Submitted:
20 August 2025
Posted:
21 August 2025
You are already at the latest version
Abstract
The questions of how life forms, whether life is an inevitable outcome, and how diverse its presentation could be remains some of the most profound in science. Investigations into the origin of life confront key issues such as uncovering key constraints and universal features of life, the plausibility of alternative biochemistries, and the transition from purely chemical systems to information-bearing, evolvable entities. Many of these issues can be associated with early cell formation and evolution. Thus, protocellular systems have emerged as a key focus of study. Here, the community can ask questions about physical constraints and the co-evolution of energy, matter, and information. The pursuit of these answers spans a wide range of disciplines, including geochemistry, statistical physics, systems and evolutionary biology, artificial life, synthetic biology, and information theory, and reflects the inherently interdisciplinary nature of origin-of-life research. This article surveys key theoretical frameworks and experimental approaches that have shaped our current understanding, while outlining the major unresolved challenges that continue to drive the field forward. It also summarizes and contextualizes the articles in this special issue that address these questions.
Keywords:
origins of life
; evolutionary transitions
; phase transitions
; information
; error thresholds
; protocells
1. Introduction
Is life a rare, perhaps improbable event in the universe? Is it the outcome of a sequence of fortunate accidents tightly constrained by planetary conditions and the chemistry of its environment? Or is it an almost inevitable outcome of complex geochemistry and deep time? As we seek to understand how life emerged on Earth, we also face broader questions about the likelihood of life elsewhere. Can the study of our own biosphere, together with growing knowledge of our solar system and the discovery of thousands of exoplanets, offer clues about the conditions necessary for life to arise and evolve? Could there be multiple pathways through which life could have taken hold, some potentially diverging dramatically from the biochemical framework with which we are familiar [1,2,3,4]? Is life necessarily tied to information processing, replication and the principle of natural selection, or could other organizing principles give rise to living systems [5,6]? Furthermore, can advances in synthetic biology and artificial life—whether created in vitro in the lab or modeled in silico through computational simulations—reveal the generative mechanisms that underpin life itself [7,8]? These questions lie at the heart of one of the most profound frontiers of science: understanding the origins, nature, and potential ubiquity of life in the cosmos.
The questions outlined above have been the subject of intense inquiry since the origin of life (OOL) became a recognized scientific problem. This transition, from philosophical speculation to empirical investigation, gained momentum during the mid-20th century and was often considered the golden age of OOL research. The early work of Oparin on prebiotic systems [9,10] and the groundbreaking experiments by Stanley Miller in 1953 on amino acid synthesis [11,12] and Joan Oró on nucleobase synthesis [13,14], laid the foundation for modern studies, and other scholars expanded on the initial concepts [15]. It was soon understood that the chemical scenario also required an understanding of the planetary context. Comets were soon proposed as candidates for the transport of extraterrestrial organic matter to Earth [16,17,18,19]. Their work marked a turning point: Life’s beginnings could now be explored through chemistry, biology, and planetary science, opening the door to rigorous hypotheses about how living systems could emerge from nonliving matter.
Building on the recognition that life’s origin poses a multifaceted scientific challenge, it becomes clear that any meaningful progress requires navigating a diverse landscape of theoretical approaches and empirical domains [20,21]. As summarized in Figure 1, this landscape spans multiple scales, from planetary and geochemical constraints to molecular evolution, combinatorial chemistry, and the emergence of metabolic organization. Research efforts increasingly draw on a broad toolkit, incorporating insights from extreme environments, exoplanet studies, prebiotic synthesis experiments, and synthetic biology.
These diverse lines of inquiry have fueled a growing synergy between empirical findings and theoretical or computational modeling. Such cross-disciplinary efforts are essential to identify the general conditions under which life-like behavior might arise, both on Earth and in other planetary settings [22,23,24]. In this regard, the planetary context provides a natural framework: from interpreting atmospheric biosignatures on exoplanets [25,26], to modeling chemical reaction networks in early Earth and interstellar environments [27,28,29,30].
Importantly, as in other domains of evolutionary biology, effective theories of the emergence of life must reconcile historical contingency with the restrictions imposed by physical laws [31]. The convergence of classical hypotheses with contemporary advances in systems chemistry, planetary science, and network theory has coalesced in the field of astrobiology, a domain uniquely positioned to explore life as a planetary and universal phenomenon [32]. Since terrestrial life emerged in our remote past, a direct experimental testing of such a deep-time event needs to be replaced by alternatives. As mentioned above, prebiotic chemistry offered the first approach to this problem, and in recent decades novel experimental implementations, including evolutionary dynamics and Darwinian selection [33,34], have been developed. Within the field of artificial life, multiple paths have been explored to investigate this concept, including artificial chemistries [35].
In 2021, the Santa Fe Institute hosted a workshop aimed at addressing fundamental questions about the possible pathways and actual processes underlying the origin of life. The meeting brought together researchers from a broad spectrum of disciplines, including thermodynamics, information theory, and synthetic biology, to explore the multifaceted nature of this problem. This Theme Issue presents a synthesis of key ideas that emerged from the discussions, highlighting the main challenges in understanding the origins of life. These range from theoretical and conceptual frameworks to planetary-scale environmental considerations, as well as experimental efforts to recreate life-like systems in the laboratory.
2. What Is Life? Is It Expected?
If we want to understand life, should we not first have an operational definition of it [3,6,8]? This seems like a reasonable question, since assessing the likelihood of life’s existence requires first knowing how to recognize it. However, despite our extensive knowledge of living systems, we are still far from reaching a consensus. With regard to this problem, the question arises as to how to distinguish between animate and inanimate systems when considering their change and evolution. Recently, methods for measuring the complexity of any molecule have been proposed1 [40,41], with interesting discussions about minerals [42,43], which have been a complicated case for OOL research for some time [44,45]. For living matter, there is general agreement that it would follow natural selection dynamics. We understand well how this theory works and how to define rigorous mathematical models [46]. Although the scope of evolution has at times been extended well beyond biology, it has raised contentious debates about its applicability. Cairns-Smith, for example, proposed that life on Earth originated through a process of natural selection that acts on inorganic crystals [47], while Pross has explored the possibility of applying Darwinian principles to inanimate physicochemical systems [48]. Although provocative, such views remain problematic, as they rely on substrates fundamentally distinct from living matter and lack the organizational features, such as heredity, variation, and regulated metabolism, that underpin Darwinian evolution in biological systems. As such, these approaches provide, at best, useful metaphors rather than plausible mechanisms for the actual origins of life.
The problem of defining life has deep historical roots, tracing back to ancient philosophy where thinkers like Aristotle sought to distinguish living from nonliving based on characteristics such as self-movement [49], growth, and reproduction. With the rise of modern science, especially after the advent of cell theory and Darwinian evolution, biological definitions emphasized structural and functional criteria, such as metabolism and natural selection. In the twentieth century, new perspectives emerged from cybernetics, systems theory, and information science, introducing concepts such as homeostasis [50], autopoiesis [51], and informational self-replication. Despite these advances, each disciplinary lens captured only part of the phenomenon, leading to a proliferation of definitions without consensus. The challenge has intensified in recent decades with developments in synthetic biology, artificial life, and astrobiology, which increasingly blur the boundaries of what qualifies as “alive” [52] and expose the limitations of traditional biological definitions [53]. In [54], the authors have examined the difficulties inherent in evaluating speculative hypotheses about the origin of life (what they call the Science of the Gaps), emphasizing how complexity and historical contingency create persistent gaps in both knowledge and conceptual frameworks. Along with the limitations imposed by current empirical approaches, metaphors and biased narrative scenarios have played an important role. By formalizing causal assumptions, they propose a more systematic approach to reasoning through these gaps, enabling a more rigorous evaluation of fragmentary and speculative origin-of-life models.
Scholars hold differing views on the inevitability of life. Some, like Christian de Duve, argue that life is a cosmic imperative, suggesting that, given the right conditions, life will emerge as a natural consequence of chemistry [55]. Others, such as Jacques Monod, emphasize the role of chance and contingency, viewing life as a highly improbable event in the universe [56]. Still others, including proponents of systems chemistry and self-organisation, propose that life arises from the inherent combinatorial dynamics of complex systems, making it statistically likely under certain constraints [57,58]. This ongoing debate reflects deeper philosophical divisions about whether life is an accident or an expected outcome of physical laws. An important component in this context is the role that the combinatorial potential of living objects plays, particularly at the molecular level. It is well known that the dynamics of large collections of objects capable of assembling or combining can easily drive a hyperbolic growth process [41,59,60,61,62]. How does this accelerated growth dynamics impact evolution and its predictability?
The combinatorial nature of evolution [63] has been argued to pose a fundamental challenge to the formulation of a predictive theory of evolutionary change (see also this problem within the context of cellular automata and computation [64,65,66,67,68]). This difficulty is made explicit through the concept of the adjacent possible. Briefly, the adjacent possible refers to the set of novel biological forms or functions that can emerge through small, incremental modifications of a system’s current state. Evolutionary innovations emerge from this constrained space of possibilities, and as new structures or functions are realized, the adjacent possible itself expands, enabling further exploration. According to Kauffman, this process is not algorithmically predictable: we cannot prestate the full repertoire of potential evolutionary trajectories or innovations, since the space of possibilities evolves in tandem with the system. Using the concept of the Adjacent Possible, Kauffman and Roli [69] propose that life is an expected consequence of the (first-order) transition from random reaction chemistry to autocatalytic chemical sets (see [70]) along with RNA sets displaying catalytic properties. These two sets would eventually cooperate to form some class of minimal (cellular) agent capable of template replication and coding.
3. Universal Constraints
A key focus for OOL research is identifying the aspects of life that can be counted on to be universal [3,6]. Much discussion has been given to the material aspects of life where debates about the centrality of carbon and a water solvent have occurred for decades [1,71,72]. The broader question here is how can we draw general inferences about origins of life, given the immense span of time since it occurred and the potentially contingent nature of the processes involved? This challenge is closely linked to the historical character of molecular evolution: did life on Earth—and possibly elsewhere—emerge through multiple distinct pathways? [3,4] Alternatively, as some studies propose, might there be universal properties of complex systems that constrain and shape the range of possible outcomes, or clear signatures of those outcomes [6,41]? Given that we possess only a single large-scale natural instance of life’s emergence, our biosphere, the lack of comparative data may seem to preclude meaningful inference.
One way to overcome this problem is to take lessons from physics, particularly cosmology. Cosmology exemplifies the power of physics-based science to probe the deep past through indirect but quantitatively robust evidence. Despite dealing with phenomena occurring billions of years ago, the standard cosmological model achieves predictive accuracy by anchoring itself in general relativity, quantum field theory, and thermodynamics. Observables such as the cosmic microwave background anisotropies, large-scale structure, and primordial nucleosynthesis abundances provide stringent constraints on early-universe physics, allowing us to infer conditions right after the Big Bang. This inferential framework parallels evolutionary biology, where Darwin’s theory of natural selection enables reconstruction of the tree of life from morphological and genetic data. Both cosmology and evolutionary theory transform present-day observations into historically coherent models, demonstrating how systems governed by contingent, path-dependent processes can still be understood through scientific analysis grounded in universal principles. Perhaps it should not be surprising to read from Leonard Susskind, a well-known cosmologist, that “modern cosmology really began with Darwin and Wallace” [73]. Similarly, the powerful views of cosmology have inspired others working in biology to build a Big Bang model for life origins and evolution [74].
One source of inspiration with high explanatory power is connected with the physics of phase transitions. This theory, a fundamental pillar of physics, explains how systems undergo abrupt, universal changes driven by small parameter shifts. Its core concepts, symmetry breaking and criticality, apply across scales, from materials to cosmology, revealing deep connections between seemingly unrelated phenomena [75,76,77]. One of the most striking aspects of the theory of phase transitions is that it derives powerful, often exact results from remarkably simple models, such as the Ising model or percolation lattices. Despite their minimal assumptions, these models capture the essential features of critical behaviour, revealing that the macroscopic properties near phase transitions depend not on microscopic details but on broad features like dimensionality and symmetries. This phenomenon, known as universality, explains why diverse systems, from magnets to fluids to the early universe, exhibit identical critical exponents and scaling laws, highlighting the deep, emergent order underlying complex phenomena.
As discussed in [70], the transition from non-living matter to living matter involves qualitative shifts in system organization, such as symmetry breaking, percolation, and bifurcation phenomena, that mirror phase transitions in physical systems. These transitions help explain how relatively simple molecular mixtures, under the right environmental and energetic conditions, could give rise to complex, self-sustaining, and evolvable chemical networks. Key processes in early evolution, including the origin of molecular chirality, the appearance of replicators, and the onset of cooperation, are framed as critical transitions driven by underlying symmetry changes or threshold effects [78]. This perspective complements traditional chemical and biochemical approaches by offering a theoretical framework based on physics that can describe the nonlinear nature of life’s emergence and the role of collective phenomena in enabling major transitions.
Another approach is to turn to physics itself, since life and the scaling laws it often presents can be rigorously derived as evolution under fundamental physical constraints [79]. For example, classic work on OOL made strong connections with thermodynamics [80,81]. Thermodynamics is fundamental to understanding biological complexity because living systems are open, far-from-equilibrium structures that maintain order by dissipating energy. The second law sets constraints on what biological systems can do, while non-equilibrium thermodynamics explains how they sustain gradients, perform work, and self-organize. From metabolism to molecular machines, thermodynamic principles reveal how life harnesses energy flows to construct and sustain complex adaptive structures. Within the context of OOL, prebiotic environments provided nonequilibrium conditions, such as thermal gradients, redox potentials, or chemical fluxes, that enabled the formation of self-organizing structures [81,82,83,84].
Within biology, particularly with respect to molecular systems, the thermodynamic picture must be consistent with the presence of thresholds [78]. This connection is made in [85], where the author presents an elegant approach to Darwinian dynamics of molecular replicators using a thermodynamic perspective. He shows that modern biomolecules, such as RNA, conform to fundamental bounds that connect fitness and thermodynamics. In particular, it generalizes the classic results by Eigen and Schuster on the error threshold [86], using the thermodynamic formalism. The bounds presented in this article are the type of constraint that should be upheld by life everywhere in the universe.
There is another way to search for potential universal principles by considering purely informational constraints. Information theory plays a central role in theoretical efforts to explain the origins of life, as it provides a formal framework for understanding how order, structure, and function can emerge and persist in physical systems far from equilibrium. At its core, life involves the storage, transmission, and transformation of information, whether in the replication of nucleic acids, the translation of genetic codes, or the regulation of metabolic networks. Shannon’s theory [87] enables researchers to quantify the information content of molecular sequences and to distinguish between randomness and functional complexity [88,89,90]. Moreover, concepts such as mutual information, entropy, and channel capacity have been used to study how early protocells might have maintained and transmitted adaptive information under noisy prebiotic conditions [91].
A remarkable feature of life as we know it is the universal nature of the information material. Every single living cell on our planet contains DNA as the substrate of heritability. Why? Why not a biosphere with multiple forms of molecular information? Perhaps one of the most celebrated universals in biology is the central dogma of molecular biology. First articulated by Francis Crick in 1958 [92,93], the central dogma was defined as a principle about the allowed flow of information in biological systems: once information has passed into a protein, it cannot flow back to nucleic acids. This formulation was not merely a description of the transcription–translation pathway but a broader statement about the asymmetry of information transfer in living systems. Within this framework, DNA serves as a stable repository of genetic instructions, which are transcribed into RNA and subsequently translated into proteins, thus linking genotype to phenotype. Later discoveries, including reverse transcription, RNA editing, and regulatory roles of noncoding RNAs, have refined, but not overturned, Crick’s central insight [94,95]. This raises deeper questions: Is the structure of information flows described by the central dogma a historical contingency of terrestrial biology, or does it reflect a universal constraint on how any living system must be organized? Theoretical work suggests that the latter may indeed be the case.
In a very elegant theoretical study, Takeuchi and Kaneko demonstrated how the fundamental asymmetries of the central dogma, namely, information flowing from genomes to enzymes and catalysis being performed only by enzymes, could have emerged spontaneously from evolutionary dynamics [96] (see also [97]). Using a mathematical model of protocells containing replicating molecules, the study demonstrates that a trade-off between acting as a catalyst and as a template results in a conflict between molecular and cellular selection. This conflict drives a symmetry breaking, causing molecules to differentiate into genomes (information carriers) and enzymes (catalysts). The process is reinforced by a feedback loop that involves reproductive value and selection strength across different levels. The findings suggest that the central dogma arises naturally from multilevel selection, especially when the molecular-level variation is high compared to the cellular-level variation. In a new paper, [98] the authors propose a generalization of the central dogma using the separation of information from function, which they connect to the broader idea of division of labor. They show that this separation has occurred multiple times throughout the history of life beyond the central dogma, such as with the specialisation of germline and somatic cells (a crucial step towards complex multicelularity).
The previous examples involved two important components of life: metabolism and information. A third ingredient in the definition of (cellular) life is the presence of compartments. Compartments have been shown to enhance life in a variety of ways, including speeding up chemical reactions through concentration and increasing evolutionary selection through increased individual fidelity and protection against destabilizing or cheating dynamics in complex reaction networks [6,99,100,101,102,103,104,105]. Physical space in modern cells has become a central role in understanding the physiology and biophysics of cells from the constraints of reaction rates, diffusion, packing limits, and macromolecular composition [106,107,108,109].For example, the smallest cells are nearly completely packed with biomolecules [108]. What consequences do these constraints have for the origin of cellular life? In [105] the authors consider a generalized interplay between function and information, in the context of encapsulation constraints. They show that the complexity of the biochemical cycle and the chemical and biophysical properties of molecules set limits on the possibility of encapsulation and connect this to known constraints on cells due to scaling relationships for energetic and molecular crowding [108,109,110,111].
4. Pathways Towards Life
On a large scale, dealing with planetary conditions for life, scientists have established the physical constraints [24], allowing us to estimate the likelihood of some Earth-like worlds [112,113,114] and establish a catalog of exoplanets within the habitable zone [115], i.e., where water can exist in a liquid state on the planet’s surface. Assuming that the right physical conditions are met, most of the origin-of-life hypotheses converge on a minimal set of physicochemical requirements [58]. Once again, a universal requirement is the presence of liquid water [116,117], which facilitates molecular motion, reaction kinetics, and stable hydrogen bonding. A suitable temperature range, typically between 0°C and 150°C, is also essential to preserve molecular structures [117] (see also [118]). In addition, life requires a diverse chemical inventory, particularly the availability of key biogenic elements, namely carbon, hydrogen, nitrogen, oxygen, phosphorus, and sulfur (CHNOPS) [119,120]. These are required to form the basis for essential biomolecules such as nucleic acids, proteins, and lipids [58].
The chemical and geochemical details associated with the origin of life on Earth have a long history with commitments to and debates about what conditions are essential [1,71,72,100]. Recently, a variety of specific environments have been proposed as candidates for the evolution of specific aspects of life. These include polymerisation of functional and informational polymers in wet/dry cycles [121] or atmospheric aerosols [122], and energy-harnessing membranes in hydrothermal vents [101]. In terms of what is possible, there have been many recent attempts to investigate alternative chemical systems on Earth or other planetary bodies [123,124] and to unravel the detailed history of the biochemical evolutionary history [125,126,127,128,129]. However, a persistent challenge remains: the necessary conditions for these processes often appear incompatible and are seldom found together in a single environment. This has limited the plausibility of models that attempt to explain the emergence of living systems from non-living matter within a singular, unified setting.
To address this, a broader perspective is emerging, one that frames the origin of life within the planetary-scale complexity of the Hadean Earth [130,131]. This view suggests that the diverse and dynamic environments of the early Earth collectively contributed to the emergence of life. Chemically rich microenvironments, though potentially uninhabitable to early life forms, may have provided critical building blocks. Minerals are thought to have served as catalytic surfaces and, at the same time, both local mixing and global transport processes, as well as weathering or geological activity, could have connected spatially separated chemical reactions, enabling a distributed network of prebiotic chemistry. Within this geochemical mosaic, molecular diversity combined with selective processes in specific locales probably drove the accumulation and refinement of complex organic molecules required for life [130].
How were the essential components for life listed above mobilized and concentrated in prebiotic environments on the early Earth? In [132], the authors identify planetary processes that probably enabled the availability of these elements in forms suitable for supporting key prebiotic reactions. In this context, terrestrial acidic hot springs and deep-sea hydrothermal vents emerge as particularly favorable settings due to their potential to drive abiotic nitrogen reduction, release reactive phosphorus compounds, and leach key transition metals from igneous rocks. The findings also have broader implications, suggesting that similar geochemical conditions on early Mars and other terrestrial planets could have supported independent pathways to life.
Jacques Monod said that "every living being is […] a fossil" [56], which means that the evolutionary past leaves a distinguishable trace within living entities. In some cases, the geochemical context is part of this trace. Can the analysis of cellular structures and cellular networks tell us something about early and alternative scenarios for the emergence of cellular life? An example is provided by the phylogenomic analysis of universal inorganic cellular components [133]. This study supports the conjecture that the first cells might have started in geothermal environments rich in phosphate and potassium. The crucial concept is that, because early protocells lacked ion pumps and impermeable membranes, their internal chemistry reflected the composition of their surroundings. The analysis suggests that life likely originated in anoxic, metal-enriched geothermal ponds, which have left a fossil trace in the internal composition of modern cells.
Similarly, the study of metabolic networks from an evolutionary perspective offers a highly valuable approach to identify origins [129,134,135,136,137]. The first cells emerged and evolved within the context of early Earth described above, giving rise to the Last Universal Common Ancestor (LUCA). Reconstructing this early metabolic network is key to understanding the transition from prebiotic chemistry to fully functional cellular life. By tracing conserved enzymatic cores and ancient metabolic pathways, we can uncover the robust biochemical logic and environmental constraints that guided early evolution [137,138]. This network serves as a molecular fossil, offering a window into how life first captured energy and processed information within a self-sustaining, organized system.
Are the paths towards life describable as linear chains of events? In [139] a novel way to think about how biosynthetic pathways originated and evolved is proposed, while [140] addresses the deep roots of metabolism before LUCA. Both contributions converge on a critical distinction in OOL research: the difference between the mere prebiotic availability of metabolic intermediates and the emergence of structured biosynthetic pathways. Although several compounds found in extant metabolism can indeed form abiotically under prebiotic conditions, this chemical convergence does not by itself imply the existence of energetically coupled, directional pathways. In contrast, biosynthetic pathways are defined by stepwise transformations of a single precursor, enabling energy transduction and regulatory control, characteristics absent in a purely chemical network lacking genetic and enzymatic constraints. Together, these works challenge the notion that modern metabolism could have arisen simply through the co-occurrence of metabolites, underscoring the need for mechanisms capable of stabilizing and propagating reaction sequences, such as genetic encoding and selective catalysis.
5. Beyond Terrestrial Life: Exobiology, Xenobiology, and Virtual Life
In addition to studying potential origins of “life as we know it” on Earth, there are strands of research that study "life as it could be". This includes alternative terrestrial life forms obtained using engineering living matter (through synthetic biology and xenobiology), possible extraterrestrial life forms (exobiology or astrobiology) and life forms that exist in virtual environments (in silico life). This can exploit the possibilities of building, simulating, or synthesizing living or lifelike systems that are very different from terrestrial life.
Life elsewhere, extraterrestrial life, could be so different that its underlying principles have nothing to do with our understanding of living matter. This is the problem of "life as we don’t know it" [4,141,142,143]: can we articulate a universal approach that could safely explain the properties of unknown life? These are important questions, particularly within the growing field of astrobiology [3,32,144,145,146,147]. The potential diversity of life forms that could have emerged and evolved elsewhere raises two relevant questions. The first is how we can study potential scenarios for the origins of life using experiments. The second question is: What kinds of scales, case studies, model systems, and theoretical approaches can capture the universal properties of the problem?
Returning to Earth, synthetic biology is an engineering discipline that constructs living systems based on modified underlying principles of terrestrial life. However, designs sometimes depart from their technological and biological counterparts [148]. Xenobiology alters the genetic code, expanding the amino acid palette used to construct proteins and introducing novel nucleotides beyond the standard ACGT sequence in DNA, thereby increasing the possibilities for space exploration. In [149], a perspective review of one approach to achieving these xenobiological ends is presented. In an in vitro approach, cell-free expression systems (CFS) allow the creation and investigation of biological reactions outside living cells, synthetic pathways, alternative metabolisms, different information replication processes and compartmentalization processes [150]. These “roads not taken” can help provide clues about the deterministic (necessity) versus random (chance) and origins of their natural counterparts.
Systems biology techniques can provide valuable insights into the origins of life by enabling researchers to construct simplified, controllable models of early living systems. By reconstructing minimal cells, or protocells, from the bottom up, scientists can test hypotheses about how life-like behavior could emerge from nonliving components. These experimental platforms help identify which molecular characteristics and organisational principles are essential for life, explore plausible prebiotic pathways, and reveal the conditions under which key transitions, such as replication, metabolism, and compartmentalisation [151,152], can occur. In essence, synthetic biology turns the question of the origin of life into an experimentally tractable problem and allows the incorporation of different scales, from cells to ecosystems [153,154].
Demonstrating what a rich field systems biology in general, and protocells in particular, provide for investigating mechanisms relevant to OOL questions, this special issue includes three articles devoted to different aspects of protocellular research. Das et al [155] investigate the dynamics of an experimental system that incorporates ecological interactions among protocell populations. They use heterogeneous populations of protocells that have the simpler membranes expected during OOL. Such membranes can spontaneously self-assemble, and each can have a different chemical makeup. In a population of protocells with different membrane compositions, cells can interact differentially, both synergistically and antagonistically, leading to complex population dynamics. The authors show that heterogeneous systems exhibit emergent growth properties compared to homogeneous systems, suggesting how prebiotic and early biotic diversity could be beneficial. On the theoretical side, these diverse populations and their importance in OOL are studied in [156] using a simulation platform as a way to investigate how metabolic regulation can be used to enable simple adaptive or learning behaviors, without the need for genetics. By extending consumer-resource models to include stochastic evolution, individual-level novelty, and short-term memory, the study shows that simple, pre-genetic forms of adaptation and learning can significantly affect protocell survival during their lifetimes. The findings highlight the importance of metabolic regulation and agent-like behavior in shaping the evolutionary dynamics of early life.
Thomsen et al. [157] took another simulation-based protocell approach to study the molecular mechanisms directly related to metabolism, with results compared to values of wetlab-derived parameters. The simulation enables a detailed examination of the combined fitness of different processes that have opposing requirements. The use of simulation in OOL research has its advantages: experiments can typically be run more readily, and the experimenter has ready access to all the state parameter values at each time step, allowing in-depth analyses. It also has disadvantages, particularly the need to implement adequate underlying physical and chemical mechanisms so that a simulated evolutionary process has sufficiently diverse and powerful underlying properties to exploit, without hard-coding in desired or expected outcomes. Wet-lab and simulation experiments can provide valuable complementary approaches for investigating complex behaviours and emergent properties.
The field of Artificial Life goes beyond synthetic biology’s use of standard or modified biological components to study alternative biochemical systems, and inorganic systems to investigate various fundamental processes of life. Such studies are relevant both for exobiology (life on other planets that exploit alternative chemical pathways) and to prebiotic processes relevant to origins of life (for example, crystals and clays [44,47]).
A step further away from terrestrial-type life is to move from the physical (material) to the computational (abstract) domain, into the realm of virtual life. Whether virtual life (in contrast to a mere simulation of material life) is possible depends at least in part on the chosen definition of life. In this regard, however, it does not differ from the possibility of any forms of life that deviate sufficiently from the common instantiation, for example, the possibility of inorganic material life. In [158], Stepney provides a requirement-based definition of life that does not exclude the possibility of virtual life and discusses how a virtual system might fulfill these requirements. Since these requirements are necessarily abstracted away from the specifics of terrestrial life, they provide a different lens on possible origins and on the possibility of partial life that fulfills some but not all of the requirements (see also [159]). For example, artificial life need not originate through an evolutionary process (unless one includes the evolution of the artificer).
6. Discussion
Discovering life elsewhere in the universe or creating it in the laboratory would constitute a profound scientific breakthrough, reshaping our understanding of biology, evolution, and the uniqueness of life on Earth. In the search for extraterrestrial life, two promising avenues are actively being pursued. The first involves exploring our solar system, particularly environments such as Mars, Europa, and Enceladus, where liquid water and geochemical activity can support microbial ecosystems. The second focuses on detecting biosignatures from exoplanets, using next-generation telescopes to identify atmospheric gases or surface features that may indicate biological activity.
By examining the early environments of Earth, the planetary processes, and the biochemical systems, and comparing them with what we know about other planets in our solar system and beyond, we can begin to understand the prerequisites for life. Recent discoveries of exoplanets, some within habitable zones with intriguing chemistry [160,161,162], further challenge us to consider whether life is an inevitable outcome of planetary evolution or an astonishing cosmic fluke. It also pushes us to have better generative theories and experiments to rule in or out particular measurements, to guide and propose new measurements, and to serve as a test bed for exploring ideas.
On the experimental side, efforts to create life in the lab are converging on the challenge of constructing a minimal, self-sustaining, and self-replicating protocellular system. This endeavour is inherently multidisciplinary, involving prebiotic chemistry, synthetic biology, systems chemistry, and biophysics, as well as philosophy [163]. Multiple potential pathways are under active investigation, each offering complementary insights into how nonliving matter might transition into life [33,164,165,166]. As discussed in previous sections, these approaches are informed by diverse sources of evidence—geological, biochemical, and theoretical—and their integration may ultimately illuminate viable routes to the emergence of life.
What is life then? This question illustrates how hard the theoretical side of the problem is, and typically raises multiple answers [53,167,168]. One way to gain traction on this problem is to define the entire space of possibilities within a category: the space of possible living systems. We have attempted to do it by considering cellular replicators as restriction to an important class of objects that have some degree of embodiment (usually a closed membrane), can store and/or manipulate information, and have a metabolism. In Figure 2, we present our morphospace of replicators, defined qualitatively. The location of each case study along the three axes (spatial, informational, and metabolic complexity) is relative to other systems. The distance between each example does not define a metric space. On the right, the whole space is depicted, spanning many orders of magnitude from simple autocatalytic reactions (such as the Formose reaction [169], lower left corner) to modern cells in the upper right corner. The smaller cube indicates the subset of systems that involve “synthetic” designs, and the details of this space are shown in the left cube.
The space of possible replicators serves to define what a replicator is. Any possible embodied system that can exhibit autonomous replication of some kind has a place in the space. There is no single answer, and different kinds of complex replicators coexist within the boundaries of this space. This reflects intrinsic differences in how replication occurs, ranging from the genetically controlled mechanisms present in modern cells to synthetic systems that rely on the coupling between metabolism and membrane deformation.
As our understanding of potential scenarios for the emergence of life advances, we progressively illuminate different regions of the space of possibilities. Current bottom-up approaches to the origins-of-life (OOL) problem have yielded partial, yet highly encouraging, insights. In particular, our knowledge of how diverse interactions between soft matter, protometabolic networks, and information-carrying molecules operate is improving rapidly. It is also increasingly clear that explaining the rise of the first cellular replicators will require integrating both evolutionary dynamics and self-organization into theoretical and engineering frameworks [184,185,186,187,188]. In this context, we anticipate that major advances in protocell research may depend on combining evolutionary dynamics with Darwinian selection, an idea strongly supported by the successful evolution of lipid droplets in chemorobotic platforms [33]. The use of morphospaces has also proved valuable since their inception for exploring the limits of the possible. Observed systems tend to cluster along an imaginary axis connecting two opposed vertices, from minimal to maximal cellular complexity. This pattern may suggest that, despite their different origins, the three key components evolve in a synergistic manner. In contrast, empty regions of the morphospace, such as the one shown in Figure 2, point to domains of possibilities not occupied by any known system. Why is this so? In some cases, physical or chemical constraints may forbid these regions from being realized. In others, evolutionary processes may have never reached them, perhaps due to historical contingencies or environmental limitations.
We close with more questions than answers, but that is the nature of science. Whatever the answers are, they will likely emerge at the crossroads of multiple disciplines and may bring unexpected surprises.
Acknowledgments
The authors thank the NSF RCN for Exploration of Life’s Origins (NSF Grant No. 1745355) for supporting the working group where all this started to take shape. RS thanks the members of the Complex Systems Lab for useful discussions and to the Departament de Recerca i Universitats de la Generalitat de Catalunya for its support under grant 2021 SGR 0075. CPK thanks the Charities Aid Foundation of Canada (CAF) for supporting this work.
References
- Benner, S.A. Defining life. Astrobiology 2010, 10, 1021–1030. [Google Scholar] [CrossRef]
- Woese, C.R. A new biology for a new century. Microbiology and molecular biology reviews 2004, 68, 173–186. [Google Scholar] [CrossRef]
- Kempes, C.P.; Krakauer, D.C. The multiple paths to multiple life. Journal of Molecular Evolution 2021, 89, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Grefenstette, N.; Chou, L.; Colón-Santos, S.; Fisher, T.M.; Mierzejewski, V.; Nural, C.; Sinhadc, P.; Vidaurri, M.; Vincent, L.; Weng, M.M. Life as We Don’t Know It. Astrobiology 2024, 24, S–186. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.I.; Davies, P.C. The algorithmic origins of life. Journal of the Royal Society Interface 2013, 10, 20120869. [Google Scholar] [CrossRef]
- Solé, R.; Kempes, C.P.; Corominas-Murtra, B.; De Domenico, M.; Kolchinsky, A.; Lachmann, M.; Libby, E.; Saavedra, S.; Smith, E.; Wolpert, D. Fundamental constraints to the logic of living systems. Interface Focus 2024, 14, 20240010. [Google Scholar] [CrossRef]
- Langton, C.G. Artificial life: An overview; MIT press, 1997.
- Szostak, J.W. Attempts to define life do not help to understand the origin of life. Journal of Biomolecular Structure and Dynamics 2012, 29, 599–600. [Google Scholar] [CrossRef]
- Oparin, A. Proiskhozhdenie zhizni; Moskovskii Rabochii, 1924. (First presentation of Oparin’s heterotrophic origin-of-life hypothesis).
- Oparin, A. The Origin of Life; Macmillan, 1938. (English edition of Vozniknovenie zhizni na zemle, first Russian ed. 1936).
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef]
- Bada, J.L.; Lazcano, A. Prebiotic soup–revisiting the miller experiment. Science 2003, 300, 745–746. [Google Scholar] [CrossRef]
- Oró, J. Synthesis of adenine from ammonium cyanide. Biochemical and biophysical research communications 1960, 2, 407–412. [Google Scholar] [CrossRef]
- Oró, J. Mechanism of synthesis of adenine from hydrogen cyanide under possible primitive Earth conditions. Nature 1961, 191, 1193–1194. [Google Scholar] [CrossRef]
- Schopf, J.W. Pioneers of Origin of Life Studies—Darwin, Oparin, Haldane, Miller, Oró—And the Oldest Known Records of Life. Life 2024, 14, 1345. [Google Scholar] [CrossRef] [PubMed]
- Oró, J. Comets and the formation of biochemical compounds on the primitive Earth. Nature 1961, 190, 389–390. [Google Scholar] [CrossRef]
- Oro, J.; Mills, T.; Lazcano, A. Comets and the formation of biochemical compounds on the primitive Earth–A review. Origins of Life and Evolution of the Biosphere 1991, 21, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Sagan, C. The search for extraterrestrial life. Scientific American 1994, 271, 92–99. [Google Scholar] [CrossRef]
- Chyba, C.F.; Thomas, P.J.; Brookshaw, L.; Sagan, C. Cometary delivery of organic molecules to the early Earth. Science 1990, 249, 366–373. [Google Scholar] [CrossRef]
- Walker, S.I. Origins of life: a problem for physics, a key issues review. Reports on Progress in Physics 2017, 80, 092601. [Google Scholar] [CrossRef]
- Goldenfeld, N.; Woese, C. Life is physics: evolution as a collective phenomenon far from equilibrium. Annual Review of Condensed Matter Physics 2011, 2, 375–399. [Google Scholar] [CrossRef]
- Seager, S. Exoplanet habitability. Science 2013, 340, 577–581. [Google Scholar] [CrossRef]
- Takeuchi, N.; Hogeweg, P.; Kaneko, K. Conceptualizing the origin of life in terms of evolution. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 2017, 375, 20160346. [Google Scholar] [CrossRef]
- Lingam, M.; Loeb, A. Physical constraints on the likelihood of life on exoplanets. International Journal of Astrobiology 2018, 17, 116–126. [Google Scholar] [CrossRef]
- Schwieterman, E.W.; Kiang, N.Y.; Parenteau, M.N.; Harman, C.E.; DasSarma, S.; Fisher, T.M.; Arney, G.N.; Hartnett, H.E.; Reinhard, C.T.; Olson, S.L.; et al. Exoplanet biosignatures: a review of remotely detectable signs of life. Astrobiology 2018, 18, 663–708. [Google Scholar] [CrossRef]
- Walker, S.I.; Bains, W.; Cronin, L.; DasSarma, S.; Danielache, S.; Domagal-Goldman, S.; Kacar, B.; Kiang, N.Y.; Lenardic, A.; Reinhard, C.T.; et al. Exoplanet biosignatures: future directions. Astrobiology 2018, 18, 779–824. [Google Scholar] [CrossRef]
- Solé, R.V.; Munteanu, A. The large-scale organization of chemical reaction networks in astrophysics. Europhysics Letters 2004, 68, 170. [Google Scholar] [CrossRef]
- Garcia-Sanchez, M.; Jimenez-Serra, I.; Puente-Sanchez, F.; Aguirre, J. The emergence of interstellar molecular complexity explained by interacting networks. Proceedings of the National Academy of Sciences 2022, 119, e2119734119. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ruz, M.; Jiménez-Serra, I.; Aguirre, J. A theoretical approach to the complex chemical evolution of phosphorus in the interstellar medium. The Astrophysical Journal 2023, 956, 47. [Google Scholar] [CrossRef]
- Fisher, T.; Janin, E.; Walker, S. A Complex Systems Approach to Exoplanet Atmospheric Chemistry: New Prospects for Ruling Out the Possibility of Alien Life-As-We-Know-It. arXiv 2023. arXiv 2025, arXiv:2310.05359[astro-ph.EP], 2023. [Google Scholar]
- Smith, T.F.; Morowitz, H.J. Between history and physics. Journal of Molecular Evolution 1982, 18, 265–282. [Google Scholar] [CrossRef]
- Chyba, C.F.; Hand, K.P. Astrobiology: the study of the living universe. Annual Review of Astronomy and Astrophysics 2005, 43, 31–74. [Google Scholar] [CrossRef]
- Gutierrez, J.M.P.; Hinkley, T.; Taylor, J.W.; Yanev, K.; Cronin, L. Evolution of oil droplets in a chemorobotic platform. Nature Communications 2014, 5, 5571. [Google Scholar] [CrossRef]
- Points, L.J.; Taylor, J.W.; Grizou, J.; Donkers, K.; Cronin, L. Artificial intelligence exploration of unstable protocells leads to predictable properties and discovery of collective behavior. Proceedings of the National Academy of Sciences 2018, 115, 885–890. [Google Scholar] [CrossRef]
- Banzhaf, W.; Yamamoto, L. Artificial chemistries; MIT Press, 2015.
- Jaeger, J. Assembly theory: What it does and what it does not do. Journal of Molecular Evolution 2024, 92, 87–92. [Google Scholar] [CrossRef]
- Abrahão, F.S.; Hernández-Orozco, S.; Kiani, N.A.; Tegnér, J.; Zenil, H. Assembly theory is an approximation to algorithmic complexity based on LZ compression that does not explain selection or evolution. PLOS Complex Systems 2024, 1, e0000014. [Google Scholar] [CrossRef]
- Kempes, C.P.; Lachmann, M.; Iannaccone, A.; Fricke, G.M.; Chowdhury, M.R.; Walker, S.I.; Cronin, L. Assembly theory and its relationship with computational complexity. arXiv, 2024; arXiv:2406.12176. [Google Scholar]
- Lynch, M. Complexity myths and the misappropriation of evolutionary theory. Proceedings of the National Academy of Sciences 2025, 122, e2425772122. [Google Scholar] [CrossRef]
- Marshall, S.M.; Mathis, C.; Carrick, E.; Keenan, G.; Cooper, G.J.; Graham, H.; Craven, M.; Gromski, P.S.; Moore, D.G.; Walker, S.I.; et al. Identifying molecules as biosignatures with assembly theory and mass spectrometry. Nature communications 2021, 12, 3033. [Google Scholar] [CrossRef]
- Sharma, A.; Czégel, D.; Lachmann, M.; Kempes, C.P.; Walker, S.I.; Cronin, L. Assembly theory explains and quantifies selection and evolution. Nature 2023, 622, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Hazen, R.M.; Burns, P.C.; Cleaves, H.J.; Downs, R.T.; Krivovichev, S.V.; Wong, M.L. Molecular assembly indices of mineral heteropolyanions: some abiotic molecules are as complex as large biomolecules. Journal of the Royal Society Interface 2024, 21, 20230632. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.I.; Mathis, C.; Marshall, S.; Cronin, L. Experimentally measured assembly indices are required to determine the threshold for life. Journal of the Royal Society Interface 2024, 21, 20240367. [Google Scholar] [CrossRef]
- Cairns-Smith, A.G. Genetic Takeover and the Mineral Origins of Life; Cambridge University Press, 1982.
- Cleland, C.E. Epistemological issues in the study of microbial life: alternative terran biospheres? Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences 2007, 38, 847–861. [Google Scholar] [CrossRef] [PubMed]
- Schuster, P. The mathematics of Darwin’s theory of evolution: 1859 and 150 years later. In The Mathematics of Darwin’s Legacy; Springer, 2011; pp. 27–66.
- Cairns-Smith, A.G. The origin of life and the nature of the primitive gene. Journal of Theoretical Biology 1966, 10, 53–88. [Google Scholar] [CrossRef]
- Pross, A. Toward a general theory of evolution: Extending Darwinian theory to inanimate matter. Journal of Systems Chemistry 2011, 2, 1. [Google Scholar] [CrossRef]
- Byers, S. Life as “self-motion”: Descartes and “the Aristotelians” on the soul as the life of the body. Review of Metaphysics 2006, 59, 723–755. [Google Scholar]
- Cannon, W.B. Organization for physiological homeostasis. Physiological Reviews 1929, 9, 399–431. [Google Scholar] [CrossRef]
- Maturana, H.R.; Varela, F.J. Autopoeisis and Cognition: the realization of the living; D. Reidel, 1980.
- Lachmann, M.; Walker, S. Life!=alive. Aeon 2019. [Google Scholar]
- Bender, R.; Kofman, K.; y Arcas, B.A.; Levin, M. What Lives? A meta-analysis of diverse opinions on the definition of life. arXiv 2025, arXiv:2505.15849. [Google Scholar] [CrossRef]
- Smith, E.; Kubica, A. Science of the Gaps. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- de Duve, C. Life as a cosmic imperative? Philosophical Transactions of the Royal Society A 2011, 369, 620–623. [Google Scholar] [CrossRef]
- Monod, J. Chance and Necessity; Collins, 1971.
- Hazen, R.M. Chance, necessity and the origins of life: a physical sciences perspective. Philosophical Transactions of the Royal Society A 2017, 375, 20160353. [Google Scholar] [CrossRef]
- Deamer, D. First life: Discovering the connections between stars, cells, and how life began; Univ of California Press, 2011.
- Johansen, A.; Sornette, D. Finite-time singularity in the dynamics of the world population and economic indices. Physica A: Statistical Mechanics and its Applications 2001, 294, 465–502. [Google Scholar] [CrossRef]
- Hanel, R.; Kauffman, S.A.; Thurner, S. Phase transitions in random catalytic networks. Physical Review E 2005, 72, 036117. [Google Scholar] [CrossRef]
- Youn, H.; Bettencourt, L.M.A.; Strumsky, D.; Lobo, J. Invention as a combinatorial process: evidence from U.S. patents. Journal of the Royal Society Interface 2015, 12, 20150272. [Google Scholar] [CrossRef]
- Sole, R.; Amor, D.R.; Valverde, S. On singularities and black holes in combination-driven models of technological innovation networks. Plos one 2016, 11, e0146180. [Google Scholar] [CrossRef]
- Kauffman, S.A. Investigations; Oxford University Press: New York, 2000. [Google Scholar]
- Wolfram, S. Nature 1984, 311, 419–424. [CrossRef]
- Wolfram, S. Physical Review Letters 1985, 54, 735–738. [CrossRef]
- Wolfram, S. A New Kind of Science; Wolfram Media: Champaign, IL, 2002. [Google Scholar]
- Ilachinski, A. Cellular Automata: A Discrete Universe; World Scientific: Singapore, 2001. [Google Scholar]
- Israeli, N.; Goldenfeld, N. Computational irreducibility and the predictability of complex physical systems. Physical review letters 2004, 92, 074105. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, S.A.; Roli, A. Is the emergence of life and of agency expected? Philosophical Transactions of the Royal Society B, 2025; This issue. [Google Scholar]
- Solé, R.; de Domenico’, M. Phase transitions in the origins of life. Philosophical Transactions of the Royal Society B 2025, This issue, 20160353. [Google Scholar]
- Pace, N.R. The universal nature of biochemistry. Proceedings of the National Academy of Sciences 2001, 98, 805–808. [Google Scholar] [CrossRef]
- Bains, W. Many chemistries could be used to build living systems. Astrobiology 2004, 4, 137–167. [Google Scholar] [CrossRef] [PubMed]
- Susskind, L. The cosmic landscape: String theory and the illusion of intelligent design; Back Bay Books, 2008.
- Koonin, E.V. The Biological Big Bang model for the major transitions in evolution. Biology Direct 2007, 2, 21. [Google Scholar] [CrossRef]
- Stanley, H.E. Introduction to Phase Transitions and Critical Phenomena; Oxford University Press, 1971.
- Goldenfeld, N. Lectures on Phase Transitions and the Renormalization Group; Addison-Wesley: Reading, MA, 1992. [Google Scholar]
- Solé, R. Phase transitions; Princeton University Press, 2011.
- Jeancolas, C.; Malaterre, C.; Nghe, P. Thresholds in origin of life scenarios. Iscience 2020, 23. [Google Scholar] [CrossRef] [PubMed]
- Kempes, C.P.; Koehl, M.; West, G.B. The scales that limit: the physical boundaries of evolution. Frontiers in Ecology and Evolution 2019, 7, 242. [Google Scholar] [CrossRef]
- Schrödinger, E. What is Life? The Physical Aspect of the Living Cell; Cambridge University Press: Cambridge, UK, 1944. [Google Scholar]
- Morowitz, H.J. Energy Flow in Biology: Biological Organization as a Problem in Thermal Physics; Academic Press: New York, 1968. [Google Scholar]
- Morowitz, H.; Smith, E. Energy flow and the organization of life. Complexity 2007, 13, 51–59. [Google Scholar] [CrossRef]
- Smith, E. Thermodynamics of natural selection I: Energy flow and the limits on organization. Journal of theoretical biology 2008, 252, 185–197. [Google Scholar] [CrossRef]
- Smith, E. Thermodynamics of natural selection II: Chemical Carnot cycles. Journal of theoretical biology 2008, 252, 198–212. [Google Scholar] [CrossRef]
- Kolchinsky, A. Thermodynamics of Darwinian selection in molecular replicators. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Eigen, M.; Gardiner, W.; Schuster, P.; Winkler-Oswatitsch, R. The origin of genetic information. Scientific American 1981, 244, 88–119. [Google Scholar] [CrossRef] [PubMed]
- Ash, R.B. Information theory; Courier Corporation, 2012.
- Küppers, B.O. Information and the Origin of Life; Mit Press, 1990.
- Adami, C. Information theory in molecular biology. Physics of Life Reviews 2004, 1, 3–22. [Google Scholar] [CrossRef]
- Walker, S.I.; Davies, P.C.; Ellis, G.F. The informational architecture of the cell: a systems view. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 2017, 375, 20160392. [Google Scholar]
- Farnsworth, K.D.; Ellis, G.F.; Jaeger, J. Living through downward causation: from molecules to ecosystems. Interface Focus 2013, 3, 20130062. [Google Scholar]
- Crick, F.H. On protein synthesis. Symposia of the Society for Experimental Biology 1958, 12, 138–163. [Google Scholar]
- Crick, F. Central dogma of molecular biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef]
- Thieffry, D.; Sarkar, S. Forty years under the central dogma. Trends in biochemical sciences 1998, 23, 312–316. [Google Scholar] [CrossRef]
- Ille, A.M.; Lamont, H.; Mathews, M.B. The Central Dogma revisited: Insights from protein synthesis, CRISPR, and beyond. Wiley Interdisciplinary Reviews: RNA 2022, 13, e1718. [Google Scholar] [CrossRef]
- Takeuchi, N.; Kaneko, K. The origin of the central dogma through conflicting multilevel selection. Proceedings of the Royal Society B 2019, 286, 20191359. [Google Scholar] [CrossRef]
- Takeuchi, N.; Hogeweg, P.; Kaneko, K. The origin of a primordial genome through spontaneous symmetry breaking. Nature communications 2017, 8, 250. [Google Scholar] [CrossRef]
- Takeuchi, N.; Kunihiko, K. Generalising the Central Dogma as a cross-hierarchical principle of biology. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Plum, A.M.; Kempes, C.P.; Peng, Z.; Baum, D.A. Spatial structure supports diversity in prebiotic autocatalytic chemical ecosystems. npj Complexity 2025, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.; Morowitz, H.J. The Origin and Nature of Life on Earth: The Emergence of the Fourth Geosphere; Cambridge University Press, 2016.
- Lane, N.; Martin, W.F. The origin of membrane bioenergetics. Cell 2012, 151, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Hogeweg, P. Multilevel selection in models of prebiotic evolution II: a direct comparison of compartmentalization and spatial self-organization. PLoS computational biology 2009, 5, e1000542. [Google Scholar] [CrossRef]
- Saha, R.; Pohorille, A.; Chen, I.A. Molecular crowding and early evolution. Origins of Life and Evolution of Biospheres 2014, 44, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Wächtershäuser, G. Before enzymes and templates: theory of surface metabolism. Microbiological reviews 1988, 52, 452–484. [Google Scholar] [CrossRef]
- Christopher, P. Kempes, D.A.; Mathis, C. How hard is it to encapsulate life? The general constraints on encapsulation. Philosophical Transactions of the Royal Society B.
- Rivas, G.; Ferrone, F.; Herzfeld, J. Life in a crowded world: Workshop on the biological implications of macromolecular crowding. EMBO reports 2004, 5, 23–27. [Google Scholar] [CrossRef]
- Zhou, H.X.; Rivas, G.; Minton, A.P. Macromolecular crowding and confinement: biochemical, biophysical, and potential physiological consequences. Annu. Rev. Biophys. 2008, 37, 375–397. [Google Scholar] [CrossRef]
- Kempes, C.P.; Wang, L.; Amend, J.P.; Doyle, J.; Hoehler, T. Evolutionary tradeoffs in cellular composition across diverse bacteria. The ISME journal 2016, 10, 2145–2157. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Kempes, C.P. Metabolic scaling in small life forms. arXiv 2023, arXiv:2403.00001. [Google Scholar] [CrossRef]
- Kempes, C.P.; Dutkiewicz, S.; Follows, M.J. Growth, metabolic partitioning, and the size of microorganisms. Proceedings of the National Academy of Sciences 2012, 109, 495–500. [Google Scholar] [CrossRef]
- Kempes, C.P.; van Bodegom, P.M.; Wolpert, D.; Libby, E.; Amend, J.; Hoehler, T. Drivers of bacterial maintenance and minimal energy requirements. Frontiers in microbiology 2017, 8, 31. [Google Scholar] [CrossRef]
- Anglada-Escudé, G.; Amado, P.J.; Barnes, J.; Berdiñas, Z.M.; Butler, R.P.; Coleman, G.A.; de La Cueva, I.; Dreizler, S.; Endl, M.; Giesers, B.; et al. A terrestrial planet candidate in a temperate orbit around Proxima Centauri. nature 2016, 536, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Jin, M.; Lingam, M.; Airapetian, V.S.; Ma, Y.; van der Holst, B. Atmospheric escape from the TRAPPIST-1 planets and implications for habitability. Proceedings of the National Academy of Sciences 2018, 115, 260–265. [Google Scholar] [CrossRef]
- Madhusudhan, N. Exoplanetary atmospheres: key insights, challenges, and prospects. Annual Review of Astronomy and Astrophysics 2019, 57, 617–663. [Google Scholar] [CrossRef]
- Hill, M.L.; Bott, K.; Dalba, P.A.; Fetherolf, T.; Kane, S.R.; Kopparapu, R.; Li, Z.; Ostberg, C. A catalog of habitable zone exoplanets. The Astronomical Journal 2023, 165, 34. [Google Scholar] [CrossRef]
- Ball, P. Life’s matrix: a biography of water; Univ of California Press, 2001.
- Westall, F.; Brack, A. The importance of water for life. Space Science Reviews 2018, 214, 50. [Google Scholar] [CrossRef]
- do Nascimento Vieira, A.; Kleinermanns, K.; Martin, W.F.; Preiner, M. The ambivalent role of water at the origins of life. FEBS letters 2020, 594, 2717–2733. [Google Scholar] [CrossRef]
- Williams, R.J. The chemical elements of life. J. Chem. Soc. Dalton Trans 1991, 539–546. [Google Scholar] [CrossRef]
- Da Silva, J.F.; Williams, R.J.P. The biological chemistry of the elements: the inorganic chemistry of life; Oxford University Press, 2001.
- Damer, B.; Deamer, D. The hot spring hypothesis for an origin of life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef]
- Dobson, C.M.; Ellison, G.B.; Tuck, A.F.; Vaida, V. Atmospheric aerosols as prebiotic chemical reactors. Proceedings of the National Academy of Sciences 2000, 97, 11864–11868. [Google Scholar] [CrossRef]
- Goldford, J.E.; Hartman, H.; Smith, T.F.; Segrè, D. Remnants of an ancient metabolism without phosphate. Cell 2017, 168, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.B.; Drew, A.; Malloy, J.F.; Walker, S.I. Seeding biochemistry on other worlds: Enceladus as a case study. Astrobiology 2021, 21, 177–190. [Google Scholar] [CrossRef]
- Petrov, A.S.; Bernier, C.R.; Hsiao, C.; Norris, A.M.; Kovacs, N.A.; Waterbury, C.C.; Stepanov, V.G.; Harvey, S.C.; Fox, G.E.; Wartell, R.M.; et al. Evolution of the ribosome at atomic resolution. Proceedings of the National Academy of Sciences 2014, 111, 10251–10256. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.S.; Gulen, B.; Norris, A.M.; Kovacs, N.A.; Bernier, C.R.; Lanier, K.A.; Fox, G.E.; Harvey, S.C.; Wartell, R.M.; Hud, N.V.; et al. History of the ribosome and the origin of translation. Proceedings of the National Academy of Sciences 2015, 112, 15396–15401. [Google Scholar] [CrossRef]
- Goldman, A.D.; Kacar, B. Cofactors are remnants of life’s origin and early evolution. Journal of molecular evolution 2021, 89, 127–133. [Google Scholar] [CrossRef]
- Garcia, A.K.; Kaçar, B. How to resurrect ancestral proteins as proxies for ancient biogeochemistry. Free Radical Biology and Medicine 2019, 140, 260–269. [Google Scholar] [CrossRef]
- Kaçar, B. Reconstructing early microbial life. Annual Review of Microbiology 2024, 78, 463–492. [Google Scholar] [CrossRef]
- Stüeken, E.E.; Anderson, R.; Bowman, J.; Brazelton, W.; Colangelo-Lillis, J.; Goldman, A.; Som, S.; Baross, J. Did life originate from a global chemical reactor? Geobiology 2013, 11, 101–126. [Google Scholar] [CrossRef] [PubMed]
- Kitadai, N.; Maruyama, S. Origins of building blocks of life: A review. Geoscience Frontiers 2018, 9, 1117–1153. [Google Scholar] [CrossRef]
- Galloway, T.; Baidya, A.S.; Cousins, C.R.; Stüeken, E.E. Planetary sources of bio-essential nutrients on a prebiotic world. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Mulkidjanian, A.Y.; Bychkov, A.Y.; Dibrova, D.V.; Galperin, M.Y.; Koonin, E.V. Origin of first cells at terrestrial, anoxic geothermal fields. Proceedings of the National Academy of Sciences 2012, 109, E821–E830. [Google Scholar] [CrossRef]
- Preiner, M.; Xavier, J.C.; Sousa, F.L.; Zimorski, V.; Neubeck, A.; Lang, S.Q.; Greenwell, H.C.; Kleinermanns, K.; Tüysüz, H.; McCollom, T.M.; et al. Serpentinization: connecting geochemistry, ancient metabolism and industrial hydrogenation. Life 2018, 8, 41. [Google Scholar] [CrossRef]
- Xavier, J.C.; Kauffman, S. Small-molecule autocatalytic networks are universal metabolic fossils. Philosophical Transactions of the Royal Society A 2022, 380, 20210244. [Google Scholar] [CrossRef]
- Harrison, S.A.; Rammu, H.; Liu, F.; Halpern, A.; Nunes Palmeira, R.; Lane, N. Life as a guide to its own origins. Annual Review of Ecology, Evolution, and Systematics 2023, 54, 327–350. [Google Scholar] [CrossRef]
- Mrnjavac, N.; Schwander, L.; Brabender, M.; Martin, W.F. Chemical antiquity in metabolism. Accounts of Chemical Research 2024, 57, 2267–2278. [Google Scholar] [CrossRef]
- Weiss, M.C.; Preiner, M.; Xavier, J.C.; Zimorski, V.; Martin, W.F. The last universal common ancestor between ancient Earth chemistry and the onset of genetics. PLoS genetics 2018, 14, e1007518. [Google Scholar] [CrossRef]
- Negrón-Mendoza, A.; Hernández-Morales, R.; Lazcano, A. Can the origin of biosynthetic routes be explained by a Frankenstein’s monster-like spontaneous assembly of prebiotic reactants? Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Carbonell, P.; Peretó, J. Before LUCA: Unearthing the chemical roots of metabolism. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Wächtershäuser, G. Life as we don’t know it. Science 2000, 289, 1307–1308. [Google Scholar] [CrossRef]
- Cleland, C.E. The Quest for a Universal Theory of Life: Searching for Life as We Don’t Know It; Cambridge University Press, 2019.
- Walker, S.I. Life as No One Knows it: The Physics of Life’s Emergence; Penguin, 2024.
- Schopf, J.W. Cradle of Life: The Discovery of Earth’s Earliest Fossils; Princeton University Press, 1999.
- Marais, D.D.; Walter, M. Astrobiology: exploring the origins, evolution, and distribution of life in the universe. Annual Review of Ecology and Systematics 1999, 30, 397–420. [Google Scholar] [CrossRef]
- Smith, H.H.; Hyde, A.S.; Simkus, D.N.; Libby, E.; Maurer, S.E.; Graham, H.V.; Kempes, C.P.; Sherwood Lollar, B.; Chou, L.; Ellington, A.D.; et al. The grayness of the origin of life. Life 2021, 11, 498. [Google Scholar] [CrossRef] [PubMed]
- Asche, S.; Bautista, C.; Boulesteix, D.; Champagne-Ruel, A.; Mathis, C.; Markovitch, O.; Peng, Z.; Adams, A.; Dass, A.V.; Buch, A.; et al. What it takes to solve the Origin (s) of Life: An integrated review of techniques. arXiv 2023, arXiv:2308.11665. [Google Scholar] [CrossRef]
- Solé, R.V.; Macia, J. Expanding the landscape of biological computation with synthetic multicellular consortia. Natural Computing 2013, 12, 485–497. [Google Scholar] [CrossRef]
- Hofmann, M.; Abdo1, F.; Borkowski, O.; Kushwaha, M. Reconstituting alternative life using the test-bed of cell-free systems. Philosophical Transactions of the Royal Society B.
- Noireaux, V.; Liu, A.P. The new age of cell-free biology. Annual review of biomedical engineering 2020, 22, 51–77. [Google Scholar] [CrossRef]
- Gánti, T. Chemoton Theory vol 2: theory of living systems; Kluwer, 2003.
- Solé, R.V.; Munteanu, A.; Rodriguez-Caso, C.; Macía, J. Synthetic protocell biology: from reproduction to computation. Philosophical Transactions of the Royal Society B 2007, 362, 1727–1739. [Google Scholar] [CrossRef]
- Solé, R.; Maull, V.; Amor, D.R.; Mauri, J.P.; Núria, C.P. Synthetic ecosystems: From the test tube to the biosphere. ACS Synthetic Biology 2024, 13, 3812–3826. [Google Scholar] [CrossRef]
- Maull, V.; Pla Mauri, J.; Conde Pueyo, N.; Solé, R. A synthetic microbial Daisyworld: planetary regulation in the test tube. Journal of The Royal Society Interface 2024, 21, 20230585. [Google Scholar] [CrossRef]
- Das, S.; Pal, R.; Rajamani, S. Dynamical interactions among protocell populations: Implications for membrane-mediated chemical evolution. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Shirt-Ediss, B.; Ferrero-Fernández, A.; Martino, D.D.; Bich, L.; Moreno, A.; Ruiz-Mirazo, K. Modelling the prebiotic origins of regulation and agency in evolving protocell ecologies. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Thomsen, K.R.; Kolchinsky, A.; Rasmussen, S. Protocellular energy transduction, information, and fitness. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Stepney, S. Towards Origins of Virtual Artificial Life: an overview. Philosophical Transactions of the Royal Society B 2025, This issue. [Google Scholar]
- Bartlett, S.; Wong, M.L. Defining Lyfe in the Universe: From Three Privileged Functions to Four Pillars. Life 2020, 10, 42. [Google Scholar] [CrossRef]
- Kane, S.R.; Hill, M.L.; Kasting, J.F.; Kopparapu, R.K.; Quintana, E.V.; Barclay, T.; Batalha, N.M.; Borucki, W.J.; Ciardi, D.R.; Haghighipour, N.; et al. A catalog of Kepler habitable zone exoplanet candidates. The Astrophysical Journal 2016, 830, 1. [Google Scholar] [CrossRef]
- Bohl, A.; Lawrence, L.; Lowry, G.; Kaltenegger, L. Probing the Limits of Habitability: A Catalog of Rocky Exoplanets in the Habitable Zone. arXiv 2025, arXiv:2501.14054. [Google Scholar]
- Madhusudhan, N.; Constantinou, S.; Holmberg, M.; Sarkar, S.; Piette, A.A.; Moses, J.I. New Constraints on DMS and DMDS in the Atmosphere of K2-18 b from JWST MIRI. The Astrophysical Journal Letters 2025, 983, L40. [Google Scholar] [CrossRef]
- Bedau, M.A.; Cleland, C.E. The nature of life; Cambridge University Press, 2018.
- Henson, A.; Gutierrez, J.M.P.; Hinkley, T.; Tsuda, S.; Cronin, L. Towards heterotic computing with droplets in a fully automated droplet-maker platform. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 2015, 373, 20140221. [Google Scholar] [CrossRef]
- Vincent, L.; Berg, M.; Krismer, M.; Saghafi, S.T.; Cosby, J.; Sankari, T.; Vetsigian, K.; Cleaves, H.J.; Baum, D.A. Chemical ecosystem selection on mineral surfaces reveals long-term dynamics consistent with the spontaneous emergence of mutual catalysis. Life 2019, 9, 80. [Google Scholar] [CrossRef]
- Baum, D.A.; Vetsigian, K. An experimental framework for generating evolvable chemical systems in the laboratory. Origins of Life and Evolution of Biospheres 2017, 47, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Sagan, C. Definitions of life. In The Nature of Life: Classical and Contemporary Perspectives from Philosophy and Science; 2010; pp. 303–306. [Google Scholar]
- Knuuttila, T.; Loettgers, A. What are definitions of life good for? Transdisciplinary and other definitions in astrobiology. Biology & Philosophy 2017, 32, 1185–1203. [Google Scholar] [CrossRef]
- Szathmáry, E. The evolution of replicators. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 2000, 355, 1669–1676. [Google Scholar] [CrossRef]
- Lee, K.J.; McCormick, W.D.; Pearson, J.E.; Swinney, H.L. Experimental observation of self-replicating spots in a reaction–diffusion system. Nature 1994, 369, 215–218. [Google Scholar] [CrossRef]
- Hodgman, C.E.; Jewett, M.C. Cell-free synthetic biology: thinking outside the cell. Metabolic engineering 2012, 14, 261–269. [Google Scholar] [CrossRef]
- Lancet, D.; Zidovetzki, R.; Markovitch, O. Systems protobiology: origin of life in lipid catalytic networks. Journal of The Royal Society Interface 2018, 15, 20180159. [Google Scholar] [CrossRef]
- Gánti, T. Chemoton theory: theory of living systems; Springer, 2003.
- Macía, J.; Solé, R.V. Protocell self-reproduction in a spatially extended metabolism–vesicle system. Journal of theoretical biology 2007, 245, 400–410. [Google Scholar] [CrossRef]
- Macia, J.; Solé, R.V. Synthetic Turing protocells: vesicle self-reproduction through symmetry-breaking instabilities. Philosophical Transactions of the Royal Society B: Biological Sciences 2007, 362, 1821–1829. [Google Scholar] [CrossRef]
- Fellermann, H.; Solé, R.V. Minimal model of self-replicating nanocells: a physically embodied information-free scenario. Philosophical Transactions of the Royal Society B: Biological Sciences 2007, 362, 1803–1811. [Google Scholar] [CrossRef]
- Li, M.; Huang, X.; Mann, S. Spontaneous Growth and Division in Self-Reproducing Inorganic Colloidosomes. Small 2014, 10, 3291–3298. [Google Scholar] [CrossRef]
- Fellermann, H.; Rasmussen, S.; Ziock, H.J.; Solé, R.V. Life cycle of a minimal protocell—a dissipative particle dynamics study. Artificial Life 2007, 13, 319–345. [Google Scholar] [CrossRef] [PubMed]
- Barge, L.M.; Cardoso, S.S.; Cartwright, J.H.; Cooper, G.J.; Cronin, L.; De Wit, A.; Doloboff, I.J.; Escribano, B.; Goldstein, R.E.; Haudin, F.; et al. From chemical gardens to chemobrionics. Chemical Reviews 2015, 115, 8652–8703. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.S.; Cartwright, J.H.; Čejková, J.; Cronin, L.; De Wit, A.; Giannerini, S.; Horváth, D.; Rodrigues, A.; Russell, M.J.; Sainz-Díaz, C.I.; et al. Chemobrionics: From self-assembled material architectures to the origin of life. Artificial Life 2020, 26, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Langton, C.G. Studying artificial life with cellular automata. Physica D: nonlinear phenomena 1986, 22, 120–149. [Google Scholar] [CrossRef]
- Penrose, L.S. Self-reproducing machines. Scientific American 1959, 200, 105–117. [Google Scholar] [CrossRef]
- Morowitz, H.J. A model of reproduction. American Scientist 1959, 47, 261–263. [Google Scholar]
- Sole, R.V. Evolution and self-assembly of protocells. The international journal of biochemistry & cell biology 2009, 41, 274–284. [Google Scholar]
- Stano, P.; Luisi, P.L. Achievements and open questions in the self-reproduction of vesicles and synthetic minimal cells. Chemical Communications 2010, 46, 3639–3653. [Google Scholar] [CrossRef]
- Noireaux, V.; Maeda, Y.T.; Libchaber, A. Development of an artificial cell, from self-organization to computation and self-reproduction. Proceedings of the National Academy of Sciences 2011, 108, 3473–3480. [Google Scholar] [CrossRef]
- Xu, C.; Hu, S.; Chen, X. Artificial cells: from basic science to applications. Materials today 2016, 19, 516–532. [Google Scholar] [CrossRef] [PubMed]
- Hirschi, S.; Ward, T.R.; Meier, W.P.; Müller, D.J.; Fotiadis, D. Synthetic biology: bottom-up assembly of molecular systems. Chemical Reviews 2022, 122, 16294–16328. [Google Scholar] [CrossRef] [PubMed]
| 1 |
Figure 1.
The many paths to the origins of life. Several approaches to the problem of how life originated, evolved and became complex are outlined here. These include, among others: (a) prebiotic experiments using simple mixtures of molecules, such as this recreation of Titan’s atmosphere; (b) extended Miller-like experiments with concomitant formation of embodied structures; (c) evolutionary dynamics of chemical droplets using a combination of in vitro selection and AI; (d) the analysis of primitive Earth environments; (e) the exploration of exoplanets; (f) extreme life forms, such as those found in ocean vents; (g) engineering minimal genomes, such Mycoplasma JVC1-syn1.0; (h) cell-free systems obtained from cells or reconstituted from purified components. and (i) spatially extended artificial life models (here Lenia); (j) combinatorial chemistry studies; (k) evolutionary aspects of metabolic networks and the diverse components associated to them, inferred from comparative genomics studies and (j) full-model simulation of cells using (still limited) computational approximations to molecular dynamics that try to replicate (on short time scales) some of the supramolecular processes taking place in the crowded cellular environment (illustration by D. Goodsell).
Figure 1.
The many paths to the origins of life. Several approaches to the problem of how life originated, evolved and became complex are outlined here. These include, among others: (a) prebiotic experiments using simple mixtures of molecules, such as this recreation of Titan’s atmosphere; (b) extended Miller-like experiments with concomitant formation of embodied structures; (c) evolutionary dynamics of chemical droplets using a combination of in vitro selection and AI; (d) the analysis of primitive Earth environments; (e) the exploration of exoplanets; (f) extreme life forms, such as those found in ocean vents; (g) engineering minimal genomes, such Mycoplasma JVC1-syn1.0; (h) cell-free systems obtained from cells or reconstituted from purified components. and (i) spatially extended artificial life models (here Lenia); (j) combinatorial chemistry studies; (k) evolutionary aspects of metabolic networks and the diverse components associated to them, inferred from comparative genomics studies and (j) full-model simulation of cells using (still limited) computational approximations to molecular dynamics that try to replicate (on short time scales) some of the supramolecular processes taking place in the crowded cellular environment (illustration by D. Goodsell).

Figure 2.
Life as a space of the possible. This figure presents a qualitative morphospace in which three distinct aspects of complexity—related to extant, synthetic, and theoretical self-replicating systems—are used to position a range of case studies relative to one another. Included are modern cells, synthetic protocells, minimal-genome cells, theoretical models of self-replicating entities, spatially distributed cellular automata, and thermodynamic machines. The right panel shows the entire morphospace, highlighting a large void that may represent either unexplored or fundamentally inaccessible regions. The left panel provides a close-up view of a region populated by synthetic droplets, protocells, and various theoretical constructs. Some non-cellular systems are also indicated using black circles as a reference. Within the synthetic cube, we indicate: (1) self-replicating spots [170], cell-free systems (SS) [171], lipid composome [172], evolved droplets [33], the chemoton [173], Rashevsky droplets [174], Turing protocells [175], synthetic nanocells [176], replicating inorganic colloidosomes, black sphere) [177] or the Los Alamos Bug (LABug) [178]. We also indicate chemical gardens (Chem Gardens) as another corner of spatially-extended self-organizing chemical systems [179,180]. Additionally, three classic examples of abstract models of self-reproducing machines or cells are also indicated, namely: (i) Langton’s loop [181] based on a cellular automaton running on a two-dimensional lattice, (ii) Penrose blocks [182] based on physical blocks that interact and self-assemble into larger structures and can display "reproduction" and (iii) Morowitz electromechanical machine [183].
Figure 2.
Life as a space of the possible. This figure presents a qualitative morphospace in which three distinct aspects of complexity—related to extant, synthetic, and theoretical self-replicating systems—are used to position a range of case studies relative to one another. Included are modern cells, synthetic protocells, minimal-genome cells, theoretical models of self-replicating entities, spatially distributed cellular automata, and thermodynamic machines. The right panel shows the entire morphospace, highlighting a large void that may represent either unexplored or fundamentally inaccessible regions. The left panel provides a close-up view of a region populated by synthetic droplets, protocells, and various theoretical constructs. Some non-cellular systems are also indicated using black circles as a reference. Within the synthetic cube, we indicate: (1) self-replicating spots [170], cell-free systems (SS) [171], lipid composome [172], evolved droplets [33], the chemoton [173], Rashevsky droplets [174], Turing protocells [175], synthetic nanocells [176], replicating inorganic colloidosomes, black sphere) [177] or the Los Alamos Bug (LABug) [178]. We also indicate chemical gardens (Chem Gardens) as another corner of spatially-extended self-organizing chemical systems [179,180]. Additionally, three classic examples of abstract models of self-reproducing machines or cells are also indicated, namely: (i) Langton’s loop [181] based on a cellular automaton running on a two-dimensional lattice, (ii) Penrose blocks [182] based on physical blocks that interact and self-assemble into larger structures and can display "reproduction" and (iii) Morowitz electromechanical machine [183].
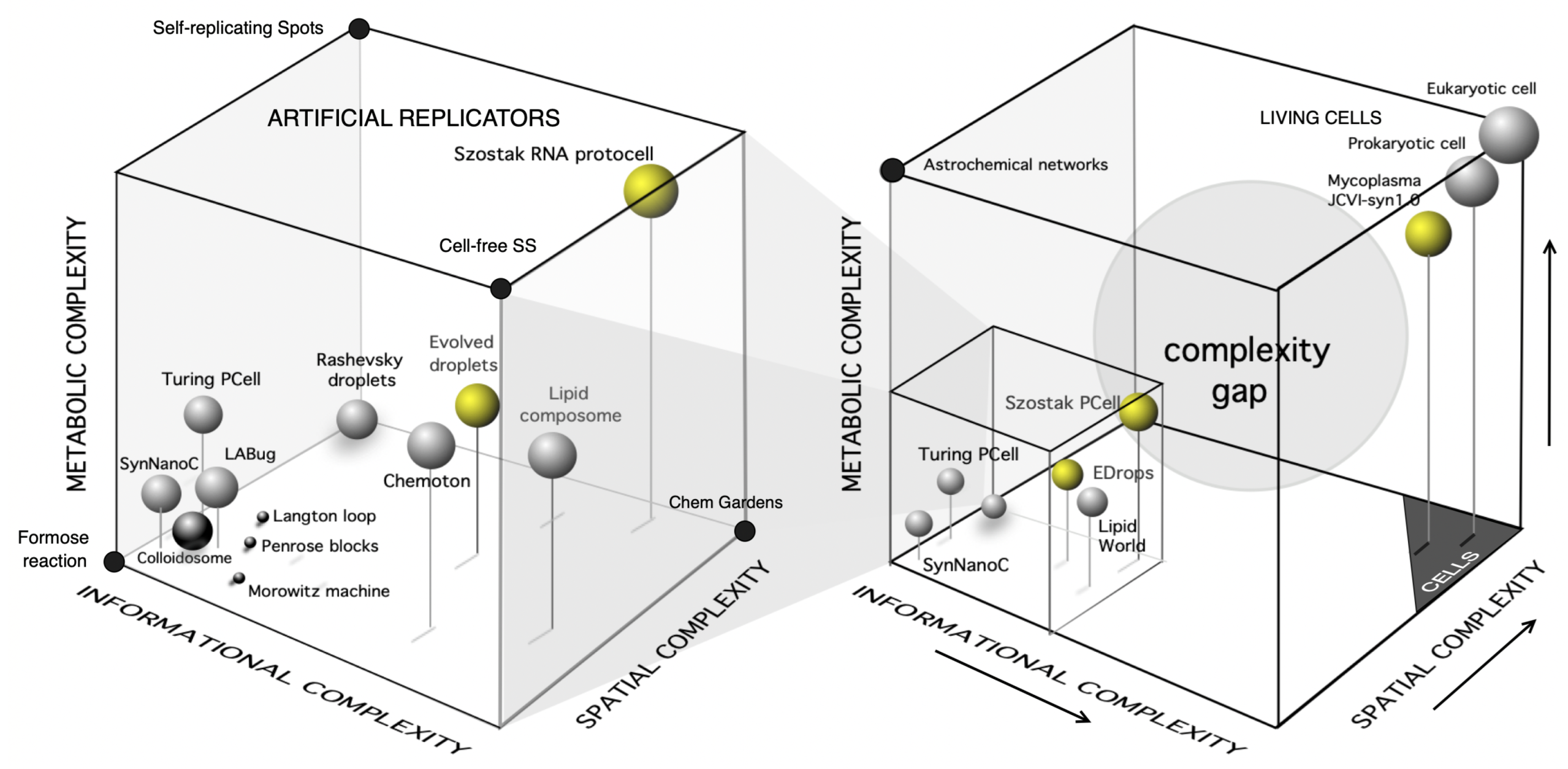
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



