Submitted:
27 August 2025
Posted:
28 August 2025
You are already at the latest version
Abstract
Scorpion venom has emerged as a promising source of anticancer compounds due to its diverse bioactive components, including neurotoxins, antimicrobial peptides, and enzymes. Proteomic analyses have characterized venom composition in several species, while further functional assays have clarified their anticancer mechanisms. This review synthesizes current knowledge on scorpion venom-derived peptides with demonstrated anticancer activity, which selectively target ion channels, induce apoptosis, or disrupt tumor microenvironments. Where available, we highlight proteomic studies that have identified these components and discuss their structural features relevant to drug design. We also examine clinical applications and the challenges in translating venom peptides into therapies. The crucial and growing role of proteomics in this field, particularly for venom fractionation, component identification, and structural characterization, is critically evaluated.
Keywords:
proteomics
; cancer
; scorpion venom
; natural products
; anticancer peptides
; targeted therapy
; drug delivery
1. Introduction
1.1. The Global Cancer Burden
Cancer remains a leading cause of mortality worldwide, with 20 million new cases and 9.7 million deaths reported in 2022 [1]. Projections suggest these numbers will increase significantly by 2040, with an estimated 29.9 million new cases annually, driven by population growth, aging, and changing exposure patterns to risk factors [1]. Despite significant advances in cancer treatment over the past decades, the need for more effective, targeted, and less toxic therapeutic options continues to drive research in this field.
Cancer is inherently complex, representing a group of related yet distinct diseases, each with unique molecular and cellular features [2]. This heterogeneity presents significant challenges in developing universal treatment approaches and necessitates exploring novel therapeutic strategies. The financial burden of cancer treatment, estimated to exceed $245 billion annually by 2030 in the United States alone [3], further emphasizes the need for innovative and cost-effective treatment options.
1.2. Current Challenges in Cancer Treatment
Despite significant advancements in oncology, traditional cancer therapies continue to face substantial limitations that affect their efficacy and patient outcomes.
Chemotherapeutic agents, while effective against rapidly dividing cancer cells, often lack specificity, leading to collateral damage to healthy tissues. This non-selectivity results in a range of adverse effects, including myelosuppression, which increases infections risk, gastrointestinal toxicity, manifesting as mucositis and diarrhea; cardiotoxicity, particularly with anthracyclines like doxorubicin; neurotoxicity, secondary malignancies, and fertility issues. These side effects can significantly impair a patient's quality of life and may necessitate dose reductions or treatment discontinuation [4].
Drug resistance represents another major challenge in cancer treatment. Tumors may exhibit intrinsic resistance or develop acquired resistance over time. Mechanisms contributing to resistance include enhanced drug efflux via ATP-binding cassette (ABC) transporters, alterations in drug targets, increased DNA repair capabilities, activation of alternative survival pathways, metabolic reprogramming, and modifications within the tumor microenvironment [2,5,6,7,8]. These adaptations often necessitate changes in therapeutic strategies, which may lead to additional complications and diminished patient outcomes.
The blood-brain barrier (BBB) presents a significant challenge in treating brain tumors. Its selective permeability restricts the entry of many therapeutic agents into the central nervous system (CNS). Although tumors can disrupt the BBB, leading to the formation of a heterogeneous blood-tumor barrier (BTB), drug delivery remains inconsistent and often inadequate. This limitation hampers the efficacy of systemic therapies for brain malignancies [9,10,11,12].
The increasing costs of cancer treatments, particularly advanced therapies such as targeted agents and immunotherapies, pose significant barriers to access, especially in low- and middle-income countries. The financial burden extends beyond drug prices, including the need for specialized infrastructure and personnel. These economic challenges contribute to disparities in cancer care and outcomes globally.
1.3. Natural Products in Cancer Therapy
The limitations of conventional cancer treatments have driven renewed interest in natural products as sources of novel bioactive compounds. Historically, natural products have played a crucial role in drug discovery, with approximately 32.5% of approved drugs between 1981 and 2019 being derived from or inspired by natural sources [13], with plant-derived agents, such as taxanes and vinca alkaloids, transforming oncology practice. Animal venoms are an underexplored frontier, providing evolutionarily refined peptides with high specificity for ion channels and receptors, which are often dysregulated in cancer [14]. These venom-derived compounds exhibit distinct pharmacological advantages, including structural complexity enabling selective target engagement, multi-modal mechanisms of action that may circumvent drug resistance, and therapeutic indices often superior to synthetic molecules due to their biological optimization through natural selection [15,16,17,18,19,20].
1.4. Historical Use of Venom in Medicine
The therapeutic application of animal venoms has been documented for thousands of years across various cultures and medical traditions. Ancient Egyptian, Chinese, and Greek medical texts document the use of venoms to treat diverse pathologies, including pain, inflammatory conditions, neurological disorders, cardiovascular diseases, and cancer therapy. This empirical knowledge has provided valuable leads for contemporary drug discovery, yielding several clinically approved venom-derived therapeutics [14,15]. The journey from traditional remedy to approved drug is exemplified by captopril, the first venom-derived drug approved by the FDA in 1981. It was developed from the Brazilian Bothrops jararaca viper´s venom, revolutionizing hypertension treatment as the first angiotensin-converting enzyme inhibitor [21,22]. Similarly, the anticoagulant eptifibatide (Integrilin®), developed from the pygmy rattlesnake (Sistrurus miliarius barbourin) venom [23], the analgesic ziconotide (Prialt®), from the marine cone snail Conus magus [24], and exenatide (Byetta®) derived from Gila monster (Heloderma suspectum) venom for the treatment of type 2 diabetes [25] demonstrate venom´s capacity to address critical therapeutic needs. These successes have catalyzed the investigation of lesser-studied venoms, particularly scorpion toxins, for their unique pharmacologic properties, including anticancer potential.
1.5. The Promise of Scorpion Venom
Scorpion venoms have emerged as a promising source of anticancer compounds due to their distinctive biochemical characteristics. They present high-affinity targeting of ion channels and receptors overexpressed in tumors, along with the ability to cross the blood-brain barrier, enabling CNS tumor targeting. Furthermore, their multi-component compositions act synergistically, while their evolutionarily optimized molecular structures confer selective cytotoxicity against cancer cells.
This review provides a systematic analysis of the proteomic complexity of scorpion venoms, their multimodal mechanisms of anticancer activity, and their potential for translational applications in oncology.
2. Composition and Biochemistry of Scorpion Venom
Scorpion venom represents a rich reservoir of bioactive compounds, comprising diverse proteins, peptides, enzymes, nucleotides, lipids, inorganic salts, free amino acids, and small molecules, demonstrating remarkable specificity in targeting cellular pathways [26,27]. Among these, proteins and peptides are the most pharmacologically significant, particularly for anticancer applications due to their high target selectivity and low off-target toxicity [28,29].
2.1. Proteomic Characterization of Scorpion Venom
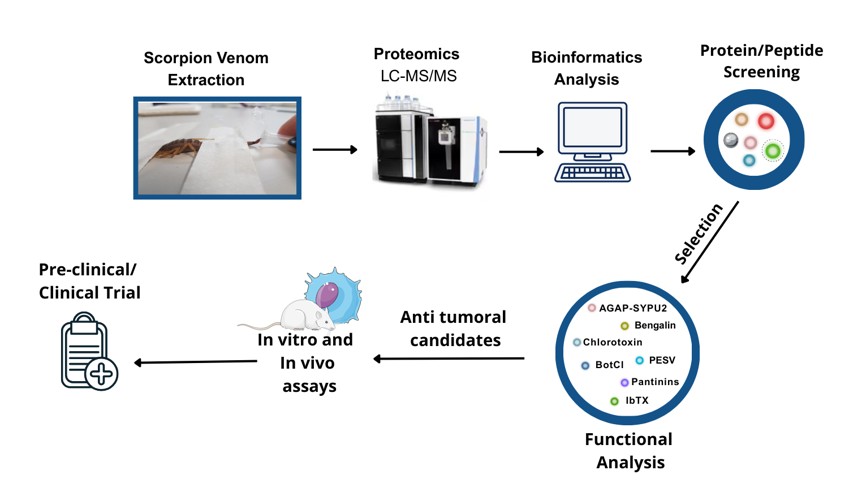
Recent advances in high-throughput proteomics have revolutionized the characterization of scorpion venom, enabling comprehensive profiling of its complex molecular composition. State-of-the-art analytical techniques, such as nano-scale liquid chromatography coupled with tandem mass spectrometry (LC-MS/MS), combined with advanced bioinformatics pipelines, have enabled researchers to identify and quantify hundreds of unique proteins, peptides, and bioactive molecules within venom samples [30,31] (Table 1).
The key steps in venom proteomics typically begin with venom fractionation through chromatographic methods, such as high-performance liquid chromatography (HPLC) or size exclusion chromatography (SEC), followed by high-resolution mass spectrometric analysis for peptide sequencing and structural elucidation. Database mining using UniProt and VenomZone, together with bioinformatics analysis, is then employed to identify venom components and facilitate the discovery of previously unknown bioactive peptides with therapeutic potential, including anticancer properties [80]. This workflow is summarized in Figure 1.
Proteomic analyses across different species have provided insights into venom diversity and revealed evolutionary patterns in venom biochemistry, highlighting conserved elements that may serve as valuable leads for drug development [81,82]. This natural diversity in venom composition constitutes a vast molecular natural library of bioactive compounds, enabling targeted exploration for therapeutic applications.
Quantitative proteomics, including label-free, iTRAQ, and tandem mass tag (TMT)-based approaches, has further enhanced our understanding of venom composition by revealing the relative abundance of different constituents and their variability across species and environmental conditions [83]. These methods have also elucidated critical post-translational modifications (PTMs), such as disulfide bond formation, phosphorylation, and N-glycosylation in neurotoxins, which significantly influence protein stability and biological activity [84,85,86]. The integration of top-down (intact protein analysis) and bottom-up/shotgun (digested peptide analysis) proteomic strategies provide complementary insights into venom protein structure and function, facilitating the identification of promising therapeutic candidates.
Together, these advances not only deepen our understanding of venom composition but also accelerate the identification of bioactive candidates with anticancer potential, underscoring the importance of proteomics in drug development.
2.2. Major Protein and Peptide Components of Scorpion Venom
The protein and peptide components of scorpion venom represent a complex mixture of bioactive molecules that constitute the molecular basis for its therapeutic potential. These components can be classified into several major groups based on their distinct structural and functional characteristics.
Neurotoxins represent one of the most abundant and well-studied groups, comprising both long-chain toxins that modulate voltage-gated sodium channels and short-chain toxins that target potassium and chloride channels [29,87,88,89,90,91,92,93,94,95,96,97]. These neurotoxins have shown promise in cancer therapy due to their ability to modulate ion channels that are crucial for cancer cell proliferation and migration, and are often dysregulated in cancer cells [98,99]. These interactions can disrupt cancer cell membrane potential, ultimately triggering apoptotic pathways. Of particular interest are certain short-chain peptides that exhibit the ability to cross the blood-brain barrier and specifically bind to cancer cell membranes, making them valuable candidates for treating neurological malignancies [27,100,101,102].
In addition to neurotoxins, scorpion venom contains antimicrobial peptides that demonstrate not only antimicrobial activity [103] but also exhibit potential anticancer properties through their membrane-disrupting mechanisms [70,104,105,106]. These peptides often exhibit selective cytotoxicity against cancer cells while showing minimal effects on normal cells, a characteristic that significantly enhances their therapeutic potential [74].
The venom also contains various enzyme inhibitors, particularly those targeting proteases that play crucial roles in cancer progression and metastasis [75,107,108,109,110,111,112]. Phospholipases represent another major component, capable of modifying cell membrane composition and triggering diverse cellular responses [99,113,114,115,116,117,118,119].
The disulfide-rich peptides are of pharmacological interest due to their remarkable stability conferred by multiple disulfide bonds and their high specificity for cellular targets, making them excellent candidates for drug development [84,120,121,122]. Recent research studies have identified novel peptides with unique mechanisms of action against cancer cells, including those that modulate the expression of tumor suppressor p53 or interfere with the PI3K/Akt signaling pathway, which is frequently dysregulated in cancer [28,123,124,125]. Additionally, the venom contains various bioactive amines and polyamines that can influence cellular signaling pathways, often synergizing with other venom components to enhance their cytotoxic effects [126,127,128,129,130].
Advances in proteomics and structural biology have enabled the optimization of these venom components for therapeutic applications. Strategies such as peptide cyclization and PEGylation have improved molecular stability, while conjugation techniques have enhanced tumor targeting specificity [16,19,131]. The multi-pathway inhibitory effects of certain venom components offer promise for overcoming drug resistance in cancer treatment [125].
The enzymatic components of scorpion venom represent a sophisticated array of biological catalysts that contribute significantly to its therapeutic potential [83]. These enzyme systems comprise diverse protein classes with distinct catalytic activities, including phospholipases, hyaluronidases, metalloproteases, and serine proteases [26,132,133]. Phospholipases, particularly PLA2, play a crucial role in membrane disruption and can trigger various cellular responses relevant to cancer therapy [134]. Hyaluronidases, often referred to as “spreading factors”, facilitate the distribution of other venom components by breaking down hyaluronic acid in the extracellular matrix (ECM) [135], a property that could be particularly advantageous in designing targeted drug delivery systems for cancer treatment. Metalloproteases present in scorpion venom can modulate the ECM, thereby affecting critical processes in cancer metastasis such as cell adhesion and migration [135]. The presence of serine proteases adds further complexity to the venom's enzymatic profile, given their ability to influence blood coagulation and inflammatory responses [134]. The coordinated action of these enzyme systems creates a complex network of biochemical reactions that can be potentially harnessed for therapeutic applications. A comprehensive understanding of their kinetics and regulatory mechanisms is essential for developing effective cancer treatments, as many of these enzymes exhibit substrate specificity or target cellular pathways implicated in cancer biology [136].
3. Molecular Mechanisms of Anticancer Activity
Scorpion venom components exert anticancer effects through diverse molecular mechanisms, including induction of apoptosis, cell cycle regulation, and modulation of ion channels. Rather than acting as non-specific cytotoxins, many peptides demonstrate selectivity for malignant cells by targeting signaling pathways, surface receptors, or membrane properties that are dysregulated in cancer. The following subsections provide an overview of the principal mechanisms, illustrated by representative venom-derived molecules (Figure 2). Detailed descriptions of individual peptides are provided in Section 4.
3.1. Induction of Apoptosis
The investigation of cell death pathways induced by scorpion venom components has revealed complex molecular mechanisms of action that selectively induce cancer cell death through both intrinsic and extrinsic apoptotic pathways. The intrinsic apoptotic pathway activation involves critical disruption of mitochondrial function, as demonstrated by Smp24, a peptide derived from Scorpio maurus palmatus. This peptide disrupts mitochondrial homeostasis by reducing the mitochondrial membrane potential (ΔΨm), increasing reactive oxygen species (ROS) production, and inducing cytoskeletal reorganization in HepG2 cancer cells while exhibiting minimal toxicity toward normal LO2 liver cells [73,74]. Mitochondrial depolarization and ROS generation are hallmarks of caspase-3-mediated disruption of electron transport chain complexes I and II, a feedback mechanism that amplifies apoptotic signaling after cytochrome c release [32,137]. In addition, both Smp24 and Smp43 activate caspase-1, triggering pyroptosis in both myeloid (KG1-a) and lymphoid (CCRF-CEM) leukemia cells, again with minimal toxicity to non-tumor HaCaT cells [70].
Bengalin from Heterometrus bengalensis Koch exemplifies mitochondrial-targeting peptides, inducing cytochrome c release and caspase activation in leukemic cells [39]. Detailed proteomic and structural insights are discussed in Section 4.1.2. A similar mechanism is seen with peptide BmKn-2 from Mesobuthus martensii Karsch, which induces apoptosis in canine mammary gland tumor CHMp-5b and CHMPp-13a cell lines and in both human oral squamous carcinoma cells (HSC4) and human mouth epidermoid carcinoma cells (KB) via Bax/ Bcl-2 modulation and caspase 9 activation, while sparing normal human gingival and dental pulp cells [45,46]. Consistently, Hemiscorpius lepturus venom promotes Bax, caspase 3, and p53 overexpression alongside Bcl2 suppression in both CT26 colon carcinoma cells and xenograft tumors, with low cytotoxicity toward non-tumorigenic VERO cells [55].
Venoms also act on oncogenic signaling. Buthus martensii Karsch venom (BmK) selectively kills Raji and Jurkat lymphoma cells while sparing normal human peripheral blood lymphocytes. In Raji cells, BmK upregulated the tumor suppressor PTEN expression, decreasing Akt and Bad phosphorylation, thereby downregulating PI3K/Akt signaling. In PTEN-negative Jurkat cells, apoptosis proceeds through p27-mediated mechanisms, underscoring cell-specific vulnerabilities. Combining BmK with the Akt inhibitor LY294002 synergistically enhanced apoptosis, underscoring the therapeutic potential of venom-derived compounds in targeting oncogenic pathways [43].
The extrinsic apoptotic pathway is mediated through interactions with death receptors on cancer cell surfaces. Neopladines 1 and 2 from Tityus discrepans venom upregulate Fas ligand in SKBR3 breast cancer cells, promoting death-inducing signaling complex (DISC) formation, caspase-8 activation, and selective killing relative to non-malignant MA104 cells [67]. Similar receptor-driven effects have been reported in Androctonus crassicauda venom-treated HCT-8 colorectal cancer cells [28,32]. Activated caspase-8 also cleaves the BH3-only protein Bid, generating truncated Bid (tBid) that translocates to mitochondria, linking extrinsic and intrinsic pathways, thereby amplifying mitochondrial permeabilization and caspase-3 activation [28,32,138,139,140,141,142].
Taken together, these findings show that scorpion venoms exploit multiple cell death pathways, with selectivity for cancer cells driven by mitochondrial dysfunction, death receptor activation, and pathway cross-talk. Their multi-target nature, simultaneously inhibiting proliferation, angiogenesis, and metastasis, positions them as promising candidates for combinatorial anticancer therapies [44,143].
3.2. Cellular Signaling and Cycle Disruption
The impact of scorpion venom on cellular signal transduction pathways has been mapped through comprehensive phosphoproteomic analyses, revealing intricate networks of molecular interactions. Key targets include MAP kinases, PI3K/Akt, and JAK/STAT pathways, with venom peptides selectively inhibiting growth factor receptor signaling, disrupting crucial survival pathways in cancer cells. For example, Buthus martensii Karsch venom (BmK) upregulates PTEN in Raji lymphoma cells, suppressing PI3K/Akt via reduced Akt/Bad phosphorylation, while p27 mediates death in PTEN-negative Jurkat cells [43]. Temporal proteomics reveals sequential pathway activation, while protein-protein interaction studies highlight venom-induced disruption of cross-talk between survival pathways, potentially circumventing drug resistance.
Proteomics also clarifies cell cycle interference, identifying venom interactions with cyclins, CDKs, and checkpoint regulators. High-throughput screens show venom peptides arresting the cell cycle at G1/S or G2/M transitions through post-translational modifications, such as phosphorylation of regulatory proteins. Quantitative proteomics demonstrates differential expression of cycle-related proteins in cancer versus normal cells, supporting selective toxicity. These insights have propelled targeted strategies to halt cancer proliferation.
4. Specific Anticancer Components in Scorpion Venom
4.1. Apoptosis induction
4.1.1. TsAP-1 and TsAP-2 from Tityus Serrulatus
The 17-mer peptides TsAP-1 and TsAP-2, isolated from the venom of the Brazilian yellow scorpion Tityus serrulatus, demonstrate significant anticancer potential through selective cytotoxicity and apoptosis induction (see Section 3.1). While TsAP-1 exhibits modest activity against oral carcinoma (H157 cells, IC₅₀ >50 μM), its cationic analog TsAP-S1 shows dramatically enhanced potency (IC₅₀ = 2.5 μM in leukemia cells) through membrane disruption mechanisms [77]. TsAP-2 displays greater inherent activity, with an IC₅₀ of 4 μM in SKBR3 breast cancer cells, and its engineered analog TsAP-S2 achieves remarkable potency (IC₅₀ = 0.83 μM) while maintaining selectivity for cancer cells over normal cells. These peptides induce apoptosis through mitochondrial disruption and activation of caspase pathways, as evidenced by their ability to suppress proliferation in multiple cancer cell lines, including glioblastoma (U251-MG) and prostate adenocarcinoma (PC-3), with minimal effects on non-malignant cells [77]. Further supporting their therapeutic potential, TsAP-2 and the structurally related peptide Stigmurin from Tityus stigmurus exhibit antiproliferative effects on tumor cells while demonstrating low cytotoxicity toward normal cells, highlighting their selectivity and safety profile [78]. The enhanced activity of cationicity-modified analogs (TsAP-S1 and TsAP-S2) underscores the importance of structural optimization for improving anticancer efficacy. These findings position TsAP-1 and TsAP-2 as promising candidates for targeted cancer therapy, with their mechanisms of action rooted in membrane interaction and apoptotic pathway activation. Further research is needed to elucidate their precise molecular targets and evaluate their efficacy in vivo.
4.1.2. Bengalin from Heterometrus Bengalensis and Other Novel Peptides
Bengalin, a peptide isolated from the Indian black scorpion Heterometrus bengalensis, exemplifies how structural and proteomic analyses deepen our understanding of venom-derived anticancer compounds. Proteomic profiling has confirmed its abundance in crude venom and provided evidence of its selective enrichment in venom fractions with cytotoxic activity [144]. Structurally, Bengalin exhibits a disulfide-stabilized fold characteristic of scorpion toxins, conferring conformational stability and resistance to proteolytic degradation, properties that enhance its therapeutic potential [41]. Beyond its structural resilience, proteomic and immunoblotting studies have documented Bengalin´s dual impact on cancer cells. It induces apoptotic signaling, evidenced by Bax/Bcl-2 modulation, cytochrome c release, and caspase cascade activation, while simultaneously engaging autophagic responses through Beclin-1 and Atg upregulation and LC3 conversion [38,39].
These molecular features correlate with selective cytotoxicity by inducing apoptosis in U937 and K562 leukemic cells through multiple pathways mediated by the increase of caspase-3 activity and mitochondrial disruption pathways [39,40] and autophagic cell death via inhibition of proliferative MAPK/ERK and PI3K/AKT pathways [38,39]. Although Bengalin has shown selective cytotoxic potential for leukemic cells [144], more studies are needed to investigate the exact mechanisms of action of this compound and perform in vivo assays to confirm its anti-tumorigenic activity.
4.1.3. Neopladine 1 and Neopladine 2 from Tityus Discrepans
Neopladine 1 and 2, isolated from Tityus discrepans venom, represent novel anticancer compounds with unique mechanisms of action. These peptides exhibit selective anticancer activity against HER2-positive human breast carcinoma SKBR3 cells, inducing apoptosis while showing negligible effects on non-malignant MA104 monkey kidney cells [42,67,128]. Mass spectrometry analysis determined their molecular masses to be 29,918 Da (Neopladine 1) and 30,388 Da (Neopladine 2), and their N-terminal sequences were confirmed by Edman degradation. The peptides induce apoptosis in SKBR3 cells, with Neopladine 1 causing 6.3% apoptosis and Neopladine 2 causing 4.1% apoptosis after 5 hours of exposure; prolonged exposure increases this effect. Immunohistochemical studies indicate that neopladines bind to SKBR3 cell surfaces, upregulating FasL and Bcl-2 expression, which are critical in apoptosis signaling. Interestingly enough, the combined application of neopladines 1 and 2 reduced apoptosis but increased necrosis, suggesting complex interactions that require additional investigation [67].
4.2. Ion Channel Modulation
4.2.1. AGAP-SYPU2 from Buthus Martensii Karsch
Analgesic Anti-tumor Peptide AGAP-SYPU2, a peptide isolated from Buthus martensii Karsch (BmK) scorpion venom, exhibits dual analgesic and antitumor activities [35,145]. Shao et al. purified AGAP-SYPU2 and demonstrated its strong analgesic effects against both visceral and somatic pain, with its mechanism involving inhibition of voltage-gated sodium channels, which are critical in pain signaling. Although its onset of action is slower than morphine, it provides prolonged analgesic effects [36].
In cancer therapy, AGAP (a homolog of AGAP-SYPU2) shows promising antitumor properties. It prolonged survival by 36.05% in Ehrlich ascites tumor models and reduced tumor weight by 46.3% in S180 fibrosarcoma mouse models [36]. The peptide's anticancer activity is linked to its modulation of sodium channels such as Nav1.4, Nav1.5, and Nav1.7, which are overexpressed in cancer cells and contribute to proliferation and migration. Moreover, AGAP was able to inhibit the proliferation and migration of SHG-44 glioma cells, suggesting a shared mechanism involving sodium channel blockade [37]. The dual functionality of AGAP-SYPU2—both as an analgesic and antitumor agent—makes it clinically valuable, as it may improve patient survival without compromising quality of life. However, further research is needed to optimize its potency and evaluate its efficacy in other cancer types.
4.2.2. BotCl from Buthus Occitanus Tunetanus
BotCl, a chlorotoxin-like peptide isolated from the venom of the scorpion Buthus occitanus tunetanus, has emerged as a promising anticancer agent due to its ability to target ClC-3 chloride channels, which are overexpressed in glioblastoma and breast cancer cells [47,48]. This peptide belongs to the chlorotoxin family, characterized by four disulfide bonds that confer structural stability and high binding affinity to tumor-specific ion channels and membrane receptors, such as matrix metalloproteinase-2 (MMP-2) [47,49]. BotCl shares significant sequence homology with chlorotoxin (CTX), a well-studied scorpion venom peptide currently in clinical trials for glioma imaging and therapy, suggesting similar mechanisms of action, including inhibition of tumor cell migration and invasion [47,49].
The anticancer properties of BotCl extend beyond glioblastoma. Its ability to reduce tumor viability in breast cancer models highlights its broad-spectrum potential, possibly through mechanisms involving chloride channel blockade and disruption of tumor microenvironment signaling [47,48]. Furthermore, BotCl’s structural stability under varying pH and temperature conditions enhances its suitability for therapeutic development, including conjugation with imaging agents or cytotoxic drugs for targeted cancer therapy [48,146].
In summary, BotCl represents a novel chlorotoxin-like peptide with significant anticancer potential, supported by proteomic and structural analyses. Its ability to target ClC-3 channels and MMP-2 in glioblastoma and breast cancer positions it as a promising candidate for further preclinical and clinical evaluation, particularly in the development of tumor-specific diagnostic and therapeutic agents.
4.2.3. Iberiotoxin (IbTX) from Hottentotta Tamulus
Iberiotoxin (IbTX), a 37-amino acid peptide derived from the venom of the scorpion Hottentotta tamulus (also known as Buthus tamulus or Mesobuthus tamulus), has emerged as a potent inhibitor of voltage-gated potassium channels, particularly Kv1.1 and Kv1.3, which are overexpressed in malignancies such as glioblastoma (U87), breast (MDA-MB-231), colon (LS174), cervical (HeLa), and ovarian (A2780) cancers [57,58,59]. This toxin shares 68% sequence homology with charybdotoxin (ChTX) but exhibits superior selectivity for large-conductance calcium-activated potassium (BKCa) channels, with an equilibrium dissociation constant (Kd) of 1.16 nM, making it one of the most potent blockers of this channel family [57,60,61,147].
IbTX's anticancer potential extends beyond direct channel blockade. In breast cancer models (MCF-7, MDA-MB-231), IbTX-sensitive currents were shown to modulate proliferation under conditions of elevated intracellular calcium, such as ATP stimulation [58]. This suggests a context-dependent role in tumor signaling, where IbTX could synergize with calcium-mobilizing therapies. Additionally, structural studies using synthetic chimeric peptides, such as IbTX-ChTX hybrids revealed that the toxin's C-terminal domain is critical for Kv1.1/1.3 selectivity, providing a template for designing targeted anticancer derivatives [148].
Despite its promise, challenges remain in translating IbTX into clinical applications due to its potential off-target effects on neuronal and cardiovascular Kv channels. However, advancements in bioconjugation—such as biotinylated IbTX for imaging BKCa distribution in cancer cells—highlight its utility as a molecular tool for tumor profiling and drug development [149]. Future research should explore engineered analogs with enhanced tumor specificity and delivery systems to harness IbTX's full therapeutic potential.
In summary, IbTX represents a structurally and functionally characterized scorpion venom peptide with validated anticancer activity, supported by proteomic and electrophysiological evidence. Its ability to target oncogenic potassium channels positions it as a promising candidate for further preclinical evaluation in calcium-driven malignancies [57,58,60].
4.3. Cell Cycle Arrest
4.3.1. Gonearrestide from Androctonus Mauritanicus
Gonearrestide, an 18-amino acid peptide (2.2 kDa) isolated from the venom of Androctonus mauritanicus, represents a novel class of scorpion venom-derived compounds with potent anticancer activity. Identified through a high-throughput platform combining next-generation sequencing (NGS) transcriptomics and LC-MS/MS proteomics, this peptide was selected from 238 novel peptides discovered in scorpion venom libraries due to its selective cytotoxicity against cancer cells while sparing normal epithelial cells and erythrocytes [54].
The peptide's mechanism of action centers on inducing G1 cell cycle arrest in colorectal cancer cells (HCT116), achieved through dual modulation of cyclin-dependent kinase (CDK) regulators. Gonearrestide downregulates CDK4, a key driver of G1/S transition, while simultaneously upregulating the CDK inhibitors p21 and p27, as well as cyclin D3 [54]. This coordinated action effectively halts cancer cell proliferation, as demonstrated by RNA sequencing showing significant alterations in cell cycle-related gene expression profiles.
In preclinical validation, Gonearrestide exhibited broad-spectrum activity against multiple cancer cell lines while showing negligible toxicity to normal cells, a critical advantage over conventional chemotherapeutics. Its ability to inhibit primary colon cancer cells and solid tumors in vivo further underscores its therapeutic potential. The discovery of Gonearrestide exemplifies how integrated omics technologies (proteomics/transcriptomics) can accelerate the identification of bioactive venom peptides with precise mechanisms of action. Its cell cycle-specific targeting offers a template for developing novel anticancer agents that combine high potency with reduced off-target effects.
4.3.2. PESV from Buthus Martensii Karsch
Mass spectrometry and molecular biology analyses of the Buthus martensii Karsch (BmK) venom have identified multiple bioactive components, including polypeptides with anticancer activity [121,150]. Comprising 50-60 amino acid peptides, the Polypeptide Extract from Scorpion Venom (PESV) interferes with tumor growth through both anti-proliferative and pro-apoptotic mechanisms. In preclinical models, it inhibited the angiogenesis and suppressed tumor growth of H22 hepatocellular carcinoma in murine models [151].
In human androgen-independent DU145 prostate cancer cells, PESV induces cell cycle arrest at the G1-phase by upregulating Kip1/p27 expression while downregulating cyclin E, thereby disrupting cyclin-dependent kinase (CDK) activity, a key driver of cancer progression, which is normally regulated by CDK inhibitors (CDKIs) [152,153]. This cell-cycle blockade is accompanied by apoptosis mediated through upregulation of the proapoptotic protein Bax and downregulation of the anti-apoptotic protein Bcl-2, highlighting PESV´s dual regulatory impact on proliferation and survival pathways. Importantly, PESV demonstrates preferential cytotoxicity toward prostate cancer cells over normal prostate epithelial cells, highlighting its potential as a targeted therapeutic agent [152].
Although these findings support PESV as a promising candidate, its precise molecular targets and the identity of the active peptide(s) within the extract remain unresolved. Further studies across different prostate cancer models are needed to validate its efficacy and clarify the signaling cascades involved [152].
4.4. Membrane Disruption and Tumor Microenvironment
4.4.1. Hyaluronidase BmHYA1 from Buthus Martensii Karsch
The hyaluronidase BmHYA1, isolated from the venom of the scorpion Buthus martensi Karsch (BmK), has emerged as a promising anticancer agent due to its ability to modulate the tumor microenvironment without observed toxic side effects [42,154,155,156,157,158]. This enzyme specifically targets hyaluronic acid (hyaluronan, HA), a key glycosaminoglycan polymer in the extracellular matrix (ECM), which promotes tumor progression by facilitating cancer cell migration, invasion, adhesion, and metastasis through interactions with CD44 receptors [156]. Elevated HA levels are associated with malignant phenotypes, as it provide a structural scaffold for tumor survival and activate signaling pathways via CD44, which is often overexpressed in cancer cells and generates oncogenic variants, such as CD44v6, through alternative splicing [159,160].
BmHYA1 exerts its anticancer effects by degrading HA, thereby disrupting HA-CD44 interactions, and sensitizing cancer cells to apoptosis by disrupting CD44-mediated survival signals, including TGF-β activation and resistance to immune cytotoxicity [156]. This degradation reduces interstitial fluid pressure in tumors, enhancing the penetration of chemotherapeutic agents and improving drug delivery. In addition, BmK hyaluronidase was completely removed from triple-negative MDA-MB-231 breast cancer cells, downregulated CD44v6 expression, and inhibited HA-mediated oncogenic signaling [156]. The enzyme’s specificity for HA makes it particularly effective against HA-rich cancers, such as breast and pancreatic carcinomas. Its dual role—as a standalone antitumor agent and a facilitator of combination therapies—highlights its therapeutic potential. However, further research is needed to fully elucidate its mechanisms and optimize its clinical application.
4.4.2. RK1 from Buthus Occitanus Tunetanus
RK1, a 14-amino acid peptide isolated from the venom of the Tunisian scorpion Buthus occitanus tunetanus, represents a promising anticancer peptide with distinctive pharmacological properties. Biochemical and functional characterization has revealed that RK1 exhibits potent antitumor activity by simultaneously inhibiting cancer cell proliferation, migration, and angiogenesis without manifesting significant cytotoxicity toward normal cells, making it potentially effective against metastatic cancer. Moreover, RK1 demonstrated remarkable efficacy against glioblastoma (U87) and melanoma (IGR39) cell lines, with its mechanism of action involving the disruption of tumor cell adhesion and suppression of vascular growth, as evidenced by the chicken chorioallantoic membrane (CAM) assay [161].
Emerging evidence also indicates that RK1 may interfere with integrin-mediated pathways, which are critical for tumor cell adhesion, metastasis, and angiogenesis. This dual disintegrin-like activity, particularly on α1β1 and αvβ3 integrins, further expands its therapeutic potential by targeting the tumor microenvironment [161]. Given these multifaceted mechanisms, RK1 could serve as a foundational scaffold for developing novel anticancer therapeutics, either as a standalone agent or in combination with existing treatment modalities.
4.4.3. Vmct1 from Vaejovis Mexicanus
Vmct1 is a 13-residue non-disulfide-bridged cationic peptide (NDBP) originally identified from the venom gland transcriptome of the scorpion Vaejovis mexicanus. With a net charge of +2, ~69% hydrophobicity, and an amphipathic α-helical conformation, the native peptide exhibits antimicrobial activity but lacks significant anticancer effects. However, synthetic analogs of Vmct1 with lysine substitutions (Vmct1-K) were designed to enhance cationicity and bioactivity, resulting in potent cytotoxicity against melanoma (B16-F10), breast (MCF-7), and cervical (HeLa) cancer cell lines (IC₅₀: 3.4 - 6.2 µM), with minimal toxicity to non-tumoral VERO and red blood cells [34]. Mechanistic studies suggest a membrane-lytic mode of action. SEM images of Vmct1-K-treated cells revealed blebbing, wrinkling, and cell shrinkage, while increased propidium iodide uptake confirmed membrane disruption [34]. Together, these findings highlight Vmct1-K as a rationally optimized peptide with selective anticancer potential via direct membrane targeting.
4.4.4. AcrAP1/AcrAP2 from Androctonus Crassicauda
AcrAP1 and AcrAP2 are non-disulfide bridged peptides (NDBPs) isolated from the venom of the Arabian scorpion Androctonus crassicauda, which exhibit selective antimicrobial activity against Staphylococcus aureus and Candida albicans but exhibit no cytotoxic or antiproliferative effects on a panel of different human cancer cell lines. However, cationicity-enhanced analogues AcrAP1a and AcrAP2a, engineered by substituting polar residues with lysine, displayed significantly improved biological activity. These analogues exhibited potent antiproliferative effects against multiple human cancer cell lines, including lung adenocarcinoma (NCI-H460), breast carcinoma (MCF-7 and MDA-MB-435s), and prostate carcinoma (PC-3), with IC₅₀ values ranging from 2.1 to 3.6 µM. Interestingly enough, AcrAP1a also induced a paradoxical proliferative response in H460 and PC-3 cells at nanomolar concentrations, suggesting potential concentration-dependent dual effects that warrant further mechanistic investigation [33]. The enhanced anticancer activity of the analogues appears to correlate with their amphipathic α-helical structures and increased net positive charge, promoting interactions with negatively charged components of cancer cell membranes. While direct evidence of membrane lysis was not provided, the observed cytotoxic effects are consistent with mechanisms reported for other cationic antimicrobial peptides. The rational design of AcrAP analogs achieved by substituting neutral residues with lysine, like Vmct1-K demonstrates how structural optimization can amplify anticancer effects while minimizing off-target toxicity [34]. In summary, these findings highlight the potential of venom-derived peptide templates for the rational design of anticancer agents. The structural optimization of AcrAP1/AcrAP2 into cationic analogues demonstrates a viable strategy to enhance bioactivity while maintaining selective toxicity, although concentration-dependent paradoxical effects underscore the need for thorough preclinical evaluation.
4.4.5. Pantinins 1-3 from Pandinus Imperator
Pantinins 1-3 are a family of short, cysteine-free, α-helical cationic peptides identified via cDNA cloning from the African scorpion Pandinus imperator venom gland transcripts. Their amphipathic helices underlie broad-spectrum antimicrobial activity, including potent effects against multidrug-resistant bacteria such as vancomycin-resistant Enterococcus strains (VRE) [68], which provides a rationale to explore their repurposing in oncology. Beyond their antibacterial profile, synthetic pantinin analogs have been shown to interact selectively with negatively charged membranes, producing membranolytic activity against human cancer cell lines while sparing erythrocytes, thus demonstrating low hemolytic potential [69].
This mode of action aligns with a growing class of anticancer antimicrobial peptides (AMPs) that exploit electrostatic differences in membrane lipid composition between malignant and healthy cells. While pantinins are α-helical, their mechanism is conceptually parallel to that of engineered β-hairpin peptides such as SVS-1, which undergo membrane-induced folding into a cytotoxic conformation upon contact with tumor membranes [162]. Such comparisons highlight a broader principle: secondary structure plasticity, driven by the tumor microenvironment, underpins the selective cytotoxicity of many AMPs.
Recent reviews have highlighted the growing interest in antimicrobial peptides (AMPs) as dual-purpose agents with antimicrobial and anticancer potential [163]. Within this broader context, pantinins exemplify scorpion-derived AMPs that combine small size, a favorable safety profile, and well-defined helical structure, making them attractive scaffolds for further optimization. Strategies such as enhancing cationicity or conjugating to delivery systems may improve their tumor specificity and therapeutic index.
4.5. Multifunctional Peptides
4.5.1. Chlorotoxin from Leiurus Hebraeus and Derivatives
Chlorotoxin (CTX) is a 36-amino acid peptide originally isolated and purified from the venom of Leiurus hebraeus (formerly L. quinquestriatus hebraeus) [164]. Structural analysis by NMR revealed a compact tertiary structure comprising an α-helix packed against three antiparallel β-strands stabilized by four disulfide bonds, features that underlie its remarkable stability and high affinity for tumor cell membranes [165,166]. CTX selectively binds to gliomas and other neuroectodermal tumors, with negligible affinity for normal tissues [49,50,167,168], making it a valuable scaffold for tumor targeting.
Proteomics and mass spectrometry studies have been critical for defining CTX´s molecular interactors. Affinity-column pull-downs combined with mass spectrometry further demonstrated that matrix metalloproteinase-2 (MMP-2) as a primary binding partner of CTX and revealed its association with a membrane complex containing MT1-MMP, TIMP-2, and αvβ3 integrin, implicating CTX in the modulation of pericellular proteolysis and adhesion [49,169]. These proteomic analyses reinforced earlier electrophysiology-based findings that CTX blocks glioma-enriched ClC-3 chlorine channels involved in cell migration and cytoskeletal remodeling [170,171]. More recently, protein microarray profiling coupled with mass spectrometry validation identified cortactin, an actin-binding and Src kinase substrate linked to invasive cancers, as a novel CTX interactor, suggesting an additional mechanism for its anti-migratory effects and making it an important biomarker for invasive cancers [172]. Such mass spectrometry-based proteomics-driven discoveries have expanded the CTX interactome beyond MMP-2, ClC-3, and annexin A2, highlighting neuropilin-1 (NRP1) as another critical binding partner for tumor selectivity [173,174].
Recent proteomic surveys of Buthidae venoms have further contextualized chlorotoxin within the venom proteome. Using LC-MS/MS, Mabunda et al. (2025) identified chlorotoxin and CTX-like peptides as major cysteine-rich peptide families in Leiurus hebraeus venom. Importantly, integrative functional assays linked these CTX-containing fractions to cytotoxic and anti-migratory effects in glioma and melanoma cells, thereby bridging venom proteomics with cancer pharmacology. These findings reinforce the evolutionary conservation and therapeutic potential of CTX and highlight the value of advanced proteomics in connecting peptide abundance with functional anticancer activity [175].
Functional proteomic fragment mapping has shown that the C-terminal residues (29–36) of CTX retain partial activity, selectively inhibiting migration without fully blocking invasion [172]. This finding suggests that distinct CTX structural motifs may differentially regulate cytoskeletal versus proteolytic pathways. Internalization studies using live-cell imaging confirmed clathrin-mediated uptake of CTX derivatives [176], linking receptor binding with intracellular trafficking.
Building on these proteomics-driven insights, chlorotoxin has recently been incorporated into the design of chimeric antigen receptor (CAR) T cells. CLTX-CAR T cells employ the chlorotoxin peptide as a tumor recognition domain, redirecting T cell cytotoxicity against glioblastoma cells through MMP-2–associated binding. In a first-in-human Phase I clinical trial (NCT04214392), Barish et al. (2025) demonstrated the feasibility and safety of intracavitary administration of CLTX-CAR T cells in patients with recurrent glioblastoma. No dose-limiting toxicities or immunogenicity were observed, and three of four patients achieved transient stable disease with evidence of CAR T cell persistence and local cytokine induction in the tumor cavity. Although long-term clinical benefit remains to be established, this study represents the first clinical application of a venom-derived peptide as the targeting domain of a cellular immunotherapy and highlights the translational potential of CTX as a proteomics-validated tumor ligand [177].
From a translational perspective, synthetic CTX derivatives have been developed, notably TM-601, radiolabeled with iodine-131 (131 I-chlorotoxin), which demonstrated tumor-specific uptake and prolonged retention in Phase I/II clinical trials in malignant glioma [178,179,180]. Beyond therapy, proteomics-guided bioconjugates have supported CTX applications in diagnostics and imaging. BLZ-100 (Tumor Paint), a fluorescent CTX conjugate, enables intraoperative visualization of gliomas [181]. Nanotechnology-based delivery systems, such as CTX/mApoE-modified liposomes or MiniCTX shuttles, further enhance blood-brain barrier penetration and selective tumor targeting [182,183].
4.5.2. Maurocalcine from Scorpio Maurus Palmatus and Related Peptides
Maurocalcine (MCa), a 33-amino acid peptide isolated from the venom of Scorpio maurus palmatus, exhibits dual functionality as a cell-penetrating peptide and a modulator of intracellular calcium signaling, with promising applications in oncology [63,184,185,186]. Structural analyses revealed three disulfide bonds and a cationic surface that facilitates membrane translocation, while its binding to ryanodine receptors (RyR1) triggers Ca²⁺ release from the endoplasmic reticulum [63,187]. Further studies have mapped its interaction with RyR1’s cytoplasmic domain, identifying key residues, such as Lys20, critical for both receptor activation and cell penetration [63,66,188,189]. Interestingly enough, a short 9-amino acid derivative from MCa known as MCaUF1-9 showed very favorable cell-penetrating efficacy and may be used to specifically target cancer cells in vivo, due to its acidic pH that matches tumor acidic environments [190]. Moreover, MCa has been shown to overcome doxorubicin resistance in MDA-MB231 breast cancer cell line [64]. Another MCa derivative, Pt-1-DMCa, a platinum-maurocalcin conjugate, has been shown to induce apoptosis in human glioblastoma U87 cells through ROS-dependent modulation of the PI3K/AKT/OfoxO3a signaling pathway [65]. Furthermore, different studies have revealed novel applications for MCa in delivering therapeutic cargo to cancer cells [63,184,185,186,191]. Overall, MCa and its peptide derivatives´ ability to efficiently cross cell membranes has made it an attractive candidate for drug delivery systems.
5. Cancer-Specific Targeting Mechanisms
5.1. Blood-Brain Barrier Penetration
The ability of certain scorpion venom components to cross the blood-brain barrier (BBB) has been extensively characterized, revealing unique structural and functional mechanisms that make them valuable for targeting brain cancers. Mass spectrometry-based proteomic studies and structural studies have identified specific post-translational modifications such as C-terminal amidation and disulfide bonds that enhance BBB penetration while preserving therapeutic efficacy [192,193,194].
Chlorotoxin, discussed in detail in Section 4.5.1, exemplifies a peptide capable of penetrating the blood-brain barrier and selectively targeting gliomas.
Mechanistically, venom peptides, such as the other CTX derivative MiniCTX3, exploit endogenous transport systems such as receptor-mediated transcytosis, to cross the BBB, as revealed through protein-protein interaction networks and surface plasmon resonance (SPR) assays [182]. Other peptides, including BmK components such as BmKCT from Buthus martensii modulate BBB permeability by downregulating tight junction proteins, enhancing drug delivery to brain tumors [27,44,195]. These insights have spurred the development of targeted delivery systems, such as CTX-conjugated nanoparticles, which enhance chemotherapeutic uptake in glioblastomas while minimizing off-target effects [47,196,197].
5.2. Molecular targeting Mechanisms
Scorpion venom peptides exhibit precise molecular targeting, disrupting key pathways in cancer cells. Several studies including proteomic analysis have revealed that these peptides interact with ion channels, matrix metalloproteinases, growth factor receptors, and integrins, leading to selective cytotoxicity.
5.2.1. Ion Channel Interactions
Many scorpion venom peptides exert anticancer effects by modulating ion channels. AGAP from Buthus martensii inhibits voltage-gated sodium channels (Nav1.4, Nav1.5, Nav1.7 and Nav1.8), which are overexpressed in breast, prostate, and colon cancers, disrupting tumor proliferation, inducing apoptosis, migration, and metastasis [29,35,98,198,199,200]. Chloride channels, particularly overexpressed in gliomas, are another key target. BotCl, a chlorotoxin-like peptide from Buthus occitanus tunetanus, selectively blocks ClC-3 chloride channels in glioblastoma and breast cancer, impairing migration [48]. In addition, CTX has also been described to target MMP-2 and ClC-3 on the surface of human glioma cells, inhibiting glioma cell invasion [49,164,169,171], as well as voltage-activated chloride channels in glioma cells that facilitate cell volume changes essential for migration and invasion [201].
Potassium channels are often found to be expressed differently in cancer cells compared to normal cells. Iberiotoxin (IbTX), derived from Buthus tumulus, targets Kv1.1 and Kv1.3 potassium channels, triggering calcium dysregulation and apoptosis in cervical and ovarian cancers [58].
5.2.2. Receptor-Mediated Effects
Scorpion venom components also act through receptor-mediated mechanisms. The tetrapeptide AaTs-1 from Androctonus australis antagonizes formyl-peptide receptor-like 1 (FPRL-1) in glioblastoma, upregulating p53 while suppressing ERK/p38/JNK signaling [123]. Although the direct receptor target of BmK AGAP has not been identified, evidence suggests that it modulates receptor-associated signaling pathways. In breast cancer cells, BmK AGAP downregulates pentraxin-3 (PTX3), a molecule involved in receptor–ligand interactions within the tumor microenvironment. This leads to suppression of the NF-κB/Wnt/β-catenin axis, ultimately reducing stemness and epithelial–mesenchymal transition (EMT) [145]. Neopladines 1 and 2 from Tityus discrepans upregulate Fas ligand (FasL) in breast cancer cells, activating extrinsic apoptosis [67].
6. Immunomodulatory Effects
Scorpion venom contains a diverse array of bioactive peptides that can modulate both innate and adaptive immune responses. These peptides have been shown to influence immune cell function, including the repolarization of immune cells and the enhancement of antigen-specific responses.
6.1. Innate Immune Response Modulation
Proteomic analyses have revealed that scorpion venoms contain diverse proteins and peptides that modulate innate immune responses relevant to cancer. Several biochemical and mass spectrometry-based proteomic analyses have revealed that scorpion venom contains peptides and proteins with potential bioactive properties, including immunomodulatory effects, which can influence innate immune cells. Cota-Arce and colleagues have identified several proteins derived from the Centruroides limpidus venom, including neurotoxins, metalloproteases, phospholipases, hyaluronidases, and antimicrobial peptides. Among them, fractions CIF8 and CIF9 were able to induce the anti-inflammatory IL-10 while suppressing the pro-inflammatory IFN-γ in CD4+ T cells via Ca2+ channel modulation [202]. Moreover, they trigger innate immune crosstalk by activating macrophages and dendritic cells, evidenced by elevated IL-12 and TNF-α in co-cultures. IL-10 suppression could counteract tumor-associated immunosuppression, while Ca2+ channel-targeting toxins may disrupt cancer cell signaling [202]. Overall, this study demonstrated that these venom-derived proteins can shift the immune response toward Th1, Th2, or Th17 profiles, indicating their potential as modulators of T cell-mediated immunity that may be able to further explore their role in immune regulation relevant to cancer immunotherapy.
Another venom component, the peptide Css54 isolated from Centruroides suffusus suffusus has demonstrated significant immunomodulatory effects. Css54 enhances macrophage phagocytic activity while reshaping cytokine production. It suppresses IL-6, increases the anti-inflammatory IL-10, and modestly elevates IL-12p70 and TNF-α, with minimal impact on IFN-γ. This balanced immunomodulation suggests an ability to promote antimicrobial defense while dampening excessive inflammation [203]. These results position Css54 as a dual-function peptide with potential therapeutic applications in infection control and immune regulation, and they further raise the prospect of exploiting Css54 to modulate the tumor microenvironment by reinforcing innate immune mechanisms while preventing chronic inflammation.
The venom's ability to modulate toll-like receptor (TLR) signaling pathways is particularly noteworthy for its potential initial immune response against cancer cells. The peptide Ts1 isolated from Tityus serrulatus venom activates TLR2, TLR4, and CD14 on macrophages. This triggers MyD88-dependent NF-κB activation and TLR4-dependent, MyD88-independent c-Jun activation, as well as engagement of the ERK1/2 and p38 MAPK pathways, culminating in the release of TNF-α and IL-6 [204].
Venom-induced immune modulation has emerged as an important mechanism with potential application in cancer therapy. Several studies demonstrate that scorpion venoms can regulate macrophage polarization, shifting cells between pro-tumorigenic M2 and anti-tumorigenic M1 phenotypes [205]. Venom from Heteroctenus junceus (previously known as Rhopalurus junceus) was shown to modulate pro-inflammatory cytokine production in F3II mouse mammary tumor cells, significantly reducing IL-6 and IL-1β while elevating TNF-α and IL-12, consistent with interference in the NF-κB pathway [206]. Oral administration of H. junceus venom in F3II tumor-bearing mice suppressed tumor growth and decreased serum TNF-α levels, indicating systemic immunomodulation [207]. These dual effects on cytokine regulation suggest that venom-derived molecules may repolarize tumor-associated macrophages from an M2- to an M1-like state, thereby reprogramming the tumor microenvironment toward an anti-tumor phenotype. Supporting this concept, studies with T. serrulatus venom show that macrophage responses are mediated by TLR2/TLR4/CD14 recognition, engaging NF-κB–linked pathways, which can be negatively modulated by PPAR-γ activity [204,208]. Collectively, these findings highlight scorpion venom as a source of immunomodulatory peptides with the potential to enhance antitumor immunity through macrophage reprogramming.
In another study, the toxic fraction FTox-G50 from Androctonus australis hector venom promoted M1 macrophage polarization in adipose tissue, increasing nitric oxide NO production and upregulating IL-12p40, IL-23, and NOS2 while suppressing M2 markers Arg1 and IL-10, in a TNF-α-dependent manner [209].
This modulation of innate immunity shows potential in cancer treatment, as it can help overcome the immunosuppressive tumor microenvironment and promote anti-tumor responses without causing systemic toxicity.
6.2. Adaptive Immunity Enhancement
The impact of scorpion venom on adaptive immunity has emerged as a crucial area of investigation in cancer immunotherapy.
Notably, scorpion venom can act as an adjuvant, promoting Th1-type adaptive immunity and increasing antibody avidity, which may improve the neutralizing capacity of antibodies against systemic threats. For example, the scorpion venom from Hottentotta rugiscutis exhibits adjuvant properties by enhancing adaptive immunity, specifically promoting HBsAg-specific Th1 responses and increasing antibody avidity, which may improve neutralizing capacity against pathogens. This effect is mediated through neuroendocrine-immune interactions, including upregulation of nerve growth factor (NGF) and corticosterone (CORT), leading to activation of splenocytes and sustained IL-1β production (Santhosh et al., 2022).
Conversely, certain scorpion venom peptides selectively suppress pathogenic effector memory T lymphocytes by blocking Kv 1.3 potassium channels. For instance, HsTX1 from Heterometrus spinnifer is a potent Kv1.3 inhibitor (IC₅₀ ≈ 12 pM) [56]. Likewise, Vm24 from Vaejovis mexicanus smithi exhibits extremely high specificity and affinity for Kv1.3 (K_d ≈ 2.9 pM, ~1500-fold selectivity), attenuating human CD4⁺ T_EM cell activation, cytokine production, and proliferation upon stimulation [79,211,212].
Recent studies identified Cm28, a peptide from Centruroides margaritatus, as a high-affinity inhibitor of Kv1.2 and Kv1.3 channels. In human CD4⁺ effector memory T cells, Cm28 suppressed activation markers such as IL-2R (CD25) and CD40L, underscoring its potential as an immunomodulatory agent [51].
This mechanism, classically explored in autoimmunity, may also be exploited in cancer, where Kv1.3 contributes to shaping the tumor–immune interface. In head and neck cancer, Kv1.3high CD8⁺ tumor-infiltrating lymphocytes have been identified as functionally competent effectors [213]. Supporting this rationale, margatoxin (MgTx), another Kv1.3 inhibitor from C. margaritatus, has demonstrated both in vitro and in vivo efficacy against A549 human lung adenocarcinoma, where it suppressed tumor cell proliferation, modulated cell-cycle regulators, and significantly reduced tumor growth in xenograft models [62]. These findings suggest that Kv1.3 inhibition may reprogram immune responses within the tumor microenvironment, potentially reducing Treg activity while preserving cytotoxic T-cell function.
Beyond Kv1.3-targeting peptides, other venom-derived molecules contribute to adaptive immune modulation. For example, peptides AK and GK from Buthus martensii suppress TNF-α/EGFR/STAT3 signaling pathway, downregulating pro-inflammatory cytokines such as TNF-α and oncogenic drivers such as c-Myc while upregulating tumor suppressors (p53/PTEN). In gastric cancer models, these effects may help reverse immune evasion and restore antitumor immunity [214]. Collectively, these mechanisms underscore the potential of scorpion venom peptides to act either as immune adjuvants or as selective immunomodulators, thereby broadening their applicability in cancer immunotherapy.
6.3. Cytokine Profile Alterations
The manipulation of cytokine profiles by scorpion venom components represents a significant breakthrough in understanding their therapeutic potential. Different scorpion venoms are able to modulate cytokine networks through specific toxins, influencing both pro- and anti-inflammatory responses. FTox-G50 from Androctonus australis hector was shown to polarize adipose tissue macrophages (ATMs) toward an M1 phenotype, characterized by upregulated IL-12p40, IL-23, and NOS2 (iNOS) expression, while suppressing M2 markers such as Arginase-1 (Arg1) and IL-10. This shift is TNF-α-dependent, as demonstrated by the reversal of M1 gene expression upon etanercept (TNF-α antagonist) treatment [209]. Similarly, T. serrulatus venom (TsV) triggers systemic release of IL-1β, IL-6, TNF-α, and IFN-γ in severe envenomation cases [215]. In contrast, Leiurus macroctenus venom exhibits an atypical immunomodulatory profile, reducing pro-inflammatory cytokines (IL-6, IL-8, IL-1β) while elevating anti-inflammatory IL-4, IL-10, and IFN-γ in rat lungs, suggesting species-specific immune reprogramming [216]. The antimicrobial peptides BmKn1, BmKn2, and BmKn2-7 from Mesobuthus martensii further illustrate this duality, dampening TNF-α and IL-1β in Litopenaeus vannamei shrimp infected with Vibrio parahaemolyticus, while enhancing immune enzymes like phenoloxidase and complement component C3 [217]. Proteomic analyses of Hottentotta saulcyi venom reveal additional complexity, notably the dominance of Na⁺- and K⁺-channel–targeting peptides, as well as a substantial lipid component (~1.2% dry weight), characterized via LC-MS/MS [52].
These findings underscore the potential of venom components to recalibrate immune responses, though their clinical translation requires further exploration of dose-dependent effects and signaling pathways.
6.4. Immune Cell Activation and Regulation
Scorpion venom components selectively target immune cells, influencing their activation and functional polarization. For example, the TzII and TzIII fractions of Tityus zulianus venom selectively activate human neutrophils, inducing PKC-dependent ROS production, an effect blocked by PKC inhibition [218]. Mass spectrometry has proven indispensable in characterizing such fractions: recent MALDI-TOF and LC-MS/MS analyses revealed that specific peptide components remain non-neutralized after antivenom interaction, as shown in Odonthobuthus doriae venom [219]. These findings underscore that non-neutralized low-molecular-weight peptides may persist and retain biological activity, providing a molecular basis for their selective engagement with neutrophil membranes. However, this effect is cell-type-specific, as eosinophils show negligible respiratory burst compared to neutrophils, reflecting distinct localization and regulatory mechanisms of the NADPH oxidase complex [114]. In macrophages, the β-toxin Ts1, also known as Tsγ, from T. serrulatus venom, is recognized by TLR2, TLR4, and CD14, leading to MyD88-dependent NF-κB activation and triggering the release of IL-6, TNF-α, and nitric oxide (NO) 2011[204,220]. Ts1 has been previously sequenced as a 61-amino-acid β-toxin, confirming its identity and structural class [221]. Conversely, Meuk7–3 from Mesobuthus eupeus suppresses effector memory T cells by blocking Kv1.3 channels, a strategy relevant for autoimmune diseases [222]. The Scorpine peptide from Pandinus imperator exemplifies dual functionality, activating phagocytes while exhibiting antimicrobial properties, though its precise immune targets remain under investigation [223]. Proteomic gaps persist for several scorpion venoms, including Androctonus crassicauda, despite its medical importance and demonstrated biological activities. These studies highlight the potential of venom-derived peptides to modulate immune cell function, with implications for treating inflammatory disorders and cancer.
6.5. Potential for Immunotherapy Enhancement
Scorpion venom peptides offer promising avenues for cancer immunotherapy by targeting hallmarks of malignancy. Chlorotoxin (CTX) from Leiurus hebraeus binds glioma-specific chloride channels and MMP-2, inhibiting metastasis and enhancing blood-brain barrier penetration for drug delivery [49,146]. Its 4 kDa structure, confirmed by MALDI-TOF MS, underpins its clinical use in tumor imaging, such as CTX-Cy5.5 for fluorescence-guided surgery [44,49]. Similarly, BmK-AGAP from Buthus martensii Karsch exhibits analogous antitumor and immunomodulatory activities, though its characterization extends beyond RP-HPLC to include genomic and functional analyses (Y. Zhang et al., 2025).
The Kv1.3 blockers, such as Meuk7–3 from Mesobuthus eupeus, demonstrate translational potential by suppressing autoreactive T cells in autoimmune diseases, with MALDI-TOF and structural modeling confirming their ion channel-targeting motifs [222].
Androctonus crassicauda venom and its peptides further expand this repertoire, inducing apoptosis and cell-cycle arrest in MCF-7 cells, with caspase-3 involvement; transcriptomic and proteomic studies of A. crassicauda venom glands detail the underlying peptide diversity [33,117,122,224,225,226,227].
Challenges include scalable production, addressed via heterologous expression in Escherichia coli, and precise immune modulation to increase bioactivity capabilities and avoid cytokine storms (Y. Zhang et al., 2025).
Together, these components highlight the dual utility of venom peptides: as direct cytotoxic agents and as immune modulators to enhance checkpoint inhibitor therapies.
7. Diagnostic Applications
Scorpion venom-derived peptides have emerged as innovative tools for tumor marking and imaging, owing to their high affinity for cancer-specific targets. Among them, chlorotoxin (CTX) has been extensively studied as a tumor-targeting agent due to its ability to selectively bind glioma and other tumor cells. Building on this specificity, synthetic derivatives such as BLZ-100 (tozuleristide, Tumor Paint) have been developed by conjugating CTX to fluorescent dyes, enabling real-time intraoperative visualization of tumor margins.
Preclinical studies demonstrated that CTX-based probes selectively label glioma, medulloblastoma, sarcoma, prostate, and colorectal tumors in mouse models, with near-infrared fluorophores offering optimal intraoperative imaging [197,228]. Comparative oncology trials in dogs confirmed safety and effective tumor visualization [229], highlighting the translational value of this approach [230]. Toxicology studies in multiple species, including rodents, dogs, and non-human primates, further established a favorable safety profile [231].
In humans, BLZ-100 has progressed to Phase 2/3 trials for pediatric central nervous system tumors, demonstrating safe administration and improved surgical precision through fluorescence-guided resection. Beyond gliomas, BLZ-100 has shown promise in identifying other lesions, such as cerebral vascular malformations [232], as well as non-melanoma skin cancers and melanoma, where it achieved accurate tumor imaging at clinically feasible doses [233]. These results underscore the potential of CTX-based probes to improve tumor visualization across diverse cancer types while maintaining low systemic toxicity.
7.1. Early Detection Methods
For early detection of Hottentotta tamulus (syn. Mesobuthus tamulus) envenomation, mass spectrometry-based proteomics was able to identify a rich repertoire of low-molecular-mass toxins, particularly Na+ and K+ ion-channel toxins that comprise a majority of the venom proteome [234]. To improve detection sensitivity in plasma, acetonitrile precipitation has been used to enrich these low-abundance peptides while simultaneously depleting high-molecular-weight plasma proteins, thereby facilitating their identification by mass spectrometry [235,236].
Although the direct application of gold nanoparticle (AuNP)–antibody conjugates and localized surface plasmon resonance (LSPR) biosensors to scorpion venom toxins has not yet been reported, AuNP-based platforms are well established in clinical diagnostics for providing rapid (often <10 min) colorimetric readouts and picomolar sensitivity for diverse biomolecules [237,238,239]. This suggests that similar systems could, in principle, be adapted for the ultrasensitive detection of scorpion venom peptides in envenomed patients. Such an approach would not only accelerate diagnosis in clinical settings but also establish a foundation for broader applications in oncology and precision medicine.
The specificity of venom peptide recognition, exemplified by Tamapin, a K⁺ channel toxin targeting SK2 [240], and α-neurotoxins acting on Na⁺ channels, underscores the potential of scorpion toxins as molecular probes. In oncology, chlorotoxin from Leiurus hebraeus provides a precedent: it binds selectively to tumor-associated receptors such as MMP-2 in gliomas and has been widely employed in tumor-targeting bioconjugates for both imaging and therapeutic applications (Deshane et al., 2003).
Beyond diagnostics, this dual-use paradigm offers therapeutic opportunities. Kv10.1 (Eag1), a voltage-gated K⁺ channel aberrantly expressed in many cancers, represents a validated pharmacological target [241,242,243]. Conjugation of venom-derived peptides such as chlorotoxin or venom-derived cell-penetrating peptides (CPPs) to nanocarriers—including AuNPs, chitosan, polyethylene glycol (PEG), and polyethylenimine (PEI)—has been shown to improve tumor-selective delivery and anti-tumor efficacy while minimizing off-target effects [244,245,246]. By bridging diagnostics and therapy, this translational framework highlights the untapped potential of scorpion venom peptides in oncology and points toward their integration into next-generation precision medicine [235].
7.2. Biomarker Development
In biomarker discovery, untargeted UHPLC-QTOF-MS metabolomic profiling of hepatocellular carcinoma (HCC)-bearing mice treated with scorpion venom peptide extract (PESV) from Buthus martensii revealed 111 significantly altered serum metabolites (48 in negative ion mode, 63 in positive ion mode), highlighting major disruptions in pathways such as aminoacyl-tRNA biosynthesis, amino acid metabolism, glutathione metabolism, protein transport, protein digestion and absorption, and cAMP signaling [247]. These findings position PESV-induced metabolic signatures as potential diagnostic and therapeutic biomarkers in HCC. The observed pathway-level perturbations suggest that scorpion venom peptides could modulate key metabolic networks relevant to tumor progression and therapy. Future studies should clarify whether these shifts intersect with known cancer-related mechanisms, including immune evasion, nitrogen metabolism, or drug resistance. Bridging these metabolomic insights with venom peptide pharmacology may open avenues for developing venom-based diagnostics and therapeutic strategies in precision oncology.
7.3. Therapeutic Monitoring
The neurotoxic properties of Odontobuthus doriae venom, which disrupts ion channels and neurotransmitter activity, also make it a promising candidate for diagnostic applications. Recent advances in plasmonic biosensing technology have enabled the detection of venom-induced neurotoxicity in human serum through circular dichroism (CD) measurements. By employing an achiral gold-coated plasmonic nanostructure, researchers demonstrated that minute changes in venom concentration alter the Stern layer’s action potential, shifting the sensor’s refractive index and producing measurable CD responses. This method achieved high sensitivity (27.4 at 520 nm) and rapid detection, offering a potential tool for diagnosing envenomation or monitoring venom-derived therapeutics. Such biosensors could be adapted to detect cancer-specific biomarkers, leveraging venom components' selective binding properties to improve early diagnosis and personalized treatment strategies [248].
7.4. Integration with Current Diagnostic Tools
Integration with current diagnostic tools further expands the clinical potential of venom-based agents. For instance, chlorotoxin (CTX) has been combined with MRI and near-infrared fluorescence in preclinical studies for glioblastoma detection [49,249,250]. In addition, CTX-drug conjugates such as CTX–onconase have shown increased antitumor efficacy in glioma models compared to unconjugated mixtures [251], while CTX-decorated liposomal nanoparticles have effectively delivered siRNA or antisense oligonucleotides to glioblastoma cells in vitro and in vivo [252]. Despite these promising results, challenges remain in translating these approaches into routine clinical use, particularly regarding standardization, scalability, and regulatory approval. Nonetheless, the high specificity of CTX for tumor cells, its modular platform design, and its blood–brain-barrier permeability position it as a valuable adjunct to conventional diagnostic and therapeutic strategies for glioblastoma and related malignancies.
8. Drug Development and Delivery Systems
8.1. Peptide Modification Strategies
The development of peptide modification strategies for scorpion venom components represents a critical advancement in cancer drug development. Mass spectrometry analysis has identified key modification sites, such as disulfide bridges in chlorotoxin (CTX), which enhance structural stability and target specificity for glioma cells [253,254]. The PEGylation of CTX demonstrated that PEG-CTX conjugates significantly improved tumor-targeting efficiency, as evidenced by an 8-fold increase in glioma accumulation compared to unmodified CTX when delivered via polyamidoamine (PAMAM) dendrimer nanoparticles [255]. This modification not only enhanced systemic circulation but also preserved CTX's binding affinity to matrix metalloproteinase-2 (MMP-2), a receptor overexpressed in gliomas, enabling precise intracellular delivery of therapeutic genes such as TRAIL (tumor necrosis factor-related apoptosis-inducing ligand) [255].
Moreover, recent studies with Tityus stigmurus venom-derived peptides demonstrate that PEGylation strategies can be extended to other venom components for cancer applications, as evidenced by PLA-PEG encapsulation of Stigmurin analogs (S1 and S2), which reduced hemolysis by 20% while maintaining antiproliferative activity against macrophage-like RAW264.7 cancer cells, highlighting the dual benefit of improved safety and retained bioactivity in venom-based nanotherapeutics [256].
Cyclization of BmK peptides from Buthus martensii has been shown to enhance their therapeutic potential. In the case of Buthicyclin, a cyclic peptide derived from Defensin 4 (BmKDfsin4), the introduction of a disulfide bond between terminal cysteine residues improved structural stability and prolonged serum half-life [257]. The peptide’s high binding affinity to opioid receptors (Mu/Kappa/Delta types) and low toxicity profile (LD50 > 20 mg/kg, <5% hemolysis at 4 mg/mL) suggests its scaffold could be repurposed for targeting cancer-associated receptors, such as opioid growth factor receptor (OGFR), which is implicated in tumor proliferation and metastasis [257].
8.2. Nanoparticle-Based Delivery
Nanoparticle delivery systems have revolutionized venom peptide therapeutics by addressing challenges such as rapid degradation and poor tissue penetration while improving controlled release, stability, and cellular uptake, all with reduced toxicity [258]. Gold nanoparticles, for instance, have shown promise in cancer treatment due to their ability to be co-functionalized with targeting ligands like peptides and stabilizing agents such as polyethylene glycol (PEG), enhancing cellular uptake while maintaining stability under physiological conditions [244,245,246]. This approach reduces non-specific protein adsorption and immune clearance, positioning gold nanoparticles as promising candidates for targeted drug delivery and imaging in oncology [259].
The anticancer potential of scorpion venom has been further demonstrated through liposomal encapsulation of venoms from Androctonus bicolor, Androctonus crassicauda, and Leiurus quinquestriatus, which significantly enhances their anti-cancer efficacy against colorectal cancer cells (HCT-8). Venom-loaded liposomes exhibited superior stability, controlled release, and increased cytotoxicity compared to free venoms, as evidenced by reduced cell viability, elevated reactive oxygen species (ROS) generation, and induction of apoptosis and cell cycle arrest at the G0/G1 phase. This nano-delivery system not only improved the therapeutic index of the venoms but also minimized non-specific toxicity, underscoring its potential as a targeted and efficient strategy for cancer therapy [28].
Chitosan nanoparticles (CN) have emerged as another promising platform for venom-based cancer therapies. For example, CN enhanced antimicrobial activity of Tityus stigmurus venom (Tsv) through high encapsulation efficiency (>78%), stability, and controlled release, with a small particle size (<180 nm) and positive charge (+23 to +28 mV) that improves cellular interactions [260]. Similarly, Rebbouh et al (2020) demonstrated the efficacy of CN in detoxifying and delivering the neurotoxin Aah II from Androctonus australis hector venom. The nanoparticles achieved 96.66% encapsulation efficiency and a biphasic release profile (55% within 8 hours and 80% over 5 days), significantly reducing toxicity while enhancing immunogenicity. Immunized mice survived lethal toxin doses (up to 8 LD50), highlighting the potential of CN for venom-based therapies [261].
These properties are particularly advantageous for antitumor applications, as chitosan’s biocompatibility and mucoadhesive nature can enhance tumor targeting and retention. By encapsulating venom peptides with known anticancer properties, CN could enhance therapeutic efficacy while minimizing systemic toxicity. Such an approach leverages the dual benefits of detoxification and controlled delivery, offering a promising strategy for developing targeted cancer therapies.
8.3. Targeted Delivery Systems
Targeted delivery systems have been refined through different experimental insights. For example, chlorotoxin CTX-conjugated iron oxide nanoparticles displayed significantly enhanced internalization in glioma cells, with approximately 50% higher uptake in C6 glioma cells compared to untargeted nanoparticles, supporting their role in improving tumor-specific delivery [262]. Beyond demonstrating selective binding and uptake, such systems also open opportunities for integration with proteomic strategies to profile nanoparticle-cell surface protein interactions, enabling a deeper understanding of receptor landscapes that mediate efficiency and refining future venom-based nanocarriers [263].
A particularly relevant innovation has been the development of dual-targeting systems designed to overcome both the blood–brain barrier (BBB) and the tumor microenvironment. In this context, Yue and colleagues engineered PEGylated liposomes conjugated with OX26, an antibody against the transferrin receptor, together with CTX, thereby coupling BBB transcytosis with glioma-specific targeting [264]. The dual-targeting lipoplexes successfully transported therapeutic plasmid DNA across the BBB in vitro and in vivo, decreased C6 glioma cell viability to nearly 46% in a co-culture BBB model, and achieved marked tumor reduction and prolonged median survival (46 days versus 13 days in controls) in rat glioma models. Histopathological and immunohistochemical analyses confirmed that the therapeutic effect was mediated by targeted hTERTC27 gene expression within the tumor site, underscoring the complementary roles of OX26 in mediating BBB transport and CTX in conferring tumor selectivity [264].
Collectively, these findings illustrate how venom-derived ligands such as CTX can be integrated into multifunctional delivery platforms to improve targeting precision and therapeutic efficacy. The combination of proteomic insights into receptor expression with rational design of dual-targeted nanocarriers may pave the way for next-generation scorpion venom–based therapeutics capable of addressing both systemic delivery challenges and the heterogeneity of tumor microenvironments.
9. Clinical Studies and Trials
9.1. Preclinical Studies
Extensive preclinical research has laid a robust foundation for the clinical translation of scorpion-venom–derived peptides. Venomics analysis combining transcriptomics and proteomics, has catalogued venom constituents with high resolution, allowing the identification of chlorotoxin (CTX)-like peptides with anticancer potential. Early venomic efforts provided the essential sequence-level starting points for functional studies [167,265]. Biochemical and structural analyses subsequently mapped CTX mechanisms of action, revealing direct interactions with tumor-associated membrane proteins, such as MMP-2 and NRP1. These interactions clarified the mechanism of cellular uptake and CTX retention within glioma models [49,174].
In vivo studies in rodents and other models reinforced these findings by demonstrating selective tumor binding, effective intratumoral localization, and pronounced antitumor activity of CTX-based agents [168,174]. Imaging-enabled preclinical studies with radiolabeled or fluorescent CTX conjugates tracked biodistribution and tumor localization in real time, thereby visually substantiating CTX´s tumor-targeting capabilities [50,266]. These cumulative preclinical data collectively supported first-in-human testing of CTX-derived agents [52,174], providing the rationale for translation into early-phase clinical trials.
Another significant preclinical development in this domain is the discovery and characterization of the peptide Gonearrestide, an anticancer peptide derived from the Moroccan fat-tail Androctonus mauretanicus scorpion venom. This peptide was identified using an integrated high-throughput platform combining next-generation transcriptome sequencing (NGS) and MS/MS proteomic analysis, which enabled the screening of over 200 novel peptides and rational functional selection [54]. Gonearrestide showed potent in vitro activity, inducing G1 cell-cycle arrest and inhibiting proliferation in human colorectal cancer cells. Mechanistically, it inhibited cyclin-dependent kinase 4 (CDK4) while upregulating key cell-cycle regulators p21, p27, and cyclin D3. Its antitumor activity extended to in vivo xenograft mouse models, confirming potent reduction of tumor growth, with minimal toxicity toward noncancerous epithelial cells and erythrocytes [54]. Systematic reviews further acknowledge Gonearrestide as a leading example of venom-derived peptides with translational potential [26].
Collectively, these investigations highlight how the synergy of molecular venomics, proteomic mapping, and imaging technologies accelerates bench-to-bedside development of scorpion venom-derived peptides. This multidisciplinary approach has not only identified promising excipient peptides such as CTX and Gonearrestide but also elucidated their mechanisms of action, helping bridge molecular discovery with early-phase clinical exploration.
9.2. Clinical Trials
Phase I clinical evaluation of scorpion venom–derived compounds has marked an important step toward their translation into oncology. Among these, chlorotoxin (CTX)–based constructs are the most extensively studied, with two agents: TM-601 and BLZ-100 (Tozuleristide), advancing into first-in-human clinical trials. These compounds were initially designed to assess safety, pharmacokinetics, and tumor-targeting or intraoperative imaging capabilities, and they represent distinct but complementary applications of venom peptides in oncology (Table 2).
9.2.1. TM-601 (131 I-chlorotoxin conjugate)
TM-601 is a synthetic chlorotoxin analog radiolabeled with iodine-131 (131I), for targeted radionuclide delivery to gliomas. In a Phase I single-dose, open-label trial, 18 adult patients with recurrent high-grade gliomas received intracavitary administration of 131I-TM-601 following tumor resection [179]. The compound showed selective tumor uptake and prolonged intracavitary retention for up to 14 days, with minimal systemic distribution and rapid clearance of non-bound peptide within 24 to 48 hours. Importantly, the therapy was well tolerated, and no dose-limiting toxicities were reported. Clinical outcomes were encouraging at 180 days post-treatment, several patients exhibited stable disease or partial radiographic responses, and two remained without evidence of disease for more than 30 months [179]. These findings confirmed the safety and tumor-targeting specificity of 131I-TM-601, providing the basis for subsequent multicenter Phase II studies that further validated its tolerability and suggested signals of tumor control in subsets of patients [173].
9.2.2. BLZ-100 (Tozuleristide, “Tumor Paint”)
BLZ-100, also known as tozuleristide or “Tumor Paint,” represents a diagnostic and surgical innovation derived from chlorotoxin. It consists of chlorotoxin conjugated to a near-infrared fluorophore, enabling real-time intraoperative visualization of tumors [228]. In a Phase I dose-escalation trial in adults with newly diagnosed or recurrent gliomas, BLZ-100 demonstrated a favorable pharmacokinetic profile, absence of dose-limiting toxicities, and robust tumor-specific fluorescence that facilitated surgical delineation of tumor margins [181]. Beyond gliomas, early-phase evaluations have extended its application to breast carcinoma, neck squamous cell carcinoma, cutaneous malignancies, and cerebral vascular malformations, where it has similarly provided intraoperative contrast and improved visualization of neoplastic and malformed tissue [232,233,267,268].
Taken together, these first-in-human studies illustrate the translational trajectory of scorpion venom–derived peptides from preclinical discovery to clinical application. TM-601 highlights the potential of chlorotoxin analogs for selective delivery of therapeutic radionuclides to malignant gliomas, while BLZ-100 demonstrates their utility as tumor-targeted imaging agents capable of enhancing surgical precision. Both exemplify how venomic and proteomic research can yield clinically valuable innovations at the interface of targeted therapy and image-guided oncology.
10. Current Challenges and Future Directions
10.1. Production and Scale-up Issues
The production and scale-up of scorpion venom–derived therapeutics remain major challenges in their development as anticancer agents. The structural complexity of venom peptides, particularly their disulfide-rich scaffolds, makes large-scale synthesis and recombinant expression technically demanding. Recombinant systems, including bacterial, yeast, and insect cell hosts, have been explored to produce bioactive peptides, although maintaining proper folding and disulfide connectivity is a persistent obstacle [27,136]. Mass spectrometry and proteomic analyses have been essential for characterizing venom components and verifying recombinant products, confirming sequence integrity, disulfide patterns, and post-translational modifications. These analytical approaches support quality control and help ensure batch-to-batch consistency. However, true large-scale manufacturing remains difficult, and further advances in expression technologies, folding strategies, and purification methods are required before venom-derived peptides can be produced reliably for widespread clinical use.
10.2. Regulatory Considerations
The regulatory development of scorpion venom–derived therapeutics presents unique challenges, particularly related to product characterization, manufacturing consistency, and safety evaluation. The structural complexity of venom peptides, including their disulfide-rich scaffolds and post-translational modifications, necessitates sophisticated analytical methods to meet regulatory standards. Mass spectrometry and proteomic approaches have been valuable in supporting early-stage product characterization by providing detailed molecular fingerprints, verifying disulfide connectivity, and assessing purity. These analyses contribute to quality control and the establishment of critical quality attributes, which are central to regulatory submissions [27,136]. However, the integration of proteomic tools into regulatory processes for venom-derived therapeutics remains largely preclinical and supportive, rather than being mandated for safety monitoring or biomarker validation. Future regulatory pathways may benefit from standardized and validated proteomic methodologies; however, conventional analytical chemistry and pharmacology approaches currently remain the backbone of regulatory compliance.
10.3. Cost and Accessibility
The production and scale-up of scorpion venom–derived therapeutics face substantial hurdles that stem from the structural complexity and heterogeneity of venom peptides, many of which are disulfide-rich and require precise folding and post-translational features to retain activity. As a result, development programs increasingly favor synthetic or recombinant routes over direct venom extraction, but these approaches introduce manufacturing challenges in expression, folding, and downstream purification. Proteomics-driven venomics-centered on high-resolution LC-MS/MS has become indispensable for defining critical quality attributes (identity, purity, isoform composition, and stability) and for establishing batch comparability, thereby informing process optimization and release testing. These analytical requirements, together with the need for controlled production of correctly folded peptides, contribute to the overall complexity and cost of goods, and they help explain why only a few scorpion-toxin candidates (e.g., chlorotoxin derivatives) have advanced clinically while no scorpion-venom drug has yet reached the market. Current literature highlights these scientific and technological bottlenecks and points to ongoing advances in recombinant expression, folding strategies, and high-throughput venomics as levers to improve manufacturability and scalability, though further optimization and standardization will be needed before routine, cost-efficient production is achieved [26,269,270,271].
10.4. Research Gaps
Comprehensive venomics has clarified that major knowledge gaps still limit the development of scorpion venom–derived therapeutics for cancer. Large portions of the venom peptidome remain insufficiently characterized across species, with unresolved questions about isoform diversity, post-translational modifications, and their impact on stability and bioactivity [80,86,175,193,219,272,273,274,275,276,277]. Proteomics and transcriptomics have mapped candidate toxins and hinted at complex mechanisms, yet for lead bioactive peptides such as chlorotoxin, target engagement and downstream pathways remain debated, underscoring the need for rigorous mechanism-of-action studies in relevant cancer models and microenvironments [49,169,278,279,280]. Furthermore, translational gaps persist: there are no validated proteomic biomarkers for patient selection, pharmacodynamic readouts, or resistance monitoring in clinical settings, reflecting the absence of efficacy-focused trials to generate such data. While [¹³¹I]-TM-601 has undergone Phase I-II clinical trials for recurrent glioma, these studies were primarily safety-oriented and did not include broad biomarker development [177,179]. Addressing these gaps will require standardized, cross-lab venomics workflows, including top-down and de novo analysis, to define critical molecular features. Systematic studies of PTMs and structure–function relationships, and integrated preclinical pipelines that couple proteomics with functional assays to prioritize truly druggable venom leads [44,80,174,277,281,282].
10.5. Future Research Directions
The future development of scorpion venom–derived cancer therapeutics holds significant promise, with proteomic and transcriptomic analysis continuing to guide discovery and preclinical evaluation. Venomics approaches integrating mass spectrometry-based proteomics and sequencing have expanded the catalog of bioactive peptides, enabling the identification of novel components with potential therapeutic applications [80]. Efforts to improve specificity and efficacy increasingly rely on structural and proteomic characterization of toxin–target interactions, exemplified by ongoing studies on chlorotoxin derivatives [175]. Advances in delivery systems, including nanoparticle formulations and peptide conjugates, are being investigated to improve stability and tumor selectivity. Although not yet widely applied to scorpion toxins, nanoparticle delivery of venom-derived peptides in broader venom research demonstrates proof-of-concept potential [283].
Looking ahead, the integration of artificial intelligence with proteomic datasets offers a path toward accelerating venom peptide discovery and prioritizing candidates with favorable properties [284,285]. Combination therapies also represent a promising avenue, as proteomic profiling can reveal synergistic effects between venom peptides and established anticancer agents [44]. Finally, the application of proteomic biomarkers to stratify patients and guide treatment design remains largely speculative, but it represents an important goal for developing personalized venom-derived therapeutics. Collectively, these strategies are expected to produce more effective and targeted interventions while addressing current translational limitations.
11. Conclusion and Future Perspectives
Scorpion venom represents a highly diverse natural library of peptides and proteins with unique structural and functional characteristics, making them valuable candidates for anticancer drug development. Proteomic advances have been pivotal in uncovering this molecular diversity, enabling the identification of novel bioactive compounds and mapping their mechanisms of action at unprecedented resolution. These peptides have demonstrated the capacity to selectively target ion channels, disrupt tumor-supportive signaling pathways, induce apoptosis, and remodel the tumor microenvironment while often sparing healthy cells. In addition, the capacity of some venom-derived peptides, such as chlorotoxin, to cross the blood-brain barrier highlights their translational potential in targeting aggressive and otherwise inaccessible tumors.
Despite these advances, important challenges remain. The clinical development of venom-derived therapeutics is hindered by obstacles such as large-scale peptide synthesis, in vivo stability, pharmacokinetics limitations, delivery strategies, and the risk of off-target effects. Addressing these barriers will be essential to enable the translation from preclinical to clinical application. In this regard, the future of scorpion venom-based drug discovery is closely linked to the evolution of proteomic technologies. Single-cell venom gland proteomics promises to delineate precise cellular origins of toxins, uncovering rare peptides and clarifying the understanding of the regulation of venom biosynthesis. Top-down proteomics will allow comprehensive characterization of intact venom proteins and their native post-translational modifications, providing insights into proteoforms that directly correlate with biological activity. Furthermore, interactomics will be key to systematically mapping the binding partners of venom peptides within cancer cells, validating molecular mechanisms, and identifying opportunities for rational combination therapies.
Beyond discovery, proteomics will play a pivotal role in addressing translational challenges. Advanced mass spectrometry will ensure batch-to-batch consistency during manufacturing by confirming sequence integrity, disulfide connectivity, and overall purity. Furthermore, proteomic data will guide the rational design of optimized analogs with improved stability. reduced immunogenicity, and improved pharmacological profiles through strategies such as peptide cyclization and PEGylation. The integration of proteomic biomarkers into preclinical and clinical settings could also help stratify patients, monitor therapeutic efficacy, and quickly identify mechanisms of resistance.
Looking ahead, the integration of proteomics with transcriptomics, structural biology, computational modeling, and synthetic biology will be crucial to optimizing venom-derived therapeutics. Strategies such as nanoparticle conjugation or imaging probes, and targeted delivery strategies may enhance efficacy and minimize off-target effects. Moreover, the immunomodulatory and tumor-microenvironment–modulating properties of certain venom peptides highlight promising synergies with immunotherapies and standard treatments.
In summary, scorpion venom remains an underexploited source of bioactive molecules with significant potential in oncology. By combining high-throughput omics technologies, nanotechnology, medicinal chemistry, artificial intelligence, and translational research may transform these natural peptides into clinically relevant anticancer agents. Realizing this promise will require sustained interdisciplinary collaboration to overcome current barriers and accelerate innovation. While significant progress has been made, key proteomic gaps remain, particularly in the detection of low-abundance peptides, characterization of post-translational modifications, and integrating with other omics layers such as transcriptomics and structural biology. Future advances in multi-omics and computational approaches will be critical to translating scorpion venom peptides into effective therapeutic tools against cancers resistant to conventional treatment.
Author Contributions
S.S.S.A., I.O.C.P., K.B.E.: Conceptualization, Writing-original draft; M.R.C.A., M.R.S, N.G.A.: Writing-original draft, Writing – review & editing; Visualization; F.V.C.P. Writing – review & editing; L.K.I.: Conceptualization, Writing-original draft, Writing – review & editing; Visualization; Funding acquisition, Supervision
Funding
This work was supported by the grants 2024/16166-0, 2019/20832-7, 2017/17943-6, 2016/04000-3, and 2013/07467-1 from the São Paulo Research Foundation (FAPESP). SSSA was supported by CNPq # 131408/2019-4 (University of São Paulo, Brazil). IOCP was supported by the Coordination for the Improvement of Higher Education Personnel (CAPES, Brazil) institutional fellowship # 88887.804166/2023-00 (University of São Paulo, Brazil). MRCA was supported by CAPES #88882.442313/2019-01 (Butantan Institute, Brazil).
Acknowledgments
The authors thank Dr. Milton Nishiyama Jr. for insightful discussions. We also gratefully acknowledge Gustavo Henrique Rhor Souza Machado for his valuable assistance with figure assembly and design.
Conflicts of Interest
The authors declare no conflicts of interest
Abbreviations
The following abbreviations are used in this manuscript:
| ΔΨm | Mitochondrial membrane potential. |
| A2780 | Human Ovarian Cancer Cell Line A2780. |
| Aah II | Aah II neurotoxin. |
| AaTs-1 | Androctonus australis Toxin-1. |
| ABC | ATP-binding cassette. |
| AcrAP1 | Androctonus crassicauda Antimicrobial Peptides 1. |
| AcrAP2 | Androctonus crassicauda Antimicrobial Peptides 2. |
| AGAP | Analgesic-Antitumor Peptide. |
| AGAP-SYPU2 | Analgesic-Antitumor Peptide Variant SYPU2. |
| AI | Artificial intelligence. |
| AK | Adenylate kinase. |
| Akt | Protein Kinase B. |
| AMPK | AMP-Activated Protein Kinase. |
| AMPs | Antimicrobial Peptides. |
| AP-1 | Activator Protein 1. |
| Arg1 | Arginase-1. |
| ATMs | Tumor-Associated Macrophages. |
| ATP | Adenosine Triphosphate. |
| AuNP-antibody | Gold nanoparticle conjugated to antibody. |
| B16-F10 | Murine Melanoma Cell Line B16-F10. |
| Bax | Bcl-2-Associated X Protein. |
| BBB | Blood-Brain Barrier. |
| Bcl-2 | B-cell Lymphoma 2. |
| BKCa | Large-Conductance Calcium-Activated Potassium Channel. |
| BLZ-100 | Chlorotoxin-based Tumor Paint Bioconjugate. |
| BmHYA1 | Buthus martensii Karsch Hyaluronidase 1. |
| BmK | Buthus martensii Karsch. |
| BmKCT | Buthus martensii Karsch chlorotoxin. |
| BmKDfsin4 | Derived Defensin-4 peptide. |
| BmKn1 | Antimicrobial peptides derived from Buthus martensii Karsch venom. |
| BmKn2–7 | Anticancer peptides derived from Buthus martensii Karsch venom. |
| BotCl | Buthus occitanus tunetanus Chloride Channel Blocker. |
| BTB | Blood-tumor barrier. |
| Buthicyclin | Cyclic peptide derived from Defensin-4. |
| C3 | Complement component 3. |
| C6 | Glioma cell line. |
| Ca²⁺ | Calcium ion. |
| CAM | Chorioallantoic Membrane. |
| Caspase-3 | Effector caspase enzyme. |
| CD | Circular dichroism. |
| CD40L | CD40 ligand. |
| CD44 | Cluster of Differentiation 44. |
| CD44v6 | CD44 Variant Isoform 6. |
| CDK | Cyclin-Dependent Kinase. |
| CDK4 | Cyclin-Dependent Kinase 4. |
| CDKI | Cyclin-Dependent Kinase Inhibitor. |
| CDKIs | Cyclin-Dependent Kinase Inhibitors. |
| CDKs | Cyclin-dependent kinases. |
| cDNA | Complementary DNA. |
| ChTX | Charybdotoxin. |
| CIF8 | Cancer Inhibitory Fractions 8. |
| CIF9 | Cancer Inhibitory Fractions 9. |
| ClC-3 | Chloride Channel Protein 3. |
| Cm28 | Centruroides margaritatus peptide/toxin. |
| CN | Chitosan nanoparticles. |
| CNS | Central nervous system. |
| CORT | Cortistatin. |
| Css54 | Cationic antimicrobial peptide 54. |
| CTX | Chlorotoxin. |
| D3 | Cyclin D3. |
| DISC | Death-inducing signaling complex. |
| DNA | Deoxyribonucleic acid. |
| DU145 | Human Prostate Carcinoma Cell Line DU145. |
| ECM | Extracellular Matrix. |
| EGFR | Epidermal Growth Factor Receptor. |
| ELISA | Enzyme-linked immunosorbent assay. |
| ERK1/2 | Extracellular Signal-Regulated Kinases 1/2. |
| F3II | Murine mammary adenocarcinoma cell line F3II. |
| FasL | Apoptosis-inducing ligand binding to Fas receptor. |
| FDA | Food and Drug Administration. |
| FoxO | Forkhead Box O. |
| FPRL-1 | Formyl Peptide Receptor-Like 1. |
| FTox-G50 | Scorpion venom fraction Toxin G50. |
| G0 | Cell cycle phases of resting. |
| G1 | Gap 1 phase of the Cell Cycle. |
| GK | Glucokinase. |
| H22 | Murine Hepatocellular Carcinoma Cell Line H22. |
| H460 / NCI-H460 | Human Non-Small Cell Lung Carcinoma Cell Line. |
| HA | Hyaluronic Acid. |
| HBsAg-specific | Hepatitis B surface antigen-specific. |
| HCC | Hepatocellular carcinoma. |
| HCT116 | Human Colorectal Carcinoma Cell Line HCT116. |
| HCT-8 | Colorectal cancer cell line. |
| HeLa | Human Cervical Cancer Cell Line HeLa. |
| HER2 | Human epidermal growth receptor 2. |
| HPLC | High-Performance Liquid Chromatography. |
| HSP70 | Heat shock protein 70. |
| HSP90 | Heat shock protein 90. |
| HsTX1 | Heterometrus scaber toxin 1. |
| hTERTC27 | C-terminal 27 kDa polypeptide fragment of human telomerase reverse transcriptase. |
| HUVEC | Human Umbilical Vein Endothelial Cells. |
| IbTX | Iberiotoxin. |
| IC₅₀ | Half Maximal Inhibitory Concentration. |
| IFN-γ | Interferon-gamma. |
| IGR39 | Human Melanoma Cell Line IGR39. |
| IL-10 | Interleukin-10. |
| IL-12p40 | Interleukin-12 subunit p40. |
| IL-12p70 | Biologically active heterodimeric form of interleukin-12. |
| IL-1β | Interleukin-1 beta. |
| IL-23 | Interleukin-23. |
| IL-2R | Interleukin-2 receptor. |
| IL-4 | Interleukin-4. |
| IL-6 | Interleukin-6. |
| IL-8 | Interleukin-8. |
| iTRAQ | Isobaric tags for relative and absolute quantification. |
| JAK | Janus kinase. |
| Kd | Equilibrium Dissociation Constant. |
| Kv1.1 | Voltage-gated Potassium Channel Subfamily A Member 1. |
| Kv1.3 | Voltage-gated Potassium Channel Subfamily A Member 3. |
| Kv10.1 | Voltage-gated potassium channel 10.1. |
| LC-MS/MS | Liquid Chromatography–Tandem Mass Spectrometry. |
| LD50 | Median lethal dose. |
| LS174 | Human Colon Adenocarcinoma Cell Line LS174. |
| LSPR | Localized plasmon resonance. |
| M1 macrophages | Classically activated macrophages. |
| M2 macrophages | Alternatively activated macrophages. |
| MALDI-TOF MS | Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. |
| MAP kinases | Mitogen-activated protein kinases. |
| MAPK p38 | p38 Mitogen-Activated Protein Kinase. |
| MCa | Micaelase. |
| MCF-7 | Human Breast Cancer Cell Line MCF-7. |
| MDA-MB-231 | Human Triple-Negative Breast Cancer Cell Line. |
| MDA-MB-435s | Human Breast Carcinoma Cell Line. |
| Meuk7–3 | Peptide fraction Meuk7–3. |
| MgTX | Margatoxin. |
| MiniCTX3 | Charybdotoxin analogue peptide. |
| MMP-2 | Matrix metalloproteinase-2. |
| MRI | Magnetic resonance imaging. |
| MS | Mass spectrometry. |
| MS/MS | Tandem mass spectrometry. |
| MT1-MMP | Membrane-Type 1 Matrix Metalloproteinase. |
| mTOR | Mechanistic Target of Rapamycin. |
| MyD88 | Myeloid Differentiation Primary Response 88. |
| NADPH | Nicotinamide Adenine Dinucleotide Phosphate. |
| Nav1.4 | Voltage-gated Sodium Channel, Alpha Subunit 1.4. |
| Nav1.5 | Voltage-gated Sodium Channel, Alpha Subunit 1.5. |
| Nav1.7 | Voltage-gated Sodium Channel, Alpha Subunit 1.7. |
| Nav1.8 | Voltage-gated Sodium Channel, Alpha Subunit 1.8. |
| NCI-H460 | Human Lung Cancer Cell Line NCI-H460. |
| NDBP | Non-Disulfide Bridged Peptide. |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer. |
| NGF | Nerve growth factor. |
| NGS | Next-generation sequencing. |
| NOS2 | Inducible Nitric Oxide Synthase. |
| NRP1 | Neuropilin-1. |
| OGFR | Opioid growth factor receptor. |
| FOXO3a | Forkhead box O3a. |
| p21 | Cyclin-dependent kinase inhibitor 1A. |
| p27 | Cyclin-dependent kinase inhibitor 1B. |
| p53 | Tumor suppressor protein. |
| PAMAM | Polyamidoamine. |
| PARP | Poly polymerase. |
| PC-3 | Human Prostate Carcinoma Cell Line PC-3. |
| PEG | polyethylene glycol. |
| PEG-CTX | Polyethylene glycol–chlorotoxin conjugate. |
| PEGylation | Polyethylene glycol conjugation. |
| PESV | Polypeptide Extract from Scorpion Venom. |
| PI3K | Phosphatidylinositol 3-Kinase. |
| PLA-PEG | Polylactic acid–polyethylene glycol copolymer. |
| PLA2 | Phospholipase A2. |
| PPAR | Peroxisome Proliferator-Activated Receptor. |
| PTEN | Fosfatase supressora tumoral. |
| PTMs | Post-translational modifications. |
| RAW264.7 | Macrophage-like cell line. |
| RK1 | Scorpion-derived Peptide from Buthus occitanus tunetanus. |
| ROS | Reactive oxygen species. |
| S1 / S2 | Designations of Stigmurin analogues. |
| S180 | Murine Sarcoma Cell Line S180. |
| SDS-PAGE | Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis. |
| SEC | Size exclusion chromatography. |
| SEM | Scanning Electron Microscopy. |
| SHG-44 | Human Glioma Cell Line SHG-44. |
| SPECT | Single-Photon Emission Computed Tomography. |
| SPR | Surface plasmon resonance. |
| STAT3 | Transcription factor. |
| STAT | Signal transducer and activator of transcription. |
| SVS-1 | Anticancer Beta-Hairpin Peptide SVS-1. |
| TAMs | Tumor-associated macrophages. |
| tBid | Truncated Bid. |
| TfR | Transferrin receptor. |
| TGF-β | Transforming Growth Factor Beta. |
| Th1 / Th2 / Th17 cells | T helper cell subsets. |
| TIMPs | Tissue Inhibitors of Metalloproteinases. |
| TLR2 | Toll-Like Receptors 2. |
| TLR4 | Toll-Like Receptors 4. |
| TM-601 | Synthetic Chlorotoxin Derivative Radiolabeled with Iodine-131. |
| TME | Tumor Microenvironment. |
| TMT | Tandem mass tags. |
| TNF-α | Tumor Necrosis Factor-alpha. |
| TRAIL | Tumor necrosis factor-related apoptosis-inducing ligand. |
| TRPV1+ | Transient Receptor Potential Vanilloid 1–positive. |
| Ts1 | Major neurotoxin from Tityus serrulatus venom. |
| Tsv | Tityus stigmurus venom. |
| TzII | Tityus zulianus toxin isoforms II. |
| TzIII | Tityus zulianus toxin isoforms III. |
| U87 | Human Glioblastoma Cell Line U87. |
| UHPLC-QTOF-MS | Liquid chromatography coupled to mass spectrometry. |
| VERO | African Green Monkey Kidney Epithelial Cell Line VERO. |
| Vm24 | Peptide from Vaejovis mexicanus. |
| Vmct1 | Vaejovis mexicanus cationic peptide 1. |
| Vmct1-K | Lysine-substituted variant of Vmct1. |
| VRE | Vancomycin-Resistant Enterococcus. |
| α1β1 | Alpha-1 Beta-1 Integrin. |
| αvβ3 | Alpha-V Beta-3 Integrin. |
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2024, 74, 229–263. [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov 2022, 12, 31–46. [CrossRef]
- Mariotto, A.B.; Enewold, L.; Zhao, J.; Zeruto, C.A.; Yabroff, K.R. Medical Care Costs Associated with Cancer Survivorship in the United States. Cancer Epidemiology, Biomarkers & Prevention 2020, 29, 1304–1312. [CrossRef]
- Amjad, M.T.; Chidharla, A.; Kasi, A. Cancer Chemotherapy; 2025;
- Nikolaou, M.; Pavlopoulou, A.; Georgakilas, A.G.; Kyrodimos, E. The Challenge of Drug Resistance in Cancer Treatment: A Current Overview. Clin Exp Metastasis 2018, 35, 309–318. [CrossRef]
- Haider, T.; Pandey, V.; Banjare, N.; Gupta, P.N.; Soni, V. Drug Resistance in Cancer: Mechanisms and Tackling Strategies. Pharmacological Reports 2020, 72, 1125–1151. [CrossRef]
- Emran, T. Bin; Shahriar, A.; Mahmud, A.R.; Rahman, T.; Abir, M.H.; Siddiquee, Mohd.F.-R.; Ahmed, H.; Rahman, N.; Nainu, F.; Wahyudin, E.; et al. Multidrug Resistance in Cancer: Understanding Molecular Mechanisms, Immunoprevention and Therapeutic Approaches. Front Oncol 2022, 12. [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [CrossRef]
- Arvanitis, C.D.; Ferraro, G.B.; Jain, R.K. The Blood–Brain Barrier and Blood–Tumour Barrier in Brain Tumours and Metastases. Nat Rev Cancer 2020, 20, 26–41. [CrossRef]
- Bérard, C.; Truillet, C.; Larrat, B.; Dhermain, F.; Estève, M.-A.; Correard, F.; Novell, A. Anticancer Drug Delivery by Focused Ultrasound-Mediated Blood-Brain/Tumor Barrier Disruption for Glioma Therapy: From Benchside to Bedside. Pharmacol Ther 2023, 250, 108518. [CrossRef]
- Kato, R.; Zhang, L.; Kinatukara, N.; Huang, R.; Asthana, A.; Weber, C.; Xia, M.; Xu, X.; Shah, P. Investigating Blood–Brain Barrier Penetration and Neurotoxicity of Natural Products for Central Nervous System Drug Development. Sci Rep 2025, 15, 7431. [CrossRef]
- Shah, B.; Dong, X. Current Status of In Vitro Models of the Blood-Brain Barrier. Curr Drug Deliv 2022, 19, 1034–1046. [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J Nat Prod 2020, 83, 770–803. [CrossRef]
- Bordon, K. de C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front Pharmacol 2020, 11. [CrossRef]
- Mohamed Abd El-Aziz, T.; Soares, A.G.; Stockand, J.D. Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins (Basel) 2019, 11, 564. [CrossRef]
- Freuville, L.; Matthys, C.; Quinton, L.; Gillet, J.-P. Venom-Derived Peptides for Breaking through the Glass Ceiling of Drug Development. Front Chem 2024, 12. [CrossRef]
- Mirzaei, S.; Fekri, H.S.; Hashemi, F.; Hushmandi, K.; Mohammadinejad, R.; Ashrafizadeh, M.; Zarrabi, A.; Garg, M. Venom Peptides in Cancer Therapy: An Updated Review on Cellular and Molecular Aspects. Pharmacol Res 2021, 164, 105327. [CrossRef]
- Prashanth, J.R.; Hasaballah, N.; Vetter, I. Pharmacological Screening Technologies for Venom Peptide Discovery. Neuropharmacology 2017, 127, 4–19. [CrossRef]
- King, G.F. Venoms as a Platform for Human Drugs: Translating Toxins into Therapeutics. Expert Opin Biol Ther 2011, 11, 1469–1484. [CrossRef]
- Lewis, R.J.; Garcia, M.L. Therapeutic Potential of Venom Peptides. Nat Rev Drug Discov 2003, 2, 790–802. [CrossRef]
- Ferreira, S.H. A Bradykinin-Potentiating Factor (Bpf) Present in Venom of Bothrops Jararaca. Br J Pharmacol Chemother 1965, 24, 163-.
- Camargo, A.C.M.; Ianzer, D.; Guerreiro, J.R.; Serrano, S.M.T. Bradykinin-Potentiating Peptides: Beyond Captopril. Toxicon 2012, 59, 516–523. [CrossRef]
- Phillips, D.R.; Scarborough, R.M. Clinical Pharmacology of Eptifibatide. American Journal of Cardiology 1997, 80, B11–B20. [CrossRef]
- McIntosh, M.; Cruz, L.J.; Hunkapiller, M.W.; Gray, W.R.; Olivera, B.M. Isolation and Structure of a Peptide Toxin from the Marine Snail Conus Magus. Arch Biochem Biophys 1982, 218, 329–334. [CrossRef]
- Eng, J.; Kleinman, W.A.; Singh, L.; Singh, G.; Raufman, J.P. Isolation and Characterization of Exendin-4, an Exendin-3 Analogue, from Heloderma Suspectum Venom. Further Evidence for an Exendin Receptor on Dispersed Acini from Guinea Pig Pancreas. Journal of Biological Chemistry 1992, 267, 7402–7405.
- Ahmadi, S.; Knerr, J.M.; Argemi, L.; Bordon, K.C.F.; Pucca, M.B.; Cerni, F.A.; Arantes, E.C.; Çalışkan, F.; Laustsen, A.H. Scorpion Venom: Detriments and Benefits. Biomedicines 2020, 8, 118. [CrossRef]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion Venom Components as Potential Candidates for Drug Development. Toxicon 2015, 93, 125–135. [CrossRef]
- Al-Asmari, A.; Ullah, Z.; Al Balowi, A.; Islam, M. In Vitro Determination of the Efficacy of Scorpion Venoms as Anti-Cancer Agents against Colorectal Cancer Cells: A Nano-Liposomal Delivery Approach. Int J Nanomedicine 2017, Volume 12, 559–574. [CrossRef]
- Montero-Dominguez, P.A.; Corzo, G. Characterization of the Coupling Mechanism of Scorpion β-Neurotoxins on the Voltage-Gated Sodium Channel HNav1.6. J Biomol Struct Dyn 2023, 41, 14419–14427. [CrossRef]
- Abdel-Rahman, M.A.; Quintero-Hernández, V.; Possani, L.D. Scorpion Venom Gland Transcriptomics and Proteomics: An Overview. In Venom Genomics and Proteomics; Springer Netherlands: Dordrecht, 2014; pp. 1–17.
- Marchi, F.C.; Mendes-Silva, E.; Rodrigues-Ribeiro, L.; Bolais-Ramos, L.G.; Verano-Braga, T. Toxinology in the Proteomics Era: A Review on Arachnid Venom Proteomics. Journal of Venomous Animals and Toxins including Tropical Diseases 2022, 28. [CrossRef]
- Al-Asmari, A.K.; Riyasdeen, A.; Islam, M. Scorpion Venom Causes Upregulation of P53 and Downregulation of Bcl-x L and BID Protein Expression by Modulating Signaling Proteins Erk 1/2 and STAT3, and DNA Damage in Breast and Colorectal Cancer Cell Lines. Integr Cancer Ther 2018, 17, 271–281. [CrossRef]
- Du, Q.; Hou, X.; Ge, L.; Li, R.; Zhou, M.; Wang, H.; Wang, L.; Wei, M.; Chen, T.; Shaw, C. Cationicity-Enhanced Analogues of the Antimicrobial Peptides, AcrAP1 and AcrAP2, from the Venom of the Scorpion, Androctonus crassicauda, Display Potent Growth Modulation Effects on Human Cancer Cell Lines. Int J Biol Sci 2014, 10, 1097–1107. [CrossRef]
- Pedron, C.N.; de Oliveira, C.S.; da Silva, A.F.; Andrade, G.P.; da Silva Pinhal, M.A.; Cerchiaro, G.; da Silva Junior, P.I.; da Silva, F.D.; Torres, M.D.T.; Oliveira, V.X. The Effect of Lysine Substitutions in the Biological Activities of the Scorpion Venom Peptide VmCT1. European Journal of Pharmaceutical Sciences 2019, 136, 104952. [CrossRef]
- Richard, S.A.; Kampo, S.; Sackey, M.; Hechavarria, M.E.; Buunaaim, A.D.B. The Pivotal Potentials of Scorpion Buthus Martensii Karsch- Analgesic-Antitumor Peptide in Pain Management and Cancer. Evidence-Based Complementary and Alternative Medicine 2020, 2020. [CrossRef]
- Shao, J.-H.; Cui, Y.; Zhao, M.-Y.; Wu, C.-F.; Liu, Y.-F.; Zhang, J.-H. Purification, Characterization, and Bioactivity of a New Analgesic-Antitumor Peptide from Chinese Scorpion Buthus Martensii Karsch. Peptides (N.Y.) 2014, 53, 89–96. [CrossRef]
- Zhao, Y.; Cai, X.; Ye, T.; Huo, J.; Liu, C.; Zhang, S.; Cao, P. Analgesic-Antitumor Peptide Inhibits Proliferation and Migration of SHG-44 Human Malignant Glioma Cells. J Cell Biochem 2011, 112, 2424–2434. [CrossRef]
- Das Gupta, S.; Halder, B.; Gomes, A.; Gomes, A. Bengalin Initiates Autophagic Cell Death through ERK–MAPK Pathway Following Suppression of Apoptosis in Human Leukemic U937 Cells. Life Sci 2013, 93, 271–276. [CrossRef]
- Gupta, S. Das; Gomes, A.; Debnath, A.; Saha, A.; Gomes, A. Apoptosis Induction in Human Leukemic Cells by a Novel Protein Bengalin, Isolated from Indian Black Scorpion Venom: Through Mitochondrial Pathway and Inhibition of Heat Shock Proteins. Chem Biol Interact 2010, 183, 293–303. [CrossRef]
- Ma, R.; Mahadevappa, R.; Kwok, H.F. Venom-Based Peptide Therapy: Insights into Anti-Cancer Mechanism. Oncotarget 2017, 8, 100908–100930. [CrossRef]
- Baradaran, M.; Pashmforoosh, N. Peptides with Diverse Functions from Scorpion Venom: A Great Opportunity for the Treatment of a Wide Variety of Diseases. The Payam-e-Marefat-Kabul Education University 2023, 27, 84–99. [CrossRef]
- Sarfo-Poku, C.; Eshun, O.; Lee, K.H. Medical Application of Scorpion Venom to Breast Cancer: A Mini-Review. Toxicon 2016, 122, 109–112. [CrossRef]
- Gao, F.; Li, H.; Chen, Y.-D.; Yu, X.-N.; Wang, R.; Chen, X.-L. Upregulation of PTEN Involved in Scorpion Venom-Induced Apoptosis in a Lymphoma Cell Line. Leuk Lymphoma 2009, 50, 633–641. [CrossRef]
- Rapôso, C. Scorpion and Spider Venoms in Cancer Treatment: State of the Art, Challenges, and Perspectives. J Clin Transl Res 2017, 3, 233–249.
- Panja, K.; Buranapraditkun, S.; Roytrakul, S.; Kovitvadhi, A.; Lertwatcharasarakul, P.; Nakagawa, T.; Limmanont, C.; Jaroensong, T. Scorpion Venom Peptide Effects on Inhibiting Proliferation and Inducing Apoptosis in Canine Mammary Gland Tumor Cell Lines. Animals 2021, 11, 2119. [CrossRef]
- Satitmanwiwat, S.; Changsangfa, C.; Khanuengthong, A.; Promthep, K.; Roytrakul, S.; Arpornsuwan, T.; Saikhun, K.; Sritanaudomchai, H. The Scorpion Venom Peptide BmKn2 Induces Apoptosis in Cancerous but Not in Normal Human Oral Cells. Biomedicine & Pharmacotherapy 2016, 84, 1042–1050. [CrossRef]
- Cheng, Y.; Zhao, J.; Qiao, W.; Chen, K. Recent Advances in Diagnosis and Treatment of Gliomas Using Chlorotoxin-Based Bioconjugates. Am J Nucl Med Mol Imaging 2014, 4, 385–405.
- Jlassi, A.; Mekni-Toujani, M.; Ferchichi, A.; Gharsallah, C.; Malosse, C.; Chamot-Rooke, J.; ElAyeb, M.; Ghram, A.; Srairi-Abid, N.; Daoud, S. BotCl, the First Chlorotoxin-Like Peptide Inhibiting Newcastle Disease Virus: The Emergence of a New Scorpion Venom AMPs Family. Molecules 2023, 28, 4355. [CrossRef]
- Dardevet, L.; Rani, D.; Aziz, T.A. El; Bazin, I.; Sabatier, J.-M.; Fadl, M.; Brambilla, E.; De Waard, M. Chlorotoxin: A Helpful Natural Scorpion Peptide to Diagnose Glioma and Fight Tumor Invasion. Toxins (Basel) 2015, 7, 1079–1101. [CrossRef]
- Cohen-Inbar, O.; Zaaroor, M. Glioblastoma Multiforme Targeted Therapy: The Chlorotoxin Story. Journal of Clinical Neuroscience 2016, 33, 52–58. [CrossRef]
- Naseem, M.U.; Carcamo-Noriega, E.; Beltrán-Vidal, J.; Borrego, J.; Szanto, T.G.; Zamudio, F.Z.; Delgado-Prudencio, G.; Possani, L.D.; Panyi, G. Cm28, a Scorpion Toxin Having a Unique Primary Structure, Inhibits KV1.2 and KV1.3 with High Affinity. Journal of General Physiology 2022, 154. [CrossRef]
- Ghezellou, P.; Jakob, K.; Atashi, J.; Ghassempour, A.; Spengler, B. Mass-Spectrometry-Based Lipidome and Proteome Profiling of Hottentotta saulcyi (Scorpiones: Buthidae) Venom. Toxins (Basel) 2022, 14, 370. [CrossRef]
- Nosouhian, M.; Rastegari, A.A.; Shahanipour, K.; Ahadi, A.M.; Sajjadieh, M.S. Anticancer Potentiality of Hottentotta saulcyi Scorpion Curd Venom against Breast Cancer: An in Vitro and in Vivo Study. Sci Rep 2024, 14, 24607. [CrossRef]
- Li, B.; Lyu, P.; Xi, X.; Ge, L.; Mahadevappa, R.; Shaw, C.; Kwok, H.F. Triggering of Cancer Cell Cycle Arrest by a Novel Scorpion Venom-derived Peptide—Gonearrestide. J Cell Mol Med 2018, 22, 4460–4473. [CrossRef]
- Moradi, M.; Najafi, R.; Amini, R.; Solgi, R.; Tanzadehpanah, H.; Esfahani, A.M.; Saidijam, M. Remarkable Apoptotic Pathway of Hemiscorpius lepturus Scorpion Venom on CT26 Cell Line. Cell Biol Toxicol 2019, 35, 373–385. [CrossRef]
- LEBRUN, B.; ROMI-LEBRUN, R.; MARTIN-EAUCLAIRE, M.-F.; YASUDA, A.; ISHIGURO, M.; OYAMA, Y.; PONGS, O.; NAKAJIMA, T. A Four-Disulphide-Bridged Toxin, with High Affinity towards Voltage-Gated K+ Channels, Isolated from Heterometrus spinnifer (Scorpionidae) Venom. Biochemical Journal 1997, 328, 321–327. [CrossRef]
- Candia, S.; Garcia, M.L.; Latorre, R. Mode of Action of Iberiotoxin, a Potent Blocker of the Large Conductance Ca(2+)-Activated K+ Channel. Biophys J 1992, 63, 583–590. [CrossRef]
- Roger, S.; Potier, M.; Vandier, C.; Le Guennec, J.-Y.; Besson, P. Description and Role in Proliferation of Iberiotoxin-Sensitive Currents in Different Human Mammary Epithelial Normal and Cancerous Cells. Biochim Biophys Acta 2004, 1667, 190–199. [CrossRef]
- Aissaoui, D.; Mlayah-Bellalouna, S.; Jebali, J.; Abdelkafi-Koubaa, Z.; Souid, S.; Moslah, W.; Othman, H.; Luis, J.; ElAyeb, M.; Marrakchi, N.; et al. Functional Role of Kv1.1 and Kv1.3 Channels in the Neoplastic Progression Steps of Three Cancer Cell Lines, Elucidated by Scorpion Peptides. Int J Biol Macromol 2018, 111, 1146–1155. [CrossRef]
- Koschak, A.; Koch, R.O.; Liu, J.; Kaczorowski, G.J.; Reinhart, P.H.; Garcia, M.L.; Knaus, H.-G. [ 125 I]Iberiotoxin-D19Y/Y36F, the First Selective, High Specific Activity Radioligand for High-Conductance Calcium-Activated Potassium Channels. Biochemistry 1997, 36, 1943–1952. [CrossRef]
- Nardi, A.; Calderone, V.; Chericoni, S.; Morelli, I. Natural Modulators of Large-Conductance Calcium-Activated Potassium Channels. Planta Med 2003, 69, 885–892. [CrossRef]
- Jang, S.H.; Choi, S.Y.; Ryu, P.D.; Lee, S.Y. Anti-Proliferative Effect of Kv1.3 Blockers in A549 Human Lung Adenocarcinoma in Vitro and in Vivo. Eur J Pharmacol 2011, 651, 26–32. [CrossRef]
- Boisseau, S.; Mabrouk, K.; Ram, N.; Garmy, N.; Collin, V.; Tadmouri, A.; Mikati, M.; Sabatier, J.-M.; Ronjat, M.; Fantini, J.; et al. Cell Penetration Properties of Maurocalcine, a Natural Venom Peptide Active on the Intracellular Ryanodine Receptor. Biochimica et Biophysica Acta (BBA) - Biomembranes 2006, 1758, 308–319. [CrossRef]
- Aroui, S.; Ram, N.; Appaix, F.; Ronjat, M.; Kenani, A.; Pirollet, F.; De Waard, M. Maurocalcine as a Non Toxic Drug Carrier Overcomes Doxorubicin Resistance in the Cancer Cell Line MDA-MB 231. Pharm Res 2009, 26, 836–845. [CrossRef]
- Aroui, S.; Dardevet, L.; Najlaoui, F.; Kammoun, M.; Laajimi, A.; Fetoui, H.; De Waard, M.; Kenani, A. PTEN-Regulated AKT/FoxO3a/Bim Signaling Contributes to Human Cell Glioblastoma Apoptosis by Platinum-Maurocalcin Conjugate. Int J Biochem Cell Biol 2016, 77, 15–22. [CrossRef]
- Ram, N.; Aroui, S.; Jaumain, E.; Bichraoui, H.; Mabrouk, K.; Ronjat, M.; Lortat-Jacob, H.; De Waard, M. Direct Peptide Interaction with Surface Glycosaminoglycans Contributes to the Cell Penetration of Maurocalcine. J Biol Chem 2008, 283, 24274–24284. [CrossRef]
- D’Suze, G.; Rosales, A.; Salazar, V.; Sevcik, C. Apoptogenic Peptides from Tityus discrepans Scorpion Venom Acting against the SKBR3 Breast Cancer Cell Line. Toxicon 2010, 56, 1497–1505. [CrossRef]
- Zeng, X.-C.; Zhou, L.; Shi, W.; Luo, X.; Zhang, L.; Nie, Y.; Wang, J.; Wu, S.; Cao, B.; Cao, H. Three New Antimicrobial Peptides from the Scorpion Pandinus Imperator. Peptides (N.Y.) 2013, 45, 28–34. [CrossRef]
- Crusca, E.; Basso, L.G.M.; Altei, W.F.; Marchetto, R. Biophysical Characterization and Antitumor Activity of Synthetic Pantinin Peptides from Scorpion’s Venom. Biochimica et Biophysica Acta (BBA) - Biomembranes 2018, 1860, 2155–2165. [CrossRef]
- Elrayess, R.A.; Mohallal, M.E.; Mobarak, Y.M.; Ebaid, H.M.; Haywood-Small, S.; Miller, K.; Strong, P.N.; Abdel-Rahman, M.A. Scorpion Venom Antimicrobial Peptides Induce Caspase-1 Dependant Pyroptotic Cell Death. Front Pharmacol 2022, 12. [CrossRef]
- Yu, Z.; Han, D.; Zhang, K.; Gai, C.; Chai, X.; Zhao, Q.; Zou, Y.; Yin, L. Optimization of Antimicrobial Peptide Smp43 as Novel Inhibitors of Cancer. Bioorg Chem 2025, 161, 108561. [CrossRef]
- Deng, Z.; Gao, Y.; Nguyen, T.; Chai, J.; Wu, J.; Li, J.; Abdel-Rahman, M.A.; Xu, X.; Chen, X. The Potent Antitumor Activity of Smp43 against Non-Small-Cell Lung Cancer A549 Cells via Inducing Membranolysis and Mitochondrial Dysfunction. Toxins (Basel) 2023, 15, 347. [CrossRef]
- Chai, J.; Yang, W.; Gao, Y.; Guo, R.; Peng, Q.; Abdel-Rahman, M.A.; Xu, X. Antitumor Effects of Scorpion Peptide Smp43 through Mitochondrial Dysfunction and Membrane Disruption on Hepatocellular Carcinoma. J Nat Prod 2021, 84, 3147–3160. [CrossRef]
- Nguyen, T.; Guo, R.; Chai, J.; Wu, J.; Liu, J.; Chen, X.; Abdel-Rahman, M.A.; Xia, H.; Xu, X. Smp24, a Scorpion-Venom Peptide, Exhibits Potent Antitumor Effects against Hepatoma HepG2 Cells via Multi-Mechanisms In Vivo and In Vitro. Toxins (Basel) 2022, 14, 717. [CrossRef]
- Guo, R.; Liu, J.; Chai, J.; Gao, Y.; Abdel-Rahman, M.A.; Xu, X. Scorpion Peptide Smp24 Exhibits a Potent Antitumor Effect on Human Lung Cancer Cells by Damaging the Membrane and Cytoskeleton In Vivo and In Vitro. Toxins (Basel) 2022, 14, 438. [CrossRef]
- Rates, B.; Ferraz, K.K.F.; Borges, M.H.; Richardson, M.; De Lima, M.E.; Pimenta, A.M.C. Tityus serrulatus Venom Peptidomics: Assessing Venom Peptide Diversity. Toxicon 2008, 52, 611–618. [CrossRef]
- Guo, X.; Ma, C.; Du, Q.; Wei, R.; Wang, L.; Zhou, M.; Chen, T.; Shaw, C. Two Peptides, TsAP-1 and TsAP-2, from the Venom of the Brazilian Yellow Scorpion, Tityus serrulatus: Evaluation of Their Antimicrobial and Anticancer Activities. Biochimie 2013, 95, 1784–1794. [CrossRef]
- Daniele-Silva, A.; Machado, R.J.A.; Monteiro, N.K. V; Estrela, A.B.; Santos, E.C.G.; Carvalho, E.; Araújo Júnior, R.F.; Melo-Silveira, R.F.; Rocha, H.A.O.; Silva-Júnior, A.A.; et al. Stigmurin and TsAP-2 from Tityus stigmurus Scorpion Venom: Assessment of Structure and Therapeutic Potential in Experimental Sepsis. Toxicon 2016, 121, 10–21. [CrossRef]
- Varga, Z.; Gurrola-Briones, G.; Papp, F.; Rodríguez de la Vega, R.C.; Pedraza-Alva, G.; Tajhya, R.B.; Gaspar, R.; Cardenas, L.; Rosenstein, Y.; Beeton, C.; et al. Vm24, a Natural Immunosuppressive Peptide, Potently and Selectively Blocks Kv1.3 Potassium Channels of Human T Cells. Mol Pharmacol 2012, 82, 372–382. [CrossRef]
- Cid-Uribe, J.I.; Veytia-Bucheli, J.I.; Romero-Gutierrez, T.; Ortiz, E.; Possani, L.D. Scorpion Venomics: A 2019 Overview. Expert Rev Proteomics 2020, 17, 67–83. [CrossRef]
- Santibáñez-López, C.; Cid-Uribe, J.; Batista, C.; Ortiz, E.; Possani, L. Venom Gland Transcriptomic and Proteomic Analyses of the Enigmatic Scorpion Superstitionia donensis (Scorpiones: Superstitioniidae), with Insights on the Evolution of Its Venom Components. Toxins (Basel) 2016, 8, 367. [CrossRef]
- So, W.L.; Leung, T.C.N.; Nong, W.; Bendena, W.G.; Ngai, S.M.; Hui, J.H.L. Transcriptomic and Proteomic Analyses of Venom Glands from Scorpions Liocheles australasiae, Mesobuthus martensii, and Scorpio Maurus palmatus. Peptides (N.Y.) 2021, 146, 170643. [CrossRef]
- Delgado-Prudencio, G.; Cid-Uribe, J.I.; Morales, J.A.; Possani, L.D.; Ortiz, E.; Romero-Gutiérrez, T. The Enzymatic Core of Scorpion Venoms. Toxins (Basel) 2022, 14, 248. [CrossRef]
- Zeng, X.-C.; Luo, F.; Li, W.-X. Molecular Dissection of Venom from Chinese Scorpion Mesobuthus Martensii: Identification and Characterization of Four Novel Disulfide-Bridged Venom Peptides. Peptides (N.Y.) 2006, 27, 1745–1754. [CrossRef]
- Verano-Braga, T.; Dutra, A.A.A.; León, I.R.; Melo-Braga, M.N.; Roepstorff, P.; Pimenta, A.M.C.; Kjeldsen, F. Moving Pieces in a Venomic Puzzle: Unveiling Post-Translationally Modified Toxins from Tityus serrulatus. J Proteome Res 2013, 12, 3460–3470. [CrossRef]
- Melo-Braga, M.N. de; Moreira, R. da S.; Gervásio, J.H.D.B.; Felicori, L.F. Overview of Protein Posttranslational Modifications in Arthropoda Venoms. Journal of Venomous Animals and Toxins including Tropical Diseases 2022, 28. [CrossRef]
- Romey, G.; Chicheportiche, R.; Lazdunski, M.; Rochat, H.; Miranda, F.; Lissitzky, S. Scorpion Neurotoxin — A Presynaptic Toxin Which Affects Both Na+ and K+ Channels in Axons. Biochem Biophys Res Commun 1975, 64, 115–121. [CrossRef]
- Romey, G.; Abita, J.P.; Chicheportiche, R.; Rochat, H.; Lazdunski, M. Scorpion Neurotoxin. Biochimica et Biophysica Acta (BBA) - Biomembranes 1976, 448, 607–619. [CrossRef]
- Catterall, W.A. Neurotoxins That Act on Voltage-Sensitive Sodium Channels in Excitable Membranes. Annu Rev Pharmacol Toxicol 1980, 20, 15–43. [CrossRef]
- Gurevitz, M.; Froy, O.; Zilberberg, N.; Turkov, M.; Strugatsky, D.; Gershburg, E.; Lee, D.; Adams, M.E.; Tugarinov, V.; Anglister, J.; et al. Sodium Channel Modifiers from Scorpion Venom: Structure–Activity Relationship, Mode of Action and Application. Toxicon 1998, 36, 1671–1682. [CrossRef]
- Cestèle, S.; Yarov-Yarovoy, V.; Qu, Y.; Sampieri, F.; Scheuer, T.; Catterall, W.A. Structure and Function of the Voltage Sensor of Sodium Channels Probed by a β-Scorpion Toxin. Journal of Biological Chemistry 2006, 281, 21332–21344. [CrossRef]
- Catterall, W.A.; Cestèle, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-Gated Ion Channels and Gating Modifier Toxins. Toxicon 2007, 49, 124–141. [CrossRef]
- Kozminsky-Atias, A.; Somech, E.; Zilberberg, N. Isolation of the First Toxin from the Scorpion Buthus Occitanus Israelis Showing Preference for Shaker Potassium Channels. FEBS Lett 2007, 581, 2478–2484. [CrossRef]
- Quintero-Hernández, V.; Jiménez-Vargas, J.M.; Gurrola, G.B.; Valdivia, H.H.; Possani, L.D. Scorpion Venom Components That Affect Ion-Channels Function. Toxicon 2013, 76, 328–342. [CrossRef]
- Housley, D.M.; Housley, G.D.; Liddell, M.J.; Jennings, E.A. Scorpion Toxin Peptide Action at the Ion Channel Subunit Level. Neuropharmacology 2017, 127, 46–78. [CrossRef]
- Zou, X.; Wu, Y.; Chen, J.; Zhao, F.; Zhang, F.; Yu, B.; Cao, Z. Activation of Sodium Channel by a Novel α-Scorpion Toxin, BmK NT2, Stimulates ERK1/2 and CERB Phosphorylation through a Ca2+ Dependent Pathway in Neocortical Neurons. Int J Biol Macromol 2017, 104, 70–77. [CrossRef]
- Beraldo Neto, E.; Freitas, L.A. de; Pimenta, D.C.; Lebrun, I.; Nencioni, A.L.A. Tb1, a Neurotoxin from Tityus bahiensis Scorpion Venom, Induces Epileptic Seizures by Increasing Glutamate Release. Toxins (Basel) 2020, 12, 65. [CrossRef]
- Díaz-García, A.; Varela, D. Voltage-Gated K+/Na+ Channels and Scorpion Venom Toxins in Cancer. Front Pharmacol 2020, 11. [CrossRef]
- Wiezel, G.A.; Oliveira, I.S.; Reis, M.B.; Ferreira, I.G.; Cordeiro, K.R.; Bordon, K.C.F.; Arantes, E.C. The Complex Repertoire of Tityus Spp. Venoms: Advances on Their Composition and Pharmacological Potential of Their Toxins. Biochimie 2024, 220, 144–166. [CrossRef]
- Ali, S.; Alam, M.; Abbasi, A.; Undheim, E.; Fry, B.; Kalbacher, H.; Voelter, W. Structure-Activity Relationship of Chlorotoxin-Like Peptides. Toxins (Basel) 2016, 8, 36. [CrossRef]
- Xu, T.; Fan, Z.; Li, W.; Dietel, B.; Wu, Y.; Beckmann, M.W.; Wrosch, J.K.; Buchfelder, M.; Eyupoglu, I.Y.; Cao, Z.; et al. Identification of Two Novel Chlorotoxin Derivatives CA4 and CTX-23 with Chemotherapeutic and Anti-Angiogenic Potential. Sci Rep 2016, 6, 19799. [CrossRef]
- Majc, B.; Novak, M.; Lah, T.T.; Križaj, I. Bioactive Peptides from Venoms against Glioma Progression. Front Oncol 2022, 12. [CrossRef]
- Samat, R.; Sen, S.; Jash, M.; Ghosh, S.; Garg, S.; Sarkar, J.; Ghosh, S. Venom: A Promising Avenue for Antimicrobial Therapeutics. ACS Infect Dis 2024, 10, 3098–3125. [CrossRef]
- Harrison, P.L.; Heath, G.R.; Johnson, B.R.G.; Abdel-Rahman, M.A.; Strong, P.N.; Evans, S.D.; Miller, K. Phospholipid Dependent Mechanism of Smp24, an α-Helical Antimicrobial Peptide from Scorpion Venom. Biochimica et Biophysica Acta (BBA) - Biomembranes 2016, 1858, 2737–2744. [CrossRef]
- Harrison, P.L.; Abdel-Rahman, M.A.; Strong, P.N.; Tawfik, M.M.; Miller, K. Characterisation of Three Alpha-Helical Antimicrobial Peptides from the Venom of Scorpio maurus palmatus. Toxicon 2016, 117, 30–36. [CrossRef]
- Xia, Z.; He, D.; Wu, Y.; Kwok, H.F.; Cao, Z. Scorpion Venom Peptides: Molecular Diversity, Structural Characteristics, and Therapeutic Use from Channelopathies to Viral Infections and Cancers. Pharmacol Res 2023, 197, 106978. [CrossRef]
- Chen, Z.; Wang, B.; Hu, J.; Yang, W.; Cao, Z.; Zhuo, R.; Li, W.; Wu, Y. SjAPI, the First Functionally Characterized Ascaris-Type Protease Inhibitor from Animal Venoms. PLoS One 2013, 8, e57529–e57529. [CrossRef]
- Zhao, R.; Dai, H.; Qiu, S.; Li, T.; He, Y.; Ma, Y.; Chen, Z.; Wu, Y.; Li, W.; Cao, Z. SdPI, The First Functionally Characterized Kunitz-Type Trypsin Inhibitor from Scorpion Venom. PLoS One 2011, 6, e27548–e27548. [CrossRef]
- Song, Y.; Gong, K.; Yan, H.; Hong, W.; Wang, L.; Wu, Y.; Li, W.; Li, W.; Cao, Z. Sj7170, a Unique Dual-Function Peptide with a Specific α-Chymotrypsin Inhibitory Activity and a Potent Tumor-Activating Effect from Scorpion Venom. Journal of Biological Chemistry 2014, 289, 11667–11680. [CrossRef]
- Ding, L.; Wang, X.; Liu, H.; San, M.; Xu, Y.; Li, J.; Li, S.; Cao, Z.; Li, W.; Wu, Y.; et al. A New Kunitz-Type Plasmin Inhibitor from Scorpion Venom. Toxicon 2015, 106, 7–13. [CrossRef]
- Liu, H.; Chen, J.; Wang, X.; Yan, S.; Xu, Y.; San, M.; Tang, W.; Yang, F.; Cao, Z.; Li, W.; et al. Functional Characterization of a New Non-Kunitz Serine Protease Inhibitor from the Scorpion Lychas mucronatus. Int J Biol Macromol 2015, 72, 158–162. [CrossRef]
- Hakim, M.A.; Yang, S. Discoveries of Serine Protease Inhibitors from Scorpions. J Proteomics Bioinform 2016, 04. [CrossRef]
- Petricevich, V.L. Scorpion Venom and the Inflammatory Response. Mediators Inflamm 2010, 2010, 1–16. [CrossRef]
- El-Benna, J.; Hurtado-Nedelec, M.; Gougerot-Pocidalo, M.-A.; Dang, P.M.-C. Effects of Venoms on Neutrophil Respiratory Burst: A Major Inflammatory Function. Journal of Venomous Animals and Toxins including Tropical Diseases 2021, 27. [CrossRef]
- Soltan-Alinejad, P.; Alipour, H.; Soltani, A.; Asgari, Q.; Ramezani, A.; Mehrabani, D.; Azizi, K. Molecular Characterization and In Silico Analyses of Maurolipin Structure as a Secretory Phospholipase A2 (sPLA) from Venom Glands of Iranian Scorpio maurus (Arachnida: Scorpionida). J Trop Med 2022, 2022, 1–11. [CrossRef]
- Baradaran, M.; Salabi, F. Genome-Wide Identification, Structural Homology Analysis, and Evolutionary Diversification of the Phospholipase D Gene Family in the Venom Gland of Three Scorpion Species. BMC Genomics 2023, 24, 730. [CrossRef]
- Salabi, F.; Jafari, H. Whole Transcriptome Sequencing Reveals the Activity of the PLA2 Family Members in Androctonus Crassicauda (Scorpionida: Buthidae) Venom Gland. The FASEB Journal 2024, 38. [CrossRef]
- Díaz, C.; Lomonte, B.; Chang-Castillo, A.; Bonilla, F.; Alfaro-Chinchilla, A.; Triana, F.; Angulo, D.; Fernández, J.; Sasa, M. Venomics of Scorpion Ananteris platnicki (Lourenço, 1993), a New World Buthid That Inhabits Costa Rica and Panama. Toxins (Basel) 2024, 16, 327. [CrossRef]
- Mendoza-Tobar, L.L.; Clement, H.; Arenas, I.; Sepulveda-Arias, J.C.; Vargas, J.A.G.; Corzo, G. An Overview of Some Enzymes from Buthid Scorpion Venoms from Colombia: Centruroides margaritatus, Tityus pachyurus, and Tityus n. Sp. Aff. Metuendus. Journal of Venomous Animals and Toxins including Tropical Diseases 2024, 30. [CrossRef]
- Possani, L. Peptides and Genes Coding for Scorpion Toxins That Affect Ion-Channels. Biochimie 2000, 82, 861–868. [CrossRef]
- Li, Z.; Hu, P.; Wu, W.; Wang, Y. Peptides with Therapeutic Potential in the Venom of the Scorpion Buthus Martensii Karsch. Peptides (N.Y.) 2019, 115, 43–50. [CrossRef]
- Mikaelian, A.G.; Traboulay, E.; Zhang, X.M.; Yeritsyan, E.; Pedersen, P.L.; Ko, Y.H.; Matalka, K.Z. Pleiotropic Anticancer Properties of Scorpion Venom Peptides: Rhopalurus princeps Venom as an Anticancer Agent. Drug Des Devel Ther 2020, Volume 14, 881–893. [CrossRef]
- Aissaoui-Zid, D.; Saada, M.-C.; Moslah, W.; Potier-Cartereau, M.; Lemettre, A.; Othman, H.; Gaysinski, M.; Abdelkafi-Koubaa, Z.; Souid, S.; Marrakchi, N.; et al. AaTs-1: A Tetrapeptide from Androctonus australis Scorpion Venom, Inhibiting U87 Glioblastoma Cells Proliferation by P53 and FPRL-1 Up-Regulations. Molecules 2021, 26. [CrossRef]
- Xu, X.; Xu, H.; Ren, F.; Huang, L.; Xu, J.; Li, F. Protective Effect of Scorpion Venom Heat-Resistant Synthetic Peptide against PM2.5-Induced Microglial Polarization via TLR4-Mediated Autophagy Activating PI3K/AKT/NF-ΚB Signaling Pathway. J Neuroimmunol 2021, 355, 577567. [CrossRef]
- You, X.; Wu, Y.; Li, Q.; Sheng, W.; Zhou, Q.; Fu, W. Astragalus–Scorpion Drug Pair Inhibits the Development of Prostate Cancer by Regulating GDPD4-2/PI3K/AKT/MTOR Pathway and Autophagy. Front Pharmacol 2022, 13. [CrossRef]
- Ismail, M.; El-Asmar, M.F.; Osman, O.H. Pharmacological Studies with Scorpion (Palamneus Gravimanus) Venom: Evidence for the Presence of Histamine. Toxicon 1975, 13, 49–56. [CrossRef]
- Basu, A.; Gomes, A.; Gomes, A.; Dasgupta, S.C.; Lahiri, S.C. Histamine, 5-HT & Hyaluronidase in the Venom of the Scorpion Lychas laevifrons (Pock). Indian J Med Res 1990, 92, 371–373.
- Kwon, N.-Y.; Sung, H.-K.; Park, J.-K. Systematic Review of the Antitumor Activities and Mechanisms of Scorpion Venom on Human Breast Cancer Cells Lines (In Vitro Study). J Clin Med 2025, 14, 3181. [CrossRef]
- Evans, E.R.J.; McIntyre, L.; Northfield, T.D.; Daly, N.L.; Wilson, D.T. Small Molecules in the Venom of the Scorpion Hormurus waigiensis. Biomedicines 2020, 8, 259. [CrossRef]
- Ageitos, L.; Torres, M.D.T.; de la Fuente-Nunez, C. Biologically Active Peptides from Venoms: Applications in Antibiotic Resistance, Cancer, and Beyond. Int J Mol Sci 2022, 23. [CrossRef]
- Soares, K.S.R.; Formiga, A.L.D.; Uchôa, A.F.C.; Cardoso, A.L.M.R.; Rodrigues, J.P.C.G.; Leite, J. de P.F.B.; Silva, L.F.A.; Alves, Á.E.F.; Barbosa-Filho, J.M.; Xavier-Junior, F.H. Beyond the Peril of Envenomation: A Nanotechnology Approach for Therapeutic Venom Delivery. J Drug Deliv Sci Technol 2025, 105, 106652. [CrossRef]
- Cajado-Carvalho, D.; da Silva, C.C.F.; Kodama, R.T.; Mariano, D.O.C.; Pimenta, D.C.; Duzzi, B.; Kuniyoshi, A.K.; Portaro, F.V. Purification and Biochemical Characterization of TsMS 3 and TsMS 4: Neuropeptide-Degrading Metallopeptidases in the Tityus serrulatus Venom. Toxins (Basel) 2019, 11, 194. [CrossRef]
- Cajado Carvalho, D.; Kuniyoshi, A.K.; Kodama, R.T.; Oliveira, A.K.; Serrano, S.M.T.; Tambourgi, D. V.; Portaro, F. V. Neuropeptide Y Family-Degrading Metallopeptidases in the Tityus serrulatus Venom Partially Blocked by Commercial Antivenoms. Toxicological Sciences 2014, 142, 418–426. [CrossRef]
- Ma, Y.; Zhao, Y.; Zhao, R.; Zhang, W.; He, Y.; Wu, Y.; Cao, Z.; Guo, L.; Li, W. Molecular Diversity of Toxic Components from the Scorpion Heterometrus Petersii Venom Revealed by Proteomic and Transcriptome Analysis. Proteomics 2010, 10, 2471–2485. [CrossRef]
- Venancio, E.J.; Portaro, F.C. V; Kuniyoshi, A.K.; Carvalho, D.C.; Pidde-Queiroz, G.; Tambourgi, D. V Enzymatic Properties of Venoms from Brazilian Scorpions of Tityus Genus and the Neutralisation Potential of Therapeutical Antivenoms. Toxicon 2013, 69, 180–190. [CrossRef]
- Ding, J.; Chua, P.-J.; Bay, B.-H.; Gopalakrishnakone, P. Scorpion Venoms as a Potential Source of Novel Cancer Therapeutic Compounds. Exp Biol Med 2014, 239, 387–393. [CrossRef]
- Ricci, J.-E.; Gottlieb, R.A.; Green, D.R. Caspase-Mediated Loss of Mitochondrial Function and Generation of Reactive Oxygen Species during Apoptosis. J Cell Biol 2003, 160, 65–75. [CrossRef]
- Roy, S.; Nicholson, D.W. Cross-Talk in Cell Death Signaling. J Exp Med 2000, 192, F21-5.
- Basu, A.; Castle, V.P.; Bouziane, M.; Bhalla, K.; Haldar, S. Crosstalk between Extrinsic and Intrinsic Cell Death Pathways in Pancreatic Cancer: Synergistic Action of Estrogen Metabolite and Ligands of Death Receptor Family. Cancer Res 2006, 66, 4309–4318. [CrossRef]
- Li, H.; Zhu, H.; Xu, C.; Yuan, J. Cleavage of BID by Caspase 8 Mediates the Mitochondrial Damage in the Fas Pathway of Apoptosis. Cell 1998, 94, 491–501. [CrossRef]
- Yin, X.-M.; Wang, K.; Gross, A.; Zhao, Y.; Zinkel, S.; Klocke, B.; Roth, K.A.; Korsmeyer, S.J. Bid-Deficient Mice Are Resistant to Fas-Induced Hepatocellular Apoptosis. Nature 1999, 400, 886–891. [CrossRef]
- Luo, X.; Budihardjo, I.; Zou, H.; Slaughter, C.; Wang, X. Bid, a Bcl2 Interacting Protein, Mediates Cytochrome c Release from Mitochondria in Response to Activation of Cell Surface Death Receptors. Cell 1998, 94, 481–490. [CrossRef]
- Srairi-Abid, N.; Othman, H.; Aissaoui, D.; BenAissa, R. Anti-Tumoral Effect of Scorpion Peptides: Emerging New Cellular Targets and Signaling Pathways. Cell Calcium 2019, 80, 160–174. [CrossRef]
- Gomes, A.; Bhattacharjee, P.; Mishra, R.; Biswas, A.K.; Dasgupta, S.C.; Giri, B. Anticancer Potential of Animal Venoms and Toxins. Indian J Exp Biol 2010, 48, 93–103.
- Kampo, S.; Ahmmed, B.; Zhou, T.; Owusu, L.; Anabah, T.W.; Doudou, N.R.; Kuugbee, E.D.; Cui, Y.; Lu, Z.; Yan, Q.; et al. Scorpion Venom Analgesic Peptide, BmK AGAP Inhibits Stemness, and Epithelial-Mesenchymal Transition by Down-Regulating PTX3 in Breast Cancer. Front Oncol 2019, 9. [CrossRef]
- Dardevet, L.; Najlaoui, F.; Aroui, S.; Collot, M.; Tisseyre, C.; Pennington, M.W.; Mallet, J.-M.; De Waard, M. A Conjugate between Lqh-8/6, a Natural Peptide Analogue of Chlorotoxin, and Doxorubicin Efficiently Induces Glioma Cell Death. Biomedicines 2022, 10, 2605. [CrossRef]
- Yu, M.; Liu, S.; Sun, P.; Pan, H.; Tian, C.; Zhang, L. Peptide Toxins and Small-Molecule Blockers of BK Channels. Acta Pharmacol Sin 2016, 37, 56–66. [CrossRef]
- Giangiacomo, K.M.; Sugg, E.E.; Garcia-Calvo, M.; Leonard, R.J.; McManus, O.B.; Kaczorowski, G.J.; Garcia, M.L. Synthetic Charybdotoxin-Iberiotoxin Chimeric Peptides Define Toxin Binding Sites on Calcium-Activated and Voltage-Dependent Potassium Channels. Biochemistry 1993, 32, 2363–2370. [CrossRef]
- Bingham, J.-P.; Bian, S.; Tan, Z.-Y.; Takacs, Z.; Moczydlowski, E. Synthesis of a Biotin Derivative of Iberiotoxin: Binding Interactions with Streptavidin and the BK Ca 2+ -Activated K + Channel Expressed in a Human Cell Line. Bioconjug Chem 2006, 17, 689–699. [CrossRef]
- Wang, W.-X.; Ji, Y.-H. Scorpion Venom Induces Glioma Cell Apoptosis in Vivo and Inhibits Glioma Tumor Growth in Vitro. J Neurooncol 2005, 73, 1–7. [CrossRef]
- Sui, W.-W.; Zhang, W.-D.; Wu, L.-C.; Zhang, Y.-Y.; Wang, Z.-P.; Wang, Z.-X.; Jia, Q. [Study on the Mechanism of Polypeptide Extract from Scorpion Venom on Inhibition of Angiogenesis of H 22 Hepatoma]. Zhongguo Zhong Xi Yi Jie He Za Zhi 2014, 34, 581–586.
- Zhang, Y.Y.; Wu, L.C.; Wang, Z.P.; Wang, Z.X.; Jia, Q.; Jiang, G.S.; Zhang, W.D. Anti-Proliferation Effect of Polypeptide Extracted from Scorpion Venom on Human Prostate Cancer Cells in Vitro. J Clin Med Res 2009, 1, 24–31. [CrossRef]
- Marak, B.N.; Dowarah, J.; Khiangte, L.; Singh, V.P. A Comprehensive Insight on the Recent Development of Cyclic Dependent Kinase Inhibitors as Anticancer Agents. Eur J Med Chem 2020, 203, 112571. [CrossRef]
- Zeng, J.; Xu, C.; Chen, L.; Li, M.; Yang, J.; Nie, L.; Zhang, M.; Zhang, X.; Zhang, E. Anti-Glioma Effect of Buthus Martensii Karsch (BmK) Scorpion by Inhibiting Myeloid-Derived Suppressor Cells and Activating T Cells in Tumor Microenvironment. J Funct Foods 2024, 116, 106163. [CrossRef]
- El-Qassas, J.; Abd El-Atti, M.; El-Badri, N. Harnessing the Potency of Scorpion Venom-Derived Proteins: Applications in Cancer Therapy. Bioresour Bioprocess 2024, 11, 93. [CrossRef]
- Feng, L.; Gao, R.; Gopalakrishnakone, P. Isolation and Characterization of a Hyaluronidase from the Venom of Chinese Red Scorpion Buthus martensi. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2008, 148, 250–257. [CrossRef]
- Feng, L.; Gao, R.; Meng, J.; Gopalakrishnakone, P. Cloning and Molecular Characterization of BmHYA1, a Novel Hyaluronidase from the Venom of Chinese Red Scorpion Buthus martensi Karsch. Toxicon 2010, 56, 474–479. [CrossRef]
- Xia, X.; Liu, R.; Li, Y.; Xue, S.; Liu, Q.; Jiang, X.; Zhang, W.; Ding, K. Cloning and Molecular Characterization of Scorpion Buthus martensi Venom Hyaluronidases: A Novel Full-Length and Diversiform Noncoding Isoforms. Gene 2014, 547, 338–345. [CrossRef]
- DELPECH, B.; GIRARD, N.; BERTRAND, P.; COUREL, M. -N.; CHAUZY, C.; DELPECH, A. Hyaluronan: Fundamental Principles and Applications in Cancer. J Intern Med 1997, 242, 41–48. [CrossRef]
- Naor, D.; Sionov, R.V.; Ish-Shalom, D. CD44: Structure, Function and Association with the Malignant Process. In; 1997; pp. 241–319.
- Khamessi, O.; Ben Mabrouk, H.; ElFessi-Magouri, R.; Kharrat, R. RK1, the First Very Short Peptide from Buthus occitanus tunetanus Inhibits Tumor Cell Migration, Proliferation and Angiogenesis. Biochem Biophys Res Commun 2018, 499, 1–7. [CrossRef]
- Sinthuvanich, C.; Veiga, A.S.; Gupta, K.; Gaspar, D.; Blumenthal, R.; Schneider, J.P. Anticancer β-Hairpin Peptides: Membrane-Induced Folding Triggers Activity. J Am Chem Soc 2012, 134, 6210–6217. [CrossRef]
- Tolos (Vasii), A.M.; Moisa, C.; Dochia, M.; Popa, C.; Copolovici, L.; Copolovici, D.M. Anticancer Potential of Antimicrobial Peptides: Focus on Buforins. Polymers (Basel) 2024, 16, 728. [CrossRef]
- DeBin, J.A.; Maggio, J.E.; Strichartz, G.R. Purification and Characterization of Chlorotoxin, a Chloride Channel Ligand from the Venom of the Scorpion. American Journal of Physiology-Cell Physiology 1993, 264, C361–C369. [CrossRef]
- Lippens, G.; Najib, J.; Wodak, S.J.; Tartar, A. NMR Sequential Assignments and Solution Structure of Chlorotoxin, a Small Scorpion Toxin That Blocks Chloride Channels. Biochemistry 1995, 34, 13–21. [CrossRef]
- Correnti, C.E.; Gewe, M.M.; Mehlin, C.; Bandaranayake, A.D.; Johnsen, W.A.; Rupert, P.B.; Brusniak, M.-Y.; Clarke, M.; Burke, S.E.; De Van Der Schueren, W.; et al. Screening, Large-Scale Production and Structure-Based Classification of Cystine-Dense Peptides. Nat Struct Mol Biol 2018, 25, 270–278. [CrossRef]
- Soroceanu, L.; Gillespie, Y.; Khazaeli, M.B.; Sontheimer, H. Use of Chlorotoxin for Targeting of Primary Brain Tumors. Cancer Res 1998, 58, 4871–4879.
- Lyons, S.A.; O’Neal, J.; Sontheimer, H. Chlorotoxin, a Scorpion-derived Peptide, Specifically Binds to Gliomas and Tumors of Neuroectodermal Origin. Glia 2002, 39, 162–173. [CrossRef]
- Deshane, J.; Garner, C.C.; Sontheimer, H. Chlorotoxin Inhibits Glioma Cell Invasion via Matrix Metalloproteinase-2. J Biol Chem 2003, 278, 4135–4144. [CrossRef]
- Ullrich, N.; Bordey, A.; Gillespie, G.Y.; Sontheimer, H. Expression of Voltage-Activated Chloride Currents in Acute Slices of Human Gliomas. Neuroscience 1998, 83, 1161–1173. [CrossRef]
- McFerrin, M.B.; Sontheimer, H. A Role for Ion Channels in Glioma Cell Invasion. Neuron Glia Biol 2006, 2, 39–49.
- Dastpeyman, M.; Giacomin, P.; Wilson, D.; Nolan, M.J.; Bansal, P.S.; Daly, N.L. A C-Terminal Fragment of Chlorotoxin Retains Bioactivity and Inhibits Cell Migration. Front Pharmacol 2019, 10, 250. [CrossRef]
- Kesavan, K.; Ratliff, J.; Johnson, E.W.; Dahlberg, W.; Asara, J.M.; Misra, P.; Frangioni, J. V; Jacoby, D.B. Annexin A2 Is a Molecular Target for TM601, a Peptide with Tumor-Targeting and Anti-Angiogenic Effects. Journal of Biological Chemistry 2010, 285, 4366–4374. [CrossRef]
- Farkas, S.; Cioca, D.; Murányi, J.; Hornyák, P.; Brunyánszki, A.; Szekér, P.; Boros, E.; Horváth, P.; Hujber, Z.; Rácz, G.Z.; et al. Chlorotoxin Binds to Both Matrix Metalloproteinase 2 and Neuropilin 1. Journal of Biological Chemistry 2023, 299, 104998. [CrossRef]
- Mabunda, I.G.; Offor, B.C.; Muller, B.; Motadi, L.R.; Piater, L.A. Scorpion Venoms from the Buthidae Family: A Dual Study of Proteomic Composition and Anticancer Potentials. Toxicon 2025, 266, 108542. [CrossRef]
- Wiranowska, M.; Colina, L.O.; Johnson, J.O. Clathrin-Mediated Entry and Cellular Localization of Chlorotoxin in Human Glioma. Cancer Cell Int 2011, 11, 27. [CrossRef]
- Barish, M.E.; Aftabizadeh, M.; Hibbard, J.; Blanchard, M.S.; Ostberg, J.R.; Wagner, J.R.; Manchanda, M.; Paul, J.; Stiller, T.; Aguilar, B.; et al. Chlorotoxin-Directed CAR T Cell Therapy for Recurrent Glioblastoma: Interim Clinical Experience Demonstrating Feasibility and Safety. Cell Rep Med 2025, 6, 102302. [CrossRef]
- Mamelak, A.N.; Jacoby, D.B. Targeted Delivery of Antitumoral Therapy to Glioma and Other Malignancies with Synthetic Chlorotoxin (TM-601). Expert Opin Drug Deliv 2007, 4, 175–186. [CrossRef]
- Mamelak, A.N.; Rosenfeld, S.; Bucholz, R.; Raubitschek, A.; Nabors, L.B.; Fiveash, J.B.; Shen, S.; Khazaeli, M.B.; Colcher, D.; Liu, A.; et al. Phase I Single-Dose Study of Intracavitary-Administered Iodine-131-TM-601 in Adults With Recurrent High-Grade Glioma. Journal of Clinical Oncology 2006, 24, 3644–3650. [CrossRef]
- Gribbin, T.E.; Senzer, N.; Raizer, J.J.; Shen, S.; Nabors, L.B.; Wiranowska, M.; Fiveash, J.B. A Phase I Evaluation of Intravenous (IV) 131 I-Chlorotoxin Delivery to Solid Peripheral and Intracranial Tumors. Journal of Clinical Oncology 2009, 27, e14507–e14507. [CrossRef]
- Patil, C.G.; Walker, D.G.; Miller, D.M.; Butte, P.; Morrison, B.; Kittle, D.S.; Hansen, S.J.; Nufer, K.L.; Byrnes-Blake, K.A.; Yamada, M.; et al. Phase 1 Safety, Pharmacokinetics, and Fluorescence Imaging Study of Tozuleristide (BLZ-100) in Adults With Newly Diagnosed or Recurrent Gliomas. Neurosurgery 2019, 85, E641–E649. [CrossRef]
- Díaz-Perlas, C.; Varese, M.; Guardiola, S.; García, J.; Sánchez-Navarro, M.; Giralt, E.; Teixidó, M. From Venoms to BBB-Shuttles. MiniCTX3: A Molecular Vector Derived from Scorpion Venom. Chem Commun (Camb) 2018, 54, 12738–12741. [CrossRef]
- Formicola, B.; Dal Magro, R.; Montefusco-Pereira, C. V; Lehr, C.-M.; Koch, M.; Russo, L.; Grasso, G.; Deriu, M.A.; Danani, A.; Bourdoulous, S.; et al. The Synergistic Effect of Chlorotoxin-MApoE in Boosting Drug-Loaded Liposomes across the BBB. J Nanobiotechnology 2019, 17, 115. [CrossRef]
- Reissmann, S.; Filatova, M.P. New Generation of Cell-penetrating Peptides: Functionality and Potential Clinical Application. Journal of Peptide Science 2021, 27. [CrossRef]
- Poillot, C.; Dridi, K.; Bichraoui, H.; Pêcher, J.; Alphonse, S.; Douzi, B.; Ronjat, M.; Darbon, H.; De Waard, M. D-Maurocalcine, a Pharmacologically Inert Efficient Cell-Penetrating Peptide Analogue. Journal of Biological Chemistry 2010, 285, 34168–34180. [CrossRef]
- Poillot, C.; Bichraoui, H.; Tisseyre, C.; Bahemberae, E.; Andreotti, N.; Sabatier, J.-M.; Ronjat, M.; De Waard, M. Small Efficient Cell-Penetrating Peptides Derived from Scorpion Toxin Maurocalcine. Journal of Biological Chemistry 2012, 287, 17331–17342. [CrossRef]
- Fajloun, Z.; Kharrat, R.; Chen, L.; Lecomte, C.; Di Luccio, E.; Bichet, D.; El Ayeb, M.; Rochat, H.; Allen, P.D.; Pessah, I.N.; et al. Chemical Synthesis and Characterization of Maurocalcine, a Scorpion Toxin That Activates Ca(2+) Release Channel/Ryanodine Receptors. FEBS Lett 2000, 469, 179–185. [CrossRef]
- Estève, E.; Mabrouk, K.; Dupuis, A.; Smida-Rezgui, S.; Altafaj, X.; Grunwald, D.; Platel, J.-C.; Andreotti, N.; Marty, I.; Sabatier, J.-M.; et al. Transduction of the Scorpion Toxin Maurocalcine into Cells. Journal of Biological Chemistry 2005, 280, 12833–12839. [CrossRef]
- Ram, N.; Weiss, N.; Texier-Nogues, I.; Aroui, S.; Andreotti, N.; Pirollet, F.; Ronjat, M.; Sabatier, J.-M.; Darbon, H.; Jacquemond, V.; et al. Design of a Disulfide-Less, Pharmacologically Inert, and Chemically Competent Analog of Maurocalcine for the Efficient Transport of Impermeant Compounds into Cells. Journal of Biological Chemistry 2008, 283, 27048–27056. [CrossRef]
- Tisseyre, C.; Bahembera, E.; Dardevet, L.; Sabatier, J.-M.; Ronjat, M.; De Waard, M. Cell Penetration Properties of a Highly Efficient Mini Maurocalcine Peptide. Pharmaceuticals 2013, 6, 320–339. [CrossRef]
- Zhou, M.; Zou, X.; Cheng, K.; Zhong, S.; Su, Y.; Wu, T.; Tao, Y.; Cong, L.; Yan, B.; Jiang, Y. The Role of Cell-penetrating Peptides in Potential Anti-cancer Therapy. Clin Transl Med 2022, 12. [CrossRef]
- Delgado-Prudencio, G.; Possani, L.D.; Becerril, B.; Ortiz, E. The Dual α-Amidation System in Scorpion Venom Glands. Toxins (Basel) 2019, 11, 425. [CrossRef]
- Zeng, L.; Zhang, C.; Yang, M.; Sun, J.; Lu, J.; Zhang, H.; Qin, J.; Zhang, W.; Jiang, Z. Unveiling the Diversity and Modifications of Short Peptides in Buthus martensii Scorpion Venom through Liquid Chromatography-High Resolution Mass Spectrometry. Toxins (Basel) 2024, 16, 155. [CrossRef]
- Wu, D.; Chen, Q.; Chen, X.; Han, F.; Chen, Z.; Wang, Y. The Blood–Brain Barrier: Structure, Regulation and Drug Delivery. Signal Transduct Target Ther 2023, 8, 217. [CrossRef]
- Fan, S.; Sun, Z.; Jiang, D.; Dai, C.; Ma, Y.; Zhao, Z.; Liu, H.; Wu, Y.; Cao, Z.; Li, W. BmKCT Toxin Inhibits Glioma Proliferation and Tumor Metastasis. Cancer Lett 2010, 291, 158–166. [CrossRef]
- Agarwal, S.; Mohamed, M.S.; Mizuki, T.; Maekawa, T.; Sakthi Kumar, D. Chlorotoxin Modified Morusin–PLGA Nanoparticles for Targeted Glioblastoma Therapy. J Mater Chem B 2019, 7, 5896–5919. [CrossRef]
- Veiseh, M.; Gabikian, P.; Bahrami, S.-B.; Veiseh, O.; Zhang, M.; Hackman, R.C.; Ravanpay, A.C.; Stroud, M.R.; Kusuma, Y.; Hansen, S.J.; et al. Tumor Paint: A Chlorotoxin:Cy5.5 Bioconjugate for Intraoperative Visualization of Cancer Foci. Cancer Res 2007, 67, 6882–6888. [CrossRef]
- Mata de los Rios, N.; Gastelum-Arellanez, A.; Clement, H.; Álvarez-Cruz, K.; Romero-Terrazas, D.; Alvarado-González, C.; Hinojos-Gallardo, L.C.; Corzo, G.; Espino-Solis, G.P. Ion-Channel-Targeting Scorpion Recombinant Toxin as Novel Therapeutic Agent for Breast Cancer. Toxins (Basel) 2025, 17, 166. [CrossRef]
- Pucca, M.B.; Peigneur, S.; Cologna, C.T.; Cerni, F.A.; Zoccal, K.F.; Bordon, K. de C.F.; Faccioli, L.H.; Tytgat, J.; Arantes, E.C. Electrophysiological Characterization of the First Tityus serrulatus Alpha-like Toxin, Ts5: Evidence of a pro-Inflammatory Toxin on Macrophages. Biochimie 2015, 115, 8–16. [CrossRef]
- Chen, B.; Zhang, C.; Wang, Z.; Chen, Y.; Xie, H.; Li, S.; Liu, X.; Liu, Z.; Chen, P. Mechanistic Insights into Nav1.7-dependent Regulation of Rat Prostate Cancer Cell Invasiveness Revealed by Toxin Probes and Proteomic Analysis. FEBS J 2019, 286, 2549–2561. [CrossRef]
- Soroceanu, L.; Manning, T.J.; Sontheimer, H. Modulation of Glioma Cell Migration and Invasion Using Cl − and K + Ion Channel Blockers. The Journal of Neuroscience 1999, 19, 5942–5954. [CrossRef]
- Cota-Arce, J.M.; Zazueta-Favela, D.; Díaz-Castillo, F.; Jiménez, S.; Bernáldez-Sarabia, J.; Caram-Salas, N.L.; Dan, K.W.L.; Escobedo, G.; Licea-Navarro, A.F.; Possani, L.D.; et al. Venom Components of the Scorpion Centruroides Limpidus Modulate Cytokine Expression by T Helper Lymphocytes: Identification of Ion Channel-Related Toxins by Mass Spectrometry. Int Immunopharmacol 2020, 84, 106505. [CrossRef]
- Park, J.; Oh, J.H.; Kang, H.K.; Choi, M.-C.; Seo, C.H.; Park, Y. Scorpion-Venom-Derived Antimicrobial Peptide Css54 Exerts Potent Antimicrobial Activity by Disrupting Bacterial Membrane of Zoonotic Bacteria. Antibiotics 2020, 9, 831. [CrossRef]
- Zoccal, K.F.; Bitencourt, C. da S.; Paula-Silva, F.W.G.; Sorgi, C.A.; de Castro Figueiredo Bordon, K.; Arantes, E.C.; Faccioli, L.H. TLR2, TLR4 and CD14 Recognize Venom-Associated Molecular Patterns from Tityus serrulatus to Induce Macrophage-Derived Inflammatory Mediators. PLoS One 2014, 9, e88174. [CrossRef]
- Xu, L.; Zhang, Y.; Dai, Q.; Lin, N.; Guan, T.; Song, X.; Hong, S. Scorpion Venom Polypeptide Governs Alveolar Macrophage M1/M2 Polarization to Alleviate Pulmonary Fibrosis. Tissue Cell 2022, 79, 101939. [CrossRef]
- Yglesias-Rivera, A.; Sánchez-Rodríguez, H.; Soto-Febles, C.; Monzote, L. Heteroctenus junceus Scorpion Venom Modulates the Concentration of Pro-Inflammatory Cytokines in F3II Tumor Cells. Life 2023, 13, 2287. [CrossRef]
- Díaz-García, A.; Ruiz-Fuentes, J.L.; Frión-Herrera, Y.; Yglesias-Rivera, A.; Garlobo, Y.R.; Sánchez, H.R.; Aurrecochea, J.C.R.; López Fuentes, L.X. Rhopalurus junceus Scorpion Venom Induces Antitumor Effect in Vitro and in Vivo against a Murine Mammary Adenocarcinoma Model. Iran J Basic Med Sci 2019, 22, 759–765. [CrossRef]
- Zoccal, K.F.; Paula-Silva, F.W.G.; Bitencourt, C. da S.; Sorgi, C.A.; Bordon, K. de C.F.; Arantes, E.C.; Faccioli, L.H. PPAR-γ Activation by Tityus serrulatus Venom Regulates Lipid Body Formation and Lipid Mediator Production. Toxicon 2015, 93, 90–97. [CrossRef]
- Ait-Lounis, A.; Laraba-Djebari, F. TNF-Alpha Modulates Adipose Macrophage Polarization to M1 Phenotype in Response to Scorpion Venom. Inflamm Res 2015, 64, 929–936. [CrossRef]
- Santhosh, K.N.; Ramesh, D.; Ramesh, D.; Nagaraj, U.; Shrinidhi, S.; Thippeswamy, N.B. Scorpion Venom Exhibits Adjuvant Effect by Eliciting HBsAg-Specific Th1 Immunity through Neuro-Endocrine Interactions. Mol Immunol 2022, 147, 136–146. [CrossRef]
- Gurrola, G.B.; Hernández-López, R.A.; Rodríguez de la Vega, R.C.; Varga, Z.; Batista, C.V.F.; Salas-Castillo, S.P.; Panyi, G.; del Río-Portilla, F.; Possani, L.D. Structure, Function, and Chemical Synthesis of Vaejovis Mexicanus Peptide 24: A Novel Potent Blocker of Kv1.3 Potassium Channels of Human T Lymphocytes. Biochemistry 2012, 51, 4049–4061. [CrossRef]
- Veytia-Bucheli, J.I.; Jiménez-Vargas, J.M.; Melchy-Pérez, E.I.; Sandoval-Hernández, M.A.; Possani, L.D.; Rosenstein, Y. Kv1.3 Channel Blockade with the Vm24 Scorpion Toxin Attenuates the CD4+ Effector Memory T Cell Response to TCR Stimulation. Cell Communication and Signaling 2018, 16, 45. [CrossRef]
- Chimote, A.A.; Hajdu, P.; Sfyris, A.M.; Gleich, B.N.; Wise-Draper, T.; Casper, K.A.; Conforti, L. Kv1.3 Channels Mark Functionally Competent CD8+ Tumor-Infiltrating Lymphocytes in Head and Neck Cancer. Cancer Res 2017, 77, 53–61. [CrossRef]
- Zhu, M.; Zhang, S.; Tang, J.; Hou, H.; Wang, L.; Lin, H.; Zhang, X.; Jin, M. Two Small Peptides from Buthus martensii Hydrolysates Exhibit Antitumor Activity Through Inhibition of TNF-α-Mediated Signal Transduction Pathways. Life 2025, 15, 105. [CrossRef]
- Magalhães, M.M.; Pereira, M.E.S.; Amaral, C.F.S.; Rezende, N.A.; Campolina, D.; Bucaretchi, F.; Gazzinelli, R.T.; Cunha-Melo, J.R. Serum Levels of Cytokines in Patients Envenomed by Tityus serrulatus Scorpion Sting. Toxicon 1999, 37, 1155–1164. [CrossRef]
- Gunas, V.; Maievskyi, O.; Synelnyk, T.; Raksha, N.; Vovk, T.; Halenova, T.; Savchuk, O.; Gunas, I. Cytokines and Their Regulators in Rat Lung Following Scorpion Envenomation. Toxicon X 2024, 22, 100198. [CrossRef]
- Zeng, L.; Sun, Y.; Zhang, H.; Yi, X.; Du, R.; Chen, Z.; Wang, Q. Scorpion Venom Peptides Enhance Immunity and Survival in Litopenaeus vannamei through Antibacterial Action against Vibrio Parahaemolyticus. Front Immunol 2025, 16. [CrossRef]
- Borges, A.; Op den Camp, H.J.M.; De Sanctis, J.B. Specific Activation of Human Neutrophils by Scorpion Venom: A Flow Cytometry Assessment. Toxicology in Vitro 2011, 25, 358–367. [CrossRef]
- Abdollahnia, A.; Bahmani, K.; Aliahmadi, A.; As’habi, M.A.; Ghassempour, A. Mass Spectrometric Analysis of Odonthobuthus doriae Scorpion Venom and Its Non-Neutralized Fractions after Interaction with Commercial Antivenom. Sci Rep 2024, 14, 10389. [CrossRef]
- Zoccal, K.F.; Bitencourt, C. da S.; Secatto, A.; Sorgi, C.A.; Bordon, K. de C.F.; Sampaio, S.V.; Arantes, E.C.; Faccioli, L.H. Tityus serrulatus Venom and Toxins Ts1, Ts2 and Ts6 Induce Macrophage Activation and Production of Immune Mediators. Toxicon 2011, 57, 1101–1108. [CrossRef]
- Martin-Eauclaire, M.-F.; Bougis, P.E.; de Lima, M.E. Ts1 from the Brazilian Scorpion Tityus serrulatus: A Half-Century of Studies on a Multifunctional Beta like-Toxin. Toxicon 2018, 152, 106–120. [CrossRef]
- Shariati, S.; Mafakher, L.; Shirani, M.; Baradaran, M. Unveiling New Kv1.3 Channel Blockers from Scorpion Venom: Characterization of Meuk7–3 and in Silico Design of Its Analogs for Enhanced Affinity and Therapeutic Potential. Int J Biol Macromol 2025, 319, 145327. [CrossRef]
- López-Giraldo, E.; Carrillo, E.; Titaux-Delgado, G.; Cano-Sánchez, P.; Colorado, A.; Possani, L.D.; Río-Portilla, F. del Structural and Functional Studies of Scorpine: A Channel Blocker and Cytolytic Peptide. Toxicon 2023, 222, 106985. [CrossRef]
- Zhang, Y.; Li, X.; Tian, C.; Zhong, C.; Li, W.; Shang, X.; Zhao, M.; Zhao, Y. Engineered Bacteria-Mediated Delivery of Scorpion Venom Peptide AGAP for Targeted Breast Cancer Therapy. Curr Microbiol 2025, 82, 323. [CrossRef]
- Wang, D.; Starr, R.; Chang, W.-C.; Aguilar, B.; Alizadeh, D.; Wright, S.L.; Yang, X.; Brito, A.; Sarkissian, A.; Ostberg, J.R.; et al. Chlorotoxin-Directed CAR T Cells for Specific and Effective Targeting of Glioblastoma. Sci Transl Med 2020, 12. [CrossRef]
- Zargan, J.; Sajad, M.; Umar, S.; Naime, M.; Ali, S.; Khan, H.A. Scorpion (Androctonus crassicauda) Venom Limits Growth of Transformed Cells (SH-SY5Y and MCF-7) by Cytotoxicity and Cell Cycle Arrest. Exp Mol Pathol 2011, 91, 447–454. [CrossRef]
- Salabi, F.; Jafari, H. Differential Venom Gland Gene Expression Analysis of Juvenile and Adult Scorpions Androctonus crassicauda. BMC Genomics 2022, 23, 636. [CrossRef]
- Butte, P. V; Mamelak, A.; Parrish-Novak, J.; Drazin, D.; Shweikeh, F.; Gangalum, P.R.; Chesnokova, A.; Ljubimova, J.Y.; Black, K. Near-Infrared Imaging of Brain Tumors Using the Tumor Paint BLZ-100 to Achieve near-Complete Resection of Brain Tumors. Neurosurg Focus 2014, 36, E1–E1. [CrossRef]
- Fidel, J.; Kennedy, K.C.; Dernell, W.S.; Hansen, S.; Wiss, V.; Stroud, M.R.; Molho, J.I.; Knoblaugh, S.E.; Meganck, J.; Olson, J.M.; et al. Preclinical Validation of the Utility of BLZ-100 in Providing Fluorescence Contrast for Imaging Spontaneous Solid Tumors. Cancer Res 2015, 75, 4283–4291. [CrossRef]
- Gordon, I.; Paoloni, M.; Mazcko, C.; Khanna, C. The Comparative Oncology Trials Consortium: Using Spontaneously Occurring Cancers in Dogs to Inform the Cancer Drug Development Pathway. PLoS Med 2009, 6, e1000161–e1000161. [CrossRef]
- Parrish-Novak, J.; Byrnes-Blake, K.; Lalayeva, N.; Burleson, S.; Fidel, J.; Gilmore, R.; Gayheart-Walsten, P.; Bricker, G.A.; Crumb, W.J.; Tarlo, K.S.; et al. Nonclinical Profile of BLZ-100, a Tumor-Targeting Fluorescent Imaging Agent. Int J Toxicol 2017, 36, 104–112. [CrossRef]
- Kobets, A.J.; Nauen, D.; Lee, A.; Cohen, A.R. Unexpected Binding of Tozuleristide “Tumor Paint” to Cerebral Vascular Malformations: A Potentially Novel Application of Fluorescence-Guided Surgery. Neurosurgery 2021, 89, 204–211. [CrossRef]
- Yamada, M.; Miller, D.M.; Lowe, M.; Rowe, C.; Wood, D.; Soyer, H.P.; Byrnes-Blake, K.; Parrish-Novak, J.; Ishak, L.; Olson, J.M.; et al. A First-in-Human Study of BLZ-100 (Tozuleristide) Demonstrates Tolerability and Safety in Skin Cancer Patients. Contemp Clin Trials Commun 2021, 23, 100830. [CrossRef]
- Das, B.; Saviola, A.J.; Mukherjee, A.K. Biochemical and Proteomic Characterization, and Pharmacological Insights of Indian Red Scorpion Venom Toxins. Front Pharmacol 2021, 12. [CrossRef]
- Puzari, U.; Das, B.; Mukherjee, A.K. Advancements in Diagnostic Techniques for Scorpion Venom Identification: A Comprehensive Review. Toxicon 2025, 253, 108191. [CrossRef]
- Puzari, U.; Khan, M.R.; Mukherjee, A.K. Development of a Gold Nanoparticle-Based Novel Diagnostic Prototype for in Vivo Detection of Indian Red Scorpion (Mesobuthus Tamulus) Venom. Toxicon X 2024, 23, 100203. [CrossRef]
- Chauhan, S.; Mittal, R.; Kumar, M.; Mittal, A.; Kushwah, A.S. Gold Nanoparticle-Based Biosensors for Point-of-Care Diagnostics: A Review of Sensing Nanoparticle Applications and Future Prospects. Comb Chem High Throughput Screen 2025, 28, 417–434. [CrossRef]
- Medley, C.D.; Smith, J.E.; Tang, Z.; Wu, Y.; Bamrungsap, S.; Tan, W. Gold Nanoparticle-Based Colorimetric Assay for the Direct Detection of Cancerous Cells. Anal Chem 2008, 80, 1067–1072. [CrossRef]
- Appidi, T.; Mudigunda, S. V; Kodandapani, S.; Rengan, A.K. Development of Label-Free Gold Nanoparticle Based Rapid Colorimetric Assay for Clinical/Point-of-Care Screening of Cervical Cancer. Nanoscale Adv 2020, 2, 5737–5745. [CrossRef]
- Pedarzani, P.; D’hoedt, D.; Doorty, K.B.; Wadsworth, J.D.F.; Joseph, J.S.; Jeyaseelan, K.; Kini, R.M.; Gadre, S.V.; Sapatnekar, S.M.; Stocker, M.; et al. Tamapin, a Venom Peptide from the Indian Red Scorpion (Mesobuthus Tamulus) That Targets Small Conductance Ca2+-Activated K+ Channels and Afterhyperpolarization Currents in Central Neurons. Journal of Biological Chemistry 2002, 277, 46101–46109. [CrossRef]
- Luis, E.; Anaya-Hernández, A.; León-Sánchez, P.; Durán-Pastén, M.L. The Kv10.1 Channel: A Promising Target in Cancer. Int J Mol Sci 2022, 23. [CrossRef]
- Ouadid-Ahidouch, H.; Ahidouch, A.; Pardo, L.A. Kv10.1 K+ Channel: From Physiology to Cancer. Pflugers Arch 2016, 468, 751–762. [CrossRef]
- Cázares-Ordoñez, V.; Pardo, L.A. Kv10.1 Potassium Channel: From the Brain to the Tumors. Biochemistry and Cell Biology 2017, 95, 531–536. [CrossRef]
- Khamehchian, S.; Nikkhah, M.; Madani, R.; Hosseinkhani, S. Enhanced and Selective Permeability of Gold Nanoparticles Functionalized with Cell Penetrating Peptide Derived from Maurocalcine Animal Toxin. J Biomed Mater Res A 2016, 104, 2693–2700. [CrossRef]
- Kievit, F.M.; Veiseh, O.; Fang, C.; Bhattarai, N.; Lee, D.; Ellenbogen, R.G.; Zhang, M. Chlorotoxin Labeled Magnetic Nanovectors for Targeted Gene Delivery to Glioma. ACS Nano 2010, 4, 4587–4594. [CrossRef]
- Zhao, L.; Li, Y.; Zhu, J.; Sun, N.; Song, N.; Xing, Y.; Huang, H.; Zhao, J. Chlorotoxin Peptide-Functionalized Polyethylenimine-Entrapped Gold Nanoparticles for Glioma SPECT/CT Imaging and Radionuclide Therapy. J Nanobiotechnology 2019, 17, 30. [CrossRef]
- Zheng, T.; Zhang, Z.; Yu, Z.; Wang, H.; Lyu, X.; Han, C. Investigation on the Mechanisms of Scorpion Venom in Hepatocellular Carcinoma Model Mice via Untargeted Metabolomics Profiling. Int Immunopharmacol 2024, 138, 112578. [CrossRef]
- Mazhdi, Y.; Hamidi, S.M. Detection of Scorpion Venom by Optical Circular Dichroism Method. Sci Rep 2021, 11, 15854. [CrossRef]
- Sun, C.; Veiseh, O.; Gunn, J.; Fang, C.; Hansen, S.; Lee, D.; Sze, R.; Ellenbogen, R.G.; Olson, J.; Zhang, M. In Vivo MRI Detection of Gliomas by Chlorotoxin-Conjugated Superparamagnetic Nanoprobes. Small 2008, 4, 372–379. [CrossRef]
- Zhao, L.; Shi, X.; Zhao, J. Chlorotoxin-Conjugated Nanoparticles for Targeted Imaging and Therapy of Glioma. Curr Top Med Chem 2015, 15, 1196–1208. [CrossRef]
- WANG, X.; GUO, Z. Chlorotoxin-Conjugated Onconase as a Potential Anti-Glioma Drug. Oncol Lett 2015, 9, 1337–1342. [CrossRef]
- Costa, P.M.; Cardoso, A.L.; Mendonça, L.S.; Serani, A.; Custódia, C.; Conceição, M.; Simões, S.; Moreira, J.N.; Pereira de Almeida, L.; Pedroso de Lima, M.C. Tumor-Targeted Chlorotoxin-Coupled Nanoparticles for Nucleic Acid Delivery to Glioblastoma Cells: A Promising System for Glioblastoma Treatment. Mol Ther Nucleic Acids 2013, 2, e100. [CrossRef]
- Amen, R.A.; Abd-Ellatef, G.E.F. Scorpion Venom and Its Different Peptides Aid in Treatment Focusing on Cancer Disease with the Mechanism of Action. Trends in Pharmacy 2025, 1, 1–11. [CrossRef]
- Rjeibi, I.; Mabrouk, K.; Mosrati, H.; Berenguer, C.; Mejdoub, H.; Villard, C.; Laffitte, D.; Bertin, D.; Ouafik, L.; Luis, J.; et al. Purification, Synthesis and Characterization of AaCtx, the First Chlorotoxin-like Peptide from Androctonus australis Scorpion Venom. Peptides (N.Y.) 2011, 32, 656–663. [CrossRef]
- Huang, R.; Ke, W.; Han, L.; Li, J.; Liu, S.; Jiang, C. Targeted Delivery of Chlorotoxin-Modified DNA-Loaded Nanoparticles to Glioma via Intravenous Administration. Biomaterials 2011, 32, 2399–2406. [CrossRef]
- Thakur, A.; Alajangi, H.K.; Sharma, A.; Hwang, E.; Khajuria, A.; Kumari, L.; Jaiswal, P.K.; Lim, Y.; Singh, G.; Barnwal, R.P. Stigmurin Encapsulated PLA–PEG Ameliorates Its Therapeutic Potential, Antimicrobial and Antiproliferative Activities. Discover Nano 2025, 20, 50. [CrossRef]
- Aliakbari, F.; Rahmati, S.; Ghanbari, A.; Madanchi, H.; Rashidy-Pour, A. Identification and Designing an Analgesic Opioid Cyclic Peptide from Defensin 4 of Mesobuthus martensii Karsch Scorpion Venom with More Effectiveness than Morphine. Biomedicine & Pharmacotherapy 2025, 188, 118139. [CrossRef]
- Ghodeif, S.K.; El-Fahla, N.A.; Abdel-Rahman, M.A.; El-Shenawy, N.S. Arthropod Venom Peptides: Pioneering Nanotechnology in Cancer Treatment and Drug Delivery. Cancer Pathogenesis and Therapy 2025. [CrossRef]
- Harrison, E.; Nicol, J.R.; Macias–Montero, M.; Burke, G.A.; Coulter, J.A.; Meenan, B.J.; Dixon, D. A Comparison of Gold Nanoparticle Surface Co-Functionalization Approaches Using Polyethylene Glycol (PEG) and the Effect on Stability, Non-Specific Protein Adsorption and Internalization. Materials Science and Engineering: C 2016, 62, 710–718. [CrossRef]
- Gláucia-Silva, F.; Torres, J.V.P.; Torres-Rêgo, M.; Daniele-Silva, A.; Furtado, A.A.; Ferreira, S. de S.; Chaves, G.M.; Xavier-Júnior, F.H.; Rocha Soares, K.S.; Silva-Júnior, A.A. da; et al. Tityus Stigmurus-Venom-Loaded Cross-Linked Chitosan Nanoparticles Improve Antimicrobial Activity. Int J Mol Sci 2024, 25, 9893. [CrossRef]
- Rebbouh, F.; Martin-Eauclaire, M.-F.; Laraba-Djebari, F. Chitosan Nanoparticles as a Delivery Platform for Neurotoxin II from Androctonus australis hector Scorpion Venom: Assessment of Toxicity and Immunogenicity. Acta Trop 2020, 205, 105353. [CrossRef]
- Fu, Y.; An, N.; Li, K.; Zheng, Y.; Liang, A. Chlorotoxin-Conjugated Nanoparticles as Potential Glioma-Targeted Drugs. J Neurooncol 2012, 107, 457–462. [CrossRef]
- Zhi, D.; Yang, T.; Yang, J.; Fu, S.; Zhang, S. Targeting Strategies for Superparamagnetic Iron Oxide Nanoparticles in Cancer Therapy. Acta Biomater 2020, 102, 13–34. [CrossRef]
- Yue, P.; He, L.; Qiu, S.; Li, Y.; Liao, Y.; Li, X.; Xie, D.; Peng, Y. OX26/CTX-Conjugated PEGylated Liposome as a Dual-Targeting Gene Delivery System for Brain Glioma. Mol Cancer 2014, 13, 191. [CrossRef]
- Ward, M.J.; Ellsworth, S.A.; Nystrom, G.S. A Global Accounting of Medically Significant Scorpions: Epidemiology, Major Toxins, and Comparative Resources in Harmless Counterparts. Toxicon 2018, 151, 137–155. [CrossRef]
- Li, M.; Shao, X.; Wu, C.; Lu, D.; Liu, K.; Wang, W.; Liu, J.; Li, H.; Su, W.; Fang, L. Chlorotoxin-Derived Bicyclic Peptides for Targeted Imaging of Glioblastomas. Chemical Communications 2020, 56, 9537–9540. [CrossRef]
- Baik, F.M.; Hansen, S.; Knoblaugh, S.E.; Sahetya, D.; Mitchell, R.M.; Xu, C.; Olson, J.M.; Parrish-Novak, J.; Méndez, E. Fluorescence Identification of Head and Neck Squamous Cell Carcinoma and High-Risk Oral Dysplasia With BLZ-100, a Chlorotoxin-Indocyanine Green Conjugate. JAMA Otolaryngology–Head & Neck Surgery 2016, 142, 330. [CrossRef]
- Dintzis, S.M.; Hansen, S.; Harrington, K.M.; Tan, L.C.; Miller, D.M.; Ishak, L.; Parrish-Novak, J.; Kittle, D.; Perry, J.; Gombotz, C.; et al. Real-Time Visualization of Breast Carcinoma in Pathology Specimens From Patients Receiving Fluorescent Tumor-Marking Agent Tozuleristide. Arch Pathol Lab Med 2019, 143, 1076–1083. [CrossRef]
- Klint, J.K.; Senff, S.; Saez, N.J.; Seshadri, R.; Lau, H.Y.; Bende, N.S.; Undheim, E.A.B.; Rash, L.D.; Mobli, M.; King, G.F. Production of Recombinant Disulfide-Rich Venom Peptides for Structural and Functional Analysis via Expression in the Periplasm of E. Coli. PLoS One 2013, 8, e63865–e63865. [CrossRef]
- Ma, Y.; Lee, C.-J.; Park, J.-S. Strategies for Optimizing the Production of Proteins and Peptides with Multiple Disulfide Bonds. Antibiotics 2020, 9, 541. [CrossRef]
- Abd El-Aziz, T.M.; Soares, A.G.; Stockand, J.D. Advances in Venomics: Modern Separation Techniques and Mass Spectrometry. Journal of Chromatography B 2020, 1160, 122352. [CrossRef]
- Dias, N.B.; de Souza, B.M.; Cocchi, F.K.; Chalkidis, H.M.; Dorce, V.A.C.; Palma, M.S. Profiling the Short, Linear, Non-Disulfide Bond-Containing Peptidome from the Venom of the Scorpion Tityus obscurus. J Proteomics 2018, 170, 70–79. [CrossRef]
- Ma, Y.; He, Y.; Zhao, R.; Wu, Y.; Li, W.; Cao, Z. Extreme Diversity of Scorpion Venom Peptides and Proteins Revealed by Transcriptomic Analysis: Implication for Proteome Evolution of Scorpion Venom Arsenal. J Proteomics 2012, 75, 1563–1576. [CrossRef]
- Bordon, K.; Santos, G.; Martins, J.; Wiezel, G.; Amorim, F.; Crasset, T.; Redureau, D.; Quinton, L.; Procópio, R.; Arantes, E. Pioneering Comparative Proteomic and Enzymatic Profiling of Amazonian Scorpion Venoms Enables the Isolation of Their First α-Ktx, Metalloprotease, and Phospholipase A2. Toxins (Basel) 2025, 17, 411. [CrossRef]
- García-Villalvazo, P.E.; Jiménez-Vargas, J.M.; Lino-López, G.J.; Meneses, E.P.; Bermúdez-Guzmán, M. de J.; Barajas-Saucedo, C.E.; Delgado Enciso, I.; Possani, L.D.; Valdez-Velazquez, L.L. Unveiling the Protein Components of the Secretory-Venom Gland and Venom of the Scorpion Centruroides Possanii (Buthidae) through Omic Technologies. Toxins (Basel) 2023, 15, 498. [CrossRef]
- Daoudi, K.; Malosse, C.; Lafnoune, A.; Darkaoui, B.; Chakir, S.; Sabatier, J.; Chamot-Rooke, J.; Cadi, R.; Oukkache, N. Mass Spectrometry-based Top-down and Bottom-up Approaches for Proteomic Analysis of the Moroccan Buthus Occitanus Scorpion Venom. FEBS Open Bio 2021, 11, 1867–1892. [CrossRef]
- von Reumont, B.M.; Anderluh, G.; Antunes, A.; Ayvazyan, N.; Beis, D.; Caliskan, F.; Crnković, A.; Damm, M.; Dutertre, S.; Ellgaard, L.; et al. Modern Venomics—Current Insights, Novel Methods, and Future Perspectives in Biological and Applied Animal Venom Research. Gigascience 2022, 11. [CrossRef]
- Ojeda, P.G.; Wang, C.K.; Craik, D.J. Chlorotoxin: Structure, Activity, and Potential Uses in Cancer Therapy. Peptide Science 2016, 106, 25–36. [CrossRef]
- Mamelak, A.N. Targeted Antitumor Therapy with the Scorpion Venom Chlorotoxin. Drugs Future 2011, 36, 0615. [CrossRef]
- Blaney, E.; Demeke, M.; Kamayirese, S.; Monga, L.; Hansen, L.A.; Watts, C.R.; Lovas, S. Does Chlorotoxin Target Matrix Metalloproteinase-2 in Glioblastoma? 2025.
- Mouchbahani-Constance, S.; Sharif-Naeini, R. Proteomic and Transcriptomic Techniques to Decipher the Molecular Evolution of Venoms. Toxins (Basel) 2021, 13, 154. [CrossRef]
- Robinson, S.D.; Undheim, E.A.B.; Ueberheide, B.; King, G.F. Venom Peptides as Therapeutics: Advances, Challenges and the Future of Venom-Peptide Discovery. Expert Rev Proteomics 2017, 14, 931–939. [CrossRef]
- Joglekar, A.V.; Dehari, D.; Anjum, Md.M.; Dulla, N.; Chaudhuri, A.; Singh, S.; Agrawal, A.K. Therapeutic Potential of Venom Peptides: Insights in the Nanoparticle-Mediated Venom Formulations. Futur J Pharm Sci 2022, 8, 34. [CrossRef]
- Psenicnik, A.; Ojanguren-Affilastro, A.A.; Graham, M.R.; Hassan, M.K.; Abdel-Rahman, M.A.; Sharma, P.P.; Santibáñez-López, C.E. Optimizing Scorpion Toxin Processing through Artificial Intelligence. Toxins (Basel) 2024, 16, 437. [CrossRef]
- Liscano, Y.; Oñate-Garzón, J.; Delgado, J.P. Peptides with Dual Antimicrobial–Anticancer Activity: Strategies to Overcome Peptide Limitations and Rational Design of Anticancer Peptides. Molecules 2020, 25, 4245. [CrossRef]
Figure 1.
Workflow for the Discovery and Development of Scorpion Venom-Derived Anticancer Therapeutics.
Figure 1.
Workflow for the Discovery and Development of Scorpion Venom-Derived Anticancer Therapeutics.
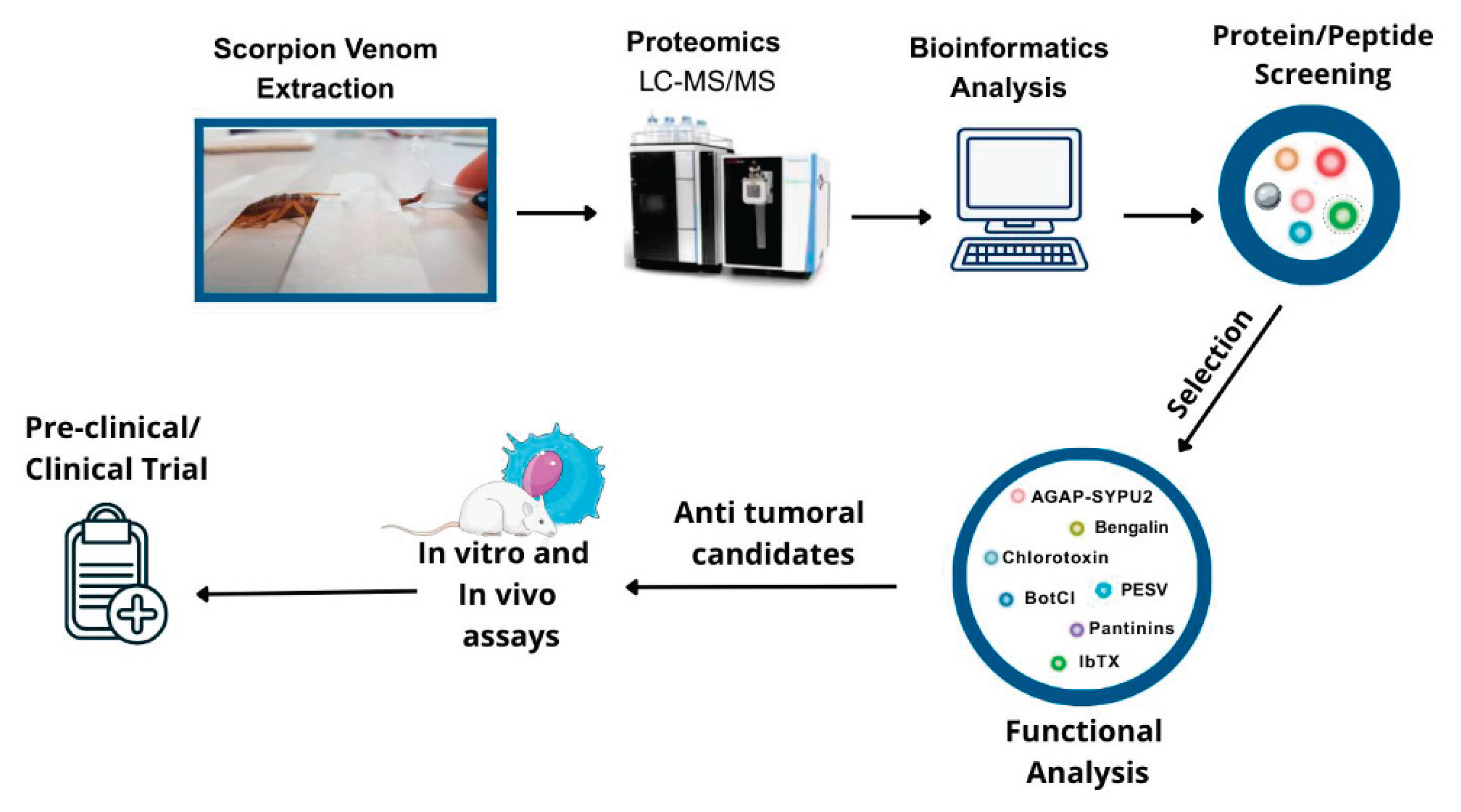
Figure 2.
Mechanisms of Anticancer Action of Selected Scorpion Venom Peptides.
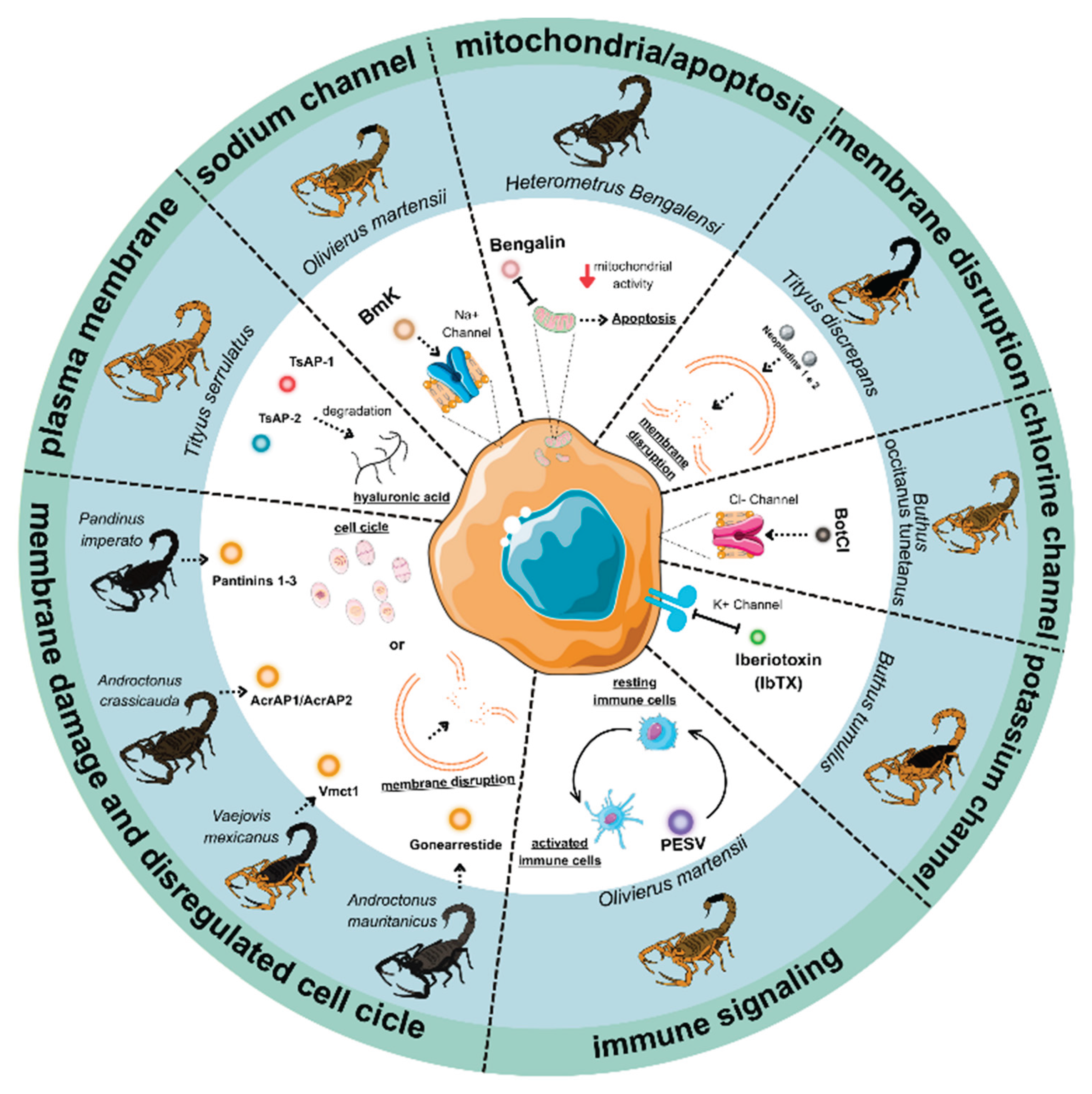

Table 1.
Scorpion venom-derived peptides and proteins with demonstrated anticancer activity.
| Peptide/Protein Name | Scorpion Source | Molecular Target | Primary Mechanism | Cancer Models Tested | Proteomics/ Mass Spectrometry Analysis | Key References |
|---|---|---|---|---|---|---|
| AcrAP1/AcrAP2 | Androctonus crassicauda | Cell membranes | They block the recognition and binding of the target DNA by the Cascade complex, preventing the subsequent degradation triggered by Cas3. | Prostate carcinoma cell line and Human lung adenocarcinoma | - | [28,32,33,34] |
| AGAP / AGAP-SYPU2 | Buthus martensii Karsch | Voltage-gated sodium channels (Nav1.4, Nav1.5, Nav1.7) | Ion channel modulation, inhibition of proliferation and migration, apoptosis | Fibrosarcoma and Human Colon Cancer | MALDI-TOF-MS | [35,36,37] |
| Bengalin | Heterometrus bengalensis | Mitochondria (Bax/Bcl-2, cytochrome c release, caspase-9/-3) | Intrinsic apoptosis: mitochondrial depolarization, cytochrome c release, PARP cleavage, HSP70/90 downregulation | Human Leukemia (histiocytic lymphoma) and chronic myelogenous leukemia | - | [38,39,40,41,42] |
| BmK crude venom | Buthus martensii Karsch | PI3K/Akt signaling, PTEN/p27 pathways | Oncogenic signaling inhibition; PTEN upregulation and Akt/Bad suppression; apoptosis in lymphoma cells | Human glioma, Human lymphoma, Human breast cancer and Human hepatoma | - | [43,44] |
| BmKn2 | Mesobuthus martensii Karsch | Mitochondria (Bax/Bcl-2, caspase-9) | Apoptosis induction, ROS generation, mitochondrial dysfunction | Human oral squamous carcinoma | - | [45,46] |
| BotCl | Buthus occitanus tunetanus | ClC-3, MMP-2 | Chlorotoxin-like; inhibition of migration and invasion in glioblastoma/breast cancer | Human glioblastoma and human breast cancer | - | [47,48,49] |
| Chlorotoxin (CTX) | Leiurus hebraeus | ClC-3 chloride channels, MMP-2 | Inhibition of tumor cell invasion and migration | Malignant glioma | - | [49,50] |
| Cm28 | Centruroides margaritatus | Potassium channel Kv1.3 | Suppress T cell activation (IL-2R and CD40L) | Not tested | - | [51] |
| Crude venom | Hottentotta saulcyi | Na+ and K+ channels | Apoptosis induction through MCF-7 cells via TNF-α and caspase-3 upregulation | Colon Cancer | LC-MS/MS | [52,53] |
| Gonearestide | Androctonus mauritanicus | Cell membranes | Induces G1 cell cycle arrest in HCT116 colorectal cancer cells by dually targeting CDK regulators. It downregulates CDK4 and upregulates the CDK inhibitors p21 and p27, along with cyclin D3. | Prostate carcinoma cell line and Human lung adenocarcinoma | LC-MS/MS and MALDI-TOF | [54] |
| Hemiscorpius lepturus crude venom | Hemiscorpius lepturus | Bax, p53, caspase-3 activation, Bcl-2 suppression | Apoptosis induction in colon carcinoma and xenografts with low toxicity to normal cells | Colorectal cancer | - | [51,55] |
| HsTX1 | Heterometrus spinnifer | Potassium channel Kv1.3 | Suppress pathogenic effector memory T lymphocyte | Not tested | MALDI-TOF | [56] |
| Iberiotoxin (IbTX) | Hottentotta tamulus (syn. Buthus/Mesobuthus tamulus) | Potassium channels Kv1.1, Kv1.3 | Blockade of K⁺ currents, Ca²⁺ dysregulation, apoptosis | Breast Cancer | - | [57,58,59,60,61]. |
| Margatoxin (MgTX) | Centruroides margaritatus | Potassium channel Kv1.3 | Suppression of proliferation and cell cycle regulation, tumor growth inhibition | Lung adenocarcinoma | - | [62] |
| Maurocalcine (MCa) | Scorpion maurus palmatus | Ryanodine receptor (RyR1) | Ca²⁺ release, cell-penetrating peptide, apoptosis and drug delivery | Breast Cancer | - | [63,64,65,66] |
| Neopladine 1 and 2 | Tityus discrepans | Fas ligand → caspase-8 → Bid/tBid | Extrinsic apoptosis via death receptor pathway; cross-talk with intrinsic mitochondrial pathway | Breast Cancer | MALDI-TOF | [67] |
| Pantinins 1-3 | Pandinus imperator | Cell membranes | Destabilizing and disrupting the membrane of target cells. | Breast adenocarcinoma and Prostate adenocarcinoma | Not specified | [68,69] |
| Smp43 | Scorpio maurus palmatus | Mitochondria | Apoptosis induction by caspase-1 activation, triggering pyroptosis | Human Lung Cancer | - | [70,71,72,73] |
| Smp24 | Scorpio maurus palmatus | Mitochondria | Apoptosis induction by reducing mitochondrial membrane potential (ΔΨm), increasing reactive oxygen species (ROS) production. Also, inducing apoptosis via caspase-1 activation triggering pyroptosis | Human Lung Cancer | - | [74,75] |
| TsAP-1 and TsAP-2 | Tityus serrulatus | Cell membranes | Membrane disruption, cytotoxicity to cancer cells, apoptosis | Human Lung Adenocarcinoma, Prostate Adenocarcinoma, Human Squamous Carcinoma | MALDI-TOF/TOF | [76,77,78] |
| TsAP-S2 | Tityus serrulatus | Mitochondria | Apoptosis induction through mitochondrial disruption and activation of caspase pathways | Not tested | ESI-MS | [77]; |
| Vm24 | Vaejovis mexicanus smithi | Potassium channel Kv1.3 | Suppress pathogenic effector memory T lymphocyte | Breast Cancer | LC-MS/MS and MALDI-TOF | [79] |
Table 2.
Clinical trials of scorpion venom-derived compounds.
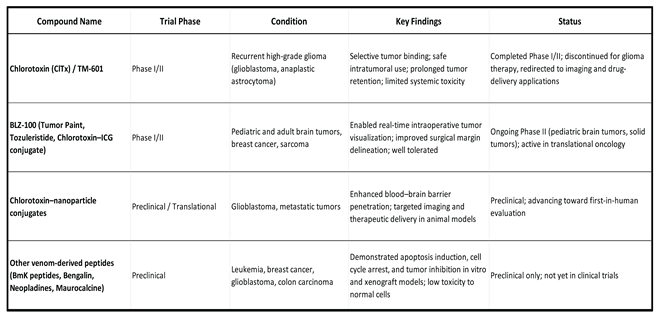 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



