Submitted:
01 September 2025
Posted:
02 September 2025
You are already at the latest version
Abstract
Background/Objectives: Gastrointestinal (GI) cancer, including colorectal, gastric, pan-creatic, hepatic, and esophageal malignancies, remains a major global health burden, with limited predictive biomarkers and targeted therapies. Omentin, also known as intelectin-1, is a secreted adipokine with anti-inflammatory, insulin-sensitizing, and immune-modulatory functions, primarily expressed in visceral adipose tissue. While omentin has been associated with favorable metabolic outcomes, its role in cancer pathogenesis appears context-dependent and remains poorly understood. This review investigates the biological functions, expression patterns, and clinical relevance of omentin across GI malignancies. Methods: A comprehensive review of the literature was conducted to evaluate the role of omentin in gastrointestinal cancers. Studies investigating omentin’s expression in tumor tissues, blood, and adipose tissue, its prognostic signifi-cance, and its therapeutic potential were analyzed. Preclinical and clinical studies ex-ploring the effects of omentin analogues or omentin-enhancing agents were also re-viewed. Results: Omentin exhibits anti-proliferative, anti-inflammatory, and an-ti-angiogenic effects within the tumor microenvironment in several GI malignancies. High intratumoral omentin expression correlates with improved prognosis in colorectal, gastric, and hepatic cancers, whereas elevated circulating levels, particularly in colorectal and pancreatic cancers, have been paradoxically associated with increased cancer risk and poor outcomes. Moreover, omentin-1 inhibits key oncogenic pathways including PI3K/Akt and NF-κB, modulates immune cell infiltration, and suppresses cancer stem cell activity. However, its different mechanism of action in systemic versus local level remains a major unresolved issue. Conclusions: Omentin demonstrates significant diagnostic, prognostic, and therapeutic potential across gastrointestinal cancers, with evidence supporting its tumor-suppressive and immunomodulatory functions. However, its con-text-dependent behavior -especially the disparity between circulating and tu-mor-localized expression- necessitates further translational studies which could validate its clinical utility and therapeutic potential across different tumor types.
Keywords:
Omentin
; Gastrointestinal Neoplasms
; Adipokines
; Tumor Microenvironment
; Bi-omarker
1. Introduction
The introduction should briefly place the study in a broad context and highlight why it is important. It should define the purpose of the work and its significance. The current state of the research field should be carefully reviewed and key publications cited. Please highlight controversial and diverging hypotheses when necessary. Finally, briefly mention the main aim of the work and highlight the principal conclusions. As far as possible, please keep the introduction comprehensible to scientists outside your particular field of research. References should be numbered in order of appearance and indicated by a numeral or numerals in square brackets—e.g., [1] or [2,3], or [4,5,6]. See the end of the document for further details on references.
Gastrointestinal (GI) cancers remain among the leading causes of cancer-related morbidity and mortality worldwide. Despite advances in surgical technique and medical oncology, outcomes for many patients with GI cancers—especially at advanced stages—remain poor [1]. This underscores the need for novel biomarkers that could aid in early detection, risk stratification, and therapeutic guidance.
Omentin, (also Omentin-1, intelectin-1, ITLN1) is an adipokine primarily produced in visceral adipose tissue, particularly within omental fat depots. In contrary to many adipokines that are pro-inflammatory, omentin is generally considered a beneficial adipose-derived factor with metabolic and anti-inflammatory roles [2]. Biologically, omentin-1 exerts insulin-sensitizing effects—enhancing insulin-mediated glucose uptake in adipocytes—and displays broad anti-inflammatory and anti-atherogenic functions [3]. Notably, circulating omentin-1 levels are inversely associated with obesity and related metabolic disorders. Omentin expression is reduced in conditions of visceral obesity, insulin resistance, and type 2 diabetes, whereas weight loss or exercise can raise omentin-1 circulating levels [4,5,6,7,8].
The observation that pro-tumor metabolic conditions coincide with low omentin has prompted interest in the possible influence of this adipokine on cancer development. It is hypothesized that dysregulation of adipokines, such as omentin, could explain why metabolic syndrome contributes to tumorigenesis in organs like the stomach, colon, and esophagus [9].
Emerging evidence suggests that omentin plays a role in cancer biology. In gastrointestinal cancers, intelectin-1 gene expression has been reported to be higher in tumor tissues compared to adjacent normal tissues, and patients with GI tumors may exhibit elevated circulating omentin-1 levels compared to healthy controls [10]. Despite growning interest, significant gaps still remain regarding the precise role of omentin-1 in the initiation and progression of GI malignancies. The prognostic and diagnostic value of circulating omentin-1 remains uncertain due to heterogeneity in existing studies’ design, small sample sizes, and variations in detection methods. Furthermore, the molecular mechanisms of interaction between omentin-1 and cancer-related pathways are only partially understood, and its potential as a therapeutic target has yet to be thoroughly investigated.
This review aims to synthesize current evidence on the biological role and clinical relevance of omentin in gastrointestinal cancers. By highlighting emerging findings and key areas of uncertainty, we seek to suggest future research directions and explore the potential for omentin as a novel biomarker and therapeutic target in GI oncology.
2. Omentin: Biochemistry and General Functions
Omentin (intelectin-1) is a secreted glycoprotein adipokine encoded by the ITLN1 gene and predominantly produced in visceral adipose tissue [11]. Humans express two omentin isoforms, omentin-1 and omentin-2, which share ~83% amino acid identity and are encoded by adjacent genes (ITLN1 and ITLN2) clustered on chromosome 1q22–q23[5]. Omentin-1 consists of 313 amino acids (~35 kDa) including a 16-amino-acid signal peptide for secretion [12]. Structurally, it contains a fibrinogen-related region and a C-terminal intelectin lectin domain, and it assembles into a disulfide-linked homotrimer (~120 kDa) as its native form [12]. This lectin-like protein binds to specific carbohydrate moieties in a calcium-dependent manner, recognizing microbial sugars (e.g. β-D-galactofuranose) but not typical mammalian glycans [12]. Such biochemical features place omentin in the X-lectin family of innate immune proteins while also functioning as an adipose-derived cytokine.
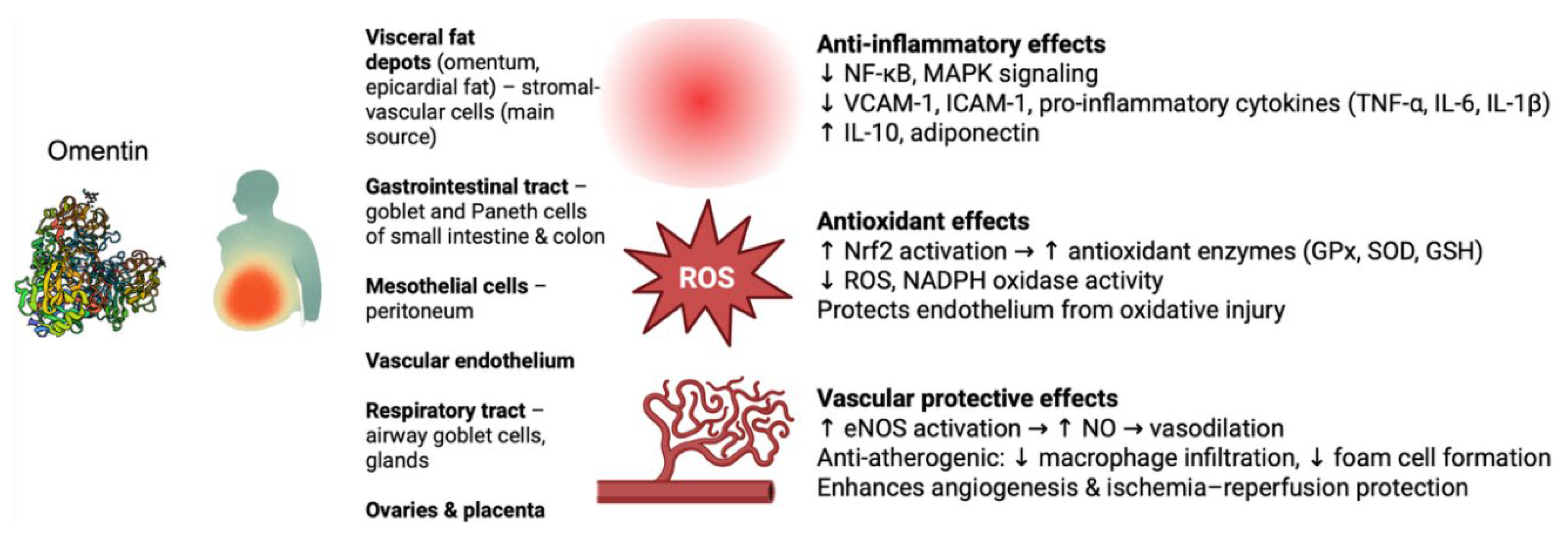
Omentin-1 is highly expressed in visceral fat depots (especially omental and epicardial fat) and is secreted into the circulation, whereas expression in subcutaneous fat is much lower [13]. Notably, omentin is produced mainly by stromal-vascular cells within adipose tissue rather than by adipocytes themselves [14]. Beyond adipose tissue, omentin is found in multiple tissues and cell types: it is expressed by mesothelial cells (e.g. in the peritoneum), endothelial and other vascular cells, and prominently by certain gut epithelial cells [14]. In the digestive tract, high omentin (intelectin-1) levels are seen in goblet and Paneth cells of the small intestine and colon, from which it is secreted into the mucus [15]. Omentin is also detected in the respiratory tract (airway goblet cells and glands), ovaries, placenta, and other tissues [16,17,18]. This wide distribution, particularly at barrier surfaces, suggests a role in immune surveillance in addition to its endocrine function as an adipokine [16,19,20].
2.1. Anti-Inflammatory and Immune Regulatory Roles
Omentin-1 exerts notable anti-inflammatory effects in various cell types and tissues. It generally acts to dampen pro-inflammatory signaling pathways and cytokine production. For example, omentin-1 inhibits the expression of adhesion molecules (VCAM-1, ICAM-1) and suppress inflammatory cytokine release in endothelial cells by blocking key stress-activated pathways (p38 MAPK, JNK) and the Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) transcription factor [12]. In human endothelial and adipose cells exposed to inflammatory stimuli (like free fatty acids or TNF-α), omentin-1 reduces NF-κB activation and lowers production of cytokines such as TNF-α, IL-6, IL-1β, and chemokine MCP-1 [21,22]. Several studies confirm that omentin’s anti-inflammatory action involves intracellular signaling modulation – for instance, activating AMP-activated protein kinase (AMPK) and Protein kinase B (Akt), while antagonizing pro-inflammatory kinases [14,23]. In obese mice, omentin derived from perivascular fat was found to increase anti-inflammatory mediators (interleukin-10 and adiponectin) and concurrently decrease levels of TNF-α, IL-6, and IL-1β in tissues by inhibiting the TXNIP/NLRP3 inflammasome pathway [24].
In addition to blunting inflammatory signaling, omentin plays a direct role in immune regulation and host defense. As a lectin, omentin can recognize and bind to carbohydrate antigens present on microbial pathogens, thereby functioning in innate immunity. It selectively binds microbial exocyclic 1,2-diol motifs (such as β-D-galactofuranose found in bacteria) and can agglutinate bacteria upon binding [20]. This agglutination not only sequesters microbes, but also promotes their clearance: for instance, human omentin-1 binding to Helicobacter pylori causes bacterial clumping and significantly enhances phagocytosis of the bacteria by macrophages [25]. Omentin-1 is present at high concentration in the intestinal mucus and on mesothelial surfaces, which suggests it contributes to the first-line immune defense by blocking pathogen entry and facilitating neutrophil/macrophage uptake of invaders. Furthermore, omentin-1 can modulate immune cell function; treating macrophages with recombinant omentin-1 in vitro was shown to attenuate lipopolysaccharide-induced expression of pro-inflammatory genes and to reduce foam cell formation (lipid accumulation) in these cells [24]. These effects were associated with activation of the Akt signaling pathway in macrophages, and pharmacologic inhibition of Akt abolished omentin’s anti-inflammatory influence [24]. Taken together, such findings indicate that omentin not only serves as an opsonin-like innate immune lectin but also actively regulates inflammatory cell signaling, biasing immune responses toward an anti-inflammatory, tissue-protective state.
2.2. Antioxidant Effects and Oxidative Stress Modulation
Another important function of omentin is the mitigation of oxidative stress in tissues. Chronic inflammation often coincides with elevated reactive oxygen species (ROS) and oxidative damage, and omentin-1 appears to counteract these processes. In vascular cells, omentin has demonstrated antioxidant effects: for example, it can inhibit NADPH oxidase-mediated superoxide production triggered by TNF-α in vascular smooth muscle cells, thereby reducing oxidative stress and inflammation in the vascular wall [26,27]. Omentin-1 also protects the endoplasmic reticulum and endothelium from oxidative injury by enhancing nitric oxide (NO) bioavailability – partly through activating peroxisome proliferator-activated receptor-δ (PPAR-δ) and AMPK, which in turn upregulates endothelial NO synthase and antioxidant defenses [28]. In models of acute inflammation, omentin was shown to activate the Nrf2 signaling pathway, a master regulator of cellular antioxidant responses. In a murine colitis study, administration of omentin-1 markedly decreased markers of oxidative damage (reducing ROS and malondialdehyde levels while increasing glutathione and superoxide dismutase activity) and concomitantly suppressed NF-κB–driven inflammation in the intestine. The protective mechanism was traced to omentin-1 activating Nrf2, which upregulated antioxidant enzymes and restored redox balance, ultimately preserving the intestinal barrier and ameliorating inflammation. This Nrf2-mediated antioxidative effect of omentin is likely relevant in other tissues as well, given that Nrf2 activation and NF-κB inhibition have been observed in omentin-treated cells under stress [29].
Direct evidence of omentin’s cytoprotective antioxidant action comes from endothelial cell experiments. Human endothelial cells pretreated with omentin-1 are significantly more resistant to oxidative injury. In-vitro assays showed that omentin-1 prevented hydrogen peroxide (H₂O₂)-induced apoptosis of endothelial cells and reduced intracellular ROS accumulation. Notably, omentin not only scavenged ROS, but also boosted the cells’ own antioxidant defenses by increasing the activity of glutathione peroxidase (GPx). The net effect was preservation of cell viability and function under oxidative challenge. Such findings underscore that omentin-1 serves as an antioxidant adipokine: by lowering oxidative stress and enhancing antioxidant enzymes, it helps shield cells (endothelium, immune cells, etc.) from oxidative damage. These antioxidant properties of omentin likely contribute to its anti-inflammatory and vascular-protective effects, since reduced oxidative stress limits the activation of inflammatory pathways and endothelial dysfunction [30].
2.3. Vascular Biology and Vascular Protective Functions
Omentin-1 plays a multifaceted role in vascular biology, generally exerting vasoprotective and anti-atherogenic effects. One key function is promoting healthy endothelial function and vasodilation. Omentin-1 stimulates the production of nitric oxide (NO) in endothelial cells, leading to endothelium-dependent relaxation of blood vessels. It has been shown to activate endothelial NO synthase (eNOS) via Akt/AMPK signaling, thereby increasing NO release. This vasodilatory action helps maintain proper vascular tone and blood flow. In experimental models of endothelial dysfunction (such as in insulin-resistant or diabetic rodents), omentin-1 treatment was able to restore normal endothelium-dependent vasorelaxation responses. For instance, arteries from diabetic rats showed improved acetylcholine-induced relaxation when treated with omentin, which requires intact perivascular adipose tissue and is likely mediated by NO and reduced oxidative stress. Thus, omentin contributes to preserving endothelial health and preventing vascular contraction or hypertension through its NO-mediated, vasodilating influence [14].
Omentin’s anti-inflammatory properties are particularly beneficial in the vasculature, where chronic inflammation drives atherosclerosis and vascular damage. Omentin-1 suppresses endothelial activation by reducing expression of adhesion molecules (ICAM-1, VCAM-1) that recruit leukocytes to the vessel wall [26,31,32]. By inhibiting NF-κB and MAPK signaling in vascular cells, omentin curtails the cascade of inflammation that leads to endothelial dysfunction and plaque formation [21]. The adipokine is therefore considered anti-atherogenic. Indeed, in-vivo studies have demonstrated that omentin can slow the development of atherosclerotic lesions. In apolipoprotein E-deficient mice (a model prone to aggressive atherosclerosis), transgenic overexpression of human omentin-1 in adipose tissue or chronic infusion of omentin significantly attenuated plaque formation in the aorta Omentin-treated ApoE⁻/⁻ mice showed smaller atherosclerotic lesions accompanied by a marked reduction in macrophage infiltration into the plaques and lower arterial levels of proinflammatory mediators like TNF-α, IL-6, and MCP-1[23,33]. This indicates that omentin curbs the inflammatory cell recruitment and cytokine milieu that drive atheroma progression. In parallel, omentin appears to influence macrophage behavior in atherosclerotic lesions: treatment of cultured human macrophages with omentin-1 reduced their uptake of oxidized LDL and formation of foam cells (cholesteryl ester-laden macrophages), which are key contributors to plaque growth [23]. Omentin also diminished LPS-induced cytokine expression in these macrophages, consistent with a shift to a less inflammatory, more cholesterol-effluxing phenotype [33]. Mechanistically, omentin’s activation of Akt in macrophages was required for these anti-atherogenic effects, aligning with its known signaling in other cell types. Beyond inhibiting atherosclerosis, omentin may promote other aspects of vascular homeostasis. It has been reported to enhance angiogenic responses in ischemic tissue and to protect the heart from ischemia–reperfusion injury, largely by activating pro-survival pathways like Phosphoinositide 3-kinase/ Protein kinase B (PI3K/Akt) and reducing oxidative/apoptotic damage in endothelial [33]. While metabolic disorders (obesity, diabetes) reduce circulating omentin levels and are associated with endothelial dysfunction, higher omentin-1 is generally correlated with better cardiovascular outcomes, reflecting its vaso-protective actions [34].
In summary, omentin serves as an anti-inflammatory, antioxidant guardian of the vasculature: it promotes endothelial-dependent vasodilation, inhibits inflammatory adhesion and infiltration in vessels, and prevents the development of atherosclerotic changes. These properties make omentin-1 an adipokine with significant cardiovascular protective potential independent of its metabolic effects.
Figure 1.
Overview of omentin expression and protective functions. Omentin, mainly secreted by visceral fat depots and expressed in gastrointestinal, mesothelial, vascular, respiratory, and reproductive tissues, exerts multiple protective effects. It suppresses inflammation (↓ NF-κB/MAPK signaling, ↓ VCAM-1/ICAM-1, ↓ TNF-α/IL-6/IL-1β, ↑ IL-10, adiponectin), reduces oxidative stress (↑ Nrf2-driven antioxidant enzymes, ↓ ROS, NADPH oxidase activity), and protects the vasculature (↑ eNOS–NO–mediated vasodilation, anti-atherogenic activity, enhanced angiogenesis and ischemia–reperfusion protection).. NF-κB, nuclear factor kappa B; MAPK, mitogen-activated protein kinase; VCAM-1, vascular cell adhesion molecule-1; ICAM-1, intercellular adhesion molecule-1; TNF-α, tumor necrosis factor alpha; IL-6, interleukin-6; IL-1β, interleukin-1 beta; IL-10, interleukin-10; Nrf2, nuclear factor erythroid 2–related factor 2; GPx, glutathione peroxidase; SOD, superoxide dismutase; GSH, glutathione; ROS, reactive oxygen species; NADPH, nicotinamide adenine dinucleotide phosphate; eNOS, endothelial nitric oxide synthase; NO, nitric oxide.
Figure 1.
Overview of omentin expression and protective functions. Omentin, mainly secreted by visceral fat depots and expressed in gastrointestinal, mesothelial, vascular, respiratory, and reproductive tissues, exerts multiple protective effects. It suppresses inflammation (↓ NF-κB/MAPK signaling, ↓ VCAM-1/ICAM-1, ↓ TNF-α/IL-6/IL-1β, ↑ IL-10, adiponectin), reduces oxidative stress (↑ Nrf2-driven antioxidant enzymes, ↓ ROS, NADPH oxidase activity), and protects the vasculature (↑ eNOS–NO–mediated vasodilation, anti-atherogenic activity, enhanced angiogenesis and ischemia–reperfusion protection).. NF-κB, nuclear factor kappa B; MAPK, mitogen-activated protein kinase; VCAM-1, vascular cell adhesion molecule-1; ICAM-1, intercellular adhesion molecule-1; TNF-α, tumor necrosis factor alpha; IL-6, interleukin-6; IL-1β, interleukin-1 beta; IL-10, interleukin-10; Nrf2, nuclear factor erythroid 2–related factor 2; GPx, glutathione peroxidase; SOD, superoxide dismutase; GSH, glutathione; ROS, reactive oxygen species; NADPH, nicotinamide adenine dinucleotide phosphate; eNOS, endothelial nitric oxide synthase; NO, nitric oxide.
3. The Role of Omentin in Metabolic Disorders
Metabolic disorders such as obesity, type 2 diabetes mellitus, metabolic syndrome, and Metabolic–associated steatotic liver disease (MASLD) are increasingly recognized as major risk factors for the initiation and progression of gastrointestinal malignancies. Chronic low-grade inflammation, hyperinsulinemia, insulin resistance, and dysregulated adipokine secretion create a systemic pro-tumorigenic environment [1,9]. These alterations promote DNA damage, oxidative stress, and angiogenesis while impairing immune surveillance, thereby facilitating tumorigenesis in organs such as the stomach, colon, liver, and pancreas [35,36].
3.1. Omentin and Obesity
Omentin-1 was first identified in stromal vascular cells of the visceral adipose tissue, and its levels have a notable inverse relationship with obesity. Numerous studies report that circulating omentin-1 is negatively correlated with body mass index (BMI), waist circumference, and adiposity in both adults and children [4,5,6]. In fact weight reduction through diet or exercise have been shown to increase circulating omentin-1 concentrations [7,8]. This obesity-related decline in omentin may be driven by the pro-inflammatory environment of excess fat, as suggested by the link between high TNF-α/IL-1 and suppressed omentin in adipose tissue [21].
3.2. Omentin, Insulin Resistance and Diabetes
Omentin is a key adipokine involved in glucose metabolism and insulin sensitivity, with accumulating evidence suggesting that its reduced levels contribute to the pathophysiology of type 2 diabetes mellitus (T2DM). In vitro studies have shown that omentin enhances insulin signal transduction by activating protein kinase B (Akt), thereby improving the insulin responsiveness of adipose tissue [11]. Therefore, a lower circulating concentration of omentin might play a mechanistic role in the development of insulin resistance [4]. Clinically, decreased plasma omentin levels have been consistently observed in patients with T2DM, while individual studies has been assessing also parameters such as fasting glucose and the HOMA-IR index [4,37]. Notably, the reduction in omentin is not limited to overweight individuals; patients with normal BMI and T2DM also display significantly lower omentin concentrations compared to healthy controls [4], indicating that the decrease is not solely attributable to obesity. Furthermore, reduced omentin levels have been linked to diabetes-related microvascular complications; patients with diabetic retinopathy, neuropathy, or nephropathy consistently show significantly lower circulating omentin compared with those without such complications, partly through mechanisms involving downregulation of vascular endothelial growth factor (VEGF). Interestingly, while the relationship between omentin and T2DM is well established, findings in type 1 diabetes mellitus (T1DM) remain inconsistent. Some studies report decreased plasma omentin levels in T1DM, whereas others have observed elevated concentrations, suggesting a divergent regulatory mechanism or role for omentin in this context [37].
3.3. Omentin and Metabolic Syndrome
Metabolic syndrome is a cluster of interconnected risk factors—including abdominal obesity, dyslipidemia, hypertension, and hyperglycemia—that predispose individuals to type 2 diabetes and cardiovascular disease. Omentin-1 appears to be closely associated with these risk factors. As previously discussed, reduced omentin-1 levels are commonly observed in conditions such as obesity, insulin resistance, and type 2 diabetes—all key components of metabolic syndrome [4,11,17]. Furthermore, omentin-1 is positively correlated with plasma levels of high-density lipoprotein (HDL) and negatively with very low-density lipoprotein (VLDL), which is a lipoprotein linked to atherogenic lipid profiles [5]. Studies investigating metabolic syndrome have consistently found that affected individuals tend to have lower circulating omentin-1 levels compared to healthy controls, a finding that aligns with omentin’s inverse association with body mass index (BMI), triglyceride levels, and insulin resistance [38,39,40]. Notably, there is also evidence that omentin-1 may have predictive value. In a prospective study of individuals with hypertension and increased risk for metabolic syndrome, lower baseline omentin-1 concentrations were associated with a higher likelihood of developing the full syndrome during follow-up [39]. Collectively, these findings suggest that omentin-1 is not only a sensitive marker of existing metabolic dysregulation, but it might also have predictive value for identifying individuals at risk of progressing to overt metabolic syndrome.
3.4. Omentin and Metabolic–Associated Steatotic Liver Disease
Metabolic-associated steatotic liver disease is associated with dysregulation of omentin-1, an insulin-sensitizing and anti-inflammatory adipokine. Clinically, patients with Metabolic dysfunction-associated steatotic liver disease (MASLD) consistently exhibit significantly lower circulating omentin-1 levels compared to healthy controls [35]. Moreover, in obese individuals, those who progress to steatohepatitis (MASH) usually show a further reduction in omentin-1 levels (independent of fibrosis), suggesting an inverse relationship between omentin and disease severity [36]. Diminished omentin-1 might exacerbate MASLD by worsening insulin resistance and inflammation: low omentin correlates with higher glycemia and Homeostatic Model Assessment of Insulin Resistance (HOMA-IR), whereas normal omentin enhances insulin-stimulated glucose uptake via Akt and suppresses macrophage NF-κB activation, thereby mitigating systemic insulin resistance and inflammatory signaling [11,35]. In vitro and animal studies support a protective role for omentin-1 in the liver. Administration of omentin-1 in high-fat diet mouse models reduces hepatic fat accumulation and improves metabolic homeostasis, largely by activating AMPK and restoring autophagy in hepatocytes [41]. Similarly, treating fat-laden hepatocyte cultures with omentin-1, reduced TNF-α levels, ER stress, and oxidative stress via inhibition of NF-κB/MAPK signaling [36]. Through these actions, omentin-1 appears to counteract the key pathophysiological drivers of MASLD – hepatic steatosis, insulin resistance, and chronic inflammation – potentially slowing progression to fibrosis. Consistently, experimental models have reported that omentin-1 therapy can ameliorate steatosis and prevent fibrotic changes [36,41]. In summary, omentin plays a protective role in MASLD pathogenesis and confers metabolic and anti-inflammatory benefits that are diagnostically and therapeutically relevant in the context of fatty liver disease.
An overview of the role of omentin in metabolic disorders is presented in Figure 2.
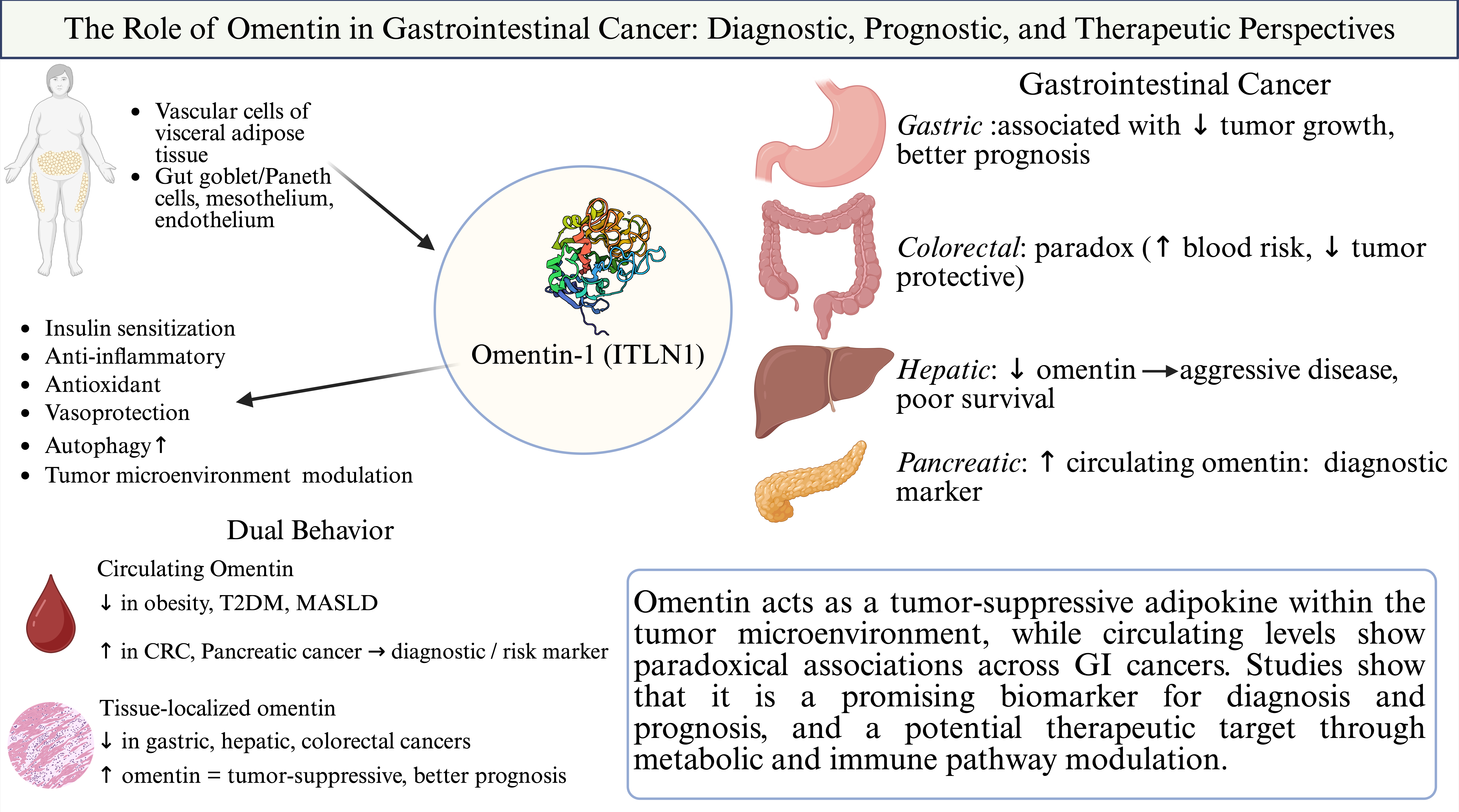
4. The Role of Omentin in Gastrointestinal Cancers
4.1. Upper Gastrointestinal Cancer
In their cross-sectional analysis of adipose tissue from upper-gastrointestinal cancer patients, Miller et al. found that omentin was the single most increased transcript in visceral adipose tissue (VAT). Compared with healthy kidney-donor controls, omentin mRNA had risen nine-fold in weight-stable cancer patients and ten-fold in cachectic patients, while remaining unchanged in subcutaneous fat. Gene-set enrichment showed that this sharp induction occurred against a background of suppressed adipogenesis and oxidative-phosphorylation programmes, suggesting that VAT responds to tumour burden by shifting away from energy-storage genes and strongly favouring omentin expression. Protein assays confirmed the transcriptional signal. Omentin concentrations in VAT were lowest in controls (≈ 20 ng mL⁻¹), intermediate in weight-stable cancer (≈ 31 ng mL⁻¹) and highest in cachectic patients (≈ 44 ng mL⁻¹), indicating a disease- and weight-loss-dependent escalation. Circulating omentin showed greater inter-individual variability, but cachectic patients still displayed significantly higher plasma levels than weight-stable counterparts. These findings suggested that omentin is selectively amplified in gastrointestinal cancer—particularly when cachexia is present—implicating it as a candidate mediator of fat–muscle cross-talk and a potential therapeutic target in cancer-associated metabolic dysfunction [42].
4.1.1. Esophageal Cancer
Esophageal cancer including squamous cell carcinoma (ESCC) and adenocarcinoma (EAC) has not been studied as extensively as other GI cancers in the context of omentin. Esophageal adenocarcinoma shares risk factors with gastric cancer (notably obesity and gastroesophageal reflux leading to intestinal metaplasia), which suggests adipokines might be relevant [43]. However, specific data on omentin-1 in esophageal cancer are limited. To date, no large clinical studies were identified that directly measure omentin levels in esophageal cancer patients versus controls.
In a multi-gene prognostic model developed for ESCC, ITLN1 emerged as one of only two protective genes contributing to a composite immune-related risk signature. Higher ITLN1 expression was associated with lower calculated risk scores and independently predicted both improved overall and recurrence-free survival across two cohorts -one based on microarray data (GSE53624) and another validated by Quantitative real-time polymerase chain reaction (qRT-PCR). Although functional experiments were not performed, the authors proposed that elevated omentin-1 may reflect a more immunocompetent tumor microenvironment, consistent with prior reports of its tumor-suppressive role in gastric and colorectal cancers [44].
Complementing these findings, a separate glycolipid metabolism-focused transcriptomic analysis in ESCC also identified ITLN1 as a protective factor. The gene was downregulated in tumor tissue compared to adjacent normal mucosa, and its expression inversely correlated with poor prognostic features. Although no functional experiments were done on ITLN1, the authors noted its known anti-inflammatory actions—particularly its ability to dampen macrophage NF-κB signaling—which fits the study’s broader finding that high-risk ESCC tumors contain fewer B-cells and NK-cells and a more immunosuppressive micro- environment. They suggested that loss of omentin-1 is part of a metabolic and immune shift toward poorer prognosis, whereas retaining or restoring omentin-1 activity might help maintain anti-tumor immunity in ESCC [45].
In contrast, studies of esophageal adenocarcinoma have positioned ITLN1 not as a dynamic biomarker of progression, but rather as a stable marker of Barrett’s esophagus (BE)—the metaplastic precursor lesion. A single-cell RNA-sequencing study demonstrated that ITLN1 is consistently enriched in goblet-cell–like populations within BE, but does not change meaningfully across progression stages (non-dysplastic BE, low- and high-grade dysplasia, and focal adenocarcinoma). In short, the paper confirmed that omentin-1 is a characteristic transcript of Barrett’s metaplasia, but it provided no evidence that changes in its level signal, drive, or predict the transition to esophageal adenocarcinoma [46].
This interpretation is supported by transcriptomic review in 2021, which identified ITLN1 as a reliable goblet-cell marker in BE based on earlier single-cell datasets, but did not implicate it in the pathogenesis or clinical behavior of esophageal adenocarcinoma. No recurrent mutations, copy-number alterations, or prognostic roles were found to ITLN1 in EAC, suggesting that it serves solely as a BE cell-type marker and is not linked directly to the initiation, progression, or treatment of esophageal cancer [47].
In summary, existing evidence suggest divergent roles in omentin biology across esophageal cancer subtypes. In ESCC, omentin-1 appears to function as a tumor-suppressive and immune-favorable marker, suggesting a potential mechanistic role that remains to be elucidated. In contrast, in esophageal adenocarcinoma, omentin-1 serves chiefly as a molecular identifier of metaplastic BE cells without dynamic changes during neoplastic progression.
4.1.2. Gastric Cancer
The role of omentin-1 in gastric cancer has been explored both at the tissue and mechanistic levels, with consistent evidence pointing toward a tumor-suppressive function. In a single-center study of 196 gastric adenocarcinoma patients, Zheng et al. demonstrated an approximately eight-fold increase in ITLN1 mRNA in tumors compared to paired normal mucosa, confirmed by qPCR (p < 0.001). Immunohistochemical analysis localized the protein to goblet cells of intestinal metaplasia and to malignant glands—never to native gastric epithelium—suggesting that ITLN1 expression is induced as part of the CDX2-driven intestinal transdifferentiation program. Indeed, a strong positive correlation was observed between nuclear CDX2 and cytoplasmic omentin-1 staining (p < 0.001).
Clinically, ITLN1 expression was strongly associated with a favorable tumor phenotype. Among ITLN1-positive tumors, 80% exhibited intestinal-type histology compared to 61% of ITLN1-negative cases. Similarly, 88% of ITLN1-positive tumors were well or moderately differentiated (G1/G2) versus only 60% among ITLN1-negatives. Limited depth of invasion (T1/T2) was observed in 87% of ITLN1-positive cases versus 68% of those lacking ITLN1 expression. Nodal involvement (N+) occurred in just 8% of ITLN1-positive tumors, compared to 34% in the negative group, while distant metastases (M1) were present in 25% of ITLN1-positive cases, compared to 53% in ITLN1-negative patients. Functionally, ITLN1-high tumors demonstrated significantly lower cellular proliferation, with Ki-67 indices decreasing in parallel with rising ITLN1 expression (p = 0.001). Moreover, expression of heparanase -an extracellular matrix-degrading enzyme implicated in metastatic invasion- was inversely correlated with ITLN1 levels (p < 0.001). These biologic and pathologic features translated into survival outcomes: patients with ITLN1-positive tumors had a median overall survival of 44 months, substantially higher than the 25 months observed in ITLN1-negative cases. In multivariate Cox regression controlling for TNM stage, grade, and nodal status, absence of ITLN1 expression independently predicted worse prognosis, doubling the hazard of death (HR = 2.25; 95% CI: 1.23–3.98; p = 0.001). Collectively, the authors suggested that omentin is more than a passive marker which is induced during CDX2-mediated intestinal metaplasia; as it may actively participate in suppressing tumor proliferation, invasion, and dissemination. Thus, they suggested that it might serve as a viable immunohistochemical marker and potential effector for therapeutic modulation in gastric cancer [48].
Li et al. investigated how omentin affects the behavior of gastric cancer cells, by increasing ITLN1 levels in SGC-7901 and AGS gastric cancer cell lines. Forced ITLN1 expression activated a molecular chain: it silenced the PI3K→AKT→IKK axis, kept NF-κB in the cytoplasm, and thereby lifted NF-κB-mediated repression of the HNF4A promoter. Elevated Hepatocyte nuclear factor 4 alpha (HNF4α) then trapped β-catenin at the membrane, lowered TOP/FOP luciferase output, and reduced the activity of several genes involved in cancer progression (including AXIN2, CCND2, RUNX2, and MMP3).
This molecular shift was reflected in distinct biological behaviors. Gastric cancer cells overexpressing ITLN1 formed fewer colonies, migrated more slowly in scratch assays, and generated smaller subcutaneous tumors in mice—along with a 70% reduction in lung metastases. In contrast, ITLN1-deficient cells showed the opposite behavior, which was reversed when HNF4α was reintroduced, confirming a functional link. In clinical samples, both ITLN1 and HNF4α were upregulated in 90 primary gastric tumors compared to adjacent normal mucosa. However, their expression declined significantly in tumors with poor differentiation, deeper invasion, lymph node involvement, and advanced TNM stage. The expression levels of ITLN1 and HNF4α showed strong correlation (R = 0.82), and high expression of either gene was associated with significantly improved overall survival (p < 0.001). Based on these findings, the authors suggested a tumor-suppressive role for omentin-1 in gastric cancer, acting through inhibition of PI3K/NF-κB signaling, restoration of HNF4α, and suppression of β-catenin activity. Loss of ITLN1 was correlated to a more aggressive tumor phenotype and worse prognosis, suggesting its relevance as both a prognostic marker and therapeutic target [49].
4.2. Hepato-Pancreato-Biliary (HPB) Cancer
4.2.1. Hepatocellular Cancer
Emerging data suggest that omentin-1 plays a tumor-suppressive role in hepatocellular carcinoma (HCC), supported by mechanistic insights, histopathological findings, and clinical observations.
Zhang et al. demonstrated that omentin-1 exerts pro-apoptotic effects in HCC cell lines through a Sirt1-dependent p53 acetylation pathway. In vitro, exogenous omentin-1 at concentrations of 1–2 µg/mL significantly reduced proliferation and increased apoptosis in HepG2 and HuH-7 cells. Omentin-1 enhanced acetylation of p53 without altering its transcript levels, suggesting stabilization through post-translational modification. Silencing Sirt1 reversed these effects, identifying it as the primary deacetylase inhibited by omentin-1. Downstream, omentin-1 triggered mitochondrial apoptotic signaling, while also activated the JNK pathway selectively. Together, these findings support a model in which omentin-1, derived from visceral fat, augments hepatocyte apoptosis via a Sirt1–p53–JNK axis, linking adipokine signaling to anti-tumor mechanisms in the liver. This pathway suggests a potential therapeutic target that links metabolic signals from visceral fat to tumour suppression in the liver. [50]
Li et al. studied a cohort of 149 patients undergoing surgical resection for HCC. Quantitative analyses revealed that ITLN1 expression was significantly reduced in 78% of tumors compared to matched non-neoplastic liver tissue. Low ITLN1 levels correlated with larger tumor size (≥5 cm), poor differentiation, capsular invasion, cirrhosis, and vascular infiltration—hallmarks of biologically aggressive disease. Notably, recurrence rates after resection were nearly four times higher in ITLN1-low tumors than in ITLN1-high ones. Survival analyses further emphasized the prognostic impact: patients with high ITLN1 expression exhibited a 5-year overall survival of 56% and disease-free survival of 36%, compared to 32% and 21%, respectively, in the ITLN1-low group. In multivariate Cox regression adjusting for tumor burden, fibrosis, and vascular features, high ITLN1 remained an independent predictor of improved outcome, halving the hazard of death and recurrence (HR ≈ 0.5). Although the mechanistic pathways were not explored in this study, the authors referenced ITLN1’s known roles in modulating HNF4α and β-catenin signaling as possible avenues for its anti-tumor function. They concluded that diminished ITLN-1 is a hallmark of biologically aggressive HCC and that restoring or mimicking its activity might offer a novel therapeutic target [51].
Furthermore, Karabulut et al. identified ITLN1 as a functional receptor for bovine lactoferrin (LF), mediating its internalization and anti-tumor actions in liver cancer. Using HepG2, Hep3B, and SK-Hep1 cells, the authors demonstrated that LF binds to cell-surface ITLN1 with high affinity, triggering endocytosis and initiating downstream anti-cancer programs. LF-INTL1 engagement activated p38 MAPK and JNK pathways, leading to mitochondrial apoptosis, as well as ERK-mediated p53 upregulation and G1-phase cell cycle. Moreover, silencing ITLN1abolished LF uptake and downstream signaling, underscored the importance of ITLN1 in these processes. In an orthotopic nude-mouse model, oral LF (100–200 mg/kg/day) prevented hepatic tumor outgrowth, preserved body weight, and suppressed pro-angiogenic (VEGF/CD31) and cell-cycle markers (CDK2/4). Histological analysis confirmed near-complete absence of tumor foci in LF-treated livers. Collectively, the authors proposed ITLN1 as a dual-purpose molecule: (i) a predictive marker of LF sensitivity and (ii) a therapeutic target through which ligand-based or mimetic therapies might be developed to enhance apoptosis, cell-cycle control, and angiogenesis suppression in HCC [52].
4.2.2. Pancreatic Cancer
The potential role of omentin-1 in pancreatic adenocarcinoma (PA) has been explored primarily through serum-based studies, with an emphasis on its diagnostic and prognostic implications. In a prospective case–control study, Karabulut et al. measured circulating omentin-1 levels in 33 treatment-naïve patients with histologically confirmed PA and compared them to 30 age-, sex-, and BMI-matched healthy controls. Serum omentin-1 concentrations were markedly elevated in the cancer cohort, with a median of approximately 9.6 ng/mL versus 1.6 ng/mL in controls—an approximate eight-fold increase (p < 0.001). This elevation did not correlate with demographic factors, systemic inflammation, or conventional tumor markers (CA19-9, CEA), but was significantly higher in patients with large primary tumors (≥ 4 cm, p = 0.03), suggesting a link between omentin-1 and tumor burden.
Despite its strong diagnostic separation, omentin-1 did not demonstrate prognostic value. Overall survival in the cohort was 41 weeks, and neither Kaplan–Meier nor multivariate Cox regression analyses identified omentin-1 as a significant predictor of outcome. Instead, prognosis was determined by classical factors such as age > 60 years, ECOG performance status, presence of liver metastases, and response to gemcitabine-based chemotherapy. Based on these results, the authors proposed omentin-1 as a promising adjunct diagnostic biomarker for PA, but not as a reliable predictor of prognosis or treatment response [53].
Further supporting its potential diagnostic relevance, a study by Głuszek et al. compared serum omentin-1 levels across 25 patients with pancreatic ductal adenocarcinoma (PDAC), 10 with chronic pancreatitis (CP), and 36 healthy individuals. Omentin-1 was significantly elevated in PDAC cases (mean ~582 ng/mL) compared to controls (~462 ng/mL; p < 0.05), while patients with CP showed intermediate levels without statistically significant differences from either group. Within the PDAC cohort, omentin-1 levels did not correlate with TNM stage, resectability, age, or BMI. Receiver operating characteristic (ROC) analysis yielded an Area under the curve (AUC) of 0.73, with 65% sensitivity and 78% specificity, suggesting moderate diagnostic accuracy; yet, no association with clinical outcome was found [54].
4.3. Colorectal Cancer
The EPIC-Potsdam case-cohort analysis using baseline plasma from a random sub-cohort of 2,295 adults and 251 incident cases accrued over 10.4 years, provided prospective evidence that circulating omentin-1 is positively—rather than inversely—associated with colorectal-cancer development. Notably, a monotonic dose-response relationship was observed: participants in the highest omentin quartile (median ≈ 563 ng/mL) had a 2.3-fold higher CRC risk than those in the lowest quartile (≈ 287 ng/mL), and each doubling of omentin-1 was associated with nearly a two-fold increase in relative risk (RR ≈ 1.98).
Subgroup analyses revealed a significant interaction with obesity status. In non-obese individuals (BMI < 30 kg/m²), the omentin–CRC association was pronounced (RR per doubling ≈ 2.3), whereas in obese individuals (BMI ≥ 30 kg/m²), no significant relationship was found (RR ≈ 1.1). These findings suggest that omentin-1 may act via distinct biological mechanisms depending on adiposity—possibly through differential activation of PI3K-Akt signaling, genomic instability, or immune surveillance in lean versus obese tissue microenvironments. Despite its positive correlations with protective factors such as HDL-C and adiponectin, omentin-1 did not confer cancer protection, highlighting a potential dual or context-dependent role. Overall, the author team positioned omentin-1 as a metabolically modulated biomarker that independently enhances colorectal-cancer risk stratification, particularly among lean individuals and underscored the need for mechanistic studies to unravel its paradoxical tumor-promoting effects in a non-obese context [55].
A case-control study involving 358 patients with histologically confirmed colorectal adenocarcinoma and 286 colonoscopy-verified healthy controls, found that circulating omentin-1 levels are significantly elevated in CRC and reflected tumor invasiveness. Mean plasma omentin-1 was approximately double in cancer patients compared to controls (≈ 67 ng/mL vs 33 ng/mL, P = 0.005). Stratification by tertiles revealed that individuals in the highest omentin-1 group had an adjusted 5.8-fold increased odds of CRC versus those in the lowest tertile, even after controlling for confounders. Diagnostic performance was high, with an area under the ROC curve of 0.884; at a threshold of 56.9 ng/mL, omentin-1 demonstrated 81% sensitivity and 70% specificity.
High circulating omentin-1 was also associated with more aggressive disease features: levels above the control-defined threshold were significantly linked to deeper tumor invasion (pT3/4) and advanced TNM stage (III–IV). However, omentin-1 showed no association with tumor size, side, grade, nodal status, or distant metastasis. Notably, in the CRC cohort, omentin-1 levels were not correlated with BMI, glucose, or lipid markers—indicating that its elevation reflects cancer biology rather than metabolic syndrome or obesity. Taken together, the authors underlined the role of omentin-1 as a robust and independent biomarker for CRC risk and progression, with potential applications in early detection and disease monitoring in clinical practice [56].
Feng et al. evaluated the diagnostic and prognostic significance of plasma omentin-1 in 319 patients undergoing curative resection for colorectal cancer, compared with 300 age- and sex-matched healthy controls. Pre-operative omentin-1 levels were markedly elevated in CRC patients (mean ≈ 69 ng/mL) relative to controls (≈ 38 ng/mL, P < 0.001), and although levels declined significantly post-operatively, they remained above baseline despite no evidence of disease on imaging. Cross-sectional analyses linked higher circulating omentin-1 to adverse pathological features: patients with TNM stage III/IV disease and nodal metastases had significantly higher levels than those with stage I/II or node-negative tumors (P < 0.01).
Using a plasma threshold of 50 ng/mL, multivariate Cox regression models adjusting for TNM stage, tumor grade, lymph-node status, and vascular invasion identified omentin-1 as an independent predictor of recurrence (HR ≈ 3.3, P < 0.001). During a median follow-up of 24 months, the cohort experienced a 30.6% rate of recurrence or metastasis and a 19.3% mortality rate. Elevated omentin-1 retained its prognostic value in fully adjusted models, predicting both recurrence (OR ≈ 3.3) and 2-year overall mortality (OR ≈ 2.1), with comparable strength to classical predictors such as nodal involvement and vascular tumor emboli. Based on these findings, omentin was suggested as dual-purpose biomarker in CRC: a state marker reflecting disease presence and biological aggressiveness, and a prognostic tool for stratifying risk of early postoperative relapse and death, potentially informing surveillance and adjuvant treatment decisions [57].
Maeda et al. focused on Transmembrane protein 207 (TMEM207), a previously uncharacterized transmembrane protein that was found to be an intracellular chaperone for omentin-1. Immunohistochemical analysis of 216 CRC specimens revealed TMEM207 expression in only 18% of tumors overall but in 64% of mucinous carcinomas, indicating a histotype-specific pattern. Notably, TMEM207-positive tumors exhibited a significantly lower incidence of lymph-node metastasis (2/38, 5.3%) compared to TMEM207-negative counterparts (54/178, 30.3%; P < 0.001), suggesting a potential role for TMEM207 in limiting metastatic dissemination.
Biochemical assays, including co-immunoprecipitation and in situ proximity-ligation, confirmed a physical interaction between TMEM207 and omentin-1 within the cytoplasm—likely in the endoplasmic reticulum–Golgi axis—suggesting that TMEM207 facilitates proper omentin-1 processing and secretion. Functional knockdown of TMEM207 in CRC cell lines (SW480, RCM-1) led to polyubiquitination and proteasomal degradation of omentin-1, significantly reducing extracellular omentin levels and, by inference, diminishing its tumor-suppressive availability in the microenvironment. The authors proposed a mechanistic axis, in which TMEM207 deficiency drives functional omentin-1 loss, compromising anti-metastatic signaling and facilitating nodal dissemination. These findings introduced TMEM207 as a potential therapeutic target, with future interventions aimed at restoring TMEM207 function or exogenous omentin-1 supplementation to limit metastatic progression in colorectal cancer [58].
A review by the same authors consolidated emerging evidence that omentin-1 (intelectin-1) may function as a tumor-suppressive adipokine in colorectal cancer, with its local bioavailability tightly linked to the processing chaperone TMEM207. While circulating omentin-1 is typically reduced in obesity—a pattern shared with adiponectin—the review highlights colorectal proteomic data indicating that tumor omentin-1 expression is over 40-fold higher in stage IV long-term survivors compared to patients with rapid disease progression, suggesting a favorable prognostic role. Additionally, TMEM207 positivity correlated with significantly fewer nodal metastases and, in stage III/IV mucinous tumors, conferred a substantial survival benefit, with 5-year disease-free survival approximately doubling (p = 0.014). Taken together, these findings suggest that preserved TMEM207-mediated omentin-1 secretion may characterize a less aggressive, mucin-rich CRC phenotype and serve as a favorable prognostic marker. Conversely, loss of this pathway may represent a molecular link between visceral obesity and more invasive colorectal carcinogenesis [59].
Zhang et al. demonstrated that colorectal tumors themselves are capable of producing omentin-1, establishing a tumor-autonomous source for this adipose-derived factor. Matched tumor and adjacent normal mucosa from 24 CRC patients revealed approximately 5-fold higher ITLN1 mRNA in the malignant tissue, accompanied by markedly increased protein levels as shown by immunohistochemistry. Notably, omentin-1 staining localized exclusively to the cytoplasm of neoplastic glandular epithelium, with no signal in adjacent non-malignant cells. Functionally, colorectal-cancer cell lines recapitulated this pattern: SW480 cells exhibited integral ITLN1 transcription and secreted progressively increasing amounts of omentin-1 into the culture medium over 48 hours, despite retaining a substantial intracellular pool—implying active synthesis, storage, and secretion. These data highlighted an autocrine or paracrine regulatory loop in which tumor-derived omentin-1 may modulate the local microenvironment.
The authors proposed this tumor-driven omentin-1 production as a mechanistic explanation for the frequent elevation of circulating omentin-1 in CRC patients—despite systemic reductions in obesity—and implied a functional role beyond biomarker status. Given omentin-1’s known capacity to activate PI3K-Akt and JNK pathways, its autocrine secretion may support proliferative and anti-apoptotic signaling in CRC. Thus, targeting the synthesis or downstream signaling of tumor-derived omentin-1 could represent a novel therapeutic strategy in colorectal cancer, particularly in settings where systemic omentin suppression is absent [60].
Ji et al. isolated CD133⁺ tumor-initiating cells from the SW480 colorectal-carcinoma line and demonstrated that omentin-1 acts directly on this stem-cell compartment. Exposure to 1 or 2 µg ml⁻¹ recombinant omentin-1 reduced sphere-forming cell growth and drove cell apoptosis; the effects intensified with both dose and duration (up to 48 h). Pharmacological blockade of phosphatidylinositol-3-kinase with LY294002 further depressed colony-forming ability and raised cell death, and when combined with omentin-1 produced an additive suppressive effect.
Mechanistically, omentin-1 markedly reduced phosphorylated-Akt without altering total Akt, confirming inactivation of the PI3K–Akt pathway. The synergy with LY294002 supports that this signaling axis is a primary target of omentin’s action in colorectal cancer stem-like cells. Taken together, the data demonstrated that omentin-1 is as a negative regulator of tumor-initiating capacity in CRC via PI3K–Akt suppression, suggesting that adipokine deficiency -common in visceral obesity- may diminish inhibitory signals on cancer stem-like cells and thus promote CRC progression [61].
Uyeturk et al. investigated circulating omentin-1 levels in 45 patients with stage III colon carcinoma who had undergone curative resection followed by standard FOLFOX-4 adjuvant chemotherapy. Two months after treatment completion -at a time point verified as recurrence-free via imaging and biomarker evaluation- plasma omentin-1 concentrations were significantly elevated compared to 35 matched healthy controls (median 618 pg/mL vs 376 pg/mL; p < 0.001). This post-treatment rise could not be explained by classical metabolic confounders: fasting glucose, lipid profile, BMI, renal function, and hemoglobin levels were comparable between groups, and omentin-1 did not correlate with any of these parameters within the cancer cohort. These findings suggest a cancer- or treatment-related modulation of omentin-1, independent of metabolic status.
The authors proposed that this post-treatment upregulation reflects a shift in adipokine signaling induced by malignancy or its eradication—potentially reversing the suppression typically seen in visceral obesity. They outlined two potential, non-mutually exclusive interpretations: (i) omentin-1 elevation may represent a compensatory, anti-tumorigenic host response triggered by cytoreductive therapy; or (ii) conversely, it may reflect tumor–adipose cross-talk that promotes residual disease survival. Given that omentin-1 can exhibit both pro- and anti-tumoral functions depending on microenvironmental context, the precise biological role and prognostic implications of this post-treatment elevation remain to be fully elucidated [62].
A case–control study in 358 patients with colorectal cancer (CRC) and 286 age- and BMI-matched controls examined the functional missense variant rs2274907 A>T (Val109Asp) in the ITLN1 gene to assess its contribution to cancer susceptibility and adipokine regulation. The frequency of the minor A allele was similar between patients and controls (26.1% vs 23.4%, P > 0.10), and the distribution of genotypes (TT 55% vs 57%, AT 37% vs 35%, AA 8% in both groups) showed no significant differences, remaining consistent with Hardy–Weinberg equilibrium (χ² = 0.73, P = 0.69). Adjusted logistic regression controlling for metabolic and lifestyle confounders showed no significant association with CRC under either a dominant (AT + AA vs TT: OR = 1.01; 95 % CI: 0.73–1.41; P = 0.95) or recessive model (AA vs TT + AT: OR = 0.93; 95 % CI: 0.52–1.67; P = 0.82). Importantly, circulating omentin-1 levels remained significantly elevated in CRC patients compared to controls (median 196 vs 158 ng/mL; p = 0.005) irrespective of genotype, suggesting that tumor-associated overexpression is not genetically predetermined by this single nucleotide polymorphism.
However, a gene–environment interaction was observed with respect to adiposity. While obesity alone (BMI ≥ 25 kg/m²) conferred an approximately two-fold increase in CRC risk compared to lean TT homozygotes (adjusted OR = 1.99; 95 % CI: 1.06–6.27; P = 0.041), obese individuals carrying at least one A allele (AT or AA) exhibited a three-fold increase in risk (OR = 3.03; 95 % CI: 1.17–8.25; P = 0.022). Furthermore, within the cancer cohort, AA homozygotes showed an over-representation of aggressive features: 67 % had deep mural invasion (pT3–T4) versus 39 % of T-allele carriers (P < 0.001), and 59 % had advanced-stage disease (stage III–IV) compared to 34 % of T-carriers (P = 0.019). These data indicate that while rs2274907 A>T does not independently influence CRC susceptibility, the A allele acts as a risk and aggressiveness modifier in the context of obesity. The authors highlighted this complex gene–environment interaction between adipokine-related genetic variants, obesity, and colorectal cancer biology, underscoring the need for further mechanistic studies and exploration of their potential role in personalized risk assessment models [63].
In a case–control study, Fazeli et al. investigated plasma omentin-1 levels in 39 newly diagnosed colorectal cancer patients compared to 30 age- and sex-matched healthy controls. They observed a dramatic increase in circulating omentin-1 among CRC patients, of approximately 203 ng/mL versus just 9 ng/mL in controls—a nearly 20-fold elevation (p < 0.0001). This substantial difference remained highly significant even after controlling for body mass index, waist-to-hip ratio, age, metabolic syndrome status, and lipid levels, indicating that the rise in omentin-1 was not attributable to obesity or metabolic dysfunction.
Within the CRC group, omentin-1 showed weak positive correlation with total and LDL cholesterol but was unrelated to glucose levels, BMI, or TNM stage. These findings suggest that omentin-1 elevation is an early and stage-independent feature of CRC biology, not simply a marker of tumor burden or body composition. The authors proposed that omentin-1 might serve as a diagnostic biomarker—and potentially a mechanistic contributor—to colorectal tumorigenesis via pathways distinct from the insulin-resistance and obesity-driven adipokine axis traditionally implicated in cancer risk [64].
Furthermore, a translational study highlighted the tumor-suppressive role of omentin-1 in colorectal cancer. In 229 primary CRC specimens, ITLN1 protein was absent or very low in 54.6 % of tumors, while it was consistently present in adjacent normal tissue. Data from The Cancer Genome Atlas (TCGA) showed that patients with high ITLN1 expression had a 49 % lower risk of death than those with low expression (HR = 0.51, p = 0.01). In a separate patient cohort, median survival dropped from 83.3 months in ITLN1-positive tumors to 64.1 months in ITLN1-negative ones (p = 0.001), and ITLN1 remained an independent prognostic factor after adjustment for other variables.
To investigate functionally, CRC cells were engineered to express normal amounts of omentin-1 and implanted into mice. These tumors grew significantly less—by ~50 %—compared to control cells, even though in vitro cell growth and migration were barely affected. This suggested that omentin-1 works mainly by modifying the tumor microenvironment rather than directly inhibiting tumor-cell proliferation. Flow cytometry and immunohistochemistry revealed that tumors expressing omentin-1 had 58 % fewer immunosuppressive myeloid-derived suppressor cells (MDSCs), 64 % fewer endothelial progenitor cells (EPCs), and double amount of tumor-infiltrating natural killer (NK) cells. The remaining MDSCs were also less suppressive: they produced 40 % less nitric oxide (NOS2) and 35 % less reactive oxygen species, allowing T-cells to proliferate more effectively. Mechanistically, omentin-1 activated PI3K/AKT/GSK3β signaling, stabilized the antioxidant protein Nrf2, and suppressed NF-κB activity, leading to sharp drops in IL-17D (↓68 %) and CXCL2 (↓72 %) secretion—two cytokines that normally attract MDSCs and EPCs. Blocking the PI3K pathway reversed these effects, confirming that omentin-1 acts through this route. Collectively, these data showed that approximately half of CRCs downregulate ITLN1 expression, potentially facilitating immune evasion and neoangiogenesis. Reconstitution of omentin-1 restores a more immunogenic and less vascularized tumor microenvironment, offering a promising therapeutic strategy aimed not at tumor cells per se, but at the stromal and immune components that enable tumor progression [65].
In a proteomic study of stage-IV colorectal cancer, omentin-1 emerged as one of the strongest biomarkers distinguishing long-term survivors from those with rapid disease progression. Kim et al. analyzed tumor samples to identify proteins differentially expressed between patients with good prognosis (survival > 5 years) and those with poor prognosis (death ≤ 25 months). Out of 1,044 quantified proteins, 175 showed ≥2-fold differences, and ITLN1 had the largest increase in the good-prognosis group, with a 41.4-fold rise in isotope-coded affinity tag signal. Follow-up validation confirmed these findings. Western blotting showed an 11.5-fold higher ITLN1 protein level in long-surviving tumors (p = 0.03), and an independent patient set reproduced this with a near-identical 11.6-fold difference (p < 0.05). When combined with four other top-performing proteins (FABP1, TPM2, TAGLN, and VCP) in a logistic regression model, ITLN1 helped generate a near-perfect classification (AUC = 1.00), though ITLN1 alone still yielded an AUC of 0.69—the highest individual contribution.
Histologically, strong cytoplasmic ITLN1 staining was seen in 33 % of good-prognosis tumors but in none of the poor-prognosis group, aligning with the biochemical data. Functional pathway analysis placed ITLN1 among a small cluster of proteins enriched in favorable cases, contrasting with the dominant up-regulation of cytoskeletal and migration-related pathways in aggressive tumors. Altogether, based on these results, the authors identified omentin-1 as a robust prognostic discriminator in advanced CRC, with expression levels that differ ~40-fold between survival extremes. Their data suggested that preserved ITLN1 may reflect or contribute to a less invasive tumor phenotype, and supported its use as a molecular marker for outcome prediction in metastatic colorectal cancer [66].
A comprehensive overview of key preclinical and clinical studies evaluating the role of omentin-1 (ITLN1) across gastrointestinal malignancies is provided in Table 1.
5. Directions and Research Needs
5.1. What Is Missing About Omentin’s Role in GI Cancers?
Authors should discuss the results and how they can be interpreted from the perspective of previous studies and of the working hypotheses. The findings and their implications should be discussed in the broadest context possible. Future research directions may also be highlighted.
Despite substantial advances in understanding the biological role and clinical implications of omentin in gastrointestinal cancers, several critical knowledge gaps remain, limiting its definitive application as a biomarker or therapeutic target.
In esophageal cancer, key gaps remain in understanding how omentin is altered and what functional role it plays. One priority is to gather data on circulating omentin-1 levels in esophageal adenocarcinoma (EAC) and squamous cell carcinoma (ESCC) patients, using patient cohorts and adjusting for obesity-related factors. Esophageal adenocarcinoma often arises in the context of visceral obesity, which itself lowers circulating omentin-1 levels, whereas ESCC patients are typically non-obese; thus, future studies should measure omentin-1 in EAC and ESCC while controlling for visceral adiposity and insulin resistance to detect true cancer-specific changes [43]. Another open question is the cellular distribution of ITLN1 in esophageal tissues. Applying transcriptomics or advanced immunohistochemistry along the esophageal epithelium (from normal mucosa through Barrett’s metaplasia to carcinoma) could map which cell types express ITLN1 and how this varies with disease stage [46]. A third gap is the lack of functional studies defining omentin’s role in esophageal tumor biology and how it affects tumor growth or modulates pathways like inflammation and apoptosis.
Regarding gastric cancer, future research priorities include the prospective validation of ITLN1 as a routine prognostic biomarker across diverse patient groups and treatment modalities. Additionally, serum-based studies with adequate power are needed to clarify whether circulating omentin-1 changes after curative gastrectomy or during adjuvant therapy. It is also important to investigate recombinant omentin or HNF4α activators in preclinical models, such as patient-derived gastric-cancer organoids and xenografts [49]. Finally, combination-therapy studies should test for synergy between omentin pathway activation and immune-checkpoint blockade, particularly in MSI-high disease.
For hepatocellular carcinoma, research efforts should focus on the prospective validation of ITLN1 as a prognostic biomarker across HBV-, HCV- and MASLD-related HCC. Pharmacokinetic and pharmacodynamic evaluations of recombinant omentin-1 in vivo models of liver cancer are also warranted to better characterize its therapeutic potential. Further research should explore combination strategies evaluating omentin pathway activation with current first-line agents, such as atezolizumab plus bevacizumab or lenvatinib, to test for additive or synergistic effects. Moreover, large cohorts are required for examine its role as serum biomarker for minimal-residual disease or early recurrence post-resection [51].
In pancreatic cancer, several gaps need to be addressed. The source of markedly elevated circulating omentin-1 in patients remains unknown, necessitating further investigation to determine whether this increase is driven by tumor-derived production, interactions with cancer-associated adipocytes, or a broader systemic inflammatory response. [53]. Mechanistic studies are absent, making it crucial to investigate whether omentin-1 plays a causal role in pancreatic cancer biology using appropriate in vitro and in vivo models. The utility of serum omentin-1 as a diagnostic biomarker also requires validation in large, prospective studies to confirm its accuracy in differentiating early-stage cancer from non-malignant conditions like chronic pancreatitis [54].
In the context of colorectal cancer, a key challenge is understanding why high blood levels of omentin-1 are linked to an increased cancer risk, while high levels of omentin-1 within the tumor itself predict better patient survival. This paradox suggests a clear difference between omentin circulating in the bloodstream -which might simply indicate conditions like obesity-and omentin acting locally within tumor. Resolving this contradiction likely involves studying the molecule TMEM207, which helps omentin fold properly and function effectively. It is believed that TMEM207 ensures omentin is correctly structured and released specifically in the tumor environment, allowing it to act as an anti-cancer agent. If this is true, circulating omentin might be inactive or simply a marker, while TMEM207-regulated omentin within the tumor microenvironment may exert active anti-tumor effects. The tumor-suppressive effects of locally active omentin-1 likely involve modulation of the tumor microenvironment, specifically through the reduction of immunosuppressive myeloid-derived suppressor cells (MDSCs) and the inhibition of angiogenesis via suppression of endothelial progenitor cells (EPCs). Understanding exactly how omentin-1 prevents these cells from promoting tumor growth is vital. Additionally, omentin-1 might strongly counteract cancer stem cells and support tumor-suppressive microRNAs, which are small molecules that help regulate cancer growth. These cellular-level effects might explain why local omentin-1 is protective and could be especially beneficial for patients at higher risk, such as those with diabetes. Overall, addressing the systemic (blood-based) versus local (tumor-based) paradox of omentin requires integrating molecular (TMEM207), environmental (MDSCs/EPCs), and cellular (cancer stem cells) mechanisms [58].
5.2. Proposals for Future Research Focusing on Translational and Clinical Studies
To effectively translate the current understanding of omentin into clinical practice, future research must focus on validating its biomarker potential, clarifying unresolved biological inconsistencies, and investigating its viability as a therapeutic target.
A key translational priority is validating omentin as a robust clinical biomarker through large-scale, prospective clinical studies designed to assess its prognostic and diagnostic value. For instance, prospective cohort trials examining ITLN1 expression in gastric cancer, hepatocellular carcinoma, and colorectal cancer using standardized immunohistochemical protocols are essential for ensuring reproducible and clinically meaningful results. Another priority is to investigate depot-specific expression of omentin in cancer, since visceral adipose tissue is the primary source of omentin, whereas subcutaneous adipose tissue expresses it at much lower levels. Whether differential expression between VAT and subcutaneous fat contributes to cancer development, progression, or cachexia remains unknown, and future studies should address this as a potential determinant of systemic versus tumor-localized omentin activity. Similarly, in malignancies like pancreatic cancer, where elevated circulating omentin-1 levels indicate possible diagnostic utility, carefully designed case-control studies are needed to assess its sensitivity and specificity, particularly when combined with established biomarkers such as CA19-9. Standardizing assay methodologies -including Enzyme-linked immunosorbent assay (ELISA) protocols for circulating omentin-1 quantification- is essential to ensure data consistency, reproducibility, and comparability across different studies. Importantly, future investigations should also elucidate the relationship between omentin-1 expression and surgical outcomes, including the impact of different resection strategies on circulating and tissue omentin-1 levels. Understanding whether surgical cytoreduction modifies omentin signaling -either through tumor removal or changes in systemic inflammation -may provide critical insights into its role as a dynamic biomarker.
Moreover, addressing existing biological paradoxes associated with omentin-1 remains a critical obstacle in clinical translation. A prominent challenge is the contradictory evidence surrounding omentin-1 in CRC, wherein elevated circulating levels appear associated with increased risk, while increased tumor-localized expression correlates with protective effects. Comprehensive cohort studies that concurrently measure circulating omentin-1, tumor-localized ITLN1 expression, its chaperone protein TMEM207, and detailed patient metabolic profiles are thus needed. These integrated analyses can elucidate the mechanistic distinctions underlying systemic versus localized omentin dynamics. Moreover, the underlying origin of increased circulating omentin-1 in pancreatic and colorectal cancers should be explored using biologically relevant in vitro and ex vivo models that simulate tumor–host interactions. Advanced single-cell RNA sequencing should be employed to map ITLN1 expression and its receptors within the tumor microenvironment, clarifying the cellular origin and targets of omentin-1 signaling across gastrointestinal malignancies.
Lastly, translational efforts prioritize the systematic investigation of omentin as a therapeutic agent. Given its established metabolic and immune-regulatory role, future research should explore potential synergistic effects between omentin signaling enhancement and existing therapeutic approaches, such as metabolic modulators (e.g., metformin) or immune checkpoint inhibitors (e.g., PD-1 blockade). In malignancies where omentin-1 demonstrates tumor-suppressive activity—such as gastric cancer, hepatocellular carcinoma, and specific subtypes of colorectal cancer—direct therapeutic approaches using recombinant omentin-1 or newly developed omentin-mimetic compounds should be evaluated in preclinical models. Additionally, targeting the TMEM207–omentin-1 signaling axis represents an innovative therapeutic strategy. Moreover, the TMEM207–omentin-1 signaling pathway offers a novel therapeutic target. Developing small molecules or biologics that enhance or stabilize the TMEM207–omentin interaction may promote endogenous production of functional omentin-1, thereby enhancing its anti-metastatic and tumor-suppressive effects. Such targeted therapeutic strategies hold substantial promise for personalized therapeutic approaches in gastrointestinal oncology.
6. Conclusions
Omentin is an adipokine with emerging relevance in gastrointestinal malignancies, demonstrating anti-inflammatory, antioxidant, and tumor-suppressive effects within the local tumor microenvironment. In colorectal, gastric, hepatic, and pancreatic cancer, its expression patterns and clinical associations suggest both diagnostic and prognostic potential, though findings remain context dependent. Further research is needed to clarify its tissue-specific roles and translate into effective biomarkers or targeted therapeutic strategies.
Author Contributions
Author Contributions: Conceptualization, A.M. and D.S.; methodology, A.M., M.F. and I.L.; software, A.K. and A.M.K.; validation, I.L., A.Ka. and D.T.; formal analysis, A.M. and A.K.; investigation, A.M., M.F., M.E.K. and A.M.; resources, K.G.T. and D.S.; data curation, A.M., A.K. and A.Ka.; writing—original draft preparation, A.M., A.K., M.E.K. and A.M.; writing—review and editing, I.L., M.F., A.Ka., D.T., K.G.T. and D.S.; visualization, A.M. and A.M.; supervision, I.L., K.G.T. and D.S.; project administration, I.L. and D.S.; funding acquisition, not applicable. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
None to declare.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Akt | Protein kinase B |
| AMPK | AMP-activated protein kinase |
| AUC | Area under the curve |
| BE | Barrett’s esophagus |
| CA19-9 | Carbohydrate antigen 19-9 |
| CEA | Carcinoembryonic antigen |
| CDX2 | Caudal-type homeobox 2 |
| CP | Chronic pancreatitis |
| CRC | Colorectal cancer |
| CSC | Cancer stem cell |
| CXCL2 | C-X-C motif chemokine ligand 2 |
| EAC | Esophageal adenocarcinoma |
| EPC | Endothelial progenitor cell |
| ERK | Extracellular signal–regulated kinase |
| ESCC | Esophageal squamous cell carcinoma |
| HCC | Hepatocellular carcinoma |
| HNF4α | Hepatocyte nuclear factor 4 alpha |
| HOMA-IR | Homeostatic model assessment of insulin resistance |
| HR | Hazard ratio |
| IHC | Immunohistochemistry |
| ITLN1 | Intelectin-1 (gene encoding omentin-1) |
| JNK | c-Jun N-terminal kinase |
| LF | Lactoferrin |
| MAPK | Mitogen-activated protein kinase |
| MASLD | Metabolic-associated steatotic liver disease |
| MASH | Metabolic-associated steatohepatitis |
| MDSC | Myeloid-derived suppressor cell |
| miRNA | MicroRNA |
| NF-κB | Nuclear factor kappa B |
| NK cell | Natural killer cell |
| Nrf2 | Nuclear factor erythroid 2–related factor 2 |
| OR | Odds ratio |
| PA | Pancreatic adenocarcinoma |
| PDAC | Pancreatic ductal adenocarcinoma |
| PI3K | Phosphoinositide 3-kinase |
| PPAR-δ | Peroxisome proliferator-activated receptor delta |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| RFS | Recurrence-free survival |
| ROC | Receiver operating characteristic |
| ROS | Reactive oxygen species |
| SIRT1 | Sirtuin 1 |
| SNP | Single-nucleotide polymorphism |
| TCGA | The Cancer Genome Atlas |
| TMEM207 | Transmembrane protein 207 |
| TNF-α | Tumor necrosis factor alpha |
| TNM | Tumor–node–metastasis (staging) |
| TOP/FOP | TCF reporter constructs for Wnt/β-catenin signaling |
| VAT | Visceral adipose tissue |
| VCAM-1 | Vascular cell adhesion molecule 1 |
| VEGF | Vascular endothelial growth factor |
| VLDL | Very low–density lipoprotein |
References
- Lu, L.; Mullins, C.S.; Schafmayer, C.; Zeißig, S.; Linnebacher, M. A Global Assessment of Recent Trends in Gastrointestinal Cancer and Lifestyle-Associated Risk Factors. Cancer Communications 2021, 41, 1137–1151. [Google Scholar] [CrossRef]
- Tan, B.K.; Adya, R.; Randeva, H.S. Omentin: A Novel Link Between Inflammation, Diabesity, and Cardiovascular Disease. Trends in Cardiovascular Medicine 2010, 20, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-Y.; Chan, L.; Zhou, S.-W. Omentin: Linking Metabolic Syndrome and Cardiovascular Disease. Curr Vasc Pharmacol 2014, 12, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhu, L.; Zheng, M.; Fan, C.; Li, Y.; Zhang, D.; He, Y.; Yang, H. Changes of Serum Omentin-1 Levels in Normal Subjects, Type 2 Diabetes and Type 2 Diabetes with Overweight and Obesity in Chinese Adults. Ann Endocrinol (Paris) 2014, 75, 171–175. [Google Scholar] [CrossRef] [PubMed]
- de Souza Batista, C.M.; Yang, R.-Z.; Lee, M.-J.; Glynn, N.M.; Yu, D.-Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin Plasma Levels and Gene Expression Are Decreased in Obesity. Diabetes 2007, 56, 1655–1661. [Google Scholar] [CrossRef]
- Rothermel, J.; Lass, N.; Barth, A.; Reinehr, T. Link between Omentin-1, Obesity and Insulin Resistance in Children: Findings from a Longitudinal Intervention Study. Pediatr Obes 2020, 15, e12605. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Catalán, V.; Ortega, F.; Gómez-Ambrosi, J.; Ricart, W.; Frühbeck, G.; Fernández-Real, J.M. Circulating Omentin Concentration Increases after Weight Loss. Nutr Metab (Lond) 2010, 7, 27. [Google Scholar] [CrossRef]
- Saremi, A.; Asghari, M.; Ghorbani, A. Effects of Aerobic Training on Serum Omentin-1 and Cardiometabolic Risk Factors in Overweight and Obese Men. J Sports Sci 2010, 28, 993–998. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. [Google Scholar] [CrossRef]
- Paval, D.R.; Di Virgilio, T.G.; Skipworth, R.J.E.; Gallagher, I.J. The Emerging Role of Intelectin-1 in Cancer. Front Oncol 2022, 12, 767859. [Google Scholar] [CrossRef]
- Yang, R.-Z.; Lee, M.-J.; Hu, H.; Pray, J.; Wu, H.-B.; Hansen, B.C.; Shuldiner, A.R.; Fried, S.K.; McLenithan, J.C.; Gong, D.-W. Identification of Omentin as a Novel Depot-Specific Adipokine in Human Adipose Tissue: Possible Role in Modulating Insulin Action. Am J Physiol Endocrinol Metab 2006, 290, E1253–1261. [Google Scholar] [CrossRef]
- Dec, P.; Poniewierska-Baran, A.; Modrzejewski, A.; Pawlik, A. The Role of Omentin-1 in Cancers Development and Progression. Cancers 2023, 15, 3797. [Google Scholar] [CrossRef]
- Fain, J.N.; Sacks, H.S.; Buehrer, B.; Bahouth, S.W.; Garrett, E.; Wolf, R.Y.; Carter, R.A.; Tichansky, D.S.; Madan, A.K. Identification of Omentin mRNA in Human Epicardial Adipose Tissue: Comparison to Omentin in Subcutaneous, Internal Mammary Artery Periadventitial and Visceral Abdominal Depots. Int J Obes (Lond) 2008, 32, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Sena, C.M. Omentin: A Key Player in Glucose Homeostasis, Atheroprotection, and Anti-Inflammatory Potential for Cardiovascular Health in Obesity and Diabetes. Biomedicines 2024, 12, 284. [Google Scholar] [CrossRef]
- Washimi, K.; Yokose, T.; Yamashita, M.; Kageyama, T.; Suzuki, K.; Yoshihara, M.; Miyagi, Y.; Hayashi, H.; Tsuji, S. Specific Expression of Human Intelectin-1 in Malignant Pleural Mesothelioma and Gastrointestinal Goblet Cells. PLoS One 2012, 7, e39889. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, B.; Hao, C.; Huang, X.; Li, X.; Huang, Y.; Luo, Z. Omentin-A Novel Adipokine in Respiratory Diseases. International Journal of Molecular Sciences 2018, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Czechowski, P.; Hagemann, T.; Ghosh, A.; Sun, W.; Dong, H.; Noé, F.; Niersmann, C.; Reinisch, I.; Wolfrum, C.; Herder, C.; et al. Expression of Intelectin-1, Also Known as Omentin-1, Is Related to Clinical Phenotypes Such as Overweight, Obesity, Insulin Resistance, and Changes after Bariatric Surgery. Sci Rep 2024, 14, 22286. [Google Scholar] [CrossRef] [PubMed]
- Peña-Cano, M.I.; Valencia-Ortega, J.; Morales-Ávila, E.; Díaz-Velázquez, M.F.; Gómez-Díaz, R.; Saucedo, R. Omentin-1 and Its Relationship with Inflammatory Factors in Maternal Plasma and Visceral Adipose Tissue of Women with Gestational Diabetes Mellitus. J Endocrinol Invest 2022, 45, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Nonnecke, E.B.; Castillo, P.A.; Dugan, A.E.; Almalki, F.; Underwood, M.A.; De La Motte, C.A.; Yuan, W.; Lu, W.; Shen, B.; Johansson, M.E.V.; et al. Human Intelectin-1 (ITLN1) Genetic Variation and Intestinal Expression. Sci Rep 2021, 11, 12889. [Google Scholar] [CrossRef]
- Andresen, S.; Fantone, K.; Chapla, D.; Rada, B.; Moremen, K.W.; Pierce, M.; Szymanski, C.M. Human Intelectin-1 Promotes Cellular Attachment and Neutrophil Killing of Streptococcus Pneumoniae in a Serotype-Dependent Manner. Infect Immun 90, e00682-21. [CrossRef]
- Yamawaki, H.; Kuramoto, J.; Kameshima, S.; Usui, T.; Okada, M.; Hara, Y. Omentin, a Novel Adipocytokine Inhibits TNF-Induced Vascular Inflammation in Human Endothelial Cells. Biochem Biophys Res Commun 2011, 408, 339–343. [Google Scholar] [CrossRef]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose Tissue-Derived Omentin-1 Function and Regulation. In Comprehensive Physiology; Terjung, R., Ed.; Wiley, 2017; pp. 765–781 ISBN 978-0-470-65071-4.
- Hiramatsu-Ito, M.; Shibata, R.; Ohashi, K.; Uemura, Y.; Kanemura, N.; Kambara, T.; Enomoto, T.; Yuasa, D.; Matsuo, K.; Ito, M.; et al. Omentin Attenuates Atherosclerotic Lesion Formation in Apolipoprotein E-Deficient Mice. Cardiovasc Res 2016, 110, 107–117. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Z.; Qian, G.; Zhou, J. Omentin-1 Attenuates Adipose Tissue Inflammation via Restoration of TXNIP/NLRP3 Signaling in High-Fat Diet-Induced Obese Mice. Fundam Clin Pharmacol 2020, 34, 721–735. [Google Scholar] [CrossRef]
- Sigal, M.; Reinés, M. del M.; Müllerke, S.; Fischer, C.; Kapalczynska, M.; Berger, H.; Bakker, E.R.M.; Mollenkopf, H.-J.; Rothenberg, M.E.; Wiedenmann, B.; et al. R-Spondin-3 Induces Secretory, Antimicrobial Lgr5+ Cells in the Stomach. Nat Cell Biol 2019, 21, 812–823. [Google Scholar] [CrossRef]
- Kazama, K.; Usui, T.; Okada, M.; Hara, Y.; Yamawaki, H. Omentin Plays an Anti-Inflammatory Role through Inhibition of TNF-α-Induced Superoxide Production in Vascular Smooth Muscle Cells. Eur J Pharmacol 2012, 686, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Queiroz, M.; Azul, L.; Seiça, R.; Sena, C.M. Omentin: A Novel Therapeutic Approach for the Treatment of Endothelial Dysfunction in Type 2 Diabetes. Free Radic Biol Med 2021, 162, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Fang, S.; Liu, X.; Li, J.; Wang, X.; Cui, J.; Chen, T.; Li, Z.; Yang, F.; Tian, J.; et al. Omentin-1 Protects against High Glucose-Induced Endothelial Dysfunction via the AMPK/PPARδ Signaling Pathway. Biochem Pharmacol 2020, 174, 113830. [Google Scholar] [CrossRef] [PubMed]
- Tao, M.; Yan, W.; Chen, C.; Tang, M.; Zhao, X.; Feng, Q.; Fei, X.; Fu, Y. Omentin-1 Ameliorates Experimental Inflammatory Bowel Disease via Nrf2 Activation and Redox Regulation. Life Sci 2023, 328, 121847. [Google Scholar] [CrossRef] [PubMed]
- Binti Kamaruddin, N.A.; Fong, L.Y.; Tan, J.J.; Abdullah, M.N.H.; Singh Cheema, M.; Bin Yakop, F.; Yong, Y.K. Cytoprotective Role of Omentin Against Oxidative Stress-Induced Vascular Endothelial Cells Injury. Molecules 2020, 25, 2534. [Google Scholar] [CrossRef]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of Anti-Inflammatory Adipokines in Obesity-Related Diseases. Trends Endocrinol Metab 2014, 25, 348–355. [Google Scholar] [CrossRef]
- Zhong, X.; Li, X.; Liu, F.; Tan, H.; Shang, D. Omentin Inhibits TNF-α-Induced Expression of Adhesion Molecules in Endothelial Cells via ERK/NF-κB Pathway. Biochem Biophys Res Commun 2012, 425, 401–406. [Google Scholar] [CrossRef]
- Watanabe, K.; Watanabe, R.; Konii, H.; Shirai, R.; Sato, K.; Matsuyama, T.-A.; Ishibashi-Ueda, H.; Koba, S.; Kobayashi, Y.; Hirano, T.; et al. Counteractive Effects of Omentin-1 against Atherogenesis†. Cardiovasc Res 2016, 110, 118–128. [Google Scholar] [CrossRef]
- Shibata, R.; Ouchi, N.; Takahashi, R.; Terakura, Y.; Ohashi, K.; Ikeda, N.; Higuchi, A.; Terasaki, H.; Kihara, S.; Murohara, T. Omentin as a Novel Biomarker of Metabolic Risk Factors. Diabetol Metab Syndr 2012, 4, 37. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, S.; Ke, Y.; Li, Q.; Shen, C.; Ruan, Y.; Wu, K.; Hu, J.; Liu, S. Association of Circulating Omentin Level and Metabolic-Associated Fatty Liver Disease: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2023, 14. [Google Scholar] [CrossRef]
- Salvoza, N.; Giraudi, P.; Gazzin, S.; Bonazza, D.; Palmisano, S.; de Manzini, N.; Zanconati, F.; Raseni, A.; Sirianni, F.; Tiribelli, C.; et al. The Potential Role of Omentin-1 in Obesity-Related Metabolic Dysfunction-Associated Steatotic Liver Disease: Evidence from Translational Studies. Journal of Translational Medicine 2023, 21, 906. [Google Scholar] [CrossRef] [PubMed]
- As Habi, A.; Sadeghi, M.; Arab, A.; Hajianfar, H. The Association between Omentin and Diabetes: A Systematic Review and Meta-Analysis of Observational Studies. Diabetes Metab Syndr Obes 2019, 12, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Wang, X.; Bu, P. Omentin-1 Is Associated with Carotid Atherosclerosis in Patients with Metabolic Syndrome. Diabetes Res Clin Pract 2011, 93, 21–25. [Google Scholar] [CrossRef]
- Cetin Sanlialp, S.; Nar, G.; Nar, R. Relationship between Circulating Serum Omentin-1 Levels and Nascent Metabolic Syndrome in Patients with Hypertension. J Investig Med 2022, 70, 780–785. [Google Scholar] [CrossRef]
- Varona, J.F.; Ortiz-Regalón, R.; Sánchez-Vera, I.; López-Melgar, B.; García-Durango, C.; Castellano Vázquez, J.M.; Solís, J.; Fernández-Friera, L.; Vidal-Vanaclocha, F. Soluble ICAM 1 and VCAM 1 Blood Levels Alert on Subclinical Atherosclerosis in Non Smokers with Asymptomatic Metabolic Syndrome. Arch Med Res 2019, 50, 20–28. [Google Scholar] [CrossRef]
- Huang, Z.; Luo, L.; Xiao, Z.; Xiong, M.; Wen, Z. Omentin-1 Mitigates Non-Alcoholic Fatty Liver Disease by Preserving Autophagy through AMPKα/mTOR Signaling Pathway. Sci Rep 2024, 14, 31464. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.; Dreczkowski, G.; Ramage, M.I.; Wigmore, S.J.; Gallagher, I.J.; Skipworth, R.J.E. Adipose Depot Gene Expression and Intelectin-1 in the Metabolic Response to Cancer and Cachexia. J Cachexia Sarcopenia Muscle 2020, 11, 1141–1153. [Google Scholar] [CrossRef]
- Huang, F.-L.; Yu, S.-J. Esophageal Cancer: Risk Factors, Genetic Association, and Treatment. Asian Journal of Surgery 2018, 41, 210–215. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, Y.; Zhang, Z.; Zhang, Z.; Zhang, G.; Wang, F.; Che, Y.; Fang, L.; Zhang, Y.; Sun, N.; et al. Identification of a Prognostic Immune Signature for Esophageal Squamous Cell Carcinoma to Predict Survival and Inflammatory Landscapes. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Zhu, L.; Liang, F.; Han, X.; Ye, B.; Xue, L. Machine Learning-Based Glycolipid Metabolism Gene Signature Predicts Prognosis and Immune Landscape in Oesophageal Squamous Cell Carcinoma. J Cell Mol Med 2025, 29, e70434. [Google Scholar] [CrossRef] [PubMed]
- Busslinger, G.A.; de Barbanson, B.; Oka, R.; Weusten, B.L.A.; de Maat, M.; van Hillegersberg, R.; Brosens, L.A.A.; van Boxtel, R.; van Oudenaarden, A.; Clevers, H. Molecular Characterization of Barrett’s Esophagus at Single-Cell Resolution. Proceedings of the National Academy of Sciences 2021, 118, e2113061118. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, S.; Jonas, C.; Wenzel, M.C.; Velazquez Camacho, O.; Arolt, C.; Zhao, Y.; Büttner, R.; Quaas, A.; Plum, P.S.; Hillmer, A.M. Genomic and Transcriptomic Characteristics of Esophageal Adenocarcinoma. Cancers (Basel) 2021, 13, 4300. [Google Scholar] [CrossRef]
- Zheng, L.; Weng, M.; Qi, M.; Qi, T.; Tong, L.; Hou, X.; Tong, Q. Aberrant Expression of Intelectin-1 in Gastric Cancer: Its Relationship with Clinicopathological Features and Prognosis. J Cancer Res Clin Oncol 2012, 138, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhao, X.; Xiao, Y.; Mei, H.; Pu, J.; Xiang, X.; Jiao, W.; Song, H.; Qu, H.; Huang, K.; et al. Intelectin 1 Suppresses Tumor Progression and Is Associated with Improved Survival in Gastric Cancer. Oncotarget 2015, 6, 16168–16182. [Google Scholar] [CrossRef]
- Zhang, Y.-Y.; Zhou, L.-M. Omentin-1, a New Adipokine, Promotes Apoptosis through Regulating Sirt1-Dependent P53 Deacetylation in Hepatocellular Carcinoma Cells. European Journal of Pharmacology 2013, 698, 137–144. [Google Scholar] [CrossRef]
- Li, J.; Tao, H.-S.; Yuan, T.; Huang, Z.-Y.; Zhang, E.-L. Intelectin-1 Is a Novel Prognostic Biomarker for Hepatocellular Carcinoma. Medicine (Baltimore) 2023, 102, e36474. [Google Scholar] [CrossRef] [PubMed]
- Cidem, A.; Chang, G.R.-L.; Yen, C.-C.; Chen, M.-S.; Yang, S.-H.; Chen, C.-M. Lactoferrin Targeting INTL1 Receptor Inhibits Hepatocellular Carcinoma Progression via Apoptosis and Cell Cycle Signaling Pathways. Sci Rep 2024, 14, 31210. [Google Scholar] [CrossRef]
- Karabulut, S.; Afsar, C.U.; Karabulut, M.; Alis, H.; Bozkurt, M.A.; Aydogan, F.; Serilmez, M.; Tas, F. Clinical Significance of Serum Omentin-1 Levels in Patients with Pancreatic Adenocarcinoma. BBA Clin 2016, 6, 138–142. [Google Scholar] [CrossRef]
- Kiczmer, P.; Szydło, B.; Seńkowska, A.P.; Jopek, J.; Wiewiora, M.; Piecuch, J.; Ostrowska, Z.; Świętochowska, E. Serum Omentin-1 and Chemerin Concentrations in Pancreatic Cancer and Chronic Pancreatitis. Folia medica Cracoviensia 2018. [Google Scholar] [CrossRef]
- Aleksandrova, K.; di Giuseppe, R.; Isermann, B.; Biemann, R.; Schulze, M.; Wittenbecher, C.; Fritsche, A.; Lehmann, R.; Menzel, J.; Weikert, C.; et al. Circulating Omentin as a Novel Biomarker for Colorectal Cancer Risk: Data from the EPIC–Potsdam Cohort Study. Cancer Research 2016, 76, 3862–3871. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Y.; Deng, L.; Wang, Y.; Li, Y.; Chen, M. The Association between Chinese Patients’ Elevated Omentin-1 Levels, Their Clinicopathological Features, and the Risk of Colorectal Cancer. Int J Clin Exp Pathol 2019, 12, 2264–2274. [Google Scholar]
- Feng, Z.; Sun, H.; Liu, P.; Shi, W.; Han, W.; Ma, L. Analysis of the Expression of Plasma Omentin-1 Level in Colorectal Cancer and Its Correlation with Prognosis. Transl Cancer Res 2020, 9, 6479–6486. [Google Scholar] [CrossRef]
- Maeda, K.; Saigo, C.; Kito, Y.; Sakuratani, T.; Yoshida, K.; Takeuchi, T. Expression of TMEM207 in Colorectal Cancer: Relation between TMEM207 and Intelectin-1. J Cancer 2016, 7, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, K.; Maeda, K.; Saigo, C.; Kito, Y.; Yoshida, K.; Takeuchi, T. Adiponectin and Intelectin-1: Important Adipokine Players in Obesity-Related Colorectal Carcinogenesis. International Journal of Molecular Sciences 2017, 18, 866. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, X.; Chen, M. Autocrine Action of Adipokine Omentin-1 in the SW480 Colon Cancer Cell Line. Oncol Lett 2020, 19, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Wan, L.; Zhang, Q.; Chen, M.; Zhao, X. The Effect of Omentin-1 on the Proliferation and Apoptosis of Colon Cancer Stem Cells and the Potential Mechanism. J BUON 2019, 24, 91–98. [Google Scholar]
- Uyeturk, U.; Alcelik, A.; Aktas, G.; Tekce, B.K. Post-Treatment Plasma Omentin Levels in Patients with Stage III Colon Carcinoma. J BUON 2014, 19, 681–685. [Google Scholar]
- Zhang, Y.; Zhao, X.; Li, Y.; Wang, Y.; Chen, M. Association between the Omentin-1 Gene Rs2274907 A>T Polymorphism and Colorectal Cancer in the Chinese Han Population: A Case-Control Study. J Int Med Res 2021, 49, 03000605211006522. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, M.S.; Dashti, H.; Akbarzadeh, S.; Assadi, M.; Aminian, A.; Keramati, M.R.; Nabipour, I. Circulating Levels of Novel Adipocytokines in Patients with Colorectal Cancer. Cytokine 2013, 62, 81–85. [Google Scholar] [CrossRef]
- Chen, L.; Jin, X.-H.; Luo, J.; Duan, J.-L.; Cai, M.-Y.; Chen, J.-W.; Feng, Z.-H.; Guo, A.M.; Wang, F.-W.; Xie, D. ITLN1 Inhibits Tumor Neovascularization and Myeloid Derived Suppressor Cells Accumulation in Colorectal Carcinoma. Oncogene 2021, 40, 5925–5937. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Kang, U.-B.; Lee, H.; Jung, J.-H.; Lee, S.-T.; Yu, M.-H.; Kim, H.; Lee, C. Profiling of Differentially Expressed Proteins in Stage IV Colorectal Cancers with Good and Poor Outcomes. Journal of Proteomics 2012, 75, 2983–2997. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
Schematic representation of the role of omentin in metabolic disorders. Akt, protein kinase B; TNF-α, tumor necrosis factor alpha; IL-1, interleukin-1; NF-κB, nuclear factor kappa B; HDL, high-density lipoprotein; VLDL, very low-density lipoprotein; AMPK, AMP-activated protein kinase.
Figure 2.
Schematic representation of the role of omentin in metabolic disorders. Akt, protein kinase B; TNF-α, tumor necrosis factor alpha; IL-1, interleukin-1; NF-κB, nuclear factor kappa B; HDL, high-density lipoprotein; VLDL, very low-density lipoprotein; AMPK, AMP-activated protein kinase.
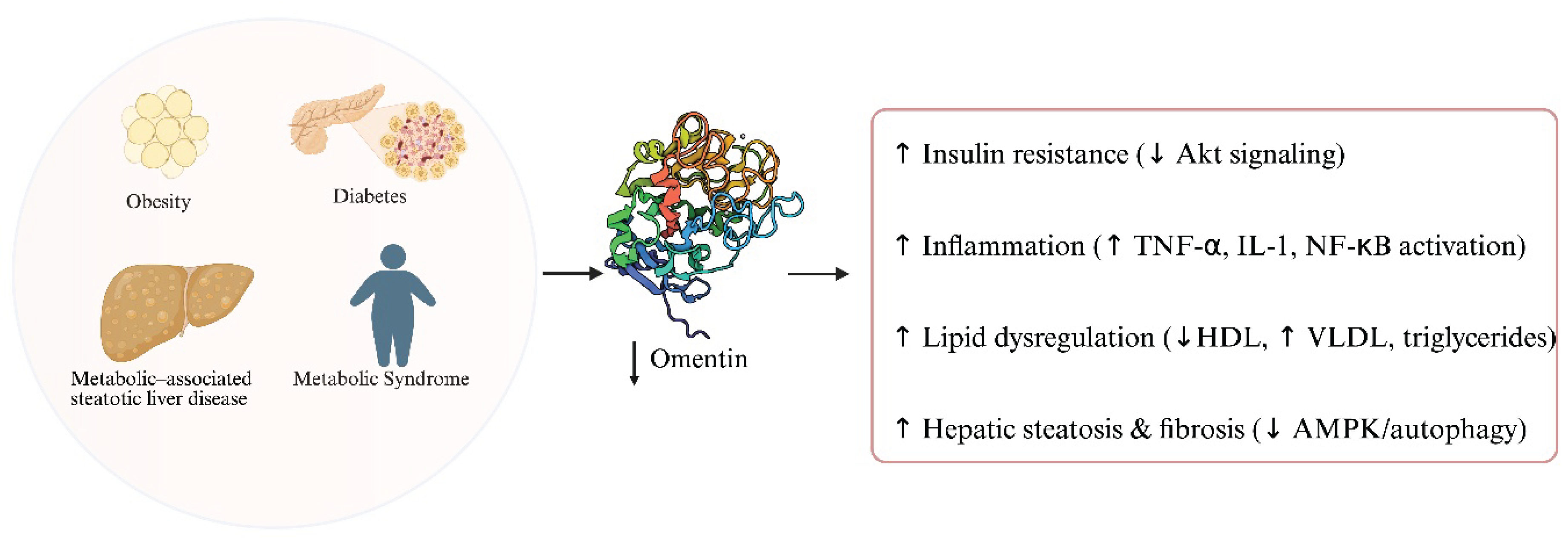

Table 1.
Structured summary of published studies investigating omentin in gastrointestinal cancers including experimental model used, key findings, proposed biological role and relevant clinical implications. Abbreviations: CRC: Colorectal cancer; ESCC: Esophageal squamous cell carcinoma; EAC: Esophageal adenocarcinoma; BE: Barrett’s esophagus; ITLN1: Intelectin-1; OS: Overall survival; RFS: Recurrence-free survival; qRT-PCR: Quantitative real-time polymerase chain reaction; IHC: Immunohistochemistry; ELISA: Enzyme-linked immunosorbent assay; PA: Pancreatic adenocarcinoma; PDAC: Pancreatic ductal adenocarcinoma; CP: Chronic pancreatitis; AUC: Area under the curve; CSC: Cancer stem cell; CD133+: Stem cell marker; HNF4α: Hepatocyte nuclear factor 4 alpha; NF-κB: Nuclear factor kappa-light-chain-enhancer of activated B cells; PI3K: Phosphoinositide 3-kinase; Akt: Protein kinase B; TOP/FOP: TCF reporter constructs for Wnt signaling; miRNA: MicroRNA; HR: Hazard ratio; OR: Odds ratio; SNP: Single-nucleotide polymorphism; TCGA: The Cancer Genome Atlas; WB: Western blot; TMA: Tissue microarray; TMEM207: Transmembrane protein 207; EPC: Endothelial progenitor cell; MDSC: Myeloid-derived suppressor cell; NOS2: Nitric oxide synthase 2; ROS: Reactive oxygen species; CXCL2, IL-17D: Cytokines involved in immune recruitment.
Table 1.
Structured summary of published studies investigating omentin in gastrointestinal cancers including experimental model used, key findings, proposed biological role and relevant clinical implications. Abbreviations: CRC: Colorectal cancer; ESCC: Esophageal squamous cell carcinoma; EAC: Esophageal adenocarcinoma; BE: Barrett’s esophagus; ITLN1: Intelectin-1; OS: Overall survival; RFS: Recurrence-free survival; qRT-PCR: Quantitative real-time polymerase chain reaction; IHC: Immunohistochemistry; ELISA: Enzyme-linked immunosorbent assay; PA: Pancreatic adenocarcinoma; PDAC: Pancreatic ductal adenocarcinoma; CP: Chronic pancreatitis; AUC: Area under the curve; CSC: Cancer stem cell; CD133+: Stem cell marker; HNF4α: Hepatocyte nuclear factor 4 alpha; NF-κB: Nuclear factor kappa-light-chain-enhancer of activated B cells; PI3K: Phosphoinositide 3-kinase; Akt: Protein kinase B; TOP/FOP: TCF reporter constructs for Wnt signaling; miRNA: MicroRNA; HR: Hazard ratio; OR: Odds ratio; SNP: Single-nucleotide polymorphism; TCGA: The Cancer Genome Atlas; WB: Western blot; TMA: Tissue microarray; TMEM207: Transmembrane protein 207; EPC: Endothelial progenitor cell; MDSC: Myeloid-derived suppressor cell; NOS2: Nitric oxide synthase 2; ROS: Reactive oxygen species; CXCL2, IL-17D: Cytokines involved in immune recruitment.
| Authors and Year | Cancer Type | Model | Findings | Role | Clinical Implication |
| Zhang et al., 2020 | ESCC | ESCC (microarray + qRT-PCR) | ↑ ITLN1 linked to better OS/RFS; protective gene in 6-gene immune signature | Protective, prognostic | Favorable immune profile and improved survival |
| Liang et al., 2021 | ESCC | ESCC tumor and adjacent normal tissue | ↓ ITLN1 in tumors; associated with better immune contexture and lower immune risk score | Immune-modulating, prognostic | Immune biomarker; favorable prognosis |
| Busslinger et al., 2021 | EAC | Barrett’s esophagus and dysplasia/EAC tissue | ITLN1 expressed in BE epithelium; unchanged in progression to dysplasia or adenocarcinoma | Marker of metaplasia | Diagnostic utility in Barrett’s; no role in malignant transformation |
| Hoppe et al., 2021 | EAC | Review of Barrett’s esophagus and EAC data | ITLN1 marks goblet-like BE cells; no expression change in EAC | Marker of cell identity | Limited diagnostic/prognostic value in EAC |
| Zheng et al., 2012 | Gastric | qPCR, IHC, survival analysis on tumors and normal tissue | ITLN1 mRNA 8× higher in tumors; associated with CDX2 expression, favorable histology, lower stage, improved survival (HR = 2.25 for ITLN1−) | Tumor suppressor downstream of CDX2 | Prognostic marker and therapeutic target for intestinal-type GC |
| Li et al., 2015 | Gastric | In vitro (SGC-7901, AGS), in vivo (xenografts) | ITLN1 inhibits PI3K/NF-κB, restores HNF4α, suppresses β-catenin; reduces invasion/metastasis; positively correlates with OS (R = 0.82) | Tumor-suppressive adipokine | Dual biomarker with HNF4α; therapeutic target |
| Zhang et al., 2013 | HCC | HepG2, HuH-7 cell lines | Omentin-1 stabilizes p53 via SIRT1 inhibition; activates caspase-mediated apoptosis via JNK | Tumor suppressor | Potential therapeutic via p53 reactivation |
| Li et al., 2023 | hCC | 149 HCC patients (tissue analysis) | ↓ ITLN1 in 78% of tumors; associated with aggressive features and worse survival (HR ≈ 0.5) | Prognostic biomarker | Risk stratification tool post-resection |
| Cidem et al., 2024 | HCC | HepG2, Hep3B, SK-Hep1 + orthotopic mice | LF requires omentin-1 for uptake; triggers apoptosis, cell-cycle arrest, inhibits angiogenesis | Therapeutic co-factor | Predictive biomarker and therapeutic enhancer for LF-based treatment |
| Karabulut et al., 2016 | Pancreatic | 33 PA patients vs 30 controls (ELISA) | ↑ Omentin-1 (~9.6 vs ~1.6 ng/mL); correlates with tumor size, not OS or chemo response | Diagnostic marker | Potential biomarker for detection, not for prognosis |
| Kiczmer et al., 2018 | Pancreatic | 25 PDAC, 10 CP, 36 controls (serum) | ↑ Omentin-1 in PDAC vs controls; AUC 0.73; no correlation with TNM stage or BMI | Diagnostic adjunct | Support diagnosis; limited prognostic use |
| Aleksandrova et al., 2016 | CRC | Prospective cohort (n=2,295 subcohort + 251 CRC cases) | High baseline omentin-1 associated with increased CRC risk, especially in non-obese | Risk biomarker | Stratify risk, particularly in lean individuals |
| Zhao et al., 2019 | CRC | Case-control (358 CRC vs 286 controls) | ↑ Omentin-1 in CRC; associated with advanced stage (pT3/4, TNM III–IV); AUC 0.88 | Diagnostic and staging marker | Non-invasive diagnostic tool with good sensitivity |
| Feng et al., 2020 | CRC | Prospective cohort (319 CRC vs 300 controls) | Elevated pre-op omentin-1 predicts recurrence (HR 3.3) and mortality (HR 2.1) | Prognostic biomarker | Post-operative monitoring and treatment |
| Kawashima et al., 2016 | CRC | Review of mechanistic and IHC data | TMEM207 stabilizes omentin-1; loss linked to aggressive, non-mucinous CRC | Facilitator of tumor suppression | TMEM207–omentin axis: prognostic and therapeutic value |
| Maeda et al., 2016 | CRC | IHC + in vitro (siRNA) | TMEM207 loss reduces omentin secretion; correlates with nodal metastasis | Post-translational regulator | Potential target for therapy and prognosis |
| Zhang et al., 2020 | CRC | CRC tissue + SW480 cells | Tumors overexpress omentin-1; secreted in autocrine/paracrine fashion | Context-dependent role | Targeting tumor-derived omentin alters CRC progression |
| Ji et al., 2019 | CRC | CD133⁺ SW480 cells | Omentin-1 inhibits PI3K/Akt; reduces stemness and promotes apoptosis | Cancer stem cell suppressor | Adjuvant therapy targeting cancer stem cells |
| Uyeturk et al., 2014 | CRC | Stage III post-FOLFOX patients (n=45) | ↑ Omentin-1 after chemo; not correlated with metabolic parameters | Reactive or compensatory marker | Host response to therapy |
| Zhang et al., 2021 | CRC | Genetic association study (SNP) | Variant not associated with CRC overall; modifies risk in obesity | Gene–environment interaction | Stratify risk in obese patients based on genotype |
| Fazeli et al., 2013 | CRC | 39 CRC vs 30 controls | ≈20× higher plasma omentin-1 in CRC; independent of BMI or stage | Carcinogenesis biomarker | Early detection |
| Chen et al., 2021 | CRC | TCGA + IHC (n=229) | Low ITLN1 linked to worse OS; re-expression reduces EPCs and MDSCs | Microenvironment modulator | Therapeutic restoration enhances anti-tumor immunity |
| Kim et al., 2012 | CRC | Stage IV CRC proteomics | ITLN1 41× higher in long-term survivors; validated by WB/IHC | Prognostic biomarker | Survival indicator; guides treatment intensity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



