Submitted:
11 September 2025
Posted:
11 September 2025
You are already at the latest version
Abstract
Lipid metabolism is a vital biological process essential for human health, encompassing key pathways necessary for the survival and homeostasis of all organisms. Liver X Recep-tors (LXRs) are extensively acknowledged as pivotal regulators of lipid homeostasis and inflammatory responses. Pharmacological activation of liver X receptor (LXR) has been shown to increase expression of ApoE and ABCA1 reducing neurodegeneration in murine models of Alzheimer´s disease. Because previous reports determined that Nectandra reticu-lata (Lauraceae) extract has agonistic LXRs activity, the objective of this study was to de-termine the metabolites present in this extract and to evaluate their in silico and in vitro agonistic activity. Identification was done by reversed phase (RP) ultra-high performance liquid chromatography (UHPLC) with a diode array detector (DAD) and high resolution mass spectrometry with electrospray ionization (ESI-HR-MS). In silico study was performed by Auto Dock Vina. The in vitro agonist activity was evaluated using real-time re-verse transcription-polymerase chain reaction (qRT-PCR) to determine the RNA expression of ApoE and ABCA1. The chromatographic analysis revealed the presence of three glycosylated flavonols: The in silico study showed that isolated flavonoids generate a hydrogen bond with T302 and T316 (LXRα and LXRβ respectively). The in vitro study showed that the flavonoids increased the expression of mRNA of both APOE and ABCA1 target genes of LXRs, as observed by qRT-PCR. The bioactive flavonoids isolated in this study possess documented antioxidant effect, when combined with their LXR agonist activity, they become promising bioactive candidates for use in nutraceutical formulations aimed at promoting brain health and anti-inflammatory effects.
Keywords:
liver X receptors
; inflammatory response
; Nectandra
; glycosylated flavonoids
; molecular docking
; agonistic activity
1. Introduction
Dementia is a clinical syndrome characterized by a progressive decline in cognitive functions, arising as a manifestation of various underlying pathologies, such as neurodegenerative diseases, where inflammatory processes can trigger or exacerbate them by increasing neuronal death that drives disease progression [1]. Alzheimer Diseases (AD) is the most common form of dementia and people with such disease have progressive decline in memory, thinking, communication skills, and learning capacity [2]. The histopathology of AD is related to both the injury and death of cholinergic neurons and the formation of protein aggregates [2]. Current evidence indicates that there is no a single pathway that produces this disease [3]. Different hypotheses have been proposed about the etiology of this pathology, each of them with a feature in common that involves factors that trigger the formation of two types of protein aggregates (neurofibrillary tangles and amyloid plaques). In particular, there is a hypothesis that postulates that the development of AD is associated with changes related to lipid regulation, since patients with this pathology have substantial increase in cholesterol levels [4]. Because of the complexity of the AD etiology multiple metabolic pathways are involved and then must be studied [5].
In recent years, some research has focused on an emerging target called Liver X receptors (LXRs). LXRs are ligand-activated transcription factors, which regulate the expression of genes involved in the flow and transport of lipids [6,7]. These receptors are related to the regulation of the ApoE and ABCA1, which not only helps to eliminate β-amyloid peptide (Aβ), but also modulate both cholesterol levels and the inhibition of amyloid precursor protein (APP) production [7]. This means that when the LXRs are activated by an agonist, they regulate the amount of cholesterol in the membrane and therefore reduce the production of Aβ with the help of ApoE and ABC1 modulation [6,7,8]. With this in mind, metabolites with LXRs agonist action may be candidates for more advanced studies if therapeutic options for AD are seek.
Previous studies, carried out by our research group have shown LXRs agonist activity of Nectandra reticulata leaf extract [9]. We already reported the presence of quercitrin in a N. reticulata leaf extract, compound that was identified by means of the comparison of retention time and UV-vis maxima of a chromatographic signal against those of an authentic standard [10]. In the mentioned work neither further identification strategies were attempted for quercitrin nor for other metabolites. Besides that, report, no additional information on the metabolites of this species is currently available; however, studies on metabolites have been conducted for other species within the genus. Some of the metabolites described for the Nectandra genus are flavonoids, lignoids, phenylpropanoids, steroids, terpenoids, and alkaloids [11,12]. Many of them have activities such as anti-inflammatory, anti-fatty liver disease, and antioxidant.
Quercitrin affects positively important biological pathways in the progression of AD, being widely documented the antioxidant and free radical scavenger activity [13], together with its capability of inhibiting acetylcholinesterase activity [14]. Besides that, quercitrin reduces the accumulation of amyloid-β plaques in AD model mice [15]. Considering that the leaf extract of N. reticulata has LXRs agonist activity and that its metabolic profile has not fully been characterized, we conducted this research with the aim of confirming the identity of quercitrin and identifying other metabolites present in an ethanolic extract of N. reticulata, by reversed phase-ultra high resolution liquid chromatography (RP-UHPLC) coupled to a diode array detector (DAD) and to a high resolution mass spectrometer detector with an electrospray ionization source (ESI-HR-MS/MS). To determine the agonist effect of the identified compounds we carried out in silico studies, based on molecular docking, to explain the LXRs agonist activity. Furthermore, we followed the ApoE and ABCA1 mRNA expression, both genes direct transcriptional targets of LXRα and LXRβ.
2. Results and Discussion
In this research, an ethanolic extract from N. reticulata was analyzed to identify its most abundant compounds and to test them for in silico LXRs agonist activity, as well as them in vitro agonist activity of both APOE and ABCA1 target genes of LXRs. Three compounds were found as the main representatives: quercitrin (quercetin-3-O-rhamnoside), afzelin (kaempferol-3-O-rhamnoside), and either kaempferol-3-O-(6''-p-coumarylglucoside) or kaempferol-7-O-(6''-p-coumarylglucoside). Despite high resolution mass spectrometry data were crucial to annotate such compounds, acidic hydrolysis was useful to rule out if e.g. kaempferol or luteolin were part of a compound, since both have the same molecular weight. When doing such acidic hydrolysis, a special emphasis was placed to find the conditions in which the compound was successfully hydrolyzed while ensuring the released constituents remained stable and soluble [16]. Furthermore, the identified compounds were found to have LXRs agonistic activity as shown not only by the degree of the interaction between them and the amino acids threonine 302 and threonine 316 (LXRα and LXRβ, respectively) but also because of the increased expression of mRNA of both APOE and ABCA1 target genes of LXRs as induced by the presence of such isolated compounds.
2.1. Identification of the Metabolites
The chromatographic profile of N. reticulata crude extract showed three compounds with characteristic UV-vis spectra of glycosylated flavonoids (Table 1) [17].
Those three compounds accounted for 99.35% (Figure 1) of the chromatographic area recorded at 355 nm. UV-vis spectra of flavonoids are typically characterized by two absorption maxima (this is, Band II and Band I) [18]. Depending on the wavelength of the maxima, different types of flavonoids can be differentiated. The maximum UV observed for compounds 1-3 at 260-268 and at 314-349 (Table 1) is characteristic of either flavones or flavonols [19]. From literature it is known that these types of flavonoids have Band I between 300-380 nm and Band II between 240-280 nm, with hypsochromic shifts if they are either glycosylated or methylated [18]. Additionally, the molecular ions of the compounds 1-3 ([M+H]+ or [M-H]-) together with the daughter ions can be found in the same Table 1. Taking into account that the molecular mass of flavones or flavonols ranges from 220 to 301 a.m.u. [20], the greater masses observed for compounds 1-3 are indicative of flavones or flavonols being glycosylated or with substitution patterns in their structure.
Compound 1, with a retention time of 7.80 min, was the most abundant with a relative area of 84.46% (Figure 1).
This compound had two absorbance maxima at 260 and 349 nm and showed a molecular ion in the negative ion mode at m/z 447.0936 ([M-H]-) (Table 1). The fragmentation of the molecular ion peak showed the detachment of a rhamnosyl fragment, thus yielding an aglycone at m/z 300.0273, which is likely to be a product of an homolytic fragmentation of the parent ion. In the positive ion mode, no signal corresponding to the molecular ion was observed, instead a signal corresponding to the aglycone was present at m/z 303.0509 [M-rhamnose+H]+. These signals suggest that compound 1 has quercetin as a flavonol in its structure. When the N. reticulata crude extract was hydrolyzed, the aglicone quercetin appeared as the most abundant reaction product (Rt at 12.29 min, UVmax at 257 and 368 nm; chromatographic and spectroscopic data coincident to those of authentic quercetin) (Figure 2 and Table 2). It was concluded, therefore, that compound 1 corresponded to quercitrin (quercetin-3-O-rhamnoside) (Figure 3a).
Compound 2 in the crude extract of N. reticulata was found at a retention time of 9.44 min, with absorbance maxima at 260 and 341 nm, and relative area of 8.00% (Figure 1). It showed a molecular ion peak at m/z 431.0980 in the negative ion mode. Fragmentation of that parent ion yielded an ion at m/z 284.0323, as a result of a loss of rhamnose ([M-rhamnose-2H]-●) [21]. In contrast, in the positive ion mode, the molecular ion peak was not found, only the loss of a sugar moiety to yield the aglycone at m/z 287.0561 [M-rhamnose+H]+ was observed. The ion at m/z 284.0323 in the negative ion mode, together with the ion in the positive mode at m/z 287.0561, and the UV maxima are indicative of the presence of either kaempferol or luteolin in the structure of compound 2. When hydrolyzing the crude extract, the aglycone kaempferol was found (Retention time and UV maxima of the released aglycone equal to those of authentic kaempferol). Therefore, it was concluded that compound 2 corresponded to afzelin (kaempferol-3-O-rhamnoside). Both compounds 1 and 2 have been previously reported in other species of the Nectandra genus, such as N. glabrescens, N. grandiflora, and N. amazonum [21,22,23,24]. Furthermore, compound 1 was previously reported by us in an extract of N. reticulata [10].
Compound 3 had a retention time of 14.26 min, UV-VIS maxima at 268 and 314 nm, and relative area of 6.89% (Figure 1). This compound, in the positive ion mode, exhibited a signal of the molecular ion peak and a loss of a coumaroyl glucoside at m/z 595.1464 [M+H]+ and m/z 287.0559 [M-coumaryl glucoside+H]+, respectively. In the negative ion mode, compound 3 revealed the ions at m/z 593.1296 [M-H]- and at m/z 285.0396 [M-coumaryl glucoside-H]-. Additionally, in positive ion mode, it yielded a signal at m/z 309.0981 [M-kaempferol+H]+ that is common in metabolites that have a coumaroyl glucoside decoration in their structure [25]. Daughter ions, in both negative and positive ion mode, showed that there is a flavonoid with a mass of 286 a.m.u. in the structure. As mentioned above, the only possible aglycone (product of the hydrolysis reaction) is kaempferol. Furthermore, the presence of p-coumaric acid was verified in the hydrolyzed extract by comparison with the authentic standard. With these analysis conditions, however, it was not possible to state if the p-coumaroyl glucoside substitution is in the 3- or 7- position of the flavonoid scaffold. Studies of the genus Nectandra have shown the presence of some kampferol derivatives, glycosilated in position 3- such as kaempferol-3-O-α-rhamnopyranoside and kaempferol-3-O-α-(3,4-di-E-p-coumaroyl)-rhamnopyranoside [26]. Some studies of the Lauraceae family indicate that most of the elucidated compounds have the substitution in the position 3- of the flavonoid nucleus, in fact, the flavonoids quercetin and kaempferol are the most common flavonoids [11,12]. If the biosynthesis of glycosylated flavonoids in plants is considered, it is known that the most common substitution is also at the carbon 3 [27,28]. It is also known that there are enzymes, such as UDP-flavonoid-3-O-glucosyltransferase that exhibit strict regio-selectivity for the position 3 [30]. This information together allows us to conclude that compound 3 is more likely to be kaempferol-3-O-(6''-p-coumarylglucoside) than kaempferol-7-O-(6''-p-coumarylglucoside).
2.2. Evaluation of Hydrolysis Conditions.
Identification presented in the previous section required the confirmation of the aglycones by hydrolysis. Acidic hydrolysis of the crude extract allowed us not only to identify the aglycones of the glycosylated flavonoids present in the mixture, but also revealed an important effect of the composition of the hydrolysis solvent mixture. To confirm the presence of quercetin, p-coumaric acid, and to rule out the presence of luteolin or kaempferol, authentic standards were analyzed under the same chromatographic conditions that the crude extract and the hydrolysates were studied.
When hydrolysis was performed with either 50 or 75% (v/v) methanol, a complete hydrolysis of compound 1 was not observed (Figure 2). Instead, when done at 80% (v/v) methanol compound 1 was fully hydrolyzed. Furthermore, while compound 2 was not fully hydrolyzed at either of the tested methanol concentrations, compound 3 was hydrolyzed the most at 50% (v/v) methanol. As a result of the hydrolysis process, the area of the released quercetin and kaempferol increased when using 80% (v/v) methanol. In contrast, a greater chromatographic area was found for p-coumaric acid when the hydrolysis was carried out with 50% (v/v) methanol. Under the tested conditions no luteolin was observed. We hypothesize that the different chromatographic area of quercetin, kaempferol, and p-coumaric acid as result of varying the percentage of methanol is the result of not only the extent of the hydrolysis but also of differences on solubility of the released compounds under the hydrolysis.
2.3. Molecular Docking into the Agonist Binding Sites of LXRα and LXRβ Receptors.
Previous studies reported the agonist activity of different flavonoids against LXRs, including cyanidin [29], quercetin, and apigenin [17,30]. A previous detailed study indicates that the hydrogen bond with threonine 302 and threonine 316 (LXRα and LXRβ respectively) would be involved in the agonist activity of the studied compounds. Because of that, re-docking was performed with the protein data bank (PDB) files for hLXRα-LBD (3IPQ) and hLXRβ-LBD (1PQ6) and the crystal compound GW3965, to ensure that the conditions of the grid box and settings were able to reproduce the binding modes as reported in the literature [17].
Likewise, molecular docking of the natural flavonoids, quercetin and apigenin, was carried out. As expected from the work by Fouache et al [17], an effective hydrogen bond for both LXRα and LXRβ was observed. After this, the binding modes of the compounds identified in the crude extract of N. reticulata were predicted. Those binding modes with the lowest energy are presented (Figure 4). Because it was not possible to determine the substitution of the p-coumaroyl glucoside in compound 3, both possibilities [kaempferol-3-O-(6''-p-coumarylglucoside) and kaempferol-7-O-(6''-p-coumarylglucoside)] were tested. In Figure 4, the hydrogen bond related to the agonist activity of the elucidated compounds with T302 and T316 can be observed. Quercitrin, afzelin, and kaempferol-7-O-(6''-p-coumarylglucoside) show a hydrogen bond with LXRα and LXRβ with less than 3.0 Å, this hydrogen bond with T302 and T316 has been linked to agonist activity. This distance of the hydrogen bond is a criterion to define the strength of interaction and the ligand-receptor affinity [31]. Kaempferol 3-O-(6''-p-coumarylglucoside) shows a hydrogen bond with a distance lower than 3.0 Å with LXRβ, but greater than 3.0 Å with LXRα, which means that the interaction between such kaempferol-3-O-glycoside and the LXRα is not enough to state that a hydrogen bond occurs [31]. To sum up, there is a relationship between the agonist activity of the extract and the flavonoid-type compounds identified in it.
2.4. In Vitro Upregulation of LXRs Target Genes by N. reticulata Extracted Compounds.
The treatment with 2 μM GW3965 (synthetic agonist of LXRs) increased the mRNA expression of both ApoE (P < 0.001) and ABCA1 (P < 0.0001) as compared to the vehicle treatment (Figure 5). Regarding to the compounds of interest, it was found that 2 μM quercitrin (compound 1) increased the expression of both ApoE (P < 0.0001) and ABCA1 (P < 0.0001) as compared to the vehicle treatment (Figure 5). In addition, 5 μM quercetin (the hydrolyzed product of quercitrin) also increased the expression of both ApoE (P < 0.05) and ABCA1 (P < 0.05) (Figure 5). These results suggest that quercitrin is more active than its hydrolyzed counterpart, quercetin, suggesting that its glycosylation promotes its agonist activity. Regarding to afzelin (compound 2) and its hydrolyzed product kaempferol, 2 μM of afzelin increased the expression of ABCA1 (P<0.01) without significantly changing ApoE levels as compared to the vehicle. In contrast, 2 μM kaempferol increased the expression of both ApoE (P < 0.0001) and ABCA1 (P < 0.001) (Figure 5), suggesting that kaempferol is more active in the absence of the glycosylated group.
3. Material and Methods
3.1. Chemicals and Standards.
Luteolin was purchased from Fluka® Analytical (France); both quercitrin (quercetin-3-O-rhamnoside) and kaempferol (China), p-coumaric acid (United Kingdom), and quercetin (India) were obtained from SIGMA. Afzelin (kaempferol-3-O-rhamnoside) was obtained from the extract of N. reticulata by traditional chromatographic techniques. All those standards, in solid state, had more than 95% (w/w) purity. Methanol and acetonitrile, both HPLC grade, were purchased from Honeywell (Muskegon, Michigan, USA). LiChropur HPLC-MS grade formic acid was purchased from Merck (Darmstadt, Germany). Supelco LiChrosolv HPLC-MS grade acetonitrile was purchased from Sigma-Aldrich (Darmstadt, Germany).
3.2. Plant Material and Extraction.
N. reticulata leaves were collected in Granada (Cundinamarca, Colombia), 31 km from Bogotá D.C. A specimen of the species was deposited and identified by the Herbario Nacional de Colombia at Universidad Nacional de Colombia whose code is COL547368. Subsequently, leaves were dried at room temperature, ground to a particle size lower than 2 mm, extracted by percolation with 96% (v/v) aqueous ethanol, and rotavaporated at 40 °C.
3.3. Acid Hydrolysis of the Plant Extract.
The plant extract (50 mg) was hydrolyzed by reflux at 80 °C for 2 h, using 5 mL of 1.2 M HCl and 2 mg of ascorbic acid in a hydro alcoholic mixture of methanol [32]. In addition, solubility tests were carried out modifying the proportion of methanol in the reaction solvent, testing aqueous solutions of methanol at 50, 75 or 80% (v/v). After the reaction time was completed, the mixture was cooled to room temperature, the pH was set to 4-6, the product was dried and re-suspended up to 10.0 mL with 100% (v/v) methanol in a volumetric flask. Finally, the hydrolysis reaction product was filtered using a 0.45 µm filter and analyzed by RP-UHPLC-DAD.
3.4. Identification of Metabolites
The samples (crude extract and hydrolysis reaction products) were analyzed on a Dionex Ultimate 3000 system (Thermo Scientific, San Jose, California, USA). The system was equipped with a pump, an autosampler, and a photodiode array detector (DAD). The stationary phase was a Hypersyl Gold RP column (Thermo Scientific; 150 mm x 2.1 mm id; 1.9 µm particle size), operated at 40 °C. The eluents were water/formic acid (99.9/0.1 v/v) (eluent A) and acetonitrile (eluent B). The elution program was 0-4 min, linear gradient from 5 to 20% B; 4-10 min, isocratic at 20% B; 10-12 min, linear gradient from 20 to 25% B; 12-16 min, isocratic at 25% B; 16-22 min, linear gradient from 25 to 52% B; 22-26 min, isocratic at 52% B; 26-30 min, linear gradient from 52 to 70% B; 30-33 min, linear gradient from 70 to 100% B; 33-36 min, isocratic at 100% B; 36-41 min, linear gradient from 100 to 5% B; 41-46 min, isocratic at 5% B. The flow rate was 400 μL/min with an injection volume of 5 μL.
In addition, the crude extract was analyzed on an Agilent Technologies 1260 liquid chromatography system coupled to a Q-TOF 6545 quadrupole time-of-flight mass analyzer with electrospray ionization. For the chromatographic separation, the same conditions of the previously described program were used, only the detection system was changed. Mass spectrometry detection was performed in full scan in both positive and negative ESI mode from 100 to 1,100 m/z, with a source voltage of 3,000 V, and an ion transfer tube temperature of 290 °C.
The identification was based on the analysis of retention time and the elution order, UV-Vis spectra, HR-MS/MS data, reaction products after acidic hydrolysis, comparison of chromatographic, spectroscopic, and spectrometric features of chromatographic peaks with standards, and comparison with the literature.
3.5. Molecular Docking.
The identified compounds were tested for their ability to bind to either LXRα or LXRβ by molecular docking, using AutoDock Vina (ADV, v.1.2). The 3D-structure of LXRα and LXRβ (PDB ID: 1PQ6 and 3IPQ, respectively) was obtained from the Protein Data Bank. The pdbqt files were prepared by removing both water molecules and all ligands included in the crystal and only polar hydrogen atoms were set. Visualization of the binding mode predictions were performed with UCSF Chimera 1.15 molecular viewer. Next, a docking grid box with a size of 25 × 25 × 25 Å was created. A molecular re-docking of GW3965 into LXRα and LXRβ (1PQ6 and 3IPQ) was performed to validate the methodology previously described. In addition, a molecular docking of the active compounds quercetin and apigenin was conducted as part of the validation studies [33]. Subsequently, molecular docking analysis was conducted on the molecules identified from the extract and interactions between ligands and nuclear receptors were analyzed.
3.6. In Vitro Upregulation of LXRs Target Genes by N. reticulata Extracted Compounds
To corroborate the LXR agonistic action of pure compounds extracted from N. reticulata we evaluated the expression of both ApoE and ABCA1 genes in the U87 cell line after treatment with the control (vehicle), the synthetic agonist of LXRs (GW3965) as positive control, and compounds from N. reticulata. Induction of ApoE mRNA in U87 cells has been used previously to research nuclear receptor activation [18].
3.7. Cell Culture and Viability.
Human U87 cell line (human glioma) was grown in Dulbecco's Modified Eagle's Medium (DMEM; Invitrogen) supplemented with 10% fetal bovine serum (FBS, Biowest.) and 1% (w/v) penicillin–streptomycin (Santa Cruz Biotechnology, Inc.) in a humidified atmosphere at 37 °C in 5% CO2.
3.8. Real-Time Reverse Transcription-Polymerase Chain Reaction (qRT-PCR).
To monitor the in vitro gene expression of LXRs target genes, qRT-PCR was performed. Briefly, total RNA from U87 cells was extracted using the RNA easy Kit (Qiagen) according to the manufacturer’s instructions. The RNA was quantified using the NanoDropTM 2000 (Thermo Scientific, San Jose, California, USA). For RNA detection and quantitation, the Luna® Universal One-Step qRT-PCR kit (E3005, New England Biolabs) was used, according to the manufacturer’s protocol. Primer pairs (Microgen, South Korea) for APOE (forward CCTCAAGAGCTGGTTCGAG, reverse TCGGCGTTCAGTGATTGTC), ABCA1 (forward GCTTTCAATCATCCCCTGAA, reverse CAGGTGTT TGCTTTGCTGA), and GADPF (forward GACCTGCCGTCTAGAAAAACC, reverse ACCACCTGGTGCTCAGTGTAG) were designed to hybridize to unique regions of the appropriate gene sequence. Statistical significance was calculated by a one-way ANOVA, and the data are presented as means + standard error of mean (SEM, n = 3).
4. Conclusions
We report the flavonoid composition of an ethanolic extract of Nectandra reticulata. The main compound corresponds to quercitrin, the other two compounds are afzelin and either kaempferol 3-O- or 7-O-(6''-p-coumaryl glucoside). By molecular docking, it was possible to identify that quercitrin, afzelin, and kaempferol-7-O-(6''-p-coumarylglucoside) are capable of binding to the amino acids threonine 302 and threonine 316 (LXRα and LXRβ respectively), which are related to the LXRs agonist activity. The studied flavonoids increased the expression of mRNA of both APOE and ABCA1 target genes of LXRs, as observed by qRT-PCR. All in all, the identified compounds could be related to the agonistic activity of the ethanolic extract of N. reticulata. It would be important to evaluate the activity of these isolated compounds and verify whether they have antagonistic, additive, or synergistic interaction. The isolated flavonoids, identified as LXR agonists, are capable of modulating lipid homeostasis and regulating inflammatory processes. In conjunction with their demonstrated antioxidant activity, they may serve as suitable agents for incorporation into nutraceuticals or dietary supplements.
Author Contributions
J.P.T: Investigation (lead); Methodology (lead); Data curation (equal). F.L.V: Supervision (equal); Data curation (equal). A.S.H: Supervision (equal); Data curation (equal) Funding acquisition (equal); Project administration (equal). C.E.N.C: Supervision (equal); Data curation (equal); Writing-review; editing (equal). M.A.M: Conceptualization (equal); Funding acquisition (equal); Project administration (equal); Supervision (equal); Writing-review; editing (equal). Furthermore, all data were generated in-house, and no paper mill was used. All authors agree to be accountable for all aspects of work ensuring integrity and accuracy.
Funding
This work was supported by the Ministerio de Ciencia, Tecnología e Innovación, Colombia [grants number 727-2018 and 734-2018].
Data Availability Statement
The underlying data supporting the findings of this study are available from the authors upon reasonable request.
Acknowledgments
The authors thanks Ministerio de Ciencia, tecnología e Innovación, Colombia for financial support, and Ministerio de Ambiente y Desarrollo Sostenible by Permission No. 121 of January 22, 2016 (modification Otrosí No. 21) and permission request to access genetic resources and their derivative products No. RGE 46 (Article 6, Law 1955 2019).
Conflicts of Interest
There are no conflicts to declare.
Abbreviations
The following abbreviations are used in this manuscript:
| LXRs | Liver X Receptors |
| ApoE | Apolipoprotein E |
| ABCA1 | ABC transporter protein member 1 |
| AD | Alzheimer Diseases |
| RP | Reversed Phase |
| UHPLC | Ultra High Performance Liquid Chromatography |
| DAD | Diode array detector |
| ESI-HR-MS | High Resolution Mass Spectrometry with Electrospray ionization |
| qRT-PCR | Real Time Reverse Transcription Polymerase Chain Reaction |
| RNA | Ribonucleic acid |
| mRNA | Messenger Ribonucleic Acid |
| LXRα | Liver X Receptors alpha |
| LXRβ | Liver X Receptors beta |
| a.m.u. | Atomic Mass Unit |
| HPLC | High Performance Liquid Chromatography |
| HCl | Hydrocloryc acid |
| Q-TOF | Quadrupole Time of Flight |
References
- World Health Organization, Dementia. https://www.who.int/news-room/fact-sheets/detail/dementia, (accessed November 16, 2024).
- B. Duthey, A public health approach to innovation, 2013, 6, 74. https://www.medbox.org/pdf/5e148832db60a2044c2d5489, (accessed January 9, 2025).
- M. Calabrò, C. Rinaldi, G. Santoro and C. Crisafulli, AIMS Neurosci., 2020, 8, 86–132. [CrossRef]
- E. Mohandas, V. Rajmohan and B. Raghunath, Indian J. Psychiatry, 2009, 51, 55–61. [CrossRef]
- A. G. Sandoval-Hernández, L. Buitrago, H. Moreno, G. P. Cardona-Gómez and G. Arboleda, PLoS One, 2015, 10, e0145467. [CrossRef]
- D. R. Riddell, H. Zhou, T. A. Comery, E. Kouranova, C. F. Lo, H. K. Warwick, R. H. Ring, Y. Kirksey, S. Aschmies, J. Xu, K. Kubek, W. D. Hirst, C. Gonzales, Y. Chen, E. Murphy, S. Leonard, D. Vasylyev, A. Oganesian, R. L. Martone, M. N. Pangalos, P. H. Reinhart, and J. S. Jacobsen, Mol. Cell. Neurosci., 2007, 34, 621–628. [CrossRef]
- R. K. Sodhi and N. Singh, Pharmacol. Res., 2013, 72, 45–51. [CrossRef]
- R. Koldamova and I. Lefterov, Current Alzheimer Research, 2007, 4, 171–178. [CrossRef]
- A. Bustos-Rangel, J. Muñoz-Cabrera, L. Cuca, G. Arboleda, M. Ávila-Murillo, and A.G. Sandoval-Hernández, Front. Nat. Prod, 2023, 2, 1169182. [CrossRef]
- J. Pulido-Teuta, C.E. Narváez-Cuenca, and M. Avila-Murillo, RSC Adv., 2024,14, 21874-21886. [CrossRef]
- 11.S. S. Grecco, H. Lorenzi, A. G. Tempone and J. H. G. Lago, Tetrahedron: Asymmetry, 2016, 27, 793–810. [CrossRef]
- V. E. Macías-Villamizar, L. E. Cuca-Suárez, and E. D. Coy-Barrera, Bol. Latinoam. y del Caribe de Plantas Med. y Aromáticas, 2015, 14, 317-342, URL: https://www.redalyc.org/articulo.oa?id=85641104007.
- S. Rattanajarasroj and S. Unchern, Neurochem Res., 2010, 35, 1196–1205. [CrossRef]
- G. Rajamanickam and S.L. Manju, Research Square, 2022. [CrossRef]
- L. Wang, J. Sun, Z. Miao, X. Jiang, Y. Zheng, and G. Yang, NeuroReport, 2022, 33, 327-335. [CrossRef]
- T. C. Tolosa, H. Rogez, E. M. Silva and J. N. S. Souza, J. Braz. Chem. Soc., 2018, 29, 2475–2481. [CrossRef]
- M. Taniguchi, C. A. LaRocca, J. D. Bernat and J. S. Lindsey, J. Nat. Prod., 2023, 86, 1087–1119. [CrossRef]
- T. J. Mabry, K. R. Markham and M. B. Thomas, in The Systematic Identification of Flavonoids, eds. T. J. Mabry, K. R. Markham and M. B. Thomas, Springer, Berlin, Heidelberg, 1970, pp. 41–164.
- J. Greenham, J.B. Harborne, and C.A. Williams, Phytochem. Anal., 2003, 14,100-118. [CrossRef]
- N. Fabre, I. Rustan, E. de Hoffmann and J. Quetin-Leclercq, J. Am. Soc. Mass Spectrom., 2001, 12, 707–715. [CrossRef]
- S. Kazuno, M. Yanagida, N. Shindo and K. Murayama, Anal. Biochem., 2005, 347, 182–192. [CrossRef]
- J. M. Barbosa-Filho, M. Yoshida and O. R. Gottlieb, Phytochem., 1989, 28, 2209-221. [CrossRef]
- F. R. Garcez, W. S. Garcez, M. Martins and A. C. Cruz, Planta Med., 1999, 65, 775. [CrossRef]
- A. B. Ribeiro, V. da S. Bolzani, M. Yoshida, L. S. Santos, M. N. Eberlin and D. H. S. Silva, J. Braz. Chem. Soc., 2005, 16, 526–530. [CrossRef]
- D. F. Felipe, L. Z. S. Brambilla, C. Porto, E. J. Pilau and D. A. G. Cortez, Molecules, 2014, 19, 15720–15734. [CrossRef]
- G. A. Conserva, T. A. Costa-Silva, L. M. Quirós-Guerrero, L. Marcourt, J.-L. Wolfender, E. F. Queiroz, A. G. Tempone and J. H. G. Lago, Chem-Biol. Interact., 2021, 349, 109661. [CrossRef]
- J. Xiao, T. S. Muzashvili and M. I. Georgiev, Biotechnol. Adv, 2014, 32, 1145–1156. [CrossRef]
- R. Yin, B. Messner, T. Faus-Kessler, T. Hoffmann, W. Schwab, M.-R. Hajirezaei, V. von Saint Paul, W. Heller and A. R. Schäffner, J. Exp. Bot., 2012, 63, 2465–2478. [CrossRef]
- Y. Jia, M. H. Hoang, H.-J. Jun, J. H. Lee and S.-J. Lee, Bioorg. & Med. Chem. Lett., 2013, 23, 4185–4190. [CrossRef]
- Y. Hu, Y. Yang, Y. Yu, G. Wen, N. Shang, W. Zhuang, D. Lu, B. Zhou, B. Liang, X. Yue, F. Li, J. Du and X. Bu, J. Med. Chem., 2013, 56, 6033–6053. [CrossRef]
- T. K. Harris and A. S. Mildvan, Proteins: Struct., Funct., and Bioinform., 1999, 35, 275–282. [CrossRef]
- A. M. Nuutila, K. Kammiovirta, and K.-M. Oksman-Caldentey, Food Chem., 2002, 76, 519–525. [CrossRef]
- Fouache, N. Zabaiou, C. De Joussineau, L. Morel, S. Silvente-Poirot, A. Namsi, G. Lizard, M. Poirot, M. Makishima, S. Baron, J.-M. A. Lobaccaro and A. Trousson, J. Steroid Biochem. Mol. Biol., 2019, 190, 173–182. [CrossRef]
- S. Mallick, P. A. Marshall, C. E. Wagner, M. C. Heck, Z. L. Sabir, M. S. Sabir, C. M. Dussik, A. Grozic, I. Kaneko and P. W. Jurutka, ACS Chem. Neurosci., 2021, 12, 857–871. [CrossRef]
Figure 1.
Chromatographic profile of the ethanolic extract of Nectandra reticulata as recorded at 355 nm.
Figure 1.
Chromatographic profile of the ethanolic extract of Nectandra reticulata as recorded at 355 nm.
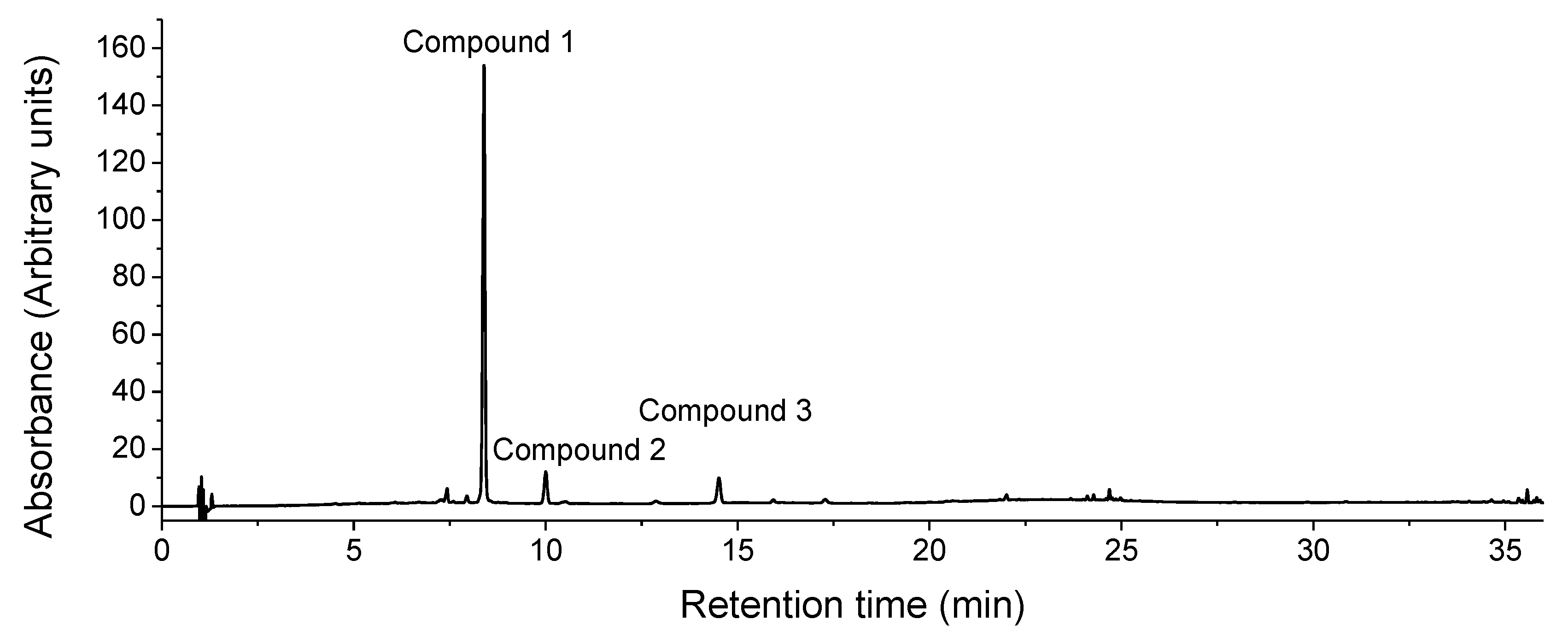
Figure 2.
Chromatographic profile of the different hydrolysates as recorded at 310 nm between 0 and 20 min. (A) Non-hydrolyzed extract. (B) Hydrolysis at 80% (v/v) methanol. (C) Hydrolysis at 75% (v/v) methanol. (D) Hydrolysis at 50% (v/v) methanol.
Figure 2.
Chromatographic profile of the different hydrolysates as recorded at 310 nm between 0 and 20 min. (A) Non-hydrolyzed extract. (B) Hydrolysis at 80% (v/v) methanol. (C) Hydrolysis at 75% (v/v) methanol. (D) Hydrolysis at 50% (v/v) methanol.
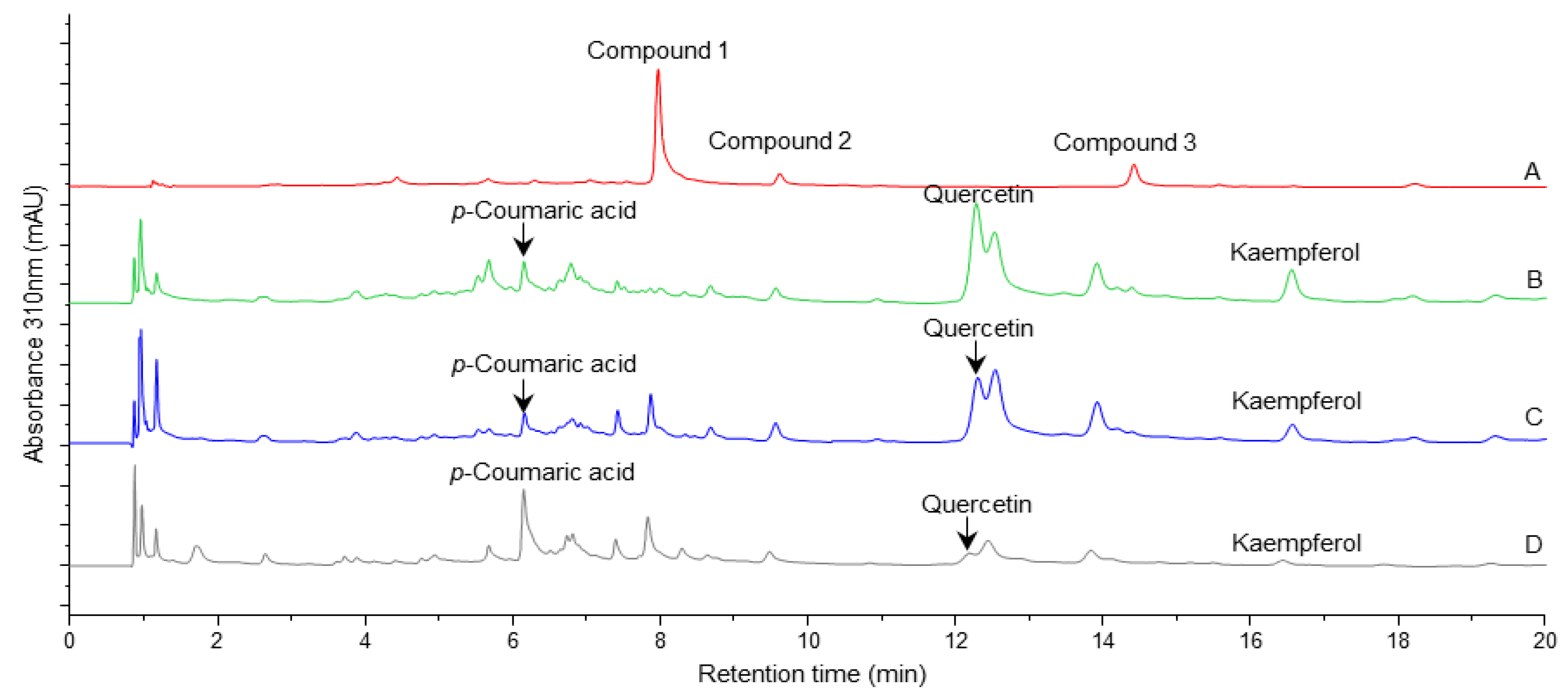
Figure 3.
Chemical structure of the marker compounds (A) quercitrin [quercetin-3-rhamnoside]. (B) afzelin [(kaempferol-3-O-rhamnoside]. (C) kaempferol-3-O-(6''-p-coumarylglucoside) or kaempferol-7-O-(6''-p-coumarylglucoside).
Figure 3.
Chemical structure of the marker compounds (A) quercitrin [quercetin-3-rhamnoside]. (B) afzelin [(kaempferol-3-O-rhamnoside]. (C) kaempferol-3-O-(6''-p-coumarylglucoside) or kaempferol-7-O-(6''-p-coumarylglucoside).
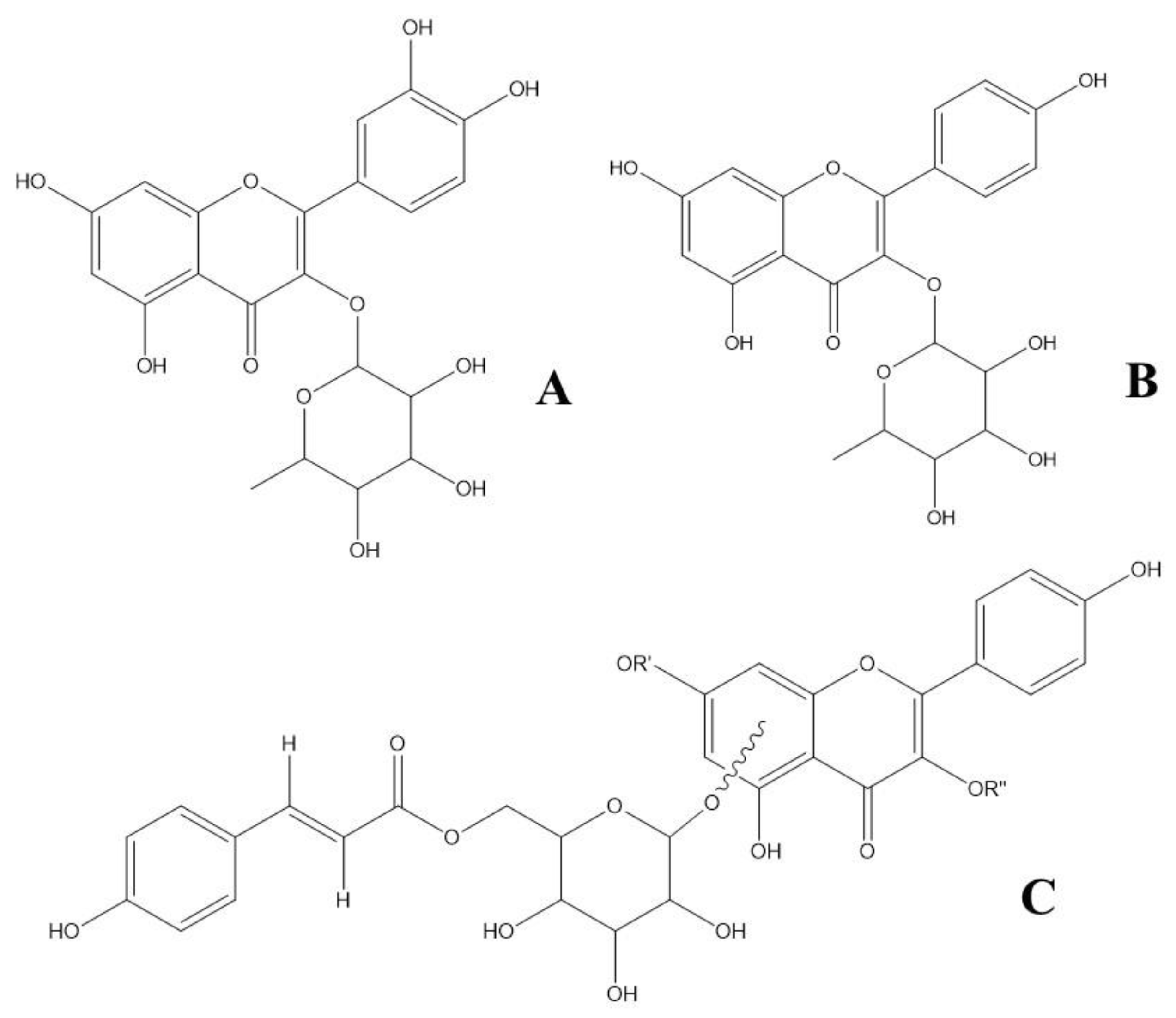
Figure 4.
Three-dimensional representation of flavonoids in the LXRα or LXRβ ligand-binding pocket, analyzing the hydrogen bond with T302 or T316, respectively.
Figure 4.
Three-dimensional representation of flavonoids in the LXRα or LXRβ ligand-binding pocket, analyzing the hydrogen bond with T302 or T316, respectively.
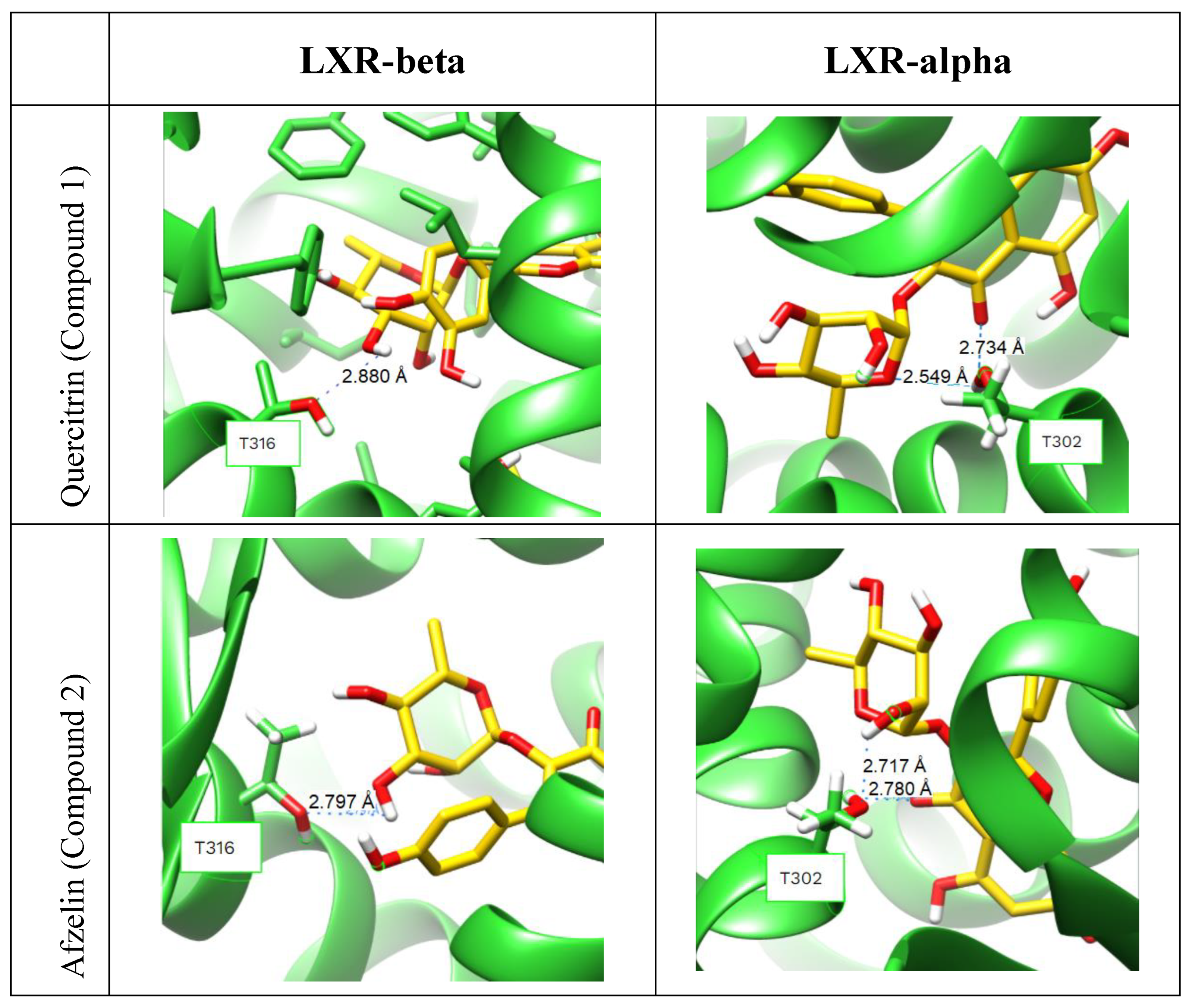
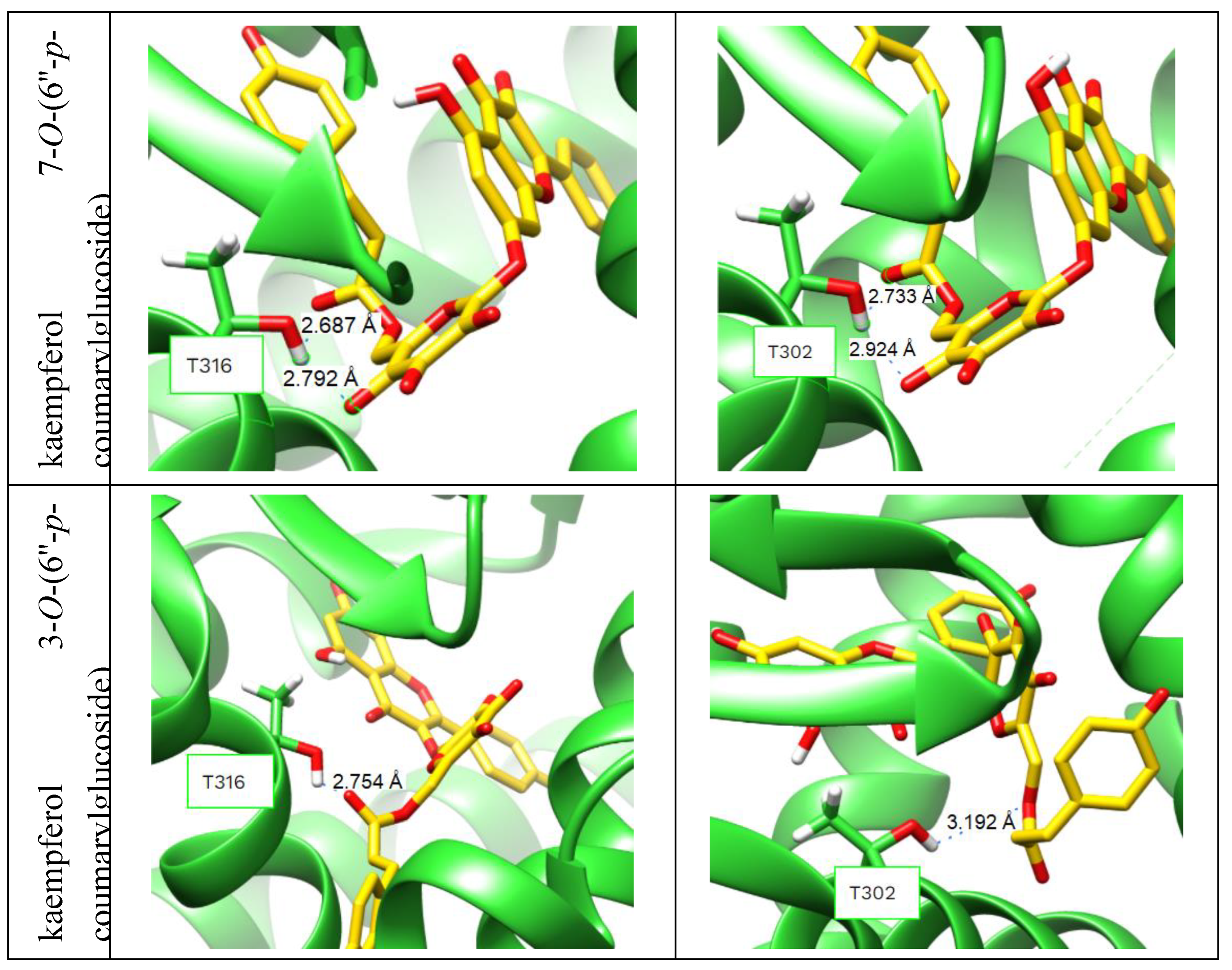
Figure 5.
ABCA1 and ApoE mRNA levels. U87 cells were treated with a control (vehicle) and the flavonoid compounds. qRT-PCR were used to examine the mRNA levels and fold-changes was calculated by the 2-ΔΔCt method as compared with non-infection cell control and using endogenous GAPDH mRNA level for normalization. (A) The fold change of ABCA1 in mRNA levels; (B) The fold change of ApoE in mRNA levels. The data are shown the as the mean ± SE from three sets of independent experiments. ∗P<0.05; ∗∗P<0.01; ∗∗∗P<0.001; ∗∗∗∗P<0.0001.
Figure 5.
ABCA1 and ApoE mRNA levels. U87 cells were treated with a control (vehicle) and the flavonoid compounds. qRT-PCR were used to examine the mRNA levels and fold-changes was calculated by the 2-ΔΔCt method as compared with non-infection cell control and using endogenous GAPDH mRNA level for normalization. (A) The fold change of ABCA1 in mRNA levels; (B) The fold change of ApoE in mRNA levels. The data are shown the as the mean ± SE from three sets of independent experiments. ∗P<0.05; ∗∗P<0.01; ∗∗∗P<0.001; ∗∗∗∗P<0.0001.
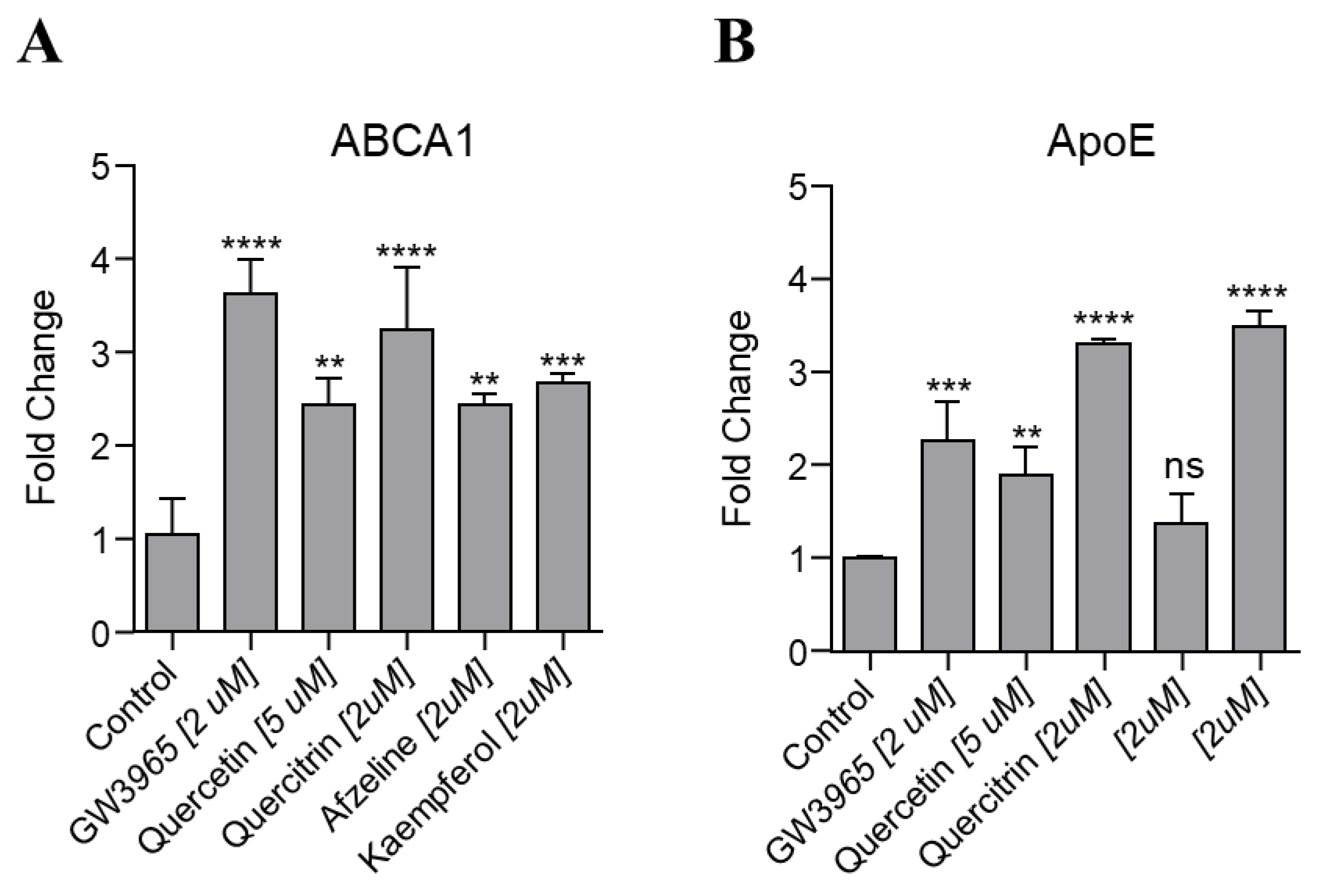

Table 1.
Identification of markers in the ethanolic extract by UHPLC-DAD and UHPLC-ESI-HR-MS/MS.
| Peak no. | Retention time (min) | UVmax λ (nm) | MS | MS2 | Molecular formula | AME (ppm) |
Tentative annotation | ||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 7.80 | 260; 349 | 447.0936 | 300.0273 | C21H20O11 | 1.8 | Quercitrin | ||
| [M-H]- | [M-rhamnose-H]-● | ||||||||
| 303.0509 | |||||||||
| [M-rhamnose+H]+ | |||||||||
| 2 | 9.44 | 266; 341 | 431.0980 | 284.0323 | C21H20O10 | -0.5 | Afzelin | ||
| [M-H]- | [M-rhamnose-H]-● | ||||||||
| 287.0561 | |||||||||
| [M-rhamnose+H]+ | |||||||||
| 3 | 14.26 | 229; 268; 314 |
593.1296 | 285.0396 | C30H26O13 | -0.2 | Kaempferol 3-(6''-p- coumarylglucoside) or Kaempferol 7-(6''- p-coumarylglucoside) | ||
| [M-H]- | [M-coumarylglucoside-H]- | ||||||||
| 595.1464 | 309.0981 | ||||||||
| [M+H]+ | [M-kaempferol+H]+ | ||||||||
| 287.0559 | |||||||||
| [M-coumarylglucoside+H]+ | |||||||||
*AME: Accurate mass error (ppm).
Table 2.
Aglicones present in the extract after hydrolysis.
| Compound | Standard retention time (min) |
Standard UV λ (nm) | Hydrolyzed extract retention time (min) |
UV λ (nm) |
|---|---|---|---|---|
| Quercetin | 12.30 | 202; 257; 368 | 12.29 | 203; 257; 368 |
| Kaempferol | 16.59 | 197; 267; 367 | 16.57 | 197; 267; 366 |
| Luteolin | 12.63 | 208; 256; 348 | 12.54* | 230; 310* |
| p-Coumaric acid | 6.15 | 227; 310 | 6.16 | 229; 311 |
*There is no match between the standard and the reaction product.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



