Submitted:
29 August 2025
Posted:
29 August 2025
You are already at the latest version
Abstract
Treatment options for major depression are limited: only about 1/3 of patients achieve remission with first line treatments. More importantly, there are no established predictive markers to identify patients who may be treatment refractory. Research from us and others have established markers related to metabolic disturbances (increased BMI, increased triglyceride levels), inflammatory markers (C-reactive protein, CRP), autonomic disturbances (reduced blood pressure, reduced heart rate variability), brain morphology changes (increased volume of the choroid plexus and brain ventricle volumes). Approximately 1/3 of patients with depression may express these characteristics. These features can mechanistically be linked to a reduced sensitivity of peripheral mineralocorticoid receptors and as a consequence increased release of renin from the kidney, increased production of angiotensin and increased adrenal release of aldosterone. The primary CNS target of aldosterone is a key center of autonomic and affect regulation, i.e. the nucleus of the solitary tract (NTS), which is also the entry point of the vagus nerve, which provides signals from baroreceptors and chemoreceptors. In search of a mechanism to overcome this pathology we identified a molecule, which is derived from the licorice plant glycyrrhiza glabra, namely glycyrrhizin and it biologically active metabolite enoxolone. These molecules have been demonstrated to reverse some of these markers by a well understood dual mechanism: It inhibits the enzyme 11beta hydroxysteroid- dehydrogenase type 2 (11betaHSD2) and inhibits the toll like receptor 4 (TLR4), i.e. the ligand for lipopolysaccharide (LPS) and trigger of innate immunity. As a consequence, patients which show an increase in inflammation markers, an increase in aldosterone or a low blood pressure, are a preferential target population for this compound. Importantly, these patients can be identified as likely treatment refractory BEFORE a treatment is initiated. Clinically, these patients are primarily young females or patients with a history of childhood trauma. A combination of enoxolone with standard antidepressants may therefore avoid a trial and error approach in these subjects and will benefit them by achieving recovery faster.
Keywords:
autonomic nervous system
; sympathetic
; parasympathetic
; vagus
; blood pressure
; baroreceptor
; nucleus of the solitary tract
; area postrema
; aldosterone
; cortisol
; 11beta hydroxysteroid-dehydrogenase type 2
; toll like receptor 4 (TLR4)
; glycyrrhizin
; glycyrrhetinic acid
; enoxolone
; therapy refractory depression
; neuroinflammation
Introduction
STAR-D, a large open label study in patients with depression revealed that first line treatment with the standard antidepressant, sertraline, a selective serotonin reuptake inhibitor (SSRI) leads to remission on only approx. 30% of patients [1]. Apparently, standard antidepressants only work for a small subgroup of patients with depression. Established markers to identify these subjects do not exist, but a number of important demographic, clinical and biomarker characteristics have been reported.
A well characterized clinical differentiation may help with the identification of biological subtypes of more difficult to treat patients. That is the distinction between melancholic and atypical depression. Atypical depression vs. other forms is associated with worse treatment outcome [2,3]. Atypical depression shows hypersomnia, hyperphagia, emotional irritability and somatic complaints [4].
Melancholic depression shows largely opposite features, including weight- and appetite loss, early morning awakening, psychomotor changes and a specific melancholic mood, sometimes referred to as “feeling of feelinglessness”. Differential biological pathways underlie these subtypes, which involve endocrine regulation (HPA-axis and renin-angiotensin-aldosterone system [RAAS]), metabolic status, and autonomic characteristics, including blood pressure and inflammation parameters [4,5,6,7]. Atypical depression has been recognized as a form, which goes along with a reduced noradrenergic activity and normal to reduced cortisol plasma concentration [8,9], whereas an increase in HPA axis activity and noradrenergic activity is related to melancholia.
Despite the fact that melancholic depression is the textbook form of this disorder, it appears to contrast with the foundational neurobiological explanation of the neurobiology of depression, i.e., the idea of a monoamine deficit in depression. This hypothesis was based on the efficacy of norepinephrine reuptake- and monoamine oxidase- inhibitors [8] and the observation that reserpine, a compound which depletes noradrenaline, induces depression [10]. Interestingly, reserpine was utilized as a medication to treat hypertension, i.e., to lower blood pressure, with one important adverse event: fatigue. In accordance some studies reported reduced sympathetic activity was reported in patients with clinical depression [11,12,13], as assessed with electrodermal skin conductance determination. In contrast, increased muscle sympathetic nervous activity (MSNA) was observed in severely depressed patients, who are often melancholic, compared to controls, and MSNA correlated with the severity of depression in these subjects. Interestingly, MSNA could be reduced by the treatment with the SSRI sertraline [14]. Major depression, which is associated with increased sympathetic nervous system activity can therefore go along with arterial hypertension [15] and, as outlined below, may respond better to standard antidepressants.
Several newer studies revealed a number of additional differentiating characteristics between atypical and melancholic depression, namely an increase in inflammation and metabolic characteristics, including an increase in body mass index (BMI) and triglyceride levels in the atypical form [16,17,18]. Interestingly, autonomic markers, including higher heart rate and low cardio-autonomic balance (derived from respiratory sinus arrhythmia and pre-ejection period; higher values are associated with higher parasympathetic activity) where predictive of lower low density lipoprotein (LDL) two years later, indicating that metabolic changes are consequences of autonomic vulnerability factors [19].
On that basis a framework of immune-metabolic depression was developed, which shows an overlap with atypical depression symptoms. Of importance, the context of these biomarkers may be of importance: for example elderly men, who are often melancholic, show an increase in inflammation, which may be related to a different mechanism than in young women [20], who preferentially show signs of atypical depression (see below). We will focus on the relevant biomarkers in the following, starting with a simple biomarker, blood pressure.
Blood Pressure
The fact that patients with depression have rather a low vs. high blood pressure is well established [21,22]. This study of Licht et al., also demonstrated that standard antidepressants tend to increase blood pressure. Similarly, anxiety and depression were associated with low blood pressure in several large studies [23,24] including in elderly subjects [25]. Additional features of low blood pressure are fatigue or hypersomnia: this has parallels to chronic fatigue syndrome (earlier referred to as neurasthenia) [26,27].Vice versa, higher blood pressure is linked with lesser perceived stress and higher quality of life [28,29,30]. On the basis of the observation of an inverse relationship of blood pressure and wellbeing, Dworkin formulated the hypothesis of “learned hypertension” , i.e., blood pressure is upregulated based on the sense of an improved wellbeing [31]. This hypothesis was largely confirmed on the basis of long term data [32].
Blood pressure has only recently been a focus of interest as predictors of depression development, i.e., as a vulnerability marker: Low blood pressure is associated with an increased risk to develop depression [33] and in particular for atypical depression, but appears to be protective against melancholic depression [34]. Low blood pressure is also a risk factor for therapy non-response in depression [35], primarily in women [36]. This is supported by data comparing outpatients with hospitalized patients with depression, the latter plausibly with more refractory forms, which show a reduced blood pressure in comparison to outpatients [37] despite the fact that waist-hip ratio and lipid levels were also higher in this population [37]. In addition, these subjects showed high triglycerides and low HDL. These additional parameters may have contributed to the assumed therapy refractoriness.
Dynamic changes of blood pressure in the course of the day should also be considered. A drop of more than 10 % of systolic blood pressure during sleep defines a so called dipper status, which is associated with lower cardiovascular risk in comparison to non-dippers. Non-dipper status is often observed in patients with hypertension, who also show higher anxiety and depression levels [38], sleep disturbance [39], or shorter sleep duration [40]. However, one study found a more pronounced blood pressure dip [41] in patients with with higher depression severity. Use of sedating medications in the study population was made responsible for that.
Support for a causal relationship between low blood pressure and depressed mood and not only an association is implied by observations that pharmacological interventions with the alpha adrenergic compound midodrine [42], increased salt intake or the MR agonist fludrocortisone improve orthostatic reactions [43] and depressive symptoms [44,45]. Influences of vagal function can be suggested from findings in patients with vasovagal syncope, who did not respond to pharmacological or non-pharmacological treatment [44]. These patients showed higher depression and anxiety ratings; even subsyndromal orthostatic hypotension is associated with cognitive dysfunction, increased hopelessness [46] and increased risk of anxiety and depression [47].
Renin-Angiotensin Aldosterone System:
Aldosterone is a steroid hormone best known for its blood pressure regulating effect. It is regulated via the sympathetic nervous system by beta adrenergic activation of the renin-angiotensin II cascade; in addition in the short term the HPA axis is involved, as ACTH also leads to aldosterone release (see [48]). Potassium can directly release aldosterone from the adrenal cortex. Mineralocorticoids, including aldosterone, have been recognized to affect brain function, focusing on autonomic, not emotional regulation [49]. This is despite the fact the Selye, who brought the “stress” concept into neurobiology, pointed out the importance of mineralocorticoid, like aldosterone as a stress hormone [50]. Selye already identified the proinflammatory effect of mineralocorticoids. Only more recently was aldosterone’s role in emotional regulation recognized [35,48,51,52,53] and its pro-inflammatory role further supported [54].
The role of aldosterone as a mediator to induce depression was established: acute stress and experimental models of depression, including tryptophan depletion and magnesium-depletion, lead to an increase in plasma aldosterone in animal models [55,56,57]. This points to an involvement of glutamatergic and serotonergic mechanisms. Acute stress also leads to an increase in aldosterone in healthy volunteers [58,59].
The fact of an increased level of aldosterone in patients with depression is now well established [60,61,62,63,64]. A causal relationship is implied by the observation that aldosterone leads to depressive and anxiety symptoms, as for example demonstrated in patients with primary hyperaldosteronism, and symptoms improve after its treatment [65,66]. Besides depression and anxiety, somatic complaints are common in these patients. Also, animal data show that subchronic infusion of aldosterone leads to depression and anxiety like behavior [51]. Similarly, polymorphisms of the renin-angiotensin aldosterone system, i.e., of the angiotensin converting enzyme (ACE) and the angiotensin II (ATII) receptor are associated with worse treatment outcome for the more biologically active genotypes [67]. Important in our context, a high ratio of saliva aldosterone/cortisol appears to be related to lesser therapy response [35] in hospitalized patients with depression.
Of note, the increase in aldosterone as observed with an aldosterone releasing adenoma or aldosterone administration is associated with an increase in blood pressure and increased BMI. Increased BMI and increase aldosterone/renin ratio were positively, but independently associated with the severity of depression symptoms [68]. Interestingly, in the latter study an increase in blood pressure was associated with less anxiety in men, which may point to a counterregulatory mechanism of blood pressure on anxiety, possibly via baroreceptor activation (see below). This is in also in line with the above described protective effect of a higher blood pressure against the development of depression.
What is the mechanism of aldosterone to induce depression? Aldosterone has a fairly specific neuronal target, i.e., nucleus of the solitary tract (NTS). The NTS regulates autonomic function, as outlined below, an projects to anatomical targets related to motivation, like the nucleus accumbens; interoception, like the insula; as well as prefrontal cortical areas, like the anterior cingulate [69,70,71]. Specifically, aldosterone acts at neurons, which co-express MR and the enzyme 11-beta-hydroxysteroid dehydrogenase type 2 (11beta-HSD2). This enzyme rapidly degrades cortisol intracellularly and therefore lets aldosterone to compete at the MR, which otherwise would be occupied with the much higher concentrated cortisol. In the absence of 11betaHSD2, as for example in the hippocampus, cortisol/corticosterone is the main ligand at the MR. These neurons of the NTS, which co-express MR and 11beta HSD2, overlap with those of vagus nerve afferents [72] and are involved in salt appetite [73]. This is relevant as salt appetite is increased in depression and anxiety [74,75,76] and can be used as a behavioral marker for central MR activation. The NTS is also activated via the baroreceptor, which indirectly inhibits sympathetic neurons in the rostroventerolateral medulla (RVLM) [77]. At the same time baroreceptor activation increases parasympathetic output. Aldosterone inhibits this mechanism [78], therefore activating the sympathetic and inhibiting the parasympathetic nervous system. Interestingly, baroreceptor activation may induce an anti-inflammatory action [79], which will be covered later. Another nucleus which may be involved in sympathetic regulation of aldosterone is the paraventricular nucleus of the hypothalamus, which, similar to the NTS, coexpresses 11betaHSD2 and MR. Inhibition of 11betaHSD2 leads to an activation of the PVN, which goes along with an increase in HPA axis activity, an increase in blood pressure as well as an increase in renal sympathetic activity [80]. In contrast to the area postrema/NTS complex, this nucleus is protected by the blood brain barrier, which may dampen the effect of aldosterone at this nucleus.
What Leads to an Increase in Aldosterone?
Aldosterone release is activated by low blood pressure (via RAAS activation) and low plasma sodium. The low blood pressure as a trigger needs to be explored. One potential explanation is that lower blood pressure is related to lower peripheral mineralocorticoid receptor (MR) sensitivity: we observed evidence for that in our study [35], as the correlation curve between aldosterone and blood pressure was shifted to the right in non-responders to standard treatment, i.e., a higher aldosterone concentration is required to maintain the same blood pressure. A kidney related mechanism for this appears plausible, as a reduced MR sensitivity at the kidney leads to a reduced sodium reuptake. Alternatively, a vascular mechanism could be involved, similarly to what was proposed in patients with joint hypermobility and reduced vascular tone [81]. The same conclusion of a reduced peripheral MR activity was drawn based on neuroendocrine challenge experiments with a mixed MR-GR agonist [82,83]. In line with this, the adjunct administration of the MR agonist fludrocortisone lead to a faster clinical response to standard antidepressants in comparison to placebo [45]. At the same time, fludrocortisone reduces aldosterone plasma levels. This is most likely a peripheral phenomenon, as fludrocortisone crosses the blood brain barrier only in a very limited way [84].
A consequence of this is the activation of a compensatory mechanism: aldosterone and other MR agonists increase salt intake [85,86] Accordingly, MR antagonists reduce appetite for salt ([87,88]. Patients suffering from depression often suffer from loss of appetite, which could be caused by a subjective tastelessness of food, based on an increased salt taste threshold and increased salt preference [35]. This mechanism can be induced by salt restriction, which leads to an increase in aldosterone and as a consequence an increase in inflammation markers and metabolic markers, including triglycerides, as demonstrated in a large meta analysis [89]. Salt restriction also leads to increased signs of depression and anxiety in animal models [90,91] and in epidemiological studies [92]. In addition and already mentioned above, serotoninergic [56,93,94] and glutamatergic mechanisms [57] play a role.
A link to sleep may exist: there is a close temporal relationship between the activity of RAAS and sleep processes: the concentration of renin and aldosterone increases in synchrony with an increase in SWS [95]. Thus, the total sleep duration as well as the duration of SWS provides a possible correlate for the activity of the MR. On the other hand, conditions with sleep disturbances also show increased nightly aldosterone concentrations. These include major depression (see above) as well as obstructive sleep apnea [96] and prolonged physical strain [97].
Inflammation
Inflammation has long been described as a component of major depression [98]. Higher levels of inflammation are linked to treatment resistance to standard antidepressants [17,99]. In depressed patients, inflammatory changes are found including of CRP and IL-6, slightly less expressed change also takes place with IL-1 [100,101]. The pathophysiology of inflammation in depression is an area of active research.
One important trigger of inflammation is the activation of the toll like receptor 4 (TLR4), which leads to the release of inflammatory cytokines. The TLR4 is the receptor for lipopolysaccharide (LPS, endotoxin). An increased brain expression of TLR4 has been observed in a broad range of neuropsychiatric disturbances, including major depression [102].
Peripheral administration of LPS is an established model for depression in animal models [103,104,105] and induces depressive symptoms in humans [106]. Increased LPS levels are associated with some forms of depression [107], possible based on a “leaky gut” , which may be the consequence of autonomic dysregulation.
TLR4 receptors are regulated by a range of relevant signal pathways, including the beta adrenergic [108], act synergistically with NMDA receptor blockade to reduce white matter integrity [109] and influence the anti-inflammatory and neuroprotective role of glucocorticoids by acting on microglia [110]. Importantly, psychological stress is able to induce TLR4 expression. [111,112,113]. This leads to an increase in the expression of proinflammatory factors interleukin 1beta (IL1b), cyclooxygenase 2 (COX-2) and prostaglandin E2 (PGE2) in the brain cortex and is associated with an increase in depression related behavior as assessed in the forced swim test [113]. An activation of the nuclear factor kappa B (NFkB) inflammatory pathway was involved [114]. Blocking TLR4 receptors pharmacologically prevented an increase in TLR4 expression within the prefrontal cortex of rats [115]. Bacterial translocation from the gut may be involved in this mechanism, as implied by a suppression of this reaction by antibiosis. This action may involve monocytes and brain macrophages [116,117]. Intracerebroventricular administration of LPS leads to an increase of the expression of TLR4 at ventricular ependyma, the choroid plexus and the area postrema [117], which may lead to a feed forward cycle.
A CNS pathway has been described, which regulates peripheral inflammatory activity, i.e., the neuroimmune reflex [118,119]: inflammatory signals from the periphery are transmitted via vagal afferents to the NTS, which then leads to an activation of vagal efferents to the spleen. This vagal activity suppresses inflammatory activity, creating a feedback loop! Therefore, the key anatomical target for aldosterone is involved in the regulation of inflammation. Supporting the physiological relevance, TLR4 is expressed in astrocytes of the area postrema and NTS, i.e., the main targets for aldosterone! The involvement of astrocytes links its mechanism to the glutamatergic system [120], which is the main transmitter of the afferent vagus nerve [121]. Disturbances not only of autonomic regulation, but also interoception can be expected [71], which is reflected in the typical somatoform complaints of patients with increased inflammatory activity.
In our context the anatomical locations of TLR4 are of interest in order to identify these receptors as treatment targets. The TLR4 is expressed in astrocytes of the choroid plexus, circumventricular organs including the area postrema and the nucleus of the solitary tract [122] as well as in the nodose ganglion of the vagus nerve [123]. This makes these TLR4 receptors accessible for systemic treatments without the need to cross the blood brain barrier! Of note, the area postrema is closely connected to the NTS, the primary target for aldosterone. This is a remarkable coincidence for the mode of action of our identified intervention (see below) that the main anatomical area for aldosterone is also affected by TLR4. It shows the synergism between both the RAAS and inflammation on a physiological level. This finding has a major relevance for the treatment approach, which well be described later. A further synergism at a molecular level exists, which will be covered in the next section.
The close link between autonomic nervous system and inflammation was outlined recently with the overall perspective that activation of the SNS is proinflammatory in nature and related to reduced vagal/parasympathetic function) [124,125]. However, it later stages of a disease process, an increase in SNS activity may be beneficial [126].
RAAS and Inflammation
Aldosterone is related to inflammatory and metabolic disturbances [127,128] an correlation was seen to an increase in CRP, insulin, total cholesterol, triglycerides in young obese adults. As mentioned already a physiological interaction between aldosterone and inflammation exists via aldosterone’s action on the NTS. On a molecular level aldosterone acts synergistically at the trigger for the innate immunity, the toll like receptor 4 (TLR4) receptor [129]; furthermore, TLR4 activation by lipopolysaccharide (LPS) induces aldosterone release and inflammation at the adrenal cortex, which can be suppressed by an inhibitor of the RAAS, i.e., an angiotensin II (ATII) receptor antagonist [130]. This may create a feed forward cycle. Indeed, the proinflammatory effect of ATII and its potential role in depression is well established [131].
A direct effect of aldosterone to induce inflammation has been reported: aldosterone acts on neutrophils: the neutrophil/lymphocyte ratio is correlated with aldosterone plasma concentration in patients with primary hyperaldosteronism [132]. Aldosterone induces Th17 cells [133,134] and Th17 cells are increased in the plasma of patients with depression [135].
Role of Obesity and Metabolic Parameters
Obesity has been associated with treatment resistance [136,137], and is related to atypical depression [138,139], which consistently also shows lesser therapy response to standard antidepressants [3]. Obesity in depressed subjects is correlated with depression symptoms [18,140,141]. Obesity related metabolic changes are an increase in triglyceride- and LDL levels. The increase in LDL appears to be somewhat specific to patients with atypical depression [142].
Furthermore, CPR and smoking predict increases in lipid levels two years later in a mixed population from the NESDA study, which includes patients with anxiety and depression [143]. This indicates that inflammation leads to metabolic disturbances. Vice versa, altered lipid metabolism can activate the toll like receptor associated NLRP3 pathway: free unsaturated fatty acids, like palmitate, induce inflammation in in microvascular endothelial cells [144]. This let to a reduction of ZO-1/ZO-2 adhesion factors. RAAS-activation has a link to vascular inflammatory changes and influences inflammatory parameters in plasma [145,146]. High fat diet is associated with brain inflammation which is mediated via activation of brain RAAS [147]. Obesity induces vascular inflammation and is related to white matter dysfunction. This is prevented by the ATII antagonist canrenoate [148]. These associations show an integral mutual interplay between altered lipid metabolism, inflammation and the renin-angiotensin aldosterone system.
The question arises if manipulation of just one of the systems can be beneficial? Regarding obesity and related lipid alterations, reduction of obesity is related to beneficial clinical outcomes, but with limitations. For example, time restricted eating let to a reduced weight and reduced food addiction score in patients with food addiction, which was accompanied by a significant increase of BDNF and a reduction of lipopolysaccharide binding protein (LBP), a co-factor the TLR4-receptor, in the intervention vs. the control group, but stress levels did not change after 8 weeks of treatment [149]. Bariatric surgery improved depression [150], but may lead to higher levels of suicide [151]. Whereas obesity is associated with brain atrophy [152,153], reduction in weight re-balances sympathovagal activity [154] and improves brain morphology [153,155]. With weight loss aldosterone plasma concentration was reduced in subjects, who had a metabolic syndrome at baseline [128]. This study showed that weight loss was associated with a reduction of CRP, insulin, leptin and it reduced sympathetic activity as expressed by a reduced low frequency HRV. Furthermore, a reduction in salt appetite, as determined by urine sodium content, was observed. The change of inflammatory markers correlated with the change in aldosterone levels. However, a large study found an increase in BMI under treatment was associated with a better therapy response, in particular in patients with a low BMI at baseline [156]. Blood pressure was not reported in this study. It is not clear if the latter finding is related to an increase in appetite, as expected with the improvement of melancholic symptoms, but the association of clinical improvement with weight gain shows that increased weight is not universally associated with higher depression severity. As always, the context of the determined biomarkers is important.
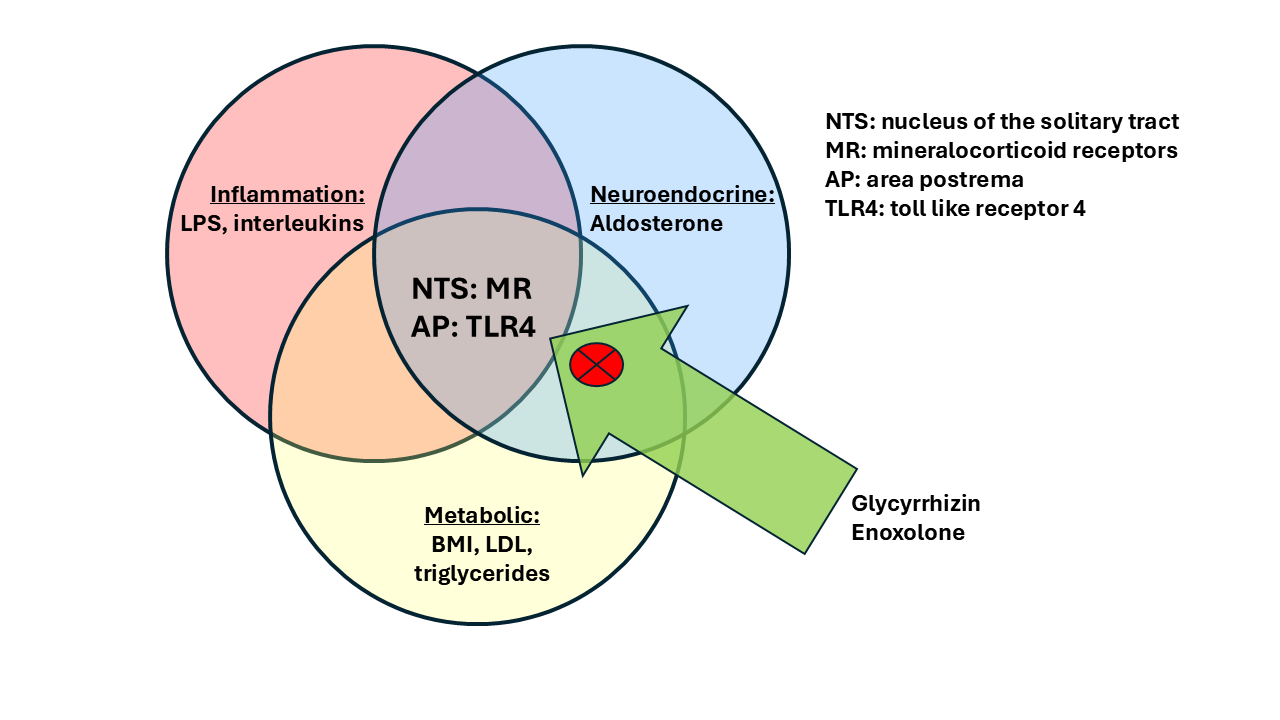
The combination of these characteristics: increased inflammation, obesity and metabolic abnormalities, autonomic dysregulation based on hyperaldosteronism and consequently central MR activation, as well as signs of atypical depression could constitute a specific biological subtype of major importance.
Clinical Relevance
We described a form of depression, which shows signs of metabolic, inflammatory and neuroendocrine alterations, including a lower level of cortisol and potentially higher level of aldosterone. This form in unfortunately characterized as “atypical”, which is true when compared to the textbook form of depression, which often characterized melancholia. Importantly and often overlooked, this form of depression is prevalent in females and in particular young females, which show atypical features in about 80 % [4,157,158]. In addition, these patients show an earlier age of onset in comparison to non-atypical patients, signs of somatization, an increased risk of drug abuse [159]. This is a crucial group of patients, which do frequently not respond to the first line psychopharmacological treatment option of SSRIs. There is evidence that they respond to compounds, which increase noradrenergic and dopaminergic activity, in particular monoamine oxidase inhibitors [160] and interestingly St. John’s wort [161,162], both of which are related to an increase in dopaminergic activity [120,163]. This is of interest as dopaminergic dysfunction appear to be a primary consequence of inflammation, as observed in depressed patients, who show signs if inflammation: Treatment with l-dopa, i.e., and increase in dopaminergic activity, was beneficial in these subjects [164]. Complementary to this mechanism, aldosterone has a direct effect on dopaminergic transmission [48,91,165], which may explain the clinical dopamine deficiency syndrome. This may point to a further understanding of this disorder as a monoamine deficit disorder, besides other characteristics.
How these changes are mediated is not entirely clear. A role of the autonomic nervous system is suggested by the observation that autonomic changes are associated with metabolic alterations: lower HRV in patients with depression is associated with metabolic disturbances [166]. Cardiac physiologic markers, including heart rate, low cardiac autonomic balance and low RSA are associated with abnormal metabolic markers [167] and even predicted the development of metabolic abnormalities, including lower HDL [19]. This may point to autonomic dysfunction as the primary disturbance. In this context, the role of vagal activity is of importance and complex: The sensory (afferent) vagus is involved in sensing satiety [168], which may be interrupted in subjects with hyperphagia. Regarding efferent vagal pathways, high levels of central obesity, as expressed by waist circumference, is consistently related to low HRV as a potential sign of low vagal activity [169]. Low HRV and low baroreflex sensitivity are also associated with MRI assessed higher liver fat content patients with early type II diabetes [170]. Accordingly, weight loss leads to an increase in HRV [154]. Vice versa, recent meta analysis of auricular vagus nerve stimulation shows beneficial effects on glucose tolerance and BMI, but the relevance of the effect size was questioned [171]. However, other studies show that vagus nerve blockade in obese subjects [172] or vagotomy in patients with obesity [173] and in mice [174] improve metabolic function. This makes actually intuitive sense, as the vagus nerve is generally regarded as a rest and digest nerve. Metabolic parameters are in fact regulated by the autonomic nervous system in complex ways [175]. An increase in sympathetic activation is associated with lipolysis and an increase in triglycerides [176]. The role of increased vagal activity in health conditions requires further study [177].
Regarding the question which of these systems should be primarily targeted to achieve a clinical benefit a recent study showed that a reduction of lipids and inflammation, may not be sufficient to improve the underlying pathophysiology: reduction of these markers with a statin as an adjunct treatment to standard antidepressants in comparison to placebo in patients with depression did not have any effect on clinical outcome [178]. These findings imply that the root cause of the disorder is upstream of inflammation and lipid dysregulation. One may consider the role of the autonomic nervous system [177].
Brain Imaging Studies
The characteristics which define a subtype with immune-metabolic changes are an increase in BMI or triglyceride levels and an increase of inflammation markers. In extension this may be associated with a reduced cortisol and increased plasma aldosterone concentration and autonomic dysfunction. Do these markers have consequences for brain morphology? We demonstrated that an increase in BMI and triglycerides are associated with an increase in the volume of the lateral ventricles of the brain [179,180] and that ventricular volumes at baseline were related to treatment outcome. Inflammation has been demonstrated as a potential cause for ventricular volume enlargement, for example in patients with multiple sclerosis [181]. More specifically, the relationship between immune-metabolic forms of depression and increased ventricular volume was confirmed in a large observational study [182]. An increase in ventricular volume was contributed to an increase in the volume and increased activity of the choroid plexus [181]. Moreover, the association between increased choroid plexus volume and inflammatory markers has been confirmed in patients with bipolar disorder [183,184] and major depression [185].
Alterations in gene expression of the choroid plexus have been observed in patients with depression, in particular of markers of transforming growth factor beta (TGFbeta) [186]. As patients with primary hyperaldosteronism have reduced TGFbeta1 plasma levels, which could be reversed with spironolactone [187], a causal link may exist. In our studies an additional feature occurred: We found an association between enlarged ventricles and higher volume of the choroid plexi with a reduction in anterior and medial parts of the corpus callosum [179,180], confirming earlier observations [188]. How these changes are mediated is not immediately clear. We proposed a model, in which the chang of the choroid plexus is a consequence of autonomic dysregulation, which leads to an increase in CSF release and potentially a compression of parts of the corpus callosum [189]. Therefore, balancing the autonomic nervous system may be a priority in order to improve these conditions.
The Role of Childhood Trauma
Childhood trauma has been associated with neuroendocrine characteristics, in particular hypocortisolism, inflammation and autonomic alterations, with a link to orthostatic dysregulation and baroreceptor dysfunction [190]. Furthermore, high aldosterone levels and low blood pressure occurs in subjects with a history of childhood trauma [191]. Traumatization affects brain morphology: Higher volumes of the brain ventricles and smaller volumes of the corpus callosum in comparison to control subjects were observed in traumatized children, who developed PTSD [192,193]. Accordingly, and in line with our studies, white matter integrity of the corpus callosum as measured by fractional anisotropy (FA) was reduced in bipolar subjects with higher levels of childhood trauma and higher levels of inflammation [194]. This study is based on the same sample which showed larger choroid plexus volumes with inflammation [184]. This observation has parallels to morphological changes in patients with depression less responsive to therapy [179,180]. It further supports the association between a history of childhood trauma and risk of antidepressant treatment refractoriness [195,196].
Possible Therapeutic Interventions
In order to understand the plausibility of a targeted pharmacological intervention it is useful to understand the physiology of the intervention for a given clinical outcome. The most relevant and broadly studied one in this context is physical exercise as a treatment of depression [197]. There is preliminary evidence that atypical depression is particularly sensitive to exercise [198]. Furthermore, the combination of exercise to sertraline increases parameters of HRV, which is regarded as an increase in vagal tone [199]. In our context, it is of interest to note that exercise does not only reduce weight, but also reduces ATII, aldosterone and norepinephrine levels in the general population [200]. This has not been studied in patients with depression. As already reported, weigh loss induced by Bariatric surgery is associated with a reduction in aldosterone concentration [201], normalization of brain morphology [202] and improvement of depressive symptoms. This supports a role of aldosterone in the characterized forms of depression.
Regarding pharmacological interventions, the MR agonist fludrocortisone showed some beneficial effect in speeding up recovery in patients with depression [45]. Interestingly, fludrocortisone leads to a suppression of aldosterone release via feedback mechanism. Alternatively, ATII antagonists have shown benefits, primarily in open label studies (see [48]). A limitation of this approach is that ATII antagonists not only reduce aldosterone levels, but also blood pressure, which may counteract its efficacy, as both signals work in opposition at the NTS.
Glycyrrhizin/Enoxolone
Glycyrrhizin Reduces Aldosterone via Inhibition of the 11betaHSD2
An alternative approach to reduce aldosterone plasma concentrations and at the same time increase blood pressure was identified [203]: Glycyrrhizin, from the licorice plant, glycyrrhiza glabra, and its active metabolite glycyrrhetinic acid (enoxolone) acts as an inhibitor of the 11betaHSD2.
Figure 1.
Source of glycyrrhizin and enoxolone: Glycyrrhizin is derived from glycyrrhiza glabra. Glycyrrhetinic acid (Enoxolone) is the active metabolite of glycyrrhizin. It is generated in the gut by hydrolysis by gut bacteria (from [255]).
Figure 1.
Source of glycyrrhizin and enoxolone: Glycyrrhizin is derived from glycyrrhiza glabra. Glycyrrhetinic acid (Enoxolone) is the active metabolite of glycyrrhizin. It is generated in the gut by hydrolysis by gut bacteria (from [255]).
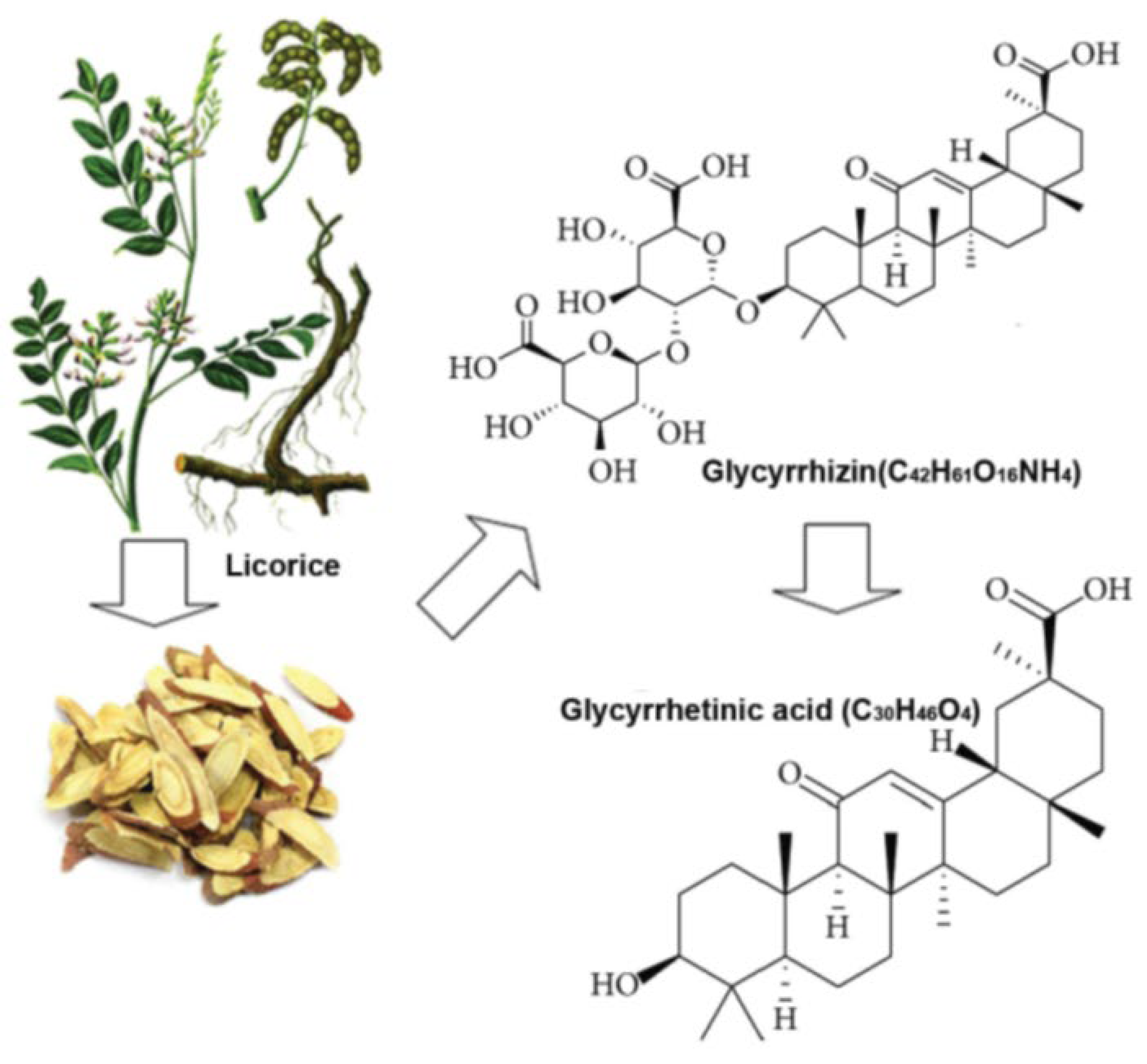
This inhibition mediates a reduction of aldosterone release (see below for details). These compounds are also antagonists at the TLR4. Glycyrrhizin and enoxolone have been widely studies for their pharmacological and biological properties [204]. Their pharmacokinetic and toxicological profile are well known. As glycyrrhizin is converted into enoxolone in the gut and only enoxolone is taken up to act systemically, we regard these compounds as equivalent and don’t differentiate their actions in the following [205]. A related compound is a modified enoxolone, carbenoxolone, which is discussed in the same way.
Enoxolone acts on 11betaHSD2. This is the same enzyme, which protects MR in the NTS and the kidney from being occupied by cortisol/corticosterone, by its intracellular metabolism into the inactive cortisone, therefore, allowing aldosterone to compete. The dose range is critical for its effect: enoxolone acts in a reversible fashion in a dose of 500 mg daily, but may be irreversible in a dose of 1500 mg daily [206]. The main target based side effects can be summarized under pseudohyperaldosteronism, i.e., an increase in blood pressure and hypokalemia [204] based on the stimulation of MR by cortisol. On the basis of these parameters a non effect dose of approx. 100 mg daily has been described [207,208]. As a consequence of the MR stimulatory effect of glycyrrhizin/enoxolone, a feedback mechanism acts to reduce aldosterone release [209,210,211,212]. Due to the MR stimulation an increase in blood pressure can be observed despite the reduction in aldosterone levels. Recently, in a crossover study in healthy subjects a dose as low as 100 mg glycyrrhizin demonstrated to reduce renin and aldosterone and to slightly increase blood pressure [213]. The blood pressure increasing effect of 11betaHSD2 inhibition is in part due to a direct action at vascular MR leading to a vascular contraction. This involves an increased sensitivity to norepinephrine and cortisol [214], as shown in healthy subjects. The vascular effect of glycyrrhizin from licorice (dose 290 -370 mg for two weeks) is reflected by an increase in aortic and popliteal pulse wave velocity, alongside the increased blood pressure and a reduced orthostatic response [215]. In addition, this study showed a reduction in low frequency heart rate variability, potentially as a sign of reduced sympathetic activity.
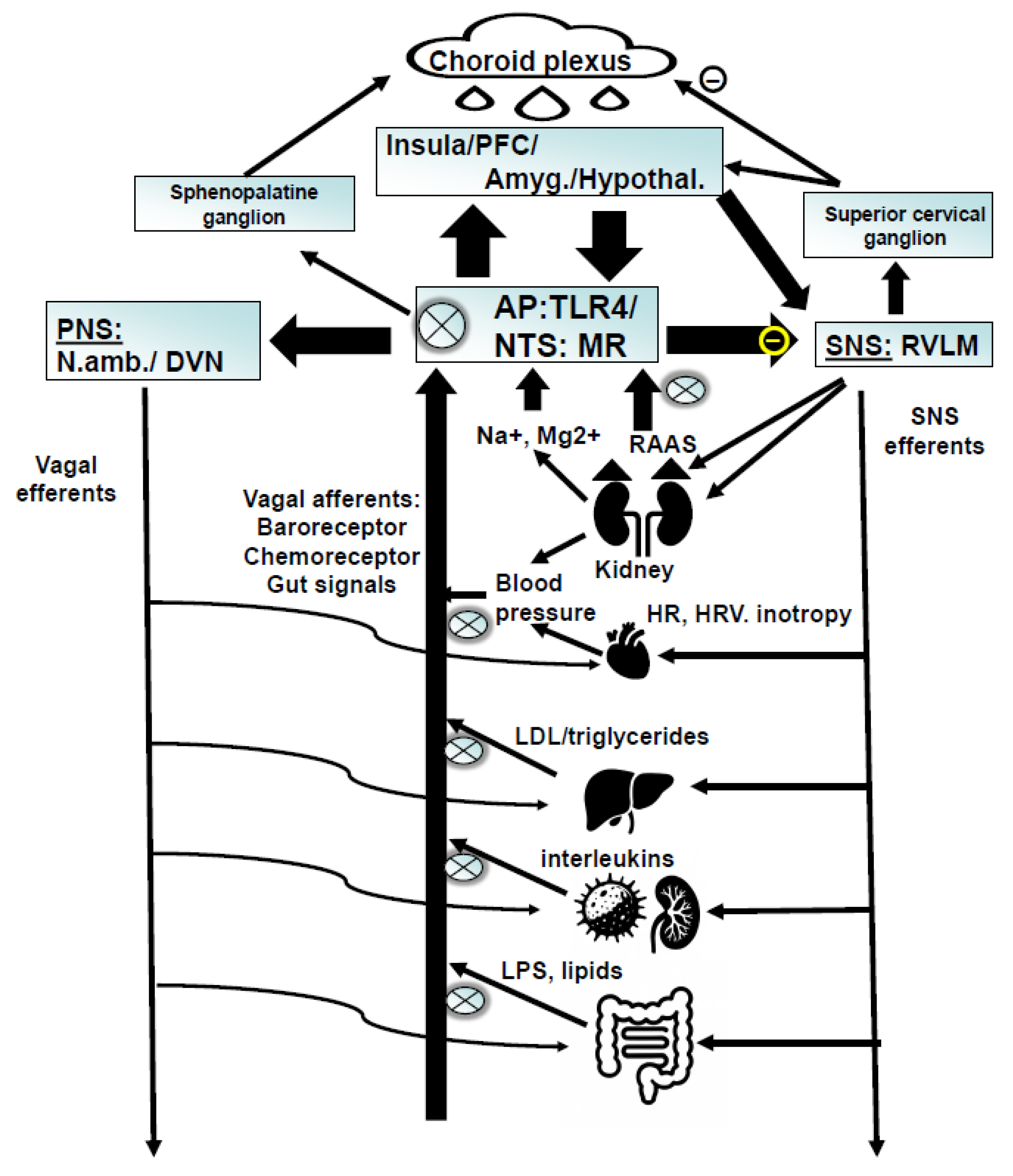
Figure 2.
Schematic characterization proposed to summarize the underlying neurobiology with targets for enoxolone: 1. Enoxolone inhibits the release of aldosterone from the adrenal cortex due the fact that it inhibits the kidney and endothelial 11betaHSD2. This allows cortisol to bind to peripheral MR, which via feedback inhibits renin release, the generation of angiotensin and finally the activation of ATII mediated aldosterone release. The reduction of aldosterone reverses the inhibition of NTS neurons, normalizes baroreceptor reflex and other inputs from the afferent vagus. As a consequence, efferent vagus activity is increased, which has effects on several levels: improvement of cardiovascular regulation; sensitization of the neuro- inflammatory reflex; improvement of metabolic activity. 2. Enoxolone also inhibits TLR4 activity with direct consequences at immune cells; sensitization of the neuro-immune reflex via vagal (nodose ganglion); reduced inflammation of the gut. PFC: prefrontal cortex; Amyg: amygdala; hypothal.: hypothalamus; NTS: nucleus of the solitary tract; AP: area postrema; PNS: parasympathetic nervous system; SNS: sympathetic nervous system; N.amb.: nucleus ambiguous; DRN: dorsal vagal nucleus; RVLM: rostroventrolateral medulla; HR: heart rate; HRV: heart rate variability; RAAS: renin-angiotensin-aldosterone system; LDL: low density lipoprotein. MR: mineralocorticoid receptor; TLR4: toll like receptor 4; LPS: lipopolysaccharide (endotoxin). X : target for enoxolone.
Figure 2.
Schematic characterization proposed to summarize the underlying neurobiology with targets for enoxolone: 1. Enoxolone inhibits the release of aldosterone from the adrenal cortex due the fact that it inhibits the kidney and endothelial 11betaHSD2. This allows cortisol to bind to peripheral MR, which via feedback inhibits renin release, the generation of angiotensin and finally the activation of ATII mediated aldosterone release. The reduction of aldosterone reverses the inhibition of NTS neurons, normalizes baroreceptor reflex and other inputs from the afferent vagus. As a consequence, efferent vagus activity is increased, which has effects on several levels: improvement of cardiovascular regulation; sensitization of the neuro- inflammatory reflex; improvement of metabolic activity. 2. Enoxolone also inhibits TLR4 activity with direct consequences at immune cells; sensitization of the neuro-immune reflex via vagal (nodose ganglion); reduced inflammation of the gut. PFC: prefrontal cortex; Amyg: amygdala; hypothal.: hypothalamus; NTS: nucleus of the solitary tract; AP: area postrema; PNS: parasympathetic nervous system; SNS: sympathetic nervous system; N.amb.: nucleus ambiguous; DRN: dorsal vagal nucleus; RVLM: rostroventrolateral medulla; HR: heart rate; HRV: heart rate variability; RAAS: renin-angiotensin-aldosterone system; LDL: low density lipoprotein. MR: mineralocorticoid receptor; TLR4: toll like receptor 4; LPS: lipopolysaccharide (endotoxin). X : target for enoxolone.
A direct CNS effect may not be relevant for the action of glycyrrhizin/enoxolone in the proposed dose range, but should be briefly reviewed here for completeness: The administration of glycyrrhizin intracerebroventricularly in rats leads to an activation of the hypothalamic paraventricular nucleus, which goes along with an increase in blood pressure and renal sympathetic activity [80] by direct sensitization of the PVN neurons to cortisol/corticosterone. In this context it is important to keep in mind that our attempt is to reduce aldosterone peripherally, therefore, we expect a reduced central MR activation in the projected dose range as a consequence. This targeted approach can be reached as enoxolone crosses the blood brain barrier only to a limited extend [216]. It is of interest to note that there is a potential alternative mechanism for enoxolone and the related compound carbenoxolone to affect hypothalamic activity: carbenoxolone administration into the ventromedial hypothalamus reduced hypothalamic theta oscillations in a sodium-channel dependent way. Mediation by gap junction block was suggested [217]. Involvement of gap junction blockade was also proposed for the effect of carbenoxolone to block vagally mediated hippocampal theta oscillations [218]. However, these effects are unlikely to be relevant in the proposed doses due to the limited BBB permeability.
Glycyrrhizin Reduces Inflammation via TLR4 Inhibition
Glycyrrhizin/enoxolone have been recognized as neuroprotective by inhibiting inflammation via TLR4 antagonism and reduction of high mobility group box-1 (HMGB-1) protein [116,117,219] and action at its receptor RAGE [220,221], for an overview see [102,111,222,223].
Glycyrrhizin reduces the generation of oxidative stress from neutrophils [224]. In addition and complementary to this effect, enoxolone protects against oxidative stress in intestinal epithelial cells by activating the PI3k/Akt pathway [225]. Similarly, advanced glycation endproducts (AGEs) induced oxidative stress and inflammation in umbilical cord epithelial cells, which was inhibited by glycyrrhizin [226]. In a hamster model, an extract from glycyrrhiza glabra had anti-inflammatory effects agains SARS/Cov-2 infection, which reflects direct inhibitory action at neutrophils and reduced oxidative stress [227]. A suppression of the differentiation of Th1, Th2 and Th17 cells was involved in the action of glycyrrhizin in this study. A direct effect of glycyrrhizin in T-cells was confirmed in another experiment [228]. Overall, it appears that one target of glycyrrhizin/enoxolone is immune cells.
An indirect immune suppression can be implied by the action of these compounds in the function of the NTS. The neuroimmune reflex [118,119] establishes a feedback mechanism to suppress increased peripheral inflammation. The key anatomical structures are the vagus afferents, connecting to the vagus nerve and the efferent vagus from the nucleus ambiguous and dorsal vagal nucleus. These efferents to the spleen suppress immune response, as determined by interleukin release, via cholinergic alpha-7 nicotinergic receptors.
This association between the central element of the anti-inflammatory reflex and its association with the area postrema leads to an additional important anatomical location relevant for the anti-inflammatory effect of enoxolone: TLR4 is expressed in astrocytes of the area postrema and NTS, the latter being the main target for aldosterone! This demonstrates an intriguing synergism between the main effects of enoxolone to affect NTS activity on the one hand by targeting astrocytes via TLR4 and neurons via 11betaHSD2.
Behavioral Effects of Glycyrrhizin/Enoxolone
An animal model (single prolonged stress) leads to an induction TLR4 and HMGB1 expression in the basolateral amygdala, which was accompanied by neuroinflammation. These changes could be inhibited by glycyrrhizin [111,112] Glycyrrhizin blocks the effect of subchronic restraint stress: this form of stress induction inhibits locomotor activity, which can be reversed by glycyrrhizin [229]. The behavioral effect was accompanied by an increase in corticosterone release. Interestingly, the beneficial behavioral effect was observable for rats treated for 10 days, but not for several weeks. In another study glycyrrhizin, as active component of an extract, reduced salt appetite and increased HPA axis activity, when administered orally in a dose of 150 mg/kg per day for two weeks. The extract had a concentration of glycyrrhizin of approx. 7% [230]. The reduced salt appetite is a sign of reduced central MR activation, as intended. This was accompanied by an anxiolytic effect and, again, an increase in corticosterone release. The increase in aldosterone was restricted to unstressed, not stressed rats [231]. A non-significant reduction in aldosterone was observed, which overall points to a reduction in the aldosterone/corticosterone ratio.
Glycyrrhizin counteracts depression like behavior in mice administered with LPS, the ligand of the TLR4-receptor [232]. Zhang et al. (2014) and Kamisili [233] pointed to the importance of the inhibition of the NFkB-IL-17-pathway in the action of glycyrrhizin for brain protection. This is relevant, as brain morphological changes, mediated by inflammatory processes, may be mediated by this inflammatory pathway. Action at the choroid plexus structures appear to be involved and may mediate depression [183]. The choroid plexus appears to be an entry point of IL-17 producing Th-17 T-helper cells, which play a key role in demyelination [234]. We are currently researching if enoxolone leads to changes in brain morphology in patients with depression and for which patients this may be related to treatment outcome (https://www.clinicaltrials.gov/study/NCT05570110).
In addition, the same inflammatory pathway is also relevant for the beneficial behavioral effect of glycyrrhizin via improved integrity of the gut [235,236]. In line with this with these observations, a glycyrrhizin containing extract downregulated the expression of angiotensin converting enzyme 2 (ACE2)[231], potentially due to the reduced inflammatory stress [237].
The expected reduction of aldosterone induced by this compound may also induce an anti-inflammatory effect. Glycyrrhizin demonstrated anti-inflammatory properties in an ischemia model via inhibition of TLR4 receptors, which was similar to effects observed with the ATII antagonist candesartan [238]. This point back to the synergism of aldosterone and inflammatory activity via aldosterone’s action to boost TLR4 activity [129].
Effect of Glycyrrhizin/Enoxolone on Metabolic Parameters
Glycyrrhizin demonstrated an effect on metabolic parameters: in an animal model (doxorubine induced cardiomyopathy) triglyceride levels decrease, whereas HDL levels increase [239], in addition to an increase in blood pressure. Four week treatment with an ethanolic extract of licorice let to a similar reduction in triglycerides, plasma glucose and weight gain in high fat diet (for 10 weeks) induced metabolic disturbance in rats. This was accompanied by changes in gut flora, liver steatosis and signs of reduced liver inflammation [240]. Oral administration of glycyrrhetinic acid (enoxolone) leads to an improved lipid profile and a reduced weight gain in a high fat diet model [241]. The effect on lipid metabolism and inflammation may involve the stabilization of ACE2 in the liver [242] in an LPS/D-galactosaminase model. It may also involve a change in sympathetic hepatic nerve activity, which may potentially be mediated via the gap-junction inhibitory effect of these compounds, as described for carbenoxolone [243]. Finally, metabolic changes in obesity involve higher levels of free unsaturated fatty acids, which induce inflammation and a reduction of ZO-1/ZO-1 adhesion factors in microvascular endothelial cells. These changes could be reversed with the administration of glycyrrhizin [144].
Glycyrrhizin Reverses Catecholamine Depletion – Autonomic Activity as Primary Driver?
Back to the starting point focusing on catecholamines and autonomic regulation: Schildkraut’s framework of the catecholamine deficiency hypothesis of depression may best apply to patients with fatigue and/or low blood pressure! The overlap between these symptoms is well documented [199,244]. Ammonium glycyrrhizinate (100 and 150 mg/kg) intraperitoneally, i.p.) has demonstrated a reversal of behavioral changes induced by reserpine [245], potentially mediated by an inhibition of monoamine oxidase. In line with this observation, an antidepressant effect of glycyrrhizin could be inhibited by antagonism at noradrenergic and dopaminergic receptors: reduced immobility was observed in the forced swim at a dose of 150 mg/kg extract containing approx. 7% glycyrrhizin. 75 mg/kg or 300 mg/kg were not effective, supporting an optimal dose range. The effect could be blocked by sulpiride (D2 antagonist) or prazosin (alpha1 antagonist) [246]. In a further experiment, utilizing a norepinephrine depletion model with fusaric acid, a dose dependent reversal of anxiety related behavior was observed with the administration of glycyrrhizin, which was also accompanied by an increase in brain metabolic signs (increase in ATP) and integrity (increase in brain derived neurotrophic factor, BDNF) [247]. Doses of 200 mg/kg and 300 mg/kg orally (dissolved in destilled water), but not 100 mg/kg were effective on these parameters.
Clinical Studies:
How do the reported biological activities of glycyrrhizin translate into clinical outcome? Beneficial effects in animal models of depression and anxiety were reported above. The number of clinical observations is, however, limited. Glycyrrhizin is a common ingredient in Japanese, Chinese and Ayurvedic medicine, for example of the Japanese Yokukansan and showed beneficial effects on depressive symptoms in a placebo-controlled trial [248,249]. It is, however, difficult to attribute this to any specific ingredient. In clinical studies in patients with depression glycyrrhizin in a defined concentration as component of an extract from glycyrrhiza glabra demonstrated antidepressant activity in comparison to a historic control [203]. The effect was most pronounced in a patient group with a low vs. higher systolic blood pressure (median split at 127 mmHg). Clinical improvement in the glycyrrhizin treated subjects was associated with a reduced heart rate and a shortening of sleep duration. The anti-inflammatory effect of glycyrrhizin is another important element of its efficacy: In a randomized placebo controlled trial with adjunct treatment of glycyrrhizin to an SSRI an overall statistically significant benefit of glycyrrhizin vs. placebo was observed, but that effect was mainly restricted to subjects with a higher level of C-reactive protein as a sign of systemic inflammation [250]. Subjects were primarily male (89%) and had on average a normal BMI of approx. 23.4, i.e., did not show metabolic abnormalities. Besides a significant clinical improvement, the actively treated subjects showed a reduction in TNFalpha and IL1beta, which was significantly different from the placebo group. An interims analysis from an ongoing randomized placebo-controlled study with pure enoxolone, 100 mg vs. placebo, in hospitalized patients with depression [251] confirmed target engagement by a significant reduction of the plasma aldosterone/cortisol ratio and of plasma CRP. Treatment responders showed preferentially higher CRP levels at baseline as well as a history of more childhood abuse. Put together, we provided evidence for an antidepressant effect of glycyrrhizin/enoxolone in combination with standard antidepressants in a population, which can be identified on the basis of biological (inflammation, autonomic disturbances, high aldosterone/cortisol levels) and anamnestic (childhood trauma) parameters. We hypothesize that the assessments of these biomarkers can be done BEFORE any standard antidepressant is applied, which may avoid the clinical expression of treatment refractoriness. However, these subjects can be regarded as treatment refractory based on their underlying biology. Therefore, enoxolone/glycyrrhizin could be a first line treatment for subjects with high risk of treatment resistance.
Limitation
The current summary of the effects of glycyrrhizin/enoxolone focus on the overlap with a specific form of depression, which is less responsive to standard antidepressants and which show signs of inflammation, increased aldosterone levels, low cortisol and low blood pressure. The motivation to study these compounds was to find an intervention which reverse these biomarker changes and the underlying neurobiology, therefore improving clinical outcome. We therefore reported aspects of enoxolone, which are supportive of this concept, and are the main characteristics of this compound. Other properties have not been discussed in detail. These include the antiviral effect of these compounds [252], anticancer, liver protective, SARS-Cov2 targeting and also 11betaHSD1 inhibiting properties [253] other properties, for an overview see [204,254].
Conclusion
We identified biomarkers related to treatment resistance to standard antidepressants in depression. These include autonomic (low blood pressure, low heart rate variability), inflammation (higher CRP), metabolic (higher triglycerides and higher BMI) and neuroendocrine (high aldosterone/cortisol ratio) markers. The underlying neurobiology points to a dysfunction of peripheral MR activity, which leads to pontine autonomic dysregulation. Glycyrrhizin/enoxolone has specific properties to overcome the underlying neurobiology. The clinical effect in a specific subgroup is supported by preliminary data. Additional studies are ongoing to further test these findings.
References
- Trivedi, M.H.; Rush, A.J.; Wisniewski, S.R.; Nierenberg, A.A.; Warden, D.; Ritz, L.; Norquist, G.; Howland, R.H.; Lebowitz, B.; McGrath, P.J. , et al. Evaluation of outcomes with citalopram for depression using measurement-based care in STAR*D: implications for clinical practice. Am J Psychiatry 2006, 163, 28–40. [Google Scholar] [CrossRef]
- Fava, M.; Rush, A.J.; Alpert, J.E.; Balasubramani, G.K.; Wisniewski, S.R.; Carmin, C.N.; Biggs, M.M.; Zisook, S.; Leuchter, A.; Howland, R. , et al. Difference in treatment outcome in outpatients with anxious versus nonanxious depression: a STAR*D report. Am J Psychiatry 2008, 165, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.W.; McGrath, P.J.; Fava, M.; Wisniewski, S.R.; Zisook, S.; Cook, I.; Nierenberg, A.A.; Trivedi, M.H.; Balasubramani, G.K.; Warden, D. , et al. Do atypical features affect outcome in depressed outpatients treated with citalopram? Int J Neuropsychopharmacol 2010, 13, 15–30. [Google Scholar] [CrossRef]
- Murck, H. Atypical depression spectrum disorder - neurobiology and treatment. Acta Neuropsychiatrica 2003, 15, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Lamers, F.; Bot, M.; Jansen, R.; Chan, M.K.; Cooper, J.D.; Bahn, S.; Penninx, B.W. Serum proteomic profiles of depressive subtypes. Transl Psychiatry 2016, 6, e851. [Google Scholar] [CrossRef]
- Lamers, F.; Vogelzangs, N.; Merikangas, K.R.; de Jonge, P.; Beekman, A.T.; Penninx, B.W. Evidence for a differential role of HPA-axis function, inflammation and metabolic syndrome in melancholic versus atypical depression. Mol Psychiatry 2013, 18, 692–699. [Google Scholar] [CrossRef]
- Milaneschi, Y.; Lamers, F.; Berk, M.; Penninx, B. Depression Heterogeneity and Its Biological Underpinnings: Toward Immunometabolic Depression. Biol Psychiatry 2020, 88, 369–380. [Google Scholar] [CrossRef]
- Gold, P.W.; Wong, M.L. Re-assessing the catecholamine hypothesis of depression: the case of melancholic depression. Mol Psychiatry 2021, 26, 6121–6124. [Google Scholar] [CrossRef]
- Wong, M.L.; Kling, M.A.; Munson, P.J.; Listwak, S.; Licinio, J.; Prolo, P.; Karp, B.; McCutcheon, I.E.; Geracioti, T.D., Jr.; DeBellis, M.D. , et al. Pronounced and sustained central hypernoradrenergic function in major depression with melancholic features: relation to hypercortisolism and corticotropin-releasing hormone. Proc Natl Acad Sci U S A 2000, 97, 325–330. [Google Scholar] [CrossRef]
- Schildkraut, J.J. The catecholamine hypothesis of affective disorders: a review of supporting evidence. Am J Psychiatry 1965, 122, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Thorell, L.H. Valid electrodermal hyporeactivity for depressive suicidal propensity offers links to cognitive theory. Acta Psychiatr Scand 2009, 119, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Ward, N.G.; Doerr, H.O.; Storrie, M.C. Skin conductance: a potentially sensitive test for depression. Psychiatry Res 1983, 10, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Mestanikova, A.; Ondrejka, I.; Mestanik, M.; Hrtanek, I.; Snircova, E.; Tonhajzerova, I. Electrodermal Activity in Adolescent Depression. Adv Exp Med Biol 2016, 935, 83–88. [Google Scholar] [CrossRef]
- Scalco, A.Z.; Rondon, M.U.; Trombetta, I.C.; Laterza, M.C.; Azul, J.B.; Pullenayegum, E.M.; Scalco, M.Z.; Kuniyoshi, F.H.; Wajngarten, M.; Negrao, C.E. , et al. Muscle sympathetic nervous activity in depressed patients before and after treatment with sertraline. J Hypertens 2009, 27, 2429–2436. [Google Scholar] [CrossRef]
- Scalco, A.Z.; Scalco, M.Z.; Azul, J.B.; Lotufo Neto, F. Hypertension and depression. Clinics (Sao Paulo) 2005, 60, 241–250. [Google Scholar] [CrossRef]
- Shelton, R.C.; Pencina, M.J.; Barrentine, L.W.; Ruiz, J.A.; Fava, M.; Zajecka, J.M.; Papakostas, G.I. Association of obesity and inflammatory marker levels on treatment outcome: results from a double-blind, randomized study of adjunctive L-methylfolate calcium in patients with MDD who are inadequate responders to SSRIs. J Clin Psychiatry 2015, 76, 1635–1641. [Google Scholar] [CrossRef]
- Vreijling, S.R.; Chin Fatt, C.R.; Williams, L.M.; Schatzberg, A.F.; Usherwood, T.; Nemeroff, C.B.; Rush, A.J.; Uher, R.; Aitchison, K.J.; Kohler-Forsberg, O. , et al. Features of immunometabolic depression as predictors of antidepressant treatment outcomes: pooled analysis of four clinical trials. Br J Psychiatry 2024, 224, 89–97. [Google Scholar] [CrossRef]
- Penninx, B.; Lamers, F.; Jansen, R.; Berk, M.; Khandaker, G.M.; De Picker, L.; Milaneschi, Y. Immuno-metabolic depression: from concept to implementation. Lancet Reg Health Eur 2025, 48, 101166. [Google Scholar] [CrossRef]
- Licht, C.M.; de Geus, E.J.; Penninx, B.W. Dysregulation of the autonomic nervous system predicts the development of the metabolic syndrome. J Clin Endocrinol Metab 2013, 98, 2484–2493. [Google Scholar] [CrossRef]
- Vogelzangs, N.; Duivis, H.E.; Beekman, A.T.; Kluft, C.; Neuteboom, J.; Hoogendijk, W.; Smit, J.H.; de Jonge, P.; Penninx, B.W. Association of depressive disorders, depression characteristics and antidepressant medication with inflammation. Transl Psychiatry 2012, 2, e79. [Google Scholar] [CrossRef] [PubMed]
- Licht, C.M.; de Geus, E.J.; Seldenrijk, A.; van Hout, H.P.; Zitman, F.G.; van Dyck, R.; Penninx, B.W. Depression is associated with decreased blood pressure, but antidepressant use increases the risk for hypertension. Hypertension 2009, 53, 631–638. [Google Scholar] [CrossRef]
- Montano, D. Depressive symptoms and blood pressure. Journal of Psychophysiology 2019. [Google Scholar] [CrossRef]
- Hildrum, B.; Mykletun, A.; Stordal, E.; Bjelland, I.; Dahl, A.A.; Holmen, J. Association of low blood pressure with anxiety and depression: the Nord-Trondelag Health Study. J Epidemiol Community Health 2007, 61, 53–58. [Google Scholar] [CrossRef]
- Schaare, H.L.; Blochl, M.; Kumral, D.; Uhlig, M.; Lemcke, L.; Valk, S.L.; Villringer, A. Associations between mental health, blood pressure and the development of hypertension. Nat Commun 2023, 14, 1953. [Google Scholar] [CrossRef]
- Stroup-Benham, C.A.; Markides, K.S.; Black, S.A.; Goodwin, J.S. Relationship between low blood pressure and depressive symptomatology in older people. J Am Geriatr Soc 2000, 48, 250–255. [Google Scholar] [CrossRef]
- Newton, J.L.; Sheth, A.; Shin, J.; Pairman, J.; Wilton, K.; Burt, J.A.; Jones, D.E. Lower ambulatory blood pressure in chronic fatigue syndrome. Psychosom Med 2009, 71, 361–365. [Google Scholar] [CrossRef]
- Halls Dally, J.F. Nervous exhaustion and low blood pressure. The British Medial Journal 1925, 634–635. [Google Scholar] [CrossRef]
- Herrmann-Lingen, C.; Meyer, T.; Bosbach, A.; Chavanon, M.L.; Hassoun, L.; Edelmann, F.; Wachter, R. Cross-Sectional and Longitudinal Associations of Systolic Blood Pressure With Quality of Life and Depressive Mood in Older Adults With Cardiovascular Risk Factors: Results From the Observational DIAST-CHF Study. Psychosom Med 2018, 80, 468–474. [Google Scholar] [CrossRef]
- Berendes, A.; Meyer, T.; Hulpke-Wette, M.; Herrmann-Lingen, C. Association of elevated blood pressure with low distress and good quality of life: results from the nationwide representative German Health Interview and Examination Survey for Children and Adolescents. Psychosom Med 2013, 75, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Hassoun, L.; Herrmann-Lingen, C.; Hapke, U.; Neuhauser, H.; Scheidt-Nave, C.; Meyer, T. Association between chronic stress and blood pressure: findings from the German Health Interview and Examination Survey for Adults 2008-2011. Psychosom Med 2015, 77, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, B.R.; Filewich, R.J.; Miller, N.E.; Craigmyle, N.; Pickering, T.G. Baroreceptor activation reduces reactivity to noxious stimulation: implications for hypertension. Science 1979, 205, 1299–1301. [Google Scholar] [CrossRef]
- Rau, H.; Elbert, T. Psychophysiology of arterial baroreceptors and the etiology of hypertension. Biological psychology 2001, 57, 179–201. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.W.; Chang, Y.; Lim, S.W.; Cho, J.; Kim, H.N.; Kim, K.B.; Kim, J.; Kim, Y.H.; Shin, D.W.; Oh, K.S. , et al. Bidirectional association between blood pressure and depressive symptoms in young and middle-age adults: A cohort study. Epidemiol Psychiatr Sci 2020, 29, e142. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.S.; Berntson, J.; Polanka, B.M.; Stewart, J.C. Cardiovascular Risk Factors as Differential Predictors of Incident Atypical and Typical Major Depressive Disorder in US Adults. Psychosom Med 2018, 80, 508–514. [Google Scholar] [CrossRef]
- Buttner, M.; Jezova, D.; Greene, B.; Konrad, C.; Kircher, T.; Murck, H. Target-based biomarker selection - Mineralocorticoid receptor-related biomarkers and treatment outcome in major depression. J Psychiatr Res 2015, 66-67, 24–37. [Google Scholar] [CrossRef]
- Engelmann, J.; Murck, H.; Wagner, S.; Zillich, L.; Streit, F.; Herzog, D.P.; Braus, D.F.; Tadic, A.; Lieb, K.; Muller, M.B. Routinely accessible parameters of mineralocorticoid receptor function, depression subtypes and response prediction: a post-hoc analysis from the early medication change trial in major depressive disorder. World J Biol Psychiatry 2022, 1–12. [Google Scholar] [CrossRef]
- Luppino, F.S.; Bouvy, P.F.; Giltay, E.J.; Penninx, B.W.; Zitman, F.G. The metabolic syndrome and related characteristics in major depression: inpatients and outpatients compared: metabolic differences across treatment settings. Gen Hosp Psychiatry 2014, 36, 509–515. [Google Scholar] [CrossRef]
- Sunbul, M.; Sunbul, E.A.; Kosker, S.D.; Durmus, E.; Kivrak, T.; Ileri, C.; Oguz, M.; Sari, I. Depression and anxiety are associated with abnormal nocturnal blood pressure fall in hypertensive patients. Clin Exp Hypertens 2014, 36, 354–358. [Google Scholar] [CrossRef]
- Zhao, S.; Fu, S.; Ren, J.; Luo, L. Poor sleep is responsible for the impaired nocturnal blood pressure dipping in elderly hypertensive: A cross-sectional study of elderly. Clin Exp Hypertens 2018, 40, 582–588. [Google Scholar] [CrossRef]
- Okajima, K.; Yamanaka, G.; Oinuma, S.; Kikichi, T.; Yamanaka, T.; Otsuka, K.; Cornelissen, G. Even mild depression is associated with among-day blood pressure variability, including masked non-dipping assessed by 7-d/24-h ambulatory blood pressure monitoring. Clin Exp Hypertens 2015, 37, 426–432. [Google Scholar] [CrossRef]
- Lederbogen, F.; Gernoth, C.; Hamann, B.; Kniest, A.; Heuser, I.; Deuschle, M. Circadian blood pressure regulation in hospitalized depressed patients and non-depressed comparison subjects. Blood Press Monit 2003, 8, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.T.; Huang, C.C. Midodrine hydrochloride in patients on hemodialysis with chronic hypotension. Ren Fail 1996, 18, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Medow, M.S.; Stewart, J.M.; Sanyal, S.; Mumtaz, A.; Sica, D.; Frishman, W.H. Pathophysiology, diagnosis, and treatment of orthostatic hypotension and vasovagal syncope. Cardiol Rev 2008, 16, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Gracie, J.; Newton, J.L.; Norton, M.; Baker, C.; Freeston, M. The role of psychological factors in response to treatment in neurocardiogenic (vasovagal) syncope. Europace 2006, 8, 636–643. [Google Scholar] [CrossRef]
- Otte, C.; Hinkelmann, K.; Moritz, S.; Yassouridis, A.; Jahn, H.; Wiedemann, K.; Kellner, M. Modulation of the mineralocorticoid receptor as add-on treatment in depression: a randomized, double-blind, placebo-controlled proof-of-concept study. J Psychiatr Res 2010, 44, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Czajkowska, J.; Ozhog, S.; Smith, E.; Perlmuter, L.C. Cognition and hopelessness in association with subsyndromal orthostatic hypotension. J Gerontol A Biol Sci Med Sci 2010, 65, 873–879. [Google Scholar] [CrossRef]
- Perlmuter, L.C.; Sarda, G.; Casavant, V.; O’Hara, K.; Hindes, M.; Knott, P.T.; Mosnaim, A.D. A review of orthostatic blood pressure regulation and its association with mood and cognition. Clin Auton Res 2012, 22, 99–107. [Google Scholar] [CrossRef]
- Murck, H.; Schussler, P.; Steiger, A. Renin-angiotensin-aldosterone system: the forgotten stress hormone system: relationship to depression and sleep. Pharmacopsychiatry 2012, 45, 83–95. [Google Scholar] [CrossRef]
- de Kloet, E.R.; Van Acker, S.A.; Sibug, R.M.; Oitzl, M.S.; Meijer, O.C.; Rahmouni, K.; de Jong, W. Brain mineralocorticoid receptors and centrally regulated functions. Kidney Int 2000, 57, 1329–1336. [Google Scholar] [CrossRef]
- Selye, H. Stress and inflammation. Am J Proctol 1953, 4, 229–230. [Google Scholar]
- Hlavacova, N.; Wes, P.D.; Ondrejcakova, M.; Flynn, M.E.; Poundstone, P.K.; Babic, S.; Murck, H.; Jezova, D. Subchronic treatment with aldosterone induces depression-like behaviours and gene expression changes relevant to major depressive disorder. Int J Neuropsychopharmacol 2012, 15, 247–265. [Google Scholar] [CrossRef]
- Joels, M.; de Kloet, E.R. 30 YEARS OF THE MINERALOCORTICOID RECEPTOR: The brain mineralocorticoid receptor: a saga in three episodes. J Endocrinol 2017, 234, T49–T66. [Google Scholar] [CrossRef]
- Goldstein, D.S. Stress and the “extended” autonomic system. Auton Neurosci 2021, 236, 102889. [Google Scholar] [CrossRef]
- Gomez-Sanchez, E.P. Mineralocorticoid receptors in the brain and cardiovascular regulation: minority rule? Trends Endocrinol Metab 2011, 22, 179–187. [Google Scholar] [CrossRef]
- Franklin, M.; Bermudez, I.; Hlavacova, N.; Babic, S.; Murck, H.; Schmuckermair, C.; Singewald, N.; Gaburro, S.; Jezova, D. Aldosterone increases earlier than corticosterone in new animal models of depression: is this an early marker? J Psychiatr Res 2012, 46, 1394–1397. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.; Bermudez, I.; Murck, H.; Singewald, N.; Gaburro, S. Sub-chronic dietary tryptophan depletion--an animal model of depression with improved face and good construct validity. J Psychiatr Res 2012, 46, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.; Hlavacova, N.; Babic, S.; Pokusa, M.; Bermudez, I.; Jezova, D. Aldosterone Signals the Onset of Depressive Behaviour in a Female Rat Model of Depression along with SSRI Treatment Resistance. Neuroendocrinology 2015, 102, 274–287. [Google Scholar] [CrossRef]
- Gideon, A.; Sauter, C.; Fieres, J.; Berger, T.; Renner, B.; Wirtz, P.H. Kinetics and Interrelations of the Renin Aldosterone Response to Acute Psychosocial Stress: A Neglected Stress System. J Clin Endocrinol Metab 2020, 105. [Google Scholar] [CrossRef] [PubMed]
- Makatsori, A.; Duncko, R.; Moncek, F.; Loder, I.; Katina, S.; Jezova, D. Modulation of neuroendocrine response and non-verbal behavior during psychosocial stress in healthy volunteers by the glutamate release-inhibiting drug lamotrigine. Neuroendocrinology 2004, 79, 34–42. [Google Scholar] [CrossRef]
- Murck, H.; Held, K.; Ziegenbein, M.; Kunzel, H.; Koch, K.; Steiger, A. The renin-angiotensin-aldosterone system in patients with depression compared to controls--a sleep endocrine study. BMC Psychiatry 2003, 3, 15. [Google Scholar] [CrossRef]
- Emanuele, E.; Geroldi, D.; Minoretti, P.; Coen, E.; Politi, P. Increased plasma aldosterone in patients with clinical depression. Arch Med Res 2005, 36, 544–548. [Google Scholar] [CrossRef]
- Nowacki, J.; Wingenfeld, K.; Kaczmarczyk, M.; Chae, W.R.; Salchow, P.; Abu-Tir, I.; Piber, D.; Hellmann-Regen, J.; Otte, C. Cardiovascular risk and steroid hormone secretion after stimulation of mineralocorticoid and NMDA receptors in depressed patients. Translational Psychiatry 2020, 10, 109. [Google Scholar] [CrossRef]
- Izakova, L.; Hlavacova, N.; Segeda, V.; Kapsdorfer, D.; Morovicsova, E.; Jezova, D. Salivary aldosterone, cortisol and their morning to evening slopes in patients with depressive disorder and healthy subjects: acute episode and follow up six months after reaching remission. Neuroendocrinology 2020. [Google Scholar] [CrossRef]
- Segeda, V.; Izakova, L.; Hlavacova, N.; Bednarova, A.; Jezova, D. Aldosterone concentrations in saliva reflect the duration and severity of depressive episode in a sex dependent manner. J Psychiatr Res 2017, 91, 164–168. [Google Scholar] [CrossRef]
- Murck, H.; Adolf, C.; Schneider, A.; Schlageter, L.; Heinrich, D.; Ritzel, K.; Sturm, L.; Quinkler, M.; Beuschlein, F.; Reincke, M. , et al. Differential effects of reduced mineralocorticoid receptor activation by unilateral adrenalectomy vs mineralocorticoid antagonist treatment in patients with primary aldosteronism - Implications for depression and anxiety. J Psychiatr Res 2021, 137, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Kunzel, H.E. Psychopathological symptoms in patients with primary hyperaldosteronism--possible pathways. Horm Metab Res 2012, 44, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Bondy, B.; Baghai, T.C.; Zill, P.; Schule, C.; Eser, D.; Deiml, T.; Zwanzger, P.; Ella, R.; Rupprecht, R. Genetic variants in the angiotensin I-converting-enzyme (ACE) and angiotensin II receptor (AT1) gene and clinical outcome in depression. Prog Neuropsychopharmacol Biol Psychiatry 2005, 29, 1094–1099. [Google Scholar] [CrossRef]
- Murck, H.; Schlageter, L.; Schneider, A.; Adolf, C.; Heinrich, D.; Quinkler, M.; Beuschlein, F.; Reincke, M.; Kunzel, H. The potential pathophysiological role of aldosterone and the mineralocorticoid receptor in anxiety and depression - Lessons from primary aldosteronism. J Psychiatr Res 2020, 130, 82–88. [Google Scholar] [CrossRef]
- Shekhtman, E.; Geerling, J.C.; Loewy, A.D. Aldosterone-sensitive neurons of the nucleus of the solitary tract: multisynaptic pathway to the nucleus accumbens. J Comp Neurol 2007, 501, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Forstenpointner, J.; Maallo, A.M.S.; Elman, I.; Holmes, S.; Freeman, R.; Baron, R.; Borsook, D. The solitary nucleus connectivity to key autonomic regions in humans. Eur J Neurosci 2022, 56, 3938–3966. [Google Scholar] [CrossRef]
- Critchley, H.D.; Harrison, N.A. Visceral influences on brain and behavior. Neuron 2013, 77, 624–638. [Google Scholar] [CrossRef]
- Shin, J.W.; Geerling, J.C.; Loewy, A.D. Vagal innervation of the aldosterone-sensitive HSD2 neurons in the NTS. Brain Res 2009, 1249, 135–147. [Google Scholar] [CrossRef]
- Geerling, J.C.; Loewy, A.D. Sodium depletion activates the aldosterone-sensitive neurons in the NTS independently of thirst. Am J Physiol Regul Integr Comp Physiol 2007, 292, R1338–1348. [Google Scholar] [CrossRef]
- Goldstein, P.; Leshem, M. Dietary sodium, added salt, and serum sodium associations with growth and depression in the U.S. general population. Appetite 2014, 79, 83–90. [Google Scholar] [CrossRef]
- Ileri-Gurel, E.; Pehlivanoglu, B.; Dogan, M. Effect of acute stress on taste perception: in relation with baseline anxiety level and body weight. Chem Senses 2013, 38, 27–34. [Google Scholar] [CrossRef]
- Heath, T.P.; Melichar, J.K.; Nutt, D.J.; Donaldson, L.F. Human taste thresholds are modulated by serotonin and noradrenaline. J Neurosci 2006, 26, 12664–12671. [Google Scholar] [CrossRef]
- Guyenet, P.G.; Stornetta, R.L.; Souza, G.; Abbott, S.B.G.; Brooks, V.L. Neuronal Networks in Hypertension: Recent Advances. Hypertension 2020, 76, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Monahan, K.D.; Leuenberger, U.A.; Ray, C.A. Aldosterone impairs baroreflex sensitivity in healthy adults. Am J Physiol Heart Circ Physiol 2007, 292, H190–197. [Google Scholar] [CrossRef] [PubMed]
- Bassi, G.S.; Brognara, F.; Castania, J.A.; Talbot, J.; Cunha, T.M.; Cunha, F.Q.; Ulloa, L.; Kanashiro, A.; Dias, D.P.; Salgado, H.C. Baroreflex activation in conscious rats modulates the joint inflammatory response via sympathetic function. Brain Behav Immun 2015, 49, 140–147. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Kang, Y.M.; Yu, Y.; Wei, S.G.; Schmidt, T.J.; Johnson, A.K.; Felder, R.B. 11beta-hydroxysteroid dehydrogenase type 2 activity in hypothalamic paraventricular nucleus modulates sympathetic excitation. Hypertension 2006, 48, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Csecs, J.L.L.; Dowell, N.G.; Savage, G.K.; Iodice, V.; Mathias, C.J.; Critchley, H.D.; Eccles, J.A. Variant connective tissue (joint hypermobility) and dysautonomia are associated with multimorbidity at the intersection between physical and psychological health. Am J Med Genet C Semin Med Genet 2021, 187, 500–509. [Google Scholar] [CrossRef]
- Juruena, M.F.; Cleare, A.J.; Papadopoulos, A.S.; Poon, L.; Lightman, S.; Pariante, C.M. Different responses to dexamethasone and prednisolone in the same depressed patients. Psychopharmacology (Berl) 2006, 189, 225–235. [Google Scholar] [CrossRef]
- Juruena, M.F.; Pariante, C.M.; Papadopoulos, A.S.; Poon, L.; Lightman, S.; Cleare, A.J. The role of mineralocorticoid receptor function in treatment-resistant depression. J Psychopharmacol 2013, 27, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Vogt, W.; Fischer, I.; Ebenroth, S.; Appel, S.; Knedel, M.; Lucker, P.W.; Rennekamp, H. [Pharmacokinetics of 9 -fluorhydrocortisone]. Arzneimittelforschung 1971, 21, 1133–1143. [Google Scholar]
- Fluharty, S.J.; Epstein, A.N. Sodium appetite elicited by intracerebroventricular infusion of angiotensin II in the rat: II. Synergistic interaction with systemic mineralocorticoids. Behav Neurosci 1983, 97, 746–758. [Google Scholar] [CrossRef]
- Wolf, G.; Handal, P.J. Aldosterone-induced sodium appetite: dose-response and specificity. Endocrinology 1966, 78, 1120–1124. [Google Scholar] [CrossRef]
- Sakai, R.R.; McEwen, B.S.; Fluharty, S.J.; Ma, L.Y. The amygdala: site of genomic and nongenomic arousal of aldosterone-induced sodium intake. Kidney Int 2000, 57, 1337–1345. [Google Scholar] [CrossRef]
- Sakai, R.R.; Nicolaidis, S.; Epstein, A.N. Salt appetite is suppressed by interference with angiotensin II and aldosterone. Am J Physiol 1986, 251, R762–768. [Google Scholar] [CrossRef]
- Graudal, N.A.; Hubeck-Graudal, T.; Jurgens, G. Effects of low sodium diet versus high sodium diet on blood pressure, renin, aldosterone, catecholamines, cholesterol, and triglyceride. The Cochrane database of systematic reviews 2020, 12, CD004022. [Google Scholar] [CrossRef]
- Leshem, M. Low dietary sodium is anxiogenic in rats. Physiol Behav 2011, 103, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Grippo, A.J.; Moffitt, J.A.; Beltz, T.G.; Johnson, A.K. Reduced hedonic behavior and altered cardiovascular function induced by mild sodium depletion in rats. Behav Neurosci 2006, 120, 1133–1143. [Google Scholar] [CrossRef]
- Shimizu, Y.; Kadota, K.; Koyamatsu, J.; Yamanashi, H.; Nagayoshi, M.; Noda, M.; Nishimura, T.; Tayama, J.; Nagata, Y.; Maeda, T. Salt intake and mental distress among rural community-dwelling Japanese men. Journal of physiological anthropology 2015, 34, 26. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.H.; Calvird, M.; Gordon, R.D.; Taylor, P.J.; Ward, G.; Pimenta, E.; Young, R.; Stowasser, M. Effects of two selective serotonin reuptake inhibitor antidepressants, sertraline and escitalopram, on aldosterone/renin ratio in normotensive depressed male patients. J Clin Endocrinol Metab 2011, 96, 1039–1045. [Google Scholar] [CrossRef]
- Vivas, L.; Godino, A.; Dalmasso, C.; Caeiro, X.E.; Macchione, A.F.; Cambiasso, M.J. Neurochemical Circuits Subserving Fluid Balance and Baroreflex: A Role for Serotonin, Oxytocin, and Gonadal Steroids. In Neurobiology of Body Fluid Homeostasis: Transduction and Integration, De Luca, L.A., Jr., Menani, J.V., Johnson, A.K., Eds. Boca Raton (FL), 2014.
- Charloux, A.; Gronfier, C.; Lonsdorfer-Wolf, E.; Piquard, F.; Brandenberger, G. Aldosterone release during the sleep-wake cycle in humans. Am J Physiol 1999, 276, E43–E49. [Google Scholar] [CrossRef] [PubMed]
- Loh, H.H.; Lim, Q.H.; Chai, C.S.; Goh, S.L.; Lim, L.L.; Yee, A.; Sukor, N. Influence and implications of the renin-angiotensin-aldosterone system in obstructive sleep apnea: An updated systematic review and meta-analysis. J Sleep Res 2023, 32, e13726. [Google Scholar] [CrossRef]
- Opstad, P.K.; Oktedalen, O.; Aakvaag, A.; Fonnum, F.; Lund, P.K. Plasma renin activity and serum aldosterone during prolonged physical strain. The significance of sleep and energy deprivation. Eur J Appl Physiol Occup Physiol 1985, 54, 1–6. [Google Scholar] [CrossRef]
- Zorrilla, E.P.; Luborsky, L.; McKay, J.R.; Rosenthal, R.; Houldin, A.; Tax, A.; McCorkle, R.; Seligman, D.A.; Schmidt, K. The relationship of depression and stressors to immunological assays: a meta-analytic review. Brain Behav Immun 2001, 15, 199–226. [Google Scholar] [CrossRef]
- Arteaga-Henriquez, G.; Simon, M.S.; Burger, B.; Weidinger, E.; Wijkhuijs, A.; Arolt, V.; Birkenhager, T.K.; Musil, R.; Muller, N.; Drexhage, H.A. Low-Grade Inflammation as a Predictor of Antidepressant and Anti-Inflammatory Therapy Response in MDD Patients: A Systematic Review of the Literature in Combination With an Analysis of Experimental Data Collected in the EU-MOODINFLAME Consortium. Frontiers in psychiatry 2019, 10, 458. [Google Scholar] [CrossRef]
- Howren, M.B.; Lamkin, D.M.; Suls, J. Associations of depression with C-reactive protein, IL-1, and IL-6: a meta-analysis. Psychosom Med 2009, 71, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Pace, T.W.; Mletzko, T.C.; Alagbe, O.; Musselman, D.L.; Nemeroff, C.B.; Miller, A.H.; Heim, C.M. Increased stress-induced inflammatory responses in male patients with major depression and increased early life stress. Am J Psychiatry 2006, 163, 1630–1633. [Google Scholar] [CrossRef]
- Garcia Bueno, B.; Caso, J.R.; Madrigal, J.L.; Leza, J.C. Innate immune receptor Toll-like receptor 4 signalling in neuropsychiatric diseases. Neurosci Biobehav Rev 2016, 64, 134–147. [Google Scholar] [CrossRef]
- Bay-Richter, C.; Janelidze, S.; Hallberg, L.; Brundin, L. Changes in behaviour and cytokine expression upon a peripheral immune challenge. Behav Brain Res 2011, 222, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Frenois, F.; Moreau, M.; O’Connor, J.; Lawson, M.; Micon, C.; Lestage, J.; Kelley, K.W.; Dantzer, R.; Castanon, N. Lipopolysaccharide induces delayed FosB/DeltaFosB immunostaining within the mouse extended amygdala, hippocampus and hypothalamus, that parallel the expression of depressive-like behavior. Psychoneuroendocrinology 2007, 32, 516–531. [Google Scholar] [CrossRef]
- Yirmiya, R. Endotoxin produces a depressive-like episode in rats. Brain Res 1996, 711, 163–174. [Google Scholar] [CrossRef]
- Vedder, H.; Schreiber, W.; Schuld, A.; Kainz, M.; Lauer, C.J.; Krieg, J.C.; Holsboer, F.; Pollmacher, T. Immune-endocrine host response to endotoxin in major depression. J Psychiatr Res 2007, 41, 280–289. [Google Scholar] [CrossRef]
- Maes, M.; Kubera, M.; Leunis, J.C. The gut-brain barrier in major depression: intestinal mucosal dysfunction with an increased translocation of LPS from gram negative enterobacteria (leaky gut) plays a role in the inflammatory pathophysiology of depression. Neuro endocrinology letters 2008, 29, 117–124. [Google Scholar]
- Kizaki, T.; Shirato, K.; Sakurai, T.; Ogasawara, J.E.; Oh-ishi, S.; Matsuoka, T.; Izawa, T.; Imaizumi, K.; Haga, S.; Ohno, H. Beta2-adrenergic receptor regulate Toll-like receptor 4-induced late-phase NF-kappaB activation. Mol Immunol 2009, 46, 1195–1203. [Google Scholar] [CrossRef]
- Glezer, I.; Zekki, H.; Scavone, C.; Rivest, S. Modulation of the innate immune response by NMDA receptors has neuropathological consequences. J Neurosci 2003, 23, 11094–11103. [Google Scholar] [CrossRef] [PubMed]
- Glezer, I.; Simard, A.R.; Rivest, S. Neuroprotective role of the innate immune system by microglia. Neuroscience 2007, 147, 867–883. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.; Wu, G.; Jiang, Z. Glycyrrhizin Treatment Facilitates Extinction of Conditioned Fear Responses After a Single Prolonged Stress Exposure in Rats. Cell Physiol Biochem 2018, 45, 2529–2539. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Wang, Y.; Chen, K.; Long, Z.; Zou, J. Ketamine Alleviates Depressive-Like Behaviors via Down-Regulating Inflammatory Cytokines Induced by Chronic Restraint Stress in Mice. Biol Pharm Bull 2017, 40, 1260–1267. [Google Scholar] [CrossRef]
- Garate, I.; Garcia-Bueno, B.; Madrigal, J.L.; Bravo, L.; Berrocoso, E.; Caso, J.R.; Mico, J.A.; Leza, J.C. Origin and consequences of brain Toll-like receptor 4 pathway stimulation in an experimental model of depression. J Neuroinflammation 2011, 8, 151. [Google Scholar] [CrossRef]
- Garate, I.; Garcia-Bueno, B.; Madrigal, J.L.; Caso, J.R.; Alou, L.; Gomez-Lus, M.L.; Mico, J.A.; Leza, J.C. Stress-induced neuroinflammation: role of the Toll-like receptor-4 pathway. Biol Psychiatry 2013, 73, 32–43. [Google Scholar] [CrossRef]
- Garate, I.; Garcia-Bueno, B.; Madrigal, J.L.; Caso, J.R.; Alou, L.; Gomez-Lus, M.L.; Leza, J.C. Toll-like 4 receptor inhibitor TAK-242 decreases neuroinflammation in rat brain frontal cortex after stress. J Neuroinflammation 2014, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Wisor, J.P.; Clegern, W.C.; Schmidt, M.A. Toll-like receptor 4 is a regulator of monocyte and electroencephalographic responses to sleep loss. Sleep 2011, 34, 1335–1345. [Google Scholar] [CrossRef]
- Xia, Y.; Yamagata, K.; Krukoff, T.L. Differential expression of the CD14/TLR4 complex and inflammatory signaling molecules following i.c.v. administration of LPS. Brain Res 2006, 1095, 85–95. [Google Scholar] [CrossRef]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. Reflex control of immunity. Nat Rev Immunol 2009, 9, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Vance, K.M.; Ribnicky, D.M.; Hermann, G.E.; Rogers, R.C. St. John’s Wort enhances the synaptic activity of the nucleus of the solitary tract. Nutrition 2014, 30, S37–42. [Google Scholar] [CrossRef]
- Sartor, D.M.; Verberne, A.J. The role of NMDA and non-NMDA receptors in the NTS in mediating three distinct sympathoinhibitory reflexes. Naunyn Schmiedebergs Arch Pharmacol 2007, 376, 241–252. [Google Scholar] [CrossRef]
- Nakano, Y.; Furube, E.; Morita, S.; Wanaka, A.; Nakashima, T.; Miyata, S. Astrocytic TLR4 expression and LPS-induced nuclear translocation of STAT3 in the sensory circumventricular organs of adult mouse brain. J Neuroimmunol 2015, 278, 144–158. [Google Scholar] [CrossRef]
- Hosoi, T.; Okuma, Y.; Matsuda, T.; Nomura, Y. Novel pathway for LPS-induced afferent vagus nerve activation: possible role of nodose ganglion. Auton Neurosci 2005, 120, 104–107. [Google Scholar] [CrossRef]
- Halaris, A. Inflammation and depression but where does the inflammation come from? Curr Opin Psychiatry 2019, 32, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Kenney, M.J.; Ganta, C.K. Autonomic nervous system and immune system interactions. Comprehensive Physiology 2014, 4, 1177–1200. [Google Scholar] [CrossRef]
- Pongratz, G.; Straub, R.H. The sympathetic nervous response in inflammation. Arthritis Res Ther 2014, 16, 504. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.N.; Tepper, P.; Barinas-Mitchell, E.; Woodard, G.A.; Sutton-Tyrrell, K. Serum aldosterone is associated with inflammation and aortic stiffness in normotensive overweight and obese young adults. Clin Exp Hypertens 2012, 34, 63–70. [Google Scholar] [CrossRef]
- Cooper, J.N.; Fried, L.; Tepper, P.; Barinas-Mitchell, E.; Conroy, M.B.; Evans, R.W.; Mori Brooks, M.; Woodard, G.A.; Sutton-Tyrrell, K. Changes in serum aldosterone are associated with changes in obesity-related factors in normotensive overweight and obese young adults. Hypertension research : official journal of the Japanese Society of Hypertension 2013, 36, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Bay-Richter, C.; Hallberg, L.; Ventorp, F.; Janelidze, S.; Brundin, L. Aldosterone synergizes with peripheral inflammation to induce brain IL-1beta expression and depressive-like effects. Cytokine 2012, 60, 749–754. [Google Scholar] [CrossRef]
- Sanchez-Lemus, E.; Murakami, Y.; Larrayoz-Roldan, I.M.; Moughamian, A.J.; Pavel, J.; Nishioku, T.; Saavedra, J.M. Angiotensin II AT1 receptor blockade decreases lipopolysaccharide-induced inflammation in the rat adrenal gland. Endocrinology 2008, 149, 5177–5188. [Google Scholar] [CrossRef]
- Benicky, J.; Sanchez-Lemus, E.; Honda, M.; Pang, T.; Orecna, M.; Wang, J.; Leng, Y.; Chuang, D.M.; Saavedra, J.M. Angiotensin II AT1 receptor blockade ameliorates brain inflammation. Neuropsychopharmacology 2011, 36, 857–870. [Google Scholar] [CrossRef]
- Libianto, R.; Hu, J.; Chee, M.R.; Hoo, J.; Lim, Y.Y.; Shen, J.; Li, Q.; Young, M.J.; Fuller, P.J.; Yang, J. A Multicenter Study of Neutrophil-to-Lymphocyte Ratio in Primary Aldosteronism. J Endocr Soc 2020, 4, bvaa153. [Google Scholar] [CrossRef]
- Herrada, A.A.; Contreras, F.J.; Marini, N.P.; Amador, C.A.; Gonzalez, P.A.; Cortes, C.M.; Riedel, C.A.; Carvajal, C.A.; Figueroa, F.; Michea, L.F. , et al. Aldosterone promotes autoimmune damage by enhancing Th17-mediated immunity. J Immunol 2010, 184, 191–202. [Google Scholar] [CrossRef]
- Odermatt, A.; Kratschmar, D.V. Tissue-specific modulation of mineralocorticoid receptor function by 11beta-hydroxysteroid dehydrogenases: an overview. Mol Cell Endocrinol 2012, 350, 168–186. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef]
- Kloiber, S.; Ising, M.; Reppermund, S.; Horstmann, S.; Dose, T.; Majer, M.; Zihl, J.; Pfister, H.; Unschuld, P.G.; Holsboer, F. , et al. Overweight and obesity affect treatment response in major depression. Biol Psychiatry 2007, 62, 321–326. [Google Scholar] [CrossRef]
- Jantaratnotai, N.; Mosikanon, K.; Lee, Y.; McIntyre, R.S. The interface of depression and obesity. Obes Res Clin Pract 2017, 11, 1–10. [Google Scholar] [CrossRef]
- Lasserre, A.M.; Glaus, J.; Vandeleur, C.L.; Marques-Vidal, P.; Vaucher, J.; Bastardot, F.; Waeber, G.; Vollenweider, P.; Preisig, M. Depression with atypical features and increase in obesity, body mass index, waist circumference, and fat mass: a prospective, population-based study. JAMA psychiatry 2014, 71, 880–888. [Google Scholar] [CrossRef]
- Lasserre, A.M.; Strippoli, M.F.; Glaus, J.; Gholam-Rezaee, M.; Vandeleur, C.L.; Castelao, E.; Marques-Vidal, P.; Waeber, G.; Vollenweider, P.; Preisig, M. Prospective associations of depression subtypes with cardio-metabolic risk factors in the general population. Mol Psychiatry 2017, 22, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.E.; Freedland, K.E.; Carney, R.M.; Stetler, C.A.; Banks, W.A. Pathways linking depression, adiposity, and inflammatory markers in healthy young adults. Brain Behav Immun 2003, 17, 276–285. [Google Scholar] [CrossRef]
- van Reedt Dortland, A.K.; Vreeburg, S.A.; Giltay, E.J.; Licht, C.M.; Vogelzangs, N.; van Veen, T.; de Geus, E.J.; Penninx, B.W.; Zitman, F.G. The impact of stress systems and lifestyle on dyslipidemia and obesity in anxiety and depression. Psychoneuroendocrinology 2013, 38, 209–218. [Google Scholar] [CrossRef]
- van Reedt Dortland, A.K.; Giltay, E.J.; van Veen, T.; van Pelt, J.; Zitman, F.G.; Penninx, B.W. Associations between serum lipids and major depressive disorder: results from the Netherlands Study of Depression and Anxiety (NESDA). J Clin Psychiatry 2010, 71, 729–736. [Google Scholar] [CrossRef]
- van Reedt Dortland, A.K.; Giltay, E.J.; van Veen, T.; Zitman, F.G.; Penninx, B.W. Longitudinal relationship of depressive and anxiety symptoms with dyslipidemia and abdominal obesity. Psychosom Med 2013, 75, 83–89. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Li, X.; Zhang, Y.; Gulbins, E.; Zhang, Y. Enhancement of endothelial permeability by free fatty acid through lysosomal cathepsin B-mediated Nlrp3 inflammasome activation. Oncotarget 2016, 7, 73229–73241. [Google Scholar] [CrossRef]
- Pacurari, M.; Kafoury, R.; Tchounwou, P.B.; Ndebele, K. The Renin-Angiotensin-aldosterone system in vascular inflammation and remodeling. Int J Inflam 2014, 2014, 689360. [Google Scholar] [CrossRef]
- Duprez, D.A. Role of the renin-angiotensin-aldosterone system in vascular remodeling and inflammation: a clinical review. J Hypertens 2006, 24, 983–991. [Google Scholar] [CrossRef] [PubMed]
- de Kloet, A.D.; Pioquinto, D.J.; Nguyen, D.; Wang, L.; Smith, J.A.; Hiller, H.; Sumners, C. Obesity induces neuroinflammation mediated by altered expression of the renin-angiotensin system in mouse forebrain nuclei. Physiol Behav, 2014. [Google Scholar] [CrossRef]
- Pires, P.W.; McClain, J.L.; Hayoz, S.F.; Dorrance, A.M. Mineralocorticoid receptor antagonism prevents obesity-induced cerebral artery remodeling and reduces white matter injury in rats. Microcirculation 2018, 25, e12460. [Google Scholar] [CrossRef] [PubMed]
- Irani, H.; Abiri, B.; Khodami, B.; Yari, Z.; Lafzi Ghazi, M.; Hosseinzadeh, N.; Saidpour, A. Effect of time restricted feeding on anthropometric measures, eating behavior, stress, serum levels of BDNF and LBP in overweight/obese women with food addiction: a randomized clinical trial. Nutr Neurosci 2024, 27, 577–589. [Google Scholar] [CrossRef]
- Law, S.; Dong, S.; Zhou, F.; Zheng, D.; Wang, C.; Dong, Z. Bariatric surgery and mental health outcomes: an umbrella review. Front Endocrinol (Lausanne) 2023, 14, 1283621. [Google Scholar] [CrossRef] [PubMed]
- Peterhansel, C.; Petroff, D.; Klinitzke, G.; Kersting, A.; Wagner, B. Risk of completed suicide after bariatric surgery: a systematic review. Obes Rev 2013, 14, 369–382. [Google Scholar] [CrossRef]
- Zade, D.; Beiser, A.; McGlinchey, R.; Au, R.; Seshadri, S.; Palumbo, C.; Wolf, P.A.; DeCarli, C.; Milberg, W. Apolipoprotein epsilon 4 allele modifies waist-to-hip ratio effects on cognition and brain structure. J Stroke Cerebrovasc Dis 2013, 22, 119–125. [Google Scholar] [CrossRef]
- Willette, A.A.; Kapogiannis, D. Does the brain shrink as the waist expands? Ageing Res Rev 2015, 20, 86–97. [Google Scholar] [CrossRef]
- Pontiroli, A.E.; Merlotti, C.; Veronelli, A.; Lombardi, F. Effect of weight loss on sympatho-vagal balance in subjects with grade-3 obesity: restrictive surgery versus hypocaloric diet. Acta Diabetol 2013, 50, 843–850. [Google Scholar] [CrossRef]
- Hassenstab, J.J.; Sweet, L.H.; Del Parigi, A.; McCaffery, J.M.; Haley, A.P.; Demos, K.E.; Cohen, R.A.; Wing, R.R. Cortical thickness of the cognitive control network in obesity and successful weight loss maintenance: a preliminary MRI study. Psychiatry Res 2012, 202, 77–79. [Google Scholar] [CrossRef]
- Dreimuller, N.; Lieb, K.; Tadic, A.; Engelmann, J.; Wollschlager, D.; Wagner, S. Body mass index (BMI) in major depressive disorder and its effects on depressive symptomatology and antidepressant response. J Affect Disord 2019, 256, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Angst, J.; Gamma, A.; Benazzi, F.; Silverstein, B.; Ajdacic-Gross, V.; Eich, D.; Rossler, W. Atypical depressive syndromes in varying definitions. Eur Arch Psychiatry Clin Neurosci 2005. [Google Scholar] [CrossRef]
- Novick, J.S.; Stewart, J.W.; Wisniewski, S.R.; Cook, I.A.; Manev, R.; Nierenberg, A.A.; Rosenbaum, J.F.; Shores-Wilson, K.; Balasubramani, G.K.; Biggs, M.M. , et al. Clinical and demographic features of atypical depression in outpatients with major depressive disorder: preliminary findings from STAR*D. J Clin Psychiatry 2005, 66, 1002–1011. [Google Scholar] [CrossRef]
- Matza, L.S.; Revicki, D.A.; Davidson, J.R.; Stewart, J.W. Depression with atypical features in the National Comorbidity Survey: classification, description, and consequences. Arch Gen Psychiatry 2003, 60, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Quitkin, F.M.; Stewart, J.W.; McGrath, P.J.; Tricamo, E.; Rabkin, J.G.; Ocepek-Welikson, K.; Nunes, E.; Harrison, W.; Klein, D.F. Columbia atypical depression. A subgroup of depressives with better response to MAOI than to tricyclic antidepressants or placebo. Br J Psychiatry Suppl 1993, 30–34. [Google Scholar] [CrossRef]
- Mannel, M.; Kuhn, U.; Schmidt, U.; Ploch, M.; Murck, H. St. John’s wort extract LI160 for the treatment of depression with atypical features - a double-blind, randomized, and placebo-controlled trial. J Psychiatr Res 2010, 44, 760–767. [Google Scholar] [CrossRef]
- Murck, H.; Fava, M.; Alpert, J.; Nierenberg, A.A.; Mischoulon, D.; Otto, M.W.; Zajecka, J.; Mannel, M.; Rosenbaum, J.F. Hypericum extract in patients with MDD and reversed vegetative signs: re-analysis from data of a double-blind, randomized trial of hypericum extract, fluoxetine, and placebo. Int J Neuropsychopharmacol 2005, 8, 215–221. [Google Scholar] [CrossRef]
- Schroeder, C.; Tank, J.; Goldstein, D.S.; Stoeter, M.; Haertter, S.; Luft, F.C.; Jordan, J. Influence of St John’s wort on catecholamine turnover and cardiovascular regulation in humans. Clinical pharmacology and therapeutics 2004, 76, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Bekhbat, M.; Li, Z.; Dunlop, B.W.; Treadway, M.T.; Mehta, N.D.; Revill, K.P.; Lucido, M.J.; Hong, C.; Ashchi, A.; Wommack, E.C. , et al. Sustained effects of repeated levodopa (L-DOPA) administration on reward circuitry, effort-based motivation, and anhedonia in depressed patients with higher inflammation. Brain Behav Immun 2025, 125, 240–248. [Google Scholar] [CrossRef]
- Roitman, M.F.; Patterson, T.A.; Sakai, R.R.; Bernstein, I.L.; Figlewicz, D.P. Sodium depletion and aldosterone decrease dopamine transporter activity in nucleus accumbens but not striatum. Am J Physiol 1999, 276, R1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.X.; Penninx, B.; de Geus, E.J.C.; Lamers, F.; Kuan, D.C.; Wright, A.G.C.; Marsland, A.L.; Muldoon, M.F.; Manuck, S.B.; Gianaros, P.J. Associations of immunometabolic risk factors with symptoms of depression and anxiety: The role of cardiac vagal activity. Brain Behav Immun 2018, 73, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Licht, C.M.; Vreeburg, S.A.; van Reedt Dortland, A.K.; Giltay, E.J.; Hoogendijk, W.J.; DeRijk, R.H.; Vogelzangs, N.; Zitman, F.G.; de Geus, E.J.; Penninx, B.W. Increased sympathetic and decreased parasympathetic activity rather than changes in hypothalamic-pituitary-adrenal axis activity is associated with metabolic abnormalities. J Clin Endocrinol Metab 2010, 95, 2458–2466. [Google Scholar] [CrossRef]
- Warne, J.P.; Foster, M.T.; Horneman, H.F.; Pecoraro, N.C.; Ginsberg, A.B.; Akana, S.F.; Dallman, M.F. Afferent signalling through the common hepatic branch of the vagus inhibits voluntary lard intake and modifies plasma metabolite levels in rats. J Physiol 2007, 583, 455–467. [Google Scholar] [CrossRef]
- Windham, B.G.; Fumagalli, S.; Ble, A.; Sollers, J.J.; Thayer, J.F.; Najjar, S.S.; Griswold, M.E.; Ferrucci, L. The Relationship between Heart Rate Variability and Adiposity Differs for Central and Overall Adiposity. J Obes 2012, 2012, 149516. [Google Scholar] [CrossRef]
- Ziegler, D.; Strom, A.; Kupriyanova, Y.; Bierwagen, A.; Bonhof, G.J.; Bodis, K.; Mussig, K.; Szendroedi, J.; Bobrov, P.; Markgraf, D.F. , et al. Association of Lower Cardiovagal Tone and Baroreflex Sensitivity With Higher Liver Fat Content Early in Type 2 Diabetes. J Clin Endocrinol Metab 2018, 103, 1130–1138. [Google Scholar] [CrossRef]
- Hua, K.; Usichenko, T.; Cummings, M.; Bernatik, M.; Willich, S.N.; Brinkhaus, B.; Dietzel, J. Effects of auricular stimulation on weight- and obesity-related parameters: a systematic review and meta-analysis of randomized controlled clinical trials. Front Neurosci 2024, 18, 1393826. [Google Scholar] [CrossRef]
- Apovian, C.M.; Shah, S.N.; Wolfe, B.M.; Ikramuddin, S.; Miller, C.J.; Tweden, K.S.; Billington, C.J.; Shikora, S.A. Two-Year Outcomes of Vagal Nerve Blocking (vBloc) for the Treatment of Obesity in the ReCharge Trial. Obes Surg 2017, 27, 169–176. [Google Scholar] [CrossRef]
- Catinis, A.M.; Hinojosa, A.J.; Leonardi, C.; Cook, M.W. Hepatic Vagotomy in Patients With Obesity Leads to Improvement of the Cholesterol to High-Density Lipoprotein Ratio. Obes Surg 2023, 33, 3740–3745. [Google Scholar] [CrossRef]
- Lubaczeuski, C.; Balbo, S.L.; Ribeiro, R.A.; Vettorazzi, J.F.; Santos-Silva, J.C.; Carneiro, E.M.; Bonfleur, M.L. Vagotomy ameliorates islet morphofunction and body metabolic homeostasis in MSG-obese rats. Braz J Med Biol Res 2015, 48, 447–457. [Google Scholar] [CrossRef]
- Zou, J.; Li, J.; Wang, X.; Tang, D.; Chen, R. Neuroimmune modulation in liver pathophysiology. J Neuroinflammation 2024, 21, 188. [Google Scholar] [CrossRef] [PubMed]
- Geerling, J.J.; Boon, M.R.; Kooijman, S.; Parlevliet, E.T.; Havekes, L.M.; Romijn, J.A.; Meurs, I.M.; Rensen, P.C. Sympathetic nervous system control of triglyceride metabolism: novel concepts derived from recent studies. J Lipid Res 2014, 55, 180–189. [Google Scholar] [CrossRef]
- Murck, H. Neuroendocrine and Autonomic Dysregulation in Affective Disorders. In Handbook of the Biology and Pathology of Mental Disorders, Springer: 2025; pp. 1-23.
- Otte, C.; Chae, W.R.; Dogan, D.Y.; Piber, D.; Roepke, S.; Cho, A.B.; Trumm, S.; Kaczmarczyk, M.; Brasanac, J.; Wingenfeld, K. , et al. Simvastatin as Add-On Treatment to Escitalopram in Patients With Major Depression and Obesity: A Randomized Clinical Trial. JAMA psychiatry 2025. [Google Scholar] [CrossRef]
- Murck, H.; Luerweg, B.; Hahn, J.; Braunisch, M.; Jezova, D.; Zavorotnyy, M.; Konrad, C.; Jansen, A.; Kircher, T. Ventricular volume, white matter alterations and outcome of major depression and their relationship to endocrine parameters - A pilot study. World J Biol Psychiatry 2020, 1–15. [Google Scholar] [CrossRef]
- Murck, H.; Fava, M.; Cusin, C.; Fatt, C.C.; Trivedi, M. Brain ventricle and choroid plexus morphology as predictor of treatment response in major depression: Findings from the EMBARC study. Brain Behav Immun Health 2024, 35, 100717. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, V.; Gonzalez-Escamilla, G.; Ciolac, D.; Albrecht, P.; Kury, P.; Gruchot, J.; Dietrich, M.; Hecker, C.; Muntefering, T.; Bock, S. , et al. Translational value of choroid plexus imaging for tracking neuroinflammation in mice and humans. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef]
- Hagenberg, J.; Be, C.s.g.; group, O.s.; Bruckl, T.M.; Erhart, M.; Kopf-Beck, J.; Kodel, M.; Rehawi, G.; Roh-Karamihalev, S.; Sauer, S. , et al. Dissecting depression symptoms: Multi-omics clustering uncovers immune-related subgroups and cell-type specific dysregulation. Brain Behav Immun 2025, 123, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Bravi, B.; Melloni, E.M.T.; Paolini, M.; Palladini, M.; Calesella, F.; Servidio, L.; Agnoletto, E.; Poletti, S.; Lorenzi, C.; Colombo, C. , et al. Choroid plexus volume is increased in mood disorders and associates with circulating inflammatory cytokines. Brain Behav Immun 2023, 116, 52–61. [Google Scholar] [CrossRef]
- Cao, Y.; Lizano, P.; Deng, G.; Sun, H.; Zhou, X.; Xie, H.; Zhan, Y.; Mu, J.; Long, X.; Xiao, H. , et al. Brain-derived subgroups of bipolar II depression associate with inflammation and choroid plexus morphology. Psychiatry and clinical neurosciences 2023, 77, 613–621. [Google Scholar] [CrossRef]
- Althubaity, N.; Schubert, J.; Martins, D.; Yousaf, T.; Nettis, M.A.; Mondelli, V.; Pariante, C.; Harrison, N.A.; Bullmore, E.T.; Dima, D. , et al. Choroid plexus enlargement is associated with neuroinflammation and reduction of blood brain barrier permeability in depression. Neuroimage Clin 2022, 33, 102926. [Google Scholar] [CrossRef]
- Turner, C.A.; Thompson, R.C.; Bunney, W.E.; Schatzberg, A.F.; Barchas, J.D.; Myers, R.M.; Akil, H.; Watson, S.J. Altered choroid plexus gene expression in major depressive disorder. Frontiers in human neuroscience 2014, 8, 238. [Google Scholar] [CrossRef]
- Carvajal, C.A.; Herrada, A.A.; Castillo, C.R.; Contreras, F.J.; Stehr, C.B.; Mosso, L.M.; Kalergis, A.M.; Fardella, C.E. Primary aldosteronism can alter peripheral levels of transforming growth factor beta and tumor necrosis factor alpha. J Endocrinol Invest 2009, 32, 759–765. [Google Scholar] [CrossRef]
- Macmaster, F.P.; Carrey, N.; Marie Langevin, L. Corpus callosal morphology in early onset adolescent depression. J Affect Disord 2013, 145, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Murck, H.; Lehr, L.; Jezova, D. A viewpoint on aldosterone and BMI related brain morphology in relation to treatment outcome in patients with major depression. J Neuroendocrinol 2023, 35, e13219. [Google Scholar] [CrossRef] [PubMed]
- Dempster, K.S.; O’Leary, D.D.; MacNeil, A.J.; Hodges, G.J.; Wade, T.J. Linking the hemodynamic consequences of adverse childhood experiences to an altered HPA axis and acute stress response. Brain Behav Immun 2021, 93, 254–263. [Google Scholar] [CrossRef]
- Terock, J.; Hannemann, A.; Klinger-Konig, J.; Janowitz, D.; Grabe, H.J.; Murck, H. The neurobiology of childhood trauma-aldosterone and blood pressure changes in a community sample. World J Biol Psychiatry 2022, 1–9. [Google Scholar] [CrossRef]
- De Bellis, M.D.; Zisk, A. The biological effects of childhood trauma. Child Adolesc Psychiatr Clin N Am 2014, 23, 185–222. [Google Scholar] [CrossRef] [PubMed]
- De Bellis, M.D.; Keshavan, M.S.; Shifflett, H.; Iyengar, S.; Beers, S.R.; Hall, J.; Moritz, G. Brain structures in pediatric maltreatment-related posttraumatic stress disorder: a sociodemographically matched study. Biol Psychiatry 2002, 52, 1066–1078. [Google Scholar] [CrossRef]
- Cao, Y.; Lizano, P.; Li, M.; Colic, L.; Chand, T.; Javaheripour, N.; Sun, H.; Deng, G.; Zhou, X.; Long, X. Associations among bipolar II depression white matter subgroups, inflammation, symptoms and childhood maltreatment. Nature Mental Health 2025, 1–11. [Google Scholar] [CrossRef]
- Nelson, J.; Klumparendt, A.; Doebler, P.; Ehring, T. Childhood maltreatment and characteristics of adult depression: meta-analysis. Br J Psychiatry 2017, 210, 96–104. [Google Scholar] [CrossRef]
- Lippard, E.T.C.; Nemeroff, C.B. The Devastating Clinical Consequences of Child Abuse and Neglect: Increased Disease Vulnerability and Poor Treatment Response in Mood Disorders. Am J Psychiatry 2020, 177, 20–36. [Google Scholar] [CrossRef] [PubMed]
- Noetel, M.; Sanders, T.; Gallardo-Gomez, D.; Taylor, P.; Del Pozo Cruz, B.; van den Hoek, D.; Smith, J.J.; Mahoney, J.; Spathis, J.; Moresi, M. , et al. Effect of exercise for depression: systematic review and network meta-analysis of randomised controlled trials. BMJ 2024, 384, e075847. [Google Scholar] [CrossRef]
- Rethorst, C.D.; Tu, J.; Carmody, T.J.; Greer, T.L.; Trivedi, M.H. Atypical depressive symptoms as a predictor of treatment response to exercise in Major Depressive Disorder. J Affect Disord 2016, 200, 156–158. [Google Scholar] [CrossRef]
- Toni, G.; Belvederi Murri, M.; Piepoli, M.; Zanetidou, S.; Cabassi, A.; Squatrito, S.; Bagnoli, L.; Piras, A.; Mussi, C.; Senaldi, R. , et al. Physical Exercise for Late-Life Depression: Effects on Heart Rate Variability. Am J Geriatr Psychiatry 2016, 24, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Baffour-Awuah, B.; Man, M.; Goessler, K.F.; Cornelissen, V.A.; Dieberg, G.; Smart, N.A.; Pearson, M.J. Effect of exercise training on the renin-angiotensin-aldosterone system: a meta-analysis. J Hum Hypertens 2024, 38, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Berney, M.; Vakilzadeh, N.; Maillard, M.; Faouzi, M.; Grouzmann, E.; Bonny, O.; Favre, L.; Wuerzner, G. Bariatric Surgery Induces a Differential Effect on Plasma Aldosterone in Comparison to Dietary Advice Alone. Front Endocrinol (Lausanne) 2021, 12, 745045. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, G.; Xu, M.; Cai, W.; Zhu, Q.; Qian, L.; Zhang, Y.E.; Yuan, K.; Liu, J.; Li, Q. , et al. Recovery of brain structural abnormalities in morbidly obese patients after bariatric surgery. Int J Obes (Lond) 2016, 40, 1558–1565. [Google Scholar] [CrossRef]
- Murck, H.; Lehr, L.; Hahn, J.; Braunisch, M.C.; Jezova, D.; Zavorotnyy, M. Adjunct Therapy With Glycyrrhiza Glabra Rapidly Improves Outcome in Depression-A Pilot Study to Support 11-Beta-Hydroxysteroid Dehydrogenase Type 2 Inhibition as a New Target. Frontiers in psychiatry 2020, 11, 605949. [Google Scholar] [CrossRef]
- Cosmetic Ingredient Review Expert, P. Final report on the safety assessment of Glycyrrhetinic Acid, Potassium Glycyrrhetinate, Disodium Succinoyl Glycyrrhetinate, Glyceryl Glycyrrhetinate, Glycyrrhetinyl Stearate, Stearyl Glycyrrhetinate, Glycyrrhizic Acid, Ammonium Glycyrrhizate, Dipotassium Glycyrrhizate, Disodium Glycyrrhizate, Trisodium Glycyrrhizate, Methyl Glycyrrhizate, and Potassium Glycyrrhizinate. Int J Toxicol 2007, 26 Suppl 2, 79–112. [Google Scholar] [CrossRef]
- Yamamura, Y.; Kawakami, J.; Santa, T.; Kotaki, H.; Uchino, K.; Sawada, Y.; Tanaka, N.; Iga, T. Pharmacokinetic profile of glycyrrhizin in healthy volunteers by a new high-performance liquid chromatographic method. J Pharm Sci 1992, 81, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Krahenbuhl, S.; Hasler, F.; Frey, B.M.; Frey, F.J.; Brenneisen, R.; Krapf, R. Kinetics and dynamics of orally administered 18 beta-glycyrrhetinic acid in humans. J Clin Endocrinol Metab 1994, 78, 581–585. [Google Scholar] [CrossRef]
- Isbrucker, R.A.; Burdock, G.A. Risk and safety assessment on the consumption of Licorice root (Glycyrrhiza sp.), its extract and powder as a food ingredient, with emphasis on the pharmacology and toxicology of glycyrrhizin. Regul Toxicol Pharmacol 2006, 46, 167–192. [Google Scholar] [CrossRef]
- van Gelderen, C.E.; Bijlsma, J.A.; van Dokkum, W.; Savelkoul, T.J. Glycyrrhizic acid: the assessment of a no effect level. Hum Exp Toxicol 2000, 19, 434–439. [Google Scholar] [CrossRef]
- Sigurjonsdottir, H.A.; Franzson, L.; Manhem, K.; Ragnarsson, J.; Sigurdsson, G.; Wallerstedt, S. Liquorice-induced rise in blood pressure: a linear dose-response relationship. J Hum Hypertens 2001, 15, 549–552. [Google Scholar] [CrossRef]
- Forslund, T.; Fyhrquist, F.; Froseth, B.; Tikkanen, I. Effects of licorice on plasma atrial natriuretic peptide in healthy volunteers. J Intern Med 1989, 225, 95–99. [Google Scholar] [CrossRef]
- Epstein, M.T.; Espiner, E.A.; Donald, R.A.; Hughes, H. Effect of eating liquorice on the renin-angiotensin aldosterone axis in normal subjects. Br Med J 1977, 1, 488–490. [Google Scholar] [CrossRef]
- Armanini, D.; Lewicka, S.; Pratesi, C.; Scali, M.; Zennaro, M.C.; Zovato, S.; Gottardo, C.; Simoncini, M.; Spigariol, A.; Zampollo, V. Further studies on the mechanism of the mineralocorticoid action of licorice in humans. J Endocrinol Invest 1996, 19, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Af Geijerstam, P.; Joelsson, A.; Radholm, K.; Nystrom, F.H. A low dose of daily licorice intake affects renin, aldosterone, and home blood pressure in a randomized crossover trial. Am J Clin Nutr 2024, 119, 682–691. [Google Scholar] [CrossRef]
- Walker, B.R.; Connacher, A.A.; Webb, D.J.; Edwards, C.R. Glucocorticoids and blood pressure: a role for the cortisol/cortisone shuttle in the control of vascular tone in man. Clin Sci (Lond) 1992, 83, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Hautaniemi, E.J.; Tahvanainen, A.M.; Koskela, J.K.; Tikkakoski, A.J.; Kahonen, M.; Uitto, M.; Sipila, K.; Niemela, O.; Mustonen, J.; Porsti, I.H. Voluntary liquorice ingestion increases blood pressure via increased volume load, elevated peripheral arterial resistance, and decreased aortic compliance. Scientific reports 2017, 7, 10947. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, M.; Imamura, S.; Kawakami, Z.; Ikarashi, Y.; Kase, Y. The blood-brain barrier permeability of 18beta-glycyrrhetinic acid, a major metabolite of glycyrrhizin in Glycyrrhiza root, a constituent of the traditional Japanese medicine yokukansan. Cell Mol Neurobiol 2012, 32, 1139–1146. [Google Scholar] [CrossRef]
- Iigaya, K.; Onimaru, H.; Ikeda, K.; Iizuka, M.; Izumizaki, M. Cellular mechanisms of synchronized rhythmic burst generation in the ventromedial hypothalamus. Pflugers Arch 2025, 477, 131–145. [Google Scholar] [CrossRef]
- Broncel, A.; Bocian, R.; Klos-Wojtczak, P.; Konopacki, J. Hippocampal theta rhythm induced by vagal nerve stimulation: The effect of modulation of electrical coupling. Brain Res Bull 2019, 152, 236–245. [Google Scholar] [CrossRef]
- Schrofelbauer, B.; Raffetseder, J.; Hauner, M.; Wolkerstorfer, A.; Ernst, W.; Szolar, O.H. Glycyrrhizin, the main active compound in liquorice, attenuates pro-inflammatory responses by interfering with membrane-dependent receptor signalling. Biochem J 2009, 421, 473–482. [Google Scholar] [CrossRef]
- Kong, Z.H.; Chen, X.; Hua, H.P.; Liang, L.; Liu, L.J. The Oral Pretreatment of Glycyrrhizin Prevents Surgery-Induced Cognitive Impairment in Aged Mice by Reducing Neuroinflammation and Alzheimer’s-Related Pathology via HMGB1 Inhibition. J Mol Neurosci 2017, 63, 385–395. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, F.; Li, W.; Li, W.; Hu, Y.C.; Li, S.; Zhu, J.H.; Zhou, M.; Hang, C.H. Glycyrrhizic acid confers neuroprotection after subarachnoid hemorrhage via inhibition of high mobility group box-1 protein: a hypothesis for novel therapy of subarachnoid hemorrhage. Med Hypotheses 2013, 81, 681–685. [Google Scholar] [CrossRef]
- Sun, X.; Zeng, H.; Wang, Q.; Yu, Q.; Wu, J.; Feng, Y.; Deng, P.; Zhang, H. Glycyrrhizin ameliorates inflammatory pain by inhibiting microglial activation-mediated inflammatory response via blockage of the HMGB1-TLR4-NF-kB pathway. Exp Cell Res 2018, 369, 112–119. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, Y.; Weng, Z.; Zhou, T.; Feng, T.; Lin, Y. Glycyrrhizin protects brain against ischemia-reperfusion injury in mice through HMGB1-TLR4-IL-17A signaling pathway. Brain Res 2014, 1582, 176–186. [Google Scholar] [CrossRef]
- Akamatsu, H.; Komura, J.; Asada, Y.; Niwa, Y. Mechanism of anti-inflammatory action of glycyrrhizin: effect on neutrophil functions including reactive oxygen species generation. Planta Med 1991, 57, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Wang, F.; Zhu, J.; Wang, S.; Liu, Y.; Xu, J.; Zhao, Q.; Qin, Y.; Si, W.; Zhang, J. 18Beta-Glycyrrhetinic Acid Attenuates H(2)O(2)-Induced Oxidative Damage and Apoptosis in Intestinal Epithelial Cells via Activating the PI3K/Akt Signaling Pathway. Antioxidants (Basel) 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhu, M.M.; Zhang, M.H.; Wang, R.S.; Tan, X.B.; Song, J.; Ding, S.M.; Jia, X.B.; Hu, S.Y. Protection of glycyrrhizic acid against AGEs-induced endothelial dysfunction through inhibiting RAGE/NF-kappaB pathway activation in human umbilical vein endothelial cells. J Ethnopharmacol 2013, 148, 27–36. [Google Scholar] [CrossRef]
- Rizvi, Z.A.; Babele, P.; Sadhu, S.; Madan, U.; Tripathy, M.R.; Goswami, S.; Mani, S.; Kumar, S.; Awasthi, A.; Dikshit, M. Prophylactic treatment of Glycyrrhiza glabra mitigates COVID-19 pathology through inhibition of pro-inflammatory cytokines in the hamster model and NETosis. Front Immunol 2022, 13, 945583. [Google Scholar] [CrossRef]
- Bordbar, N.; Karimi, M.H.; Amirghofran, Z. The effect of glycyrrhizin on maturation and T cell stimulating activity of dendritic cells. Cell Immunol 2012, 280, 44–49. [Google Scholar] [CrossRef]
- Ainsah, O.; Nabishah, B.M.; Osman, C.B.; Khalid, B.A. Short- and long-term effects of glycyrrhizic acid in repetitive stress. Clin Exp Pharmacol Physiol 1999, 26, 444–448. [Google Scholar] [CrossRef]
- Murck, H.; Karailiev, P.; Karailievova, L.; Puhova, A.; Jezova, D. Treatment with Glycyrrhiza glabra Extract Induces Anxiolytic Effects Associated with Reduced Salt Preference and Changes in Barrier Protein Gene Expression. Nutrients 2024, 16. [Google Scholar] [CrossRef]
- Jezova, D.; Karailiev, P.; Karailievova, L.; Puhova, A.; Murck, H. Food Enrichment with Glycyrrhiza glabra Extract Suppresses ACE2 mRNA and Protein Expression in Rats-Possible Implications for COVID-19. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Wu, T.Y.; Liu, L.; Zhang, W.; Zhang, Y.; Liu, Y.Z.; Shen, X.L.; Gong, H.; Yang, Y.Y.; Bi, X.Y.; Jiang, C.L. , et al. High-mobility group box-1 was released actively and involved in LPS induced depressive-like behavior. J Psychiatr Res 2015, 64, 99–106. [Google Scholar] [CrossRef]
- Kamisli, S.; Ciftci, O.; Taslidere, A.; Basak Turkmen, N.; Ozcan, C. The beneficial effects of 18beta-glycyrrhetinic acid on the experimental autoimmune encephalomyelitis (EAE) in C57BL/6 mouse model. Immunopharmacol Immunotoxicol 2018, 40, 344–352. [Google Scholar] [CrossRef]
- Reboldi, A.; Coisne, C.; Baumjohann, D.; Benvenuto, F.; Bottinelli, D.; Lira, S.; Uccelli, A.; Lanzavecchia, A.; Engelhardt, B.; Sallusto, F. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat Immunol 2009, 10, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cai, T.T.; Shen, Y.; Zhou, X.B.; Chen, T.; Xu, Q. Si-Ni-San, a traditional Chinese prescription, and its active ingredient glycyrrhizin ameliorate experimental colitis through regulating cytokine balance. Int Immunopharmacol 2009, 9, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Fang, D.; Li, L.; Chen, L.; Li, Q.; Gong, F.; Fang, M. Glycyrrhizin ameliorates experimental colitis through attenuating interleukin-17-producing T cell responses via regulating antigen-presenting cells. Immunol Res 2017, 65, 666–680. [Google Scholar] [CrossRef]
- Visvanathan, R.; Houghton, M.J.; Williamson, G. Impact of Glucose, Inflammation and Phytochemicals on ACE2, TMPRSS2 and Glucose Transporter Gene Expression in Human Intestinal Cells. Antioxidants (Basel) 2025, 14. [Google Scholar] [CrossRef] [PubMed]
- Barakat, W.; Safwet, N.; El-Maraghy, N.N.; Zakaria, M.N. Candesartan and glycyrrhizin ameliorate ischemic brain damage through downregulation of the TLR signaling cascade. Eur J Pharmacol 2014, 724, 43–50. [Google Scholar] [CrossRef]
- Garg, M.; Singhal, T.; Sharma, H. Cardioprotective effect of ammonium glycyrrhizinate against doxorubicin-induced cardiomyopathy in experimental animals. Indian J Pharmacol 2014, 46, 527–530. [Google Scholar] [CrossRef]
- Liu, F.; Tang, X.; Mao, B.; Zhang, Q.; Zhao, J.; Cui, S.; Chen, W. Ethanol Extract of Licorice Alleviates HFD-Induced Liver Fat Accumulation in Association with Modulation of Gut Microbiota and Intestinal Metabolites in Obesity Mice. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Park, M.; Lee, J.H.; Choi, J.K.; Hong, Y.D.; Bae, I.H.; Lim, K.M.; Park, Y.H.; Ha, H. 18beta-glycyrrhetinic acid attenuates anandamide-induced adiposity and high-fat diet induced obesity. Mol Nutr Food Res 2014, 58, 1436–1446. [Google Scholar] [CrossRef]
- Zhou, L.; Yi, Y.; Lin, B.; Qiu, Z.; Wang, C.; Li, Y. Glycyrrhizic acid mitigates hepatocyte steatosis and inflammation through ACE2 stabilization via dual modulation of AMPK activation and MDM2 inhibition. Eur J Pharmacol 2025, 1002, 177817. [Google Scholar] [CrossRef] [PubMed]
- Seseke, F.G.; Gardemann, A.; Jungermann, K. Signal propagation via gap junctions, a key step in the regulation of liver metabolism by the sympathetic hepatic nerves. FEBS Lett 1992, 301, 265–270. [Google Scholar] [CrossRef]
- Rowe, P.C.; Bou-Holaigah, I.; Kan, J.S.; Calkins, H. Is neurally mediated hypotension an unrecognised cause of chronic fatigue? Lancet 1995, 345, 623–624. [Google Scholar] [CrossRef] [PubMed]
- Otari, K.S.; Shete, R.V.; Bhutada, R.N.; Upasani, C.D. Effect of ammonium glycyrrhizinate on haloperidol- and reserpine- induced neurobehavioral alterations in experimental paradigms. Orient Pharm Exp Med 2011, 11, 153–160. [Google Scholar] [CrossRef]
- Dhingra, D.; Sharma, A. Antidepressant-like activity of Glycyrrhiza glabra L. in mouse models of immobility tests. Prog Neuropsychopharmacol Biol Psychiatry 2006, 30, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Ahmed-Farid, O.A.; Haredy, S.A.; Niazy, R.M.; Linhardt, R.J.; Warda, M. Dose-dependent neuroprotective effect of oriental phyto-derived glycyrrhizin on experimental neuroterminal norepinephrine depletion in a rat brain model. Chem Biol Interact 2019, 308, 279–287. [Google Scholar] [CrossRef]
- Komiyama, M.; Ozaki, Y.; Wada, H.; Yamakage, H.; Satoh-Asahara, N.; Yasoda, A.; Sunagawa, Y.; Morimoto, T.; Tamaki, S.; Masahiro, S. , et al. Randomized double-blind placebo-controlled multicenter trial for the effects of a polyherbal remedy, Yokukansan (YiganSan), in smokers with depressive tendencies. BMC Complement Med Ther 2022, 22, 311. [Google Scholar] [CrossRef]
- Ikarashi, Y.; Mizoguchi, K. Neuropharmacological efficacy of the traditional Japanese Kampo medicine yokukansan and its active ingredients. Pharmacol Ther 2016, 166, 84–95. [Google Scholar] [CrossRef]
- Cao, Z.Y.; Liu, Y.Z.; Li, J.M.; Ruan, Y.M.; Yan, W.J.; Zhong, S.Y.; Zhang, T.; Liu, L.L.; Wu, R.; Wang, B. , et al. Glycyrrhizic acid as an adjunctive treatment for depression through anti-inflammation: A randomized placebo-controlled clinical trial. J Affect Disord 2020, 265, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Murck, H.; Mahmoudi, S.T.; Jezova, D. Inhibition of the 11beta-Hydroxysteroid-Dehydrogenase Type 2 (11betaHSD2) Reduces Aldosterone/Cortisol Ratio and Affects Treatment Outcome Depending on Childhood Trauma History and Inflammation. Biological Psychiatry 2025, 97, S213. [Google Scholar] [CrossRef]
- Huan, C.; Chen, C.; Xu, W.; Guo, T.; Pan, H.; Gao, S. Study on Antiviral Activities of Glycyrrhizin. International Journal of Biomedical Engineering and Clinical Science 2020, 6, 68–70. [Google Scholar] [CrossRef]
- Seckl, J. 11beta-Hydroxysteroid dehydrogenase and the brain: Not (yet) lost in translation. J Intern Med 2024, 295, 20–37. [Google Scholar] [CrossRef]
- Ding, H.; Deng, W.; Ding, L.; Ye, X.; Yin, S.; Huang, W. Glycyrrhetinic acid and its derivatives as potential alternative medicine to relieve symptoms in nonhospitalized COVID-19 patients. J Med Virol 2020. [Google Scholar] [CrossRef]
- Huang, Q.C.; Wang, M.J.; Chen, X.M.; Yu, W.L.; Chu, Y.L.; He, X.H.; Huang, R.Y. Can active components of licorice, glycyrrhizin and glycyrrhetinic acid, lick rheumatoid arthritis? Oncotarget 2016, 7, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



