Submitted:
25 August 2025
Posted:
27 August 2025
You are already at the latest version
Abstract
Since abiotic stress is an unfavourable environmental factor that significantly affects plant growth and development, plants have evolved over time to adapt to these stresses, thereby enhancing their survival capabilities. As one of the largest transcription factor families in plants, bZIP transcription factors are involved in plant growth and development, plant hormone signalling transduction, secondary metabolism synthesis pathways, and responses to abiotic stress. Numerous studies have shown that bZIP transcription factors, upon activation by external environmental signals, can either act independently or through interactions with other proteins to bind to the A-box (TACGTA), the C-box (GACGTC), and the G-box (CACGTG) cis-acting elements in the promoter regions of downstream target non-biotic stress genes, thereby regulating the expression of downstream stress response genes and modulating plant tolerance to non-biotic stress as well as secondary metabolic pathways in plants. Additionally, bZIP transcription factors are involved in plant hormone signalling pathways in response to non-biotic stress. For example, abscisic acid, jasmonic acid, brassinosteroids, and salicylic acid. In this review, we discuss the structure, classification, and mechanisms of action of plant bZIP transcription factors, their roles in plant hormone regulation, and the mechanisms by which bZIP transcription factors respond to abiotic stresses such as drought, salt stress, high/low temperatures, nutrient deficiencies, heavy metal stress and light stress. Additionally, we summarise the role of bZIP transcription factors in regulating the biosynthesis of various secondary metabolites, such as flavonoids, terpenoids, alkaloids, phenolic acids and lignans. Finally, the role of bZIP transcription factors and non-coding RNAs (ncRNA) in enhancing tolerance to abiotic stress was discussed. In the future, we will focus on studying bZIP transcription factors as core regulators of downstream gene networks associated with abiotic stress to enhance plant tolerance to abiotic stress and improve crop yields, thereby addressing food security issues.
Keywords:
bZIP transcription factor
; plant hormone signalling
; abiotic stress
; plant growth and development
; secondary metabolite biosynthetic pathways
1. Introduction
Transcription factors are proteins that can bind to specific nucleotide sequences upstream of a gene, and these proteins regulate the transcription of these genes and thus have a regulatory effect on plant growth and development, biotic and abiotic stresses, secondary metabolites [1,2,3,4]. Transcription factors contain four types of structural domains, such as DNA-binding structural domains, transcriptional regulatory structural domains, oligomerisation sites and nuclear localisation structural domains [5,6,7]. Transcription factors can regulate the expression of downstream genes by interacting with other transcription factors, or enhance or repress the expression of genes by binding to the promoter sequences of downstream target genes, thus improving the tolerance of plants to stresses and abiotic stresses, and they play important roles in regulating the physiological and biochemical processes of plants and even the whole life activities [8,9,10,11] (Figure 1). bZIP (basic region/leucine zipper motif) transcription factors are one of the largest families of transcription factors in plants, and they are induced to be expressed in response to abiotic stresses, such as drought, salt, cold, high temperature, nutrient stress, heavy metal stress, and high light stresses, and induced to be expressed in response to biotic stresses. They also play important roles in regulating plant growth and development and secondary metabolite biosynthetic pathways [12,13]. It has been shown that when activated by external environmental signals, bZIP transcription factors can regulate the transcription level of target genes by interacting with other transcription factors or by directly interacting with cis-acting elements in the promoter regions of the adversity-associated genes [14,15,16]. bZIP transcription factors can recognize cis-acting elements with the core sequence ACGT, such as CACGTG (G box), GACGTC (C box), TACGTA (A box) and so on. The bZIP transcription factor is involved in regulating the expression of downstream adversity response-related genes through these elements, thereby regulating plant tolerance to abiotic adversity and biotic adversity [17,18]. Nowadays, researchers have identified a large number of bZIP transcription factors in plants, many of which are involved in plant abiotic stress response and regulate plant resistance under abiotic stress, such as Arabidopsis thaliana L., Oryza sativa L., Zea mays L., Glycine max L. and so on (Table 1). It has been found that overexpression or silencing of the expression of these transcription factor genes can increase plant resistance to abiotic and biotic stresses [12,13,14]. Therefore, the study of bZIP transcription factors is crucial for the future improvement of crop survival under stress conditions. In the future, these research results can not only provide a theoretical basis and support for the improvement of crop resistance, but also be utilised as direct genetic resources.
Phytohormones not only play an important role in the growth and development of plants, but also play an important role in disease, biotic and abiotic stresses resistance signalling pathways in plants [19,20]. Currently, the resistance-related hormones that have been studied in plants include abscisic acid (ABA), salicylic acid (SA), jasmonic acid (JA), ethylene (ET), cytokinin (CTK), brassinosteroid (BR), strigolactones (SL) and cytokinin (CTK), among others. There are many genes directly involved in resistance in plants, and the pathways that regulate the expression of these genes are very complex [21,22,23,24]. The opening of each resistance signalling pathway requires the concerted efforts of many upstream key regulators. When plants are subjected to external abiotic and biotic stresses, specific signalling molecules such as phytohormones, Ca2+ and NO- in their bodies bind to receptor proteins in plant cells, regulating downstream transcription factors to interact with other proteins, and thus activating or repressing the expression of target genes by binding to the promoters of the downstream resistance genes concerned [25,26,27,28]. The bZIP transcription factors, a common family of transcription factors in plants, they usually act as positive or negative regulators in the phytohormone-mediated regulation of disease, stress resistance and plant growth and development signalling [29,30]. The bZIP transcription factors are multifunctional in plant, thus they can participate in a wide range of hormone-mediated signalling pathways. Sometimes, bZIP transcription factors also interact with other transcription factors to co-regulate hormone-mediated signalling pathways [31,32].
In recent years, research reviews on bZIP transcription factors have been published, but most of the published reviews focus only on the discovery, structure, classification, functional diversity, and certain adversity regulatory mechanisms of bZIP transcription factors, with a very poor understanding of the bZIP regulatory network and bZIP transcription factors regulate plant responses to abiotic stresses through plant hormones [33,34]. For example, Han et al [12] reviewed the structure, classification, biological functions and regulatory mechanisms of bZIP transcription factors are described, and the molecular mechanisms by which bZIP transcription factors regulate the biosynthesis of flavonoids, terpenoids, alkaloids, phenolic acids and lignin are elaborated. Liu et al [13] reviewed the structural features and functional characterisation of the bZIP transcription factor group, the bZIP transcription factor complex and its molecular regulatory mechanisms associated with salt stress resistance, and the regulation of transcription factors in plant salt stress resistance are summarised and discussed. Guo et al [14] reviewed the latest discussions on the interaction of bZIP with other proteins to regulate abiotic stresses, such as cold, heat, drought, and salt. However, some studies have shown that bZIP transcription factors can indirectly affect plant growth and development and tolerance to abiotic stress by participating in the regulation of plant hormone signal transduction. At the same time, the regulatory effect of exogenous plant growth regulators also requires the participation of bZIP transcription factors [35,36]. Other studies have shown that bZIP transcription factors can interact with non-coding RNAs (ncRNAs) to jointly regulate the expression of downstream genes, thus affecting the tolerance of plants under stress conditions [37,38].
In summary, there is little summary literature on the effects of bZIP transcription factors on plant stress tolerance signalling pathways through the hormonal pathway and the cross-cutting network of bZIP transcription factors and ncRNA that promotes normal plant growth and development and enhances abiotic stress in plants. Therefore, it is highly necessary for us to conduct a more comprehensive review of plant bZIP transcription factors to provide a comprehensive perspective for future in-depth studies of bZIP transcription factors. To fill this knowledge gap, this review firstly introduces the structure, classification and evolution of bZIP transcription factors. Secondly, the roles of bZIP transcription factors involved in plant hormones in plant response to abiotic stresses are summarised. Thirdly, we systematically summarised the molecular mechanisms of bZIP transcription factors involved in plant resistance to various abiotic stresses. Finally, the effects of bZIP transcription factors on the biosynthesis of secondary metabolites in plants as well as the cross-network between bZIP transcription factors and ncRNAs for normal growth and development and enhancement of abiotic stresses are discussed. The current research problems are also presented and their future research directions are envisioned. In this sense, it can serve as an effective reference point for understanding the regulatory network of plant bZIP transcription factors and their related functions, providing valuable insights for stress tolerance breeding applications and theoretical support for future global food security.
1.1. The Structure of the Plant bZIP Transcription Factors
As one of the largest transcription factor families in plants, bZIP transcription factors are widely present in various eukaryotic organisms. To date, scientists have discovered and identified members of the bZIP gene family in many plants, most of which are involved in plant growth and development, abiotic stress, biotic stress, and the regulation of secondary metabolism in plants [39,40]. The bZIP transcription factors are named after the conserved bZIP structural domain and have a relatively conserved structure consisting of 60-80 amino acids, and the structural domain consists of two specialised structures, including a basic region and a leucine zip region [41,42]. The basic region is usually a nuclear localisation signal consisting of about 16 amino acids followed by an invariant N-X7-R/K sequence, which is responsible for recognising and binding to specific sequences on the promoter to perform biological functions, and it is a highly conserved region [43,44,45,46]. As shown in the figure, the bZIP domain is divided into two closely adjacent parts. The first is a highly conserved basic region consisting of approximately 20 amino acid residues, which contains a nuclear localisation signal and an N-X7-R/K motif capable of binding to DNA; the second is a leucine zipper region, where each repeat unit consists of 7 amino acid residues, with leucine or a related hydrophobic amino acid at position 7, and the number of repeat units varies from 3 to 8 (Figure 2A). Each bZIP domain forms a continuous α-helix, and the α-helices of two bZIP proteins interlock due to the hydrophobicity of their side chains, forming a zipper structure (Figure 2B) [24,47]. bZIP proteins must form homodimers or heterodimers to function properly.
1.2. The Classification of the Plant bZIP Transcription Factors
The classification method of bZIP transcription factor is relatively diverse and varies among different species (Table 2). The number of bZIP members in different species ranges from 9 to 227, and the number of bZIP subfamilies is between 10 and 20, with only a few species having bZIP subfamilies smaller than 10 or larger than 20. The functions of different subfamilies vary greatly, but these bZIP transcription factors play very important roles in plant stress response and growth and development [48,49,50,51].
Currently, classification is primarily based on amino acid richness, DNA binding sites, conserved sequences, and phylogenetic relationships. Classification based on specific amino acid richness divides the bZIP family in Brassica rapa into 9 groups, each of which is rich in glutamine (Q), aspartic acid (D), proline (P), asparagine (N), serine (S), glycine (G), and those with only a simple domain, no simple domain, or only one transmembrane domain [48]. Based on the amino acid sequences of the bZIP basic structure domain and hinge region, as well as the DNA-binding sites, bZIP genes in rice can be classified into 11 groups, I–XI, with each group containing 7, 3, 6, 22, 3, 14, 16, 3, 11, 3, and 1 OsbZIP genes, respectively [49]. Tomato bZIP genes can be divided into 9 groups, A-I, containing 16, 12, 6, 2, 12, 2, 3, 4, and 12 SlbZIP genes, respectively [52]. Grape (Vitis vinifera) bZIP genes can be divided into 11 groups: A-I, J, and U, with each group containing 13, 3, 10, 5, 4, 6, 2, 1, 2, 7, and 2 VvbZIP genes, respectively [53]. According to the specificity of the conserved basic structural region binding to DNA, plant bZIP proteins can be classified into four types, namely, G-box-binding bZIP proteins, C-box-binding bZIP proteins, bZIP proteins binding to both G-box and C-box, and bZIP proteins that do not form dimers with DNA [54].
By selecting Arabidopsis thaliana L., Zea Mays and other species, a number of amino acid sequences were found in bZIP family transcription factor proteins, and therefore it was hypothesized that these amino acid sequences might be bZIP family transcription factors conserved amino acid sequences. As shown in the Figure S1, amino acid sequences KTLRRLAQNREAARKSRLRKKAYIQNLET is a conserved amino acid sequences of the bZIP family transcription factors (Supplementary Figure S1). To investigate the phylogenetic and evolutionary relationships of bZIP transcription factors in different species, we constructed phylogenetic trees of the amino acid sequences of some bZIP TF in Arabidopsis thaliana, Oryza sativa, Glycine max, Panicum virgatum, Phragmites australis and Sorghum bicolor. using MEGA 11.0 software [55]. Among them, we found that OsbZIP54 and OsbZIP56 and OsbZIP24 were highly related, PvTGAL9-Like isoform X1 and PvTGAL9-Like isoform X4 were highly related, PvLG2 isoform X1 and PvLG2 isoform X2 were highly related, AtTGA9 and AtT27G7.2 were highly related, PvPVAP13 and PvTGAL4-Like isoform X3 were highly related. At the same time, we also found that MYB transcription factors between different species have strong phylogenetic relationships. Such as, PhTGA2.3-Like isoform X1, PhTGA2.3-Like isoform X2 and OsbZIP64 were highly related, SbTGAL4 isoform X5 and MfTGAL4-Like isoform X3 were highly related, it is possible that bZIP TF have similar functions in different species. The bZIP TF are involved in regulating plant growth and development, secondary metabolism and response to abiotic and biotic stress (Supplementary Figure S2).
1.3. The Mechanism of Action of bZIP Transcription Factors
Three of the four core nucleotide flanking bases determine the binding specificity of plant bZIP transcription factor proteins. They recognise the cis-acting element (Abscisic acid responsive elements) ABRE whose core sequence is an ACGT palindrome. bZIP transcription factors are involved in ABA signalling mainly by binding to the promoter regions of genes containing ABRE [56]. The structure of the plant bZIP transcription factor determines its function. bZIP binds preferentially to DNA sequences containing the ACGT core, such as the A-box (TACGTA), the C-box (GACGTC), and the G-box (CACGTG), and is able to promote or repress downstream expression of relevant abiotic stress genes, which enables the plant to survive in complex abiotic environments better [57]. Additionally, some bZIP transcription factors can also bind to non-palindromic DNA sequences. For example, other non-palindromic cis-acting elements such as the PB (TGAAAA) element and the H-box (CCTACC) element [58]. The GmbZIP44 transcription factor in soybeans can specifically bind to the cis-element GLM (GTGAGTCAT) [59].
The bZIP transcription factors can alter their specificity and affinity for DNA binding through phosphorylation modifications, dimerisation, or interactions with other proteins, while also affecting the stability of bZIPs themselves. bZIP can specifically bind to the promoter sequence of target genes through homodimer or heterodimer forms. The binding site is located in the basic region of bZIP, and this specific binding can activate or inhibit the transcription of target genes [60]. The bZIP TF family members respond to abscisic acid (ABA) signals through cis-acting elements in Arabidopsis, such as ABRE. When ABA is recognised by specific receptors, protein kinase II or serine/threonine kinase transduces the ABA signal, phosphorylating bZIP transcription factors to induce the expression of downstream genes. Following post-translational modification, bZIP is activated and participates in responses to various stress signals and plant growth and development. The upstream region of bZIP promoters contains cis-acting elements associated with hormone regulation, such as gibberellic acid (GA), ABA, indoleacetic acid (IAA), and methyl jasmonate (MeJA) response elements, indicating that bZIP expression can be regulated by multiple plant hormones [56,57,58,59,60].
Phosphorylation is an important regulatory mechanism of bZIP transcription factors and plays an important role in the ABA pathway and stress response in plants. The negative charge generated by phosphorylation can modify the conformation of proteins and their repulsive and attractive forces [60,61,62]. When abiotic or biotic stress signals are detected by plants, bZIP transcription factors can be activated by sucrose non-fermenting 1-related protein kinase (SnRK), mitogen-activated protein kinase (MAPK), and calmodulin-domain protein kinases (CDPK), thereby participating in plant stress responses and growth regulation (Figure 3).
Ubiquitination, one of the post-translational modifications of bZIP transcription factors, is also an important mechanism for bZIP biological activity. The degradation of bZIP is mediated by the ubiquitin-26S proteasome. The MdbZIP44 transcription factor is degraded by the ubiquitin-26S proteasome system in Malus domestica, thereby inhibiting the anthocyanin biosynthesis regulated by MdbZIP44 [63]. The bZIP transcription factor can interact with other transcription factors or proteins to regulate downstream gene expression, thereby regulating plant stress resistance by enhancing or inhibiting gene expression. OsbZIP23 can be activated by OsSAPK2 phosphorylation, while OsPP2C49 can inhibit OsbZIP23 transcription, indicating that bZIP transcription factor interacts with these two enzymes to regulate the rice ABA signalling pathway and stress resistance [64]. Zhang et al. reported the ThbZIP1 transcription factor of binds to the cis-elements C-box, G-box, and A-box of stress response gene promoters under ABA, salt, and drought stress, thereby regulating the expression of downstream genes in Tamarix hispida [65].
2. Role of bZIP Transcription Factors Involved Phytohormone in Plant Response to Abiotic Stresses
Phytohormones have an important impact on plant growth and development and on the regulation of plant tolerance to abiotic stresses. It was found that bZIP can indirectly affect plant growth and development as well as abiotic stress by participating in the regulation of phytohormone signalling. Expression of bZIP transcription factor genes is altered in the presence of exogenous phytohormones such as salicylic acid (SA), abscisic acid (ABA), jasmonic acid (JA), cytokinin (6-BA), gibberellin (GA) and ethylene (ET) [16,66,67,68]. Zhang et al. found Expression of the wheat TabZIP1 gene was up-regulated after exogenous JA and ET treatments, whereas there was no significant change in the expression of this gene under the influence of exogenous SA [69]. Han et al. identified 66 putative bZIP transcription factor genes in the genome of Glycyrrhiza uralensis, and quantified the gene expression levels of the 66 bZIP family members under abiotic stress treatments using qRT-PCR, which is responsive to ABA stress, was also found to have a regulatory role in the synthesis of secondary metabolites by Glycyrrhiza uralensis [70]. Wang et al. reported a SAPK10-bZIP72-AOC pathway, through which ABA promotes JA biosynthesis to synergistically inhibit rice seed germination. SAPK10-dependent phosphorylation enhances the stability of the bZIP72 protein as well as its binding ability to the AOC promoter, increasing the transcription of AOC and the concentration of endogenous JA. Blocking JA biosynthesis significantly reduced the sensitivity of ABA to rice seed germination [71,72]. Wang et al. identified some bZIP transcription factor genes from whole genome sequences of V. radiata and V. angularis, respectively. A number of cis-acting regulatory elements (CARE) involved in abiotic stress and phytohormone responses were detected in the putative promoter region of the bZIP genes. To analysis of bZIP genes expression profiles under drought, salt and ABA stress using quantitative real-time PCR revealed that most of the bZIP genes had altered expression [73]. Zhang et al. identified a novel TabZIP14-B gene involved as a positive regulator in mediating abiotic stress response in wheat. TabZIP14-B is transcriptionally activated and binds to the ABA response element (ABRE) in yeast. TabZIP14-B was expressed in roots, stems, leaves, and young spikes and was up-regulated by treatments with exogenous ABA, salt, freezing temperature, and polyethylene glycol (PEG) stress. Overexpressing TabZIP14-B showed enhanced tolerance to salt, freezing stress, and ABA sensitivity in Arabidopsis [74]. Zhou et al. Identified 66 bZIP family members in Bletilla striata. Some of the BsbZIPs were highly expressed under low temperature, injury, oxidative stress and GA treatment. In addition, BsbZIP13 was located in the nucleus. Yeast two-hybrid (Y2H) and bimolecular fluorescence complementation (BiFC) experiments showed that BsbZIP13 could interact with multiple BsSnRK2 [75]. A total of eight gene family members were identified in cucumber (Cucumis sativus L.) by Lu et al. All eight CsABF/AREB genes were responsive to ABA and JA, with CsABF7 showing the highest expression under ABA and Me-JA treatments. Drought and salt stress significantly induced the expression of CsABF1, CsABF2, CsABF7 and CsABF8. Drought and NaCl stress significantly induced the expression of CsABF1, CsABF2, CsABF7 and CsABF8 [76]. Chai et al. identified a soybean bZIP gene GmbZIP60. GmbZIP60 is located in the nucleus. GmbZIP60 is induced by salt stress, drought stress, and various phytohormone treatments, including ABA, ET, and JA. Overexpression of GmbZIP60 in transgenic soybean and rice enhanced their tolerance to salt stress and drought stress. Chromatin immunoprecipitation-qPCR (ChIP-qPCR) analysis further confirmed that GmbZIP60 could directly bind to the promoters of abiotic stress-related genes induced by ABA, ET, JA and SA [77]. He et al. isolated a bZIP gene GmbZIP19 from soybean, and the expression of GmbZIP19 was significantly induced by ABA, JA, and SA, but decreased under salt and drought stress conditions. By constructing a soybean transient GmbZIP19 overexpression line and performing chromatin immunoprecipitation experiments, and found that GmbZIP19 was able to bind to the promoters of marker genes induced by ABA, JA, ET and SA in soybeans [78]. In conclusion, bZIP TF genes expression are altered under the regulation of plant hormone, such as SA, ABA, JA, 6-BA and ET.
3. Role of bZIP Transcription Factors in Response to Abiotic Stresses
Abiotic stress has a significant impact on plant growth, development, and yield. These stresses disrupt plant physiological and biochemical processes, leading to growth inhibition, metabolic disorders, and even cellular damage and death [79,80]. To cope with abiotic stress, plants have evolved a series of complex adaptive mechanisms, including osmotic regulation, reactive oxygen species (ROS) scavenging, hormone regulation, and gene expression regulation [81]. Therefore, to understand the mechanisms of abiotic stress is of great importance for protecting crop yields. With the continuous improvement of whole-genome sequencing technology, bZIP transcription factors have been found to play an important role in abiotic stress in many higher plants, such as Oryza sativa, Zea mays, and Glycine max (Figure 4) (Table 3).
3.1. bZIP Transcription Factors in Response to Drought Stress
Drought has a huge impact on plants. It can hinder plant growth, reduce yields, and affect physiological processes such as photosynthesis and water metabolism. In addition, drought can alter soil microbial communities, affecting interactions between plants and microorganisms, increasing plant mortality and the risk of ecosystem fires [82,83]. Pan et al. isolated the bZIP family gene SlbZIP38 from tomato (Solanum lycopersicum cv. Ailsa Craig). SlbZIP38 is differentially expressed in different organs of tomato plants and is downregulated by drought, salt stress, and ABA. Overexpression of SlbZIP38 plants significantly reduces tomato tolerance to drought and salt stress. Overexpression of SlbZIP38 plants reduces chlorophyll and free proline content in leaves but increases malondialdehyde content [84]. Li et al. identified the soybean bZIP gene GmTRAB1, whose transcripts are upregulated under drought, ABA, and oxidative stress conditions. Overexpression of GmTRAB1 enhanced the tolerance of transgenic Arabidopsis and soybean root hairs to osmotic stress. However, RNAi of GmTRAB1 expression in soybean root hairs increased their drought sensitivity. GmTRAB1 also enhances the sensitivity of transgenic plants to ABA and regulates ABA-mediated stomatal closure under drought stress. Additionally, GmTRAB1 stimulates the transcriptional accumulation of drought, ABA and antioxidant-related genes in response to drought stress [85]. Wang et al. identified a bZIP transcription factor named CmbZIP9 from the leaf transcriptome of Chrysanthemum mongolicum and found that overexpressing CmbZIP9 in tobacco reduced malondialdehyde (MDA) content under drought stress, increased peroxidase (POD) and superoxide dismutase (SOD) activity, and elevated the expression levels of stress-related genes, thereby enhancing the drought tolerance of the transgenic tobacco lines [86]. Wu et al. identified a bZIP transcription factor named EDT1 from Oryza sativa. The EDT1 protein contains a bZIP domain, a putative nuclear localization signal (NLS), and six conserved phosphorylation sites. Overexpressing EDT1 rice plants exhibited significantly enhanced drought tolerance and the expression of stress-related genes such as OsbZIP12, SNAC1, OsLEA3, OsbZIP16, OsbZIP10 and OsABI2 was upregulated in EDT1-overexpressing rice lines. These results indicate that EDT1 plays a positive role in rice drought tolerance [82].
3.2. Molecular Mechanisms of bZIP Transcription Factors Associated with Salt Stress
The visible symptoms of salt damage are chlorosis of leaf tips, followed by leaf scorching, browning, and leaf death. This leads to slow plant growth, poor root development, sterility, and reduced seed yield. Excessive salt content causes physiological disorders in plants and can even lead to death (Figure 5) [10,87]. He et al. cloned a new F-bZIP transcription factor ZmbZIP76 from the maize inbred line He344. The expression of ZmbZIP76 in maize is significantly induced by high salt, osmotic stress and ABA. Overexpression of ZmbZIP76 enhances the tolerance of transgenic plants to salt and osmotic stress. ZmbZIP76 upregulates the expression of a series of non-biotic stress response genes by binding to ACGT elements, thereby enhancing ROS scavenging capacity, increasing abscisic acid levels, proline content, K+/Na+ ratios, reducing water loss rates, and minimizing membrane damage, thereby enhancing plant salt tolerance [88]. Wei et al. found that JrbZIP40 is strongly induced under both drought and salt stress. JrbZIP40 is localized to the cell nucleus and possesses transcriptional activation activity. Overexpression of JrbZIP40 in transgenic Arabidopsis seedlings significantly enhanced their resistance to salt and drought stress. JrbZIP40 can bind to the promoters of JrHB7 and JrATG8G and activate their expression, thereby enhancing the plant's resistance to salt and drought stress. The results elucidate the regulatory network and biological functions of JrbZIP40 in drought and salt tolerance [89]. Wang et al. identified and analyzed 49 bZIP transcription factor genes in the potato genome, several of which are induced under different stress conditions. The expression of StbZIP25, which is homologous to the AtbZIP36/ABF2 gene in Arabidopsis, was significantly upregulated under salt stress treatment. The StbZIP25 protein is localized in the cell nucleus and functions as a transcription activator. Overexpression of StbZIP25 enhances the salt tolerance of Arabidopsis [90]. Liu et al. identified an OsbZIP71 transcription factor from rice. OsbZIP71 can bind to the G-box motif but lacks transcriptional activity. Drought, polyethylene glycol (PEG), and ABA treatments strongly induced the expression of OsbZIP71, while salt treatment inhibited its expression. Overexpressing-OsbZIP71 plants showed significantly enhanced tolerance to drought, salt, and PEG osmotic stress. RNAi knockdown Osbzip71 lines had the opposite effect. Some abiotic stress-related genes OsVHA-B, OsNHX1, COR413-TM1 and OsMYB4 were upregulated in the overexpressed OsbZIP71 lines but downregulated in the Osbzip71 lines. OsbZIP71 can directly bind to the promoters of OsNHX1 and COR413-TM1 in vivo. These results suggest that OsbZIP71 may play an important role in rice drought and salt tolerance [91].
3.3. bZIP Transcription Factors Involved in Plant Response to Temperature Stress
Temperature is a major environmental factor affecting plant growth and development and fruit quality after harvest. Suitable temperatures are conducive to plant growth. Both high and low temperatures can cause varying degrees of stress on plants, affecting physiological processes such as photosynthesis, respiration, and water metabolism, which in turn impact plant growth, flowering, fruiting and yield [92,93,94].
3.3.1. bZIP Transcription Factors and High-Temperature Stress
When temperatures exceed the optimal temperature, the enzyme systems involved in physiological and biochemical reactions are impaired, metabolic activity is hindered, and this in turn affects normal plant growth and development [95]. Shen et al. reported that the bZIP family member CabZIP63 regulates the expression of CaWRKY40 in Capsicum annuum. Silencing the CabZIP63 gene in pepper plants significantly reduced their resistance to ralstonia solanacearum inoculation (RSI) and tolerance to high temperature-high humidity (HTHH), accompanied by downregulation of immunity or heat-related genes CaPR1, CaNPR1, CaDEF1 and CaHSP24. Overexpression of CabZIP63 plants induced allergen-mediated cell death and increased the expression of the detected immunity and heat-tolerance-related marker genes. In conclusion, CabZIP63 is involved in plant immunity or heat-tolerance biological processes [96]. Chen et al. identified OsbZIP33 is significantly upregulated under drought, high salinity, H₂O₂ and high-temperature stress conditions in Oryza sativa L.. Overexpressing-OsbZIP33 plants exhibited significantly enhanced drought tolerance. Under drought conditions, the expression levels of representative downstream drought-induced genes in transgenic rice plants were significantly higher than those in wild-type plants. Studies indicate that OsbZIP33 is induced by exogenous ABA. Overexpression of OsbZIP33 increases the sensitivity of transgenic plants to ABA compared to wild-type plants. In summary, the results indicate that OsbZIP33 regulates rice drought tolerance and plant tolerance under high-temperature conditions [97]. Qiu et al. found that OsbZIP14 is a key heat-responsive transcription factor gene. After knocking out Osbzip14 gene in the rice (ZH-11), the knockout mutant exhibited dwarfism and reduced tillering during the grain filling stage. Under high-temperature treatment, the expression of OsbZIP58, a key gene regulating the accumulation of rice seed storage proteins (SSPs), was upregulated in Osbzip14 mutants. Additionally, OsbZIP14 directly can interact with OsbZIP58. The results indicate that under high-temperature stress, OsbZIP14 regulates plant tolerance to high temperatures through synergistic interactions with OsbZIP58 and OsbZIP14 [98]. Jiang et al. demonstrated that the nuclear-localized proteins BdibZIP44 and BdibZIP53 are involved in heat tolerance mechanisms. These proteins exhibit inducible expression under heat stress conditions in Brachypodium distachyon. Overexpression BdibZIP44 and BdibZIP53 plants enhanced antioxidant enzyme activity including catalase (CAT), superoxide dismutase (SOD), and peroxidase (POD), reinforcing their positive regulatory role in adapting to high-temperature stress. These antioxidants collectively enhance the removal of harmful ROS. In summary, BdibZIP44 and BdibZIP53 play a crucial role in regulating the robust defense mechanisms that enable plants to withstand high-temperature stress [99].
3.3.2. bZIP Transcription Factors and Low-Temperature Stress
When environmental temperatures are between the minimum and optimal temperatures, physiological and biochemical reactions within plants accelerate as temperatures rise, metabolic activity increases, thereby accelerating growth and development (Figure 5) [100,101]. A bZIP transcription factor named CsbZIP18 was found to be strongly induced under cold conditions by Yao et al. Compared with wild-type, overexpressing-CsbZIP18 plants showed reduced sensitivity to ABA, increased relative electrolyte leakage (REL), and decreased maximum quantum efficiency of photosystem II (Fv/Fm) under freezing conditions in Arabidopsis. Under cold stress, the expression of ABA homeostasis and signal transduction-related genes, as well as the abiotic stress-induced genes RD22, RD26 and RAB18, are suppressed in the overexpression lines. These results suggest that CsbZIP18 negatively regulates plant freezing tolerance via an ABA-dependent pathway [102]. Liang et al. identified the TabZIP transcription factor TaABI5 in wheat, whose ectopic expression upregulated the expression of multiple cold-response genes in Arabidopsis. TaABI5 can enhance the plant's cold tolerance. TaABI5 is located in the cell nucleus and TaABI5 interacted with the cold-tolerance transcription factor TaICE1 in a yeast two-hybrid system. In summary, this study demonstrates that the TaABI5 influences the cold tolerance of plants [103]. Liu et al. isolated a member of the bZIP transcription factor family OsbZIP52/RISBZ5 from Oryza sativa. The expression of OsbZIP52 gene is strongly induced by low temperatures. OsbZIP52 is a nuclear-localized protein. OsbZIP52 functions as a transcription activator and can specifically bind to the G-box promoter motif. Overexpression of OsbZIP52 plants exhibit significantly enhanced sensitivity to cold and drought stress. Several abiotic stress-related genes, including OsLEA3, OsTPP1, Rab25, are downregulated in OsbZIP52-overexpressing lines. In conclusion, OsbZIP52/RISBZ5 may play a negative regulatory role under cold and drought stress conditions [104]. A cold-induced transcription factor, ABF1 was identified in the bZIP family by Shu et al. Knockout of abf1 resulted in reduced cold tolerance, increased ion leakage, and decreased proline content in rice, while ABF1-overexpressing lines exhibited the opposite trend. The SnRK2 protein kinase SAPK10 can phosphorylate ABF1, enhancing its DNA-binding ability to the G-box cis-element of the TPS2 promoter, thereby increasing TPS2 transcription levels and endogenous trehalose content. Exogenous trehalose application also enhances the cold tolerance of abf1 mutant lines. In summary, a new pathway “SAPK10-ABF1-TPS2” regulates trehalose homeostasis to improve rice cold tolerance [105].
3.4. Role of bZIP Transcription Factors in Response to Nutritional Element Stress
Plant growth and health depend on adequate and balanced nutrition. Nutrition affects plants in many ways, including growth, development, disease resistance, yield and quality. Nutritional deficiencies can lead to stunted growth, discoloured leaves, reduced fruit production, and even plant death. Excessive nutrition can also be harmful to plants, affecting their normal growth and development [106,107]. Zinc has a significant impact on plants. It indirectly influences the synthesis of auxin within crops. When crops are deficient in zinc, the auxin content in stems and buds decreases, growth stagnates, and plants become stunted. Additionally, zinc acts as an activator for many enzymes, exerting a broad influence on plant carbon and nitrogen metabolism, thereby aiding photosynthesis. Furthermore, zinc enhances plants stress tolerance. Increases seed weight and alters the seed-to-stem ratio [108,109]. Assunção et al. identified the bZIP transcription factor genes bZIP19 and bZIP23 in Arabidopsis, which regulate plant adaptation to low zinc supply. bZIP19 and bZIP23 are associated with the promoter regions of the ZIP4 gene, which belongs to the zinc-responsive transporter ZIP family induced by zinc deficiency. The bzip19/bzip23 double mutant is highly sensitive to zinc deficiency. This is because the bzip19/bzip23 mutant cannot induce the expression of a small group of genes that constitute the primary response to zinc deficiency, including additional ZIP metal transporter genes. In summary, the AtbZIP19 and AtbZIP23 transcription factors are involved in zinc regulation in higher plants [110]. Lilay et al. discovered that the bZIP19 and bZIP23 transcription factors are central regulatory factors in the zinc deficiency response in Arabidopsis. Their targets include members of the ZIP (Zrt/Irt-like protein) transporter family, which are involved in cellular zinc absorption and are upregulated in response to zinc deficiency. The zinc deficiency-sensitive phenotype of bzip19/bzip23 double mutants indicates that the zinc-dependent activity of bZIP19 and bZIP23 is regulated by zinc at the protein level in the nucleus, while adequate intracellular zinc suppresses their activity. Additionally, bZIP19 and bZIP23 transcription factors exhibit significant functional redundancy [111].
Iron is an essential trace element for plant growth and participates in many important physiological processes in plants, including chlorophyll synthesis, photosynthesis, respiration, and enzyme activity. Iron has poor mobility within plants, and iron deficiency can cause symptoms such as leaf yellowing and shoot death [112]. Mankotia et al. reported that the bZIP transcription factor family member HY5 (Elongated Hypocotyl 5) plays an important role in the iron deficiency signaling pathway of Arabidopsis. hy5 mutants fail to produce an optimal iron deficiency response and exhibit root growth defects under iron-deficient conditions. Compared to WT plants under iron-deficient conditions, the induction of genes involved in the iron absorption pathway (FIT-FER-LIKE IRON DEFICIENCY-INDUCED TRANSCRIPTION FACTOR, FRO2-FERRIC REDUCTION OXIDASE 2, and IRT1-IRON-REGULATED TRANSPORTER1) is reduced in hy5 mutants. These results indicate that bZIP transcription factor HY5 plays an important role in regulating the iron deficiency response in Arabidopsis [113].
3.5. bZIP Transcription Factors Involved in Plant Response to Heavy Metals Stress
The effects of heavy metals on plants are mainly reflected in their physiological and biochemical activities, growth and development, and ultimately their yield and quality [114]. Under heavy metal stress, plants undergo a series of physiological and biochemical reactions, including reduced antioxidant enzyme activity, membrane lipid peroxidation, and cell apoptosis. These reactions affect plant growth and metabolism, leading to symptoms such as stunted growth, leaf chlorosis, and dwarfism, and may even result in plant death [115]. A bZIP gene BnbZIP2 was cloned from ramie by Huang et al. The BnbZIP2 protein is localized in both the nucleus and cytoplasm. BnbZIP2 expression is significantly higher in female and male flowers, and its expression is induced by drought, high salt and ABA treatment. Transgenic overexpressing-BnbZIP2 Arabidopsis plants exhibit higher sensitivity to drought and cadmium stress during seed germination, while their tolerance to high salt stress during seed germination and plant development is higher than that of WT plants. Therefore, the BnbZIP2 can be involved in plant regulation of cadmium [116]. Sedum plumbizincicola is a cadmium (Cd)/zinc (Zn)/lead (Pb) hyperaccumulator native to southeastern China, with potential application value in plant-based remediation of heavy metal-contaminated soils. Lu et al. identified hormone-responsive, abiotic and biotic stress-responsive, and plant development-related cis-acting elements in the SpbZIP promoter sequences, and most SpbZIP genes showed upregulated expression levels under cadmium stress conditions. SpbZIP60 is localised in the cell nucleus. Overexpression of SpbZIP60 enhances the cadmium tolerance of transgenic Arabidopsis by inhibiting ROS accumulation, protecting photosynthetic organs, and reducing cadmium content [117]. Chai et al. identified a bZIP transcription factor gene GmbZIP152, which is significantly induced in soybeans under stress conditions such as Sclerotinia sclerotiorum, plant hormones, salt, drought, and heavy metal stress. Overexpression of GmbZIP152 enhances soybean resistance to Sclerotinia sclerotiorum and tolerance to salt, drought, and heavy metal stress in Arabidopsis. Compared to the WT, the antioxidant enzyme-related genes AtCAT1, AtSOD and AtPOD1 and their enzymatic activities in OE-GmbZIP152 are induced by Sclerotinia sclerotiorum, salt, drought and heavy metal stress. In conclusion, GmbZIP152 plays a crucial role in soybean response to heavy metal stress [118]. Hou et al. identified four single nucleotide polymorphisms (SNPs) and 30 candidate genes associated with Pb tolerance in maize seedlings. The ZmbZIP107 transcription factor was identified as a key gene controlling Pb tolerance in maize. Two 5'UTR variants of ZmbZIP107 affect its expression levels and Pb tolerance across different maize varieties. The ZmbZIP107 protein is specifically localized to the cell nucleus and ZmbZIP107 mRNA expression is highest in the root system of maize seedlings. Heterologous expression of ZmbZIP107 enhances rice tolerance to lead stress and reduces lead absorption by the root system [119]. These data indicates that bZIP transcription factor can suppress ROS accumulation in the heavy metal stress response of plants, reduce intracellular heavy metal content and alleviate the damage caused by heavy metals to plants.
3.6. bZIP Transcription Factors Involved in Plant Response to High Light Stress
High light has a dual effect on plants. On the one hand, light is the basis for photosynthesis in plants, providing them with energy and promoting growth. On the other hand, excessive or inappropriate light can damage plants, such as leaf scorching and sunburn on fruits, and even inhibit their growth and development [120,121]. Liu et al. identified 56 mulberry bZIP transcription factors and classified them into 12 groups through phylogenetic analysis. Numerous stress response elements were identified in the promoter regions of bZIP genes. Additionally, by analyzing the expression profiles of RNA sequencing data, MabZIP43 and MabZIP24 were identified as responding to high temperature, saline-alkali, and strong light stress, respectively. The results indicate that the bZIP gene family plays a significant role in stress responses [122]. Stracke et al. have discovered that the bZIP transcription factor ELONGATED HYPOCOTYL5 (HY5) is essential for the transcriptional activation of the PRODUCTION OF FLAVONOL GLYCOSIDES(PFG)PFG1/MYB12 and PFG3/MYB111 genes under ultraviolet (UV)-B and visible light conditions. A synthetic protein composed of HY5 and the VP16 activation domain is sufficient to activate the expression of PFG1/MYB12 in plants. However, despite the significant reduction in CHS levels in the myb11/myb12/myb111 triple mutant under dark and constant light conditions, leading to reduced UV-B tolerance, overexpression of PFG1/MYB12 enhances tolerance [123]. AtbZIP16 and AthZIP68 are two putative G-group bZIP transcription factors in Arabidopsis, and the other three members of the G-group bZIPs are GBF1-3, which can bind to G-boxes. Shen et al. reports that AtbZIP16 and AthZIP68 can bind to cis-elements containing the ACGT core sequence. Additionally, AtbZIP16 and AtbZIP68 can form homodimers and also form heterodimers with other members of the G group. AtbZIP16 and AthZIP68 are localized to the nuclei of onion epidermal cells. In Arabidopsis, AthZIP16 and AthZIP68 may also be involved in the light-responsive processes mediated by GBF1-3 [124]. In summary, these results indicate that bZIP transcription factors can regulate plant responses to light stress either independently or in interaction with other proteins, with the aim of improving plant tolerance to light stress.
4. bZIP Transcription Factors Mediated Control of the Plant Secondary Metabolite Biosynthetic Pathways
The bZIP family transcription factors not only play important roles in regulating plant growth and development and abiotic stress response, but also participate in the regulation of plant primary and secondary metabolites, such as flavonoids, terpenoids, alkaloid, phenolic acids and lignans [125,126,127,128,129]. Plant secondary metabolites are diverse in type and structure, and can generally be classified into three major categories: terpenoids, phenolics (such as flavonoids), and nitrogen-containing compounds (such as alkaloids) [130,131]. Secondary metabolites perform various complex biological functions, playing crucial roles in plants responses to environmental stress, interspecific competition and coevolution, attracting insects for pollination, and defending against predator attacks [132]. We will discuss in detail the research progress on the biosynthesis of these secondary metabolites regulated by bZIP transcription factors (Table 4).
4.1. bZIP Transcription Factors Are Involved in Flavonoids Biosynthesis in Plants
Flavonoids play a significant role in the prevention and treatment of cardiovascular and cerebrovascular diseases. Many studies have shown that bZIP transcription factors can participate in the biosynthesis pathway of flavonoids (Figure 6) [133]. Han et al. isolated and identified the GbbZIP08 gene in Ginkgo biloba. GbbZIP08 is localized in the plant cell nucleus. The total flavonoid, kaempferol, and anthocyanin content in transgenic tobacco was significantly higher than that in the WT. The expression of structural genes in the flavonoid biosynthetic pathway was significantly upregulated. In summary, GbbZIP08 overexpression promotes flavonoid accumulation in plants and increases the transcriptional levels of genes related to flavonoid synthesis [134]. Zhou et al. identified 62 DcbZIP genes in Dendrobium catenatum, grouped into 13 subfamilies. Among these, the expression of DcbZIP3, DcbZIP6, DcbZIP28 was induced by low temperatures in both leaves and stems. DcbZIP3, DcbZIP6, DcbZIP28 contains the bZIP conserved domain and is localized to the cell nucleus. DcbZIP6 shows a significant negative correlation with PAL2 (palmitoyl-CoA), which is involved in flavonoid metabolism. Additionally, DcbZIP28 shows a significant negative correlation with multiple metabolism-related genes in the polysaccharide metabolism pathway, including 6-phosphofructokinase PFKA1, aldose-6-phosphate reductase ALDO2 and fructose kinase SCRK5. These results suggest that DcbZIP6 and DcbZIP28 primarily participate in flavonoid or polysaccharide metabolism [135]. Han et al. screened the genome-wide distribution of bZIP transcription factors from the Ginkgo biloba L. database, identifying a total of 40 bZIP genes, which were classified into 10 subclasses. GbbZIP08 and GbbZIP15 exhibited the highest expression levels in mature leaves, while GbbZIP13 showed high expression levels in all eight tissues. Correlation analysis and phylogenetic tree analysis indicated that GbbZIP08 and GbbZIP15 are involved in flavonoid biosynthesis. Among them, GbbZIP08 showed significantly up-regulated expression at 4°C. In summary, GbbZIP08 and GbbZIP15 are involved in regulating flavonoid biosynthesis and abiotic stress responses [136]. Zhang et al. cloned and functionally characterized the receptor-like kinase OsRLCK160 and the bZIP transcription factor OsbZIP48, which positively regulate the accumulation of flavonoids, through a metabolite-based genome-wide association study. OsRLCK160 phosphorylates OsbZIP48, thereby regulating flavonoid accumulation and participating in rice UV-B tolerance. These studies indicate that OsRLCK160 and OsbZIP48 play a significant role in regulating flavonoid accumulation in rice [137].
4.2. bZIP Transcription Factors Are Involved in Terpenoids Biosynthesis in Plants
Terpenoids are essential components for plant growth and development and also play a role in plants adaptation to and defense against environmental stress [138]. Terpenoids, such as monoterpenes and diterpenes, play a crucial role in plants responses to abiotic stressors like drought, salinity, and pest infestations. For example, some terpenoids act as signaling molecules, attracting natural enemies to prey on insects that feed on plants, thereby protecting the plants themselves [139]. Michael et al. demonstrated that the elongated hypocotyl (HY5) acts as a bZIP transcription factor and plays a key role in the light-mediated transcriptional regulation of the terpene synthase AtTPS03. Expression analysis of the hy5-215 mutant and HY5 overexpression lines revealed that HY5 functions as a positive regulator of AtTPS03. Electrophoresis mobility shift assay (EMSA) studies showed that HY5 can bind to the promoter of AtTPS03. In summary, HY5 is essential for light-mediated transcriptional regulation of AtTPS03 in Arabidopsis to achieve terpenoid biosynthesis [140]. Xu et al. found that after treatment with the plant hormones MeJA and ABA, the expression level of a TF in Tanacetum cinerariifolium significantly increased, and it was named TcbZIP60. TcbZIP60 is localized in the cell nucleus and can directly bind to the E-box/G-box motifs on the promoters of the pyrethrin synthesis genes TcCHS and TcAOC, thereby activating their expression. The transient overexpression of TcbZIP60 increased the expression levels of the pyrethrin synthesis genes, leading to a significant accumulation of pyrethrin. Silencing Tcbzip60 significantly downregulates pyrethrin accumulation and the expression of its related genes. In summary, TcbZIP60 regulates the terpenoid and MeJA pathways involved in pyrethrin synthesis [141]. Wu et al. identified 105 SrbZIP genes at the whole-genome level and classified them into 12 subfamilies using bioinformatics methods in Stevia rebaudiana Bertoni. Three major cis-acting elements were identified in the SrbZIP promoter region, including development-related elements, defense and stress response elements. Additionally, qRT-PCR analysis was performed to assess the expression levels of 15 SrbZIP genes under various treatments, including light exposure, abiotic stress, plant hormones and different tissues. Among these, three SrbZIP genes SrbZIP54, SrbZIP63 and SrbZIP32 were predicted to be associated with the synthesis of terpenoid compounds involved in stress responses in Stevia rebaudiana. In conclusion, the bZIP TF SrbZIP54, SrbZIP63, and SrbZIP32 can play important regulatory roles in terpenoid compound synthesis [142].
Artemisinin, a secondary metabolite accumulated in Artemisia annua, is a sesquiterpene compound that can treat malaria. Zhang et al. identified six members of the a subfamily from 64 bZIP transcription factors highly expressed in glandular hairs, named AabZIP1 to AabZIP6. When these transcription factors and the ADS and CYP71AV1 genes were transformed into tobacco leaves, it was found that AabZIP1 could directly bind to the ABRE promoter element of the ADS and CYP71AV1 genes and activate the expression of the two key enzyme genes [143]. Overexpression of AabZIP1 significantly increases artemisinin content, indicating that the transcription factor AabZIP1 positively regulates artemisinin synthesis. Dual luciferase assay results showed that ABA enhances AabZIP1's transcriptional activity on the ADS and CYP71AV1 promoters. The ABA receptor protein AaPLY9 was isolated from Artemisia annua plants where AabZIP1 was overexpressed and artemisinin content was significantly increased, indicating that AabZIP1 plays a key role in ABA-induced artemisinin synthesis (Figure 7) [143,144].
4.3. bZIP Transcription Factors Are Involved in Alkaloid Biosynthesis in Plants
Alkaloids are an important class of secondary metabolites produced by plants during long-term evolution. They have multiple physiological and ecological functions for plants, including regulating cell proliferation and differentiation, influencing plant morphogenesis, responding to environmental stress, and defending against pests and diseases [145,146]. A total of 65 IibZIP genes were identified in the Isatis indigotica genome, distributed across seven chromosomes with high conservation and classified into 11 subgroups. Among these, 17 IibZIP genes were co-expressed with 79 transcription factors, with most of these genes associated with plant responses to abiotic stress and hormone signaling. The 17 IibZIP genes are associated with the regulation of 110 metabolites. Additionally, IibZIP23, IibZIP38 and IibZIP51 are closely associated with six metabolites, including three alkaloids (quinoline alkaloid Stylopine, indole alkaloid Tabersonine, and indole-3-acetic acid), the flavonoid myricetin 3-O-galactoside, and two major metabolites, 2-hydroxy-6-aminopurine and 3-dehydroquinic acid [147]. Overexpression of CrMYC2 induces the expression of the bZIP transcription factor CrGBF and reduces the accumulation of alkaloids in the hairy roots of Catharanthus roseus. Meanwhile, CrGBF1 and CrGBF2 form homodimers and heterodimers, thereby inhibiting the transcriptional activity of key terpenoid indole alkaloids (TIA) pathway gene promoters. The reason is that CrGBFs may antagonize the transcriptional activation of CrMYC2 by competitively binding to the T/G box in the target promoter and/or by forming non-DNA-binding complexes through protein-protein interactions, thereby preventing CrMYC2 from binding to its target promoter [148]. In summary, bZIP transcription factors can regulate the biosynthesis of terpenoid compounds, laying the foundation for future research on secondary metabolism genetic engineering in medicinal plants.
4.4. bZIP Transcription Factors Are Involved in Phenolic Acids in Plants
Phenolic acids have multiple effects on plants, including antioxidant properties, defense mechanisms, UV protection, structural support and allelochemical effects. They can influence plant growth, development, and resistance to adverse conditions. Phenolic acids may also affect the distribution of soil microorganisms, thereby impacting soil quality and ecosystem health [149,150,151]. ABA can effectively increase the content of phenolic acids and tanshinones in Salvia miltiorrhiza, but its regulatory mechanism remains unclear. Shi et al. conducted a deep transcriptomic analysis of ABA-treated Salvia miltiorrhiza root hairs. The expression levels of single genes encoding key enzymes such as PAL, TAT, C4H, RAS, CYP98A14 and GGPPS were significantly higher than those in the control group. Candidate laccases with expression patterns similar to RAS and CYP98A14 in phenolic acid biosynthesis, as well as potential P450 single genes involved in tanshinones biosynthesis, were identified. Additionally, they identified 1,250 TFs representing 35 families, with MYB, AP2/ERF, bHLH, and bZIP being the most abundant. Therefore, transcription factors from these families can participate in the regulation of phenolic acid and tanshinones compounds [152]. Ibáñez et al. analyzed the Prunus salicina Lindl. reference genome and identified 60 genes encoding bZIP proteins. Based on the predicted amino acid sequences, PsabZIP genes were distributed across 12 groups. RNA sequencing analysis of fruits from two Prunus salicina. varieties at different developmental stages detected 53 expressed bZIP-encoding genes, indicating their role in fruit development. Through correlation analysis of bZIP transcription factor expression with nine phenolic compounds, 29 genes were found to be significantly associated with the biosynthesis of these compounds, suggesting that bZIP transcription factors may act as regulatory factors in the biosynthesis of phenolic compounds in Prunus salicina Lindl [153].
4.5. bZIP Transcription Factors Are Involved in Lignin in Plants
Lignin is widely present in the cells of woody plants, herbaceous plants, and all vascular plants, playing a role in strengthening plant tissues. Lignin is the second most abundant plant-based natural substance in nature, following cellulose. Lignin is typically found in the highest concentrations in the trunks of trees, where it serves as an important substance for enhancing plant resilience and keeping trees upright and stable [154,155,156]. Lignin plays a crucial role in defending against drought stress. Tu et al. demonstrated that VlbZIP30 mediates drought tolerance by activating the expression of lignin biosynthesis genes and increasing lignin deposition in grapevine. Overexpressing VlbZIP30 grape plants exhibited lignin deposition under control conditions, which was attributed to the upregulation of VvPRX4 and VvPRX72 expression. Overexpression of VlbZIP30 enhanced drought tolerance, characterized by reduced leaf water loss rate under drought conditions, maintained effective photosynthetic rate, and increased lignin content in leaves. VlbZIP30 directly binds to the G-box cis-elements in the promoters of the lignin biosynthesis gene VvPRX N1 and the drought response gene VvNAC17, thereby regulating their expression. In summary, VlbZIP30 is associated with lignin synthesis and drought tolerance in grapevine [157]. Juice sac granulation (a physiological disorder) causes significant postharvest losses in Citrus maxima. Juice sac granulation is closely associated with lignin accumulation. The results of Li et al.'s study indicate that lignin content in NC (near the seed) and FC (far from the seed) juice sacs generally increases from 157 DPA after flowering to 212 DPA, reaching a maximum at 212 DPA. Additionally, lignin content in NC juice sacs was higher than in FC juice sacs. Following data assembly and bioinformatics analysis, 11 DEGs were identified as hub genes for lignin biosynthesis. Among these, PAL (phenylalanine ammonia-lyase), HCT (hydroxycinnamoyl-CoA shikimate/quinic acid hydroxycinnamoyltransferase), 4CL2 (4-coumaroyl: CoA ligase), C4H (cinnamic acid 4-hydroxylase), C3'H (p-coumaric acid 3-hydroxylase), and CCoAOMT1 (caffeoyl-CoA 3-oxomethyltransferase) were the most differentially expressed DEGs in granulated juice sacs. Co-expression analysis revealed that the expression patterns of transcription factors such as bHLH130 are highly correlated with lignin formation, indicating that bHLH130 can regulate lignin accumulation in plants [158].
5. bZIP Transcription Factors Interact with ncRNAs to Regulate of Abiotic Stress in Plants
Non-coding RNAs (ncRNAs) are RNAs that do not code for proteins, including rRNAs, tRNAs, snRNAs, snoRNAs, LncRNAs, microRNAs (miRNAs) and so on [159,160]. ncRNAs are RNAs with known functions that share the common characteristic of being transcribed from the genome but are not translated into proteins, and are able to carry out their biological functions at the RNA level [161,162]. Baruah et al. identified 12807 Long Non-coding RNAs (lncRNAs) in Capsicum annuum tissues under abiotic stress conditions. 30 lncRNAs were found to act as precursors for 10 miRNAs in Capsicum annuum, and 1807 lncRNAs interacted with 194 miRNAs targeting 621 Capsicum annuum mRNAs. The expected role of LncRNAs under different abiotic stress conditions by interacting with miRNAs and regulating stress-responsive bZIP transcription factors in Capsicum annuum [163]. Baloglu et al. identified 64 genes encoding bZIP transcription factors in the cucumber genome. genome-wide expression analysis of the bZIP transcription factors genes showed that the 64 CsbZIP genes were differentially expressed in at least one of the ten sampled tissues. A total of four CsbZIP genes showed high expression values in leaf, flower and root tissues. Computerised miRNA and target transcript analyses revealed that a total of 21 CsbZIP transcription factors were targeted by 38 plant miRNAs. CsbZIP20 and CsbZIP22 were the most frequently targeted genes by miR165 and miR166 family members, respectively. The expression of ten CsbZIP transcription factors genes in drought-stressed cucumber root and leaf tissues was also analysed by quantitative RT-PCR. The expression of these genes was found to be increased in root tissues at 24 h after PEG treatment [164]. Neysanian et al. evaluated the effectiveness of foliar sprays of selenates and selenium nanoparticles (SeNPs) on tomato plants to study the mitigation of risks associated with water deficit conditions. Tomato plants were treated with SeNPs at concentrations of 0 and 4 mg L-1; after the third spray, the plants were exposed to water deficit conditions. Foliar sprays of SeNPs not only improved growth, yield, and developmental transition to flowering but also mitigated the deleterious risks associated with water deficit conditions. Under normal irrigation conditions, miR172 expression was slightly increased in SeNP-treated plants, whereas miR172 showed a down-regulation under drought stress. The bZIP transcription factor and CRTISO gene were up-regulated after SeNP and drought treatments. Drought stress significantly increased H2O2 accumulation, which was alleviated by SeNPs. Foliar sprays of Se or SeNPs had a similar tendency to alleviate the negative effects of drought stress on membrane integrity [165]. Two small RNA libraries were generated from micropropagated ‘Hamburg muscadine grape’ (V. vinifera) plants under normal and low temperature conditions by Sun et al. Forty-four cold-induced miRNAs were identified by differential expression miRNAs (DEMs) analysis, of which 13 belonged to up-regulated DEMs and 31 to down-regulated DEMs. Prediction of the target genes of DEMs indicated that miRNAs may regulate bZIP transcription factors under cold stress [166]. In summary, the bZIP transcription factor is capable of interacting with ncRNAs and plays an important role in abiotic stress response.
6. Conclusion and Prospects
The bZIP transcription factor is one of the largest transcription factor families in plants. It plays a key role in the response of plants to abiotic stress, plant secondary metabolism and plant growth and development. At present, scientists are mainly focusing on the response of bZIP transcription factors to common abiotic stresses such as drought, cold, heat, salt, heavy metals, nutritional stress and light stress. The structure, classification and function of bZIP transcription factors have been studied extensively. The bZIP transcription factors of different species have been mined and identified using bioinformatics methods, and the evolutionary relationship and functional correlation between bZIP transcription factors of different species have been analysed. A number of important results have been obtained. At the same time, there has been a more detailed understanding of the downstream target genes which are under their control. However, there are several shortcomings in studying the bZIP family transcription factors in plants. For example, there has been little research on the homology and evolution of bZIP transcription factors between different crops, such as Oryza sativa, Glycine max and so on. Therefore, some important bZIP transcription factor genes may be overlooked. The second point is that the current family classification based on the number of bZIP transcription factor structural domains is crude. It lacks a brief description of their functions. In the future, the functional classification should be based on the functional differences of different family members and on the combination of structural and functional classification. This will help to understand the known bZIP transcription factor genes faster and at the same time predict the unknown bZIP genes so that more functions of bZIP transcription factor genes can be discovered. The third point is that, at present, the focus of researchers is only on the function of bZIP transcription factor genes, and in the future, the functional verification of bZIP transcription factor genes will be more important. At present, researchers are only focusing on the function of bZIP transcription factor genes. The future is to combine the functional verification of bZIP transcription factor genes with the actual use of breeding, to apply them to production practice, to improve the resistance of plants, and ultimately to achieve the purpose of high yield.
Up to now, research on MYB transcription factors has been conducted for more than 30 years, but most of the research results are still in the stage of theoretical research. Advanced biotechnological means should be used to conduct in-depth research on the mechanism of bZIP transcription factors regulating the production of plant secondary metabolites, so as to comprehensively improve the resilience of the crop and the economic value of the potential, which will be of great significance to the agricultural economy. Moreover, most of the research on bZIP transcription factors nowadays focuses on agricultural crops as well as fruits and vegetables, while there are relatively few relevant studies in the field of medicinal plants. The slow development of traditional Chinese medicine is largely due to its unclear mechanism of action. If we can form a specific and clear understanding of the mechanism of bZIP transcription factors, it is equivalent to a fundamental understanding of the microscopic composition of traditional Chinese medicine, which will play an important role in the further study of bZIP transcription factors in medicinal plants, and will make the study of its structure, function, and metabolic pathway much easier.
In conclusion, plant growth and development and the regulation of plant responses to abiotic stresses are controlled by the bZIP family of transcription factors, and bZIP transcription factors also can regulate secondary metabolism biosynthetic pathways in plants. In the future, in order to improve plant traits and provide the theoretical basis and germplasm resources for breeding superior varieties, we will pay more attention to controlling the expression of bZIP family transcription factor genes through molecular breeding. Traditional breeding methods to improve plant traits are time-consuming, labour-intensive and uncertain. They cannot meet the needs of modern plant breeding. In addition to being involved in the regulation of plant growth and development, bZIP transcription factors also play an important role in plant response to and tolerance of abiotic stress, and the study of bZIP transcription factors provides an important theoretical basis for breeding varieties that are resistant to stress, thereby maintaining stable crop yields and providing a solid foundation for global food security in the future.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1 Conserved amino acid structural domains of bZIP transcription factors in Arabidopsis thaliana, soybean and other species. The red boxes indicate possible conserved structural domains in the figure. Figure S2 Phylogenetic tree bZIPs family transcription factor proteins in Arabidopsis thaliana, Oryza sativa, Glycine max and so on. Phylogenetic tree of bZIPs transcription factors proteins in selected angiosperms. The bZIPs family gene sequences of plants were retrieved using the NCBI (https://www.ncbi.nlm.nih.gov/) and China National Rice Data Center (https://www.ricedata.cn/). The phylogenetic tree was constructed using MEGA version 11.0 with the bootstrap method based on full amino acid sequences. Numbers next to the branches show the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates).
Author Contributions
All the authors contributed to the present form of the manuscript. Z.M.: Supervision, Writing–original draft, Writing –review & editing. W.J.: Writing – review & editing. Y.Z.: Supervision, Writing –original draft, Writing – review & editing.
Funding Information
This research was supported by open Access funding provided by the Max Planck Society.
Data Availability
Data will be made available on request.
Conflicts of Interest
The authors declare that they have no conflicts of interest associated with this work.
References
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.K. Abiotic stress responses in plants. Nat Rev Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, L.; Jiang, W. Understanding AP2/ERF Transcription Factor Responses and Tolerance to Various Abiotic Stresses in Plants: A Comprehensive Review. Int J Mol Sci. 2024, 25, 893. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Hu, L. WRKY Transcription Factor Responses and Tolerance to Abiotic Stresses in Plants. Int J Mol Sci. 2024, 25, 6845. [Google Scholar] [CrossRef] [PubMed]
- Javed, T.; Gao, S.J. WRKY transcription factors in plant defense. Trends Genet. 2023, 39, 787–801. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, L. MicroRNA: A Dynamic Player from Signalling to Abiotic Tolerance in Plants. Int J Mol Sci. 2023, 24, 11364. [Google Scholar] [CrossRef]
- Ma, Z.; Wu, T.; Huang, K.; Jin, Y.M.; Li, Z.; Chen, M.; Yun, S.; Zhang, H.; Yang, X.; Chen, H.; Bai, H.; Du, L.; Ju, S.; Guo, L.; Bian, M.; Hu, L.; Du, X.; Jiang, W. A Novel AP2/ERF Transcription Factor, OsRPH1, Negatively Regulates Plant Height in Rice. Front Plant Sci. 2020, 11, 709. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, L.; Zhong, Y. Structure, evolution, and roles of MYB transcription factors proteins in secondary metabolite biosynthetic pathways and abiotic stresses responses in plants: a comprehensive review. Front. Plant Sci 2025, 16, 1626844. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Jin, Y.M.; Wu, T.; Hu, L.; Zhang, Y.; Jiang, W.; Du, X. OsDREB2B, an AP2/ERF transcription factor, negatively regulates plant height by conferring GA metabolism in rice. Front Plant Sci. 2022, 13, 1007811. [Google Scholar] [CrossRef]
- Huang, S.; Ma, Z.; Hu, L.; Huang, K.; Zhang, M.; Zhang, S.; Jiang, W.; Wu, T.; Du, X. Involvement of rice transcription factor OsERF19 in response to ABA and salt stress responses. Plant Physiol Biochem. 2021, 167, 22–30. [Google Scholar] [CrossRef]
- Zhong, Y.; Chi, H.; Wu, T.; Fan, W.; Su, H.; Li, R.; Jiang, W.; Du, X.; Ma, Z. Diversity of rhizosphere microbial communities in different rice varieties and their diverse adaptive responses to saline and alkaline stress. Front Microbiol. 2025, 16, 1537846. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Wang, C.; Yang, X.; Wang, L.; Ye, J.; Xu, F.; Liao, Y.; Zhang, W. Role of bZIP transcription factors in the regulation of plant secondary metabolism. Planta. 2023, 258, 13. [Google Scholar]
- Liu, H.; Tang, X.; Zhang, N.; Li, S.; Si, H. Role of bZIP Transcription Factors in Plant Salt Stress. Int J Mol Sci. 2023, 24, 7893. [Google Scholar] [CrossRef]
- Guo, Z.; Dzinyela, R.; Yang, L.; Hwarari, D. bZIP Transcription Factors: Structure, Modification, Abiotic Stress Responses and Application in Plant Improvement. Plants (Basel). 2024, 13, 2058. [Google Scholar]
- Jahan, T.; Huda, M.N.; Zhang, K.; He, Y.; Lai, D.; Dhami, N.; Quinet, M.; Ali, M.A.; Kreft, I.; Woo, S.H.; Georgiev, M.I.; Fernie, A.R.; Zhou, M. Plant secondary metabolites against biotic stresses for sustainable crop protection. Biotechnol Adv. 2025, 79, 108520. [Google Scholar] [CrossRef] [PubMed]
- Aizaz, M.; Lubna; Jan, R.; Asaf, S.; Bilal, S.; Kim, K.M.; Al-Harrasi, A. Regulatory Dynamics of Plant Hormones and Transcription Factors under Salt Stress. Biology (Basel). 2024, 13, 673. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, Y.; Feng, B.; Li, P.; Li, C.; Zhu, C.Y.; Ren, S.N.; Wang, H.L. Regulatory networks of bZIPs in drought, salt and cold stress response and signaling. Plant Sci. 2025, 352, 112399. [Google Scholar] [CrossRef]
- De Souza, C.R.B.; Serrão, C.P.; Barros, N.L.F.; Dos Reis, S.P.; Marques, D.N. Plant bZIP Proteins: Potential use in Agriculture - A Review. Curr Protein Pept Sci. 2024, 25, 107–119. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. Salicylic acid in plant immunity and beyond. Plant Cell. 2024, 36, 1451–1464. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, Y.; Yang, S. Regulatory Networks Underlying Plant Responses and Adaptation to Cold Stress. Annu Rev Genet. 2024, 58, 43–65. [Google Scholar] [CrossRef] [PubMed]
- Supriya, L.; Dake, D.; Woch, N.; Gupta, P.; Gopinath, K.; Padmaja, G.; Muthamilarasan, M. Sugar sensors in plants: Orchestrators of growth, stress tolerance, and hormonal crosstalk. J Plant Physiol. 2025, 307, 154471. [Google Scholar] [CrossRef]
- Qi, J.; Mao, Y.; Cui, J.; Lu, X.; Xu, J.; Liu, Y.; Zhong, H.; Yu, W.; Li, C. The role of strigolactones in resistance to environmental stress in plants. Physiol Plant. 2024, 176, e14419. [Google Scholar] [CrossRef]
- Xu, L.; Zhu, Y.; Wang, Y.; Zhang, L.; Li, L.; Looi, L.J.; Zhang, Z. The potential of melatonin and its crosstalk with other hormones in the fight against stress. Front Plant Sci. 2024, 15, 1492036. [Google Scholar] [CrossRef]
- Wang, X.; Wen, H.; Suprun, A.; Zhu, H. Ethylene Signaling in Regulating Plant Growth, Development, and Stress Responses. Plants (Basel). 2025, 14, 309. [Google Scholar] [CrossRef]
- Dröge-Laser, W.; Snoek, B.L.; Snel, B.; Weiste, C. The Arabidopsis bZIP transcription factor family-an update. Curr Opin Plant Biol. 2018, 45, 36–49. [Google Scholar] [CrossRef]
- Pacifici, E.; Polverari, L.; Sabatini, S. Plant hormone cross-talk: the pivot of root growth. J Exp Bot. 2015, 66, 1113–1121. [Google Scholar] [CrossRef]
- Lutova, L.A.; Dodueva, I.E.; Lebedeva, M.A.; Tvorogova, V.E. Transcription Factors in Developmental Genetics and the Evolution of Higher Plants. Genetika. 2015, 51, 539–557. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Jin, L.; Peng, R. Crosstalk between Ca2+ and Other Regulators Assists Plants in Responding to Abiotic Stress. Plants (Basel). 2022, 11, 1351. [Google Scholar] [CrossRef] [PubMed]
- Janiak, A.; Kwaśniewski, M.; Szarejko, I. Gene expression regulation in roots under drought. J Exp Bot. 2016, 67, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Wu, M.; Hu, W.; Liu, R.; Yan, H.; Xiang, Y. Genome-Wide Identification and Expression Analyses of the bZIP Transcription Factor Genes in moso bamboo (Phyllostachys edulis). Int J Mol Sci. 2019, 20, 2203. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.J.; Zhu, Y.X. Transcription factor families in Arabidopsis: major progress and outstanding issues for future research. Curr Opin Plant Biol. 2006, 9, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Gangappa, S.N.; Botto, J.F. The Multifaceted Roles of HY5 in Plant Growth and Development. Mol Plant. 2016, 9, 1353–1365. [Google Scholar] [CrossRef]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, R.; Guo, C.; Hou, H.; Wang, X.; Gao, H. Evolutionary and Expression Analyses of the Apple Basic Leucine Zipper Transcription Factor Family. Front Plant Sci. 2016, 7, 376. [Google Scholar] [CrossRef]
- Aerts, N.; Hickman, R.; Van Dijken, A.J.H.; Kaufmann, M.; Snoek, B.L.; Pieterse, C.M.J.; Van Wees, S.C.M. Architecture and dynamics of the abscisic acid gene regulatory network. Plant J. 2024, 119, 2538–2563. [Google Scholar] [CrossRef] [PubMed]
- Martignago, D.; Da, S.F.V.; Lombardi, A.; Gao, H.; Korwin, K.P.; Galbiati, M.; Tonelli, C.; Coupland, G.; Conti, L. The bZIP transcription factor AREB3 mediates FT signalling and floral transition at the Arabidopsis shoot apical meristem. PLoS Genet. 2023, 19, e1010766. [Google Scholar]
- Meng, X.B.; Zhao, W.S.; Lin, R.M.; Wang, M.; Peng, Y.L. Identification of a novel rice bZIP-type transcription factor gene, OsbZIP1, involved in response to infection of Magnaporthe grisea. Plant Molecular Biology Reporter. 2005, 23, 301–302. [Google Scholar] [CrossRef]
- Vesely, P.W.; Staber, P.B.; Hoefler, G.; Kenner, L. Translational regulation mechanisms of AP-1 proteins. Mutat Res. 2009, 682, 7–12. [Google Scholar] [CrossRef]
- Golldack, D.; Lüking, I.; Yang, O. Plant tolerance to drought and salinity: stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Gai, Y.; Li, L.; Liu, B.; Ma, H.; Chen, Y.; Zheng, F.; Sun, X.; Wang, M.; Jiao, C.; Li, H. Distinct and essential roles of bZIP transcription factors in the stress response and pathogenesis in Alternaria alternata. Microbiol Res. 2022, 256, 126915. [Google Scholar] [CrossRef] [PubMed]
- Weltmeier, F.; Rahmani, F.; Ehlert, A.; Dietrich, K.; Schütze, K.; Wang, X.; Chaban, C.; Hanson, J.; Teige, M.; Harter, K.; Vicente-Carbajosa, J.; Smeekens, S.; Dröge-Laser, W. Expression patterns within the Arabidopsis C/S1 bZIP transcription factor network: availability of heterodimerization partners controls gene expression during stress response and development. Plant Mol Biol. 2009, 69, 107–119. [Google Scholar] [CrossRef]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP Research Group. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Alves, M.S.; Dadalto, S.P.; Gonçalves, A.B.; De Souza, G.B.; Barros, V.A.; Fietto, L.G. Plant bZIP transcription factors responsive to pathogens: a review. Int J Mol Sci. 2013, 14, 7815–7828. [Google Scholar] [CrossRef]
- Kang, S.G.; Price, J.; Lin, P.C.; Hong, J.C.; Jang, J.C. The arabidopsis bZIP1 transcription factor is involved in sugar signaling, protein networking, and DNA binding. Mol Plant. 2010, 3, 361–373. [Google Scholar] [CrossRef]
- Li, H.; Li, L.; Shang, G.G.; Jia, C.; Deng, S.; Noman, M.; Liu, Y.; Guo, Y.; Han, L.; Zhang, X.; Dong, Y.; Ahmad, N.; Du, L.; Li, H.; Yang, J. Genome-wide identification and expression analysis of bZIP gene family in Carthamus tinctorius L. Sci Rep. 2020, 10, 15521. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr Opin Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Yang, H.; Yan, Y.; Wei, Y.; Tie, W.; Ding, Z.; Zuo, J.; Peng, M.; Li, K. Genome-wide characterization and analysis of bZIP transcription factor gene family related to abiotic stress in cassava. Sci Rep. 2016, 6, 22783. [Google Scholar] [CrossRef]
- Keller, W.; König, P.; Richmond, T.J. Crystal structure of a bZIP/DNA complex at 2.2 A: determinants of DNA specific recognition. J Mol Biol. 1995, 254, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Jung, H.J.; Park, J.I.; Yang, T.J.; Nou, I.S. Transcriptome analysis of newly classified bZIP transcription factors of Brassica rapa in cold stress response. Genomics. 2014, 104, 194–202. [Google Scholar] [CrossRef]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice. Plant Physiol. 2008, 146, 333–350. [Google Scholar] [CrossRef]
- Li, X.; Gao, S.; Tang, Y.; Li, L.; Zhang, F.; Feng, B.; Fang, Z.; Ma, L.; Zhao, C. Genome-wide identification and evolutionary analyses of bZIP transcription factors in wheat and its relatives and expression profiles of anther development related TabZIP genes. BMC Genomics. 2015, 16, 976. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Chen, X.; Yang, T.B.; Cheng, Q.; Cheng, Z.M. Genome-Wide Identification of bZIP Family Genes Involved in Drought and Heat Stresses in Strawberry (Fragaria vesca). Int J Genomics. 2017, 2017, 3981031. [Google Scholar] [CrossRef]
- Li, D.; Fu, F.; Zhang, H.; Song, F. Genome-wide systematic characterization of the bZIP transcriptional factor family in tomato (Solanum lycopersicum L.). BMC Genomics. 2015, 16, 771. [Google Scholar] [CrossRef]
- Liu, J.; Chen, N.; Chen, F.; Cai, B.; Dal, S.S.; Tornielli, G.B.; Pezzotti, M.; Cheng, Z.M. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine (Vitis vinifera). BMC Genomics. 2014, 15, 281. [Google Scholar] [CrossRef] [PubMed]
- Izawa, T.; Foster, R.; Chua, N.H. Plant bZIP protein DNA binding specificity. J Mol Biol. 1993, 230, 1131–1144. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Narusaka, Y.; Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Furihata, T.; Abe, H.; Narusaka, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Interaction between two cis-acting elements, ABRE and DRE, in ABA-dependent expression of Arabidopsis rd29A gene in response to dehydration and high-salinity stresses. Plant J. 2003, 34, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhou, H.; Xu, Y.; Huang, D.; Wu, B.; Xing, W.; Chen, D.; Xuv, B.; Song, S. Comprehensive analysis of bZIP transcription factors in passion fruit. iScience. 2023, 26, 106556. [Google Scholar] [CrossRef]
- Kim, S.; Kang, J.Y.; Cho, D.I.; Park, J.H.; Kim, S.Y. ABF2, an ABRE-binding bZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef]
- Liao, Y.; Zou, H.F.; Wei, W.; Hao, Y.J.; Tian, A.G.; Huang, J.; Liu, Y.F.; Zhang, J.S.; Chen, S.Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta. 2008, 228, 225–240. [Google Scholar] [CrossRef]
- Schütze, K.; Harter, K.; Chaban, C. Post-translational regulation of plant bZIP factors. Trends Plant Sci. 2008, 13, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Muñiz, G.M.N.; Giammaria, V.; Grandellis, C.; Téllez, I.M.T.; Ulloa, R.M.; Capiati, D.A. Characterization of StABF1, a stress-responsive bZIP transcription factor from Solanum tuberosum L. that is phosphorylated by StCDPK2 in vitro. Planta. 2012, 235, 761–778. [Google Scholar] [CrossRef]
- Meng, D.; Cao, H.; Yang, Q.; Zhang, M.; Borejsza, W.E.; Wang, H.; Dandekar, A.M.; Fei, Z.; Cheng, L. SnRK1 kinase-mediated phosphorylation of transcription factor bZIP39 regulates sorbitol metabolism in apple. Plant Physiol. 2023, 192, 2123–2142. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. Apple bZIP transcription factor MdbZIP44 regulates abscisic acid-promoted anthocyanin accumulation. Plant Cell Environ. 2018, 41, 2678–2692. [Google Scholar] [CrossRef]
- Zong, W.; Tang, N.; Yang, J.; Peng, L.; Ma, S.; Xu, Y.; Li, G.; Xiong, L. Feedback Regulation of ABA Signaling and Biosynthesis by a bZIP Transcription Factor Targets Drought-Resistance-Related Genes. Plant Physiol. 2016, 171, 2810–2825. [Google Scholar] [CrossRef]
- Zhang, L.; Xie, J.; Wang, L.; Si, L.; Zheng, S.; Yang, Y.; Yang, H.; Tian, S. Wheat TabZIP8, 9, 13 participate in ABA biosynthesis in NaCl-stressed roots regulated by TaCDPK9-1. Plant Physiol Biochem. 2020, 151, 650–658. [Google Scholar] [CrossRef]
- Meng, X.B.; Zhao, W.S.; Lin, R.M.; Wang, M.; Peng, Y.L. Identification of a novel rice bZIP-type transcription factor gene, OsbZIP1, involved in response to infection of Magnaporthe grisea. Plant Molecular Biology Reporter. 2025, 23, 301–302. [Google Scholar] [CrossRef]
- Berendzen, K.W.; Weiste, C.; Wanke, D.; Kilian, J.; Harter, K.; Dröge-Laser, W. Bioinformatic cis-element analyses performed in Arabidopsis and rice disclose bZIP- and MYB-related binding sites as potential AuxRE-coupling elements in auxin-mediated transcription. BMC Plant Biol. 2012, 12, 125. [Google Scholar] [CrossRef]
- Zhang, J.; Qian, J.Y.; Bian, Y.H.; Liu, X.; Wang, C.L. Transcriptome and Metabolite Conjoint Analysis Reveals the Seed Dormancy Release Process in Callery Pear. Int J Mol Sci. 2022, 23, 2186. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, G.; Xia, N.; Wang, X.J.; Huang, L.L.; Kang, Z.S. Cloning and characterization of a bZIP transcription factor gene in wheat and its expression in response to stripe rust pathogen infection and abiotic stresses. Physiological and Molecular Plant Pathology. 2008, 73, 88–94. [Google Scholar] [CrossRef]
- Han, Y.; Hou, Z.; He, Q.; Zhang, X.; Yan, K.; Han, R.; Liang, Z. Genome-Wide Characterization and Expression Analysis of bZIP Gene Family Under Abiotic Stress in Glycyrrhiza uralensis. Front Genet. 2021, 12, 754237. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Y.; Zhang, S.; Xie, K.; Zhang, C.; Xi, Y.; Sun, F. Genome-wide analysis of the abiotic stress-related bZIP family in switchgrass. Mol Biol Rep. 2020, 47, 4439–4454. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, Y.; Qiu, J.; Wang, H.; Wang, S.; Tang, L.; Tong, X.; Zhang, J. Abscisic acid promotes jasmonic acid biosynthesis via a 'SAPK10-bZIP72-AOC' pathway to synergistically inhibit seed germination in rice (Oryza sativa). New Phytol. 2020, 228, 1336–1353. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, J.; Li, X.; Wang, S.; Wu, J. Salt and drought stress and ABA responses related to bZIP genes from V. radiata and V. angularis. Gene. 2018, 651, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, L.; Xia, C.; Gao, L.; Hao, C.; Zhao, G.; Jia, J.; Kong, X. A Novel Wheat C-bZIP Gene, TabZIP14-B, Participates in Salt and Freezing Tolerance in Transgenic Plants. Front Plant Sci. 2017, 8, 710. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Zhao, G.; Zheng, S.; Xie, S.; Lu, C.; Liu, S.; Wang, Z.; Niu, J. Comprehensive Functional Analysis of the bZIP Family in Bletilla striata Reveals That BsbZIP13 Could Respond to Multiple Abiotic Stresses. Int J Mol Sci. 2023, 24, 15202. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Qiao, Y.; Pan, X.; Chen, X.; Su, W.; Li, A.; Li, X.; Liao, W. Genome-Wide identification and expression analysis of CsABF/AREB gene family in cucumber (Cucumis sativus L.) and in response to phytohormonal and abiotic stresses. Sci Rep. 2025, 15, 15757. [Google Scholar] [CrossRef]
- Chai, M.; Yang, F.; Cai, S.; Liu, T.; Xu, X.; Huang, Y.; Xi, X.; Yang, J.; Cao, Z.; Sun, L.; Dou, D.; Fang, X.; Yan, M.; Cai, H. Overexpression of the Transcription Factor GmbZIP60 Increases Salt and Drought Tolerance in Soybean (Glycine max). Int J Mol Sci. 2025, 26, 3455. [Google Scholar] [CrossRef]
- He, Q.; Cai, H.; Bai, M.; Zhang, M.; Chen, F.; Huang, Y.; Priyadarshani, S.V.G.N.; Chai, M.; Liu, L.; Liu, Y.; Chen, H.; Qin, Y. A Soybean bZIP Transcription Factor GmbZIP19 Confers Multiple Biotic and Abiotic Stress Responses in Plant. Int J Mol Sci. 2020, 21, 4701. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat Rev Mol Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.Y.; Qin, F.; Li, J.; Ding, Y.; Shi, Y.; Wang, Y.; Yang, Y.; Guo, Y.; Zhu, J.K. Plant abiotic stress response and nutrient use efficiency. Sci China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS Homeostasis in Abiotic Stress Tolerance in Plants. Int J Mol Sci. 2020, 21, 5208. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhang, M.; Zhang, H.; Huang, K.; Chen, M.; Chen, C.; Yang, X.; Li, Z.; Chen, H.; Ma, Z.; Zhang, X.; Jiang, W.; Du, X. Identification and characterization of EDT1 conferring drought tolerance in rice. Journal of plant biology. 2019, 62, 39–47. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz, U.B.; García, C.P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef]
- Pan, Y.; Hu, X.; Li, C.; Xu, X.; Su, C.; Li, J.; Song, H.; Zhang, X.; Pan, Y. SlbZIP38, a Tomato bZIP Family Gene Downregulated by Abscisic Acid, Is a Negative Regulator of Drought and Salt Stress Tolerance. Genes (Basel). 2017, 8, 402. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Q.Y.; Xu, P.; Wang, X.H.; Dai, S.J.; Liu, Z.N.; Xu, M.; Cao, X.; Cui, X.Y. GmTRAB1, a Basic Leucine Zipper Transcription Factor, Positively Regulates Drought Tolerance in Soybean (Glycine max. L). Plants (Basel). 2024, 13, 3104. [Google Scholar] [CrossRef]
- Wang, X.; Meng, Y.; Zhang, S.; Wang, Z.; Zhang, K.; Gao, T.; Ma, Y. Characterization of bZIP Transcription Factors in Transcriptome of Chrysanthemum mongolicum and Roles of CmbZIP9 in Drought Stress Resistance. Plants (Basel). 2024, 13, 2064. [Google Scholar] [CrossRef]
- Zhou, H.; Shi, H.; Yang, Y.; Feng, X.; Chen, X.; Xiao, F.; Lin, H.; Guo, Y. Insights into plant salt stress signaling and tolerance. J Genet Genomics. 2024, 51, 16–34. [Google Scholar] [CrossRef]
- He, L.; Wu, Z.; Wang, X.; Zhao, C.; Cheng, D.; Du, C.; Wang, H.; Gao, Y.; Zhang, R.; Han, J.; Xu, J. A novel maize F-bZIP member, ZmbZIP76, functions as a positive regulator in ABA-mediated abiotic stress tolerance by binding to ACGT-containing elements. Plant Sci. 2024, 341, 111952. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Jiao, C.; Li, X.Y.; Li, X.; Zhang, X.Y.; Wang, Y.; Song, Y.; Yang, K.; Liu, Y.; Zhang, J.; Liu, M.; Liang, Y.L.; Wang, X.; Li, H.; Jia, P.; Zhang, X.; Qi, G.; Dong, Q. Identification and functional characterization of JrbZIP40 in walnut reveals its role in salt and drought stress tolerance in transgenic Arabidopsis seedlings. BMC Plant Biol. 2025, 25, 807. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guo, C.; Li, Z.; Sun, J.; Wang, D.; Xu, L.; Li, X.; Guo, Y. Identification and Analysis of bZIP Family Genes in Potato and Their Potential Roles in Stress Responses. Front Plant Sci. 2021, 12, 637343. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol Biol. 2014, 84, 19–36. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, S. Surviving and thriving: How plants perceive and respond to temperature stress. Dev Cell. 2022, 57, 947–958. [Google Scholar] [CrossRef]
- Suzuki, N. Temperature Stress and Responses in Plants. Int J Mol Sci. 2019, 20, 2001. [Google Scholar] [CrossRef]
- Kan, Y.; Mu, X.R.; Gao, J.; Lin, H.X.; Lin, Y. The molecular basis of heat stress responses in plants. Mol Plant. 2023, 16, 1612–1634. [Google Scholar] [CrossRef]
- Perrella, G.; Bäurle, I.; van, Z.M. Epigenetic regulation of thermomorphogenesis and heat stress tolerance. New Phytol. 2022, 234, 1144–1160. [Google Scholar] [CrossRef]
- Shen, L.; Liu, Z.; Yang, S.; Yang, T.; Liang, J.; Wen, J.; Liu, Y.; Li, J.; Shi, L.; Tang, Q.; Shi, W.; Hu, J.; Liu, C.; Zhang, Y.; Lin, W.; Wang, R.; Yu, H.; Mou, S.; Hussain, A.; Cheng, W.; Cai, H.; He, L.; Guan, D.; Wu, Y.; He, S. Pepper CabZIP63 acts as a positive regulator during Ralstonia solanacearum or high temperature-high humidity challenge in a positive feedback loop with CaWRKY40. J Exp Bot. 2016, 67, 2439–2451. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Dai, X.J.; Gu, Z.Y. OsbZIP33 is an ABA-dependent enhancer of drought tolerance in rice. Crop Science. 2015, 55, 1673–1685. [Google Scholar] [CrossRef]
- Qiu, F.; Zheng, Y.; Lin, Y.; Woldegiorgis, S.T.; Xu, S.; Feng, C.; Huang, G.; Shen, H.; Xu, Y.; Kabore, M.A.F.; Ai, Y.; Liu, W.; He, H. Integrated ATAC-Seq and RNA-Seq Data Analysis to Reveal OsbZIP14 Function in Rice in Response to Heat Stress. Int J Mol Sci. 2023, 24, 5619. [Google Scholar] [CrossRef]
- Jiang, M.; Li, P. Unraveling the evolution of the ATB2 subgroup basic leucine zipper transcription factors in plants and decoding the positive effects of BdibZIP44 and BdibZIP53 on heat stress in Brachypodium distachyon. Plant Physiol Biochem. 2025, 222, 109708. [Google Scholar] [CrossRef] [PubMed]
- Sadura, I.; Janeczko, A. Brassinosteroids and the Tolerance of Cereals to Low and High Temperature Stress: Photosynthesis and the Physicochemical Properties of Cell Membranes. Int J Mol Sci. 2021, 23, 342. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wu, A.; He, Y.; Li, F.; Wei, C. Genome-wide characterization of the basic leucine zipper transcription factors in Camellia sinensis. Tree Genetics & Genomes. 2018, 14, 27. [Google Scholar]
- Yao, L.; Hao, X.; Cao, H.; Ding, C.; Yang, Y.; Wang, L.; Wang, X. ABA-dependent bZIP transcription factor, CsbZIP18, from Camellia sinensis negatively regulates freezing tolerance in Arabidopsis. Plant Cell Rep. 2020, 39, 553–565. [Google Scholar] [CrossRef]
- Liang, Y.; Xia, J.; Jiang, Y.; Bao, Y.; Chen, H.; Wang, D.; Zhang, D.; Yu, J.; Cang, J. Genome-Wide Identification and Analysis of bZIP Gene Family and Resistance of TaABI5 (TabZIP96) under Freezing Stress in Wheat (Triticum aestivum). Int J Mol Sci. 2022, 23, 2351. [Google Scholar] [CrossRef]
- Liu, C.; Wu, Y.; Wang, X. bZIP transcription factor OsbZIP52/RISBZ5: a potential negative regulator of cold and drought stress response in rice. Planta. 2012, 235, 1157–1169. [Google Scholar] [CrossRef]
- Shu, Y.; Zhang, W.; Tang, L.; Li, Z.; Liu, X.; Liu, X.; Liu, W.; Li, G.; Ying, J.; Huang, J.; Tong, X.; Hu, H.; Zhang, J.; Wang, Y. ABF1 Positively Regulates Rice Chilling Tolerance via Inducing Trehalose Biosynthesis. Int J Mol Sci. 2023, 24, 11082. [Google Scholar] [CrossRef]
- Usmani, L.; Shakil, A.; Khan, I.; Alvi, T.; Singh, S.; Das, D. Brassinosteroids in Micronutrient Homeostasis: Mechanisms and Implications for Plant Nutrition and Stress Resilience. Plants (Basel). 2025, 14, 598. [Google Scholar] [CrossRef]
- Hossain, M.S.; Garcia, C.P.; Mühling, K.H. Editorial: Mineral nutrition and plant stress tolerance. Front Plant Sci. 2024, 15, 1461651. [Google Scholar] [CrossRef]
- Assunção, A.G.L. The F-bZIP-regulated Zn deficiency response in land plants. Planta. 2022, 256, 108. [Google Scholar] [CrossRef]
- Thiébaut, N.; Hanikenne, M. Zinc deficiency responses: bridging the gap between Arabidopsis and dicotyledonous crops. J Exp Bot. 2022, 73, 1699–1716. [Google Scholar] [CrossRef] [PubMed]
- Assunção, A.G.; Herrero, E.; Lin, Y.F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.; van, E.M.; Fiers, M.; Schat, H.; Aarts, M.G. Arabidopsis thaliana transcription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc Natl Acad Sci U S A. 2010, 107, 10296–10301. [Google Scholar] [CrossRef]
- Lilay, G.H.; Castro, P.H.; Campilho, A.; Assunção, A.G.L. The Arabidopsis bZIP19 and bZIP23 Activity Requires Zinc Deficiency - Insight on Regulation From Complementation Lines. Front Plant Sci. 2019, 9, 1955. [Google Scholar] [CrossRef]
- Liang, G. Iron uptake, signaling, and sensing in plants. Plant Commun. 2022, 3, 100349. [Google Scholar] [CrossRef]
- Mankotia, S.; Singh, D.; Monika, K.; Kalra, M.; Meena, H.; Meena, V.; Yadav, R.K.; Pandey, A.K.; Satbhai, S.B. ELONGATED HYPOCOTYL 5 regulates BRUTUS and affects iron acquisition and homeostasis in Arabidopsis thaliana. Plant J. 2023, 114, 1267–1284. [Google Scholar] [CrossRef] [PubMed]
- Morkunas, I.; Woźniak, A.; Mai, V.C.; Rucińska, S.R.; Jeandet, P. The Role of Heavy Metals in Plant Response to Biotic Stress. Molecules. 2018, 23, 2320. [Google Scholar] [CrossRef] [PubMed]
- Goncharuk, E.A.; Zagoskina, N.V. Heavy Metals, Their Phytotoxicity, and the Role of Phenolic Antioxidants in Plant Stress Responses with Focus on Cadmium: Review. Molecules. 2023, 28, 3921. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhou, J.; Jie, Y.; Xing, H.; Zhong, Y.; Yu, W.; She, W.; Ma, Y.; Liu, Z.; Zhang, Y. A Ramie bZIP Transcription Factor BnbZIP2 Is Involved in Drought, Salt, and Heavy Metal Stress Response. DNA Cell Biol. 2016, 35, 776–786. [Google Scholar] [CrossRef]
- Lu, Z.; Qiu, W.; Jin, K.; Yu, M.; Han, X.; He, X.; Wu, L.; Wu, C.; Zhuo, R. Identification and Analysis of bZIP Family Genes in Sedum plumbizincicola and Their Potential Roles in Response to Cadmium Stress. Front Plant Sci. 2022, 13, 859386. [Google Scholar] [CrossRef]
- Chai, M.; Fan, R.; Huang, Y.; Jiang, X.; Wai, M.H.; Yang, Q.; Su, H.; Liu, K.; Ma, S.; Chen, Z.; Wang, F.; Qin, Y.; Cai, H. GmbZIP152, a Soybean bZIP Transcription Factor, Confers Multiple Biotic and Abiotic Stress Responses in Plant. Int J Mol Sci. 2022, 23, 10935. [Google Scholar] [CrossRef]
- Hou, F.; Liu, K.; Zhang, N.; Zou, C.; Yuan, G.; Gao, S.; Zhang, M.; Pan, G.; Ma, L.; Shen, Y. Association mapping uncovers maize ZmbZIP107 regulating root system architecture and lead absorption under lead stress. Front Plant Sci. 2022, 13, 1015151. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.; Fang, Y.; Gao, Y.; Yang, S.; Su, J.; Ni, J.; Teng, Y.; Bai, S. Phosphorylated transcription factor PuHB40 mediates ROS-dependent anthocyanin biosynthesis in pear exposed to high light. Plant Cell. 2024, 36, 3562–3583. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Ke, X.; Yang, X.; Liu, Y.; Hou, X. Plants response to light stress. J Genet Genomics. 2022, 49, 735–747. [Google Scholar] [CrossRef]
- Liu, Q.; Fang, H.; Zhou, H.; Wang, X.; Hou, Z. Identification and Characterization of bZIP Gene Family Combined Transcriptome Analysis Revealed Their Functional Roles on Abiotic Stress and Anthocyanin Biosynthesis in Mulberry (Morus alba). Horticulturae. 2025, 11, 694. [Google Scholar] [CrossRef]
- Stracke, R.; Favory, J.J.; Gruber, H.; Bartelniewoehner, L.; Bartels, S.; Binkert, M.; Funk, M.; Weisshaar, B.; Ulm, R. The Arabidopsis bZIP transcription factor HY5 regulates expression of the PFG1/MYB12 gene in response to light and ultraviolet-B radiation. Plant Cell Environ. 2010, 33, 88–103. [Google Scholar]
- Shen, H.; Cao, K.; Wang, X. AtbZIP16 and AtbZIP68, two new members of GBFs, can interact with other G group bZIPs in Arabidopsis thaliana. BMB Rep. 2008, 41, 132–8. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules. 2018, 23, 762. [Google Scholar] [CrossRef]
- Bonta, R.K. Dietary Phenolic Acids and Flavonoids as Potential Anti-Cancer Agents: Current State of the Art and Future Perspectives. Anticancer Agents Med Chem. 2020, 20, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Cai, W.; Zhang, H.H.; Zhong, Y.S.; Fang, J.; Zhang, W.Y.; Mo, L.; Wang, L.C.; Yu, C.H. Selaginella uncinata flavonoids ameliorated ovalbumin-induced airway inflammation in a rat model of asthma. J Ethnopharmacol. 2017, 195, 71–80. [Google Scholar] [CrossRef]
- Jiao, W.H.; Cheng, B.H.; Shi, G.H.; Chen, G.D.; Gu, B.B.; Zhou, Y.J.; Hong, L.L.; Yang, F.; Liu, Z.Q.; Qiu, S.Q.; Liu, Z.G.; Yang, P.C.; Lin, H.W. Dysivillosins A-D, Unusual Anti-allergic Meroterpenoids from the Marine Sponge Dysidea villosa. Sci Rep. 2017, 7, 8947. [Google Scholar] [CrossRef]
- Wangchuk, P.; Sastraruji, T.; Taweechotipatr, M.; Keller, P.A.; Pyne, S.G. Anti-inflammatory, Anti-bacterial and Anti-acetylcholinesterase Activities of two Isoquinoline Alkaloids-Scoulerine and Cheilanthifoline. Nat Prod Commun. 2016, 11, 1801–1804. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Jadaun, J.; Chandra, M.; Sangwan, N. Medicinal plant transcriptomes: The new gateways for accelerated understanding of plant secondary metabolism. Plant Genet Resour. 2016, 14, 256–269. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, S.; Li, R.; Zhao, Q. Unraveling the specialized metabolic pathways in medicinal plant genomes: a review. Front Plant Sci. 2024, 15, 1459533. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Rashmi, R.; Toppo, V.; Chole, P.B.; Banadka, A.; Sudheer, W.N.; Nagella, P.; Shehata, W.F.; Al-Mssallem, M.Q.; Alessa, F.M.; Almaghasla, M.I.; Rezk, A.A. Plant Secondary Metabolites: The Weapons for Biotic Stress Management. Metabolites. 2023, 13, 716. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Borek, C. Antioxidant health effects of aged garlic extract. J Nutr. 2001, 131, 1010S–5S. [Google Scholar] [CrossRef]
- Han, H.; Dong, L.; Zhang, W.; Liao, Y.; Wang, L.; Wang, Q.; Ye, J.; Xu, F. Ginkgo biloba GbbZIP08 transcription factor is involved in the regulation of flavonoid biosynthesis. J Plant Physiol. 2023, 287, 154054. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Lu, C.; Zhou, F.; Zhu, Y.; Jiang, W.; Zhou, A.; Shen, Y.; Pan, L.; Lv, A.; Shao, Q. Transcription factor DcbZIPs regulate secondary metabolism in Dendrobium catenatum during cold stress. Physiol Plant. 2024, 176, e14501. [Google Scholar] [CrossRef]
- Han, H.; Xu, F.; Li, Y.; Yu, L.; Fu, M.; Liao, Y.; Yang, X.; Zhang, W.; Ye, J. Genome-wide characterization of bZIP gene family identifies potential members involved in flavonoids biosynthesis in Ginkgo biloba L. Sci Rep. Sci Rep. 2021, 11, 23420. [Google Scholar] [CrossRef]
- Zhang, F.; Huang, J.; Guo, H.; Yang, C.; Li, Y.; Shen, S.; Zhan, C.; Qu, L.; Liu, X.; Wang, S.; Chen, W.; Luo, J. OsRLCK160 contributes to flavonoid accumulation and UV-B tolerance by regulating OsbZIP48 in rice. Sci China Life Sci. 2022, 65, 1380–1394. [Google Scholar] [CrossRef]
- Tholl, D. Biosynthesis and biological functions of terpenoids in plants. Adv Biochem Eng Biotechnol. 2015, 148, 63–106. [Google Scholar]
- Zheng, X.; Li, P.; Lu, X. Research advances in cytochrome P450-catalysed pharmaceutical terpenoid biosynthesis in plants. J Exp Bot. 2019, 70, 4619–4630. [Google Scholar]
- Michael, R.; Ranjan, A.; Kumar, R.S.; Pathak, P.K.; Trivedi, P.K. Light-regulated expression of terpene synthase gene, AtTPS03, is controlled by the bZIP transcription factor, HY5, in Arabidopsis thaliana. Biochem Biophys Res Commun. 2020, 529, 437–443. [Google Scholar] [PubMed]
- Xu, Z.; Zeng, T.; Li, J.; Zhou, L.; Li, J.; Luo, J.; Zheng, R.; Wang, Y.; Hu, H.; Wang, C. TcbZIP60 positively regulates pyrethrins biosynthesis in Tanacetum cinerariifolium. Front Plant Sci. 2023, 14, 1133912. [Google Scholar]
- Wu, M.; Chen, J.; Tang, W.; Jiang, Y.; Hu, Z.; Xu, D.; Hou, K.; Chen, Y.; Wu, W. Genome-Wide Identification and Expression Analysis of bZIP Family Genes in Stevia rebaudiana. Genes (Basel). 2023, 14, 1918. [Google Scholar] [PubMed]
- Zhang, F.; Fu, X.; Lv, Z.; Lu, X.; Shen, Q.; Zhang, L.; Zhu, M.; Wang, G.; Sun, X.; Liao, Z.; Tang, K. A basic leucine zipper transcription factor, AabZIP1, connects abscisic acid signaling with artemisinin biosynthesis in Artemisia annua. Mol Plant. 2015, 8, 163–175. [Google Scholar]
- Banerjee, A.; Roychoudhury, A. Abscisic-acid-dependent basic leucine zipper (bZIP) transcription factors in plant abiotic stress. Protoplasma. 2017, 254, 3–16. [Google Scholar]
- Facchini, P.J. Regulation of alkaloid biosynthesis in plants. Alkaloids Chem Biol. 2006, 63, 1–44. [Google Scholar]
- Bhambhani, S.; Kondhare, K.R.; Giri, A.P. Diversity in Chemical Structures and Biological Properties of Plant Alkaloids. Molecules. 2021, 26, 3374. [Google Scholar]
- Jiang, M.; Wang, Z.; Ren, W.; Yan, S.; Xing, N.; Zhang, Z.; Li, H.; Ma, W. Identification of the bZIP gene family and regulation of metabolites under salt stress in isatis indigotica. Front Plant Sci. 2022, 13, 1011616. [Google Scholar]
- Sui, X.; Singh, S.K.; Patra, B.; Schluttenhofer, C.; Guo, W.; Pattanaik, S.; Yuan, L. Cross-family transcription factor interaction between MYC2 and GBFs modulates terpenoid indole alkaloid biosynthesis. J Exp Bot. 2018, 69, 4267–4281. [Google Scholar] [PubMed]
- Stuper-Szablewska, K.; Perkowski, J. Phenolic acids in cereal grain: Occurrence, biosynthesis, metabolism and role in living organisms. Crit Rev Food Sci Nutr. 2019, 59, 664–675. [Google Scholar]
- Vinogradova, N.; Vinogradova, E.; Chaplygin, V.; Mandzhieva, S.; Kumar, P.; Rajput, V.D.; Minkina, T.; Seth, C.S.; Burachevskaya, M.; Lysenko, D.; Singh, R.K. Phenolic Compounds of the Medicinal Plants in an Anthropogenically Transformed Environment. Molecules. 2023, 28, 6322. [Google Scholar]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Res Int. 2020, 132, 109114. [Google Scholar] [CrossRef]
- Shi, M.; Hua, Q.; Kai, G.Y. Comprehensive transcriptomic analysis in response to abscisic acid in Salvia miltiorrhiza. Plant Cell Tissue Organ Cult. 2021, 147, 389–404. [Google Scholar] [CrossRef]
- Ibáñez, J.; Battistoni, B.; Fiol, A.; Dare, A.P.; Ballesta, P.; Ahumada, S.; Meisel, L.A.; Allan, A.; Espley, R.; Pacheco, I. Systematic characterization of the bZIP transcription factor family of Japanese plum (Prunus salicina Lindl.) and their potential role in phenolic compound biosynthesis. Scientia Horticulturae. 2025, 341, 113962. [Google Scholar]
- Moura, J.C.; Bonine, C.A.; de Oliveira, F.V.J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J Integr Plant Biol. 2010, 52, 360–376. [Google Scholar]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu Rev Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. Int J Mol Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Wang, X.; Yin, W.; Wang, Y.; Li, Y.; Zhang, G.; Li, Z.; Song, J.; Wang, X. Grapevine VlbZIP30 improves drought resistance by directly activating VvNAC17 and promoting lignin biosynthesis through the regulation of three peroxidase genes. Hortic Res. 2020, 7, 150. [Google Scholar] [CrossRef]
- Li, X.; Huang, H.; Rizwan, H.M.; Wang, N.; Jiang, J.; She, W.; Zheng, G.; Pan, H.; Guo, Z.; Pan, D.; Pan, T. Transcriptome Analysis Reveals Candidate Lignin-Related Genes and Transcription Factors during Fruit Development in Pomelo (Citrus maxima). Genes (Basel). 2022, 13, 845. [Google Scholar] [PubMed]
- Nemeth, K.; Bayraktar, R.; Ferracin, M.; Calin, G.A. Non-coding RNAs in disease: from mechanisms to therapeutics. Nat Rev Genet. 2024, 25, 211–232. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; Gingeras, T.R.; Guttman, M.; Hirose, T.; Huarte, M.; Johnson, R.; Kanduri, C.; Kapranov, P.; Lawrence, J.B.; Lee, J.T.; Mendell, J.T.; Mercer, T.R.; Moore, K.J.; Nakagawa, S.; Rinn, J.L.; Spector, D.L.; Ulitsky, I.; Wan, Y.; Wilusz, J.E.; Wu, M. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat Rev Mol Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, A.T.; Blevins, T.; Swiezewski, S. Long Noncoding RNAs in Plants. Annu Rev Plant Biol. 2021, 72, 245–271. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Sunwoo, H.; Spector, D.L. Long noncoding RNAs: functional surprises from the RNA world. Genes Dev. 2009, 23, 1494–1504. [Google Scholar] [CrossRef]
- Baruah, P.M.; Krishnatreya, D.B.; Bordoloi, K.S.; Gill, S.S.; Agarwala, N. Genome wide identification and characterization of abiotic stress responsive lncRNAs in Capsicum annuum. Plant Physiol Biochem. 2021, 162, 221–236. [Google Scholar] [CrossRef]
- Baloglu, M.C.; Eldem, V.; Hajyzadeh, M.; Unver, T. Genome-wide analysis of the bZIP transcription factors in cucumber. PLoS One. 2014, 9, e96014. [Google Scholar] [CrossRef] [PubMed]
- Neysanian, M.; Iranbakhsh, A.; Ahmadvand, R.; Ardebili, Z.O.; Ebadi, M. Selenium nanoparticles conferred drought tolerance in tomato plants by altering the transcription pattern of microRNA-172 (miR-172), bZIP, and CRTISO genes, upregulating the antioxidant system, and stimulating secondary metabolism. Protoplasma. 2024, 261, 735–747. [Google Scholar] [CrossRef]
- Sun, X.; Fan, G.; Su, L.; Wang, W.; Liang, Z.; Li, S.; Xin, H. Identification of cold-inducible microRNAs in grapevine. Front Plant Sci. 2015, 6, 595. [Google Scholar] [CrossRef]
- Corrêa, L.G.; Riaño-Pachón, D.M.; Schrago, C.G.; dos Santos, R.V.; Mueller-Roeber, B.; Vincentz, M. The role of bZIP transcription factors in green plant evolution: adaptive features emerging from four founder genes. PLoS One. 2008, 3, e2944. [Google Scholar] [CrossRef]
- Wei, K.; Chen, J.; Wang, Y.; Chen, Y.; Chen, S.; Lin, Y.; Pan, S.; Zhong, X.; Xie, D. Genome-wide analysis of bZIP-encoding genes in maize. DNA Res. 2012, 19, 463–476. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Shi, H.; Guo, M.; Chai, M.; He, Q.; Yan, M.; Cao, D.; Zhao, L.; Cai, H.; Qin, Y. Evolutionary and expression analyses of soybean basic Leucine zipper transcription factor family. BMC Genomics. 2018, 19, 159. [Google Scholar] [CrossRef]
- Jin, Z.; Xu, W.; Liu, A. Genomic surveys and expression analysis of bZIP gene family in castor bean (Ricinus communis L.). Planta. 2014, 239, 299–312. [Google Scholar] [CrossRef]
- Li, Y.; Yao, G.; Tang, Y.; Lu, X.; Qiao, X.; Wang, C. Genome-Wide Survey and Expression Analysis of the Basic Leucine Zipper (bZIP) Gene Family in Eggplant (Solanum melongena L.). Horticulturae. 2022, 8, 1153. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, P.; Sharma, D.; Verma, S.K.; Halterman, D.; Kumar, A. Genome-wide identification and expression profiling of basic leucine zipper transcription factors following abiotic stresses in potato (Solanum tuberosum L.). PLoS One. 2021, 16, e0247864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, W.; Qi, X.; Liu, Z.; Xie, W.; Wang, Y. Genome-wide identification, expression profiling, and SSR marker development of the bZIP transcription factor family in Medicago truncatula. Biochemical Systematics and Ecology. 2015, 61, 218–228. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Y.; Zhang, S.; Xie, K.; Zhang, C.; Xi, Y.; Sun, F. Genome-wide analysis of the abiotic stress-related bZIP family in switchgrass. Mol Biol Rep. 2020, 47, 4439–4454. [Google Scholar] [CrossRef]
- Wang, X.L.; Zhong, Y.; Cheng, Z.M.; Xiong, J.S. Divergence of the bZIP Gene Family in Strawberry, Peach, and Apple Suggests Multiple Modes of Gene Evolution after Duplication. Int J Genomics. 2015, 2015, 536943. [Google Scholar] [CrossRef]
- Liu, Y.; Chai, M.; Zhang, M.; He, Q.; Su, Z.; Priyadarshani, S.V.G.N.; Liu, L.; Dong, G.; Qin, Y. Genome-Wide Analysis, Characterization, and Expression Profile of the Basic Leucine Zipper Transcription Factor Family in Pineapple. Int J Genomics. 2020, 2020, 3165958. [Google Scholar] [CrossRef]
- Zhao, K.; Chen, S.; Yao, W.; Cheng, Z.; Zhou, B.; Jiang, T. Genome-wide analysis and expression profile of the bZIP gene family in poplar. BMC Plant Biol. 2021, 21, 122. [Google Scholar] [CrossRef]
- Jing, Y.; Yu, Y.; Wang, H.; Wu, Y.; Li, C.H. Genome-wide identification and expression analysis of the bZIP gene family in silver birch (Betula pendula Roth.). J. For. Res. 2022, 33, 1615–1636. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, C.; Zhu, Q.; Guo, F.; Chai, R.; Wang, M.; Deng, X.; Dong, T.; Meng, X.; Zhu, M. Genome- and transcriptome-widesystematic characterization of bZIP transcription factor family identifies promising members involved in abiotic stress responsein sweetpotato. Sci. Hortic. 2022, 303, 111185. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, J.; Zhang, B.; Vanitha, J.; Ramachandran, S.; Jiang, S.Y. Genome-wide expansion and expression divergence of the basic leucine zipper transcription factors in higher plants with an emphasis on sorghum. J Integr Plant Biol. 2011, 53, 212–231. [Google Scholar] [CrossRef]
- Zhang, Z.; Quan, S.; Niu, J.; Guo, C.; Kang, C.; Liu, J.; Yuan, X. Genome-Wide Identification, Classification, Expression and Duplication Analysis of bZIP Family Genes in Juglans regia L. Int J Mol Sci. Int J Mol Sci. 2022, 23, 5961. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, D.; Jia, L.; Huang, X.; Ma, G.; Wang, S.; Zhu, M.; Zhang, A.; Guan, M.; Lu, K.; Xu, X.; Wang, R.; Li, J.; Qu, C. Genome-Wide Identification and Structural Analysis of bZIP Transcription Factor Genes in Brassica napus. Genes (Basel). 2017, 8, 288. [Google Scholar] [CrossRef]
- Zheng, Q.; Meng, X.; Fan, X.; Chen, S.; Sang, K.; Yu, J.; Zhou, Y.; Xia, X. Regulation of PILS genes by bZIP transcription factor TGA7 in tomato plant growth. Plant Sci. 2025, 352, 112359. [Google Scholar] [CrossRef]
- Nardeli, S.M.; Arge, L.W.P.; Artico, S.; de Moura, S.M.; Tschoeke, D.A.; de Freitas, Guedes, F.A.; Grossi-de-Sa, M.F.; Martinelli, A.P.; Alves-Ferreira, M. Global gene expression profile and functional analysis reveal the conservation of reproduction-associated gene networks in Gossypium hirsutum. Plant Reprod. 2024, 37, 215–227. [Google Scholar] [CrossRef]
- Liu, X.; Chu, Z. Genome-wide evolutionary characterization and analysis of bZIP transcription factors and their expression profiles in response to multiple abiotic stresses in Brachypodium distachyon. BMC Genomics. 2015, 16, 227. [Google Scholar] [CrossRef]
- Pourabed, E.; Ghane, G.F.; Soleymani, M.P.; Razavi, S.M.; Shobbar, Z.S. Basic leucine zipper family in barley: genome-wide characterization of members and expression analysis. Mol Biotechnol. 2015, 57, 12–26. [Google Scholar] [CrossRef]
- Mirzaei, K.; Bahramnejad, B.; Fatemi, S. Genome⁃wide identification and characterization of the bZIP gene family in potato (Solanum tuberosum). Plant Gene. 2020, 24, 100257–100267. [Google Scholar] [CrossRef]
- Liu, J.X.; Srivastava, R.; Che, P.; Howell, S.H. Salt stress responses in Arabidopsis utilize a signal transduction pathway related to endoplasmic reticulum stress signaling. Plant J. 2007, 51, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Veerabagu, M.; Kirchler, T.; Elgass, K.; Stadelhofer, B.; Stahl, M.; Harter, K.; Mira-Rodado, V.; Chaban, C. The interaction of the Arabidopsis response regulator ARR18 with bZIP63 mediates the regulation of PROLINE DEHYDROGENASE expression. Mol Plant. 2014, 7, 1560–1577. [Google Scholar] [CrossRef] [PubMed]
- Van, L.J.; Blomme, J.; Kulkarni, S.R.; Cannoot, B.; De, W.N.; Eeckhout, D.; Persiau, G.; Van, D.S.E.; Vercruysse, L.; Vanden Bossche, R.; Heyndrickx, K.S.; Vanneste, S.; Goossens, A.; Gevaert, K.; Vandepoele, K.; Gonzalez, N.; Inzé, D.; De, J.G. Functional characterization of the Arabidopsis transcription factor bZIP29 reveals its role in leaf and root development. J Exp Bot. 2016, 67, 5825–5840. [Google Scholar]
- Tang, W.; Page, M. Transcription factor AtbZIP60 regulates expression of Ca2+ -dependent protein kinase genes in transgenic cells. Mol Biol Rep. 2013, 40, 2723–2732. [Google Scholar] [CrossRef]
- Lakra, N.; Nutan, K.K.; Das, P.; Anwar, K.; Singla-Pareek, S.L.; Pareek, A. A nuclear-localized histone-gene binding protein from rice (OsHBP1b) functions in salinity and drought stress tolerance by maintaining chlorophyll content and improving the antioxidant machinery. J Plant Physiol. 2015, 176, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Lakra, N.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. A unique bZIP transcription factor imparting multiple stress tolerance in Rice. Rice (N Y). 2019, 12, 58. [Google Scholar] [CrossRef]
- Mukherjee, K.; Choudhury, A.R.; Gupta, B.; Gupta, S.; Sengupta, D.N. An ABRE-binding factor, OSBZ8, is highly expressed in salt tolerant cultivars than in salt sensitive cultivars of indica rice. BMC Plant Biol. 2006, 6, 18. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, J.; Li, Z.; Qiao, J.; Quan, R.; Wang, J.; Huang, R.; Qin, H. SALT AND ABA RESPONSE ERF1 improves seed germination and salt tolerance by repressing ABA signaling in rice. Plant Physiol. 2022, 189, 1110–1127. [Google Scholar] [CrossRef]
- Amir, H.M.; Lee, Y.; Cho, J.I.; Ahn, C.H.; Lee, S.K.; Jeon, J.S.; Kang, H.; Lee, C.H.; An, G.; Park, P.B. The bZIP transcription factor OsABF1 is an ABA responsive element binding factor that enhances abiotic stress signaling in rice. Plant Mol Biol. 2010, 72, 557–566. [Google Scholar] [CrossRef]
- Joo, J.; Lee, Y.H.; Song, S.I. Overexpression of the rice basic leucine zipper transcription factor OsbZIP12 confers drought tolerance to rice and makes seedlings hypersensitive to ABA. Plant Biotechnology Reports. 2014, 8, 431–441. [Google Scholar] [CrossRef]
- Chen, H.; Chen, W.; Zhou, J.; He, H.; Chen, L.; Chen, H.; Deng, X.W. Basic leucine zipper transcription factor OsbZIP16 positively regulates drought resistance in rice. Plant Sci 2012, 193-194, 8–17. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, B.; Ren, J.; Zhou, Y.; Han, Y.; Niu, S.; Zhang, Y.; Shi, Y.; Zhou, J.; Yang, C.; Ma, X.; Liu, X.; LuoV, Y.; Jin, C.; Luo, J. OsbZIP18, a Positive Regulator of Serotonin Biosynthesis, Negatively Controls the UV-B Tolerance in Rice. Int J Mol Sci. 2022, 23, 3215. [Google Scholar] [CrossRef]
- Wang, B.X.; Xu, B.; Liu, Y.; Li, J.F.; Sun, Z.G.; Chi, M.; Xing, Y.G.; Yang, B.; Li, J.; Liu, J.B.; Chen, T.M.; Fang, Z.W.; Lu, B.G.; Xu, D.Y.; Babatunde, K.B. A novel mechanisms of the signaling cascade associated with the SAPK10-bZIP20 NHX1 synergistic interaction to enhance tolerance of plant to abiotic stress in rice (Oryza sativa L.). Plant Science. 2022, 323, 111393. [Google Scholar]
- Zong, W.; Yang, J.; Fu, J.; Xiong, L. Synergistic regulation of drought-responsive genes by transcription factor OsbZIP23 and histone modification in rice. J Integr Plant Biol. 2020, 62, 723–729. [Google Scholar] [CrossRef]
- Shimizu, H.; Sato, K.; Berberich, T.; Miyazaki, A.; Ozaki, R.; Imai, R.; Kusano, T. LIP19, a basic region leucine zipper protein, is a Fos-like molecular switch in the cold signaling of rice plants. Plant Cell Physiol. 2005, 46, 1623–1634. [Google Scholar] [CrossRef]
- Lilay, G.H.; Castro, P.H.; Guedes, J.G.; Almeida, D.M.; Campilho, A.; Azevedo, H.; Aarts, M.G.M.; Saibo, N.J.M.; Assunção, A.G.L. Rice F-bZIP transcription factors regulate the zinc deficiency response. J Exp Bot. 2020, 71, 3664–3677. [Google Scholar] [CrossRef]
- Park, S.H.; Jeong, J.S.; Lee, K.H.; Kim, Y.S.; Choi, Y.D.; Kim, J.K. OsbZIP23 and OsbZIP45, members of the rice basic leucine zipper transcription factor family, are involved in drought tolerance. Plant Biotechnology Reports. 2015, 9, 89–96. [Google Scholar] [CrossRef]
- Tang, N.; Ma, S.; Zong, W.; Yang, N.; Lv, Y.; Yan, C.; Guo, Z.; Li, J.; Li, X.; Xiang, Y.; Song, H.; Xiao, J.; Li, X.; Xiong, L. MODD Mediates Deactivation and Degradation of OsbZIP46 to Negatively Regulate ABA Signaling and Drought Resistance in Rice. Plant Cell. 2016, 28, 2161–2177. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Niu, X.L.; Yang, S.H.; Li, Y.X.; Liu, L.L.; Tang, W.; Liu, Y.S. Research on heat and drought tolerance in rice (Oryza sativa L.) by overexpressing transcription factor OsbZIP60. Scientia Agricultura Sinica. 2011, 44, 4142–4149. [Google Scholar]
- Yang, S.; Xu, K.; Chen, S.; Li, T.; Xia, H.; Chen, L.; Liu, H.; Luo, L. A stress-responsive bZIP transcription factor OsbZIP62 improves drought and oxidative tolerance in rice. BMC Plant Biol. 2019, 19, 260. [Google Scholar] [CrossRef]
- Yoon, S.; Lee, D.K.; Yu, I.J.; Kim, Y.S.; Choi, Y.D.; Kim, J.K. Overexpression of the OsbZIP66 transcription factor enhances drought tolerance of rice plants. Plant Biotechnology Reports, 2017, 11, 53–62. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Y.; Wang, Y.; Li, J.; Sun, Z.; Chi, M.; Xing, Y.; Xu, B.; Yang, B.; Li, J.; Liu, J.; Chen, T.; Fang, Z.; Lu, B.; Xu, D.; Babatunde, K.B. OsbZIP72 is involved in transcriptional gene-regulation pathway of abscisic acid signal transduction by activating rice high-affinity potassium transporter OsHKT1;1. Rice Science, 2021, 28, 257–267. [Google Scholar]
- Liu, C.; Schläppi, M.R.; Mao, B.; Wang, W.; Wang, A.; Chu, C. The bZIP73 transcription factor controls rice cold tolerance at the reproductive stage. Plant Biotechnol J. 2019, 17, 1834–1849. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Li, M.; Yang, S.; Gao, C.; Su, Y.; Zeng, X.; Jiao, Z.; Xu, W.; Zhang, M.; Xia, K. miR2105 and the kinase OsSAPK10 co-regulate OsbZIP86 to mediate drought-induced ABA biosynthesis in rice. Plant Physiol. 2022, 189, 889–905. [Google Scholar] [CrossRef]
- Wang, C.; Lu, G.; Hao, Y.; Guo, H.; Guo, Y.; Zhao, J.; Cheng, H. ABP9, a maize bZIP transcription factor, enhances tolerance to salt and drought in transgenic cotton. Planta. 2017, 246, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ma, Q.; Jin, X.; Peng, X.; Liu, J.; Deng, L.; YanV, H.; Sheng, L.; JiangV, H.; Cheng, B. A novel maize homeodomain-leucine zipper (HD-Zip) I gene, Zmhdz10, positively regulates drought and salt tolerance in both rice and Arabidopsis. Plant Cell Physiol. 2014, 55, 1142–1156. [Google Scholar] [CrossRef]
- Wang, Z.; Cheng, K.; Wan, L.; Yan, L.; Jiang, H.; Liu, S.; Lei, Y.; Liao, B. Genome-wide analysis of the basic leucine zipper (bZIP) transcription factor gene family in six legume genomes. BMC Genomics. 2015, 16, 1053. [Google Scholar] [CrossRef]
- Bi, C.; Yu, Y.; Dong, C.; Yang, Y.; Zhai, Y.; Du, F.; Xia, C.; Ni, Z.; Kong, X.; Zhang, L. The bZIP transcription factor TabZIP15 improves salt stress tolerance in wheat. Plant Biotechnol J. 2021, 19, 209–211. [Google Scholar] [CrossRef]
- Han, N.N.; Fan, S.Y.; Zhang, T.T.; et al. SlHY5 is a necessary regulator of the cold acclimation response in tomato. Plant Growth Regul. 2020, 91, 1–12. [Google Scholar] [CrossRef]
- Hsieh, T.H.; Li, C.W.; Su, R.C.; Cheng, C.P.; Sanjaya; Tsai, Y.C.; Chan, M.T. A tomato bZIP transcription factor, SlAREB, is involved in water deficit and salt stress response. Planta. 2010, 231, 1459–1473. [Google Scholar] [CrossRef]
- Kaminaka, H.; Näke, C.; Epple, P.; Dittgen, J.; Schütze, K.; Chaban, C.; Holt, B.F.; Merkle, T.; Schäfer, E.; Harter, K.; Dangl, J.L. bZIP10-LSD1 antagonism modulates basal defense and cell death in Arabidopsis following infection. EMBO J. 2006, 25, 4400–4411. [Google Scholar] [CrossRef]
- Okada, A.; Okada, K.; Miyamoto, K.; Koga, J.; Shibuya, N.; Nojiri, H.; Yamane, H. OsTGAP1, a bZIP transcription factor, coordinately regulates the inductive production of diterpenoid phytoalexins in rice. J Biol Chem. 2009, 284, 26510–26518. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Nishizawa, Y.; Minami, E.; Nojiri, H.; Yamane, H.V.; Okada, K. Overexpression of the bZIP transcription factor OsbZIP79 suppresses the production of diterpenoid phytoalexin in rice cells. J Plant Physiol. 2015, 173, 19–27. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, S.; Liu, Z.; Wang, L.; Bi, Y. Both HY5 and HYH are necessary regulators for low temperature-induced anthocyanin accumulation in Arabidopsis seedlings. J Plant Physiol. 2011, 168, 367–374. [Google Scholar] [CrossRef]
- Zhang, F.; Xiang, L.; Yu, Q.; Zhang, H.; Zhang, T.; Zeng, J.; Geng, C.; Li, L.; Fu, X.; Shen, Q.; Yang, C.; Lan, X.; Chen, M.; Tang, K.; Liao, Z. ARTEMISININ BIOSYNTHESIS PROMOTING KINASE 1 positively regulates artemisinin biosynthesis through phosphorylating AabZIP1. J Exp Bot. 2018, 69, 1109–1123. [Google Scholar] [CrossRef]
- Hao, X.; Zhong, Y.; Nï, T.H.W.; Fu, X.; Yan, T.; Shen, Q.; Chen, M.; Ma, Y.; Zhao, J.; Osbourn, A.; Li, L.; Tang, K. Light-Induced Artemisinin Biosynthesis Is Regulated by the bZIP Transcription Factor AaHY5 in Artemisia annua. Plant Cell Physiol. 2019, 60, 1747–1760. [Google Scholar] [CrossRef]
- Lv, Z.; Guo, Z.; Zhang, L.; Zhang, F.; Jiang, W.; Shen, Q.; Fu, X.; Yan, T.; Shi, P.; Hao, X.; Ma, Y.; Chen, M.; Li, L.; Zhang, L.; Chen, W.; Tang, K. Interaction of bZIP transcription factor TGA6 with salicylic acid signaling modulates artemisinin biosynthesis in Artemisia annua. J Exp Bot. 2019, 70, 3969–3979. [Google Scholar] [CrossRef]
- Deng, C.; Shi, M.; Fu, R.; Zhang, Y.; Wang, Q.; Zhou, Y.; Wang, Y.; Ma, X.; Kai, G. ABA-responsive transcription factor bZIP1 is involved in modulating biosynthesis of phenolic acids and tanshinones in Salvia miltiorrhiza. J Exp Bot. 2020, 71, 5948–5962. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Z.; Ji, A.; Luo, H.; Song, J. Genomic survey of bZIP transcription factor genes related to tanshinone biosynthesis in Salvia miltiorrhiza. Acta Pharm Sin B. 2018, 8, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Wakamatsu, S.; Sakuta, M. Characterization of SBZ1, a soybean bZIP protein that binds to the chalcone synthase gene promoter. Plant Biotechnol. 2008, 25, 131–140. [Google Scholar] [CrossRef]
- Dröge, L.W.; Kaiser, A.; Lindsay, W.P.; Halkier, B.A.; Loake, G.J.; Doerner, P.; Dixon, R.A.; Lamb, C. Rapid stimulation of a soybean protein-serine kinase that phosphorylates a novel bZIP DNA-binding protein, G/HBF-1, during the induction of early transcription-dependent defenses. EMBO J. 1997, 16, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Anguraj, V.A.K.; Renaud, J.; Kagale, S.; Dhaubhadel, S. GmMYB176 Regulates Multiple Steps in Isoflavonoid Biosynthesis in Soybean. Front Plant Sci. 2019, 10, 562. [Google Scholar] [CrossRef]
- Akagi, T.; Katayama-Ikegami, A.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. Seasonal abscisic acid signal and a basic leucine zipper transcription factor, DkbZIP5, regulate proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2012, 158, 1089–1102. [Google Scholar] [CrossRef]
- Sibéril, Y.; Benhamron, S.; Memelink, J.; Giglioli-Guivarc'h, N.; Thiersault, M.; Boisson, B.; Doireau, P.; Gantet, P. Catharanthus roseus G-box binding factors 1 and 2 act as repressors of strictosidine synthase gene expression in cell cultures. Plant Mol Biol. 2001, 45, 477–488. [Google Scholar] [CrossRef]
- Fricke, J.; Hillebrand, A.; Twyman, R.M.; Prüfer, D.; Schulze, G.C. Abscisic acid-dependent regulation of small rubber particle protein gene expression in Taraxacum brevicorniculatum is mediated by TbbZIP1. Plant Cell Physiol. 2013, 54, 448–464. [Google Scholar] [CrossRef]
- Seong, E.S.; Lee, JG.; Chung, I.M.; Yu, CY. Changes of phenolic compounds in LebZIP2-overexpressing transgenic plants. Indian J Biochem Biophys. 2019, 56, 484–491. [Google Scholar]
- Shi, M.; Du, Z.; Hua, Q.; Kai, G. CRISPR/Cas9-mediated targeted mutagenesis of bZIP2 in Salvia miltiorrhiza leads to promoted phenolic acid biosynthesis. Ind Crops Prod. 2021, 167, 113560. [Google Scholar] [CrossRef]
- Matousek, J.; Kocábek, T.; Patzak, J.; Stehlík, J.; Füssy, Z.; Krofta, K.; Heyerick, A.; Roldán-Ruiz, I.; Maloukh, L.; De Keukeleire, D. Cloning and molecular analysis of HlbZip1 and HlbZip2 transcription factors putatively involved in the regulation of the lupulin metabolome in hop (Humulus lupulus L.). J Agric Food Chem. 2010, 58, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Malacarne, G.; Coller, E.; Czemmel, S.; Vrhovsek, U.; Engelen, K.; Goremykin, V.; Bogs, J.; Moser, C. The grapevine VvibZIPC22 transcription factor is involved in the regulation of flavonoid biosynthesis. J Exp Bot. 2016, 67, 3509–3522. [Google Scholar] [CrossRef] [PubMed]
- Collin, A.; Daszkowska-Golec, A.; Kurowska, M.; Szarejko, I. Barley ABI5 (Abscisic Acid INSENSITIVE 5) Is Involved in Abscisic Acid-Dependent Drought Response. Front Plant Sci. 2020, 11, 1138. [Google Scholar] [CrossRef]
- Li, Y.; Xu, P.; Chen, G.; Wu, J.; Liu, Z.; Lian, H. FvbHLH9 Functions as a Positive Regulator of Anthocyanin Biosynthesis by Forming a HY5-bHLH9 Transcription Complex in Strawberry Fruits. Plant Cell Physiol. 2020, 61, 826–837. [Google Scholar] [CrossRef]
- Akagi, T.; Ikegami, A.; Tsujimoto, T.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. DkMyb4 is a Myb transcription factor involved in proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2009, 151, 2028–2045. [Google Scholar] [CrossRef]
- Akagi, T.; Katayama, I.A.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. Seasonal abscisic acid signal and a basic leucine zipper transcription factor, DkbZIP5, regulate proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2012, 158, 1089–1102. [Google Scholar] [CrossRef]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. Apple bZIP transcription factor MdbZIP44 regulates abscisic acid-promoted anthocyanin accumulation. Plant Cell Environ. 2018, 41, 2678–2692. [Google Scholar] [CrossRef]
- An, J.P.; Zhang, X.W.; Liu, Y.J.; Wang, X.F.; You, C.X.; Hao, Y.J. ABI5 regulates ABA-induced anthocyanin biosynthesis by modulating the MYB1-bHLH3 complex in apple. J Exp Bot. 2021, 72, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Su, J.; Zhu, Y.; Yao, G.; Allan, A.C.; Ampomah, D.C.; Shu, Q.; Lin, W.K.; Zhang, S.; Wu, J. The involvement of PybZIPa in light-induced anthocyanin accumulation via the activation of PyUFGT through binding to tandem G-boxes in its promoter. Hortic Res. 2019, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Zhao, Y.; Yang, J.; He, Y.; Li, G.; Ma, W.; Huang, X.; Su, J. Transcription factor PyHY5 binds to the promoters of PyWD40 and PyMYB10 and regulates its expression in red pear 'Yunhongli No. 1'. Plant Physiol Biochem. 2020, 154, 665–674. [Google Scholar] [CrossRef]
- Xu, J.; Wu, S.; Xu, Y.H.; Ge, Z.; Sui, C.; Wei, J. Overexpression of BcbZIP134 negatively regulates the biosynthesis of saikosaponins. Plant Cell Tissue Organ Cult. 2019, 137, 297–308. [Google Scholar] [CrossRef]
- Wang, C.C.; Meng, L.H.; Gao, Y.; Grierson, D.; Fu, D.Q. Manipulation of Light Signal Transduction Factors as a Means of Modifying Steroidal Glycoalkaloids Accumulation in Tomato Leaves. Front Plant Sci. 2018, 9, 437. [Google Scholar] [CrossRef]
- Chang, C.; Liu, Z.; Wang, Y.; Tang, Z.; Yu, F. A bZIP transcription factor, CaLMF, mediated light-regulated camptothecin biosynthesis in Camptotheca acuminata. Tree Physiol. 2019, 39, 372–380. [Google Scholar] [CrossRef]
Figure 1.
The bZIP transcription factors involved in the regulatory mode in plants growth and development, abiotic and biotic stress response.
Figure 1.
The bZIP transcription factors involved in the regulatory mode in plants growth and development, abiotic and biotic stress response.
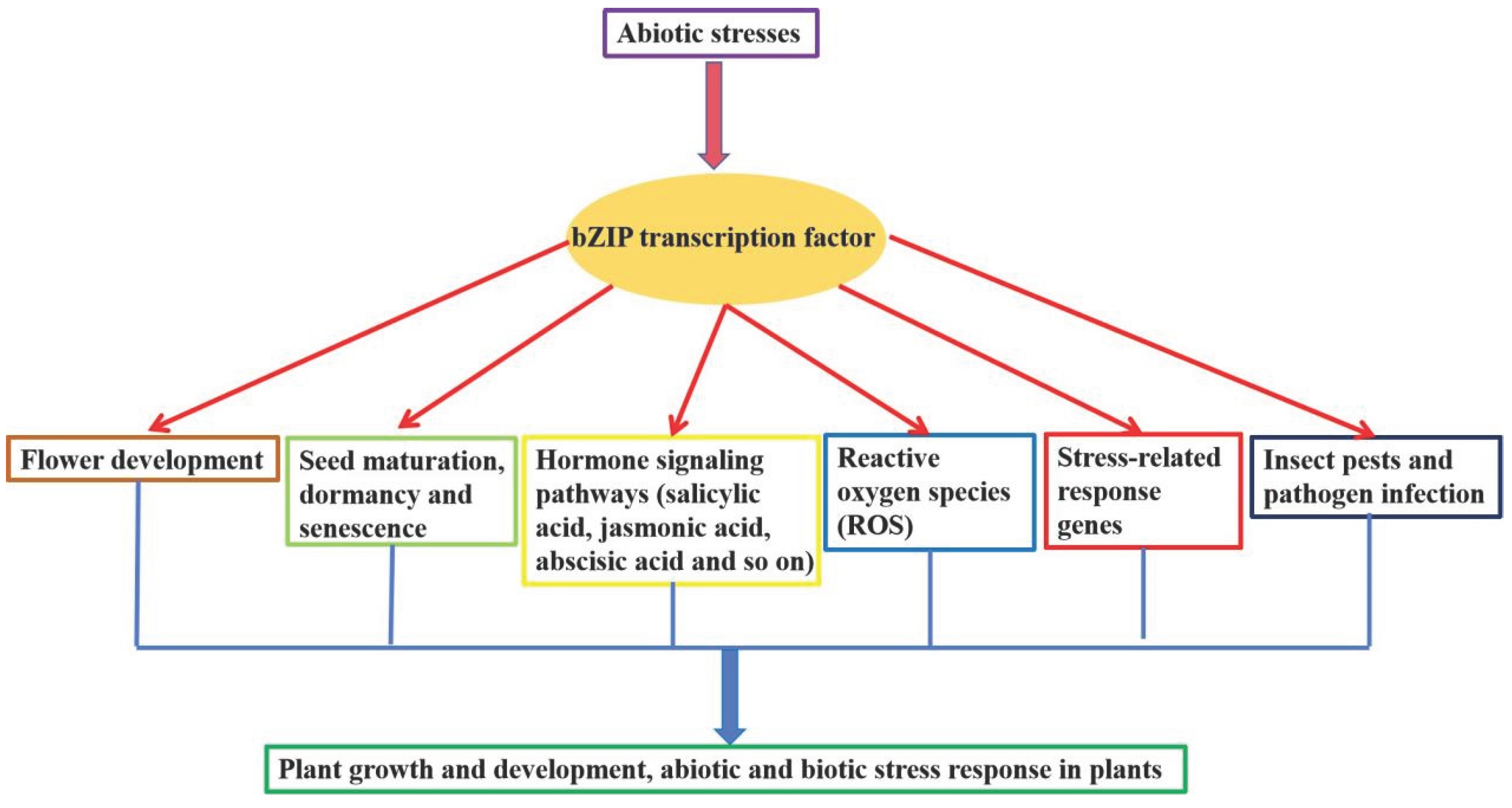
Figure 2.
(A) Primary structure of the bZIP domain. Schematic representation of the bZIP transcription factors domain, consisting of a basic DNA-binding region (blue) and the leucine zipper region (green) respectively. (B) Structural model of a dimer of the bZIP transcription factors domains, which is derived by PDB website (www. rcsb.org). The leucine zip region is composed of one or more repeats containing 7 amino acid residues, where the 7th position of each repeat is leucine, and the rest of the amino acids include isoleucine, methionine, valine, methionine and so on. This region can mediate the formation of homo- or heterodimers of bZIP transcription factor proteins with transcriptional activation or repression functions. Other hydrophobic residues may be present in positions 3 and 4. The leucine zip region allows the bZIP transcription factor protein to bind to DNA in a dimerised form by forming an amphipathic α-helix structure.
Figure 2.
(A) Primary structure of the bZIP domain. Schematic representation of the bZIP transcription factors domain, consisting of a basic DNA-binding region (blue) and the leucine zipper region (green) respectively. (B) Structural model of a dimer of the bZIP transcription factors domains, which is derived by PDB website (www. rcsb.org). The leucine zip region is composed of one or more repeats containing 7 amino acid residues, where the 7th position of each repeat is leucine, and the rest of the amino acids include isoleucine, methionine, valine, methionine and so on. This region can mediate the formation of homo- or heterodimers of bZIP transcription factor proteins with transcriptional activation or repression functions. Other hydrophobic residues may be present in positions 3 and 4. The leucine zip region allows the bZIP transcription factor protein to bind to DNA in a dimerised form by forming an amphipathic α-helix structure.
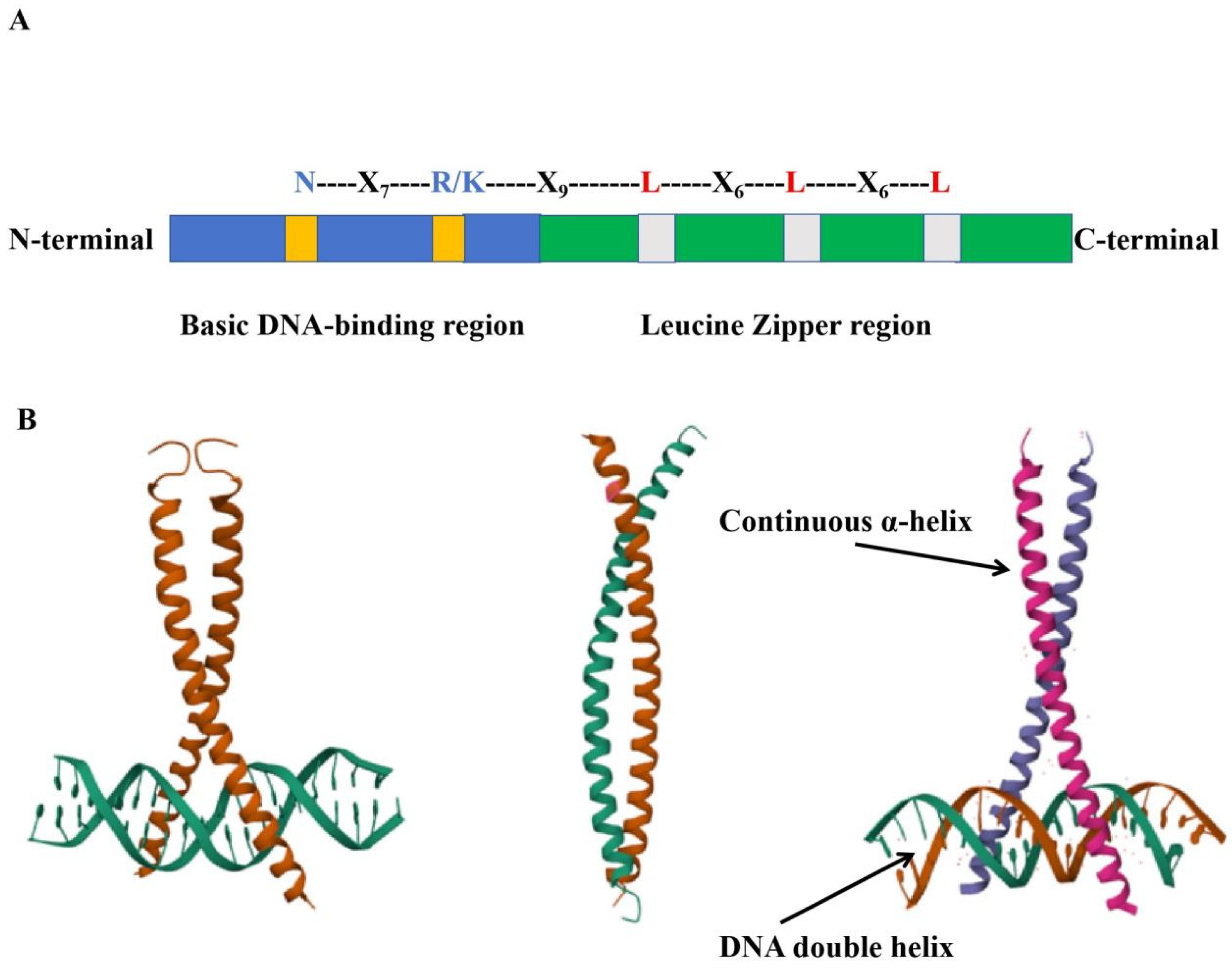
Figure 3.
The bZIP transcription factor regulatory mechanism. The arrows in the figure indicate the route of action.
Figure 3.
The bZIP transcription factor regulatory mechanism. The arrows in the figure indicate the route of action.
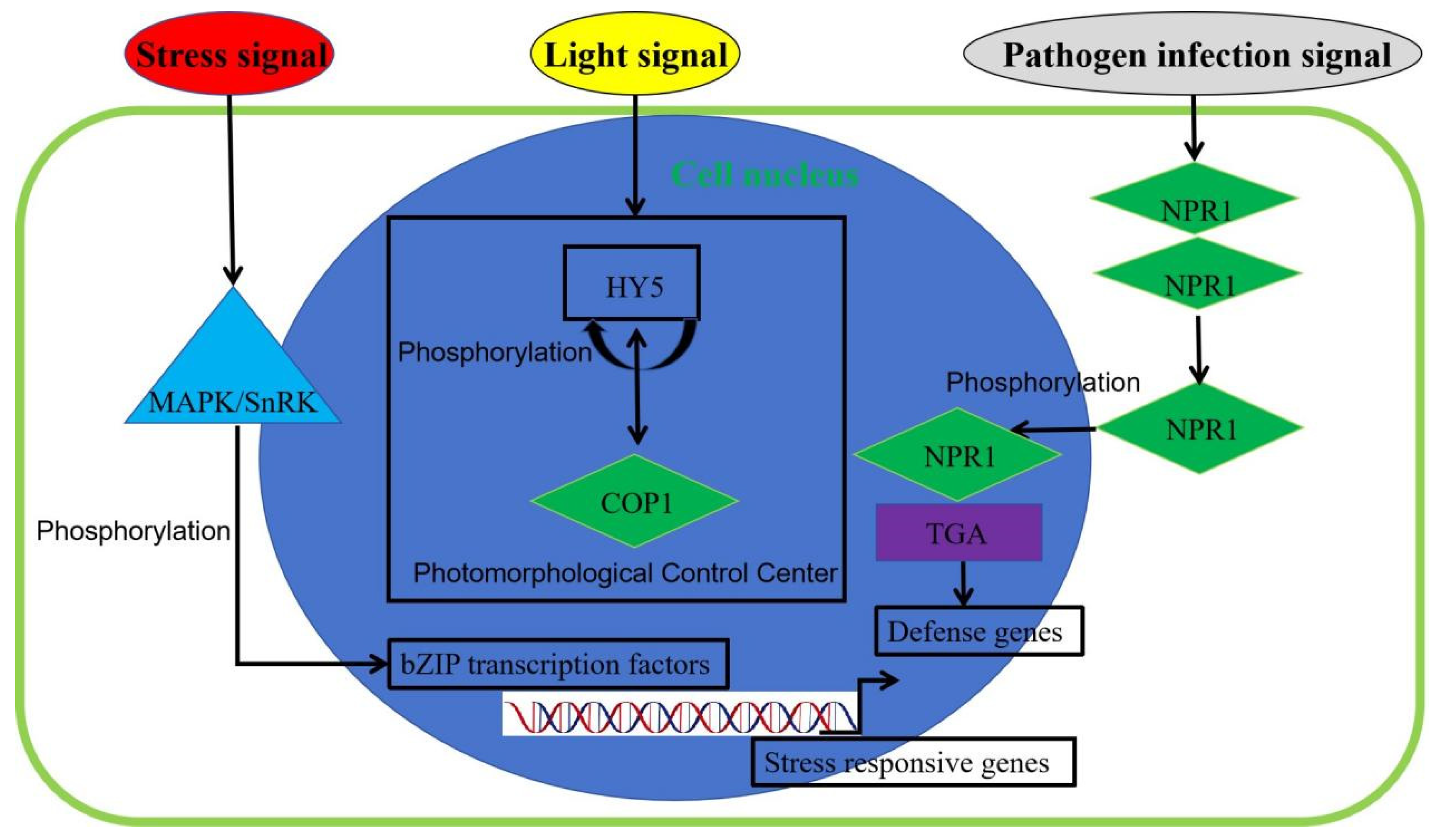
Figure 4.
Regulatory function of bZIP transcription factors under abiotic stress in plants. Multiple members of the bZIP transcription factor family enhance plant resistance to abiotic stresses such as high temperature, low temperature, salt, and drought by participating in osmotic regulation, reactive oxygen species scavenging and redox balance, hormone signal transduction, and other pathways.
Figure 4.
Regulatory function of bZIP transcription factors under abiotic stress in plants. Multiple members of the bZIP transcription factor family enhance plant resistance to abiotic stresses such as high temperature, low temperature, salt, and drought by participating in osmotic regulation, reactive oxygen species scavenging and redox balance, hormone signal transduction, and other pathways.
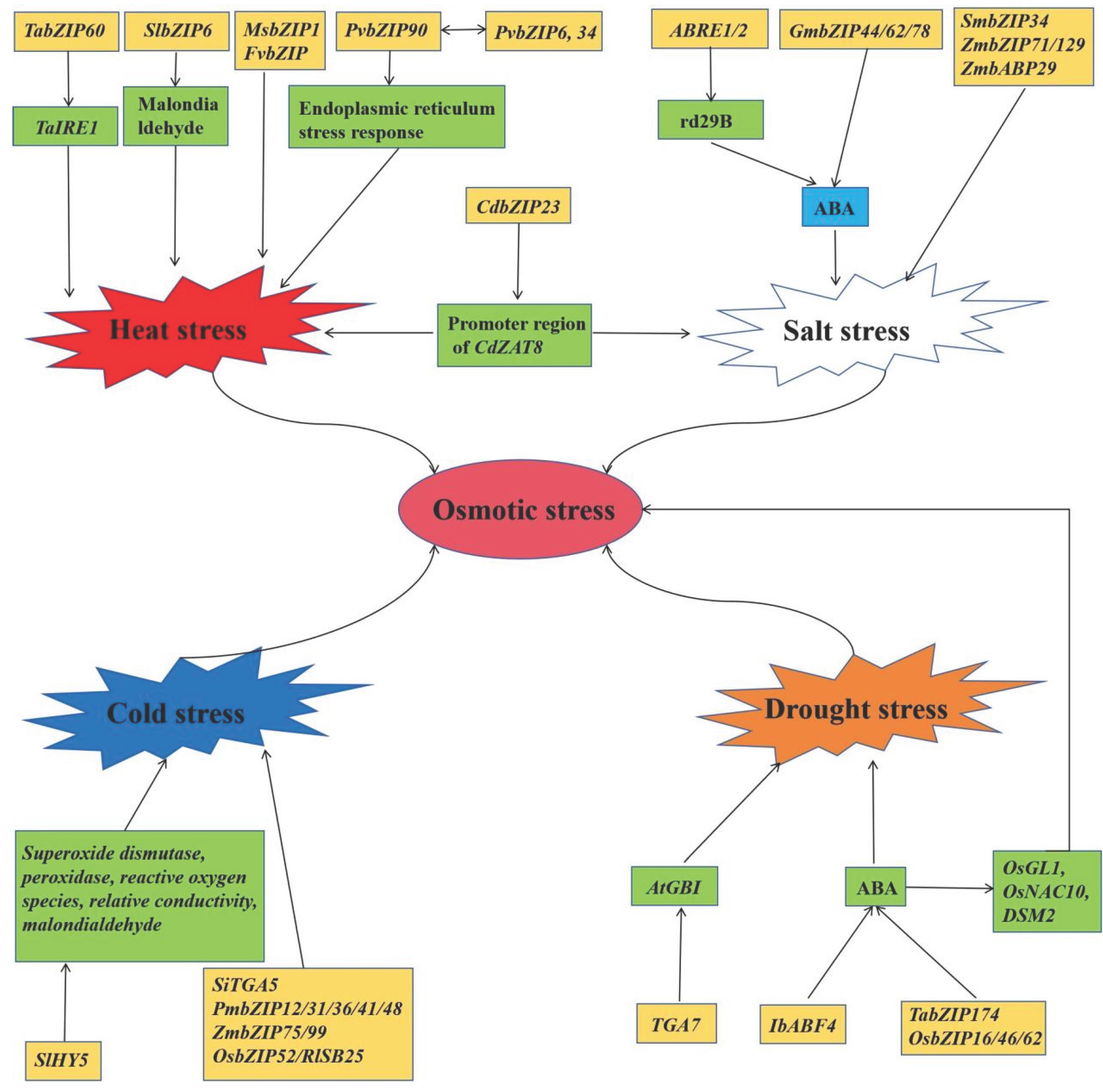
Figure 5.
The bZIP transcription factors involved in abiotic stress response pathways. The bZIP transcription factors participate in pathways related to salt stress and low temperature stress in plants. (Ⅰ). The bZIP TF specifically binds to the TGACGG element to regulate the expression of downstream salt stress response genes; (ⅠⅠ). The bZIP TF binds to the promoter of the OsNHX1 anti-transporter gene, transporting Na+ and K+; (ⅠⅡ). The bZIP TF activates the activity of antioxidant enzymes (such as SOD and APX) and removes ROS from plants; (Ⅳ). bZIP TF respond to ABA signal transduction; bZIP transcription factors recognise the ACGT sequence in the promoter region of ABA-induced genes, thereby regulating downstream COR genes. (V). Under low-temperature conditions, oxidative stress occurs, and ethylene response factors (ethylene response factor, AP2/ERF) regulate downstream low-temperature-related genes by activating bZIP transcription factors.
Figure 5.
The bZIP transcription factors involved in abiotic stress response pathways. The bZIP transcription factors participate in pathways related to salt stress and low temperature stress in plants. (Ⅰ). The bZIP TF specifically binds to the TGACGG element to regulate the expression of downstream salt stress response genes; (ⅠⅠ). The bZIP TF binds to the promoter of the OsNHX1 anti-transporter gene, transporting Na+ and K+; (ⅠⅡ). The bZIP TF activates the activity of antioxidant enzymes (such as SOD and APX) and removes ROS from plants; (Ⅳ). bZIP TF respond to ABA signal transduction; bZIP transcription factors recognise the ACGT sequence in the promoter region of ABA-induced genes, thereby regulating downstream COR genes. (V). Under low-temperature conditions, oxidative stress occurs, and ethylene response factors (ethylene response factor, AP2/ERF) regulate downstream low-temperature-related genes by activating bZIP transcription factors.
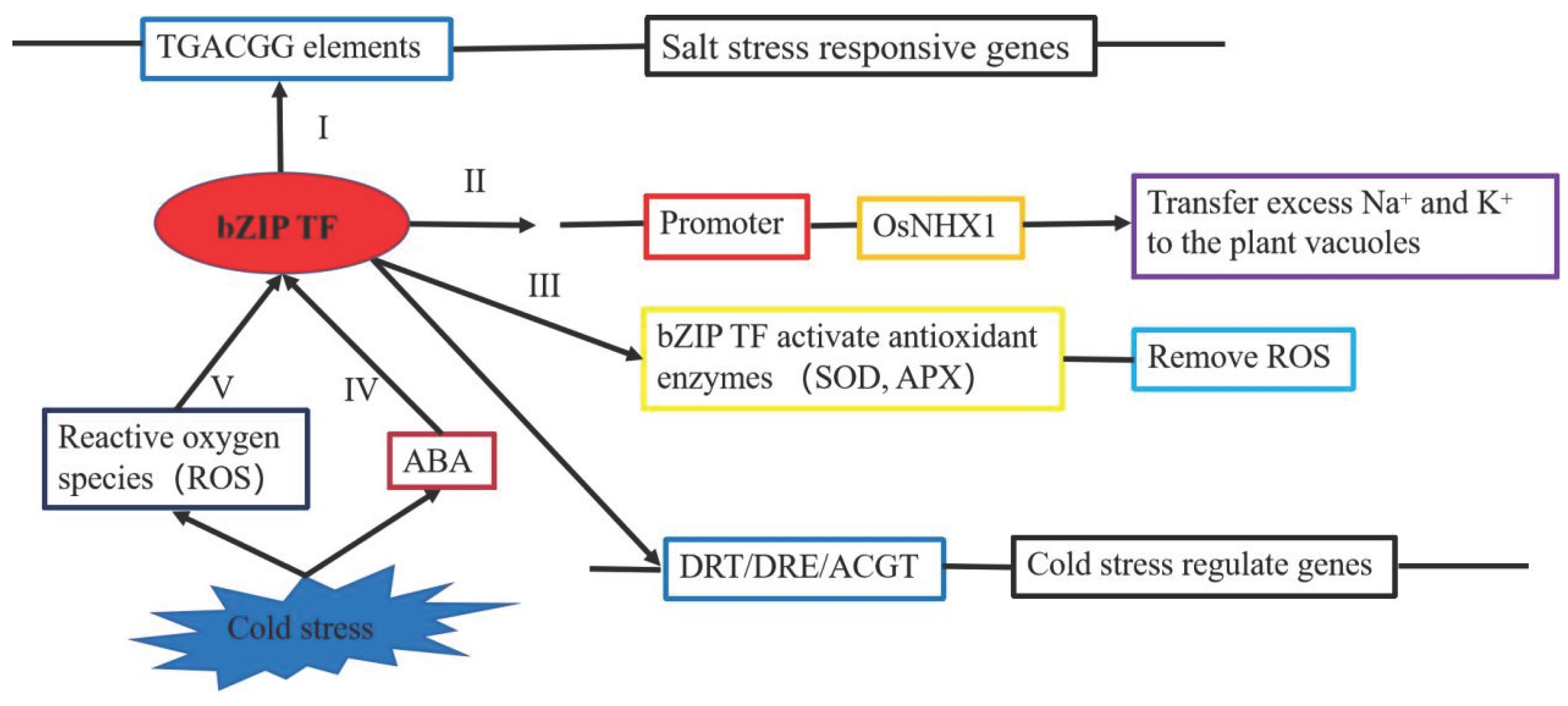
Figure 6.
Schematic diagram of the bZIP transcription factors participate in flavonoid biosynthetic pathway in plants.
Figure 6.
Schematic diagram of the bZIP transcription factors participate in flavonoid biosynthetic pathway in plants.
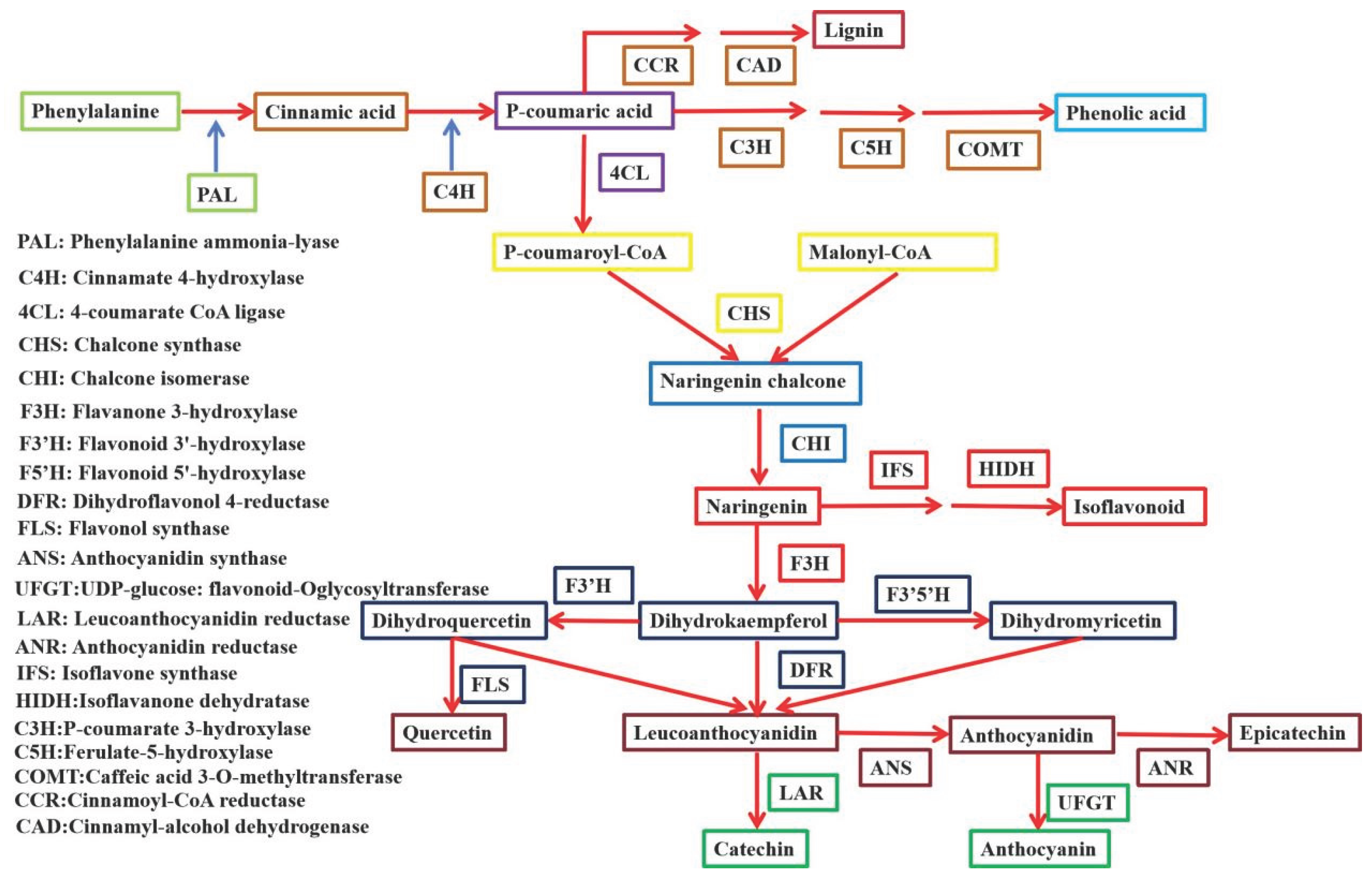
Figure 7.
The bZIP transcription factors participate in the ABA-mediated regulation mechanism of artemisinin biosynthesis. ABA binds to the receptor protein AaPLY9, causing a conformational change and forming a complex with the 2C-type serine/threonine residue protein phosphatase (PP2C), thereby inhibiting the phosphatase activity of PP2C and activating the activity of the sucrose non-glycolytic protein kinase (SnRK2). SnRK2 activates the downstream transcription factor AabZIP1 through protein phosphorylation. AabZIP1 binds to the cis-acting element ABRE to activate the expression of the ADS and CYP71AV1 genes. Through a series of intermediate products, artemisinin is ultimately synthesised.
Figure 7.
The bZIP transcription factors participate in the ABA-mediated regulation mechanism of artemisinin biosynthesis. ABA binds to the receptor protein AaPLY9, causing a conformational change and forming a complex with the 2C-type serine/threonine residue protein phosphatase (PP2C), thereby inhibiting the phosphatase activity of PP2C and activating the activity of the sucrose non-glycolytic protein kinase (SnRK2). SnRK2 activates the downstream transcription factor AabZIP1 through protein phosphorylation. AabZIP1 binds to the cis-acting element ABRE to activate the expression of the ADS and CYP71AV1 genes. Through a series of intermediate products, artemisinin is ultimately synthesised.
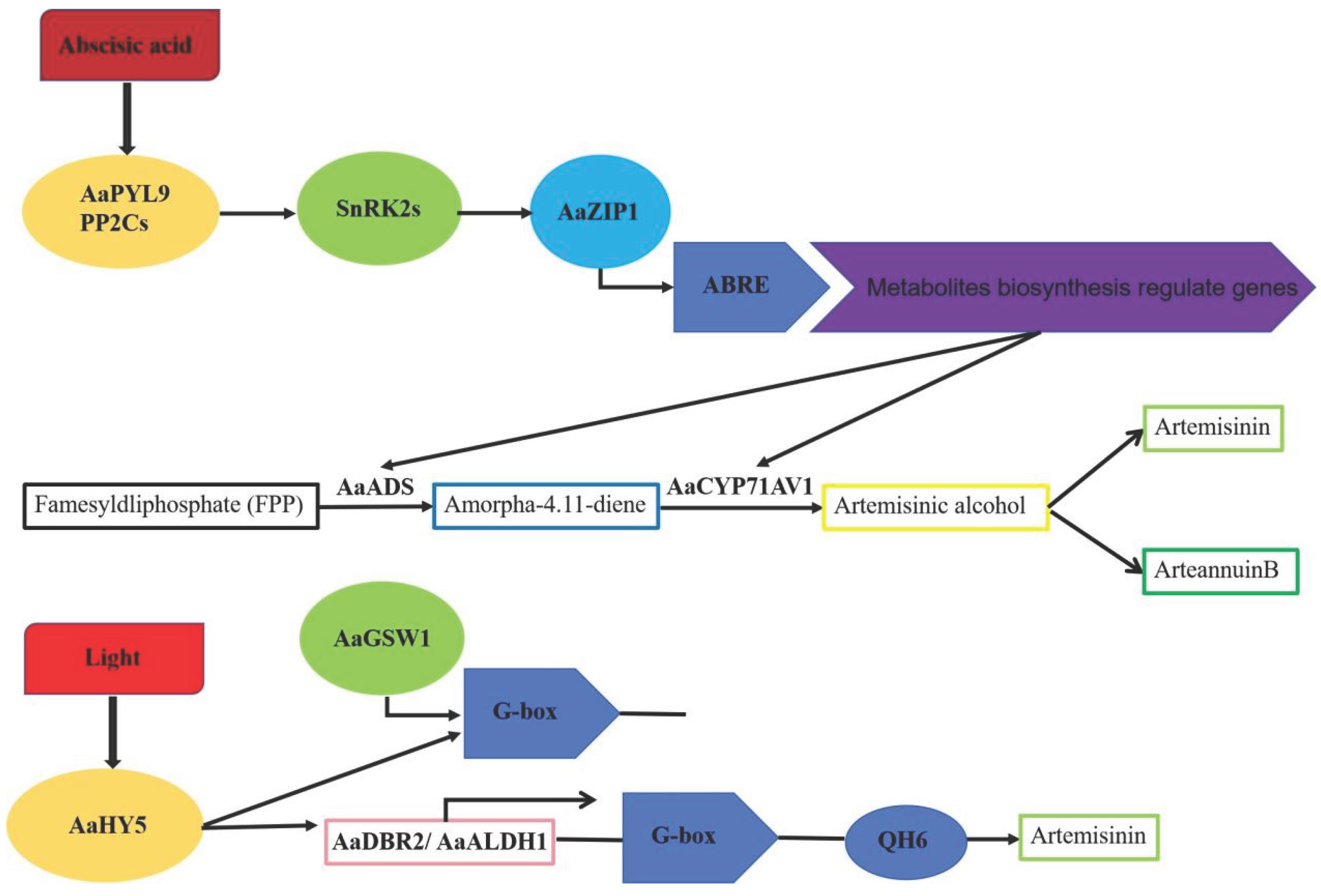
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



