis, making it particularly well suited for mechanistic studies, structure–activity relationship investigations, and the generation of purified peptides for multiple applications. Industrially, hydrolysis can be scaled through batch or continuous reactor systems, but costs remain heavily dependent on enzyme price, stability, and the need for inactivation and downstream purification. Furthermore, hydrolysates often require debittering due to the release of hydrophobic peptides, a factor that complicates direct use in food matrices.
Fermentation, in contrast, offers a low-cost and sustainable strategy, since microorganisms grow on inexpensive substrates and continuously generate proteolytic enzymes in situ. This approach is especially advantageous when the goal is to enrich whole flours or fermented foods with peptides, while simultaneously improving sensory and nutritional attributes (e.g., reduction of antinutritional factors, enhanced digestibility, and flavor development). Fermentation is also aligned with consumer expectations for natural and “clean-label” products. However, fermentation is less predictable than enzymatic hydrolysis; peptide yield and bioactivity depend strongly on strain selection, inoculum size, substrate composition, and environmental parameters such as pH, temperature, and moisture content. Moreover, batch-to-batch variability and the risk of contamination remain critical challenges in large-scale applications.
In terms of regulatory and market perspectives, enzymatic hydrolysates are closer to purified nutraceuticals, allowing more straightforward characterization, standardization, and dose definition. Fermented products, on the other hand, fit more readily into functional foods but typically deliver complex peptide mixtures whose exact contributions to health effects are more difficult to isolate. A promising direction for future development is therefore the integration of both approaches, for example by combining controlled enzymatic pretreatment with fermentation, in order to maximize peptide yield and multifunctionality while mitigating the drawbacks of each method.
4. Bioactivities
The growing global emphasis on sustainable nutrition and disease prevention has placed plant-derived proteins at the forefront of scientific interest, particularly those from pulses and pseudocereals. These protein sources are increasingly recognized not only for their nutritional value but also for their capacity to generate bioactive compounds with significant health-promoting properties. Of particular interest is the potential of these proteins to serve as precursors for BAPs.
Pulses such as soybean, chickpea, pea, lupin, and common bean, along with pseudocereals like quinoa, amaranth, and buckwheat, have demonstrated promising bioactivities through a wide range of mechanisms. Numerous studies have highlighted their protective roles against chronic and degenerative diseases, including cardiovascular disorders, type 2 diabetes, and inflammation-related conditions. These molecules can influence metabolic pathways by modulating immune responses, improving insulin sensitivity, lowering blood pressure via ACE inhibition, and exerting antioxidant or antimicrobial effects (Figure 4).
The functional attributes of these peptides are not only dependent on their amino acid composition but also on processing methods, which significantly influence their chain length, solubility, and bioactivity. Thus, the degree of hydrolysis and the physicochemical properties of the resulting peptide fractions are critical determinants of their efficacy. Overall, the ability to derive multifunctional BAPs from pulses and pseudocereals opens avenues for the development of next-generation functional foods and nutraceuticals.
The following sections provide a comprehensive overview of the bioactivities investigated for hydrolysates and peptides produced through enzymatic techniques (Table 1) and microbial fermentation (Table 2).
4.1. Antioxidant Activity
Recent studies have increasingly highlighted the potent antioxidant potential of peptides derived from pulses and pseudocereals, revealing multiple underlying mechanisms through which they exert their bioactivity [57]. These peptides demonstrate the capacity to directly scavenge reactive oxygen species (ROS), chelate redox-active metal ions, and modulate intracellular antioxidant defenses. What is particularly compelling is the role of specific amino acid residues, especially hydrophobic or aromatic ones like tyrosine and tryptophan, which appear to enhance radical scavenging efficacy, especially when located in short peptide sequences [77,78].
In cellular systems (damage model of mouse skeletal muscle myoblast), peptides isolated from Tartary buckwheat have been shown to significantly reduce ROS accumulation and lipid peroxidation [79]. This protective effect seems to be linked to the stabilization of mitochondrial membrane potential and a concurrent rise in catalase activity, pointing toward preservation of mitochondrial integrity under oxidative stress. Similar protective responses have been observed in hepatocytes treated with novel pea-derived peptides, including YLVN, EEHLCFR, and TFY [80]. These peptides, obtained by a series of purification steps including ultrafiltration, ion exchange chromatography, G25 gel filtration chromatography, and reversed-phase chromatography, not only demonstrated strong in vitro DPPH and ABTS radical scavenging activity but also enhanced cell viability under H₂O₂-induced oxidative stress. Their antioxidant effects extended to the prevention of intracellular ROS accumulation and the upregulation of key enzymes involved in antioxidant defense, such as superoxide dismutase (SOD) and catalase (CAT).
Moreover, computational docking studies suggest that these peptides may activate the Keap1–Nrf2 signaling pathway, a central regulator of cellular redox homeostasis. By disrupting Keap1-mediated inhibition of Nrf2, the peptides potentially facilitate increased expression of downstream antioxidant enzymes like heme oxygenase-1 (HO-1), thereby amplifying endogenous cytoprotection mechanisms.
The antioxidant potential of these peptides has also been demonstrated in vivo. In hypertensive rat models, oral administration of a red quinoa protein hydrolysate led to a significant increase in tissue glutathione levels alongside a marked reduction in malondialdehyde (MDA), a lipid peroxidation marker [58]. These outcomes reflect an enhanced systemic antioxidant status and further support the relevance of these peptides beyond cellular models.
Additional findings suggest that metal chelation is another critical aspect of the antioxidant function of legume-derived peptides. By binding redox-active metals such as iron and copper, these peptides may effectively inhibit reactions that generate harmful hydroxyl radicals. Collectively, the evidence points to a multifaceted antioxidant action, ranging from direct radical scavenging and metal chelation to the activation of redox-sensitive signaling pathways. Peptides derived from bean protein isolate and phaseolin have shown notable metal chelating activity following enzymatic hydrolysis [49]. Among the resulting size-fractionated peptides, those in the 0.7–1.0 kDa range exhibited the highest copper chelation (up to 82%), while iron binding was strongest in select isolate fractions, though generally lower. Phaseolin peptides appeared to be the main contributor to both antioxidant and copper chelating effects.
4.2. Anti-Inflammatory Effect
BAPs from legumes and pseudocereals can suppress inflammatory signaling, chiefly by downregulating NF-κB and pro-inflammatory cytokines. Quinoa albumin fractions (rich in the anti-inflammatory lunasin, a 43-amino-acid peptide [81]) reduced IL-1β-induced NF-κB/IL-8 expression by up to 74% [57]. Importantly, in the same study, simulated gastrointestinal digestion appears to enhance these effects; digests of quinoa and buckwheat protein fractions and peptides significantly reduced IL-1β–driven cytokine release in Caco-2 cells. Collectively, the evidence indicates that digestion-released peptides, with low-molecular-weight fragments (≤ 14 kDa), may exert cytoprotective effects by mitigating NF-κB–driven transcriptional activation in response to inflammatory stimuli.
A recent study on the Panhandle Pride white bean cultivar, developed by the University of Nebraska, demonstrated that the <3 kDa peptide fraction markedly attenuates TNF-α–induced inflammatory and oxidative responses in human vascular endothelial cells [82]. Among all tested molecular weight fractions, only the <3 kDa filtrate significantly suppressed TNF-α–driven expression of key inflammatory biomarkers, including intracellular adhesion molecule-1 (ICAM-1), vascular adhesion molecule-1 (VCAM-1), and the chemokine MCP-1. This fraction, enriched in γ-glutamyl peptides and hydrophobic dipeptides, retained its bioactivity following simulated gastrointestinal digestion. Notably, the digested filtrate continued to downregulate VCAM-1, E-selectin, and MCP-1 expression at peptide concentrations as low as 250 μg/mL.
Comparable anti-inflammatory potential has been observed in lupin protein hydrolysate. Using a Caco-2/THP-1 macrophage co-culture model to simulate intestinal absorption, the hydrolysate at 0.1–0.5 mg/mL effectively counteracted LPS-induced upregulation of pro-inflammatory cytokine mRNA (TNF-α, IL-1β, IL-6), restoring expression levels to those of unstimulated controls [52]. Notably, IL-1β mRNA levels were reduced below baseline, indicating a pronounced anti-inflammatory effect.
The immunomodulatory properties of lupin protein hydrolysate have also been confirmed in vivo. In a murine model of experimental autoimmune encephalomyelitis, an established model of multiple sclerosis characterized by elevated pro-inflammatory cytokine production during the acute phase, prophylactic administration of lupin protein hydrolysate at 100 mg/kg significantly attenuated disease severity. Treated mice maintained higher functional performance scores relative to controls, exhibiting milder neurological deficits such as unilateral hind-limb paralysis or impaired righting reflex [53].
4.3. Antihypertensive Effect
Many legumes and pseudocereal-derived peptides lower blood pressure by targeting the renin–angiotensin system and enhancing vasodilatory pathways. The dominant mechanism is ACE inhibition, which prevents formation of the vasoconstrictor angiotensin II and preserves vasodilator bradykinin. In vitro studies have identified many ACE-inhibitory sequences encrypted in these proteins. For example, a novel pentapeptide SAPPP was purified from quinoa bran globulin hydrolysate with an ACE-inhibitory IC₅₀ of 915 μM, while the activity of SAPPP after the digestion was calculated to be 926 μM [55]. In spontaneously hypertensive rats, orally administered pulse-derived peptides have shown clear antihypertensive effects [60]. A tetrapeptide from enzymatically treated soymilk, FFYY, given at only 80 µg/kg body weight per day, significantly attenuated the rise in systolic and diastolic blood pressure in SHR over 3 weeks, without adverse effects. The tetrapeptide showed strong inhibitory activity to the ACE-enzyme with an inhibitory concentration of 1.9 µM. Similarly, Daliri et al. (2020) demonstrated that a single oral dose of a fermented soybean protein hydrolysate produced a rapid antihypertensive effect in SHR [75]. Systolic blood pressure fell by 25 mmHg and diastolic by 40 mmHg within 6 hours of the hydrolysate administration, comparable to the ACE inhibitor drug captopril. Mechanistically, fermented soybean protein hydrolysate feeding acutely inhibited serum ACE activity in the rats while elevating vasoprotective mediators (increased nitric oxide and superoxide dismutase levels, reduced malondialdehyde). Notably, the soy peptide treatment remodelled the gut microbiome of hypertensive rats, suggesting a gut-mediated contribution to its antihypertensive action (see Section 4.6).
Quinoa proteins have likewise yielded antihypertensive peptides. In vitro, quinoa protein hydrolysates obtained through high pressure-assisted enzymatic hydrolysis displayed strong ACE-inhibitory activity [56]. The < 3 kDa permeate, characterized by a higher degree of hydrolysis, was particularly effective at concentrations of 0.1 and 0.5 mg/mL, showing greater inhibition than either non-hydrolyzed proteins or hydrolysates produced under atmospheric pressure. This fraction contained several peptides with potent ACE-inhibitory potential, among which GSHWPFGGK, FSIAWPR, and PWLNFK exhibited the highest PeptideRanker scores.
Similarly, amaranth proteins have been shown to release potent antihypertensive peptides upon fermentation. Hydrolysates from Amaranthus hypochondriacus seeds fermented with Enterococcus faecium LR9 displayed the strongest ACE inhibitory activity (79%) at a concentration of 0.00625 mg/mL, outperforming those obtained with Leuconostoc mesenteroides 18C6 (68%) and enzymatic hydrolysis with Alcalase (69%) [73]. Peptide identification by tandem mass spectrometry revealed 125 sequences derived mainly from agglutinin, cystatin, 11S globulin, and trypsin inhibitor. Notably, the highest degree of proteolysis did not correspond to the strongest inhibition, suggesting that specific peptide release, rather than overall hydrolysis, determines ACE inhibitory capacity.
4.4. Antidiabetic Effect
A key mechanism in antidiabetic effect is inhibition of dipeptidyl peptidase-IV (DPP-IV), the enzyme that degrades incretin hormones like GLP-1 [83]. By inhibiting DPP-IV, food-derived peptides prolong endogenous GLP-1 action, thereby enhancing insulin secretion and improving glycemic control. Several studies have identified potent DPP-IV inhibitory peptides from legume and pseudocereal proteins.
Lupin-derived peptides have garnered attention for multi-faceted antidiabetic effects. A recent study characterized an Alcalase-generated lupin protein hydrolysate (LPH) and demonstrated its anti-diabetic activity via DPP-IV modulation [50]. Although lupin protein consumption was previously known to improve glucose metabolism, direct evidence of DPP-IV inhibition was lacking. The study confirmed that LPH potently inhibits DPP-IV in vitro and in cultured cells, leading to prolonged GLP-1 activity. The authors demonstrated that more than 140 peptides derived from lupin protein hydrolysate (LPH) are capable of trans-epithelial intestinal transport. The bioactivity of these peptides was further validated in vivo, where LPH supplementation inhibited DPP-IV activity and exerted glucose-lowering effects both in mice (100 mg/kg) and in humans following a 28-day intervention with a beverage containing 1 g of hydrolysate per 200 mL serving.
Inhibition of digestive enzymes such as α-glucosidase and α-amylase represents another antidiabetic mechanism of legume-derived peptides. Chickpea peptides, in particular, display strong α-glucosidase inhibitory activity. Ma et al. optimized a fermentation-based method to produce chickpea peptides with enhanced antidiabetic activity [74]. The fermentation with microbial strains generated low-molecular-weight fractions that suppressed yeast α-glucosidase activity by over 58% at 5 mg/mL, with Lactobacillus acidophilus–fermented chickpea producing the most potent inhibitors.
Soybean-derived peptides exhibit dual enzyme inhibition [59]. Germinated soybean produced 5–10 kDa fractions with strong DPP-IV inhibitory activity (IC₅₀ 0.91 mg/mL), alongside significant inhibition of salivary α-amylase and intestinal α-glucosidases. The active sequences, ranging from 6 to 18 amino acids, were predominantly derived from β-conglycinin, glycinin, and P34 probable thiol protease.
Beyond digestive enzyme inhibition, certain pulse-derived peptides modulate glucose metabolism through effects on insulin signaling and hepatic pathways [51]. Hydrolyzed γ-conglutin (Cgh) from Andean lupin, originates from the Andean region of Ecuador, completely inhibited DPP-IV activity at 5 mg/mL and enhanced glucose uptake 6.5-fold in an enterocyte/adipocyte co-culture by promoting GLUT-4 translocation. Additionally, in an enterocyte/hepatocyte system, Cgh reduced gluconeogenesis by 50% and downregulated phosphoenolpyruvate carboxykinase expression.
4.5. Antimicrobial Capacity
Antimicrobial peptides are generally classified into four main subgroups based on charge and structure: anionic, cationic α-helical, cationic β-sheet, and extended cationic peptides. Anionic antimicrobial peptides often interact with microbial membranes through metal ion–mediated salt bridges. Cationic α-helical peptides are amphipathic and disrupt membranes after adopting α-helical conformations. Cationic β-sheet antimicrobial peptides are stabilized by intramolecular disulfide bonds; their bactericidal action relies on electrostatic binding to negatively charged microbial surfaces followed by membrane permeabilization. Extended cationic AMPs, rich in residues such as proline, arginine, or tryptophan, lack defined secondary structures but insert into and destabilize membranes through hydrophobic and electrostatic interactions. Indeed, several studies in the past few years have isolated plant defence peptides or generated hydrolysates from pulses that are active against bacteria. Hydrolysates of adzuki bean and mung bean prepared with peptidase from Aspergillus oryzae showed fraction-dependent antibacterial activity against major foodborne pathogens [47]. Hydrolysate fraction from adzuki bean achieved 76% inhibition of Salmonella typhimurium, while mung bean peptides fraction inhibited 71% of Staphylococcus aureus at 2 mg/ mL, effects comparable to gentamicin. LC–MS/MS identified short, cationic peptides, with lysine-rich sequences most effective against S. aureus and proline motifs linked to Salmonella inhibition.
Osman et al. produced an Alcalase hydrolysate of cowpea seed proteins and fractionated it by size, finding strong antimicrobial action against both Gram-positive and Gram-negative pathogens [48]. MIC for cowpea peptides (in the <1 kDa fraction) was as low as 25 µg/mL against E. coli, P. aeruginosa, and S. pyogenes, while 100–150 µg/mL was needed against S. aureus and Listeria (which are Gram-positive and generally more peptide-resistant). Even at those levels, the cowpea hydrolysate was highly effective, reducing bacterial loads substantially. Electron microscopy of treated bacteria revealed membrane damage and leakage, confirming a lytic mode of action. Mass spectrometry analysis identified multiple small di-peptides (180–370 Da) and one penta-peptide (659 Da) in the active cowpea fraction. Among the ten peptides identified in positive ion mode, three contained arginine residues, classifying them as cationic peptides with net positive charge. Similarly, two of the five peptides detected in negative ion mode also carried arginine, confirming their cationic nature.
Complementarily and beyond growth inhibition, a red kidney bean peptide extract at the MIC (1.25 mg/mL) suppressed biofilm formation by up to 87% and markedly reduced quorum sensing–regulated virulence factors in P. aeruginosa (pyocyanin and pyoverdine) [84]. These findings suggest a dual mechanism: direct antimicrobial effects and anti-virulence activity that may limit resistance development.
4.6. Prebiotic Effect
Prebiotics were long regarded solely as non-digestible carbohydrates that stimulate beneficial gut microbes. The latest consensus from the International Scientific Association for Probiotics and Prebiotics (ISAPP), however, defines them as “a substrate that is selectively utilized by host microorganisms conferring a health benefit.” [85]. This broader definition opens the possibility for non-carbohydrate compounds, including peptides, to be considered prebiotics when they modulate the gut microbiota in a health-promoting manner. As noted above, fermented soy protein hydrolysate (P-SPI) not only lowered blood pressure in SHR but also markedly improved gut microbiota diversity and balance. In Daliri et al.’s experiment, in hypertensive rats, fermented soy protein hydrolysates restored microbial diversity and reduced the Firmicutes/Bacteroidetes ratio, a dysbiosis marker associated with hypertension. Chronic supplementation enriched short-chain fatty acid (e.g. propionate) producing genera, while decreasing pro-inflammatory taxa such as Streptococcaceae and Erysipelotrichales.
Other studies echo these findings. After simulated gastrointestinal digestion, soybean-derived peptides (DPPs) stimulated the growth of Lactobacillus reuteri more effectively than non-digested soybean proteins, undigested peptides, or the MRS control medium [76]. Metabolic activity was further assessed by monitoring lactic acid production. Interestingly, lactic acid levels in the FOS and FOS + DPP groups were significantly lower than those in the DPP-only group after 8 h of incubation and remained stable thereafter, suggesting distinct fermentation dynamics when peptides are combined with carbohydrates. Peptide profiling revealed several novel sequences (ILSPL, IQLP, IAANPA, FASPA, IATSPA, and IIP), with most characterized by a proline residue at the penultimate position and isoleucine at the N-terminus
More direct evidence of prebiotic effects comes from in vivo studies. In a high-fat diet (HFD) mouse model, lupin protein hydrolysate supplementation for 12 weeks reduced weight gain, improved adipose function, and counteracted dyslipidemia and insulin resistance [54]. These effects were associated with microbiota modulation: LPH restored the abundances of specific taxa (Patescibacteria, Thermodesulfobacteriota) and increased beneficial Verrucomicrobiota, notably Akkermansia, whose abundance, reduced under HFD, was fully recovered. Akkermansia muciniphila is a key species linked to improved metabolic health and healthy balanced gut biodiversity.
5. Regulatory Hurdles: Peptides as Novel Foods
Despite the increasing interest in BAPs for their functional and health-promoting properties, their incorporation into functional foods, nutraceuticals, or supplements is subject to stringent regulatory scrutiny. European Union (EU), United States (US), Japan and China have established frameworks to ensure the safety and proper labeling of such ingredients before market authorization. This section outlines the key regulatory requirements imposed by the European Food Safety Authority (EFSA), US Food and Drug Administration (FDA), the Food Safety Commission of Japan (FSCJ), and China Food and Drug Administration (CFDA) for peptides intended as novel food ingredients or dietary supplements. Despite procedural differences, the EFSA, FDA, FSCJ, and CFDA emphasize comprehensive safety assessments, proper characterization of peptide ingredients, and substantiation of health-related claims [86,87,88]. Figure 5 summarizes the core regulatory requirements for peptide-based novel foods in the EU, US, Japan, and China.
This comparison highlights the regulatory hurdles that must be overcome before BAPs can be successfully integrated into food systems or marketed as dietary supplements. While regulatory alignment between jurisdictions is improving, scientific substantiation of safety and efficacy remains the cornerstone of market authorization for BAPs.
5.1. European Regulatory Framework
In the EU, BAPs are regulated under Regulation (EU) 2015/2283 on novel foods, which governs food ingredients not consumed to a significant extent in the EU before 15 May 1997. Additional requirements apply under Regulation (EC) 1924/2006 on nutrition and health claims made on foods if the peptide ingredient is also intended to make health claims [86,87,89,90].
EFSA requires a comprehensive safety assessment as part of the novel food dossier. This includes molecular characterization of the peptides—such as amino acid sequence, molecular weight distribution, degree of hydrolysis, and purity profile—along with a full description of the source material, enzymatic hydrolysis process, and downstream purification steps [90]. Applicants must also provide a detailed compositional analysis, including the peptide profile, residual proteins, potential contaminants (e.g., heavy metals, microbial toxins), and batch-to-batch consistency. Stability data during storage and within the intended food matrix must demonstrate that the peptide retains its integrity and function under real-world conditions [90].
The application is required to define the target food categories, the intended use levels, and the estimated dietary exposure, with these definitions to be based on population intake data. EFSA's customary approach involves the evaluation of both mean and high-percentile (95th) exposure scenarios [90].
Safety assessments typically involve in vitro and in vivo toxicology studies, including genotoxicity, subchronic (90-day) toxicity, and, if required, reproductive or developmental studies. Human clinical data, while not mandatory, are often beneficial to support safety and efficacy [90,91]. EFSA also evaluates whether the peptide has any adverse nutritional implications when added to the diet. Given that many BAPs are derived from allergenic sources (e.g., milk, soy, fish), applicants must assess potential allergenicity using in silico sequence homology tools, in vitro digestibility tests, and, where appropriate, allergen-specific immunoassays [90,91].
If a health claim is proposed (e.g., blood pressure regulation or improved satiety), it must be separately substantiated with human intervention trials in accordance with Regulation (EC) 1924/2006 [89,91]. EFSA requires that these studies demonstrate a clear cause–effect relationship between peptide consumption and the claimed physiological benefit. Once EFSA issues a favorable scientific opinion, the European Commission can authorize the peptide for use as a novel food through an implementing act published in the EU Official Journal [92].
5.2. United States Regulatory Framework
In the US, regulatory oversight of food-derived peptides depends on their intended use. Peptides incorporated into conventional foods must either be generally recognized as safe (GRAS) under the Federal Food, Drug, and Cosmetic Act, approved as food additives, or subject to a New Dietary Ingredient (NDI) Notification if intended for use in dietary supplements under the Dietary Supplement Health and Education Act (DSHEA, 1994) [93,94].
GRAS notification is the most common regulatory route. Applicants must demonstrate that the peptide ingredient is safe under its intended conditions of use, based on publicly available scientific evidence and, in most cases, the consensus of qualified experts. The submission includes detailed information on identity, manufacturing process, and specifications, including the source, production (e.g., enzymatic hydrolysis), purification, and testing for contaminants or residual solvents [93]. Use levels across different food categories and corresponding estimated daily intake based on population data must be provided. Toxicological evaluations, similar to EFSA requirements, involve genotoxicity, subchronic, and—if warranted—chronic toxicity studies, along with assessments of allergenicity and absorption/metabolism data [93,95,96].
For dietary supplements, if the peptide ingredient was not marketed in the U.S. before October 15, 1994, it qualifies as a New Dietary Ingredient (NDI) and must undergo a premarket safety notification at least 75 days before marketing. The notification includes safety evidence, manufacturing details, and labeling information. Supplement formulations must also comply with Good Manufacturing Practices (GMP) outlined in 21 CFR Part 111 [93,95,96]. Labeling and claims are regulated under 21 CFR Part 101. While structure/function claims (e.g., "supports healthy blood pressure") are permitted with a mandatory disclaimer, health claims linking the peptide to disease risk reduction require FDA pre-approval based on significant scientific agreement [93,95,96].
5.3. Japanese Regulatory Framework
In Japan, oversight of BAPs in foods is provided by the Food Safety Commission of Japan (FSCJ) in collaboration with the Ministry of Health, Labour and Welfare (MHLW) and the Consumer Affairs Agency (CAA). Japan has a dual regulatory pathway for functional foods. The Foods for Specified Health Uses (FOSHU) system grants approval to individual products containing functional ingredients proven to regulate or support physiological functions, such as blood pressure or gastrointestinal health. It requires extensive premarket evaluation, including toxicological data, human clinical trials on Japanese subjects, and detailed compositional and stability information, before authorization is granted for health claims and often require publications in local journals. This process is rigorous and time-intensive, often exceeding one year. Once approved, the product can display the official FOSHU mark, which helps consumers identify products that have passed the rigorous government evaluation [97,98,99].
In contrast, the Foods with Function Claims (FFC) pathway, introduced in 2015, allows companies to notify the CAA of scientific evidence supporting a functional claim at least 60 days before marketing, without undergoing the same level of governmental review as FOSHU. For both systems, manufacturers must provide compositional data, manufacturing details, and safety assessments consistent with FSCJ guidance, emphasizing consumer safety and scientific substantiation [98].
5.4. Chinese Regulatory Framework
China has also established a dynamic regulatory framework for foods with health benefits, including BAPs and protein hydrolysates, under the authority of the China Food and Drug Administration (CFDA) and guided by the Food Safety Law (FSL) of 2015 [97,98]. BAPs are typically considered within the health food (HE) category, which includes dietary supplements and functional foods that are intended to regulate human body functions without treating diseases. Key hurdles include classification as “special foods,” stringent requirements for human clinical evidence to substantiate health claims, and mandatory testing conducted by CFDA-recognized agencies. These tests cover toxicological safety (acute, subchronic, and chronic toxicity studies often lasting up to 330 days), efficacy screening, hygiene standards, stability testing, and detailed production process documentation and also site visits to overseas production facilities to inspect processes and take samples Approval confers the right to use the “Blue Hat” logo, a symbol of CFDA-certified health foods [97,98,99]. However, the framework is described as being in flux, with evolving policies and guidelines that require continuous monitoring. Overall, the combination of mandatory long-term testing, dossier preparation, and regulatory uncertainty constitutes a major barrier to market entry for novel peptide-based foods in China.
6. Challenges Limiting the Commercial Development of BAPs
Despite the increasing recognition of food-derived BAPs as functional components with potential health-promoting effects, their commercial development faces substantial scientific, technological, and regulatory challenges. These limitations hinder both industrial scalability and the translation of promising laboratory findings into consumer-ready products [100].
One of the primary technical obstacles lies in the preparation and purification processes. Conventional extraction and purification methods, such as solvent extraction, ultrafiltration, and chromatography, are often labor-intensive, time-consuming, and result in low peptide yields. High-purity peptides (≥99%) are frequently targeted in commercial applications, especially for therapeutic or high-value nutraceutical products; however, achieving such purity substantially increases production costs. Moreover, ultra-purification may remove other bioactive constituents that contribute synergistically to the desired health effects [88,101].
Membrane filtration systems, although scalable, are prone to operational issues such as pore blockage, reduced selectivity, and poor reproducibility across batches. Chromatographic techniques, while precise, are economically prohibitive for industrial use—accounting for up to 70% of total capital and operational expenditures in peptide bioprocessing—thus limiting their feasibility beyond laboratory-scale production [101,102].
Processing conditions further influence the integrity and efficacy of BAPs. High-temperature treatments used to denature protein precursors can negatively affect peptide profiles, leading to chemical modifications such as racemization, cyclization, and the formation of advanced glycation end-products (AGEs) through Maillard reactions. These alterations not only reduce bioactivity but can also result in undesirable sensory changes and the formation of potentially toxic intermediates.
Organoleptic properties also present a barrier to consumer acceptance. Many peptide hydrolysates exhibit a bitter taste, attributed to hydrophobic amino acid sequences, which limits their applicability in food and nutraceutical formulations [103]. Processing strategies, especially enzyme selection, use of masking agents or sequential hydrolysis, can help in reducing bitterness and improve sensory qualities, supporting broader use in food products. However, these techniques frequently incur substantial expenses and may inadvertently diminish the peptide's functional activity through structural modification or partial degradation [104,105].
Another considerable challenge is the low bioavailability of BAPs. Although many peptides demonstrate potent biological activities in vitro, these effects often fail to replicate in vivo due to enzymatic degradation during gastrointestinal transit. BAPs demonstrate a high degree of susceptibility to hydrolysis by proteolytic enzymes, including pepsin, trypsin, chymotrypsin, brush border peptidases and serum proteases. This process results in the generation of inactive fragments and a significant reduction in systemic availability. [101,106]. Additional factors influencing poor bioavailability include high molecular weight, the presence of hydrophobic or basic amino acids at the C-terminal end (e.g., lysine, arginine), and inherently low intestinal permeability—all of which collectively impair absorption and subsequent bioactivity [101].
Furthermore, BAPs are commonly obtained as complex peptide mixtures, making it difficult to ensure batch-to-batch consistency and accurate characterization. Unlike single-entity pharmaceutical compounds, these mixtures challenge standardization, stability testing, and quality assurance protocols, which are essential for regulatory compliance and product reproducibility
Lastly, the lack of robust clinical trial data remains a critical limitation. Although numerous in vitro and animal model studies provide preliminary evidence of health benefits, these findings cannot reliably predict human outcomes due to differences in absorption, distribution, metabolism, and excretion (ADME) pathways [101]. Regulatory approval for functional foods or health claims typically necessitates comprehensive human intervention studies, including clear identification of active doses, biomarkers, and long-term safety data. Main regulatory organs rejections of health claims for BAPs typically cite insufficient evidence and methodological shortcomings in the submitted dossiers.
7. Concluding Remarks
This review provided an overview of recent advances in the production and characterization of BAPs derived from pulses and pseudocereals, with particular emphasis on bio-based approaches such as enzymatic hydrolysis and microbial fermentation. These high-quality plant proteins are a sustainable raw material for producing multifunctional peptides with antioxidant, antihypertensive, antidiabetic, antimicrobial and prebiotic properties. In this context, these bio-based approaches transform affordable, protein-rich crops into added-value functional ingredients for the food and nutraceutical industries.
By including studies published over the last years, this review highlights both the technological strategies and biological evidence supporting the potential of pulse- and pseudocereal-derived peptides. Enzymatic hydrolysis remains the most reproducible method for targeted peptide release, while microbial fermentation responds to consumer demands for natural and “clean-label” products, offering additional nutritional and sensory benefits. These approaches are increasingly complemented by in silico prediction tools, which accelerate peptide discovery and optimization in BAPs production.
Despite these promising advances, challenges remain regarding process standardization, large-scale production, and regulatory validation. Further studies are needed to deepen understanding of structure–activity relationships and to assess long-term health effects through clinical trials.
The integration of BAPs into the global food and nutraceutical markets is promising but is heavily dependent on regulatory clearance. EU, US, Japan, and China all require rigorous demonstration of safety, quality, and efficacy, necessitating advanced analytical, toxicological, and clinical data. While these regulatory frameworks present certain challenges, they also ensure consumer protection and product credibility. Aligning research and development practices with these regulatory expectations not only accelerates approval timelines but also fosters trust among consumers and health professionals. In order to successfully commercialize BAPs, harmonization of global regulatory requirements and investment in standardized methodologies for the evaluation of the potential health benefits will be critical to the successful commercialization of BAPs.
Overall, the valorization of pulses and pseudocereals as sustainable protein sources for BAPs production not only expands opportunities for developing next-generation functional foods and nutraceuticals but also contributes to global strategies promoting sustainable diets and health-oriented innovation.
Collectively, these multifaceted challenges underscore the necessity for integrated approaches that combine advanced processing technologies, in vivo validation strategies, and regulatory harmonization. Such an approach is required to facilitate the translation of BAPs from bench to market.
Author Contributions
Conceptualization, M.M., V.D., Y.J.; writing—original draft preparation, M.M., V.D. and Y.J.; writing—review and editing, M.M., V.D. and Y.J.; supervision, J.D.C. and M.B.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Acknowledgments
M. Martoccia holds a PhD career grant supported by Next Generation EU-MUR (CUP C13C22000480001).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ACE | Angiotensin-I-converting enzyme |
| ADMET | Absorption, Distribution, Metabolism, Excretion, and Toxicity |
| BAP | Bioactive peptide |
| BP | Blood pressure |
| BW | Body weight |
| Cgh | Hydrolyzed γ-conglutin |
| DPP-IV | Dipeptidyl peptidase-IV |
| E:S | Enzyme-substrate ratio |
| EAA | Essential amino acid |
| EAE | Experimental autoimmune encephalomyelitis |
| HFD | High-fat diet mice |
| IC₅₀ | Half-maximal Inhibitory Concentration |
| ICAM-1 | Intercellular Adhesion Molecule 1 |
| IL-1β | Interleukin 1β |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| LAB | Lactic acid bacteria |
| LPS | Lipopolysaccharide |
| MD | Molecular dynamics |
| MM-GBSA | Molecular mechanics generalized Born surface area |
| MM-PBSA | Molecular mechanics Poisson–Boltzmann surface area |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| QSAR | Quantitative structure–activity relationship |
| RMSD | Root-mean-square deviation |
| SHR | Spontaneously hypertensive rats |
| SOD | Superoxide dismutase |
| TE | Tolox equivalents |
| TNF-α | Tumor necrosis factor α |
| VCAM-1 | Vascular Cell Adhesion Molecule 1 |
References
- Boye, J.; Zare, F.; Pletch, A. Pulse Proteins: Processing, Characterization, Functional Properties and Applications in Food and Feed. Food Research International 2010, 43, 414–431. [Google Scholar] [CrossRef]
- Nosworthy, M.G.; Yu, B.; Zaharia, L.I.; Medina, G.; Patterson, N. Pulse Protein Quality and Derived Bioactive Peptides. Front. Plant Sci. 2025, 16, 1429225. [Google Scholar] [CrossRef]
- Martín-Cabrejas, M.A. Legumes: An Overview.
- Semba, R.D.; Ramsing, R.; Rahman, N.; Kraemer, K.; Bloem, M.W. Legumes as a Sustainable Source of Protein in Human Diets. Global Food Security 2021, 28, 100520. [Google Scholar] [CrossRef]
- Hyun, J.; Kang, S.-I.; Lee, S.-W.; Amarasiri, R.P.G.S.K.; Nagahawatta, D.P.; Roh, Y.; Wang, L.; Ryu, B.; Jeon, Y.-J. Exploring the Potential of Olive Flounder Processing By-Products as a Source of Functional Ingredients for Muscle Enhancement. Antioxidants 2023, 12, 1755. [Google Scholar] [CrossRef]
- Usman, M.; Patil, P.J.; Mehmood, A.; Rehman, A.; Shah, H.; Haider, J.; Xu, K.; Zhang, C.; Li, X. Comparative Evaluation of Pseudocereal Peptides: A Review of Their Nutritional Contribution. Trends in Food Science & Technology 2022, 122, 287–313. [Google Scholar] [CrossRef]
- Kaur, H.; Shams, R.; Dash, K.K.; Dar, A.H. A Comprehensive Review of Pseudo-Cereals: Nutritional Profile, Phytochemicals Constituents and Potential Health Promoting Benefits. Applied Food Research 2023, 3, 100351. [Google Scholar] [CrossRef]
- Carbonaro, M.; Maselli, P.; Nucara, A. Structural Aspects of Legume Proteins and Nutraceutical Properties. Food Research International 2015, 76, 19–30. [Google Scholar] [CrossRef]
- Usman, M.; Patil, P.J.; Mehmood, A.; Rehman, A.; Shah, H.; Haider, J.; Xu, K.; Zhang, C.; Li, X. Comparative Evaluation of Pseudocereal Peptides: A Review of Their Nutritional Contribution. Trends in Food Science & Technology 2022, 122, 287–313. [Google Scholar] [CrossRef]
- Morales, D.; Miguel, M.; Garcés-Rimón, M. Pseudocereals: A Novel Source of Biologically Active Peptides. Critical Reviews in Food Science and Nutrition 2021, 61, 1537–1544. [Google Scholar] [CrossRef]
- Capraro, J.; Benedetti, S.D.; Heinzl, G.C.; Scarafoni, A.; Magni, C. Bioactivities of Pseudocereal Fractionated Seed Proteins and Derived Peptides Relevant for Maintaining Human Well-Being. International Journal of Molecular Sciences 2021, 22, 3543. [Google Scholar] [CrossRef]
- Hu, K.; Huang, H.; Li, H.; Wei, Y.; Yao, C. Legume-Derived Bioactive Peptides in Type 2 Diabetes: Opportunities and Challenges. Nutrients 2023, 15, 1096. [Google Scholar] [CrossRef]
- Chang, S.K.C.; Zhang, Y.; Pechan, T. Structures, Antioxidant, and Angiotensin I-Converting Enzyme (ACE)-Inhibitory Activities of Peptides Derived from Protein Hydrolysates of Three Phenolics-Rich Legume Genera. Journal of Food Science 2025, 90, e70069. [Google Scholar] [CrossRef] [PubMed]
- Fonseca Hernandez, D.; Mojica, L.; Gonzalez de Mejia, E. Legume-Derived Bioactive Peptides: Role in Cardiovascular Disease Prevention and Control. Current Opinion in Food Science 2024, 56, 101132. [Google Scholar] [CrossRef]
- Tawalbeh, D.; Al-U’datt, M.H.; Wan Ahmad, W.A.N.; Ahmad, F.; Sarbon, N.M. Recent Advances in In Vitro and In Vivo Studies of Antioxidant, ACE-Inhibitory and Anti-Inflammatory Peptides from Legume Protein Hydrolysates. Molecules 2023, 28, 2423. [Google Scholar] [CrossRef]
- Aderinola, T.A.; Duodu, K.G. Production, Health-Promoting Properties and Characterization of Bioactive Peptides from Cereal and Legume Grains. BioFactors 2022, 48, 972–992. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Liu, H.; Zhang, Y.; Zhang, S.; Liu, T.; Wang, D. Review on Plant-Derived Bioactive Peptides: Biological Activities, Mechanism of Action and Utilizations in Food Development. Journal of Future Foods 2022, 2, 143–159. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Smith, B.; Altintas, Z. Bioactive and Health-Promoting Properties of Enzymatic Hydrolysates of Legume Proteins: A Review. Critical Reviews in Food Science and Nutrition 2024, 64, 2548–2578. [Google Scholar] [CrossRef]
- Luzardo-Ocampo, I.; Gonzalez de Mejia, E. Plant Proteins and Peptides as Key Contributors to Good Health: A Focus on Pulses. Food Research International 2025, 211, 116346. [Google Scholar] [CrossRef]
- de Carvalho Oliveira, L.; Martinez-Villaluenga, C.; Frias, J.; Elena Cartea, M.; Francisco, M.; Cristianini, M.; Peñas, E. High Pressure-Assisted Enzymatic Hydrolysis Potentiates the Production of Quinoa Protein Hydrolysates with Antioxidant and ACE-Inhibitory Activities. Food Chemistry 2024, 447, 138887. [Google Scholar] [CrossRef]
- Rezvankhah, A.; Yarmand, M.S.; Ghanbarzadeh, B.; Mirzaee, H. Development of Lentil Peptides with Potent Antioxidant, Antihypertensive, and Antidiabetic Activities along with Umami Taste. Food Science & Nutrition 2023, 11, 2974–2989. [Google Scholar] [CrossRef]
- Song, H.; Wang, Q.; Shao, Z.; Wang, X.; Cao, H.; Huang, K.; Sun, Q.; Sun, Z.; Guan, X. In Vitro Gastrointestinal Digestion of Buckwheat ( Fagopyrum Esculentum Moench) Protein: Release and Structural Characteristics of Novel Bioactive Peptides Stimulating Gut Cholecystokinin Secretion. Food & Function 2023, 14, 7469–7477. [Google Scholar] [CrossRef]
- Wang, J.; Yang, T.; Yu, W.; Zhou, J.; Bian, Y.; Li, S.; Chen, Y. Pea Power against Microbes: Elucidating the Characteristics and Antimicrobial Mechanisms of Antimicrobial Peptides Obtained through Enzymatic Hydrolysis of Pea Protein Isolate. Food Bioscience 2025, 65, 106075. [Google Scholar] [CrossRef]
- Du, Z.; Comer, J.; Li, Y. Bioinformatics Approaches to Discovering Food-Derived Bioactive Peptides: Reviews and Perspectives. TrAC Trends in Analytical Chemistry 2023, 162, 117051. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; Cermeño, M.; Khalesi, M.; Kleekayai, T.; Amigo-Benavent, M. Application of in Silico Approaches for the Generation of Milk Protein-Derived Bioactive Peptides. Journal of Functional Foods 2020, 64, 103636. [Google Scholar] [CrossRef]
- Bo, W.; Chen, L.; Qin, D.; Geng, S.; Li, J.; Mei, H.; Li, B.; Liang, G. Application of Quantitative Structure-Activity Relationship to Food-Derived Peptides: Methods, Situations, Challenges and Prospects. Trends in Food Science & Technology 2021, 114, 176–188. [Google Scholar] [CrossRef]
- Vidal-Limon, A.; Aguilar-Toalá, J.E.; Liceaga, A.M. Integration of Molecular Docking Analysis and Molecular Dynamics Simulations for Studying Food Proteins and Bioactive Peptides. J. Agric. Food Chem. 2022, 70, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Brandes, N.; Ofer, D.; Peleg, Y.; Rappoport, N.; Linial, M. ProteinBERT: A Universal Deep-Learning Model of Protein Sequence and Function. Bioinformatics 2022, 38, 2102–2110. [Google Scholar] [CrossRef]
- Peredo-Lovillo, A.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Romero-Luna, H.E. Conventional and in Silico Approaches to Select Promising Food-Derived Bioactive Peptides: A Review. Food Chemistry: X 2022, 13, 100183. [Google Scholar] [CrossRef]
- Chen, D.; Shen, X.; Chen, W.; Wu, D.; Zhang, Z.; Liu, P.; Liu, Y.; Li, W.; Yang, Y. A Screening Strategy for Bioactive Peptides from Enzymolysis Extracts of Lentinula Edodes Based on Molecular Docking and Molecular Dynamics Simulation. Journal of Future Foods 2025, 5, 388–397. [Google Scholar] [CrossRef]
- Peres Fabbri, L.; Cavallero, A.; Vidotto, F.; Gabriele, M. Bioactive Peptides from Fermented Foods: Production Approaches, Sources, and Potential Health Benefits. Foods 2024, 13, 3369. [Google Scholar] [CrossRef]
- Ajayi, F.F.; Mudgil, P.; Jobe, A.; Antony, P.; Vijayan, R.; Gan, C.-Y.; Maqsood, S. Novel Plant-Protein (Quinoa) Derived Bioactive Peptides with Potential Anti-Hypercholesterolemic Activities: Identification, Characterization and Molecular Docking of Bioactive Peptides. Foods 2023, 12, 1327. [Google Scholar] [CrossRef]
- Mora-Melgem, J.A.; Arámburo-Gálvez, J.G.; Cárdenas-Torres, F.I.; Gonzalez-Santamaria, J.; Ramírez-Torres, G.I.; Arvizu-Flores, A.A.; Figueroa-Salcido, O.G.; Ontiveros, N. Dipeptidyl Peptidase IV Inhibitory Peptides from Chickpea Proteins (Cicer Arietinum L.): Pharmacokinetics, Molecular Interactions, and Multi-Bioactivities. Pharmaceuticals 2023, 16, 1109. [Google Scholar] [CrossRef]
- Quintero-Soto, M.F.; Chávez-Ontiveros, J.; Garzón-Tiznado, J.A.; Salazar-Salas, N.Y.; Pineda-Hidalgo, K.V.; Delgado-Vargas, F.; López-Valenzuela, J.A. Characterization of Peptides with Antioxidant Activity and Antidiabetic Potential Obtained from Chickpea (Cicer Arietinum L.) Protein Hydrolyzates. Journal of Food Science 2021, 86, 2962–2977. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Baraniak, B. Activities and Sequences of the Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptides Obtained from the Digested Lentil (Lens Culinaris) Globulins. International Journal of Food Science & Technology 2013, 48, 2363–2369. [Google Scholar] [CrossRef]
- Garcia-Mora, P.; Peñas, E.; Frias, J.; Gomez, R.; Martinez-Villaluenga, C. High-Pressure Improves Enzymatic Proteolysis and the Release of Peptides with Angiotensin I Converting Enzyme Inhibitory and Antioxidant Activities from Lentil Proteins. Food Chemistry 2015, 171, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Bollati, C.; Xu, R.; Boschin, G.; Bartolomei, M.; Rivardo, F.; Li, J.; Arnoldi, A.; Lammi, C. Integrated Evaluation of the Multifunctional DPP-IV and ACE Inhibitory Effect of Soybean and Pea Protein Hydrolysates. Nutrients 2022, 14, 2379. [Google Scholar] [CrossRef] [PubMed]
- Asledottir, T.; Vegarud, G.E.; Picariello, G.; Mamone, G.; Lea, T.E.; Røseth, A.; Ferranti, P.; Devold, T.G. Bioactive Peptides Identified in Pea and Faba Bean after in Vitro Digestion with Human Gastrointestinal Enzymes. Journal of Functional Foods 2023, 102, 105445. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Miralles, B.; Carrillo, W.; Hernández-Ledesma, B. In Vitro Chemopreventive Properties of Peptides Released from Quinoa (Chenopodium Quinoa Willd.) Protein under Simulated Gastrointestinal Digestion. Food Research International 2018, 105, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, S.; Moslehishad, M.; Salami, M. Antioxidant and Alpha-Glucosidase Enzyme Inhibitory Properties of Hydrolyzed Protein and Bioactive Peptides of Quinoa. International Journal of Biological Macromolecules 2022, 213, 602–609. [Google Scholar] [CrossRef]
- Ayala-Niño, A.; Rodríguez-Serrano, G.M.; González-Olivares, L.G.; Contreras-López, E.; Regal-López, P.; Cepeda-Saez, A. Sequence Identification of Bioactive Peptides from Amaranth Seed Proteins (Amaranthus Hypochondriacus Spp.). Molecules 2019, 24, 3033. [Google Scholar] [CrossRef]
- Lammi, C.; Arnoldi, A.; Aiello, G. Soybean Peptides Exert Multifunctional Bioactivity Modulating 3-Hydroxy-3-Methylglutaryl-CoA Reductase and Dipeptidyl Peptidase-IV Targets in Vitro. J. Agric. Food Chem. 2019, 67, 4824–4830. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Peptides Derived from Soy and Lupin Protein as Dipeptidyl-Peptidase IV Inhibitors: In Vitro Biochemical Screening and in Silico Molecular Modeling Study. J. Agric. Food Chem. 2016, 64, 9601–9606. [Google Scholar] [CrossRef] [PubMed]
- Boschin, G.; Scigliuolo, G.M.; Resta, D.; Arnoldi, A. ACE-Inhibitory Activity of Enzymatic Protein Hydrolysates from Lupin and Other Legumes. Food Chemistry 2014, 145, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Coscueta, E.R.; Campos, D.A.; Osório, H.; Nerli, B.B.; Pintado, M. Enzymatic Soy Protein Hydrolysis: A Tool for Biofunctional Food Ingredient Production. Food Chemistry: X 2019, 1, 100006. [Google Scholar] [CrossRef] [PubMed]
- Kiersnowska, K.; Jakubczyk, A. Bioactive Peptides Obtained from Legume Seeds as New Compounds in Metabolic Syndrome Prevention and Diet Therapy. Foods 2022, 11, 3300. [Google Scholar] [CrossRef]
- Karami, Z.; Muangnapoh, C.; Suthienkul, O.; Suriyarak, S.; Duangmal, K. Exploring Novel Peptides in Adzuki Bean and Mung Bean Hydrolysates with Potent Antibacterial Activity. Int J Food Sci Tech 2024, 59, 4829–4840. [Google Scholar] [CrossRef]
- Osman, A.; Enan, G.; Al-Mohammadi, A.-R.; Abdel-Shafi, S.; Abdel-Hameid, S.; Sitohy, M.Z.; El-Gazzar, N. Antibacterial Peptides Produced by Alcalase from Cowpea Seed Proteins. Antibiotics 2021, 10, 870. [Google Scholar] [CrossRef]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Jacinto-Hernández, C.; Alaiz, M.; Girón-Calle, J.; Vioque, J.; Dávila-Ortiz, G. Antioxidant and Metal Chelating Activities of Peptide Fractions from Phaseolin and Bean Protein Hydrolysates. Food Chemistry 2012, 135, 1789–1795. [Google Scholar] [CrossRef]
- Cruz-Chamorro, I.; Santos-Sánchez, G.; Bollati, C.; Bartolomei, M.; Capriotti, A.L.; Cerrato, A.; Laganà, A.; Pedroche, J.; Millán, F.; del Carmen Millán-Linares, M.; et al. Chemical and Biological Characterization of the DPP-IV Inhibitory Activity Exerted by Lupin (Lupinus Angustifolius) Peptides: From the Bench to the Bedside Investigation. Food Chemistry 2023, 426, 136458. [Google Scholar] [CrossRef]
- Muñoz, E.B.; Luna-Vital, D.A.; Fornasini, M.; Baldeón, M.E.; Gonzalez de Mejia, E. Gamma-Conglutin Peptides from Andean Lupin Legume (Lupinus Mutabilis Sweet) Enhanced Glucose Uptake and Reduced Gluconeogenesis in Vitro. Journal of Functional Foods 2018, 45, 339–347. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Villanueva, A.; Pedroche, J.; Millan, F.; Martin, M.E.; Millan-Linares, M.C. Antioxidant and Anti-Inflammatory Properties of Bioavailable Protein Hydrolysates from Lupin-Derived Agri-Waste. Biomolecules 2021, 11, 1458. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Chamorro, I.; Álvarez-López, A.I.; Santos-Sánchez, G.; Álvarez-Sánchez, N.; Pedroche, J.; Millán-Linares, M. del C.; Lardone, P.J.; Carrillo-Vico, A. A Lupin (Lupinus Angustifolius) Protein Hydrolysate Decreases the Severity of Experimental Autoimmune Encephalomyelitis: A Preliminary Study. International Journal of Molecular Sciences 2025, 26, 32. [Google Scholar] [CrossRef]
- Ponce-España, E.; Cruz-Chamorro, I.; Santos-Sánchez, G.; Álvarez-López, A.I.; Fernández-Santos, J.M.; Pedroche, J.; Millán-Linares, M.C.; Bejarano, I.; Lardone, P.J.; Carrillo-Vico, A. Anti-Obesogenic Effect of Lupin-Derived Protein Hydrolysate through Modulation of Adiposopathy, Insulin Resistance and Gut Dysbiosis in a Diet-Induced Obese Mouse. Biomedicine & Pharmacotherapy 2024, 178, 117198. [Google Scholar] [CrossRef]
- Wei, Y.; Liu, Y.; Li, Y.; Wang, X.; Zheng, Y.; Xu, J.; Sang, S.; Liu, Y. A Novel Antihypertensive Pentapeptide Identified in Quinoa Bran Globulin Hydrolysates: Purification, In Silico Characterization, Molecular Docking with ACE and Stability against Different Food-Processing Conditions. Nutrients 2022, 14, 2420. [Google Scholar] [CrossRef]
- de Carvalho Oliveira, L.; Martinez-Villaluenga, C.; Frias, J.; Elena Cartea, M.; Francisco, M.; Cristianini, M.; Peñas, E. High Pressure-Assisted Enzymatic Hydrolysis Potentiates the Production of Quinoa Protein Hydrolysates with Antioxidant and ACE-Inhibitory Activities. Food Chemistry 2024, 447, 138887. [Google Scholar] [CrossRef]
- Capraro, J.; Benedetti, S.D.; Heinzl, G.C.; Scarafoni, A.; Magni, C. Bioactivities of Pseudocereal Fractionated Seed Proteins and Derived Peptides Relevant for Maintaining Human Well-Being. International Journal of Molecular Sciences 2021, 22, 3543. [Google Scholar] [CrossRef]
- López-Moreno, M.; Jiménez-Moreno, E.; Márquez Gallego, A.; Vera Pasamontes, G.; Uranga Ocio, J.A.; Garcés-Rimón, M.; Miguel-Castro, M. Red Quinoa Hydrolysates with Antioxidant Properties Improve Cardiovascular Health in Spontaneously Hypertensive Rats. Antioxidants 2023, 12, 1291. [Google Scholar] [CrossRef]
- González-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Bioactive Peptides from Germinated Soybean with Anti-Diabetic Potential by Inhibition of Dipeptidyl Peptidase-IV, α-Amylase, and α-Glucosidase Enzymes. International Journal of Molecular Sciences 2018, 19, 2883. [Google Scholar] [CrossRef]
- Alauddin, M.; Amin, M.R.; Siddiquee, M.A.; Hiwatashi, K.; Shimakage, A.; Takahashi, S.; Shinbo, M.; Komai, M.; Shirakawa, H. In Silico and in Vivo Experiment of Soymilk Peptide (Tetrapeptide - FFYY) for the Treatment of Hypertension. Peptides 2024, 175, 171170. [Google Scholar] [CrossRef]
- Bhattacharjee, M.J.; Bala, A.; Khan, M.R.; Mukherjee, A.K. Functional Impact of Bioactive Peptides Derived from Fermented Foods on Diverse Human Populations. Food Chemistry 2025, 492, 145416. [Google Scholar] [CrossRef]
- Cruz-Casas, D.E.; Aguilar, C.N.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R.; Chávez-González, M.L.; Flores-Gallegos, A.C. Bioactive Protein Hydrolysates Obtained from Amaranth by Fermentation with Lactic Acid Bacteria and Bacillus Species. Heliyon 2023, 9, e13491. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; Gobbetti, M. Synthesis of the Cancer Preventive Peptide Lunasin by Lactic Acid Bacteria During Sourdough Fermentation. Nutrition and Cancer 2012, 64, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, T. Effect of Solid-State Fermentation with Bacillus Subtilis Lwo on the Proteolysis and the Antioxidative Properties of Chickpeas. International Journal of Food Microbiology 2021, 338, 108988. [Google Scholar] [CrossRef] [PubMed]
- Chiacchio, M.F.; Tagliamonte, S.; Pazzanese, A.; Vitaglione, P.; Blaiotta, G. Lactic Acid Fermentation Improves Nutritional and Functional Properties of Chickpea Flours. Food Research International 2025, 203, 115899. [Google Scholar] [CrossRef]
- Xu, Y.; Che, J.; Wang, Y.; Xiao, B.; Wei, L.; Rong, L.; Li, R. ACE Inhibitory Peptides and Flavor Compounds from Se-Enriched Bacillus Natto Fermented Chickpea. LWT 2025, 215, 117190. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Karaś, M.; Złotek, U.; Szymanowska, U. Identification of Potential Inhibitory Peptides of Enzymes Involved in the Metabolic Syndrome Obtained by Simulated Gastrointestinal Digestion of Fermented Bean (Phaseolus Vulgaris L.) Seeds. Food Research International 2017, 100, 489–496. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Karaś, M.; Baraniak, B.; Pietrzak, M. The Impact of Fermentation and in Vitro Digestion on Formation Angiotensin Converting Enzyme (ACE) Inhibitory Peptides from Pea Proteins. Food Chemistry 2013, 141, 3774–3780. [Google Scholar] [CrossRef]
- Tonini, S.; Tlais, A.Z.A.; Galli, B.D.; Helal, A.; Tagliazucchi, D.; Filannino, P.; Zannini, E.; Gobbetti, M.; Di Cagno, R. Lentils Protein Isolate as a Fermenting Substrate for the Production of Bioactive Peptides by Lactic Acid Bacteria and Neglected Yeast Species. Microbial Biotechnology 2024, 17, e14387. [Google Scholar] [CrossRef]
- Ayyash, M.; Johnson, S.K.; Liu, S.-Q.; Mesmari, N.; Dahmani, S.; Al Dhaheri, A.S.; Kizhakkayil, J. In Vitro Investigation of Bioactivities of Solid-State Fermented Lupin, Quinoa and Wheat Using Lactobacillus Spp. Food Chemistry 2019, 275, 50–58. Food Chemistry 2019, 275, 50–58. [Google Scholar] [CrossRef]
- Li, S.; Du, G.; Shi, J.; Zhang, L.; Yue, T.; Yuan, Y. Preparation of Antihypertensive Peptides from Quinoa via Fermentation with Lactobacillus Paracasei. eFood 2022, 3, e20. [Google Scholar] [CrossRef]
- Wang, P.; Ma, T. Production of Bioactive Peptides from Tartary Buckwheat by Solid-State Fermentation with Lactiplantibacillus Plantarum ATCC 14917. Foods 2024, 13, 3204. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Casas, D.E.; Ramos-González, R.; Prado-Barragán, L.A.; Iliná, A.; Aguilar, C.N.; Rodríguez-Herrera, R.; Tsopmo, A.; Flores-Gallegos, A.C. Protein Hydrolysates with ACE-I Inhibitory Activity from Amaranth Seeds Fermented with Enterococcus Faecium-LR9: Identification of Peptides and Molecular Docking. Food Chemistry 2025, 464, 141598. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Fan, X.; Wang, D.; Li, X.; Wang, X.; Yang, J.; Qiu, C.; Liu, X.; Pang, G.; Abra, R.; et al. Study on Preparation of Chickpea Peptide and Its Effect on Blood Glucose. Front. Nutr. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Daliri, E.B.-M.; Ofosu, F.K.; Chelliah, R.; Lee, B.H.; An, H.; Elahi, F.; Barathikannan, K.; Kim, J.-H.; Oh, D.-H. Influence of Fermented Soy Protein Consumption on Hypertension and Gut Microbial Modulation in Spontaneous Hypertensive Rats. Bioscience of Microbiota, Food and Health 2020, 39, 199–208. [Google Scholar] [CrossRef]
- Zhang, C.; Xia, S.; Zhang, Y.; Zhu, S.; Li, H.; Liu, X. Identification of Soybean Peptides and Their Effect on the Growth and Metabolism of Limosilactobacillus Reuteri LR08. Food Chemistry 2022, 369, 130923. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Le Maux, S.; Dubrulle, C.; Barre, C.; FitzGerald, R.J. Quinoa (Chenopodium Quinoa Willd.) Protein Hydrolysates with in Vitro Dipeptidyl Peptidase IV (DPP-IV) Inhibitory and Antioxidant Properties. Journal of Cereal Science 2015, 65, 112–118. [Google Scholar] [CrossRef]
- Pihlanto, A. Antioxidative Peptides Derived from Milk Proteins. International Dairy Journal 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Yang, M.; Yuan, P.; Xu, W.; Jiang, T.; Huang, J. Tartary Buckwheat Peptides Prevent Oxidative Damage in Differentiated SOL8 Cells via a Mitochondria-Mediated Apoptosis Pathway. Nutrients 2025, 17, 2204. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, X. Purification, Identification and Evaluation of Antioxidant Peptides from Pea Protein Hydrolysates. Molecules 2023, 28, 2952. [Google Scholar] [CrossRef]
- Ren, G.; Zhu, Y.; Shi, Z.; Li, J. Detection of Lunasin in Quinoa (Chenopodium Quinoa Willd.) and the in Vitro Evaluation of Its Antioxidant and Anti-Inflammatory Activities. J Sci Food Agric 2017, 97, 4110–4116. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Efficacy of Great Northern Beans-Derived Bioactive Compounds in Reducing Vascular Inflammation. Food Bioscience 2024, 57, 103524. [Google Scholar] [CrossRef]
- Kieffer, T.J.; McIntosh, C.H.; Pederson, R.A. Degradation of Glucose-Dependent Insulinotropic Polypeptide and Truncated Glucagon-like Peptide 1 in Vitro and in Vivo by Dipeptidyl Peptidase IV. Endocrinology 1995, 136, 3585–3596. [Google Scholar] [CrossRef] [PubMed]
- Tetteh, J.; Wereko Brobbey, D.-Y.; Osei, K.J.; Ayamah, A.; Laryea, M.K.; Darko, G.; Borquaye, L.S. Peptide Extract from Red Kidney Beans, Phaseolus Vulgaris (Fabaceae), Shows Promising Antimicrobial, Antibiofilm, and Quorum Sensing Inhibitory Effects. Biochemistry Research International 2024, 2024, 4667379. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat Rev Gastroenterol Hepatol 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Precup, G.; Marini, E.; Zakidou, P.; Beneventi, E.; Consuelo, C.; Fernández-Fraguas, C.; Garcia Ruiz, E.; Laganaro, M.; Magani, M.; Mech, A.; et al. Novel Foods, Food Enzymes, and Food Additives Derived from Food by-Products of Plant or Animal Origin: Principles and Overview of the EFSA Safety Assessment. Front. Nutr. 2024, 11. [Google Scholar] [CrossRef]
- Vettorazzi, A.; López de Cerain, A.; Sanz-Serrano, J.; Gil, A.G.; Azqueta, A. European Regulatory Framework and Safety Assessment of Food-Related Bioactive Compounds. Nutrients 2020, 12, 613. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Keskin Ulug, S.; Hong, H.; Wu, J. Regulatory Requirements of Bioactive Peptides (Protein Hydrolysates) from Food Proteins. Journal of Functional Foods 2019, 58, 123–129. [Google Scholar] [CrossRef]
- Regulation - 1924/2006 - EN - EUR-Lex. Available online: https://eur-lex.europa.eu/eli/reg/2006/1924/oj/eng (accessed on 5 August 2025).
- Regulation - 2015/2283 - EN - EUR-Lex. Available online: https://eur-lex.europa.eu/eli/reg/2015/2283/oj/eng (accessed on 5 August 2025).
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA); Turck, D. ; Bresson, J.-L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; et al. Scientific and Technical Guidance for the Preparation and Presentation of a Health Claim Application (Revision 2). EFSA Journal 2017, 15, e04680. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA) Products; Nutrition; (nda), A. ; Turck, D.; Bresson, J.-L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; et al. Guidance on the Preparation and Submission of an Application for Authorisation of a Novel Food in the Context of Regulation (EU) 2015/2283 (Revision 1)2. EFSA Journal 2021, 19, e06555. [Google Scholar] [CrossRef]
- Program, H.F. New Dietary Ingredient (NDI) Notification Process. Available online: https://www.fda.gov/food/dietary-supplements/new-dietary-ingredient-ndi-notification-process (accessed on 5 August 2025).
- Program, H.F. Generally Recognized as Safe (GRAS). Available online: https://www.fda.gov/food/food-ingredients-packaging/generally-recognized-safe-gras (accessed on 5 August 2025).
- 21 CFR Part 101 -- Food Labeling. Available online: https://www.ecfr.gov/current/title-21/part-101 (accessed on 5 August 2025).
- 21 CFR Part 111 -- Current Good Manufacturing Practice in Manufacturing, Packaging, Labeling, or Holding Operations for Dietary Supplements. Available online: https://www.ecfr.gov/current/title-21/part-111 (accessed on 5 August 2025).
- Hobbs, J.E.; Malla, S.; Sogah, E.K. Regulatory Frameworks for Functional Food and Supplements. Canadian Journal of Agricultural Economics/Revue canadienne d’agroeconomie 2014, 62, 569–594. [Google Scholar] [CrossRef]
- Patil, P.J.; Usman, M.; Zhang, C.; Mehmood, A.; Zhou, M.; Teng, C.; Li, X. An Updated Review on Food-Derived Bioactive Peptides: Focus on the Regulatory Requirements, Safety, and Bioavailability. Comprehensive Reviews in Food Science and Food Safety 2022, 21, 1732–1776. [Google Scholar] [CrossRef]
- Patel, D.; Dufour, Y.; Domigan, N. Functional Food and Nutraceutical Registration Processes in Japan and China: A Diffusion of Innovation Perspective. Journal of Pharmacy & Pharmaceutical Sciences 2008, 11, 1–11. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-Derived Bioactive Peptides in Human Health: Challenges and Opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Tao, X.; Zhang, W.; Chen, Y.; Yu, Q.; Xie, J. Food-Derived Bioactive Peptides: Production, Biological Activities, Opportunities and Challenges. Journal of Future Foods 2022, 2, 294–306. [Google Scholar] [CrossRef]
- Barati, M.; Javanmardi, F.; Mousavi Jazayeri, S.M.H.; Jabbari, M.; Rahmani, J.; Barati, F.; Nickho, H.; Davoodi, S.H.; Roshanravan, N.; Mousavi Khaneghah, A. Techniques, Perspectives, and Challenges of Bioactive Peptide Generation: A Comprehensive Systematic Review. Comprehensive Reviews in Food Science and Food Safety 2020, 19, 1488–1520. [Google Scholar] [CrossRef] [PubMed]
- Tagliamonte, S.; Oliviero, V.; Vitaglione, P. Food Bioactive Peptides: Functionality beyond Bitterness. Nutr Rev 2025, 83, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Rezvankhah, A.; Yarmand, M.S.; Ghanbarzadeh, B.; Mirzaee, H. Characterization of Bioactive Peptides Produced from Green Lentil (Lens Culinaris) Seed Protein Concentrate Using Alcalase and Flavourzyme in Single and Sequential Hydrolysis. Journal of Food Processing and Preservation 2021, 45, e15932. [Google Scholar] [CrossRef]
- Görgüç, A.; Gençdağ, E.; Yılmaz, F.M. Bioactive Peptides Derived from Plant Origin By-Products: Biological Activities and Techno-Functional Utilizations in Food Developments – A Review. Food Research International 2020, 136, 109504. [Google Scholar] [CrossRef]
- Hu, K.; Huang, H.; Li, H.; Wei, Y.; Yao, C. Legume-Derived Bioactive Peptides in Type 2 Diabetes: Opportunities and Challenges. Nutrients 2023, 15, 1096. [Google Scholar] [CrossRef]
Figure 1.
Bio-informatic approaches for BAPs prediction.
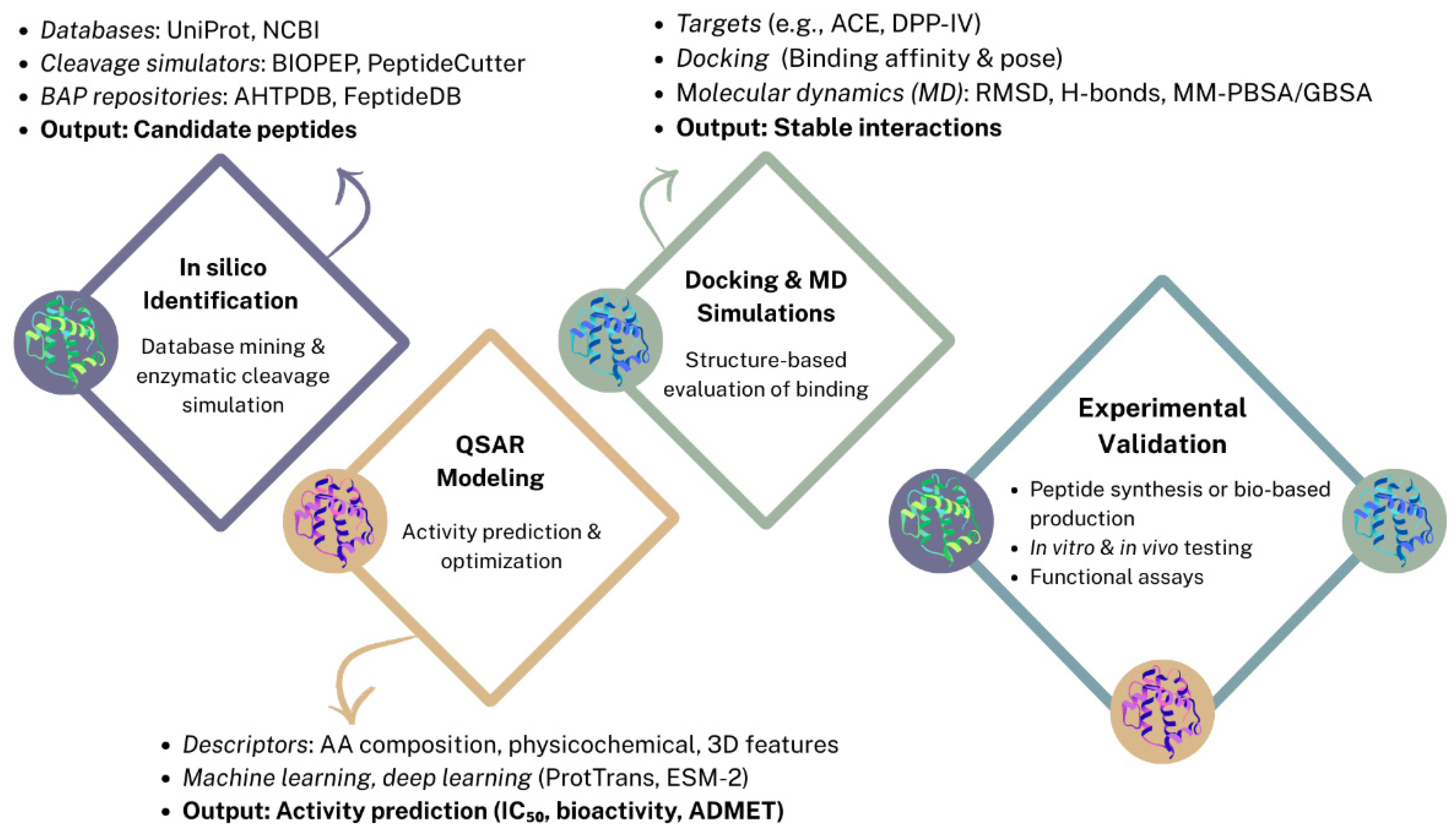
Figure 2.
Bio-based strategies for the production of BAPs.
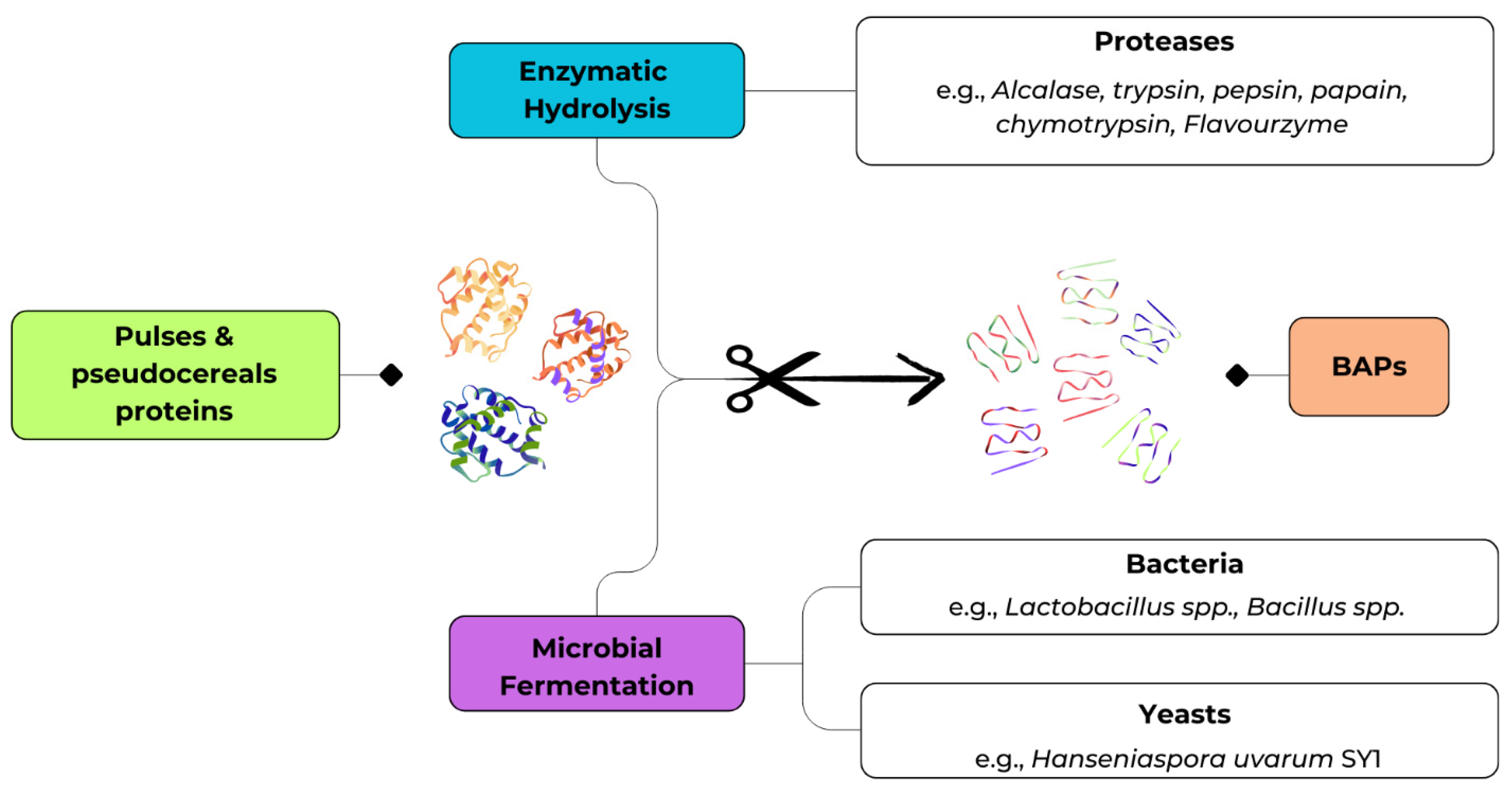
Figure 3.
Practical advantages and limitations of enzymatic hydrolysis vs. microbial fermentation for the production of BAPs from pulses and pseudocereals.
Figure 3.
Practical advantages and limitations of enzymatic hydrolysis vs. microbial fermentation for the production of BAPs from pulses and pseudocereals.
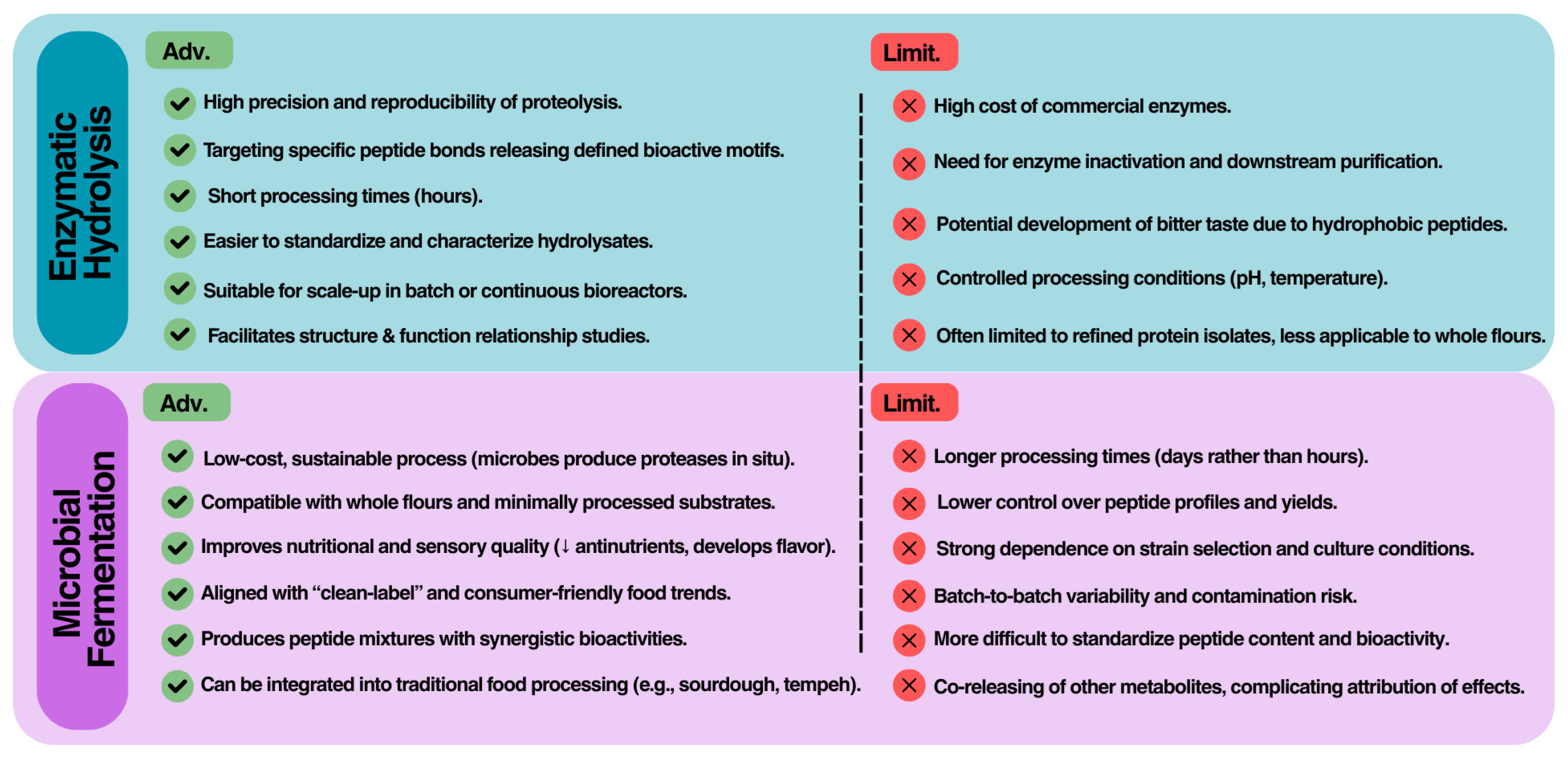
Figure 4.
Potential bioactivities from pulses and pseudocereals BAPs.
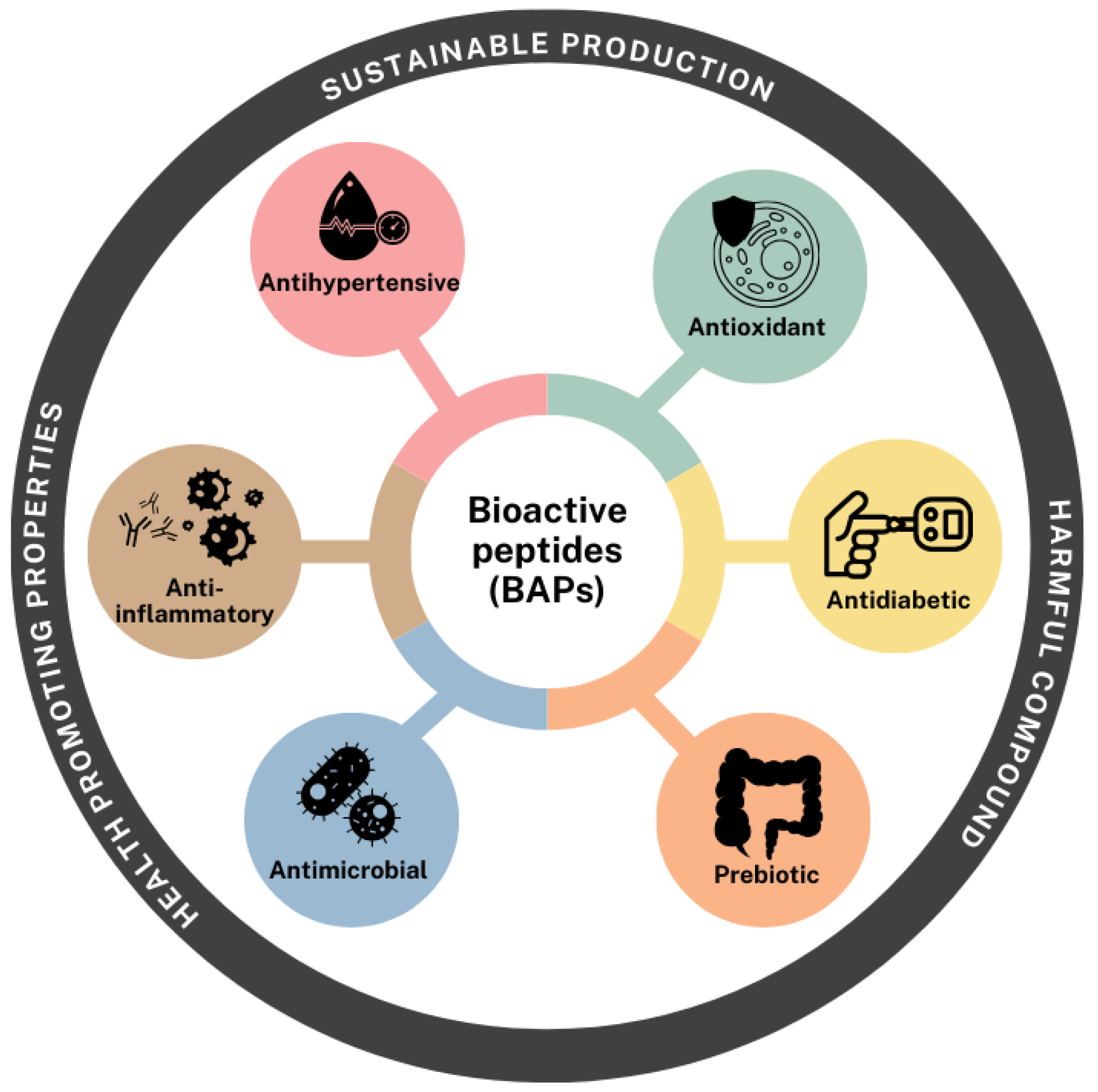
Figure 5.
Comparative overview of regulatory frameworks governing the approval of bioactive peptides as foods or health products in the European Union (EFSA), United States (FDA), Japan (FSCJ, MHLW, CAA), and China (CFDA/NMPA). The figure summarizes requirements for safety evaluation, product characterization, exposure assessment, labeling, health claims, and typical review timelines across jurisdictions.
Figure 5.
Comparative overview of regulatory frameworks governing the approval of bioactive peptides as foods or health products in the European Union (EFSA), United States (FDA), Japan (FSCJ, MHLW, CAA), and China (CFDA/NMPA). The figure summarizes requirements for safety evaluation, product characterization, exposure assessment, labeling, health claims, and typical review timelines across jurisdictions.
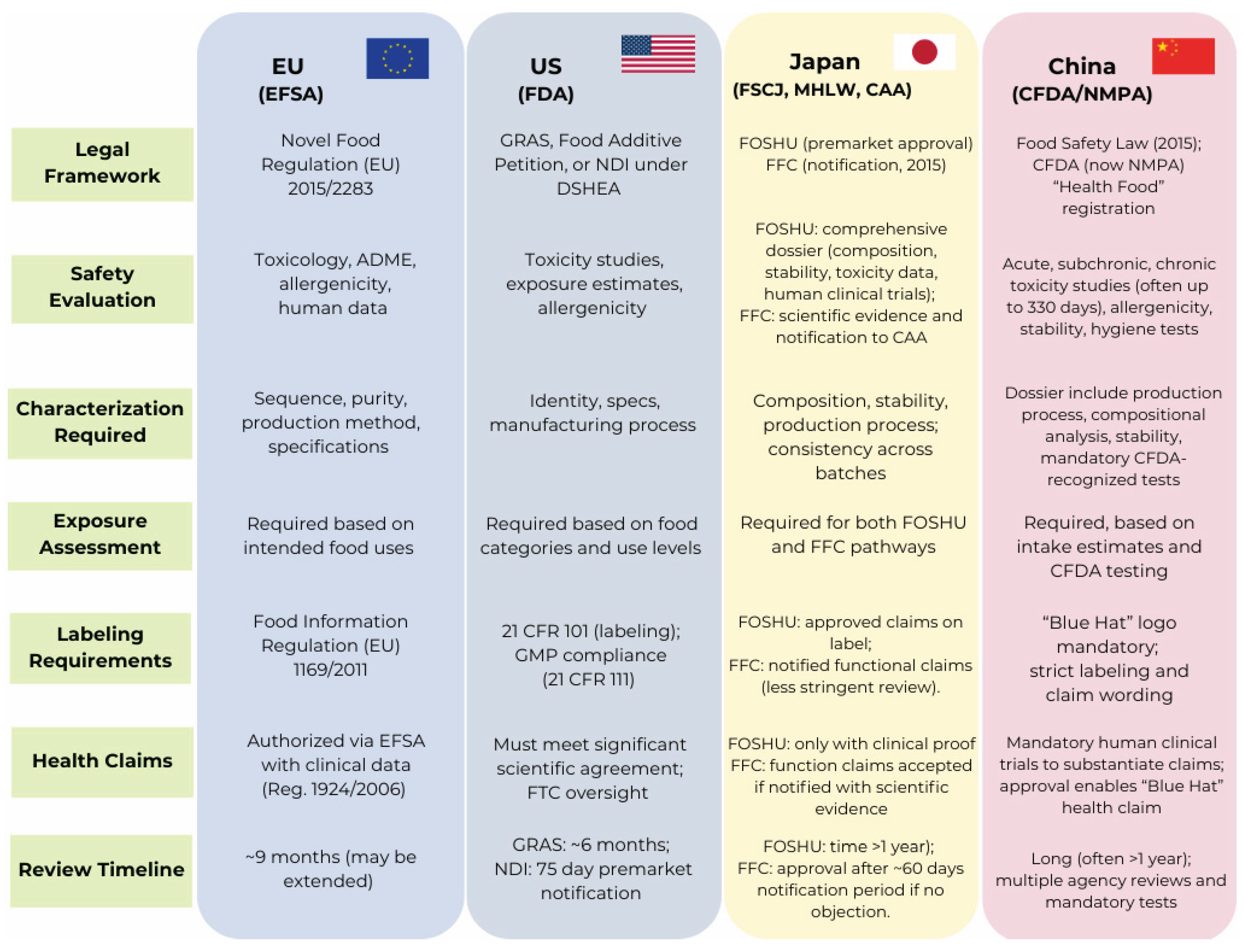
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



