Submitted:
30 August 2025
Posted:
02 September 2025
You are already at the latest version
Abstract
Metabolic engineering of mesenchymal stem/stromal cells (MSCs) represents a compelling frontier for advanced cellular therapies, enabling the precise tuning of their biological outputs. This feature paper examines the critical role of engineered culture microenvironments, specifically 3D platforms, hypoxic preconditioning, and other priming approaches, as synthetic biology strategies to guide and optimize MSC metabolic states for desired functional outcomes. We show that these non-genetic approaches significantly enhance MSC survival, immunomodulatory capacity, and regenerative potential by shifting their metabolism toward a more glycolytic phenotype. Furthermore, we propose a new paradigm of "designer" MSCs, which are programmed with synthetic circuits to sense and respond to the physiological cues of an injured microenvironment. This approach promises to transform regenerative medicine from an inconsistent field into a precise, predictable, and highly effective therapeutic discipline.
Keywords:
mesenchymal stem/stromal cells
; metabolism
; reprogramming
; glycolysis
; culture conditions
; 3D culture
; hypoxia
; preconditioning
; cell survival
; therapeutic potential
1. Introduction: The Metabolic Imperative in Mesenchymal Stem Cell Therapy and Research
Despite their immense therapeutic promise in regenerative medicine and their critical role as models in biomedical research, the full potential of mesenchymal stem/ stromal cells (MSCs) often remains elusive due to challenges in controlling their behavior. Could precise metabolic engineering, guided by tailored culture environments, be the missing link to consistently robust and predictable cellular therapies and more accurate research models?
Since their initial discovery by Friedenstein and colleagues [1] as plastic-adherent, fibroblast-like cells within bone marrow, MSCs have captivated the scientific community, emerging as one of the most intensely investigated therapeutic agents in regenerative medicine. The excitement surrounding these cells stems from a unique constellation of biological properties that position them as powerful, multifaceted "living drugs". Mesenchymal stem cells are multipotent stromal cells characterized by their adherence to plastic, specific surface marker expression (e.g., CD73, CD90, CD105), and capacity for self-renewal and differentiation into various mesodermal lineages, including osteocytes, chondrocytes, and adipocytes [2,3]. While MSCs are multipotent, and their differentiation potential is still important, especially in specific tissue engineering applications, the therapeutic potential of MSCs is now widely attributed more to their non-progenitor regulatory properties. The true "magic" of MSCs lies predominantly in their profound paracrine activity and their dynamic interactions with the host immune system. By secreting a rich cocktail of trophic factors, cytokines, and extracellular vesicles, MSCs orchestrate a pro-regenerative, anti-inflammatory, and anti-fibrotic microenvironment [4,5,6]. Crucially, they possess a remarkable immunomodulatory capacity [7,8,9], enabling them to suppress aberrant inflammatory responses by modulating the function of both innate and adaptive immune cells, including T cells, B cells, NK cells, and dendritic cells. This, combined with their low intrinsic immunogenicity, makes allogeneic "off-the-shelf" therapies feasible. This broad therapeutic potential has fueled their investigation in hundreds of clinical trials for a vast spectrum of debilitating conditions, including cardiovascular diseases, stroke, osteoarthritis, graft-versus-host disease (GvHD), and a range of autoimmune disorders [10,11]. Furthermore, their accessibility and multipotency make them invaluable tools in fundamental biomedical research, serving as in vitro models for disease mechanisms, drug screening, and understanding developmental processes [12,13].
Despite decades of promising preclinical data and fervent research, the widespread clinical translation of MSC therapies has been hampered by a stark discrepancy between their expected and actual efficacy [14,15]. The majority of clinical trials, while demonstrating a favorable safety profile, have yielded inconsistent and often modest therapeutic benefits, falling short of the transformative outcomes observed in animal models. This translational gap is not a singular issue but rather the result of several fundamental biological hurdles that collectively undermine the therapeutic potency of administered MSCs. The therapeutic efficacy of MSCs, and their utility as consistent research models, are not solely determined by their inherent multipotency or secretome, but are profoundly influenced by their metabolic state. Cellular metabolism, far from being a mere energy source, acts as a fundamental regulator of cell fate, function, and therapeutic potency [16,17]. The balance between key metabolic pathways, such as glycolysis and oxidative phosphorylation, dictates MSC proliferation rates, their capacity to differentiate along specific lineages, their survival under stress, and, therefore, their therapeutic potential [18,19,20,21]. Suboptimal metabolic conditioning in conventional in vitro culture can lead to reduced viability, impaired differentiation, and diminished paracrine activity, ultimately limiting their clinical success and hindering the reproducibility and relevance of research findings. This problem is exacerbated by the inherent biological variability of MSCs. Far from being a homogenous product, MSCs represent a heterogeneous population of cells whose functional properties are highly dependent on donor-specific attributes (e.g., age, sex, health status) [23,24,25,26,27] and the tissue of origin (e.g., bone marrow, adipose tissue, umbilical cord) [28,29,30]. This functional heterogeneity introduces significant batch-to-batch variability, making the therapeutic potency of any given MSC preparation unpredictable and clinical outcomes difficult to reproduce. Furthermore, a central paradox exists in the current manufacturing paradigm: to obtain the large cell numbers required for clinical dosing (often 1-5 million cells per kilogram of body weight), extensive ex vivo expansion is necessary. However, this prolonged culture period degrades the quality and potency of the final cell product.
To overcome these formidable challenges and finally unlock the full potential of MSCs, a paradigm shift is required—a move away from the passive administration of minimally manipulated cells toward the active, rational engineering of MSCs to create “designer” MSCs, more robust, functional, or specialized for a particular purpose, when they are later introduced into a complex in vivo environment or used in specific research assays. Culture-priming strategies are attractive as a promising tool to augment MSC functionality, and the most fundamental and effective leverage point for this engineering effort is cellular metabolism, a central regulator of cell fate, survival, and functional properties. The ability of an MSC to survive ischemic insults and to perform its therapeutic functions is inextricably linked to its metabolic plasticity to meet environmental demands. Within the context of synthetic biology, metabolic engineering involves the intentional design and manipulation of cellular metabolic pathways to achieve desired biological outcomes. This goes beyond simple observation, embracing a proactive approach to "program" cells for enhanced therapeutic performance or for predictable cellular models for research.
This feature paper will highlight how various synthetic biology approaches, particularly engineered culture environments such as 3D microenvironments and hypoxic preconditioning, alongside other targeted interventions like nutritional and genetic modifications, can precisely reprogram MSC metabolism. By detailing these strategies, we aim to underscore their potential to unlock and optimize the full therapeutic potential of MSCs for advanced regenerative medicine applications and to enhance their utility as robust models in biomedical research.
2. The Metabolic Landscape of MSCs: A Target for Engineering
The metabolic plasticity of MSCs is the engine that drives their therapeutic potential, allowing them to adapt to diverse microenvironments and execute specific functions. Understanding this metabolic landscape is therefore critical to engineering more potent and reliable cell therapies. The Figure 1 visually explains the fundamental metabolic "switches" that dictate MSC function.
2.1. The Native Niche's Metabolic Cues
Many native stem cell niches are characterized by relatively low oxygen tension [31,32]. In their native niche MSCs exist in a largely quiescent state [33,34]. To maintain the state of low proliferation while preserving their multipotent potential, MSCs primarily rely on glycolysis for energy [35,36]. This metabolic profile is a crucial adaptation to their low-oxygen environment, minimizing mitochondrial oxidative phosphorylation (OxPhos) and, therefore, the production of reactive oxygen species (ROS) [16] that are byproducts of mitochondrial respiration and can induce premature differentiation or senescence. The other benefit affecting cellular lifespan is the switch of mitochondria to cataplerotic reactions which helps to balance the various biochemical processes within the cell by regulating the flux of metabolites through the tricarboxylic acid (TCA) cycle, prevents the buildup of potentially harmful compounds, and ensures biosynthetic activity. The reliance on glycolysis also supports the cellular antioxidant pool through the pentose phosphate pathway (PPP). In other words, this glycolytic phenotype allows MSCs to maintain long-term self-renewal capacity.
2.2. Metabolic Changes Under Standard 2D Culture Expansion of MSCs
The conventional 2D culture expansion, which is critical for generating sufficient cell numbers for research and therapeutic applications, alters the bioenergetic landscape of the cells and can ultimately impact their function and quality.
Freshly isolated hMSCs are characterized by a highly glycolytic metabolism [37], a state that is known as the "Warburg-like" effect, where glucose is converted to lactate even in the presence of oxygen. Upon transfer to the standard 2D culture environment, which is characterized by a high oxygen tension (21% O2), a nutrient-rich medium with pH levels in a range of 7.2-7.4, a static, rigid, two-dimensional surface of the culture dish, which fails to replicate the complex 3D architecture, mechanical cues, and cellular interactions found in the native niche, the cells' metabolic strategy undergoes a profound change. The expansion under culture conditions induces a metabolic shift toward a state dominated by OxPhos, resulting in metabolic heterogeneity of MSCs [38]. This transition, driven by the artificial culture environment, is not a simple switch but rather a complex reprogramming of energy metabolism, involving an increased coupling between the glycolytic and TCA cycles. This metabolic and phenotypic heterogeneity becomes more pronounced with each passage, meaning that the expanded cell population is no longer uniform. Instead, it becomes a mixture of clonogenic, glycolytic cells and more mature, OXPHOS-based cells [38,39]. This heterogeneity, which exists even at the clonal level, reflects the accumulated replicative stress and the influence of the non-physiological culture environment.
These metabolic changes have significant consequences for MSC biology and can lead to a loss of their functional capacities. The increased reliance on mitochondrial respiration in a high-oxygen environment leads to a surge in ROS production, which can overwhelm the cell's antioxidant defenses. This oxidative stress, coupled with dysfunctional mitochondria and reduced autophagic/mitophagic activity contributes to a breakdown of cellular homeostasis, an acceleration of cellular senescence, a notable decrease in proliferative capacity and therapeutic efficacy [40,41].
2.3. Metabolic "Switches" Dictate Function
Upon activation, MSCs undergo profound metabolic reprogramming, shifting their energy production pathways to fuel specific functions. These metabolic "switches" are not random but are tightly regulated responses to environmental cues.
2.3.1. Glycolytic Shift for Immunomodulation.
The potent immunomodulatory capacity of MSCs is metabolically demanding and is directly linked to a switch toward aerobic glycolysis [42,43]. When MSCs encounter inflammatory signals such as interferon-gamma (IFN-γ), they ramp up their glycolytic rate via PI3-kinase/AKT pathway and its downstream effector mTORC1, increasing the production of indoleamine 2,3-dioxygenase (IDO) and prostaglandin E2 (PGE2), key immune-regulatory factors [43]. mTORC1 increased both the levels of HIF-1α protein and its transcriptional activity [37] resulting in activation of glycolytic enzymes [44]. The role of peroxisome proliferator-activated receptor (PPAR) family members in regulation of glucose metabolism and immunosuppressive properties of MSCs was demonstrated by Contreras-Lopez et al. [45] revealed that PPARβ/δ deficiency increases MSC glycolytic activity and notably enhanced their immunosuppressive capacities. Enhancement of MSC immunomodulatory abilities under the action of interferon (IFN) was also shown to be associated with glycolysis activation. Thus, upregulation of glycolytic enzymes (GLUT5, hexokinase (HK) II) caused by priming MSCs with the IFN-γ was described by Wobma et al. [46], which is consistent with the findings of mitochondrial activity decrease in MSCs after IFN-γ exposure [43].
2.3.2. Metabolism and Differentiation.
The commitment of an MSC to a specific lineage is accompanied by a distinct metabolic signature. Differentiation into both osteogenic [18,32,47,48] and adipogenic [19,48] directions require a significant increase in mitochondrial OxPhos to meet the high energetic and biosynthetic demands of these processes. In contrast, MSCs differentiated towards the chondrogenic lineage was shown to possess the reduced oxygen consumption indicating a shift towards glycolysis [32]. Chondrogenesis is particularly sensitive to the metabolic environment; MSCs differentiated under hypoxic conditions have been detected to enhance chondrogenesis while decreasing in endochondral ossification compared to cell pellets under normoxia, demonstrating how metabolic programming can precisely direct cell fate [49].
2.3.3. Metabolic Adaptation to Stress.
When transplanted into sites of injury, MSCs face a hostile microenvironment characterized by hypoxia and nutrient deprivation. Their survival hinges on their ability to rapidly adapt their metabolism. In these ischemic conditions, MSCs switch to anaerobic glycolysis to generate ATP. This survival response is critically dependent on the availability of glucose and is orchestrated by the master regulator of oxygen sensing, hypoxia-inducible factor 1-alpha (HIF-1α), which activates the transcription of genes necessary for cellular adaptation [50]. It was shown [51] that the altered metabolism in MSC-based aggregates with reduced mitochondrial potential significantly enhanced cell resistance to in vitro ischemic stress compared to adherent MSCs. These findings demonstrate that metabolic reconfiguration can be considered as a versatile tool to upregulate pro-survival pathways and increase stress resistance.
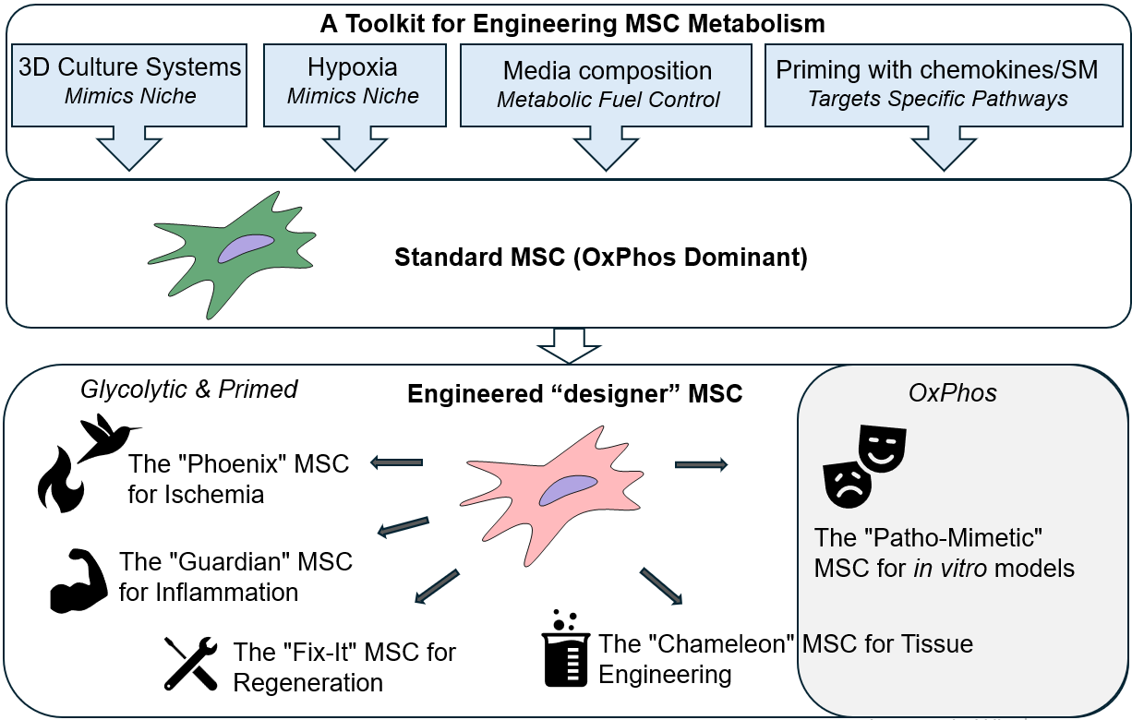
3. Engineering MSC Metabolism Through Culture Parameter Modulation
Synthetic biology's core principles are based on applying engineering concepts to biological systems, creating novel functions or optimizing existing ones [52]. While genetic engineering offers powerful tools for direct manipulation of cellular processes, its application in MSC reprogramming, particularly for clinical translation, often raises safety concerns related to potential immunogenicity, off-target effects, and long-term genomic stability [53]. Therefore, exploring alternative, non-genetic strategies for metabolic modulation becomes crucial. The metabolic state and subsequent therapeutic potential of MSCs can be profoundly influenced by modulating their ex vivo culture parameters. This strategy, often referred to as "priming" or "preconditioning," offers a powerful, non-genetic approach to engineer MSCs to be more resilient and functionally potent upon transplantation. By carefully controlling the culture environment, it's possible to guide MSCs into specific metabolic states that enhance their survival, immunomodulatory capacity, and differentiation potential.
3.1. Advanced Culture Systems: Mimicking the 3D Niche Physiological Complexity
The native stem cell niche is a sophisticated architecture composed of supportive cells and extracellular matrix (ECM) components, featuring a specific 3D topography, controlled mechanical stiffness, and gradients of oxygen and growth factors [31,54]. Traditional 2D cultures lack these crucial cell-cell and cell-matrix interactions, provide conditions only for adhesions restricted to 2D plane, resulting in overspreading and forced apical-basal polarity [55,56], which can lead to a loss of stemness during passaging and contribute to the functional heterogeneity and inconsistent therapeutic outcomes seen in clinical trials. To address these limitations, advanced 3D culture systems have been developed to more accurately recapitulate the in vivo niche, thereby enhancing the therapeutic potential of MSCs [56,57,58]. These systems generally fall into two categories: scaffold-free and scaffold-based methods.
Scaffold-free methods induce MSCs to self-aggregate into multicellular spheroids. This 3D arrangement promotes extensive cell-cell and cell-ECM interactions [59], creating in vivo resembling microenvironment and spheroid zonation that regulate cellular function. As spheroids form, natural gradients of oxygen and nutrients form, often resulting in a hypoxic core that mimics the low-oxygen conditions of the native niche [60]. This mild hypoxia was shown [61] to upregulate HIF-1α and to trigger pro-survival signaling pathways and enhance the secretion of immunomodulatory and pro-angiogenic factors, effectively "priming" the cells for transplantation into injured tissue. The significant consequence of spheroidal formation is a metabolic reconfiguration. Studies have shown that MSCs in spheroids decrease their overall metabolic activity while simultaneously increasing their survival ability [51,62,63]. Moreover, Liu et al. in their impressive report [64] demonstrated that 3D culture of MSCs within spheroids changes mitochondrial morphology, decreases mitochondrial membrane potential, increases glycolytic and anaplerotic flux, which is crucial for reacquiring a more stem-like phenotype. Numerous studies [56,57,58,59,61,64,65,66,67,68,69,70] have shown that MSCs cultured as spheroids exhibit enhanced stemness, viability, differentiation potential, immunomodulatory, angiogenic, and therapeutic effects compared to their 2D counterparts . These findings highlight spheroid culture as an effective approach to creating a native-like microenvironment and inducing metabolic reprogramming in MSCs, thereby potentiating their therapeutic potential.
Scaffold-based systems utilize biomaterials, such as hydrogels or porous solid scaffolds, to provide structural support and present defined biochemical and biophysical cues to the cells [68,71,72,73,74,75,76]. MSCs are able to sense cues from local mechanical microenvironment and transduce them into intracellular biochemical responses, a process known as mechanotransduction [77,78]. Biomaterials can be engineered to impact MSC metabolism, affecting thereby their survival and function. Thus, the significant reduction of basal metabolic activity after MSC encapsulation into alginate hydrogel has been demonstrated in numerous reports [72,76,79,80,81,82]. The higher glycolytic activity and lactate production were shown for MSCs within non-adhesive alginate hydrogel compared to cells in oxidized alginate hydrogels with the GHK peptide that promote adhesion [83]. Culturing of MSCs in alginate/hyaluronic acid hydrogel was supported both with cell amount and metabolic activity increase [84]. The MSCs encapsulated in non-adhesive alginate were found to have reversibly arrested cell cycle [81,85], to be more resistant to oxidative stress [68,76,81], more survivable [68,79,81,82], and to preserve their stemness [75,80]. It was also shown that encapsulated MSCs have enhanced paracrine activity [68,86,87]. The basal metabolic activity of MSCs cultured inside porous 3D scaffolds seems to be dependent on both the scaffold material and the porosity level which determines cell-cell interactions, but generally is higher compared to spheroid or encapsulated cultures. Thus, the metabolic activity of MSCs cultured in 3D porous scaffolds made of poly-((D,L)-lactide-ε-caprolactone)dimethacrylate (LCM) was found to be higher compared to cells in compact LCM scaffolds [71]. The basal metabolic activity of MSCs cultured for three days within plasma-based porous cryogel scaffold was similar to 2D monolayer cultures [72], however, the revealed differences in cell shape (less stretched) and reduced actin component of cytoskeleton can reflect a possible metabolic reprogramming in cells within the scaffolds. The study [88] demonstrated high metabolic activity of MSCs in polyelectrolyte multilayer film scaffold with poly-l-lysine and hyaluronic acid. Gorodetsky et al. [89] showed that during the storage of MSCs within fibrin microbeads at room temperature, pH and gas levels did not change, reflecting a reduced cell metabolism, while viability was better supported compared to cells in polystyrene carriers. The culture in porous scaffolds enhances functional properties of MSCs. It was shown [90], that MSCs in porous scaffold demonstrated the notable increase of gene expression of such growth factors and cytokines as VEGF, HGF, IGF1, FGF2, and IL10. Lian et al. [91] revealed that the paracrine effects of MSCs were stimulated by hierarchical porous structures through mechanotransductive pathways, with the specific signaling molecules involved in this mechanism being focal adhesion kinase (FAK), AKT, and yes-associated protein (YAP).
These findings collectively show that advanced 3D culture systems, such as scaffold-free spheroids and scaffold-based constructs, enhance the therapeutic potential of MSCs. They achieve this by better mimicking the native niche's physical and biochemical cues, which induces metabolic reprogramming and improves cell function compared to traditional 2D cultures.
3.2. Metabolic Modulation via Hypoxic Preconditioning
Standard laboratory culture conditions of atmospheric oxygen (~20% O2), or normoxia, are not relevant to the physiological concentrations ranging from 1% to 15% depending on in vivo niche [92], and can alter MSC metabolism and function. Culturing MSCs under hypoxic conditions (typically 1-5% O2) that mimic their native conditions has been shown to be a better choice for expansion due to the increased proliferative activity and clonogenic properties [31,32,93,94,95,96]. Moreover, MSCs cultured under hypoxic conditions display limited DNA damage [94], and upregulated expression of stemness-related genes as OCT4 and CXCR7 [96]. It was shown [16,94,97,98,99], that metabolic profile in MSCs under hypoxia is characterized by increased glycolysis, decreased OxPhos, and reduced oxidative stress, therefore is closer to the physiological metabolic mode. In addition, this metabolic shift in MSCs cultured under low oxygen tension markedly increases lifespan [94,98,100]. The underlying mechanism for the hypoxia-mediated metabolic reprogramming is stabilization of the transcription factor HIF-1α, which in turn orchestrates a metabolic shift toward glycolysis via upregulation of glycolytic enzymes [50,101]. At the same time HIF-1α decreases OxPhos by inducing pyruvate dehydrogenase kinase 1 (PDK1) which phosphorylates and, in such a way, inhibits pyruvate dehydrogenase (PDH), thereby stopping the formation of acetyl coenzyme A for TCA cycle work [101,102]. Additionally, HIF-1α upregulates the synthesis of lactate dehydrogenase (LDH) [102], which converts pyruvate to lactate and regenerates NAD+ to support glycolysis. This switch prevents excessive ROS production, while HIF-1α-null cells die from ROS toxicity under hypoxia [103]. An emerging benefit of MSC hypoxic preconditioning is their increased survivability due to reduced glucose and oxygen consumption, allowing them to live longer in an environment with limited resource availability. Increased HIF-1α also was shown to protect MSCs against cell death under conditions of oxygen-glucose deprivation via phosphorylation of PI3K, AKT and mTOR [104]. Metabolome analysis of MSCs cultured under hypoxia has identified several amino acids, fumaric acid, lactose, pyrophosphate, cholesterol, as possible mediators for enhanced cell survival [105]. Elevation of levels of essential amino acids can also reflect activation of autophagy, an important pro-survival mechanism in MSCs. HIF-1α overexpression was shown to improve in vivo survival of transplanted MSCs via autophagy induction [104,106]. HIF-1α can also upregulate protective enzymes to mitigate oxidative stress. Heme oxygenase-1 (HO-1) is transcriptionally activated by HIF-1α in response to low oxygen tension [107]. However, Beegle at al. [105] reported decreased expression of HO-1 in hypoxic-preconditioned cells. The expression of superoxide dismutase (SOD), a key antioxidant enzyme, was higher in hypoxic-preconditioned MSCs than in normoxia group [108]. Higher expression of glycolytic enzyme alpha-enolase (Eno1), glucose-6-phosphate 1-dehydrogenase (G6PD), the key enzyme in the PPP, and enzymes involved in oxidative stress resistance (peroxiredoxin-6 (Prdx6) and SOD1) in hypoxic-preconditioned MSCs was shown by Haneef et al. [109]. Interestingly, Li et al. [110] demonstrated that HIF-1α protects cells from oxidative stress-induced apoptosis through non-transcriptional mechanism. This mechanism includes HIF-1α translocation to the mitochondria, which reduces mitochondrial membrane potential, ROS production, and alters the expression of mitochondrial DNA-encoded mRNA.
Hypoxic preconditioning not only increases cell survival but also significantly affects the secretory activity of MSCs, a major contributor to their therapeutic effect. Thus, MSCs exhibited an increased expression of proangiogenic factors (VEGF, ANG, HIF-1α, PDGF) after hypoxic pretreatment [111,112,113]. HIF-1α activation in hypoxic-preconditioned MSCs has been shown to enhance angiogenic effects of the transplanted cells in a murine hindlimb ischemia model [114,115]. The upregulation of NRG-1 and involvement of NRG-1/PI3K/AKT pathway in the angiogenesis enhancement was also detected [115]. In addition, the metabolism manipulation via hypoxic preconditioning provides a promising target to enhance the immunomodulatory potential of MSCs. It promotes the reduction of pro-inflammatory cytokines (IL-6, IL-8) [116], the reduction of tumor necrosis factor-alpha (TNF-α), the increase of anti-inflammatory cytokine IL-10 and immunosuppressive factors (IDO, PGE2, PD-L1, HLA-E, HLA-G) secretion by MSCs [117,118]. Lactate production linked to hypoxia-mediated switch to glycolysis causes T-cell inhibitory action [118]. Hypoxic preconditioning also enhances the production and modify the cargo of MSC-derived exosomes and other extracellular vesicles (EVs), which carry proteins, microRNAs, and lipids that influence recipient cells [119,120,121,122,123,124]. Proteomic analysis of hypoxic preconditioned MSCs and released from them EVs showed upregulation of pathways related to glycolysis, the immune system and extracellular matrix organization [124].
The culture under low oxygen tension also affects the differentiation of MSCs. After hypoxic preconditioning, MSCs retained and even increased their ability to differentiate into chondrocytes [99,125] and adipocytes [99]. In contrast, other studies [126,127] revealed inhibited adipogenesis under hypoxia. Osteogenesis was shown to be promoted by exposure to hypoxia in some works [95,126,127], while others [128,129,130] reported reduced osteogenic differentiation. The specific hypoxic conditions (the level of oxygen tension, exposure duration, cell culture stage (proliferation or differentiation induction)) used in different studies are a major source of conflicting results.
The homing ability of MSCs is critical for their therapeutic success because it allows the transplanted cells to effectively reach and interact with damaged tissue. MSCs exposed to hypoxic conditions show increased homing ability [131,132], which can be mediated by the HGF/c-Met signaling axis [132].
Therefore, hypoxic preconditioning is a powerful and promising tool for metabolic engineering, allowing for the strategic manipulation of MSCs to improve their survival, homing, and functional output. By mimicking the native niche, this approach enhances the quality and consistency of MSCs, moving them one step closer to becoming a reliable and effective therapeutic option in regenerative medicine.
3.3. Nutrient and Media Composition
The composition of the culture medium can be strategically manipulated to control MSC metabolism.
Standard MSC culture media, such as Minimum Essential Medium alpha-modification (α-MEM) and low-glucose DMEM, typically contain glucose at concentrations of 4–5.6 mM to more closely mimic the physiological conditions of the human body, helping to maintain a balanced and more "in vivo-like" cellular metabolism. Culture in high glucose (11-25 mM) medium profoundly impacts MSCs: disturbs mitochondrial function [133], decreases proliferation rates [134,135], induces their aging [134,135,136], increases apoptosis [134], declines their immunosuppressive capabilities [137]. The underlying mechanisms involve mTOR-PI3K-Akt signaling dysregulation together with the dropping of mitochondrial membrane potential, decrease in ATP production, increased oxidative stress, and damaging of cellular structures [138]. The levels of integral regulators of glycolysis (AMP-activated protein kinase (AMPK) and phosphofructokinase-1 (PFK-1)) were found to be significantly declined under high glucose exposure [137]. High glucose also causes the decline of migration ability and osteogenic differentiation of MSCs mediated by the downregulation of SDF1, CXCR4, RUNX2, OSX, OCN, and COL1A [139]. The use of galactose as a metabolic fuel in culture media composition increases TCA cycle turnover, intracellular ROS generation, and results in aging induction in MSCs [140], and higher cell death rate in H9C2 cell lines [141]. Therefore, culture in media with high glucose, or galactose, create a pathological environment and can be used as model systems in researches addressed to cell behavior under hyperglycemia analysis, mitochondrial dysfunction study, search for novel pharmacological targets in metabolic diseases treatment, etc. Interestingly, the resistance of MSC to the short-term exposure of high glucose was with unaffected secretory activity and proliferation was reported by Weil and colleagues [142]. Beltran et al. [141] demonstrated that short-term exposure with high glucose (10 mM) can be used as a metabolic modulation procedure which increases glycolysis, reduces ROS production, and protects H9C2 cells against simulated ischemia-reperfusion injury.
Managing serum levels is a powerful tool for metabolic reprogramming in MSC culture. Reducing serum, or transitioning to serum-free media (SFM), can push MSCs away from an over-proliferative, metabolically stressed state. Thus, five day culture in SFM results in 50% reduction of MSC basal metabolic activity compared to standard culture medium with fetal calf serum (FCS) [143]. An impairment of mitochondrial metabolism was revealed [144] in MSCs exposed for 72 h to serum deprivation with ROS accumulation and upregulation of SOD as a protective mechanism. Serum deprivation during 24 h was found to causes no impact on MSC morphology, phenotype, differentiation, immunomodulatory capacities, and metabolic activity [145]. In response to serum deprivation MSCs improve their immunomodulation potency by stronger secretion of IDO and PGE2 [146], increase angiogenic factors secretion [147], and enhance exosomes production [148]. It has also been reported [149] that long-term (more than 3 weeks) serum deprivation of MSCs selects a subpopulation of early progenitor cells with longer telomeres and enhanced expression of OCT-4. SFM with high glucose was found to possess the highest fibrochondrogenic potential [150]. Reduced serum levels (5%) have been reported [151] to support osteogenic differentiation more effectively than standard concentration of 10% FCS, demonstrating the potential of media composition management in optimizing MSC differentiation strategies for advanced tissue engineering. The MSC metabolic modulation with shift to OxPhos via supplementation of culture media with 1 mM L-ascorbic acid-2-phosphate was shown [152] to improve cell growth and chondrogenic potential, essential for the articular cartilage replacement constructs manufacturing.
3.4. Cytokine Priming
The cytokine priming is a powerful strategy for metabolic reprogramming of MSCs, making them more effective for therapeutic use. This approach involves briefly exposing MSCs to inflammatory cytokines like IFN-γ and TNF-α before transplantation. The goal is to "educate" the cells to behave in a specific, beneficial way, as usual, to acquire enhanced immunomodulatory and immunosuppressive abilities [153]. It has been reported that the exposure of MSCs to IFN [154] or to the mixture of IFN-γ, TNFα, IL-1β, and IL-17 [155] led to markedly increased glycolytic rate, and boosted immunosuppressive, immunomodulatory and homing capacities in primed MSCs. The work [156] reported that priming of MSCs with the mixture of IFN-γ, TNF-α and IL-1β for 24 h enhances their immunomodulatory capacity and do not impact on viability, differentiation, and phenotype. However, there are some conflicting results in the literature about the type of glucose metabolism involved in the regulation of MSC immunomodulatory potential in response to IFN-γ stimulation. Yao et al. [157] found that IFN-γ treatment reprograms human umbilical cord derived MSCs towards and reduced glycolysis and increased aerobic oxidation, which modulates the immunosuppressive function of the primed MSCs by increasing STAT phosphorylation.
3.5. Pharmacological Priming
Pharmacological priming uses small molecules or drugs to directly target specific metabolic pathways, offering a precise and controllable way to enhance the therapeutic potential of MSCs by modulating their metabolism. Current evidences demonstrate that the switch of MSC to glycolytic metabolism enhanced their therapeutic potential can be achieved by several ways. The direct inhibition of OxPhos with oligomycin, an inhibitor of ATP synthase, was shown to induce an AMPK-dependent glycolytic switch [158] resulting in the reduction of PPARβ/δ expression level [45] and the enhancement of immunosuppressive property in treated MSCs. The glycolytic reprogramming via the induction of HIF1α by MSC treatment with O-cyclic phytosphingosine-1-phosphate, a novel chemically synthesized sphingosine metabolite, was demonstrated by Lee et al. [159]. Rhynchophylline (Rhy), a plant-derived alkaloid, enhances kinetic parameters of crucial glycolysis-associated enzymes (HK, G6PD, phosphofructokinase-1 (PFK1), and LDH), decreases PDH activity, and supports the Warburg Effect (aerobic glycolysis, increased ATP content, maintained mitochondrial membrane potential, but reduced cytochrome c oxidase activity) resulting in enhanced stemness of MSCs after Rhy treatment [160]. The possibility to induce MSC glycolytic reprogramming mediated by lysine demethylase via sodium lactate treatment with stemness enhancement was shown by Sun et al. [161].
Some studies have shown the potential of hypoxia mimetic agents for MSC metabolism modulation. Thus, the preconditioning of MSCs with deferoxamine (DFX) notably upregulated HIF-1α expression in MSCs, reprogramming their metabolic status and increasing their immunomodulatory and antioxidant capacities [162,163]. Low concentrations of desferrioxamine (DFO), a hypoxia-mimetic that inhibits HIF-1α hydroxylation via inhibition of prolyl-hydroxylases, were found [164] to reduce mitochondrial activity and apoptosis in MSCs, and to upregulate the glycolytic genes expression (HK2, PDK1, BCL2-interacting protein 3 (BNIP3), LDH).
4. Future Directions: The Next Generations of "Smart" MSCs
The metabolic engineering of MSCs, guided by synthetic biology principles, is poised to move beyond basic priming and into the realm of designing "smart," context-aware MSCs populations. Current studies reviewed here provide scientific background for future developing the next generation of metabolically engineered MSCs (Figure 2).
Synthetic biology provides a framework to design a "Phoenix" MSCs engineered for superior survival in ischemic tissues. This "smart" cell would be equipped with a synthetic genetic circuit that senses the low oxygen tension and high lactate levels characteristic of an ischemic environment. In response, this circuit would activate a rapid and robust metabolic shift from OxPhos to anaerobic glycolysis, ensuring a steady, albeit less efficient, supply of ATP for cellular function [165]. Simultaneously, the circuit would upregulate a suite of pro-survival genes (e.g., those encoding heat shock proteins and anti-apoptotic factors), enabling the cell to withstand the severe stress. This programmed resilience would allow the "Phoenix" MSC to survive the initial insult, home to the site of injury, and begin its regenerative work, a task that has proven difficult for conventional MSC therapies. A crucial extension of the "Phoenix" concept is to engineer a cell that can withstand the logistical challenges of storage and transport. By introducing a new synthetic circuit, the MSC could be placed in a state of metabolic dormancy in response to specific ambient conditions. This "ambient-stable" MSC would reduce its metabolic activity to a minimal level, effectively putting itself into a hibernation-like state [62,63,79,81,82,89]. This would eliminate the need for cumbersome and expensive cryopreservation and cold-chain logistics, allowing for an "off-the-shelf" MSC product that can be stored at room temperature and be readily available for immediate use in emergency settings.
The "Guardian" MSCs can be designed to sense and actively counter the inflammation. This MSC would switch its metabolism towards the massive production and secretion of powerful immunosuppressive molecules like IDO and PGE2 [43,117,118,146], effectively creating a localized, highly potent "living drug factory" that suppresses the damaging inflammatory cascade.
The "Fix-It" MSCs would be engineered to sense specific chemokines released by damaged tissue, such as SDF-1, to improve their homing abilities, and to release key pro-angiogenic factors for effective "jump-start" of new blood vessels formation to support tissue regeneration.
The "Chameleon" MSC would be a next-generation tool for tissue engineering, where differentiation is not a fixed, one-way process but is instead a highly tunable and inducible program. This MSC would be engineered with a synthetic regulatory system that links its metabolic state to a specific external trigger. For example, the medium composition and culture conditions customization could activate a metabolic program and differentiation pathway for more intensive chondrogenesis [150] or osteogenesis [151].
The "Patho-Mimetic" MSCs can be engineered to function as a highly representative in vitro models for specific diseases. For example, we could engineer a hyperglycemic environment by culture in high glucose to mimic a cell living in a diabetic state, including increased oxidative stress and impaired regenerative capacity. Similarly, we could create an "aged" MSC, making it a perfect model for studying age-related decline and testing senolytic drugs. This approach provides a more physiologically relevant system for research and drug development, allowing for more accurate and predictive screening of new therapies.
5. Conclusions
This feature paper has highlighted that the full potential of MSCs is not an inherent property but is, in large part, determined by their metabolic state. By applying the principles of synthetic biology, we can move beyond passive administration to the intentional engineering of MSCs, creating "smart" cells programmed for superior performance. We have demonstrated that a paradigm shift from traditional 2D culture to advanced 3D culture platforms, hypoxic preconditioning, nutrient composition management, and targeting the metabolism with chemokines or small molecules, offers a powerful, non-genetic avenue for metabolic reprogramming, which enhances the MSCs' survival, immunomodulatory capacity, and regenerative potential.
The development of the next generation of "designer" MSCs represents a compelling future for regenerative medicine. By integrating multi-functional synthetic circuits, these cells can be programmed to sense their environment, home to the site of injury, withstand hostile conditions, and orchestrate a multi-faceted repair response. While significant challenges remain, including the need for robust and scalable manufacturing processes, this approach holds the promise of transforming regenerative medicine from an inconsistent, trial-and-error field into a precise, predictable, and highly effective therapeutic discipline. The metabolic engineering of MSCs, therefore, is not just a scientific curiosity; it is the crucial next step towards unlocking their full potential and finally bringing transformative cellular therapies to the clinic.
Author Contributions
Conceptualization, N.T. and O.P.; writing—original draft preparation, N.T.; writing—review and editing, O.P.; writing—review and editing, O.T.; visualization, O.T., funding acquisition, O.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by National Research Foundation of Ukraine (project № 2021.01/0276).
References
- Friedenstein, A. J.; Chailakhjan, R. K.; Lalykina, K. S. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell and tissue kinetics 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.J.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D. G.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef]
- Molnar, V.; Pavelić, E.; Vrdoljak, K.; Čemerin, M.; Klarić, E.; Matišić, V.; Bjelica, R.; Brlek, P.; Kovačić, I.; Tremolada, C.; Primorac, D. Mesenchymal stem cell mechanisms of action and clinical effects in osteoarthritis: a narrative review. Genes 2022, 13, 949. [Google Scholar] [CrossRef] [PubMed]
- Bagno, L.L.; Salerno, A.G.; Balkan, W.; Hare, J. M. Mechanism of action of mesenchymal stem cells (MSCs): impact of delivery method. Expert Opin Biol Ther 2022, 22, 449–463. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Gou, M.; Da, L.C.; Zhang, W.Q.; Xie, H.Q. Mesenchymal stem cells for chronic wound healing: current status of preclinical and clinical studies. Tissue Eng Part B Rev 2020, 26, 555–570. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal stem cell immunomodulation: mechanisms and therapeutic potential. Trends Pharmacol Sci 2020, 41, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Vieira Paladino, F.; de Moraes Rodrigues, J.; da Silva, A.; Goldberg, A.C. The immunomodulatory potential of Wharton's jelly mesenchymal stem/stromal cells. Stem cells Int 2019, 2019, 3548917. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, Q.; Tam, P.K. H. immunomodulatory mechanisms of mesenchymal stem cells and their potential clinical applications. Int J Mol Sci 2022, 23, 10023. [Google Scholar] [CrossRef]
- Zhou, J.; Shi, Y. Mesenchymal stem/stromal cells (MSCs): origin, immune regulation, and clinical applications. Cell Mol Immunol 2023, 20, 555–557. [Google Scholar] [CrossRef]
- Kim, N.; Cho, S.G. Clinical applications of mesenchymal stem cells. Korean J Intern Med 2013, 28, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Afflerbach, A.K.; Kiri, M.D.; Detinis, T.; Maoz, B.M. mesenchymal stem cells as a promising cell source for integration in novel in vitro models. Biomolecules 2020, 10, 1306. [Google Scholar] [CrossRef]
- Milojević, M.; Rožanc, J.; Vajda, J.; Činč Ćurić, L.; Paradiž, E.; Stožer, A.; Maver, U.; Vihar, B. In vitro disease models of the endocrine pancreas. Biomedicines 2021, 9, 1415. [Google Scholar] [CrossRef]
- Galipeau, J.; Sensébé, L. Mesenchymal stromal cells: clinical challenges and therapeutic opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef]
- Robb, K.P.; Galipeau, J.; Shi, Y.; Schuster, M.; Martin, I.; Viswanathan, S. Failure to launch commercially-approved mesenchymal stromal cell therapies: what's the path forward? Proceedings of the International Society for Cell & Gene Therapy (ISCT) Annual Meeting Roundtable held in May 2023, Palais des Congrès de Paris, Organized by the ISCT MSC Scientific Committee. Cytotherapy 2024, 26, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Folmes, C.D.; Dzeja, P.P.; Nelson, T.J.; Terzic, A. Metabolic plasticity in stem cell homeostasis and differentiation. Cell Stem Cell 2012, 11, 596–606. [Google Scholar] [CrossRef]
- Zhang, H.; Menzies, K.J.; Auwerx, J. The role of mitochondria in stem cell fate and aging. Development 2018, 145, dev143420. [Google Scholar] [CrossRef]
- Shum, L.C.; White, N.S.; Mills, B.N.; Bentley, K.L.; Eliseev, R.A. Energy metabolism in mesenchymal stem cells during osteogenic differentiation. Stem Cells Dev 2016, 25, 114–122. [Google Scholar] [CrossRef]
- Meyer, J.; Salamon, A.; Mispagel, S.; Kamp, G.; Peters, K. Energy metabolic capacities of human adipose-derived mesenchymal stromal cells in vitro and their adaptations in osteogenic and adipogenic differentiation. Exp Cell Res 2018, 370, 632–642. [Google Scholar] [CrossRef]
- Sigmarsdottir, T.B.; McGarrity, S.; Yurkovich, J.T.; Rolfsson, Ó.; Sigurjónsson, Ó.E. Analyzing metabolic states of adipogenic and osteogenic differentiation in human mesenchymal stem cells via genome scale metabolic model reconstruction. Front Cell Dev Biol 2021, 9, 642681. [Google Scholar] [CrossRef] [PubMed]
- Mohammadalipour, A.; Dumbali, S.P.; Wenzel, P.L. Mitochondrial transfer and regulators of mesenchymal stromal cell function and therapeutic efficacy. Front Cell Dev Biol 2020, 8, 603292. [Google Scholar] [CrossRef]
- Liu, R.; Chang, W.; Wei, H.; Zhang, K. Comparison of the biological characteristics of mesenchymal stem cells derived from bone marrow and skin. Stem Cells Int 2016, 2016, 3658798. [Google Scholar] [CrossRef]
- Li, J.; Wu, Z.; Zhao, L.; Liu, Y.; Su, Y.; Gong, X.; Liu, F.; Zhang, L. The heterogeneity of mesenchymal stem cells: an important issue to be addressed in cell therapy. Stem Cell Res Ther 2023, 14, 381. [Google Scholar] [CrossRef]
- Trivedi, A.; Lin, M.; Miyazawa, B.; Nair, A.; Vivona, L.; Fang, X.; Bieback, K.; Schäfer, R.; Spohn, G.; McKenna, D.; Zhuo, H.; Matthay, M.A.; Pati, S. Inter- and intra-donor variability in bone marrow-derived mesenchymal stromal cells: implications for clinical applications. Cytotherapy 2024, 26, 1062–1075. [Google Scholar] [CrossRef] [PubMed]
- Mareschi, K.; Ferrero, I.; Rustichelli, D.; Aschero, S.; Gammaitoni, L.; Aglietta, M.; Madon, E.; Fagioli, F. Expansion of mesenchymal stem cells isolated from pediatric and adult donor bone marrow. J Cell Biochem 2006, 97, 744–754. [Google Scholar] [CrossRef]
- Brady, K.; Dickinson, S.C.; Hollander, A.P. Changes in chondrogenic progenitor populations associated with aging and osteoarthritis. Cartilage 2015, 6, 30S–5S. [Google Scholar] [CrossRef]
- Siegel, G.; Kluba, T.; Hermanutz-Klein, U.; Bieback, K.; Northoff, H.; Schäfer, R. Phenotype, donor age and gender affect function of human bone marrow-derived mesenchymal stromal cells. BMC Med 2013, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Al-Nbaheen, M.; Vishnubalaji, R.; Ali, D.; Bouslimi, A.; Al-Jassir, F.; Megges, M.; Prigione, A.; Adjaye, J.; Kassem, M.; Aldahmash, A. Human stromal (mesenchymal) stem cells from bone marrow, adipose tissue and skin exhibit differences in molecular phenotype and differentiation potential. Stem Cell Rev Rep 2013, 9, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Wu, X.Y.; Tong, J.B.; Yang, X.X.; Zhao, J.L.; Zheng, Q.F.; Zhao, G.B.; Ma, Z.J. Comparative analysis of human mesenchymal stem cells from bone marrow and adipose tissue under xeno-free conditions for cell therapy. Stem Cell Res Ther 2015, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.; Sorjamaa, A.; Kangasniemi, M.; Sutinen, M.; Salo, T.; Liakka, A.; Lehenkari, P.; Tapanainen, J.S.; Vuolteenaho, O.; Chen, J.C.; Lehtonen, S.; Piltonen, T.T. Niche matters: The comparison between bone marrow stem cells and endometrial stem cells and stromal fibroblasts reveal distinct migration and cytokine profiles in response to inflammatory stimulus. PLoS One 2017, 12, e0175986. [Google Scholar] [CrossRef]
- Dalloul, A. Hypoxia and visualization of the stem cell niche. Methods Mol Bio 2013, 1035, 199–205. [Google Scholar] [CrossRef]
- Pattappa, G.; Heywood, H.K.; de Bruijn, J.D.; Lee, D.A. The metabolism of human mesenchymal stem cells during proliferation and differentiation. J Cell Physiol 2011, 226, 2562–2570. [Google Scholar] [CrossRef] [PubMed]
- Shyh-Chang, N.; Daley, G.Q.; Cantley, L.C. Stem cell metabolism in tissue development and aging. Development 2013, 140, 2535–2547. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, T. Metabolic regulation of mesenchymal stem cell in expansion and therapeutic application. Biotechnol Prog 2015, 31, 468–481. [Google Scholar] [CrossRef]
- Ito, K.; Ito, K. Metabolism and the control of cell fate decisions and stem cell renewal. Annu Rev Cell Dev Biol 2016, 32, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.T.; Shih, Y.R.; Kuo, T.K.; Lee, O.K.; Wei, Y.H. Coordinated changes of mitochondrial biogenesis and antioxidant enzymes during osteogenic differentiation of human mesenchymal stem cells. Stem Cells 2008, 26, 960–968. [Google Scholar] [CrossRef]
- Ito, K.; Suda, T. Metabolic requirements for the maintenance of self-renewing stem cells. Nat Rev Mol Cell Biol 2014, 15, 243–256. [Google Scholar] [CrossRef]
- Liu, Y.; Muñoz, N.; Bunnell, B.A.; Logan, T.M.; Ma, T. Density-dependent metabolic heterogeneity in human mesenchymal stem cells. Stem Cells 2015, 33, 3368–3381. [Google Scholar] [CrossRef] [PubMed]
- Bara, J.J.; Richards, R.G.; Alini, M.; Stoddart, M.J. Concise review: Bone marrow-derived mesenchymal stem cells change phenotype following in vitro culture: implications for basic research and the clinic. Stem Cells 2014, 32, 1713–1723. [Google Scholar] [CrossRef]
- Denu, R.A.; Hematti, P. Effects of oxidative stress on mesenchymal stem cell biology. Oxid Med Cell Longev 2016, 2016, 2989076. [Google Scholar] [CrossRef]
- Denu, R.A.; Hematti, P. Optimization of oxidative stress for mesenchymal stromal/stem cell engraftment, function and longevity. Free Radic Biol Med 2021, 167, 193–200. [Google Scholar] [CrossRef]
- Contreras-Lopez, R.; Elizondo-Vega, R.; Paredes, M.J.; Luque-Campos, N.; Torres, M.J.; Tejedor, G.; Vega-Letter, A.M.; Figueroa-Valdés, A.; Pradenas, C.; Oyarce, K.; Jorgensen, C.; Khoury, M.; Garcia-Robles, M.L.A.; Altamirano, C.; Djouad, F.; Luz-Crawford, P. HIF1α-dependent metabolic reprogramming governs mesenchymal stem/stromal cell immunoregulatory functions. FASEB J 2020, 34, 8250–8264. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, X.; Muñoz, N.; Logan, T.M.; Ma, T. Commitment to aerobic glycolysis sustains immunosuppression of human mesenchymal stem cells. Stem Cells Transl Med 2019, 8, 93–106. [Google Scholar] [CrossRef]
- Lord-Dufour, S.; Copland, I.B.; Levros Jr, L.C.; Post, M.; Das, A.; Khosla, C.; Galipeau, J.; Rassart, E.; Annabi, B. Evidence for transcriptional regulation of the glucose-6-phosphate transporter by HIF-1alpha: Targeting G6PT with mumbaistatin analogs in hypoxic mesenchymal stromal cells. Stem Cells 2009, 27, 489–497. [Google Scholar] [CrossRef]
- Contreras-Lopez, R.A.; Elizondo-Vega, R.; Torres, M.J.; Vega-Letter, A.M.; Luque-Campos, N.; Paredes-Martinez, M.J.; Pradenas, C.; Tejedor, G.; Oyarce, K.; Salgado, M.; Jorgensen, C.; Khoury, M.; Kronke, G.; Garcia-Robles, M.A.; Altamirano, C.; Luz-Crawford, P.; Djouad, F. PPARβ/δ-dependent MSC metabolism determines their immunoregulatory properties. Sci Rep 2020, 10, 11423. [Google Scholar] [CrossRef] [PubMed]
- Wobma, H.M.; Tamargo, M.A.; Goeta, S.; Brown, LM.; Duran-Struuck, R.; Vunjak-Novakovic, G. The influence of hypoxia and IFN-γ on the proteome and metabolome of therapeutic mesenchymal stem cells. Biomaterials 2018, 167, 226–234. [Google Scholar] [CrossRef]
- Smith, C.O.; Eliseev, R.A. Energy metabolism during osteogenic differentiation: the role of Akt. Stem Cells Dev 2021, 30, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, C.; Khoury, M. Musculoskeletal progenitor/stromal cell-derived mitochondria modulate cell differentiation and therapeutical function. Front Immunol 2021, 12, 606781. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Chang, C.C.; Shieh, M.J.; Wang, J.P.; Chen, Y.T.; Young, T.H.; Hung, S.C. . Hypoxia enhances chondrogenesis and prevents terminal differentiation through PI3K/Akt/FoxO dependent anti-apoptotic effect. Sci Rep, 2683. [Google Scholar] [CrossRef]
- Deschepper, M.; Oudina, K.; David, B.; Myrtil, V.; Collet, C.; Bensidhoum, M.; Logeart-Avramoglou, D.; Petite, H. Survival and function of mesenchymal stem cells (MSCs) depend on glucose to overcome exposure to long-term, severe and continuous hypoxia. J Cell Mol Med 2011, 15, 1505–1514. [Google Scholar] [CrossRef]
- Tsai, A.C.; Liu, Y.; Yuan, X.; Ma, T. Compaction, fusion, and functional activation of three-dimensional human mesenchymal stem cell aggregate. Tissue Eng Part A 2015, 21, 1705–1719. [Google Scholar] [CrossRef]
- Khalil, A.S.; Collins, J.J. Synthetic biology: applications come of age. Nat Rev Genet 2010, 11, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Volarevic, V.; Markovic, B.S.; Gazdic, M.; Volarevic, A.; Jovicic, N.; Arsenijevic, N.; Armstrong, L.; Djonov, V.; Lako, M.; Stojkovic, M. Ethical and safety issues of stem cell-based therapy. Int J Med Sci 2018, 15, 36–45. [Google Scholar] [CrossRef]
- Kanazawa, S.; Okada, H.; Hojo, H.; Ohba, S.; Iwata, J.; Komura, M.; Hikita, A.; Hoshi, K. Mesenchymal stromal cells in the bone marrow niche consist of multi-populations with distinct transcriptional and epigenetic properties. Sci Rep 2021, 11, 15811. [Google Scholar] [CrossRef]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension: how 3D culture microenvironments alter cellular cues. J Cell Sci 2012, 125, 3015–3024. [Google Scholar] [CrossRef]
- Gao, Q.; Cekuc, M.S.; Ergul, Y.S.; Pius, A.K.; Shinohara, I.; Murayama, M.; Susuki, Y.; Ma, C.; Morita, M.; Chow, S.K.-H.; Goodman, S. B. 3D Culture of MSCs for Clinical Application. Bioengineering 2024, 11, 1199. [Google Scholar] [CrossRef]
- Yen, B.L.; Hsieh, C.C.; Hsu, P.J.; Chang, C.C.; Wang, L.T.; Yen, M.L. Three-Dimensional spheroid culture of human mesenchymal stem cells: offering therapeutic advantages and in vitro glimpses of the in vivo state. Stem Cells Transl Med 2023, 12, 235–244. [Google Scholar] [CrossRef]
- Kouroupis, D.; Correa, D. Increased mesenchymal stem cell functionalization in three-dimensional manufacturing settings for enhanced therapeutic applications. Front Bioeng Biotechnol 2021, 9, 621748. [Google Scholar] [CrossRef]
- Lee, E.J.; Park, S.J.; Kang, S.K.; Kim, G.H.; Kang, H.J.; Lee, S.W.; Jeon, H.B.; Kim, H.S. Spherical bullet formation via E-cadherin promotes therapeutic potency of mesenchymal stem cells derived from human umbilical cord blood for myocardial infarction. Mol Ther 2012, 20, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Pérez, J.; Ballesteros, P.; and Cerdán, S. Microscopic images of intraspheroidal pH by 1H magnetic resonance chemical shift imaging of pH sensitive indicators. Magma 2005, 18, 293–301. [Google Scholar] [CrossRef]
- Bhang, S.H.; Lee, S.; Shin, J.-Y.; Lee, T.-J.; Kim, B.-S. Transplantation of cord blood mesenchymal stem cells as spheroids enhances vascularization. Tissue Eng. Part A 2012, 18, 2138–2147. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Yan, L.; Miao, Z.; Li, E.; Wong, K.H.; Xu, R.H. Spheroidal formation preserves human stem cells for prolonged time under ambient conditions for facile storage and transportation. Biomaterials 2017, 133, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Trufanova, N.; Trufanov, O.; Bozhok, G.; Revenko, O.; Cherkashina, D.; Pakhomov, O.; Petrenko, O. Hypothermic storage of mesenchymal stromal cell-based spheroids at a temperature of 22°C. Probl Cryobiol Cryomed 2024, 34, 186–200. [Google Scholar] [CrossRef]
- Liu, Y.; Muñoz, N.; Tsai, A.C.; Logan, T.M.; Ma, T. Metabolic reconfiguration supports reacquisition of primitive phenotype in human mesenchymal stem cell aggregates. Stem Cells 2017, 35, 398–410. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, P.; Chen, L.; Wang, Y.; Wang, Z.; Zhang, B. The effects of spheroid formation of adipose-derived stem cells in a microgravity bioreactor on stemness properties and therapeutic potential. Biomaterials 2015, 41, 15–25. [Google Scholar] [CrossRef]
- Cheng, N.C.; Chen, S.Y.; Li, J.R.; Young, T.H. Short-term spheroid formation enhances the regenerative capacity of adipose-derived stem cells by promoting stemness, angiogenesis, and chemotaxis. Stem Cells Transl Med 2013, 2, 584–594. [Google Scholar] [CrossRef]
- Huang, G.S.; Dai, L.G.; Yen, B.L.; Hsu, S.H. Spheroid formation of mesenchymal stem cells on chitosan and chitosan-hyaluronan membranes. Biomaterials 2011, 32, 6929–6945. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.H.G.; McDevitt, T.C.; Cabral, J.M.S.; da Silva, C.L.; Ferreira, F.C. Tridimensional configurations of human mesenchymal stem/stromal cells to enhance cell paracrine potential towards wound healing processes. J Biotechnol 2017, 262, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Bhang, S.H.; Cho, S.W.; La, W.G.; Lee, T.J.; Yang, H.S.; Sun, A.Y.; Baek, S.H.; Rhie, J.W.; Kim, B.S. Angiogenesis in ischemic tissue produced by spheroid grafting of human adipose-derived stromal cells. Biomaterials 2011, 32, 2734–2747. [Google Scholar] [CrossRef] [PubMed]
- Ohori-Morita, Y.; Niibe, K.; Limraksasin, P.; Nattasit, P.; Miao, X.; Yamada, M.; Mabuchi, Y.; Matsuzaki, Y.; Egusa, H. Novel mesenchymal stem cell spheroids with enhanced stem cell characteristics and bone regeneration ability. Stem Cells Transl Med 2022, 11, 434–449. [Google Scholar] [CrossRef]
- Spreda, M.; Hauptmann, N.; Lehner, V.; Biehl, C.; Liefeith, K.; Lips, K.S. Porous 3D Scaffolds enhance MSC vitality and reduce osteoclast activity. Molecules 2021, 26, 6258. [Google Scholar] [CrossRef]
- Trufanova, N.; Trufanov, O.; Bozhok, G.; Oberemok, R.; Revenko, O.; Petrenko, O. Bioengineering of mesenchymal-stromal-cell-based 3D constructs with different cell organizations. Eng Proc 2024, 81, 8. [Google Scholar] [CrossRef]
- Xue, J.; Liu, Y. Mesenchymal stromal/stem cell (MSC)-based vector biomaterials for clinical tissue engineering and inflammation research: a narrative mini review. J Inflamm Res 2023, 16, 257–267. [Google Scholar] [CrossRef]
- Xing, D.; Liu, W.; Li, J.J.; Liu, L.; Guo, A.; Wang, B.; Yu, H.; Zhao, Y.; Chen, Y.; You, Z.; Lyu, C.; Li, W.; Liu, A.; Du, Y.; Lin, J. Engineering 3D functional tissue constructs using self-assembling cell-laden microniches. Acta Biomater 2020, 114, 170–182. [Google Scholar] [CrossRef]
- Pangjantuk, A.; Kaokaen, P.; Kunhorm, P.; Chaicharoenaudomrung, N.; Noisa, P. 3D culture of alginate-hyaluronic acid hydrogel supports the stemness of human mesenchymal stem cells. Sci Rep 2024, 14, 4436. [Google Scholar] [CrossRef]
- Malysz-Cymborska, I.; Golubczyk, D.; Walczak, P.; Stanaszek, L.; Janowski, M. Injectable, manganese-labeled alginate hydrogels as a matrix for longitudinal and rapidly retrievable 3D cell culture. Int J Mol Sci 2025, 26, 4574. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, C.S.; Fu, J. Forcing stem cells to behave: a biophysical perspective of the cellular microenvironment. Annu Rev Biophys 2012, 41, 519–542. [Google Scholar] [CrossRef] [PubMed]
- Raman, N.; Imran, S.A.M.; Ahmad Amin Noordin, K.B.; Zaman, W.S.W.K.; Nordin, F. Mechanotransduction in mesenchymal stem cells (MSCs) differentiation: a review. Int J Mol Sci 2022, 23, 4580. [Google Scholar] [CrossRef]
- Chen, B.; Wright, B.; Sahoo, R.; Connon, C.J. A novel alternative to cryopreservation for the short-term storage of stem cells for use in cell therapy using alginate encapsulation. Tissue Eng Part C Methods 2013, 19, 568–576. [Google Scholar] [CrossRef] [PubMed]
- de Souza, J.B.; Rosa, G.D.S.; Rossi, M.C.; Stievani, F.C.; Pfeifer, J.P.H.; Krieck, A.M.T.; Bovolato, A.L.C.; Fonseca-Alves, C.E.; Borrás, V.A.; Alves, A.L.G. In vitro biological performance of alginate hydrogel capsules for stem cell delivery. Front Bioeng Biotechnol 2021, 9, 674581. [Google Scholar] [CrossRef]
- Trufanova, N.; Hubenia, O.; Kot, Y.; Trufanov, O.; Kovalenko, I.; Kot, K.; Petrenko, O. Metabolic mode of alginate-encapsulated human mesenchymal stromal cells as a background for storage at ambient temperature. Biopreserv Biobank 2024, 2024. [Google Scholar] [CrossRef]
- Branco, A.; Tiago, A.L.; Laranjeira, P.; Carreira, M.C.; Milhano, J.C.; Dos Santos, F.; Cabral, J.M.S.; Paiva, A.; da Silva, C.L.; Fernandes-Platzgummer, A. Hypothermic Preservation of adipose-derived mesenchymal stromal cells as a viable solution for the storage and distribution of cell therapy products. Bioengineering (Basel) 2022, 9, 805. [Google Scholar] [CrossRef]
- Klontzas, M.E.; Reakasame, S.; Silva, R.; Morais, J.C.F.; Vernardis, S.; MacFarlane, R.J.; Heliotis, M.; Tsiridis, E.; Panoskaltsis, N.; Boccaccini, A.R.; Mantalaris, A. Oxidized alginate hydrogels with the GHK peptide enhance cord blood mesenchymal stem cell osteogenesis: A paradigm for metabolomics-based evaluation of biomaterial design. Acta Biomater 2019, 88, 224–240. [Google Scholar] [CrossRef]
- Tritz-Schiavi, J.; Charif, N.; Henrionnet, C.; de Isla, N.; Bensoussan, D.; Magdalou, J.; Benkirane-Jessel, N.; Stoltz, J.F.; Huselstein, C. Original approach for cartilage tissue engineering with mesenchymal stem cells. Biomed Mater Eng 2010, 20, 167–174. [Google Scholar] [CrossRef]
- Passanha, F.R.; Gomes, D.B.; Piotrowska, J.; Students of PRO3011; Moroni, L. ; Baker, M.B.; LaPointe, V.L.S. A comparative study of mesenchymal stem cells cultured as cell-only aggregates and in encapsulated hydrogels. J Tissue Eng Regen Med 2022, 16, 14–25. [Google Scholar] [CrossRef] [PubMed]
- McKinney, J.M.; Pucha, K.A.; Doan, T.N.; Wang, L.; Weinstock, L.D.; Tignor, B.T.; Fowle, K.L.; Levit, R.D.; Wood, L.B.; Willett, N.J. Sodium alginate microencapsulation of human mesenchymal stromal cells modulates paracrine signaling response and enhances efficacy for treatment of established osteoarthritis. Acta Biomater 2022, 141, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Lagneau, N.; Tournier, P.; Nativel, F.; Maugars, Y.; Guicheux, J.; Le Visage, C.; Delplace, V. Harnessing cell-material interactions to control stem cell secretion for osteoarthritis treatment. Biomaterials 2023, 296, 122091. [Google Scholar] [CrossRef]
- Liu, X.; Laurent, C.; Du, Q.; Targa, L.; Cauchois, G.; Chen, Y.; Wang, X.; de Isla, N. Mesenchymal stem cell interacted with PLCL braided scaffold coated with poly-l-lysine/hyaluronic acid for ligament tissue engineering. J Biomed Mater Res A 2018, 106, 3042–3052. [Google Scholar] [CrossRef] [PubMed]
- Gorodetsky, R.; Levdansky, L.; Gaberman, E.; Gurevitch, O.; Lubzens, E.; McBride, W.H. Fibrin microbeads loaded with mesenchymal cells support their long-term survival while sealed at room temperature. Tissue Eng Part C Methods 2011, 17, 745–755. [Google Scholar] [CrossRef]
- Czosseck, A.; Chen, M.M.; Nguyen, H.; Meeson, A.; Hsu, C.C.; Chen, C.C.; George, T.A.; Ruan, S.C.; Cheng, Y.Y.; Lin, P.J.; Hsieh, P.C.H.; Lundy, D.J. Porous scaffold for mesenchymal cell encapsulation and exosome-based therapy of ischemic diseases. J Control Release 2022, 352, 879–892. [Google Scholar] [CrossRef]
- Lian, M.; Sun, B.; Han, Y.; Yu, B.; Xin, W.; Xu, R.; Ni, B.; Jiang, W.; Hao, Y.; Zhang, X.; Shen, Y.; Qiao, Z.; Dai, K. A low-temperature-printed hierarchical porous sponge-like scaffold that promotes cell-material interaction and modulates paracrine activity of MSCs for vascularized bone regeneration. Biomaterials 2021, 274, 120841. [Google Scholar] [CrossRef]
- Samal, J.R.K.; Rangasami, V.K.; Samanta, S.; Varghese, O.P.; Oommen, O.P. Discrepancies on the role of oxygen gradient and culture condition on mesenchymal stem cell fate. Adv Healthc Mater 2021, 10, e2002058. [Google Scholar] [CrossRef]
- Dos Santos, F.; Andrade, P.Z.; Boura, J.S.; Abecasis, M.M.; da Silva, C.L.; Cabral, J.M. Ex vivo expansion of human mesenchymal stem cells: a more effective cell proliferation kinetics and metabolism under hypoxia. J Cell Physiol 2010, 223, 27–35. [Google Scholar] [CrossRef]
- Estrada, J.C.; Albo, C.; Benguría, A.; Dopazo, A.; López-Romero, P.; Carrera-Quintanar, L.; Roche, E.; Clemente, E.P.; Enríquez, J.A.; Bernad, A.; Samper, E. Culture of human mesenchymal stem cells at low oxygen tension improves growth and genetic stability by activating glycolysis. Cell Death Differ 2012, 19, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Boyette, L.B.; Creasey, O.A.; Guzik, L.; Lozito, T.; Tuan, R.S. Human bone marrow-derived mesenchymal stem cells display enhanced clonogenicity but impaired differentiation with hypoxic preconditioning. Stem Cells Transl Med 2014, 3, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Ko, Y.J.; Lee, M.W.; Park, H.J.; Park, Y.J.; Kim, D.I.; Sung, K. W.; Koo, H.H.; Yoo, K.H. Effect of low oxygen tension on the biological characteristics of human bone marrow mesenchymal stem cells. Cell Stress Chaperones 2016, 21, 1089–1099. [Google Scholar] [CrossRef]
- Muñoz, N.; Kim, J.; Liu, Y.; Logan, T.M.; Ma, T. Gas chromatography-mass spectrometry analysis of human mesenchymal stem cell metabolism during proliferation and osteogenic differentiation under different oxygen tensions. J Biotechnol 2014, 169, 95–102. [Google Scholar] [CrossRef]
- Pattappa, G.; Thorpe, S.D.; Jegard, N.C.; Heywood, H.K.; de Bruijn, J.D.; Lee, D.A. Continuous and uninterrupted oxygen tension influences the colony formation and oxidative metabolism of human mesenchymal stem cells. Tissue Eng Part C Methods 2013, 19, 68–79. [Google Scholar] [CrossRef]
- Mylotte, L.A.; Duffy, A.M.; Murphy, M.; O'Brien, T.; Samali, A.; Barry, F.; Szegezdi, E. Metabolic flexibility permits mesenchymal stem cell survival in an ischemic environment. Stem Cells 2008, 26, 1325–1336. [Google Scholar] [CrossRef]
- Fehrer, C.; Brunauer, R.; Laschober, G.; Unterluggauer, H.; Reitinger, S.; Kloss, F.; Gülly, C.; Gassner, R.; Lepperdinger, G. Reduced oxygen tension attenuates differentiation capacity of human mesenchymal stem cells and prolongs their lifespan. Aging Cell 2007, 6, 745–757. [Google Scholar] [CrossRef]
- Papandreou, I.; Cairns, R.A.; Fontana, L.; Lim, A.L.; Denko, N.C. HIF-1 mediates adaptation to hypoxia by actively downregulating mitochondrial oxygen consumption. Cell Metab 2006, 3, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L.; Roth, P.H.; Fang, H.M.; Wang, G.L. Transcriptional regulation of genes encoding glycolytic enzymes by hypoxia-inducible factor 1. J Biol Chem 1994, 269, 23757–23763. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Tchernyshyov, I.; Semenza, G.L.; Dang, C.V. HIF-1-mediated expression of pyruvate dehydrogenase kinase: a metabolic switch required for cellular adaptation to hypoxia. Cell Metab 2006, 3, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Hua, T.; Li, F.; Han, J.; Fang, J.; Xu, L.; Sun, C.; Zhang, Z.; Feng, Z.; Jiang, X. Hypoxia-inducible factor 1 α protects mesenchymal stem cells against oxygen-glucose deprivation-induced injury via autophagy induction and PI3K/AKT/mTOR signaling pathway. Am J Transl Res 2017, 9, 2492–2499. [Google Scholar]
- Beegle, J.; Lakatos, K.; Kalomoiris, S.; Stewart, H.; Isseroff, R.R.; Nolta, J.A.; Fierro, F.A. Hypoxic preconditioning of mesenchymal stromal cells induces metabolic changes, enhances survival, and promotes cell retention in vivo. Stem Cells 2015, 33, 1818–1828. [Google Scholar] [CrossRef]
- Lv, B.; Li, F.; Han, J.; Fang, J.; Xu, L; Sun, C. ; Hua, T.; Zhang, Z.; Feng, Z.; Jiang, X. Hif-1α overexpression improves transplanted bone mesenchymal stem cells survival in rat MCAO stroke model. Front Mol Neurosci 2017, 10, 80. [Google Scholar] [CrossRef]
- Lee, P.J.; Jiang, B.H.; Chin, B.Y.; Iyer, N.V.; Alam, J.; Semenza, G.L.; Choi, A.M. Hypoxia-inducible factor-1 mediates transcriptional activation of the heme oxygenase-1 gene in response to hypoxia. J Biol Chem 1997, 272, 5375–5381. [Google Scholar] [CrossRef] [PubMed]
- Peterson, K.M.; Aly, A.; Lerman, A.; Lerman, L.O.; Rodriguez-Porcel, M. Improved survival of mesenchymal stromal cell after hypoxia preconditioning: role of oxidative stress. Life Sci 2011, 88, 65–73. [Google Scholar] [CrossRef]
- Haneef, K.; Salim, A.; Hashim, Z.; Ilyas, A.; Syed, B.; Ahmed, A.; Zarina, S. Chemical hypoxic preconditioning improves survival and proliferation of mesenchymal stem cells. Appl Biochem Biotechnol 2024, 196, 3719–3730. [Google Scholar] [CrossRef]
- Li, H.S.; Zhou, Y.N.; Li, L.; Li, S.F.; Long, D.; Chen, X.L.; Zhang, J.B.; Feng, L.; Li, Y.P. HIF-1α protects against oxidative stress by directly targeting mitochondria. Redox Biol 2019, 25, 101109. [Google Scholar] [CrossRef]
- Liu, J.; Hao, H.; Xia, L.; Ti, D.; Huang, H.; Dong, L.; Tong, C.; Hou, Q.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. Hypoxia pretreatment of bone marrow mesenchymal stem cells facilitates angiogenesis by improving the function of endothelial cells in diabetic rats with lower ischemia. PLoS One 2015, 10, e0126715. [Google Scholar] [CrossRef]
- Tong, C.; Hao, H.; Xia, L.; Liu, J.; Ti, D.; Dong, L.; Hou, Q.; Song, H.; Liu, H.; Zhao, Y.; Fu, X.; Han, W. Hypoxia pretreatment of bone marrow-derived mesenchymal stem cells seeded in a collagen-chitosan sponge scaffold promotes skin wound healing in diabetic rats with hindlimb ischemia. Wound Repair Regen 2016, 24, 45–56. [Google Scholar] [CrossRef]
- Hao, D.; He, C.; Ma, B.; Lankford, L.; Reynaga, L.; Farmer, D.L.; Guo, F.; Wang, A. Hypoxic preconditioning enhances survival and proangiogenic capacity of human first trimester chorionic villus-derived mesenchymal stem cells for fetal tissue engineering. Stem Cells Int 2019, 2019, 9695239. [Google Scholar] [CrossRef]
- Yusoff, F.M.; Nakashima, A.; Kawano, K.I.; Kajikawa, M.; Kishimoto, S.; Maruhashi, T.; Ishiuchi, N.; Abdul Wahid, S.F.S.; Higashi, Y. Implantation of hypoxia-induced mesenchymal stem cell advances therapeutic angiogenesis. Stem Cells Int 2022, 2022, 6795274. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Liang, B.; Wang, H.; Hou, J.; Yuan, Q. Hypoxia pretreatment improves the therapeutic potential of bone marrow mesenchymal stem cells in hindlimb ischemia via upregulation of NRG-1. Cell Tissue Res 2022, 388, 105–116. [Google Scholar] [CrossRef]
- Kim, Y.; Jin, H.; Heo, J.; Ju, H.; Lee, H.Y.; Kim, S.; Lee, S.; Lim, J.; Jeong, S.Y.; Kwon, J.; Kim, M.; Choi, S.J.; Oh, W.; Yang, Y.S.; Hwang, H.H.; Yu, H.Y.; Ryu, C.M.; Jeon, H.B.; Shin, D.M. Small hypoxia-primed mesenchymal stem cells attenuate graft-versus-host disease. Leukemia 2018, 32, 2672–2684. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.M.; Liu, J.; Zhao, J.Y.; Xiao, L.; An, S.; Gou, Y.C.; Quan, H.X.; Cheng, Q.; Zhang, Y.L.; He, W.; Wang, Y. T.; Yu, W.J.; Huang, Y.F.; Yi, Y.T.; Chen, Y.; Wang, J. Effects of hypoxia on the immunomodulatory properties of human gingiva-derived mesenchymal stem cells. J Dent Res 2015, 94, 69–77. [Google Scholar] [CrossRef]
- Wobma, H.M.; Kanai, M.; Ma, S.P.; Shih, Y.; Li, H.W.; Duran-Struuck, R.; Winchester, R.; Goeta, S.; Brown, L.M.; Vunjak-Novakovic, G. Dual IFN-γ/hypoxia priming enhances immunosuppression of mesenchymal stromal cells through regulatory proteins and metabolic mechanisms. J Immunol Regen Med 2018, 1, 45–56. [Google Scholar] [CrossRef]
- Bister, N.; Pistono, C.; Huremagic, B.; Jolkkonen, J.; Giugno, R.; Malm, T. Hypoxia and extracellular vesicles: a review on methods, vesicular cargo and functions. J Extracell Vesicles 2020, 10, e12002. [Google Scholar] [CrossRef]
- Almeria, C.; Weiss, R.; Roy, M.; Tripisciano, C.; Kasper, C.; Weber, V.; Egger, D. Hypoxia conditioned mesenchymal stem cell-derived extracellular vesicles induce increased vascular tube formation in vitro. Front Bioeng Biotechnol 2019, 7, 292. [Google Scholar] [CrossRef]
- Ge, L.; Xun, C.; Li, W.; Jin, S.; Liu, Z.; Zhuo, Y.; Duan, D.; Hu, Z.; Chen, P.; Lu, M. Extracellular vesicles derived from hypoxia-preconditioned olfactory mucosa mesenchymal stem cells enhance angiogenesis via miR-612. J Nanobiotechnology 2021, 19, 380. [Google Scholar] [CrossRef] [PubMed]
- Phelps, J.; Hart, D.A.; Mitha, A.P.; Duncan, N.A.; Sen, A. Physiological oxygen conditions enhance the angiogenic properties of extracellular vesicles from human mesenchymal stem cells. Stem Cell Res Ther 2023, 14, 218. [Google Scholar] [CrossRef]
- Pulido-Escribano, V.; Torrecillas-Baena, B.; Camacho-Cardenosa, M.; Dorado, G.; Gálvez-Moreno, M.Á.; Casado-Díaz, A. Role of hypoxia preconditioning in therapeutic potential of mesenchymal stem-cell-derived extracellular vesicles. World J Stem Cells 2022, 14, 453–472. [Google Scholar] [CrossRef] [PubMed]
- Braga, C.L.; da Silva, L.R.; Santos, R.T.; de Carvalho, L.R.P.; Mandacaru, S.C.; de Oliveira Trugilho, M.R.; Rocco, P.R.M.; Cruz, F.F.; Silva, P.L. Proteomics profile of mesenchymal stromal cells and extracellular vesicles in normoxic and hypoxic conditions. Cytotherapy 2022, 24, 1211–1224. [Google Scholar] [CrossRef]
- Shang, J.; Liu, H.; Li, J.; Zhou, Y. Roles of hypoxia during the chondrogenic differentiation of mesenchymal stem cells. Curr Stem Cell Res Ther 2014, 9, 141–147. [Google Scholar] [CrossRef]
- Hung, S.P.; Ho, J.H.; Shih, Y.R.; Lo, T.; Lee, O.K. Hypoxia promotes proliferation and osteogenic differentiation potentials of human mesenchymal stem cells. J Orthop Res 2012, 30, 260–266. [Google Scholar] [CrossRef]
- Lennon, D.P.; Edmison, J.M.; Caplan, A.I. Cultivation of rat marrow-derived mesenchymal stem cells in reduced oxygen tension: effects on in vitro and in vivo osteochondrogenesis. J Cell Physiol 2001, 187, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Holzwarth, C.; Vaegler, M.; Gieseke, F.; Pfister, S.M.; Handgretinger, R.; Kerst, G.; Müller, I. Low physiologic oxygen tensions reduce proliferation and differentiation of human multipotent mesenchymal stromal cells. BMC Cell Biol 2010, 11, 11. [Google Scholar] [CrossRef]
- Salim, A.; Nacamuli, R.P.; Morgan, E.F.; Giaccia, A.J.; Longaker, M.T. Transient changes in oxygen tension inhibit osteogenic differentiation and Runx2 expression in osteoblasts. J Biol Chem 2004, 279, 40007–40016. [Google Scholar] [CrossRef] [PubMed]
- Malladi, P.; Xu, Y.; Chiou, M.; Giaccia, A.J.; Longaker, M.T. Effect of reduced oxygen tension on chondrogenesis and osteogenesis in adipose-derived mesenchymal cells. Am J Physiol Cell Physiol 2006, 290, C1139–C1146. [Google Scholar] [CrossRef]
- Rosová, I.; Dao, M.; Capoccia, B.; Link, D.; Nolta, J.A. Hypoxic preconditioning results in increased motility and improved therapeutic potential of human mesenchymal stem cells. Stem Cells 2008, 26, 2173–2182. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Yuan, H.; Peng, L.; Dai, Z.; Sun, Y.; Liu, R.; Li, W.; Li, J.; Zhu, C. Hypoxia preconditioning of human amniotic mesenchymal stem cells enhances proliferation and migration and promotes their homing via the HGF/C-MET signaling axis to augment the repair of acute liver failure. Tissue Cell 2024, 87, 102326. [Google Scholar] [CrossRef]
- Abu-El-Rub, E.; Almahasneh, F.; Khasawneh, R.R.; Alzu'bi, A.; Ghorab, D.; Almazari, R; Magableh, H. ; Sanajleh, A.; Shlool, H.; Mazari, M.; Bader, N.S.; Al-Momani, J. Human mesenchymal stem cells exhibit altered mitochondrial dynamics and poor survival in high glucose microenvironment. World J Stem Cells 2023, 15, 1093–1103. [Google Scholar] [CrossRef]
- Almahasneh, F.; Abu-El-Rub, E.; Khasawneh, R.R.; Almazari, R. Effects of high glucose and severe hypoxia on the biological behavior of mesenchymal stem cells at various passages. World J Stem Cells 2024, 16, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Cramer, C.; Freisinger, E.; Jones, R.K.; Slakey, D.P.; Dupin, C.L.; Newsome, E.R.; Alt, E.U.; Izadpanah, R. Persistent high glucose concentrations alter the regenerative potential of mesenchymal stem cells. Stem Cells Dev 2010, 19, 1875–1884. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lu, H.; Chen, Z.; Wang, Y.; Lin, J.; Xu, S.; Zhang, C.; Wang, B.; Yuan, Z.; Feng, X.; Jiang, X.; Pan, J. High glucose induces the aging of mesenchymal stem cells via Akt/mTOR signaling. Mol Med Rep 2017, 16, 1685–1690. [Google Scholar] [CrossRef] [PubMed]
- Khasawneh, R.R.; Abu-El-Rub, E.; Almahasneh, F.A.; Alzu'bi, A.; Zegallai, H.M.; Almazari, R.A.; Magableh, H.; Mazari, M.H.; Shlool, H.F.; Sanajleh, A.K. Addressing the impact of high glucose microenvironment on the immunosuppressive characteristics of human mesenchymal stem cells. IUBMB Life 2024, 76, 286–295. [Google Scholar] [CrossRef]
- Mateen, M.A.; Alaagib, N.; Haider, K.H. High glucose microenvironment and human mesenchymal stem cell behavior. World J Stem Cells 2024, 16, 237–244. [Google Scholar] [CrossRef]
- Duangprom, S.; Kheolamai, P.; Tantrawatpan, C.; Manochantr, S. High glucose inhibits proliferation, migration, and osteogenic differentiation of human placenta-derived mesenchymal stem cells. Sci Rep 2025, 15, 22512. [Google Scholar] [CrossRef]
- Zhang, D.; Yan, B.; Yu, S.; Zhang, C.; Wang, B.; Wang, Y.; Wang, J.; Yuan, Z.; Zhang, L.; Pan, J. Coenzyme Q10 inhibits the aging of mesenchymal stem cells induced by D-galactose through Akt/mTOR signaling. Oxid Med Cell Longev 2015, 2015, 867293. [Google Scholar] [CrossRef]
- Beltran, C.; Pardo, R.; Bou-Teen, D.; Ruiz-Meana, M.; Villena, J. A.; Ferreira-González, I.; Barba, I. Enhancing glycolysis protects against ischemia-reperfusion injury by reducing ros production. Metabolites 2020, 10, 132. [Google Scholar] [CrossRef]
- Weil, B.R.; Abarbanell, A.M.; Herrmann, J.L.; Wang, Y.; Meldrum, D.R. High glucose concentration in cell culture medium does not acutely affect human mesenchymal stem cell growth factor production or proliferation. Am J Physiol Regul Integr Comp Physiol 2009, 296, R1735–R1743. [Google Scholar] [CrossRef]
- Popov, A.; Scotchford, C.; Gran,t D. ; Sottile, V. Impact of serum source on human mesenchymal stem cell osteogenic differentiation in culture. Int J Mol Sci 2019, 20, 5051. [Google Scholar] [CrossRef]
- Giannasi, C.; Niada, S.; Della Morte, E.; Casati, S.R.; De Palma, C.; Brini, A.T. Serum starvation affects mitochondrial metabolism of adipose-derived stem/stromal cells. Cytotherapy 2023, 25, 704–711. [Google Scholar] [CrossRef]
- Asti, A.L.; Croce, S.; Valsecchi, C.; Lenta, E.; Grignano, M.A.; Gregorini, M.; Carolei, A.; Comoli, P.; Zecca, M.; Avanzini, M.A.; Rampino, T. Characterization of mesenchymal stromal cells after serum starvation for extracellular vesicle production. Appl Sci 2024, 14, 5821. [Google Scholar] [CrossRef]
- Vu, B.T.; Le, H.T.; Nguyen, K.N.; Van Pham, P. Hypoxia, serum starvation, and TNF-α can modify the immunomodulation potency of human adipose-derived stem cells. Adv Exp Med Biol, 6 November 2021. [Google Scholar] [CrossRef]
- Oskowitz, A.; McFerrin, H.; Gutschow, M.; Carter, M.L.; Pochampally, R. Serum-deprived human multipotent mesenchymal stromal cells (MSCs) are highly angiogenic. Stem Cell Res 2011, 6, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Haraszti, R.A.; Miller, R.; Dubuke, M.L.; Rockwell, H.E.; Coles, A.H.; Sapp, E.; Didiot, M.C.; Echeverria, D.; Stoppato, M.; Sere, Y.Y.; et al. Serum deprivation of mesenchymal stem cells improves exosome activity and alters lipid and protein composition. iScience 2019, 16, 230–241. [Google Scholar] [CrossRef]
- Pochampally, R.R.; Smith, J.R.; Ylostalo, J.; Prockop, D.J. Serum deprivation of human marrow stromal cells (hMSCs) selects for a subpopulation of early progenitor cells with enhanced expression of OCT-4 and other embryonic genes. Blood 2004, 103, 1647–1652. [Google Scholar] [CrossRef]
- Kwan, K.Y.C.; Li, K.; Wang, Y.Y.; Tse, W.Y.; Tong, C.Y.; Zhang, X.; Wang, D.M.; Ker, D.F.E. The Characterization of serum-free media on human mesenchymal stem cell fibrochondrogenesis. Bioengineering 2025, 12, 546. [Google Scholar] [CrossRef] [PubMed]
- Binder, B.Y.; Sagun, J.E.; Leach, J.K. Reduced serum and hypoxic culture conditions enhance the osteogenic potential of human mesenchymal stem cells. Stem Cell Rev Rep 2015, 11, 387–393. [Google Scholar] [CrossRef]
- Tee, C.A.; Roxby, D.N.; Othman, R.; Denslin, V.; Bhat, K.S.; Yang, Z.; Han, J.; Tucker-Kellogg, L.; Boyer, L.A. Metabolic modulation to improve MSC expansion and therapeutic potential for articular cartilage repair. Stem Cell Res Ther 2024, 15, 308. [Google Scholar] [CrossRef]
- Noronha, N.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res Ther 2019, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Vigo, T.; La Rocca, C.; Faicchia, D.; Procaccini, C.; Ruggieri, M.; Salvetti, M.; Centonze, D.; Matarese, G.; Uccelli, A.; MSRUN Network. IFNβ enhances mesenchymal stromal (Stem) cells immunomodulatory function through STAT1-3 activation and mTOR-associated promotion of glucose metabolism. Cell Death Dis 2019, 10, 85. [Google Scholar] [CrossRef]
- Mendt, M.; Daher, M.; Basar, R.; Shanley, M.; Kumar, B.; Wei Inng, F.L.; Acharya, S.; Shaim, H.; Fowlkes, N.; Tran, J.P.; et al. Metabolic reprogramming of GMP grade cord tissue derived mesenchymal stem cells enhances their suppressive potential in GVHD. Front Immunol 2021, 12, 631353. [Google Scholar] [CrossRef] [PubMed]
- Valencia, J.; Yáñez, R.M.; Muntión, S.; Fernández-García, M.; Martín-Rufino, J.D.; Zapata, A.G.; Bueren, J.A.; Vicente, Á.; Sánchez-Guijo, F. Improving the therapeutic profile of MSCs: Cytokine priming reduces donor-dependent heterogeneity and enhances their immunomodulatory capacity. Front Immunol 2025, 16, 1473788. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Chen, Z.; He, X.; Long, J.; Xia, X.; Li, Z.; Yang, Y.; Ao, L.; Xing, W.; Lian, Q.; Liang, H.; Xu, X. Cross talk between glucose metabolism and immunosuppression in IFN-γ-primed mesenchymal stem cells. Life Sci Alliance 2022, 5, e202201493. [Google Scholar] [CrossRef]
- Contreras-Lopez, R.; Elizondo-Vega, R.; Luque-Campos, N.; Torres, M.J.; Pradenas, C.; Tejedor, G.; Paredes-Martínez, M.J.; Vega-Letter, A.M.; Campos-Mora, M.; Rigual-Gonzalez, Y.; et al. The ATP synthase inhibition induces an AMPK-dependent glycolytic switch of mesenchymal stem cells that enhances their immunotherapeutic potential. Theranostics 2021, 11, 445–460. [Google Scholar] [CrossRef]
- Lee, H.J.; Jung, Y.H.; Choi, G.E.; Kim, J.S.; Chae, C.W.; Lim, J.R.; Kim, S.Y.; Lee, J.E.; Park, M.C.; Yoon, J.H.; Choi, M.J.; Kim, K.S.; Han, H.J. O-cyclic phytosphingosine-1-phosphate stimulates HIF1α-dependent glycolytic reprogramming to enhance the therapeutic potential of mesenchymal stem cells. Cell Death Dis 2019, 10, 590. [Google Scholar] [CrossRef]
- Kaneko, Y.; Coats, A.B.; Tuazon, J.P.; Jo, M.; Borlongan, C.V. Rhynchophylline promotes stem cell autonomous metabolic homeostasis. Cytotherapy 2020, 22, 106–113. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, X.; Dai, J.; Pan, Z.; Wu, Y.; Yu, D.; Zhu, S.; Chen, Y.; Qin, T.; Ouyang, H. Sodium lactate promotes stemness of human mesenchymal stem cells through KDM6B mediated glycolytic metabolism. Biochem Biophys Res Commun 2020, 532, 433–439. [Google Scholar] [CrossRef]
- Isildar, B.; Ozkan, S.; Neccar, D.; Koyuturk, M. Preconditioning and post-preconditioning states of mesenchymal stem cells with deferoxamine: a comprehensive analysis. Eur J Pharmacol 2025, 996, 177574. [Google Scholar] [CrossRef]
- Isildar, B.; Ozkan, S.; Sahin, H.; Ercin, M.; Gezginci-Oktayoglu, S.; Koyuturk, M. Preconditioning of human umbilical cord mesenchymal stem cells with deferoxamine potentiates the capacity of the secretome released from the cells and promotes immunomodulation and beta cell regeneration in a rat model of type 1 diabetes. Int Immunopharmacol 2024, 129, 111662. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, K.; Takami, T.; Okada, S.; Hara, K.; Matsumoto, T.; Yamamoto, N.; Yamasaki, T.; Sakaida, I. Analysis of metabolomic changes in mesenchymal stem cells on treatment with desferrioxamine as a hypoxia mimetic compared with hypoxic conditions. Stem Cells 2018, 36, 1226–1236. [Google Scholar] [CrossRef] [PubMed]
- Jaraba-Álvarez, W.V.; Uscanga-Palomeque, A.C.; Sanchez-Giraldo, V.; Madrid, C.; Ortega-Arellano, H.; Halpert, K.; Quintero-Gil, C. Hypoxia-induced metabolic reprogramming in mesenchymal stem cells: unlocking the regenerative potential of secreted factors. Front Cell Dev Biol 2025, 13, 1609082. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Metabolic phenotypes dictate msc fate and function.
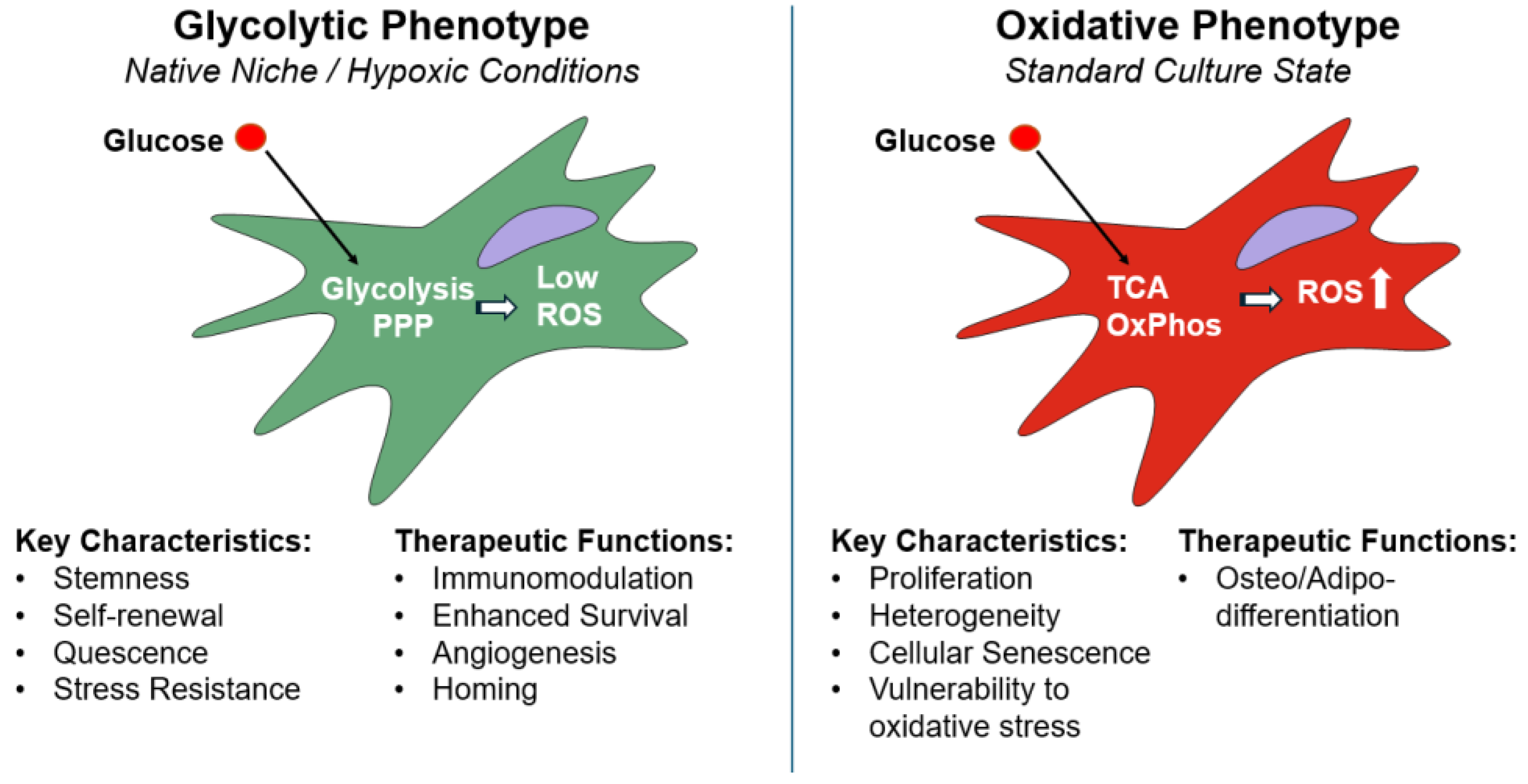
Figure 2.
The future vision: "smart" MSCs programmed to sense and respond. SM-small molecule.
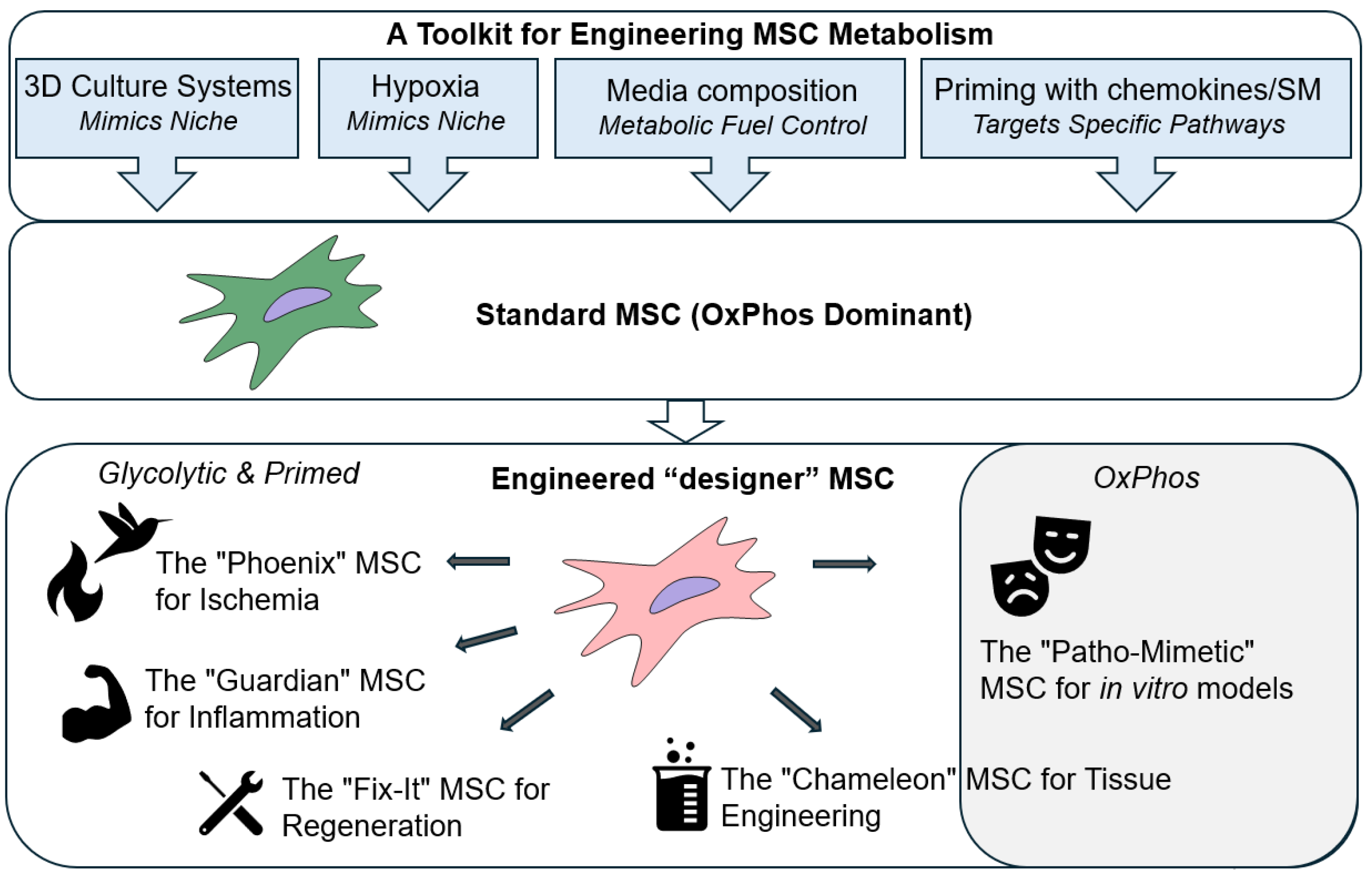
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



