Submitted:
20 September 2025
Posted:
22 September 2025
You are already at the latest version
Abstract
Major depressive disorder (MDD) is diagnosed twice as frequently in women compared to men. A disruption in brain circuits involved in emotion regulation may induce symptoms associated with MDD. Increasing evidence also indicates changes in activity within emotion regulation networks upon exposure to female gonadal hormones. Increased levels of stress reactivity in combination with low levels of protective female gonadal hormones may increase the risk for the development of MDD. GABAergic interneurons, some of which are ensheathed by perineuronal nets (PNNs), may facilitate alterations in the dynamics of neuronal networks in MDD. Microglia, which are the brain’s immune cells, are modulated by exposure to female gonadal hormones and glucocorticoids. Increased levels of stress reactivity, in combination with low levels of protective female gonadal hormones, may facilitate the aberrant modulation of GABAergic connections and the degradation of PNNs by microglia, leading to the disruption of emotion regulation circuits and the experience of psychiatric symptoms.
Keywords:
major depressive disorder
; microglia
; GABAergic system
; HPA axis
; female gonadal hormones
1. Introduction
According to the World Health Organization (WHO), major depressive disorder (MDD) is predicted to be the disorder associated with the largest disease burden by 2030 [1]. MDD also has significant economic impacts, increasing cases of workplace absenteeism, decreased productivity and unemployment [2]. By increasing the risk for death by suicide, cardiovascular diseases or stroke, depression is linked to a 20-fold increase in the risk of mortality, indicating the pressing need for practical treatment approaches [2].
The diagnosis of MDD requires the experience of at least one depressive episode[3]. According to the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5), a major depressive episode is defined by the presence of at least five symptoms within a period of 2 weeks. The core symptoms include either a depressed mood or anhedonia (loss of interest or pleasure) [4]. The secondary symptoms include changes in weight or appetite, sleeping difficulties, psychomotor retardation or agitation, loss of energy, reduced ability to think or concentrate, experiencing feelings of worthlessness or excessive guilt, and feelings of suicidality [4]. In addition to these emotional changes, evidence suggests that cognitive abilities are also disrupted in depression [5].
A variety of different mechanisms and theories have been proposed to account for the depressive symptomatology in MDD [3]. The development of depressive pathophysiology is frequently preceded by the experience of adverse life experiences associated with high levels of psychological stress, inducing disruptions in the physiological stress response [6,7]. Further, it was found that a mouse model of depression, caused by chronic stress exposure, presents increased circulation of proinflammatory cytokines together with functional and structural alterations of microglia, the brain’s resident immune cells, indicating a role for immune-related mechanisms in MDD [8,9,10]. Treatment approaches targeting mechanisms implicated in neuroplasticity, such as the rapid-acting antidepressant ketamine, also reduce symptoms in those diagnosed with treatment-resistant MDD [11]. Neuroplasticity refers to the brain’s ability to reorganize its neuronal connections through the remodeling of synaptic connections in response to external stimuli [11]. These findings indicate the modulatory effect of stimulating neuroplasticity on MDD [11].
Several pharmacological treatment approaches have shown promising results in targeting the aforementioned systems. For example, selective serotonin-reuptake inhibitors (SSRIs) lead to a reduction in symptoms as demonstrated by randomized control trials (RCT) and meta-analyses in patients with MDD [12]. Cell culture studies further reveal that SSRI administration induces a decrease in the release of proinflammatory cytokines, such as tumor necrosis factor alpha (TNF-α) and nitric oxide (NO), by microglia following an immune challenge, modeled using the inflammogen lipopolysaccharide (LPS) [13]. These findings indicate that SSRIs may exert their anti-inflammatory effects through the modulation of microglia functioning [12,13]. Further, the rapid-acting antidepressant ketamine exerts its antidepressant effects by facilitating neuroplasticity through an increased release of neurotrophic factors, such as brain-derived neurotrophic factor (BDNF) [14]. However, over half of all patients with MDD do not experience improvements despite receiving treatment with at least two different pharmacological approaches, therefore, meeting criteria for treatment-resistant depression [15].
The profile of symptoms expressed by patients with MDD is highly heterogeneous, varying significantly between patient groups [16]. Contributing to the unexplained variability in symptom representation, many clinical and preclinical studies in medical research only include male subjects or combine both sexes, covering up potential sex-specific effects [17]. The insufficient investigation and reporting of sex differences, by employing a research design or statistical testing that does not sufficiently reflect actual sex differences, or the reporting of sex differences only in some cases, may lead to a misrepresentation of sex differences [17]. MDD is diagnosed about twice as frequently in women compared to men [18]. Depression as experienced by men is predominantly reflected in externalizing behavioural symptoms such as increased levels of aggression or risk-taking behaviours, substance abuse, and increased levels of activity [19,20]. By contrast, depression in women is more frequently experienced in the form of mood changes, such as reduced interest and anhedonia, metabolic and physiological disruptions, including changes in weight and appetite, disruptions in sleep, and fatigue [21]. These findings indicate that different neurobiological mechanisms may underlie these differences in symptoms observed between sexes, prompting the need to investigate sex-specific mechanisms implicated in the pathophysiology of MDD [21].
To provide more targeted treatment approaches, a better understanding of sex-specific factors, such as monthly fluctuations across the menstrual cycle, and their influence on the mechanisms implicated in depression, is urgently needed. Therefore, in this review, we explore the impact of female gonadal hormone fluctuations and of specific female gonadal hormones on the emotion regulation networks at global network and microcircuit levels. Further, we highlight how gonadal hormone fluctuations and increases in stress reactivity in females could synergistically facilitate the development of psychiatric disorders. Microglia are critically implicated in the modulation of neuronal networks during development and in adulthood [22]. Further, the brain-resident immune cells are modulated by exposure to female gonadal hormones and glucocorticoids, the primary stress hormones in the body [23,24]. Therefore, the role of microglia, linking exposure to glucocorticoids, and withdrawal of female gonadal hormones and maladaptive restructuring of neuronal circuits, in the context of emotion regulation circuits, is specifically discussed. Finally, we present the conclusion that we draw from the summarized findings and present future perspectives outlining possibilities for closing the research gap concerning the potentially critical role of microglia in the modulation of emotion regulation circuits in MDD.
2. The Role of Emotion Regulation Networks in MDD
Using connectivity analyses following functional magnetic resonance imaging (fMRI), it was found that MDD is associated with disruptions in the neurocircuits implicated in emotion regulation [25,26]. Dysfunctional activation has been reported in a network consisting of limbic structures, including the amygdala, anterior cingulate cortex (ACC), hippocampus, hypothalamus, basal ganglia, and different components of the prefrontal cortex (PFC) [27]. Emotional regulation is facilitated through top-down control of the PFC over limbic structures [28]. The limbic system, the phylogenetically older brain region, is implicated in the generation of bottom-up, affective responses in response to external stimuli [29]. Evidence derived from studies employing functional and structural imaging indicates increased activity in bottom-up, emotion generating brain regions in depression. In contrast, inhibition by top-down prefrontal areas is disrupted [30,31].
2.1. Role of Limbic System Disruptions in MDD
In patients with MDD, the experience of psychosocial stress was found to be associated with increased amygdala activation, as assessed by fMRI [30]. The amygdala is considered the fear center of the brain, with lesions in this region inducing deficits in emotional processing and a loss of fear response in animal models [32]. Depressive symptomatology is associated with increased activity of limbic structures in response to negatively emotionally valenced stimuli. For example, results from fMRI studies indicate increased amygdala activity in response to negatively valenced stimuli, reflecting an attentional bias toward negatively valenced stimuli in depression [33]. During the encoding of novel information, increased amygdala response in depressed patients was also found to be coupled to increased hippocampal activity, potentially reflecting a bias toward negatively valenced information in the recall and formation of memories[34].
2.2. The Role of PFC Disruptions in Depression
Evidence indicates a critical role of the ventromedial (vm)PFC in emotional regulation through a downregulation of the amygdala. The vmPFC comprises parts of the ACC [35]. The vmPFC serves as a central hub in the brain’s emotion regulation network, integrating affective inputs from limbic and sensory regions, and higher-order cognitive information from PFC regions [35]. The ACC is also implicated in the regulation of the hypothalamic-pituitary-adrenal (HPA) axis, with findings indicating, for example, an association between a volumetric decrease in the rostral portion of the ACC and levels of salivary cortisol and early childhood abuse [36]. Glucocorticoids, secreted as the final step of HPA axis activation, bind to mineralocorticoid receptors (MRs) and glucocorticoid receptors (GRs) located in the ACC, thus facilitating a downregulation of HPA activity [37,38]. The modulation of HPA activity through these negative feedback mechanisms is facilitated by projections from the ACC to relay stations such as the bed nucleus of the stria terminals (BNST) and amygdala, projecting to corticotropin-releasing (CRH) in the hypothalamus[37,38].
Clinical findings suggest a role of the vmPFC in modulating the capacity for emotion regulation [39]. Employing fMRI, it was found that, participants after having experienced acute stress, as a result of exposure to the Trier Social Stress Test (TSST), displayed increased levels of activity in the vmPFC, in response to highly emotionally valenced images[39]. Further, findings revealed that increased levels of coupled functional connectivity between the amygdala and vmPFC after acute stress were associated with a smaller increase in levels of anxiety as a result of stress exposure[39]. Based on these findings, the authors concluded that the vmPFC was implicated in emotion regulation after stress exposure by modulating amygdala activity[39]. In addition, the ventrolateral (vl)PFC is involved in the modulation of emotional responses through its action on the vmPFC. In particular, the application of transcranial magnetic current stimulation (TMS) over the vlPFC reduced the experience of negative emotionality through a downregulation of vmPFC activity, thus facilitating downregulation of amygdala activity [40]. In line with these findings, reduced activity in the right inferior frontal gyrus (IFG), a sub-component of the vlPFC, was found in patients with MDD when they were instructed to downregulate their negative emotional reactions[41].
Moreover, evidence indicates that the dorsolateral (dl)PFC is critically implicated in the capacity for emotion regulation through its mutual connections with ventral PFC regions. The dlPFC is implicated in modulating attentional focus [42]. Studies further reveal a lateralization in function[42]. The left dlPFC is implicated in retaining specific task goals in working memory during cognitive tasks, enabling the focus on task-relevant information, while task irrelevant information are ignored[42]. In contrast, the right dlPFC is implicated in the redirecting of behavioural responses following the appearance of a distracting stimulus[42]. Through mutual connections between the vlPFC and the dlPFC, the dlPFC receives input from emotion generating regions, and facilitates the selection of behavioural responses appropriate to the stressor [43]. In the context of emotion regulation tasks, it was found that increased levels of functional coupling between vlPFC and dlPFC activity, through their modulatory effect on amygdala activity, are associated with a heightened capacity for cognitive reappraisal [43]. Cognitive reappraisal is described as an emotion regulation strategy including the re-interpretation of negatively valenced stimuli, resulting in the increased experience of positive emotions[43]. Based on their findings, the authors concluded that while the dlPFC maintains multiple options for response strategies in working memory, the vlPFC is implicated in the selection of the most appropriate response strategy[43]. Therefore, findings suggest a critical role for the dlPFC in emotion regulation networks through mutual interactions with regions such as the vlPFC and top-down modulation of amygdala activity.
3. The Influence of Fluctuations in Female Gonadal Hormones on Emotion Regulation Networks
3.1. Hormonal Fluctuations Along the Menstrual Cycle
Evidence indicates that exposure to gonadal hormones throughout the menstrual cycle influences the activity of key brain regions implicated in emotion regulation [44,45]. Hormonal fluctuations associated with the menstrual cycle facilitate the monthly ovulation process, which is characterized by the ovarian release of a follicle into the fallopian tube, where it can undergo fertilization [46]. Ovulation occurs due to a rapid increase in luteinizing hormone (LH) [47]. If fertilization does not occur, the corpus luteum forms from the ruptured unfertilized follicle, marking the beginning of the luteal phase[48]. The corpus luteum secretes high levels of progesterone [47]. Subsequently, a surge in both estrogen and progesterone levels occurs, reaching a peak in the mid-luteal phase, followed by a sudden decline in the late luteal phase, facilitating menstruation onset [47].
3.2. Influence of Hormonal Fluctuations Along the Menstrual Cycle on Hippocampal Structure and Function
Magnetic resonance imaging (MRI) and fMRI studies conducted in healthy women of reproductive age revealed alterations in hippocampal structural characteristics and functional connectivity throughout the menstrual cycle [44,49]. Specifically, findings revealed an increase in hippocampal volume in phases of the menstrual cycle associated with high levels of estrogen [44,49]. Women of reproductive age were exposed to structural and functional MRI, cognitive testing, and the collection of blood samples for the assessment of progesterone and estrogen during four sessions along their menstrual cycle. A bilateral increase in hippocampal volume was detected in the late follicular phase, characterized by heightened levels of estrogen, compared to the early follicular phase [49]. Functional brain activity was also assessed during the performance of cognitive tasks by exposing women of reproductive age to fMRI throughout their menstrual cycle. Findings revealed increased levels of hippocampal gray matter volume during the late follicular phase [49]. Heightened levels of gray matter volume in the hippocampus were positively correlated with estrogen levels during the late follicular phase[49] (Figure 1). Moreover, increased levels of estrogen during the late follicular phase were associated with increased connectivity of the hippocampus with prefrontal and temporal regions [49]. By contrast, in the late luteal phase, characterized by low levels of estrogen, hippocampal connectivity with prefrontal regions was reduced [49]. Additionally, it was found that this increased functional coupling between the hippocampus and prefrontal regions during high-estrogen phases of the menstrual cycle was associated with increased performance on tasks assessing memory functioning [44].
3.3. Influence of Hormonal Fluctuations Along the Menstrual Cycle on Amygdala Structure and Function
In women of the reproductive age, using MRI, an increase in gray matter volume in the dorsal part of the left amygdala was detected in the luteal phase of the menstrual cycle, characterized by a dip in estrogen levels, compared to the late follicular phase, characterized by heightened levels of estrogen [50]. Additionally, these volumetric increases in gray matter volume were linked to increased levels of negative mood following exposure to negatively valenced stimuli [50]. The results provided evidence for a positive correlation between amygdala volumetric increase and heightened levels of stress-induced negative emotionality in the luteal phase [50]. Similarly, in a different study employing fMRI, it was found that increased levels of amygdala activity in response to negatively valenced, or arousing stimuli during the early follicular phase, were associated with low levels of both estrogen and progesterone [51]. Therefore, together, these findings suggest that low levels of female gonadal hormones are associated with increased amygdala gray matter volume and functional activity [50,51] (Figure 1).
3.4. Influence of Hormonal Fluctuations Along the Menstrual Cycle on dlPFC Structure and Function
Applying fMRI during cognitive testing, dlPFC activation was further shown to vary along the menstrual cycle (Figure 1). Using fMRI, activity across different brain regions was assessed while the participants, women of reproductive age, performed cognitive testing using spatial navigation and verbal fluency tasks [44]. Further connectivity analyses were conducted to evaluate network alterations across the menstrual cycle stages [44]. It was found that during the luteal phase, characterized by heightened levels of progesterone and lower levels of estrogen, the right dlPFC displayed heightened levels of activation as assessed using fMRI across tasks [44]. As the authors found decreased levels of hippocampal activation in the luteal phase, without significant alterations in task performance, increased right dlPFC activation was hypothesized to serve as a compensatory mechanism that reduces negative effects of the decreased hippocampal activation on performance in the verbal fluency test [44]. Additionally, no effect of progesterone on activity in the left dlPFC was found [44]. Activity in the left dlPFC is considered to mainly facilitate performance in cognitive tests [44]. Similarly, in a study employing fMRI in adolescent girls, it was shown that increased levels of 17β-estradiol were associated with heightened activity in both the left and right dlPFC during an emotion regulation task[52]. Specifically, findings revealed an association between dlPFC activity and 17β-estradiol levels in the condition requiring the active down-regulation of negative emotions in response to negatively valenced stimuli[52]. In contrast, no association between dlPFC activity and 17β-estradiol levels was found in the condition requiring participants to passively view images[52]. Thus, studies implicate the modulatory role of estrogen on dlPFC functioning, facilitating an increased capacity for emotion regulation[44,52].
3.5. Synergistic Effects of Hormonal Fluctuations and Stress Reactivity on Psychiatric Symptoms
Evidence suggests that fluctuations in gonadal hormones, associated with the menstrual cycle, are related to the severity of mental health-related symptoms in women diagnosed with a psychiatric disease [53]. Findings further suggest that a disruption in HPA axis functioning may be implicated in the pathophysiology of premenstrual syndrome (PMS). PMS is characterized by the experience of adverse emotional symptoms, including depression, inattentiveness, fatigue and increased perceived stress during the late luteal phase of the menstrual cycle, which is characterized by a decline in progesterone and estrogen levels [54]. Evidence indicates decreased levels of cortisol secretion in the TSST in the context of PMS, and heightened baseline cortisol levels throughout the day, indicating dysfunctional HPA activation in the affected women [55,56]. These findings may suggest that hormonal fluctuations throughout the menstrual cycle and disruptions in HPA axis functioning may induce synergistic effects leading to an increased susceptibility to the experience of psychiatric symptoms.
4. HPA Axis
Evidence indicates that dysregulation of the physiological stress response is critically implicated in the pathophysiology of depression. The development of mood-related disorders is frequently preceded by the experience of stress-inducing life events, such as assault, loss of a loved one or financial insecurities [57]. Additionally, in humans, studies indicate altered plasma levels of cortisol, the primary stress hormone, with inconsistencies between studies. While some studies suggest increased levels of morning salivary cortisol in MDD [58,59], other studies indicate that cortisol secretion appears inconsistent and irregular within groups of patients with MDD [60].
The HPA axis comprises a series of signalling steps that induce a physiological stress response [61]. Activation of the HPA axis facilitates the neuroendocrine stress response, enabling the restoration of homeostasis throughout the body [62]. The paraventricular nucleus (PVN) of the hypothalamus receives projections from different structures in the central nervous system (CNS), such as the amygdala, contributing to modulating the stress response [63]. From the PVN projecting to the median eminence, a vascular structure connecting the hypothalamus with the pituitary gland, corticotropin-releasing hormone (CRH) and arginine vasopressin (AVP) are secreted [64,65,66]. The release of CRH and AVP from the PVN stimulates corticotropes, a group of cells located in the anterior pituitary, which are implicated in the production and secretion of adrenocorticotropic hormones (ACTH) [67,68]. The release of ACTH from the anterior pituitary induces glucocorticoid secretion from the adrenal glands [69,70].
Under normal conditions, glucocorticoids are released from the adrenal cortex in a pulsatile manner along the circadian rhythm [61,71]. While acute activation of the HPA axis is beneficial, when it becomes chronic, it can have deleterious effects throughout the body [61]. For example, chronically elevated levels of glucocorticoids induce an increased proinflammatory response that contributes to an increased risk of chronic disease conditions such as cancer, cardiovascular diseases, or neurodegenerative diseases [72,73,74]. Chronic glucocorticoids are further associated with disruptions in emotion regulation-associated regions such as the PFC and hippocampus, which are particularly dense in GRs, in humans and in animal models[75,76,77,78,79].
4.1. HPA Axis Regulation Through Harmful Feedback Mechanisms
Cortisol is implicated in a negative feedback mechanism at the level of the hypothalamus and the anterior pituitary, thus inhibiting CRH and ACTH secretion [80]. These mechanisms induce the regulation of cortisol release and the stress response through the binding to MRs and GRs [81,82]. Occupation of MRs occurs rapidly. As the binding of glucocorticoids to MRs is high already under normal physiological conditions, glucocorticoid binding to GRs is relevant to HPA regulation under acute and chronic stress [81,82]. However, under chronic stress exposure, linked to psychiatric diseases such as depression, the excessive release of glucocorticoids induces glucocorticoid insensitivity and decreased negative feedback on HPA axis functioning [81,82,83].
4.2. HPA Axis and Female Gonadal Hormones
Findings derived from preclinical research indicate a critical role of the emotion regulation circuit, as outlined above, in the modulation of HPA activity [37,81,82,83,84,85]. Projections from the hippocampus and PFC regions to the HPA axis are critically implicated in the inhibition of HPA axis activation [86,87]. Through relay stations such as the BNST and neurons in the peri-PVN area, CRH neurons in the PVN receive modulatory inputs from limbic and PFC regions, facilitating an inhibition of the stress response [84,85,86]. Studies indicate that inhibitory neurons in the BNST may mediate the inhibitory effects of projections from higher-order regions, such as the PFC and hippocampus [87]. Inhibitory projections from the BNST onto PVN neurons have been shown to exert an inhibitory effect on PVN activity, thus decreasing HPA axis activity and glucocorticoid release. For example, evidence indicates that excitatory projection neurons originating from the ventral hippocampus facilitate the inhibition of CRH cells in the PVN, modulating HPA axis activity by synapsing on GABAergic parvalbumin (PV)-positive (+) and glutamic acid decarboxylase (GAD)+ neurons in the BNST. Tracing experiments in animal models also revealed that GABAergic neurons in the BNST and peri-PVN integrate inputs from different brain areas implicated in the evaluation of stressors, projecting both excitatory and inhibitory inputs into the PVN [84,85].
Evidence further indicates a modulatory role of the central nucleus of the amygdala (CeA) on BNST neurons. Optogenetic studies conducted in animal models suggest a mutual connectivity between the CeA and BNST, with the BNST sending inhibitory output onto neurons in the amygdala [88]. By sending inhibitory projections to the BNST, the CeA facilitates the removal of inhibitory connections from the BNST to PVN neurons, thereby inducing increased levels of glucocorticoid secretion upon HPA axis activity. Evidence further indicates that chronic stress modulates modulatory inputs from the amygdala to PFC neurons [89]. Specifically, findings suggest that exposure to chronic stress is associated with increased levels of glutamatergic release from the basolateral amygdala (BLA) to the PFC [89]. Further, inhibitory connections from the BNST onto neurons in the CeA have been shown to reduce fear response [88]. However, findings further indicate a significant degree of heterogeneity in BNST nuclei and projections, with the stimulation of distinct projections from the BLA to the BNST exerting either anxiogenic or anxiolytic effects depending on the type of neuron [90].
In summary, evidence indicates that GABAergic neurons in the BNST facilitate inhibition of the HPA axis’s activity by receiving excitatory inputs from several regions across the brain. Regions implicated in the emotion regulation circuit in the brain, such as the PFC and hippocampus, are involved in the regulation of the HPA axis by sending excitatory projections to inhibitory neurons within relay stations such as the BNST.
5. GABAergic System
GABAergic neurons play a crucial role in regulating emotion regulation circuits in the brain, as well as the HPA axis. Evidence indicates a role for key brain regions forming part of the emotion regulation network in the brain, also displaying functional alterations across the menstrual cycle (reviewed above, in Section 3), in the modulation of HPA axis activity.
In the brain, regulated network activity arises from the functioning of microcircuits consisting of dense networks of inhibitory interneurons synapsing onto other inhibitory interneurons as well as excitatory pyramidal neurons spanning across several brain areas [91]. Populations of interneurons, consisting of PV+ or somatostatin (SST)+ interneurons, are critically implicated in the coordinated activity of large populations of neurons [92]. The rhythmic activation of inhibitory neurons was shown to create synchronized inhibition of excitatory pyramidal neurons [93]. In the PFC, fast-spiking PV interneurons inhibit outputs by forming synapses at the soma or axonal regions located in proximity to the soma. The rhythmic inhibition of projection neuron activity by PV interneuron activity contributes to the generation of oscillatory rhythms such as gamma and theta waves, enabling the extensive network coordination required for information processing [91,94,95]. By contrast, SST+ interneurons form inhibitory connections with other PV-expressing interneurons or dendritic branches of projection neurons [96]. SST+ neurons facilitate phasic inhibition, contributing to the modulation of networks through the gating of input signals onto specific projection neurons or the inhibition of PV+ interneurons, which are involved in modulating the outputs of projection neurons [96]. In line with the important role of inhibitory interneurons in information processing and network functioning, excitation/inhibition imbalance resulting from dysfunctional interneuronal activity is associated with neuropsychiatric diseases, causing, for example, disruptions in information processing, as well as social dysfunction [97].
While PV and SST expressing GABAergic interneurons are implicated in the coordination of neural networks through the generation of oscillatory activity, a subtype of GABAergic neurons expressing SST further forms long-range projections across brain regions [95,98]. In concert with glutamatergic projection neurons, these GABAergic projection neurons contribute to the precise temporal coordination across regions [98]. Evidence indicates a critical role for those long-range GABAergic projection neurons in the transmission of oscillatory rhythms within a region and communicated between regions [98]. Findings from preclinical studies reveal, for example, that long-range GABAergic projection neurons are implicated in the precisely timed input of firing rhythms across subregions of the hippocampus during sleep and wakefulness [98].
5.1. GABAergic System and Female Gonadal Hormones
Female ovarian hormones, derived from progesterone and testosterone, such as allopregnanolone and 17β-estradiol, were found to exert antidepressant and anxiolytic effects in animal models by modulating the GABAergic system. GABA-A receptors vary depending on various factors, such as psychological stress levels, throughout the ovarian cycle, and in psychiatric diseases, as well as in states presenting hormonal alterations, such as the postpartum period or during pregnancy [99,100]. Genetic ablation of this receptor was found to be related to depressive-like behaviour in mice [101]. Additionally, treatment with 17β-estradiol in female ovariectomized rats exposed to four weeks of chronic mild stress resulted in the upregulation of GABA-related messenger (mRNAs) and GABA levels in the amygdala, the hippocampus and PFC, as well as decreased levels of anxiety-like behaviours[102].
Gonadal hormones such as allopregnanolone and 17β-estradiol exert mood-modulating effects, for example, by acting as positive allosteric modulators (PAMs) or through the transcriptional regulation of genes encoding subunits of the GABA-A receptor [103,104]. GABA is the main inhibitory neurotransmitter in the brain, consisting of subunits surrounding pore-forming a chloride (CL-) ion gradient [105], with subunit composition and formation determining its functioning [106]. Evidence suggests that the presence of the δ subunit in GABAergic neurons plays a critical role in modifying the specific binding of gonadal hormones, including allopregnanolone, to exert mood-related effects [107,108]. Allopregnanolone was shown to reduce symptoms in a rat model of premenstrual depression, as assessed through performance in the forced swim test, by acting on the δ-GABA-A receptors in the hippocampus [108]. Further, exposure to 17β-estradiol during developmental periods has been shown to increase levels of extrasynaptic δ-GABA-A receptors affect GABAergic tone in rats and in vitro, using cultured neurons from the hippocampus[109,110].
Clinical studies further indicate a dynamic modulation of GABAergic activation by exposure to female gonadal hormones across the menstrual cycle [111]. For example, findings using magnetic resonance spectroscopy (MRS) revealed increased levels of GABAergic activity around ovulation in the PFC of reproductive-age women not using any contraceptives [112]. Additionally, the exposure to 17β-estradiol may facilitate the precise timing of GABAergic neuron firing, enabling the network activation required for emotion regulation [113]. For example, administration of 17β-estradiol in women undergoing the early follicular phase, associated with low levels of estrogen and progesterone, increases inhibitory activity within PFC regions. Network activation was measured using resting-state(rs)fMRI while participants conducted an emotion regulation task [114]. Using spectral dynamic causal modelling (spDCM) 17β-estradiol administration was associated with increased levels of inhibitory activity among PFC regions, specifically, the vlPFC and dorsomedial(dm)PFC, and parietal cortex, and with an increased inhibition of emotion generating regions, such as the amygdala [114]. Further, the increased levels of inhibitory activity within PFC regions, and suppression of amygdala activity, were associated with an increased performance in the emotion regulation task [114]. These studies indicate that 17β-estradiol may facilitate inhibitory activity within frontal regions, thus facilitating the capacity to regulate negative emotions[115].
6. Perineuronal Nets
Interneuronal functioning, integrity and connectivity is modulated by the presence of perineuronal nets (PNNs). Interneurons, particularly PV+ interneurons, are frequently enwrapped by PNNs, which modulate their firing rate and potential for rewiring [116,117]. PNNs are lattice-like structures consisting of chondroitin sulphate proteoglycans (CSPGs), forming part of the extracellular matrix, that enwrap the soma and proximal parts of neuronal dendrites [116]. Preclinical research indicates that PNNs form around interneurons in an experience-dependent manner throughout development, for example, in the visual cortex [118] .
PNNs appear to be implicated in the regulation of interneuronal functioning, modulating their excitability, and serving as a protective barrier, while simultaneously restricting their capacity for synaptic rewiring[119,120]. The ensheathment of interneurons by PNNs facilitates their protection from damaging factors, such as high levels of oxidative stress[120]. In contrast, the removal of PNNs was associated with increased levels of excitability of interneuronal cells in culture [117]. Further, a reduction in PNN components, such as brevican, neurocan, tenascin-c and tenascin-r, is implicated in the reduced occurrence of PV[121]. PV is implicated in the absorption of calcium within the cell, thereby facilitating the interneuronal capacity to fire rapidly in short intervals [122]. As a result, the loss of PV resulting, for example, from the loss of protective PNNs, may interfere with their fast-spiking properties, disrupting their capacity to precisely regulate the firing rates of other interneurons or pyramidal neurons [122,123,124]. Disruption in the functioning of GABAergic interneurons, resulting from a decline in PV expression, may modulate the excitatory/inhibitory balance toward a higher level of excitation, potentially disrupting the interneuronal capacity to form the precisely-timed neuronal oscillations critical for synchronized brain activity in emotion regulation circuits [92,121].
6.1. Modulation of PNNs by Exposure to Female Gonadal Hormones
Findings indicate a sex difference in the expression of PNNs throughout postnatal development, potentially suggesting a regulatory role of gonadal hormones on the regulation of PNN integrity. For example, evidence indicates that the pace at which the development of PNNs proceeds differs in the medial (m)PFC between male and female rats. It was found that, while in males the beginning of the pubertal period did not modulate PNN density, in females the onset of puberty was associated with a decline in PNN structures [125]. Later in development, starting around early adulthood, an increase in PNN occurrence was detected in both males and females [125]. Puberty is a period marked by strong hormonal fluctuations in women and the increased prevalence of mood-related disorders. Together, these findings suggest that hormonal changes associated with pubertal transitions in females may modulate the risk of development through the modulation of PNN functioning.
Additionally, evidence suggests that PNNs in the medial preoptic area (mPOA) are modulated by female gonadal hormones. In animal models, throughout the reproductive cycle, alterations in PNNs have been detected [126]. Additionally, it was found that interneurons enwrapped by PNN components express estrogen receptor (ER)α and progesterone receptors, indicating a role for hormonal modulation, further highlighting the role of female gonadal hormones in influencing PNNs [126].
6.2. The Role of Microglia in PNN Remodeling
Recent evidence reveals a critical role of microglia in the restructuring of PNNs [119]. Administration of the anti-depressant agent ketamine, as well as light flickering at 40 to 60Hz, increased the presence of PNN fragments within microglia, indicating phagocytosis of PNN components by microglia [119]. These findings suggest that different treatment approaches can induce microglial functional alterations, facilitating the modulation of PNNs.
Together, these findings suggest a role for PNNs in modulating interneuronal integrity and function, with the abnormal formation of PNNs potentially leading to a decline in interneuronal numbers or disrupting their capacity for synaptic rewiring [119,120]. Furthermore, microglia are implicated in the degradation of PNNs, which may be influenced by female gonadal hormones [119,126].
7. Microglia
Microglia are regulated by even subtle changes in their micro-environment, adapting their structure and function to maintain homeostasis in the CNS [127,128]. These brain-resident macrophages constitute a highly dynamic group of cells with their fine processes continuously scanning their surroundings, enabling the rapid response to signals, such as pathogens or factors released by neurons and other glial cells [129]. Further, an adjustment in the density and distribution of the microglial population occurs in response to changes in homeostasis, such as upon stress, injury or pathology, facilitating their capacity to quickly respond, notably by releasing trophic factors, cytokines or through the process of phagocytosis [127,129,130].
7.1. Microglial Involvement in Neuroplasticity
Microglia were shown to be critically involved in the cellular and molecular mechanisms underlying neuroplasticity, for example, by regulating neurogenesis and the formation, maturation and elimination of synapses [131]. During development and in adulthood, microglia shape synaptic connections in an experience-dependent manner [22,132]. The complement system, comprised of over 30 proteins implicated in the innate immune response, is implicated in the tagging of less active synapses for microglial removal [133,134]. In addition, microglia can facilitate activity and experience-dependent synapse formation and synaptic rewiring through their release of neurotrophic factors, such as BDNF [135,136]. Therefore, microglia are critically implicated in maintaining CNS homeostasis, notably through experience-dependent shaping of neuronal networks [22,136].
7.2. Microglial Modulation Through Hormonal Exposure
Gonadal hormones, derived from cholesterol, can bind to microglial receptors [137]. Cell culturing studies reveal the presence of all three of the main estrogen receptors, ERα, ERβ, and G protein–coupled estrogen receptor (GPER) on microglia [138]. Evidence indicates that 17β-estradiol, which is elevated in the follicular phase, promotes an anti-inflammatory response in microglia. For example, administration of 17β-estradiol facilitated a reduction in the release of proinflammatory cytokines in rat-derived microglial cells in primary cultures. The treatment of microglia with LPS induced in the production of various inflammatory mediators, including inducible form of NO synthase (iNOS), prostaglandin-E(2) (PGE(2)), and metalloproteinase (MMP)-9 [139]. By contrast, 17β-estradiol induced a significant reduction in these mediators[137].
7.3. Microglial Modulation by Glucocorticoids
Glucocorticoids are secreted as the final step of HPA axis activity, resulting from stress exposure [140]. By binding to GRs and MRs located on the surface of microglial glucocorticoids induce microglial morphological alterations and increased release of proinflammatory cytokines [24,141].
Additionally, findings revealing increased co-expression of the phagolysosomal marker CD68 with the microglia/macrophage marker Iba1 suggest increased levels of phagocytosis in the hypothalamus resulting from chronic stress exposure in animal models [142]. Glucocorticoid exposure may prime microglia in their response to a subsequent pro-inflammatory stimuli, increasing the release of proinflammatory cytokines in response to the second stimulus [143]. For example, it was found that exposure to 10 days of glucocorticoid administration, in vivo, followed by the extraction of microglia from the hippocampus and the administration of LPS in vitro, was associated with an increased microglial secretion of pro-inflammatory markers such as TNFα, IL-1β, IL-6 and NLRP3 by microglia [143]. Moreover, it was found that, exposure to high levels of glucocorticoids, induced through exposure to chronic stress, are associated with microglial morphological changes such as reduced levels of branching and an enlarged soma in the hypothalamus in an animal model[131]. However, findings from animal models indicate a heterogenous effect of stress on microglia depending on the region investigated, as well as the intensity and duration of the stressor applied[144]. For example, 6 hours of restraint-induced stress across three consecutive days modified microglial proliferative activity, labeled using the marker BrdU+, in a region dependent manner[144]. Significant changes in microglial proliferative activity were found in the hippocampus and hypothalamus, with strongest increases in microglial proliferation detected in the hippocampus[144]. In contrast, findings revealed no alterations in microglial proliferative changes in the PFC[144]. Additionally, it was found that the stress exposure to three hours daily for three consecutive days modulated microglia morphological features in a highly region-dependent manner[145]. Findings revealed that the presence of microglial morphological states, such as amoeboid-like, rod-like, hypertrophic or ramified, differed across the investigated regions, which included the PFC, amygdala and hippocampus[145]. Moreover, across these regions, stress exposure induced a shift in the numbers of microglia belonging to the different states, with the number of microglia across states differing between the regions after stress exposure[145]. Additionally, findings from a different study further revealed that both the sex of the animal and the type of stressor applied determine the impact of stress on microglial morphological features[146]. It was found that, at baseline, before exposing the animals to any type of stressor, microglia morphological characteristics already differed between the sexes[146]. Further, a strong shift in microglial morphological state, with microglia adopting a more amoeboid-like shape after exposure to both acute and chronic stress, was observed in females but not in males within the mPFC[146]. Moreover, the changes in microglial morphological characteristics differed depending on the type of the stressor, with varying effects depending on the duration of the stressor applied, the region investigated, and the sex of the animal [147].
In summary, findings reveal a complex relationship between the exposure to heightened levels of glucocorticoids resulting from stress exposure and alterations in microglia. While the findings derived from in vitro studies indicate a shift towards a more amoeboid and less ramified state, this shift may differ depending on factors such as stressor type, investigated region, and sex of the animal [144,145,146,147].
7.4. Microglial Role in the Shaping of Interneuronal Microcircuits
Evidence reveals that microglia are implicated in the pruning of inhibitory neuronal circuits through different mechanisms. Studies indicate a mutual interaction between GABAergic neurons and microglia, potentially determining pathological outcomes [148,149,150]. Microglia equipped with GABA-B receptors play a critical role in the shaping of GABAergic neurocircuits [151]. This microglial state was shown to be implicated in the refinement of inhibitory circuits during postnatal development in mice [151]. This reshaping process was disrupted through the knockout of GABA-B receptor in microglia, suggesting a mutual communication between GABAergic neurons and microglia in this process[151]. Similarly, in adulthood, the presence of GABA-B receptors on microglia was shown to be implicated in the communication of microglia with GABAergic neurons[152]. Further evidence indicate a role of the C3/C3aR (complement component 3/complement component 3a receptor) pathway in microglia in this synaptic pruning of inhibitory neurons[152,153]. Additionally, in a mouse model of ischemic stroke, microglia were shown to be involved in the degradation of GABAergic synapses through the release of BDNF, exerting its effect through the binding to the tropomyosin receptor kinase B (TrkB) receptor [154]. The degradation of inhibitory synapses is reflected in morphological changes with microglia adopting an amoeboid-like state, characterized by shortened processes and an enlarged soma [154,155]. Additionally, the increased expression of cluster of differentiation 11b (CD11b), the increased secretion of proinflammatory cytokines such as IL-1β and TNF-α by microglia was detected [154]. By contrast, the release of proBDNF was implicated in the disruption of glutamatergic synapses through binding of the p75^NTR receptor [154]. Therefore, several lines of evidence indicate the involvement of microglia in the shaping of interneuronal circuits during development and in adulthood.
Additionally, studies indicate that interneuronal activity may directly modulate microglial functioning, serving a protective role against the development of pathologies in some contexts. For example, activation of the GABA-B receptor on human-derived microglia, obtained through cell culturing, has been shown to reduce the release of IL-6 and IL-12p40 (interleukin-12 subunit beta) following stimulation with LPS in vitro [156]. Together, these studies reveal a critical role of the mutual interaction of microglia and the GABAergic system in guiding the shaping of GABAergic microcircuits, which may potentially have larger implications for network functioning across the brain modulated through female gonadal hormones.
In summary, exposure to physiological levels of gonadal hormones may be associated with anti-inflammatory effects on microglia. Therefore, the drop in gonadal hormones such as estrogen during the luteal phase of the menstrual cycle, in combination with the exposure to increased levels of glucocorticoids, may facilitate the increased release of proinflammatory cytokines and a shift in microglial state leading to the remodeling of emotion regulation circuits leading to the increased expression of psychiatric symptoms.
8. Conclusion and Future Perspectives
In conclusion, hormonal fluctuations across the menstrual cycle are associated with neuronal network alterations across the brain. These network alterations are the result of the coordinated activity of interneuron microcircuit facilitating the precise spatial firing of projection neurons and oscillatory activity of large groups of neurons enabling the communication across brain regions. Microglia may be critically implicated in the reshaping of these circuits, potentially, under pathological conditions contributing to a disruption in emotion regulation networks. Microglia functioning is modulated through their exposure to both glucocorticoids and female gonadal hormones, while the increased modulation of microglia during phases associated with low levels of female gonadal hormones may be associated with their detrimental modulation of intraneuronal circuits. The excessive pruning of inhibitory synapses, may lead to a shift in excitation-inhibition balance towards excitation, leading to dysregulated network activation.
In this review, the critical influence of female gonadal hormones on neurocircuits implicated in MDD is discussed. The findings summarized stress the critical need to consider fluctuating levels of female gonadal hormones for the development of novel treatment approaches for MDD. Microglia, which are modulated by both stress hormones and female gonadal hormones, and involved in the reshaping of neuronal circuits, and may perform this critical function in circuits implicated in emotion regulation. However, the understanding of the role of microglia in the reshaping of neuronal circuits across stages of the menstrual cycle and stages of differing levels of female gonadal hormones is currently limited. Future studies should focus on closing this knowledge gap, investigating the modulatory role of female gonadal hormones and increased levels of glucocorticoids on the reshaping of neurocircuits by microglia across stages of the menstrual cycle. Employing different imaging modalities in humans may facilitate a deeper understanding of this interaction of microglia with neuronal networks and the modulatory effect by levels of female gonadal hormones. The application of positron emission tomography (PET) using markers such as 18 kDa translocator protein (TSPO) enables the imaging of microglia in humans in vivo [157]. Although this approach is still lacking in specificity, new tracers are being developed, indicating that the combined use of PET with MRI or fMRI may enable, in the future, the study of microglia in humans while simultaneously taking into consideration functional and structural alterations[158]. Simultaneously, the using PET in combination with MRS may enable researchers to investigate microglial remodeling of neurocircuits in humans, while taking into account levels of GABAergic activity [159].
Abbreviation List
| GABA | gamma-aminobutyric acid |
| ACC | anterior cingulate cortex |
| ACTH | adrenocorticotropic hormones |
| AVP | arginine vasopressin |
| BDNF | brain-derived neurotrophic factor |
| BLA | basolateral amygdala |
| BNST | bed nucleus of the stria terminalis |
| C3/C3aR | complement component 3/complement component 3a receptor |
| CD11b | cluster of differentiation 11b |
| CeA | central nucleus of the amygdala |
| Cl- | chloride ion |
| CNS | central nervous system |
| CRH | corticotropin-releasing hormone |
| CSPGs | chondroitin sulphate proteoglycans |
| dlPFC | dorsolateral PFC |
| dmPFC | dorsomedial PFC |
| DSM-5 | Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition |
| ERα | estrogen receptors alpha |
| ERβ | estrogen receptors beta |
| fMRI | functional magnetic resonance imaging |
| GAD | glutamic acid decarboxylase |
| GPER | G protein–coupled estrogen receptor |
| GR | glucocorticoid receptors |
| HPA | hypothalamic pituitary adrenal axis |
| Iba1 | ionized calcium-binding adapter molecule 1 |
| IL-12p40 | interleukin 12p40 |
| IL-12p40 | interleukin-12 subunit beta |
| IL-1β | interleukin-1 beta |
| IL-6 | interleukin-6 |
| iNOS | NO synthase |
| LH | luteinizing hormone |
| LPS | lipopolysaccharide |
| MDD | major depressive disorder |
| MMP-9 | metalloproteinase-9 |
| mPFC | medial prefrontal cortex |
| mPOA | medial preoptic area |
| MR | mineralocorticoid receptors |
| MRI | magnetic resonance imaging |
| MRS | magnetic resonance spectroscopy |
| NLRP3 | NOD-like receptor family pyrin domain containing 3 |
| NO | nitric oxide |
| P75^NTR | p75 neurotrophin receptor |
| PAM | positive allosteric modulator |
| PANAS | Positive and Negative Affect Schedule |
| PET | positron emission tomography |
| PFC | prefrontal cortex |
| PGE(2) | prostaglandin-E(2) |
| PMS | premenstrual syndrome |
| PNN | perineuronal nets |
| PV | parvalbumin |
| PVN | paraventricular nucleus |
| RCT | randomized control trials |
| rs-fMRI | resting state functional magnetic imaging |
| rsFC | resting state functional connectivity |
| rsfMRI | resting state functional magnetic resonance imaging |
| spDCM | spectral dynamic causal modelling |
| SSRIs | selective serotonin reuptake inhibitors |
| SST | somatostatin |
| TMS | transcranial magnetic stimulation |
| TNFα | tumor necrosis factor alpha |
| TrkB | tropomyosin receptor kinase B |
| TSPO | 18 kDa translocator protein |
| TSST | Trier Social Stress Test |
| vlPFC | ventrolateral prefrontal cortex |
| vmPFC | ventromedial prefrontal cortex |
| WHO | World Health Organization |
References
- Zhang Y, Jia X, Yang Y, Sun N, Shi S, Wang W. Change in the global burden of depression from 1990-2019 and its prediction for 2030. J Psychiatr Res. 2024;178:16-22. [CrossRef]
- Lépine JP, Briley M. The increasing burden of depression. Neuropsychiatr Dis Treat. 2011;7(Suppl 1):3-7. [CrossRef]
- Uher R, Payne JL, Pavlova B, Perlis RH. Major Depressive Disorder in Dsm-5: Implications for Clinical Practice and Research of Changes from Dsm-Iv. Depress Anxiety. 2014;31(6):459-471. [CrossRef]
- Tolentino JC, Schmidt SL. DSM-5 Criteria and Depression Severity: Implications for Clinical Practice. Front Psychiatry. 2018;9:450. [CrossRef]
- Gonda X, Pompili M, Serafini G, Carvalho AF, Rihmer Z, Dome P. The role of cognitive dysfunction in the symptoms and remission from depression. Ann Gen Psychiatry. 2015;14(1):27. [CrossRef]
- Medina A, Seasholtz AF, Sharma V, et al. Glucocorticoid and mineralocorticoid receptor expression in the human hippocampus in major depressive disorder. J Psychiatr Res. 2013;47(3):307-314. [CrossRef]
- Wiersma JE, Hovens JGFM, van Oppen P, et al. The importance of childhood trauma and childhood life events for chronicity of depression in adults. J Clin Psychiatry. 2009;70(7):983-989. [CrossRef]
- Bisht K, Sharma K, Tremblay MÈ. Chronic stress as a risk factor for Alzheimer’s disease: Roles of microglia-mediated synaptic remodeling, inflammation, and oxidative stress. Neurobiol Stress. 2018;9:9-21. [CrossRef]
- Hellwig S, Brioschi S, Dieni S, et al. Altered microglia morphology and higher resilience to stress-induced depression-like behavior in CX3CR1-deficient mice. Brain Behav Immun. 2016;55:126-137. [CrossRef]
- Köhler CA, Freitas TH, Maes M, et al. Peripheral cytokine and chemokine alterations in depression: a meta-analysis of 82 studies. Acta Psychiatr Scand. 2017;135(5):373-387. [CrossRef]
- Kopelman J, Keller TA, Panny B, et al. Rapid neuroplasticity changes and response to intravenous ketamine: a randomized controlled trial in treatment-resistant depression. Transl Psychiatry. 2023;13(1):1-9. [CrossRef]
- Hirschfeld RMA. Efficacy of SSRIs and Newer Antidepressants in Severe Depression: Comparison With TCAs. J Clin Psychiatry. Published online 1999.
- Tynan RJ, Weidenhofer J, Hinwood M, Cairns MJ, Day TA, Walker FR. A comparative examination of the anti-inflammatory effects of SSRI and SNRI antidepressants on LPS stimulated microglia. Brain Behav Immun. 2012;26(3):469-479. [CrossRef]
- Lepack AE, Fuchikami M, Dwyer JM, Banasr M, Duman RS. BDNF Release Is Required for the Behavioral Actions of Ketamine. Int J Neuropsychopharmacol. 2015;18(1):pyu033. [CrossRef]
- Saelens J, Gramser A, Watzal V, Zarate CA, Lanzenberger R, Kraus C. Relative effectiveness of antidepressant treatments in treatment-resistant depression: a systematic review and network meta-analysis of randomized controlled trials. Neuropsychopharmacology. 2025;50(6):913-919. [CrossRef]
- Lynall ME, McIntosh AM. The Heterogeneity of Depression. Am J Psychiatry. 2023;180(10):703-704. [CrossRef]
- Garcia-Sifuentes Y, Maney DL. Reporting and misreporting of sex differences in the biological sciences. Allison DB, Zaidi M, Vorland CJ, Kahathuduwa C, eds. eLife. 2021;10:e70817. [CrossRef]
- Sloan DM, Sandt AR. Gender Differences in Depression. Womens Health. 2006;2(3):425-434. [CrossRef]
- Genuchi MC, Mitsunaga LK. Sex Differences in Masculine Depression: Externalizing Symptoms as a Primary Feature of Depression in Men. J Men’s Stud. 2015;23(3):243-251. [CrossRef]
- Rice SM, Fallon BJ, Aucote HM, Möller-Leimkühler A, Treeby MS, Amminger GP. Longitudinal sex differences of externalising and internalising depression symptom trajectories: Implications for assessment of depression in men from an online study. Int J Soc Psychiatry. 2015;61(3):236-240. [CrossRef]
- Cavanagh A, Wilson CJ, Kavanagh DJ, Caputi P. Differences in the Expression of Symptoms in Men Versus Women with Depression: A Systematic Review and Meta-analysis. Harv Rev Psychiatry. 2017;25(1):29. [CrossRef]
- Tremblay MÈ, Lowery RL, Majewska AK. Microglial interactions with synapses are modulated by visual experience. PLoS Biol. 2010;8(11):e1000527. [CrossRef]
- Baker AE, Brautigam VM, Watters JJ. Estrogen Modulates Microglial Inflammatory Mediator Production via Interactions with Estrogen Receptor β. Endocrinology. 2004;145(11):5021-5032. [CrossRef]
- Picard K, Bisht K, Poggini S, et al. Microglial-glucocorticoid receptor depletion alters the response of hippocampal microglia and neurons in a chronic unpredictable mild stress paradigm in female mice. Brain Behav Immun. 2021;97:423-439. [CrossRef]
- Berboth S, Morawetz C. Amygdala-prefrontal connectivity during emotion regulation: A meta-analysis of psychophysiological interactions. Neuropsychologia. 2021;153:107767. [CrossRef]
- Damborská A, Honzírková E, Barteček R, et al. Altered directed functional connectivity of the right amygdala in depression: high-density EEG study. Sci Rep. 2020;10(1):4398. [CrossRef]
- Helm K, Viol K, Weiger TM, et al. Neuronal connectivity in major depressive disorder: a systematic review. Neuropsychiatr Dis Treat. 2018;14:2715-2737. [CrossRef]
- Zotev V, Phillips R, Young KD, Drevets WC, Bodurka J. Prefrontal Control of the Amygdala during Real-Time fMRI Neurofeedback Training of Emotion Regulation. PLOS ONE. 2013;8(11):e79184. [CrossRef]
- Zhang JX, Bo K, Wager TD, Gross JJ. The brain bases of emotion generation and emotion regulation. Trends Cogn Sci. Published online May 29, 2025. [CrossRef]
- Siegle GJ, Thompson W, Carter CS, Steinhauer SR, Thase ME. Increased Amygdala and Decreased Dorsolateral Prefrontal BOLD Responses in Unipolar Depression: Related and Independent Features. Biol Psychiatry. 2007;61(2):198-209. [CrossRef]
- Jamieson AJ, Leonards CA, Davey CG, Harrison BJ. Major depressive disorder associated alterations in the effective connectivity of the face processing network: a systematic review. Transl Psychiatry. 2024;14(1):62. [CrossRef]
- Izquierdo A, Murray EA. Combined Unilateral Lesions of the Amygdala and Orbital Prefrontal Cortex Impair Affective Processing in Rhesus Monkeys. J Neurophysiol. 2004;91(5):2023-2039. [CrossRef]
- Costafreda SG, Brammer MJ, David AS, Fu CHY. Predictors of amygdala activation during the processing of emotional stimuli: A meta-analysis of 385 PET and fMRI studies. Brain Res Rev. 2008;58(1):57-70. [CrossRef]
- Qasim SE, Mohan UR, Stein JM, Jacobs J. Neuronal activity in the human amygdala and hippocampus enhances emotional memory encoding. Nat Hum Behav. 2023;7(5):754-764. [CrossRef]
- Vandekerckhove M, Cluydts R. The emotional brain and sleep: An intimate relationship. Sleep Med Rev. 2010;14(4):219-226. [CrossRef]
- Treadway MT, Grant MM, Ding Z, Hollon SD, Gore JC, Shelton RC. Early Adverse Events, HPA Activity and Rostral Anterior Cingulate Volume in MDD. PLOS ONE. 2009;4(3):e4887. [CrossRef]
- Herman JP, Ostrander MM, Mueller NK, Figueiredo H. Limbic system mechanisms of stress regulation: hypothalamo-pituitary-adrenocortical axis. Prog Neuropsychopharmacol Biol Psychiatry. 2005;29(8):1201-1213. [CrossRef]
- Dedovic K, Duchesne A, Andrews J, Engert V, Pruessner JC. The brain and the stress axis: the neural correlates of cortisol regulation in response to stress. NeuroImage. 2009;47(3):864-871. [CrossRef]
- Suzuki Y, Tanaka SC. Functions of the ventromedial prefrontal cortex in emotion regulation under stress. Sci Rep. 2021;11(1):18225. [CrossRef]
- He Z, Li S, Mo L, et al. The VLPFC-Engaged Voluntary Emotion Regulation: Combined TMS-fMRI Evidence for the Neural Circuit of Cognitive Reappraisal. J Neurosci. 2023;43(34):6046-6060. [CrossRef]
- Keller M, Mendoza-Quiñones R, Cabrera Muñoz A, et al. Transdiagnostic alterations in neural emotion regulation circuits – neural substrates of cognitive reappraisal in patients with depression and post-traumatic stress disorder. BMC Psychiatry. 2022;22(1):173. [CrossRef]
- Cieslik EC, Zilles K, Caspers S, et al. Is There “One” DLPFC in Cognitive Action Control? Evidence for Heterogeneity From Co-Activation-Based Parcellation. Cereb Cortex. 2013;23(11):2677-2689. [CrossRef]
- Morawetz C, Bode S, Baudewig J, Kirilina E, Heekeren HR. Changes in Effective Connectivity Between Dorsal and Ventral Prefrontal Regions Moderate Emotion Regulation. Cereb Cortex. 2016;26(5):1923-1937. [CrossRef]
- Pletzer B, Harris TA, Scheuringer A, Hidalgo-Lopez E. The cycling brain: menstrual cycle related fluctuations in hippocampal and fronto-striatal activation and connectivity during cognitive tasks. Neuropsychopharmacology. 2019;44(11):1867-1875. [CrossRef]
- Heller C, Güllmar D, Koeppel CJ, et al. Hippocampal volume and affect in response to fluctuating estrogens in menstrual cycle irregularity: a longitudinal single-subject study. Npj Womens Health. 2024;2(1):19. [CrossRef]
- Holesh JE, Bass AN, Lord M. Physiology, Ovulation. In: StatPearls. StatPearls Publishing; 2025. Accessed August 26, 2025. http://www.ncbi.nlm.nih.gov/books/NBK441996/.
- Ko CH, Wong TH, Suen JL, Lin PC, Long CY, Yen JY. Estrogen, progesterone, cortisol, brain-derived neurotrophic factor, and vascular endothelial growth factor during the luteal phase of the menstrual cycle in women with premenstrual dysphoric disorder. J Psychiatr Res. 2024;169:307-317. [CrossRef]
- Baerwald AR, Adams GP, Pierson RA. Form and function of the corpus luteum during the human menstrual cycle. Ultrasound Obstet Gynecol. 2005;25(5):498-507. [CrossRef]
- Lisofsky N, Mårtensson J, Eckert A, Lindenberger U, Gallinat J, Kühn S. Hippocampal volume and functional connectivity changes during the female menstrual cycle. NeuroImage. 2015;118:154-162. [CrossRef]
- Ossewaarde L, van Wingen GA, Rijpkema M, Bäckström T, Hermans EJ, Fernández G. Menstrual cycle-related changes in amygdala morphology are associated with changes in stress sensitivity. Hum Brain Mapp. 2013;34(5):1187-1193. [CrossRef]
- Goldstein JM, Jerram M, Poldrack R, et al. Hormonal cycle modulates arousal circuitry in women using functional magnetic resonance imaging. J Neurosci Off J Soc Neurosci. 2005;25(40):9309-9316. [CrossRef]
- Chung YS, Poppe A, Novotny S, et al. A preliminary study of association between adolescent estradiol level and dorsolateral prefrontal cortex activity during emotion regulation. Psychoneuroendocrinology. 2019;109:104398. [CrossRef]
- Handy AB, Greenfield SF, Yonkers KA, Payne LA. Psychiatric Symptoms Across the Menstrual Cycle in Adult Women: A Comprehensive Review. Harv Rev Psychiatry. 2022;30(2):100. [CrossRef]
- Ko MY, Darouian N. The Importance of an Interdisciplinary Approach in the Transition of At-Risk Patients From Hospital to Skilled Nursing Facilities: A Case Report. Cureus. 2024;16(10):e71629. [CrossRef]
- Beddig T, Reinhard I, Kuehner C. Stress, mood, and cortisol during daily life in women with Premenstrual Dysphoric Disorder (PMDD). Psychoneuroendocrinology. 2019;109:104372. [CrossRef]
- Huang Y, Zhou R, Wu M, Wang Q, Zhao Y. Premenstrual syndrome is associated with blunted cortisol reactivity to the TSST. Stress Amst Neth. 2015;18(2):160-168. [CrossRef]
- Kendler KS, Karkowski LM, Prescott CA. Causal Relationship Between Stressful Life Events and the Onset of Major Depression. Am J Psychiatry. 1999;156(6):837-841. [CrossRef]
- Bhagwagar Z, Hafizi S, Cowen PJ. Increased salivary cortisol after waking in depression. Psychopharmacology (Berl). 2005;182(1):54-57. [CrossRef]
- Hinkelmann K, Moritz S, Botzenhardt J, et al. Cognitive Impairment in Major Depression: Association with Salivary Cortisol. Biol Psychiatry. 2009;66(9):879-885. [CrossRef]
- Peeters F, Nicolson NA, Berkhof J. Levels and variability of daily life cortisol secretion in major depression. Psychiatry Res. 2004;126(1):1-13. [CrossRef]
- Green MR, McCormick CM. Sex and stress steroids in adolescence: Gonadal regulation of the hypothalamic-pituitary-adrenal axis in the rat. Gen Comp Endocrinol. 2016;234:110-116. [CrossRef]
- Goel N, Workman JL, Lee TT, Innala L, Viau V. Sex Differences in the HPA Axis. In: Terjung R, ed. Comprehensive Physiology. 1st ed. Wiley; 2014:1121-1155. [CrossRef]
- Gray TS, Carney ME, Magnuson DJ. Direct Projections from the Central Amygdaloid Nucleus to the Hypothalamic Paraventricular Nucleus: Possible Role in Stress-Induced Adrenocorticotropin Release. Neuroendocrinology. 2008;50(4):433-446. [CrossRef]
- Holmes MC, Antoni FA, Aguilera G, Catt KJ. Magnocellular axons in passage through the median eminence release vasopressin. Nature. 1986;319(6051):326-329. [CrossRef]
- Szabó K, Csányi K. The vascular architecture of the developing pituitary-median eminence complex in the rat. Cell Tissue Res. 1982;224(3):563-577. [CrossRef]
- Wiegand SJ, Price JL. Cells of origin of the afferent fibers to the median eminence in the rat. J Comp Neurol. 1980;192(1):1-19. [CrossRef]
- Luo X, Kiss A, Makara G, Lolait SJ, Aguilera G. Stress-Specific Regulation of Corticotropin Releasing Hormone Receptor Expression in the Paraventricular and Supraoptic Nuclei of the Hypothalamus in the Rat. J Neuroendocrinol. 1994;6(6):689-696. [CrossRef]
- Tse A, Lee AK, Tse FW. Ca2+ signaling and exocytosis in pituitary corticotropes. Cell Calcium. 2012;51(3):253-259. [CrossRef]
- McCartney CR, Marshall JC. Chapter 1 - Neuroendocrinology of Reproduction. In: Strauss JF, Barbieri RL, eds. Yen and Jaffe’s Reproductive Endocrinology (Eighth Edition). Elsevier; 2019:1-24.e8. [CrossRef]
- NEWMAN CB, WARDLAW SL, VAN VUGT DA, FERIN M, FRANTZ AG. Adrenocorticotropin Immunoactivity in Monkey Hypophyseal Portal Blood*. J Clin Endocrinol Metab. 1984;59(1):108-112. [CrossRef]
- Gjerstad JK, Lightman SL, Spiga F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress. 2018;21(5):403-416. [CrossRef]
- Herrero MT, Estrada C, Maatouk L, Vyas S. Inflammation in Parkinson’s disease: role of glucocorticoids. Front Neuroanat. 2015;9. [CrossRef]
- Tang S, Zhang Z, Oakley RH, et al. Intestinal epithelial glucocorticoid receptor promotes chronic inflammation–associated colorectal cancer. JCI Insight. 2021;6(24). [CrossRef]
- Pujades-Rodriguez M, Morgan AW, Cubbon RM, Wu J. Dose-dependent oral glucocorticoid cardiovascular risks in people with immune-mediated inflammatory diseases: A population-based cohort study. PLOS Med. 2020;17(12):e1003432. [CrossRef]
- Sapolsky RM. Glucocorticoids and Hippocampal Atrophy in Neuropsychiatric Disorders. Arch Gen Psychiatry. 2000;57(10):925-935. [CrossRef]
- Wang Q, Van Heerikhuize J, Aronica E, et al. Glucocorticoid receptor protein expression in human hippocampus; stability with age. Neurobiol Aging. 2013;34(6):1662-1673. [CrossRef]
- McKlveen JM, Myers B, Flak JN, et al. Role of Prefrontal Cortex Glucocorticoid Receptors in Stress and Emotion. Biol Psychiatry. 2013;74(9):672-679. [CrossRef]
- Kim EJ, Pellman B, Kim JJ. Stress effects on the hippocampus: a critical review. Learn Mem. 2015;22(9):411-416. [CrossRef]
- McEwen BS, Nasca C, Gray JD. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology. 2016;41(1):3-23. [CrossRef]
- Dogra P, Vijayashankar NP. Dexamethasone Suppression Test. In: StatPearls [Internet]. StatPearls Publishing; 2024. Accessed April 14, 2025. https://www.ncbi.nlm.nih.gov/sites/books/NBK542317/.
- Almeida FB, Pinna G, Barros HMT. The Role of HPA Axis and Allopregnanolone on the Neurobiology of Major Depressive Disorders and PTSD. Int J Mol Sci. 2021;22(11):5495. [CrossRef]
- Daskalakis NP, Meijer OC, de Kloet ER. Mineralocorticoid receptor and glucocorticoid receptor work alone and together in cell-type-specific manner: Implications for resilience prediction and targeted therapy. Neurobiol Stress. 2022;18:100455. [CrossRef]
- Heuser I, Yassouridis A, Holsboer F. The combined dexamethasone/CRH test: A refined laboratory test for psychiatric disorders. J Psychiatr Res. 1994;28(4):341-356. [CrossRef]
- Dong HW, Petrovich GD, Watts AG, Swanson LW. Basic organization of projections from the oval and fusiform nuclei of the bed nuclei of the stria terminalis in adult rat brain. J Comp Neurol. 2001;436(4):430-455. [CrossRef]
- Radley JJ, Gosselink KL, Sawchenko PE. A Discrete GABAergic Relay Mediates Medial Prefrontal Cortical Inhibition of the Neuroendocrine Stress Response. J Neurosci. 2009;29(22):7330-7340. [CrossRef]
- Herman JP, Cullinan WE, Ziegler DR, Tasker JG. Role of the paraventricular nucleus microenvironment in stress integration. Eur J Neurosci. 2002;16(3):381-385. [CrossRef]
- Cole AB, Montgomery K, Bale TL, Thompson SM. What the hippocampus tells the HPA axis: Hippocampal output attenuates acute stress responses via disynaptic inhibition of CRF+ PVN neurons. Neurobiol Stress. 2022;20:100473. [CrossRef]
- Gungor NZ, Yamamoto R, Paré D. Optogenetic study of the projections from the bed nucleus of the stria terminalis to the central amygdala. J Neurophysiol. 2015;114(5):2903-2911. [CrossRef]
- Lowery-Gionta EG, Crowley NA, Bukalo O, Silverstein S, Holmes A, Kash TL. Chronic stress dysregulates amygdalar output to the prefrontal cortex. Neuropharmacology. 2018;139:68-75. [CrossRef]
- Han RW, Zhang ZY, Jiao C, Hu ZY, Pan BX. Synergism between two BLA-to-BNST pathways for appropriate expression of anxiety-like behaviors in male mice. Nat Commun. 2024;15(1):3455. [CrossRef]
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat Rev Neurosci. 2007;8(1):45-56. [CrossRef]
- Ferguson BR, Gao WJ. PV Interneurons: Critical Regulators of E/I Balance for Prefrontal Cortex-Dependent Behavior and Psychiatric Disorders. Front Neural Circuits. 2018;12:37. [CrossRef]
- Hájos N, Paulsen O. Network mechanisms of gamma oscillations in the CA3 region of the hippocampus. Neural Netw Off J Int Neural Netw Soc. 2009;22(8):1113-1119. [CrossRef]
- Antonoudiou P, Tan YL, Kontou G, Upton AL, Mann EO. Parvalbumin and Somatostatin Interneurons Contribute to the Generation of Hippocampal Gamma Oscillations. J Neurosci. 2020;40(40):7668-7687. [CrossRef]
- McKlveen JM, Moloney RD, Scheimann JR, Myers B, Herman JP. “Braking” the Prefrontal Cortex: The Role of Glucocorticoids and Interneurons in Stress Adaptation and Pathology. Biol Psychiatry. 2019;86(9):669-681. [CrossRef]
- Cottam JCH, Smith SL, Häusser M. Target-Specific Effects of Somatostatin-Expressing Interneurons on Neocortical Visual Processing. J Neurosci. 2013;33(50):19567-19578. [CrossRef]
- Yizhar O, Fenno LE, Prigge M, et al. Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature. 2011;477(7363):171-178. [CrossRef]
- Katona L, Micklem B, Borhegyi Z, et al. Behavior-dependent activity patterns of GABAergic long-range projecting neurons in the rat hippocampus. Hippocampus. 2017;27(4):359-377. [CrossRef]
- Jie F, Yin G, Yang W, et al. Stress in Regulation of GABA Amygdala System and Relevance to Neuropsychiatric Diseases. Front Neurosci. 2018;12. [CrossRef]
- Biggio G, Cristina Mostallino M, Follesa P, Concas A, Sanna E. GABA(A) receptor function and gene expression during pregnancy and postpartum. Int Rev Neurobiol. 2009;85:73-94. [CrossRef]
- Shen Q, Lal R, Luellen BA, Earnheart JC, Andrews AM, Luscher B. GABAA receptor deficits cause HPA axis hyperactivity and antidepressant drug sensitivity reminiscent of melancholic forms of depression. Biol Psychiatry. 2010;68(6):512-520. [CrossRef]
- Tongta S, Daendee S, Kalandakanond-Thongsong S. Effects of estrogen receptor β or G protein-coupled receptor 30 activation on anxiety-like behaviors in relation to GABAergic transmission in stress-ovariectomized rats. Neurosci Lett. 2022;789:136885. [CrossRef]
- Maguire JL, Mennerick S. Neurosteroids: mechanistic considerations and clinical prospects. Neuropsychopharmacology. 2024;49(1):73-82. [CrossRef]
- Rodríguez-Palma EJ, Islas-Espinoza AM, Ramos-Rodríguez II, et al. Estradiol modulates the role of the spinal α6-subunit containing GABAA receptors in female rats with neuropathic pain. Eur J Pharmacol. 2024;974:176616. [CrossRef]
- Camille Melón L, Maguire J. GABAergic regulation of the HPA and HPG axes and the impact of stress on reproductive function. J Steroid Biochem Mol Biol. 2016;160:196-203. [CrossRef]
- Glykys J, Mann EO, Mody I. Which GABA(A) receptor subunits are necessary for tonic inhibition in the hippocampus? J Neurosci Off J Soc Neurosci. 2008;28(6):1421-1426. [CrossRef]
- Sabaliauskas N, Shen H, Molla J, et al. Neurosteroid effects at α4βδ GABAA receptors alter spatial learning and synaptic plasticity in CA1 hippocampus across the estrous cycle of the mouse. Brain Res. 2015;1621:170-186. [CrossRef]
- Zhang L, Gao YZ, Zhao CJ, Xia JY, Yang JJ, Ji MH. Reduced inhibitory and excitatory input onto parvalbumin interneurons mediated by perineuronal net might contribute to cognitive impairments in a mouse model of sepsis-associated encephalopathy. Neuropharmacology. 2023;225:109382. [CrossRef]
- Locci A, Porcu P, Talani G, et al. Neonatal estradiol exposure to female rats changes GABAA receptor expression and function, and spatial learning during adulthood. Horm Behav. 2017;87:35-46. [CrossRef]
- Pytel M, Wójtowicz T, Mercik K, et al. 17 β-estradiol modulates GABAergic synaptic transmission and tonic currents during development in vitro. Neuropharmacology. 2007;52(6):1342-1353. [CrossRef]
- De Bondt T, De Belder F, Vanhevel F, Jacquemyn Y, Parizel PM. Prefrontal GABA concentration changes in women—Influence of menstrual cycle phase, hormonal contraceptive use, and correlation with premenstrual symptoms. Brain Res. 2015;1597:129-138. [CrossRef]
- De Bondt T, De Belder F, Vanhevel F, Jacquemyn Y, Parizel PM. Prefrontal GABA concentration changes in women-Influence of menstrual cycle phase, hormonal contraceptive use, and correlation with premenstrual symptoms. Brain Res. 2015;1597:129-138. [CrossRef]
- Wójtowicz T, Lebida K, Mozrzymas JW. 17beta-estradiol affects GABAergic transmission in developing hippocampus. Brain Res. 2008;1241:7-17. [CrossRef]
- Derntl B, Eber CH, Kogler L, Rehbein E, Sundstöm-Poromaa I, Morawetz C. Estradiol modulates changes in effective connectivity in emotion regulation networks. Psychoneuroendocrinology. 2024;167:107103. [CrossRef]
- Derntl B, Eber CH, Kogler L, Rehbein E, Sundstöm-Poromaa I, Morawetz C. Estradiol modulates changes in effective connectivity in emotion regulation networks. Psychoneuroendocrinology. 2024;167:107103. [CrossRef]
- Carceller H, Guirado R, Ripolles-Campos E, Teruel-Marti V, Nacher J. Perineuronal Nets Regulate the Inhibitory Perisomatic Input onto Parvalbumin Interneurons and γ Activity in the Prefrontal Cortex. J Neurosci. 2020;40(26):5008-5018. [CrossRef]
- Dityatev A, Brückner G, Dityateva G, Grosche J, Kleene R, Schachner M. Activity-dependent formation and functions of chondroitin sulfate-rich extracellular matrix of perineuronal nets. Dev Neurobiol. 2007;67(5):570-588. [CrossRef]
- Ye Q, Miao QL. Experience-dependent development of perineuronal nets and chondroitin sulfate proteoglycan receptors in mouse visual cortex. Matrix Biol J Int Soc Matrix Biol. 2013;32(6):352-363. [CrossRef]
- Venturino A, Schulz R, Jesús-Cortés HD, et al. Microglia enable mature perineuronal nets disassembly upon anesthetic ketamine exposure or 60-Hz light entrainment in the healthy brain. Cell Rep. 2021;36(1). [CrossRef]
- Cabungcal JH, Steullet P, Morishita H, et al. Perineuronal nets protect fast-spiking interneurons against oxidative stress. Proc Natl Acad Sci. 2013;110(22):9130-9135. [CrossRef]
- Gottschling C, Wegrzyn D, Denecke B, Faissner A. Elimination of the four extracellular matrix molecules tenascin-C, tenascin-R, brevican and neurocan alters the ratio of excitatory and inhibitory synapses. Sci Rep. 2019;9(1):13939. [CrossRef]
- Bischop DP, Orduz D, Lambot L, Schiffmann SN, Gall D. Control of Neuronal Excitability by Calcium Binding Proteins: A New Mathematical Model for Striatal Fast-Spiking Interneurons. Front Mol Neurosci. 2012;5:78. [CrossRef]
- Orduz D, Bischop DP, Schwaller B, Schiffmann SN, Gall D. Parvalbumin tunes spike-timing and efferent short-term plasticity in striatal fast spiking interneurons. J Physiol. 2013;591(Pt 13):3215-3232. [CrossRef]
- Wang AS, Wan X, Storch DS, et al. Cross-species conservation in the regulation of parvalbumin by perineuronal nets. Front Neural Circuits. 2023;17. [CrossRef]
- Drzewiecki CM, Willing J, Juraska JM. Influences of age and pubertal status on number and intensity of perineuronal nets in the rat medial prefrontal cortex. Brain Struct Funct. 2020;225(8):2495-2507. [CrossRef]
- Uriarte N, Ferreño M, Méndez D, Nogueira J. Reorganization of perineuronal nets in the medial Preoptic Area during the reproductive cycle in female rats. Sci Rep. 2020;10(1):5479. [CrossRef]
- Hammond BP, Manek R, Kerr BJ, Macauley MS, Plemel JR. Regulation of microglia population dynamics throughout development, health, and disease. Glia. 2021;69(12):2771-2797. [CrossRef]
- Jäntti H, Sitnikova V, Ishchenko Y, et al. Microglial amyloid beta clearance is driven by PIEZO1 channels. J Neuroinflammation. 2022;19(1):147. [CrossRef]
- Stence N, Waite M, Dailey ME. Dynamics of microglial activation: A confocal time-lapse analysis in hippocampal slices. Glia. 2001;33(3):256-266. [CrossRef]
- St-Pierre MK, Carrier M, González Ibáñez F, et al. Ultrastructural characterization of dark microglia during aging in a mouse model of Alzheimer’s disease pathology and in human post-mortem brain samples. J Neuroinflammation. 2022;19(1):235. [CrossRef]
- Chagas L da S, Serfaty CA. The Influence of Microglia on Neuroplasticity and Long-Term Cognitive Sequelae in Long COVID: Impacts on Brain Development and Beyond. Int J Mol Sci. 2024;25(7):3819. [CrossRef]
- Ji K, Akgul G, Wollmuth LP, Tsirka SE. Microglia Actively Regulate the Number of Functional Synapses. PLOS ONE. 2013;8(2):e56293. [CrossRef]
- Ostrycharz E, Hukowska-Szematowicz B. New Insights into the Role of the Complement System in Human Viral Diseases. Biomolecules. 2022;12(2):226. [CrossRef]
- Soteros BM, Sia GM. Complement and microglia dependent synapse elimination in brain development. WIREs Mech Dis. 2022;14(3):e1545. [CrossRef]
- Huang L, Jin J, Chen K, et al. BDNF produced by cerebral microglia promotes cortical plasticity and pain hypersensitivity after peripheral nerve injury. PLOS Biol. 2021;19(7):e3001337. [CrossRef]
- Parkhurst CN, Yang G, Ninan I, et al. Microglia Promote Learning-Dependent Synapse Formation through Brain-Derived Neurotrophic Factor. Cell. 2013;155(7):1596-1609. [CrossRef]
- Smith JA, Das A, Butler JT, Ray SK, Banik NL. Estrogen or Estrogen Receptor Agonist Inhibits Lipopolysaccharide Induced Microglial Activation and Death. Neurochem Res. 2011;36(9):1587-1593. [CrossRef]
- Loiola RA, Wickstead ES, Solito E, McArthur S. Estrogen Promotes Pro-resolving Microglial Behavior and Phagocytic Cell Clearance Through the Actions of Annexin A1. Front Endocrinol. 2019;10:420. [CrossRef]
- Vegeto E, Bonincontro C, Pollio G, et al. Estrogen prevents the lipopolysaccharide-induced inflammatory response in microglia. J Neurosci Off J Soc Neurosci. 2001;21(6):1809-1818. [CrossRef]
- Herman JP, McKlveen JM, Ghosal S, et al. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr Physiol. 2016;6(2):603-621. [CrossRef]
- Tanaka J, Fujita H, Matsuda S, Toku K, Sakanaka M, Maeda N. Glucocorticoid- and mineralocorticoid receptors in microglial cells: The two receptors mediate differential effects of corticosteroids. Glia. 1997;20(1):23-37. [CrossRef]
- Chen K, Qi X, Zhu L lin, Li M li, Cong B, Li Y min. Quantitative analysis of microglia morphological changes in the hypothalamus of chronically stressed rats. Brain Res Bull. 2024;206:110861. [CrossRef]
- Frank MG, Thompson BM, Watkins LR, Maier SF. Glucocorticoids mediate stress-induced priming of microglial pro-inflammatory responses. Brain Behav Immun. 2012;26(2):337-345. [CrossRef]
- Nair A, Bonneau RH. Stress-induced elevation of glucocorticoids increases microglia proliferation through NMDA receptor activation. J Neuroimmunol. 2006;171(1-2):72-85. [CrossRef]
- Bollinger JL, Collins KE, Patel R, Wellman CL. Behavioral stress alters corticolimbic microglia in a sex- and brain region-specific manner. PLoS ONE. 2017;12(12):e0187631. [CrossRef]
- Bollinger JL, Bergeon Burns CM, Wellman CL. Differential effects of stress on microglial cell activation in male and female medial prefrontal cortex. Brain Behav Immun. 2016;52:88-97. [CrossRef]
- Gaspar R, Soares-Cunha C, Domingues AV, et al. The Duration of Stress Determines Sex Specificities in the Vulnerability to Depression and in the Morphologic Remodeling of Neurons and Microglia. Front Behav Neurosci. 2022;16. [CrossRef]
- Yu D, Jain S, Wangzhou A, et al. Microglia regulate GABAergic neurogenesis in prenatal human brain through IGF1. bioRxiv. Preprint posted online October 20, 2024:2024.10.19.619180. [CrossRef]
- Gervais É, Iloun P, Martianova E, Gonçalves Bessa AC, Rivest S, Topolnik L. Structural analysis of the microglia-interneuron interactions in the CA1 hippocampal area of the APP/PS1 mouse model of Alzheimer’s disease. J Comp Neurol. 2022;530(9):1423-1437. [CrossRef]
- Wu J, Zhang J, Chen X, et al. Microglial over-pruning of synapses during development in autism-associated SCN2A-deficient mice and human cerebral organoids. Mol Psychiatry. 2024;29(8):2424-2437. [CrossRef]
- Favuzzi E, Huang S, Saldi GA, et al. GABA-receptive microglia selectively sculpt developing inhibitory circuits. Cell. 2021;184(15):4048-4063.e32. [CrossRef]
- Chen JQA, McNamara NB, Engelenburg HJ, et al. Distinct transcriptional changes distinguish efficient and poor remyelination in multiple sclerosis. Brain J Neurol. 2025;148(6):2201-2217. [CrossRef]
- Zhang J, He H, Qiao Y, et al. Priming of microglia with IFN-γ impairs adult hippocampal neurogenesis and leads to depression-like behaviors and cognitive defects. Glia. 2020;68(12):2674-2692. [CrossRef]
- Cramer T, Gill R, Thirouin ZS, et al. Cross-talk between GABAergic postsynapse and microglia regulate synapse loss after brain ischemia. Sci Adv. 2022;8(9):eabj0112. [CrossRef]
- Paolicelli RC, Sierra A, Stevens B, et al. Microglia states and nomenclature: A field at its crossroads. Neuron. 2022;110(21):3458-3483. [CrossRef]
- Kuhn SA, van Landeghem FKH, Zacharias R, et al. Microglia express GABAB receptors to modulate interleukin release. Mol Cell Neurosci. 2004;25(2):312-322. [CrossRef]
- Rossano SM, Johnson AS, Smith A, et al. Microglia measured by TSPO PET are associated with Alzheimer’s disease pathology and mediate key steps in a disease progression model. Alzheimers Dement J Alzheimers Assoc. 2024;20(4):2397-2407. [CrossRef]
- Canário N, Jorge L, Martins R, Santana I, Castelo-Branco M. Dual PET-fMRI reveals a link between neuroinflammation, amyloid binding and compensatory task-related brain activity in Alzheimer’s disease. Commun Biol. 2022;5(1):804. [CrossRef]
- Da Silva T, Hafizi S, Rusjan PM, et al. GABA levels and TSPO expression in people at clinical high risk for psychosis and healthy volunteers: a PET-MRS study. J Psychiatry Neurosci JPN. 2019;44(2):111-119. [CrossRef]
Figure 1.
Emotion regulation network across stages of the menstrual cycle. During the late follicular phase, characterized by heightened levels of estrogen and low levels of progesterone hippocampal activity is increased, while amygdala activity is decreased, During the luteal phase, characterized by high levels of progesterone, dlPFC activity is increased.
Figure 1.
Emotion regulation network across stages of the menstrual cycle. During the late follicular phase, characterized by heightened levels of estrogen and low levels of progesterone hippocampal activity is increased, while amygdala activity is decreased, During the luteal phase, characterized by high levels of progesterone, dlPFC activity is increased.
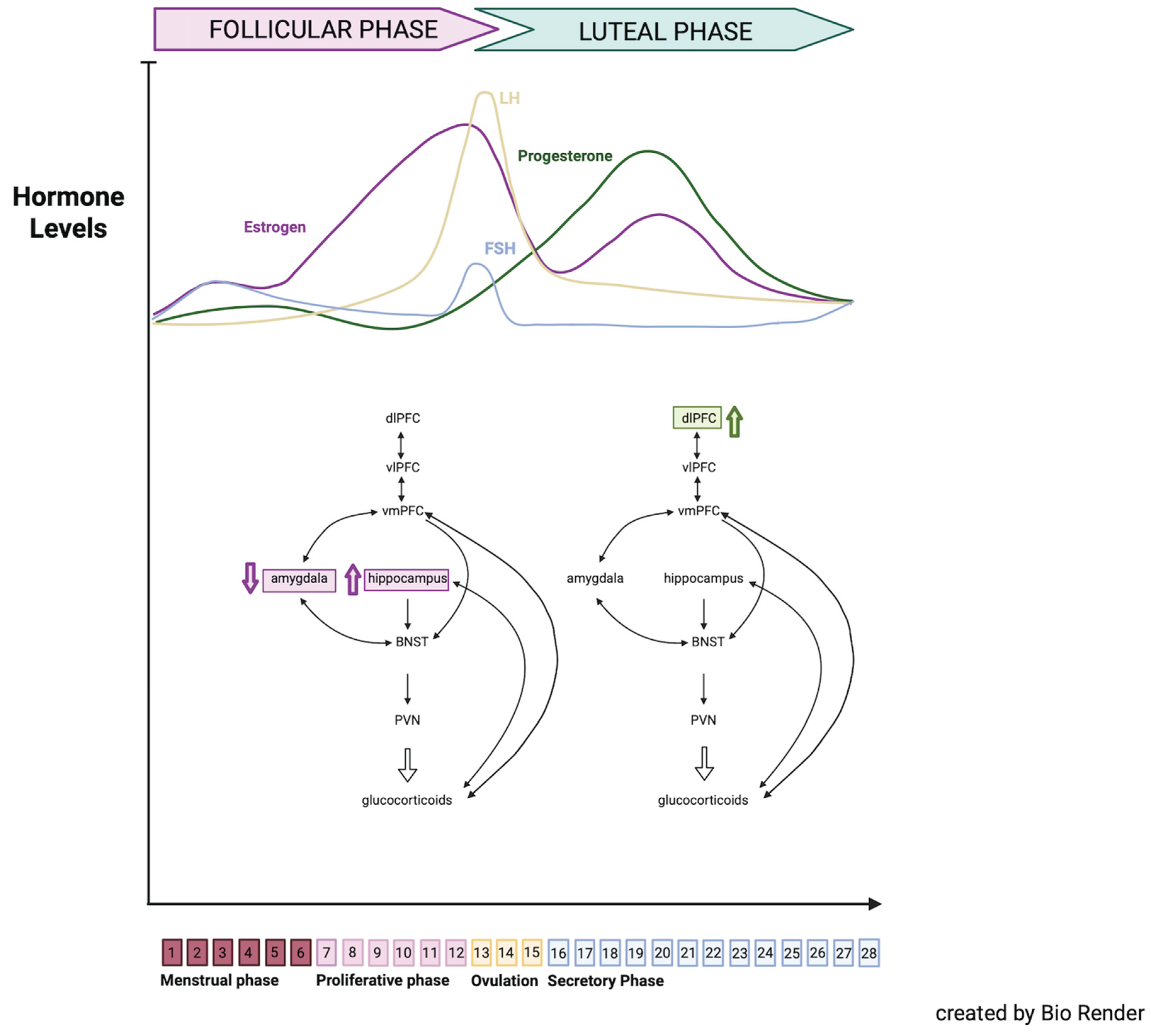
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



