Submitted:
19 August 2025
Posted:
20 August 2025
You are already at the latest version
Abstract
For eight years prior to Argentina's malaria-free certification in 2019, there were no instances of local transmission. Epidemiological surveillance focused on detecting Plasmodium vivax in areas where the last cases had been reported. During the national malaria surveillance program (2016-2018) in Salvador Mazza (Salta Province, northwestern Argentina), several neighborhoods were randomly selected for the collection of human blood samples to detect any silent circulation of P. vivax. Diagnosis of malaria parasites relied on traditional microscopy and molecular detection using blood collected on filter paper, by amplifying and sequencing a portion of the Plasmodium cytochrome b gene. An autochthonous case of P. vivax was identified in an asymptomatic 64-year-old individual in La Bendición neighborhood, Salvador Mazza. The individual had never traveled to any P. vivax-endemic region. This case is the first detected among 92 samples collected from various localities along Argentina's borders with Bolivia (northwest) and Brazil (northeast). This finding highlights the possibility of silent circulation of P. vivax in areas previously assumed to be malaria-free and raises concerns regarding the timing of the certification, prompting a reevaluation of the current situation. The extent of P. vivax circulation among asymptomatic individuals remains largely unknown. This is the first molecularly confirmed asymptomatic case of P. vivax reported following malaria-free certification in Argentina and the Southern Cone region. Currently, there is no active epidemiological or entomological surveillance in the area where the case was recorded. The permeable nature of the border facilitates disease transmission, and the lack of information about the movement of asymptomatic individuals is particularly concerning. It is imperative that the surveillance system responds effectively to maintain the status as a malaria-free country, prioritizing this within the national health system agenda due to the potential implications of silent malaria circulation and the risk of re-emergence and re-establishment.
Keywords:
asymptomatic malaria
; Argentina
; silent circulation
; parasite
; prevention of reintroduction
1. Introduction
Plasmodium vivax is a parasite widely distributed across Central and South America, Asia, and Oceania. In the American continent, this parasite is the most prevalent, and although it is less pathogenic than P. falciparum, both species still cause high morbidity and mortality in endemic areas [1,2,3]. The introduction of P. vivax in the Americas is considered a recent event from an evolutionary and dispersal perspective. It may have undergone two colonization events (pre-Columbian times and European colonization), which over time, allowed various adaptation processes involving parasites, vectors, and non-human primates interactions. These events led to the emergence of genetically related lineages, as P. simium [4].
The successful dispersion of P. vivax is mainly attributed to human migratory movements [4,5]. Additionally, inherent aspects of its biology contribute to its spread, among them the early appearance of the infectious form (gametocytes) in the human host, which is detectable from the third day after asexual forms can be detected. Also contributing are persistent relapses in the host, originated by the development of hypnozoites in the liver, and its concentration in a scarce type of red blood cell, which limits parasitemia to low densities in blood [6,7,8]. Under these factors, the control and surveillance of Plasmodium parasite infections prove to be ineffective and challenging in endemic countries lacking active programs. This is primarily due to the presence of patients with asymptomatic cases, who are mostly undetectable, neglected, and remain untreated [9,10]. Evidence has shown that asymptomatic individuals present low parasitemia, which is difficult to detect with routine field tests such as microscopy or rapid diagnostic tests (RDTs) [11]. Although the mechanisms involving submicroscopic infections are not fully understood, they may be associated with the sustainability of the disease and its endemicity, as well as the occurrence of new outbreaks [8,12,13,14].
Autochthonous malaria transmission was zero in Argentina since 2011 [15], and the country received the World Health Organization (WHO) certification as malaria-free in 2019 [3]. In past centuries, Argentina had a moderately endemic malaria area in the northwestern region, where autochthonous transmission mainly occurred. In the northwestern region P. vivax malaria transmission was typically stable with cycles over seven years. Meanwhile, in the northeastern region of Argentina, malaria transmission was unstable with epidemic outbreaks influenced by several factors related to malaria case occurrence and abundance in Brazil and Paraguay (neighboring countries). The presence and abundance of malaria cases in relation to the main vectors of malaria in both regions, Anopheles pseudopunctipennis and Anopheles darlingi (supposedly cited) was also determined by biogeography (subtropical mountainous rainforest, also called Yungas phytogeographic province in the northwest and the Paranaense biogeographic region in the northeast) and associated seasonal conditions (mainly determined by rainfall and temperature) [15]. The central and southern provinces of the country have been free of autochthonous malaria transmission with only imported malaria reported from overseas.
The prevalence of P. vivax infections was predominant in the country prior to malaria elimination. Plasmodium falciparum was also reported in the country at the beginning of 1900s and its incidence varied over the years, disappearing over time and not being found in samples since 1950. Since then, all the samples screened have only revealed the presence of P. vivax.
During the ARBOL intervention program (1996), a large campaign was carried out in Argentina and Bolivia, resulting in a marked decrease in the number of malaria cases on both sides of the border [16]. Efforts to keep autochthonous malaria cases low and, later, elimination efforts have led to a steady decline in malaria incidence until 2010, when the last 55 autochthonous malaria cases were reported in different localities in San Martín and Orán departments of Salta province, northwestern Argentina [15,16].
From 2011 onwards, the malaria cases registered were classified as imported, mainly from Bolivia, influenced by the migration of people between both countries. Between 2010 and 2017, 191 imported malaria cases were reported in the country [17]. To obtain certification as a malaria-free country, epidemiological and entomological surveillance were carried out during 2013-2018. To confirm that there had been no introduced or autochthonous cases of malaria during these years, epidemiological surveillance was carried out analyzing blood samples taken from people living in neighborhoods of Salvador Mazza locality (Salta province) and Puerto Iguazú locality (Misiones province). Also, entomological surveillance was carried out by collecting mosquitoes around dwellings in both mentioned localities to detect possible P. vivax circulation in these border areas of the country. Following these steps, the certification was obtained in 2019. In 2022, 92 blood samples from 2017-2018, ergo post-certification, were molecularly analyzed for the presence of Plasmodium infections, resulting in the detection of the first asymptomatic malaria case reported here, considered the result of local silent transmission. This manuscript describes the first case of asymptomatic malaria caused by P. vivax in Argentina. The case was initially misdiagnosed by routine microscopy but was subsequently detected by molecular techniques and confirmed as P. vivax infection.
2. Materials and Methods
2.1. Study area
For the certification process as a country free of autochthonous malaria transmission, two main strata were identified in Argentina to work on, located in both the Northwest and Northeast of the country. Stratum 1 includes certain localities in the departments of San Martín and Orán (extreme north of the province of Salta), which, being the areas where the last autochthonous malaria cases were recorded, required and still require constant epidemiological and entomological surveillance. Stratum 2 includes localities in the departments of Rivadavia, Anta, Iruya, Ledesma, Santa Bárbara, and San Pedro (south of Salta province), in Jujuy province (northwest Argentina), and Puerto Iguazú in Misiones province (extreme northeast Argentina), requiring a different type of surveillance compared to stratum 1. The surveillance system in both strata, prior to certification, depended on the National Vector Control Coordination of the Ministry of Health of the Nation. After certification, each province, through its own provincial health systems, had to take charge of surveillance in both strata (Figure 1A, B).
In Stratum 1, the locality where both the last autochthonous and imported cases were registered was Salvador Mazza in Salta province, on the border with the Republic of Bolivia. This town covers 501 km2 and has 20,670 inhabitants living in 4,725 homes under different socio-environmental conditions [18]. Some of its neighborhoods/settlements that continue to grow over time include El Arenal, Arenales, Comunidad La Bendición, Monte Sinaí, La Pista, Ferroviario 1, Ferroviario 2, El Milagro, San Cayetano, Sector 5, 28 de Marzo, El Obraje, 18 de Marzo, Nueva Esperanza, Evita, El Paraíso, Barrio Nuevo, Barrio Municipal, Barrio La Cruz, El Chorro, Barrio La Playa, Barrio San Pedro, and Barrio Centro-Norte. Of these neighborhoods/settlements, Comunidad La Bendición, El Arenal, Arenales, Nueva Esperanza, Barrio La Playa, El Chorro, Sector 5, El Obraje in Salvador Mazza, along with Barrio Ferroviario and Barrio Centro in the city of Tartagal, were selected due to the presence of autochthonous and imported malaria cases prior to certification (Figure 2) (Table 1).
In Stratum 2, the locality where both the last autochthonous and imported cases were registered was mainly Puerto Iguazú in Misiones province, on the border with the Republic of Brazil. This town covers 16 km2 and has 39,836 inhabitants living in 10,655 homes under different socio-environmental conditions [18]. Some of its neighborhoods/settlements that continue to grow over time include Villa Nueva, Villa Florida, Villa Tacuara, Obrero, Almirante Brown, Santa María del Iguazú, Barrio Villa Alta, Barrio Libertad, Bravo, Malvinas Neighborhood, Barrio Belén, Barrio San Lucas, Barrio Santa Rosa, Barrio Bicentenario, Barrio Catarata, Barrio 1 de Mayo, Barrio 25 de Mayo, Barrio Nuevo Iguazú, Barrio Alto Paraná, Barrio Hermoso, and Barrio 2000 Hectares, among others. Of these neighborhoods/settlements, Barrio 2000 Hectares, Ribera del Paraná, Villa Alta, Barrio Belén, Barrio Obrero, Barrio Santa Rosa, and Barrio San Lucas were selected due to the presence of imported and autochthonous malaria cases during the last epidemic outbreak registered in the northeast of the country prior to certification (Figure 3) (Table 1).
2.2. Epidemiological Surveillance
Since the 20th century, Argentina has had a strong epidemiological surveillance system, which has evolved in response to changing malaria transmission scenarios in the northern region of the country [15]. Epidemiological surveillance is initiated within 24 hours of reporting a malaria case and an intensive search for relatives or neighbors is carried out within approximately a 1-km radius of the malaria case' residence. The dwellings of family members and neighbors are surveyed, blood samples are collected for routine microscopy analysis, and both the houses and their peridomicile areas are sprayed with insecticides [17].
Within the National Malaria Program of the Ministry of Health of the Nation, malaria case detection in Argentina is carried out not only through passive case detection, where symptomatic individuals seek care at national or provincial health facilities, but also, and more importantly, through active case detection. This involves Ministry technicians conducting field visits to search for potential cases, primarily focusing on populations living in border areas with Bolivia, where cases were continuously recorded regardless of symptom presence. The search for positive cases encompasses entire communities. Malaria cases are diagnosed and treated according to the National Malaria Program guidelines of the National Ministry of Health [17].
Diagnosis is primarily based on routine thick blood smear microscopy. Since 2011, in preparation for certification, an additional filter paper sample was collected for molecular detection via polymerase chain reaction (PCR), which became essential for parasite confirmation. These molecular analyses were carried out in collaboration with the International Research Laboratory REHABS in South Africa. Sequencing was requested when recurrence or asymptomatic circulation of the parasite was suspected in the population under study.
2.3. Entomological Surveillance
Since 2011, when work began for certification, entomological surveillance was incorporated as a key component supporting epidemiological surveillance. Sampling of immature mosquito stages was conducted in water bodies such as the Bermejo River, Itiyuro Dam, Acambuco streams, Virgen de la Peña, and San Roquito, all within Stratum 1.
With the data obtained from the identification of the Anopheles species, added to their presence and abundance per body of water, we proceeded to characterize the breeding sites were characterized by risk levels, primarily due to the presence of malaria vector species, Anopheles pseudopunctipennis and Anopheles argyritarsis. Although entomological surveillance was not carried out in the exact neighborhood where the reported case occurred, immature forms were searched in nearby breeding sites, as well as adult mosquitoes.
2.4. Ethics Statement
This study was performed under protocols reviewed and approved by the National Ministry of Health of Argentina. All participants were informed about the objectives and procedures of the study, with voluntary participation through signed informed consent.
2.5. Malaria Diagnosis
2.5.1. Routine Microscopy
From December 2015 to November 2017, spring and summer surveys were conducted in both northern regions of Argentina, selecting these seasons due to the recorded peaks in malaria transmission. After obtaining informed consent, 92 blood samples were collected from individuals living in previously selected localities. 50 µL of blood was extracted from each person via digital puncture; thick blood smears were immediately prepared on slides and stained with GIEMSA. To visualize the presence of parasites in the asexual blood phase, slides were examined under a microscope at 1000X magnification using immersion oil. When asexual parasite stages were detected, parasite density was calculated by counting parasites per 200 leukocytes, with values expressed as parasites per µL. [18].
2.5.2. DNA Extraction and Cyt b Gene PCR Amplification
For each blood spot, DNA was extracted using the QIAgen DNeasy Blood and Tissue kit recommendations (QIAGEN, Germany). To determine the presence of Plasmodium spp., a portion of the mitochondrial cytochrome b (cyt-b) gene was amplified by Polymerase Chain Reaction (PCR), as described by Prugnolle et al. [19]. Additionally, 10 µL of the PCR product was verified by electrophoresis on a 1.5% agarose gel in TAE buffer. Once the 956 bp fragment of the cyt-b gene was obtained, it was purified. This purified DNA served as a template for sequencing. DNA sequencing of the fragment was performed by Eurofins MWG (Germany). The sequence reported in this study has been deposited in GenBank under accession number PP658217.
2.5.3. Phylogenetic Analyses
To determine the identity of the Plasmodium isolate from Argentina (accession number PP658217), we conducted a nucleotide BLAST search against the NCBI database [20,21]. The sequence exhibited a high degree of similarity (99%) to Plasmodium vivax mitochondrial genomes, confirming its species-level affiliation. Subsequently, we retrieved 35 P. vivax sequences of the cyt-b gene from GenBank, representing a broad geographical range (see Table 2). Three Plasmodium falciparum sequences (AJ276847 from Brazil, AY282957 from Ecuador, and AY282971 from Colombia) were selected to serve as outgroup taxa for phylogenetic rooting.
Multiple sequence alignment was performed using MUSCLE v3.8.31 [22], employing default settings designed to iteratively refine the alignment and balance accuracy with computational efficiency [22,23]. To evaluate the phylogenetic informativeness of the dataset and its capacity to resolve evolutionary relationships—including those involving the Argentinian isolate—we estimated the strength of the phylogenetic signal using Likelihood Mapping. This analysis was conducted in IQ-TREE v3 [24,25] using 10,000 randomly sampled quartets.
Model selection was performed to identify the most appropriate nucleotide substitution model. We applied the Akaike Information Criterion (AIC) through the Phymltest() function in the ape package [26], which interfaces with PhyML [27], all executed within the R environment [28].
Maximum likelihood phylogenetic inference was carried out using RAxML-NG v1.2.0 [29,30], employing the --all option to perform both tree search and bootstrap resampling. The analysis was based on a PHYLIP-formatted alignment of 38 sequences, using the GTR+G substitution model to account for among-site rate heterogeneity. To ensure a comprehensive exploration of tree space, ten independent tree searches were conducted. Node support was evaluated with 1,000 bootstrap replicates, and support values were calculated using the transfer bootstrap expectation (TBE) metric [31]. The analysis was executed with a random seed and parallelized using four computational threads.
3. Results
3.1. Circulation of Asymptomatic Plasmodium Vivax
An overall of 92 individuals were tested for P. vivax infection by routine microscopy and molecular techniques (PCR), from whom one person was PCR positive. All samples were negative in thick blood smear. The Plasmodium-infected individual was a retired 66-year-old male from La Bendición neighborhood, Salvador Mazza locality, Salta province near the border with Bolivia. He did not present any symptoms either before or at the time of sample collection. He reported no history of travelling abroad. He mentioned a past history of a single malaria episode in 1996 and no history of relapses.
La Bendición is located in a rural area, formerly endemic for malaria. Sixty Guarani families live in an area of 265 hectares. The migration of people is a constant dynamic throughout the year, as residents frequently travel to Bolivia for business, work and/or to visit relatives.
3.2. Phylogenetic Analyses
The phylogenetic reconstruction of P. vivax based on Cytochrome b sequences reveals a generally well-resolved topology, with several moderately to strongly supported regional clades. The Argentinian isolate (P. vivax PP658217 ARG) does not cluster within the main South American clade—which includes sequences from Colombia (AY791547 COL) and Brazil (AY791531 BRA)—but instead falls as a sister lineage to that group. Notably, it appears more closely related to the Papua New Guinea clade, which is strongly supported (bootstrap = 98). The South American clade itself is only moderately supported (bootstrap = 63), and the broader grouping that includes sequences from Mexico, Peru, Belize, and Venezuela exhibits variable support, with bootstrap values ranging from 20 to 68, reflecting uncertainty in some internal relationships. In contrast, the basal divergence between P. vivax and P. falciparum is robustly supported (bootstrap = 100). Overall, the phylogenetic position of the Argentinian isolate underscores its distinctiveness within the global P. vivax diversity (Figure 4).
4. Discussion
Between 2000 and 2020, 23 countries obtained certification as free of indigenous malaria transmission, while other countries in the process of certification showed a reduction of 82.8% of cases for the period 2010-2021 [2,32]. The elimination of indigenous malaria transmission in a country is a process that requires continuous effort, where the main concern is the resurgence of the disease in areas or countries where it is already considered eradicated [32,33]. Factors such as human mobility from endemic areas, the presence of mosquito vectors due to the availability of breeding sites linked to land use and land cover, and favorable climatic/environmental conditions (precipitation, relative humidity, and ambient temperature), represent high risk factors for malaria reintroduction [32,34,35,36].
Changes in land use are related to the reemergence of malaria by providing optimal conditions for the appearance of new breeding sites of the mosquito vector. Cuellar et al. [37] observed in the locality of San Ramón de la Nueva Orán, how the increase in land use for agriculture, with the consequent reduction of the original forest, caused a significant increase in malaria cases, being this the first work to report this situation for Argentina. Other indicators used in predicting the risk of malaria transmission, especially in areas free of the disease, are the presence, diversity, and abundance of competent vectors, their feeding behavior, and biting activity.
If financial resources are limited, entomological surveillance cannot be carried out and, therefore, increases in mosquito populations may occur. In addition, the absence of serological monitoring and the limited information on the silent circulation of the parasite in areas free of malaria transmission further complicate the situation [32,38,39]. It is important to consider that infectious malaria cases by P. vivax represent an additional challenge in both, countries with active transmission and those that have been certified as free of transmission, because this parasite species has biological characteristics such as the early appearance of gametocytes, the development of hypnozoites in the liver, and the preferential infection of reticulocytes [6,7,8]. These characteristics determine that asymptomatic individual’s positive for P. vivax are difficult to detect and act as silent reservoirs, facilitating the perpetuation and amplification of malaria transmission [8,9,10].
Karunasena et al. [33] and World Health Organization [3] reported for countries in Asia, in the Middle East, and in the Americas that the geographical context with the porous borders with countries where malaria transmission is still present represents challenges for preventing the malaria reintroduction and to maintaining the elimination efforts. This situation is also applicable to Argentina, which widely shares its border with countries such as Bolivia and Brazil, countries that have historically presented challenges in malaria control. This context shows similarities with what was reported in Sri Lanka, a country which has certified as free of malaria in 2016, although it continued to face challenges in malaria control, not only because of the difficulty to differentiate the fever symptom of malaria with viral and bacterial infections or the apparition of imported cases, but also due failures in the detection of rapid diagnostic tests in the field, which led Sri Lanka to report its first case, six years after elimination, as a result of local transmission from an imported case [33]. In Argentina, a similar situation could have occurred.
The epidemiological surveillances were carried out by the technicians of the Coordinación Nacional de Control de Vectores, households in different neighborhoods of Salvador Mazza city. In some of these dwellings, persons were registered who had malaria in the past, being at that time the cases classified as autochthonous, introduced or imported. The technicians collected the samples during spring and summer seasons in the Norwest region of Argentina, when the malaria cases were mainly registered in the past. In the northeast region of the country, the samples were collected during spring season, when the mainly epidemic peaks had occurred in the area [40,41]. Before obtaining the certification, it was supposed to be sure that there were not asymptomatic infections circulating in the endemic area of malaria in the country. The person who was positive to P. vivax was not sick, and he did not manifest any symptoms (fever), and carried out his activities normally. This person did not travel to Bolivia or any other malaria-endemic country; concluding this person acquired the infection in the same place where he lived in Argentina. Since this finding, it is important to consider there had been no cases of imported, introduced or autochthonous malaria reported from anywhere else in the area before the certification of malaria elimination. Then, the question is if there were no registered malaria cases or if there were unregistered malaria cases due to asymptomatic individuals positive for malaria who were not detected by routine analysis for the parasite. In Sri Lanka, asymptomatic malaria cases due to P. vivax parasites have been reported among foreign workers and returnees from endemic countries. Passive and active surveillance strategies were necessary to implement in order to detect these cases. Active detection has identified infections in asylum seekers, travelers, and military personnel. Diagnostic delays are frequent due to low clinical suspicion in a non-endemic context. Most current cases involved adult men aged from 21 to 50, emphasizing the need for targeted surveillance, clinician training, and coordination between health sectors to prevent reestablishment of the disease [33,42]. At a global level, it is necessary that countries, in their efforts to eliminate malaria transmission, achieve the detection of asymptomatic persons and improve the detection systems, trying to understand how these infections affect disease control strategies [8,43,44].
In Argentina, the autochthonous case was detected years later when the interaction with foreign researchers was possible. And so, an effective response to the presence of this case was not mounted to search for more asymptomatic malaria cases as well as to search for the presence of vectors of malaria. The national health system in this instance was not available to detect this situation and then to ensure that Argentina remains malaria-free.
It is important to highlight that in Argentina, as in other countries, the cost and sustainability of entomological and epidemiological surveillance activities have never been fully considered, despite the fact that maintaining national health programs is more cost-effective compared to the expenses associated with treating people sick with malaria, as well as the logistical costs of controlling and mitigating the emergence of new cases when the disease becomes re-established in a country [33,45].
In this context, molecular and phylogenomic analyses become essential tools to complement epidemiological surveillance, providing insights into the origin and potential dissemination pathways of malaria parasites. The phylogenetic reconstruction based on the cytochrome b gene revealed that the Argentinian P. vivax isolate clusters as a sister lineage to the main South American clade, showing affinity with sequences from Colombia, Brazil, Venezuela, and Peru. Interestingly, the Argentinian isolate also appears phylogenetically closer to Papua New Guinea, supported by Rodriguez et al. [5] who proposed Australasian peoples may have introduced P. vivax lineages to South America.
Our evolutionary results are preliminary, but the inclusion of more P. vivax samples from Argentina could reveal the affinities and evolutionary history of local lineages. Polymorphic variation between sequences could reveal not only neutral effects but also potential effects on parasite survival, replication, pathogenicity, or even failures in the detection of rapid diagnostic tests and the development of drug resistance. Nevertheless, it is important to recognize that this study targeted a partial region of the cyt-b gene, which is highly conserved within P. vivax but lacks sufficient resolution to detect fine-scale population structure. Through this result, we can confirm the parasite species. Therefore, future analysis involving larger genomic fragments will allow more robust inferences regarding haplotype distribution, lineage connectivity, and the evolutionary dynamics of P. vivax in the region.
The detection of an asymptomatic case during the certification of indigenous malaria transmission in Argentina raises many questions regarding the mode of infection of the person and the detection techniques used. Nevertheless, the finding made by health professionals and technicians highlights the effectiveness and relevance of the surveillance activities implemented over decades, which laid the basis for the certification obtained in 2019. World Health Organization [46], in its guidelines, mentions that if indigenous transmission is sustained for at least three consecutive years, with the occurrence of three or more indigenous cases of the same species in the same focus, or if the country fails to meet the stipulated criteria for active surveillance and response, malaria transmission could be re-established, which would require restarting the elimination process. Currently, there is no active epidemiological or entomological surveillance in the area where the case was recorded. The permeable nature of the border facilitates disease transmission, and the lack of information about the movement of asymptomatic individuals for malaria is particularly concerning. It is imperative today that the surveillance system responds effectively to protect and maintain the status as a malaria-free country in the long term, prioritizing this within the national health system agenda due to the potential implications of silent malaria circulation and the risk of re-emergence and re-establishment, with strong political commitment, adequate funding, and innovation in molecular surveillance.
5. Conclusions
Four years after the official interruption of malaria transmission, a case of P. vivax malaria was identified in a local resident of a previously endemic region in Argentina. This represented the first autochthonous case reported during the malaria elimination certification process. Although initially misdiagnosed through routine microscopy, the infection was later confirmed by molecular diagnostic methods. At the time, limitations in surveillance infrastructure hindered the timely detection of additional cases and constrained efforts to prevent further transmission. Entomological surveillance played a critical role in supporting the certification process, confirming the presence of Anopheles pseudopunctipennis in both adult and immature stages. However, since the pre-certification phase, no updated official records exist regarding this vector species, and entomological monitoring has not been maintained. Additionally, the absence of routine serological surveillance poses a challenge for the early identification of asymptomatic infections. Sustaining the status of malaria-free transmission requires robust and long-term epidemiological and entomological surveillance systems, particularly in border areas with significant population mobility. Countries that have successfully maintained malaria elimination over extended periods have done so through the implementation of comprehensive and sustained surveillance strategies, recognizing that national elimination contributes meaningfully to regional and global malaria eradication efforts.
Author Contributions
Conceptualization, M.J.D.-J. and M.O.Z.; methodology, M.J.D.-J. and M.O.Z.; formal analysis, M.J.D.-J, V.R., F.D. and N.G.-P.; investigation, M.J.D.-J., P.L.D., R.D.C.-P., B.J.P.-G, F.G.-V. and S.D.-M.; resources, M.O.Z., B.J.P.-G., F.G.-V. and R.D.C.-P.; data curation, J.L.-Z., P.L.D., N.G.-P., V.G. and F.D.; writing—original draft preparation, M.J.D.-J., J.L.-Z., S.D.-M., B.J.P.-G, F.G.-V. and V.R.; writing—review and editing, P.L.D., N.G.-P., and M.J.D.-J.; funding acquisition, M.J.D.J., B.J.P.-G., F.G.-V. and R.D.C.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially supported by Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT PICT-2014-3281). MJDJ is member of the researcher career of CONICET (Consejo Nacional de Investigaciones Científicas y Tecnológicas), Argentina. This work was also supported by the ANR (17-ERC30002) JCJC GENAD and CNRS.
Institutional Review Board Statement
This study was approved by the Ethical Committee of the Ministry of Health of Argentina (Manual de Normas y Procedimientos de Vigilancia y Control de Enfermedades de Notificación Obligatoria, Ministerio de Salud Argentina) (protocol code: CIE-10; approved on 15 November 2022).
Informed Consent Statement
Informed consent was obtained from all the subjects verbally agreed to be involved in this study.
Data Availability Statement
The datasets used during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors specially thank Dr. María Concepción Monteros and Technicians of the Control de Vectores, National Ministry of Health of Argentina, for their invaluable contribution taking the human samples as well as the mosquito’s collection at different localities situated in northern Argentina.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Battle, K.E.; Lucas, T.C.D.; Nguyen, M.; Howes, R.E.; Nandi, A.K.; Twohig, K.A.; et al. Mapping the global endemicity and clinical burden of Plasmodium vivax, 2000–17: A spatial and temporal modeling study. Lancet 2019, 394, 332–343. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2022; World Health Organization: Geneva, Switzerland, 2022; pp. 1–372. [Google Scholar]
- WHO. Countries and territories certified malaria-free by WHO. Available online: https://www.who.int/teams/global-malaria-programme/elimination/countries-and-territories-certified-malaria-free-by-who (accessed on 30 July 2025).
- Rougeron, V.; Daron, J.; Fontaine, M.C.; Prugnolle, F. Evolutionary history of Plasmodium vivax and Plasmodium simium in the Americas. Malar. J. 2022, 21, 141. [Google Scholar] [CrossRef]
- Rodrigues, P.T.; Valdivia, H.O.; de Oliveira, T.C.; Alves, J.M.P.; Duarte, A.M.R.C.; Cerutti-Junior, C.; et al. Human migration and the spread of malaria parasites to the New World. Sci. Rep. 2018, 8, 1993. [Google Scholar] [CrossRef]
- Bousema, T.; Drakeley, C. Epidemiology and infectivity of Plasmodium falciparum and Plasmodium vivax gametocytes in relation to malaria control and elimination. Clin. Microbiol. Rev. 2011, 24, 377–410. [Google Scholar] [CrossRef]
- Martin, T.C.S.; Vinetz, J.M. Asymptomatic Plasmodium vivax parasitaemia in the low-transmission setting: The role for a population-based transmission-blocking vaccine for malaria elimination. Malar. J. 2018, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Almeida, G.G.; Costa, P.A.C.; Araujo, M.d.S.; Gomes, G.R.; Carvalho, A.F.; Figueiredo, M.M.; Pereira, D.B.; Tada, M.S.; Medeiros, J.F.; Soares, I.D.S.; Carvalho, L.H.; Kano, F.S.; Castro, M.D.C.; Vinetz, J.M.; Golenbock, T.; Antonelli, L.R.D.V.; Gazzinelli, R.T. Asymptomatic Plasmodium vivax malaria in the Brazilian Amazon: Submicroscopic parasitemic blood infects Nyssorhynchus darlingi. PLoS Negl. Trop. Dis. 2021, 15, e0009077. [Google Scholar] [CrossRef] [PubMed]
- Cotter, C.; Sturrock, H.J.W.; Hsiang, M.S.; Liu, J.; Phillips, A.A.; Hwang, J.; et al. The changing epidemiology of malaria elimination: New strategies for new challenges. Lancet 2013, 382, 900–911. [Google Scholar] [CrossRef]
- Bousema, T.; Okell, L.; Felger, I.; Drakeley, C. Asymptomatic malaria infections: Detectability, transmissibility and public health relevance. Nat. Rev. Microbiol. 2014, 12, 833–840. [Google Scholar] [CrossRef]
- Recht, J.; Siqueira, A.M.; Monteiro, W.M.; Herrera, S.M.; Herrera, S.; Lacerda, M.V.G. Malaria in Brazil, Colombia, Peru and Venezuela: Current challenges in malaria control and elimination. Malar. J. 2017, 16, 1–18. [Google Scholar] [CrossRef]
- Alves, F.P.; Gil, L.H.S.; Marrelli, M.T.; Ribolla, P.E.M.; Camargo, E.P.; Pereira DaSilva, L.H. Asymptomatic carriers of Plasmodium spp. as infection source for malaria vector mosquitoes in the Brazilian Amazon. J. Med. Entomol. 2005, 42, 777–779. [Google Scholar] [CrossRef] [PubMed]
- Okell, L.C.; Bousema, T.; Griffin, J.T.; Ouedraogo, A.L.; Ghani, A.C.; Drakeley, C.J. Factors determining the occurrence of submicroscopic malaria infections and their relevance for control. Nat. Commun. 2012, 3, 1–9. [Google Scholar] [CrossRef]
- Nguyen, T.N.; von Seidlein, L.; Nguyen, T.V.; Truong, P.N.; Hung, S.D.; Pham, H.T.; et al. The persistence and oscillations of submicroscopic Plasmodium falciparum and Plasmodium vivax infections over time in Vietnam: An open cohort study. Lancet Infect. Dis. 2018, 18, 565–572. [Google Scholar] [CrossRef]
- Zaidenberg, M. The Path to Malaria Elimination in Argentina; Lambert Academic Publishing: Saarbrucken, Germany, 2015. [Google Scholar]
- Berlin, E.; Coello-Peralta, R.D.; Cedeño-Reyes, P.; Valle-Mieles, E.M.; Duque, P.L.; Zaidenberg, M.O.; Madariaga, H.; Navarro, J.C.; Dantur-Juri, M.J.; Castro, M.C. Patterns and Determinants of Imported Malaria near the Argentina–Bolivia Border, 1977–2009. Pathogens 2025, 14, 537. [Google Scholar] [CrossRef]
- Ministerio de Salud de la Nación. Eliminación del Paludismo en Argentina 2018; Ministerio de Salud de la Nación: Buenos Aires, Argentina, 2018; pp. 1–148. [Google Scholar]
- National Institute of Statistics and Censuses Argentina. Population and Housing Census 2010. Available online: https://www.indec.gob.ar/indec/web/Institucional-Indec-QuienesSomosEng (accessed on 1 March 2025).
- Prugnolle, F.; Ollomo, B.; Durand, P.; Yalcindag, E.; Arnathau, C.; Elguero, E.; et al. African monkeys are infected by Plasmodium falciparum nonhuman primate-specific strains. Proc. Natl. Acad. Sci. USA 2011, 108, 11948–11953. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinformatics 2008, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Madden, T.L.; Busby, B.; Ye, J. Reply to the paper: Misunderstood parameters of NCBI BLAST impacts the correctness of bioinformatics workflows. Bioinformatics 2018, 35, 2699–2700. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinformatics 2004, 5, 113. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Wong, T.K.; Ly-Trong, N.; Ren, H.; Baños, H.; Roger, A.J.; Susko, E.; Minh, B.Q. IQ-TREE 3: Phylogenomic inference software using complex evolutionary models. Available online: https://doi.org/10.32942/X2P62N (accessed on 30 July 2025).
- Paradis, E.; Schliep, K. ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.R-project.org/ (accessed on 30 july 2025).
- Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Flouri, T.; Izquierdo-Carrasco, F.; Darriba, D.; Aberer, A.J.; Nguyen, L.T.; Minh, B.Q.; Stamatakis, A. The phylogenetic likelihood library. Syst. Biol. 2015, 64, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Dávila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef]
- Lu, G.; Zhang, D.; Chen, J.; et al. Predicting the risk of malaria re-introduction in countries certified malaria-free: A systematic review. Malar. J. 2023, 22, 175. [Google Scholar] [CrossRef] [PubMed]
- Karunasena, V.M.; Marasinghe, M.; Koo, C.; Amarasinghe, S.; Senaratne, A.S.; Hasantha, R.; et al. The first introduced malaria case reported from Sri Lanka after elimination: Implications for preventing the re-introduction of malaria in recently eliminated countries. Malar. J. 2019, 18, 210. [Google Scholar] [CrossRef]
- WHO. Guidelines on Prevention of the Reintroduction of Malaria; World Health Organization: Geneva, Switzerland, 2007; pp. 1–51. [Google Scholar]
- Schröder, W.; Schmidt, G. Mapping the potential temperature dependent tertian malaria transmission within the ecoregions of Lower Saxony (Germany). Int. J. Med. Microbiol. 2008, 298, 38–49. [Google Scholar] [CrossRef]
- Gao, H.W.; Wang, L.P.; Liang, S.; Liu, Y.X.; Tong, S.L.; Wang, J.J.; et al. Change in rainfall drives malaria re-emergence in Anhui Province, China. PLoS One 2012, 7, e43686. [Google Scholar] [CrossRef]
- Cuellar, A.C.; Manguin, S.; Santana, M.S.; Zaidenberg, M.; Lanfri, M.; Dantur Juri, M.J. The effect of land use change in the abundance of malaria cases in Northern Argentina. 67th Annual Meeting American Society of Tropical Medicine and Hygiene, New Orleans, USA, 28 October 2018.
- Delon, F.; Mayet, A.; Thellier, M.; Kendjo, E.; Michel, R.; Ollivier, L.; et al. Assessment of the French national health insurance information system as a tool for epidemiological surveillance of malaria. J. Am. Med. Inform. Assoc. 2017, 24, 588–595. [Google Scholar] [CrossRef]
- Nasir, S.M.I.; Amarasekara, S.; Wickremasinghe, R.; Fernando, D.; Udagama, P. Prevention of re-establishment of malaria: Historical perspective and future prospects. Malar. J. 2020, 19, 452. [Google Scholar] [CrossRef]
- Bejarano, J.F.R. Distribución en altura del género Anopheles y del paludismo en la República Argentina. Rev. Sanit. Milit. Argent. 1956, 55, 4–24. [Google Scholar]
- Dantur Juri, M.J.; Moreno, M.; Prado Izaguirre, M.J.; Navarro, J.C.; Zaidenberg, M.O.; Almirón, W.R.; Claps, G.L.; Conn, J.E. Demographic history and population structure of Anopheles pseudopunctipennis in Argentina based on the mitochondrial COI gene. Parasit. Vectors 2014, 7, 423. [Google Scholar] [CrossRef] [PubMed]
- Dharmawardena, P.; Premaratne, R.; Wickremasinghe, R.; Mendis, K.; Fernando, D. Epidemiological profile of imported malaria cases in the prevention of reestablishment phase in Sri Lanka. Pathog. Glob. Health 2022, 116, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Coura, J.R.; Suárez-Mutis, M.; Ladeia-Andrade, S. A new challenge for malaria control in Brazil: Asymptomatic Plasmodium infection—A Review. Mem. Inst. Oswaldo Cruz 2006, 101, 229–237. [Google Scholar] [CrossRef]
- Barros, L.B.; Calil, P.R.; Rodrigues, P.T.; Tonini, J.; Fontoura, P.S.; Sato, P.M.; Ferreira, M.U. Clinically silent Plasmodium vivax infections in native Amazonians of northwestern Brazil: Acquired immunity or low parasite virulence? Mem. Inst. Oswaldo Cruz 2022, 117, e220175. [Google Scholar] [CrossRef]
- Shretta, R.; Avancena, A.L.; Hatefi, A. The economics of malaria control and elimination: A systematic review. Malar. J. 2016, 15, 593. [Google Scholar] [CrossRef]
- WHO. Prevention of Re-establishment of Malaria Transmission: Global Guidance; World Health Organization: Geneva, Switzerland, 2025; pp. 1–81. [Google Scholar]
Figure 1.
(A-B) Localities sampled for the detection of asymptomatic malaria cases in San Martín department (Salta province) and Iguazú department (Misiones province), northern Argentina.
Figure 1.
(A-B) Localities sampled for the detection of asymptomatic malaria cases in San Martín department (Salta province) and Iguazú department (Misiones province), northern Argentina.
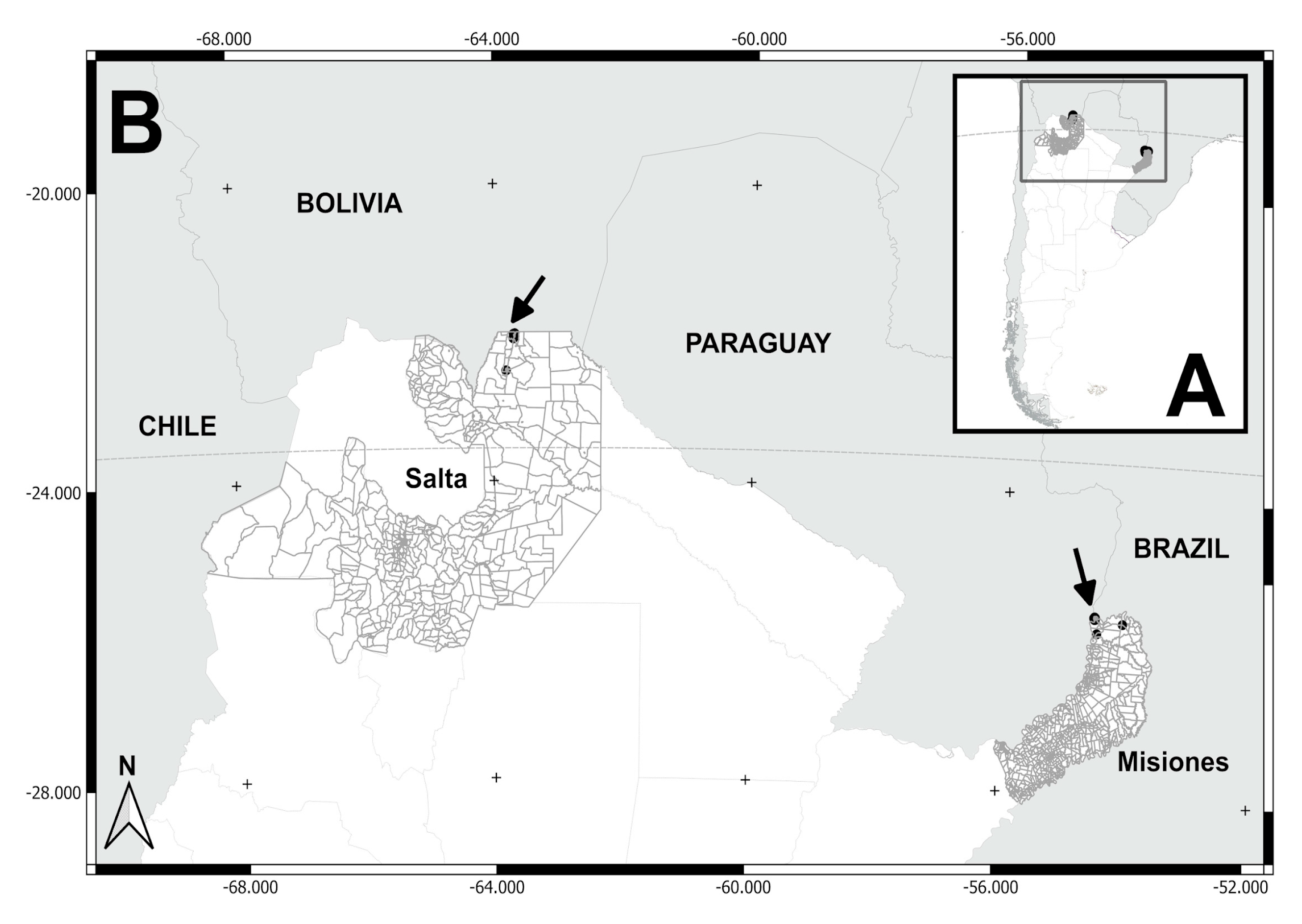
Figure 2.
(A) Localities sampled (Salvador Mazza and Tartagal) in San Martín department, Salta province. (B) Salvador Mazza locality: neighborhoods/settlements (1= La Bendición, 2= El Arenal, 3= Nueva Esperanza, 4= La Playa, 5= El Obraje, 6= El Chorro, 7= Barrio Arenales). Tartagal locality: neighborhoods/settlements sampled (8= Barrio Ferroviario, 9= Barrio Centro).
Figure 2.
(A) Localities sampled (Salvador Mazza and Tartagal) in San Martín department, Salta province. (B) Salvador Mazza locality: neighborhoods/settlements (1= La Bendición, 2= El Arenal, 3= Nueva Esperanza, 4= La Playa, 5= El Obraje, 6= El Chorro, 7= Barrio Arenales). Tartagal locality: neighborhoods/settlements sampled (8= Barrio Ferroviario, 9= Barrio Centro).
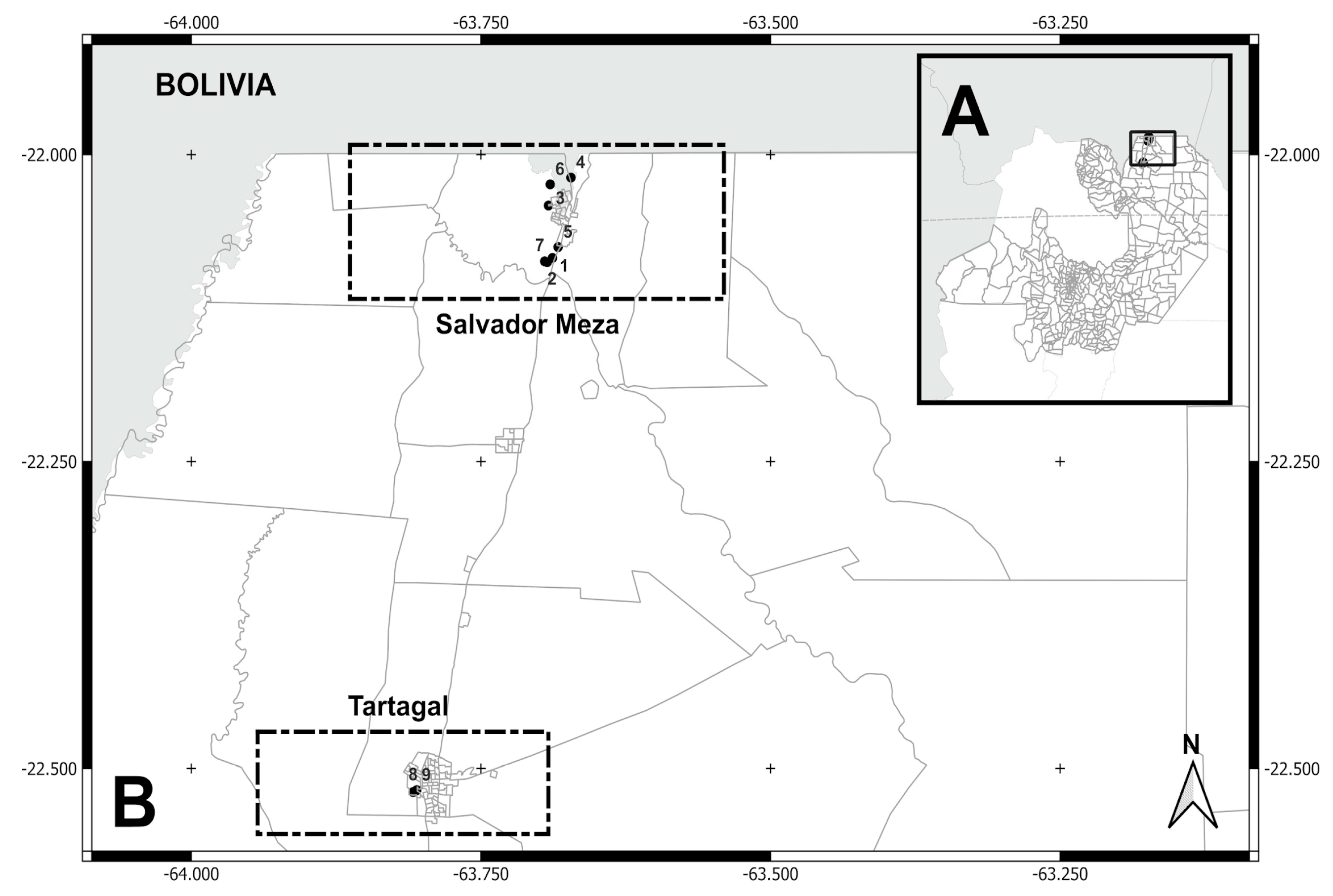
Figure 3.
(A) Localities sampled (Puerto Iguazú, Puerto Libertad, and Andrecito) in Puerto Iguazú department, Misiones province. (B) Puerto Iguazú locality: neighborhoods/settlements (10= 2000 Hectáreas, 11= Ribera del Paraná, 12= Barrio Alto Paraná, 13= Barrio Belén, 14= Barrio Obrero, 15= Barrio Santa Rosa, and 16= Barrio San Lucas). Puerto Libertad locality: neighborhoods/settlements (Barrio San Cayetano). Andrecito locality: neighborhoods/settlements (Barrio Selva 2 and Cabure-í).
Figure 3.
(A) Localities sampled (Puerto Iguazú, Puerto Libertad, and Andrecito) in Puerto Iguazú department, Misiones province. (B) Puerto Iguazú locality: neighborhoods/settlements (10= 2000 Hectáreas, 11= Ribera del Paraná, 12= Barrio Alto Paraná, 13= Barrio Belén, 14= Barrio Obrero, 15= Barrio Santa Rosa, and 16= Barrio San Lucas). Puerto Libertad locality: neighborhoods/settlements (Barrio San Cayetano). Andrecito locality: neighborhoods/settlements (Barrio Selva 2 and Cabure-í).
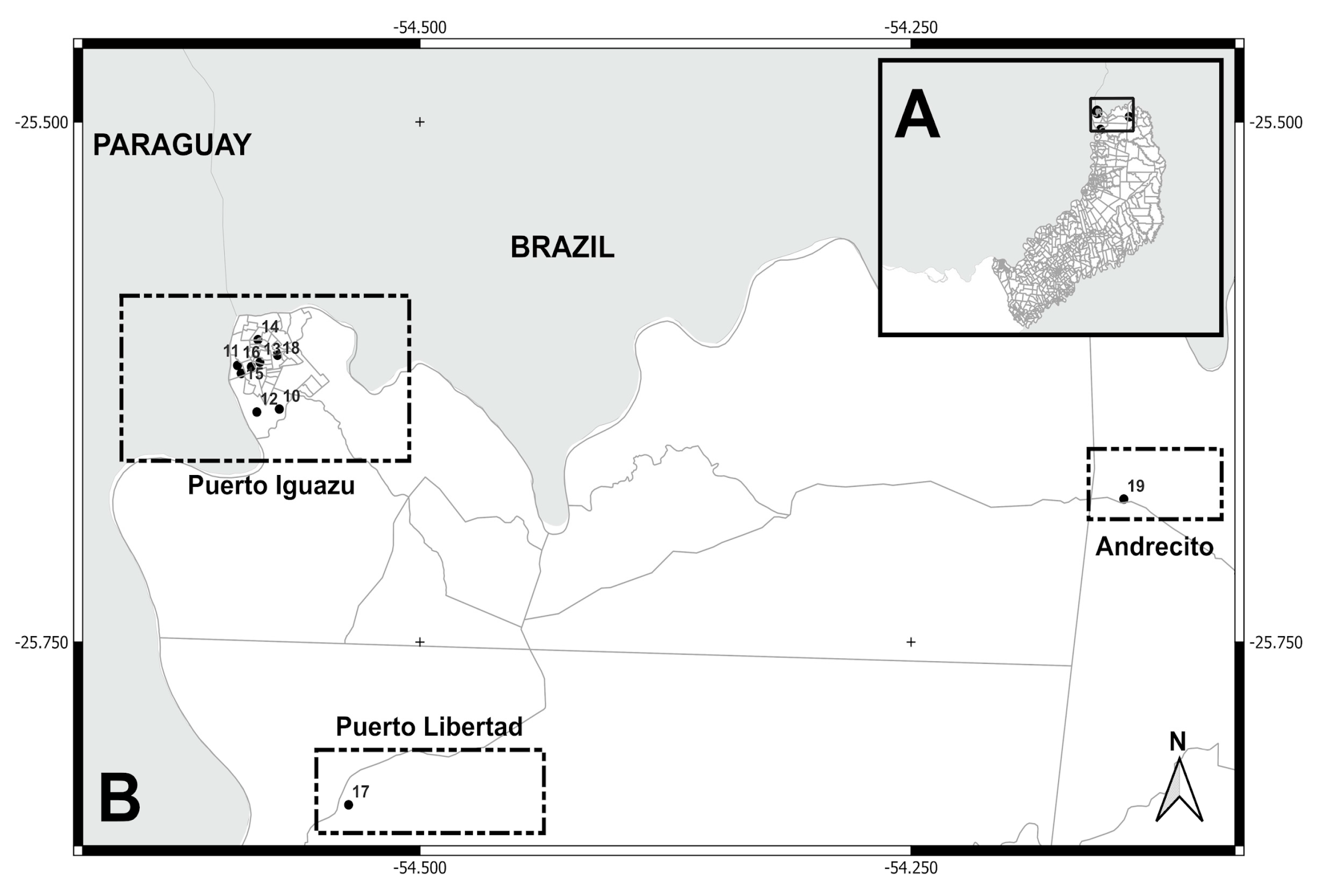
Figure 4.
Maximum likelihood phylogenetic tree of Plasmodium vivax based on Cytochrome b gene sequences. The tree was inferred using RAxML-NG under the GTR+G substitution model with 1,000 bootstrap replicates. Bootstrap support values ≥10 are shown on branches. The tree is rooted with three Plasmodium falciparum sequences (AJ276847 BRA, AY282957 ECU, and AY282971 COL). The Argentinian isolate (P. vivax PP658217 ARG) appears as a sister lineage to the main South American clade and shows closer affinity to sequences from Papua New Guinea. Tip labels include species name, GenBank accession number, and country of origin.
Figure 4.
Maximum likelihood phylogenetic tree of Plasmodium vivax based on Cytochrome b gene sequences. The tree was inferred using RAxML-NG under the GTR+G substitution model with 1,000 bootstrap replicates. Bootstrap support values ≥10 are shown on branches. The tree is rooted with three Plasmodium falciparum sequences (AJ276847 BRA, AY282957 ECU, and AY282971 COL). The Argentinian isolate (P. vivax PP658217 ARG) appears as a sister lineage to the main South American clade and shows closer affinity to sequences from Papua New Guinea. Tip labels include species name, GenBank accession number, and country of origin.
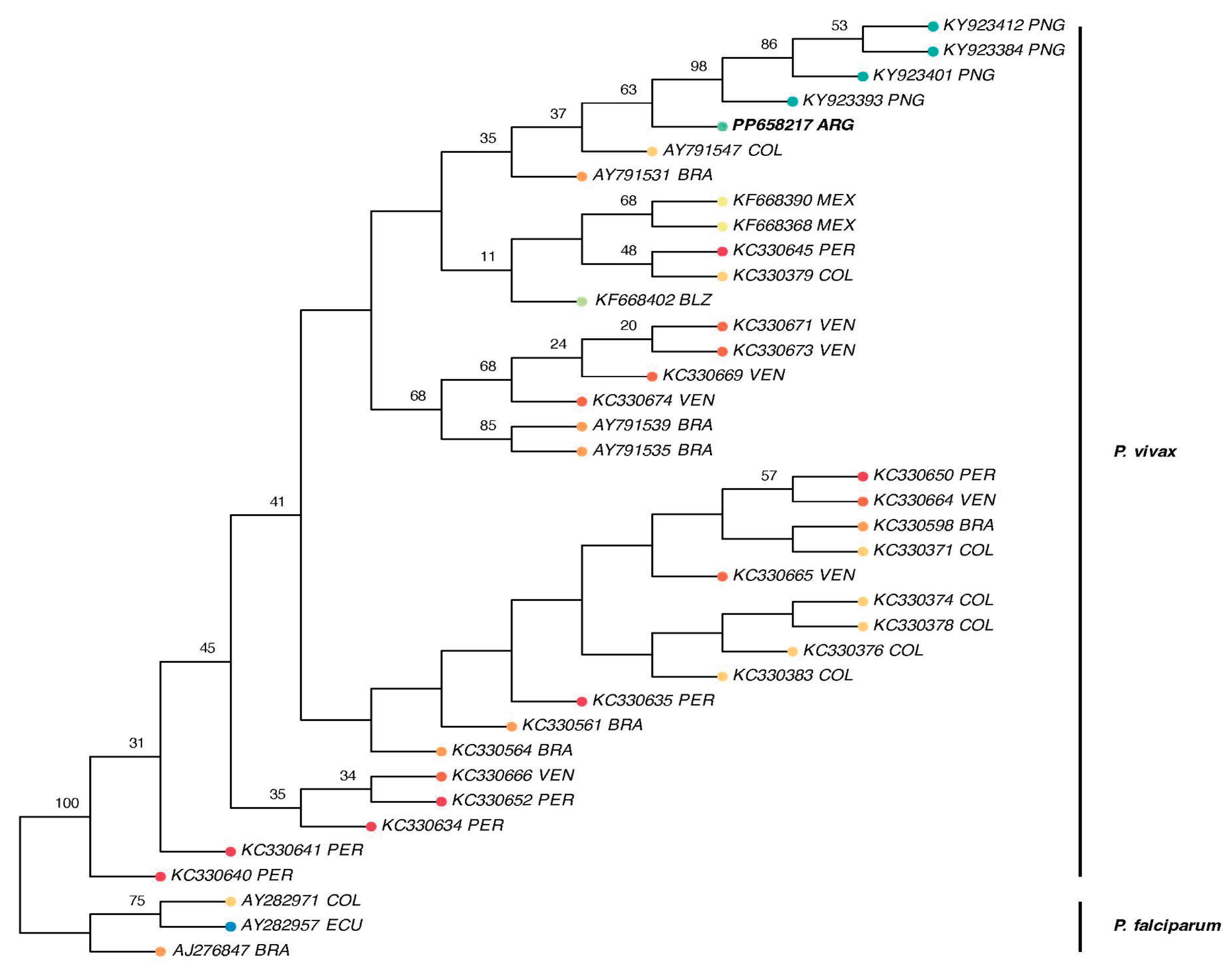

Table 1.
Localities of sample collection (biogeographical provinces and geographic coordinates), dates of collection, and number of samples processed in northern Argentina.
Table 1.
Localities of sample collection (biogeographical provinces and geographic coordinates), dates of collection, and number of samples processed in northern Argentina.
| ID | Neighborhoods, settlements / Localities | Biogeographical provinces | Latitude/longitude coordinates | Dates of collections | N samples | |
|---|---|---|---|---|---|---|
| Salta province | ||||||
| 1 | La Bendición / Salvador Mazza | Yungas ж | -22.08412, -63.68822 | November 2016 | 11 | |
| 2 | El Arenal / Salvador Mazza | -22.08693, -63.69492 | December 2013 | 3 | ||
| 3 | Nueva Esperanza/ Salvador Mazza | -22.04147, -63.69166 | December 2013 / ж 2015 | 9 | ||
| 4 | La Playa / Salvador Mazza | -22.01872, -63.67224 | December 2015 | 3 | ||
| 5 | El Obraje / Salvador Mazza | -22.07531, -63.68332 | December 2015 | 1 | ||
| 6 | El Chorro / Salvador Mazza | -22.02420, -63.69015 | December 2015 | 3 | ||
| 7 | Barrio Arenales / Salvador Mazza | -22.08756, -63.69273 | December 2015 | 11 | ||
| 8 | Barrio Ferroviario / Tartagal | -22.51909, -63.80849 | November 2016 | 1 | ||
| 9 | Barrio Centro / Tartagal | -22.51753, -63.80538 | November 2016 | 1 | ||
| ж Misiones province | ||||||
| 10 | 2000 Hectáreas / Puerto Iguazú | Paranaense ж | -25.63801, -54.57160 | August 2017 | 16 | |
| 11 | Ribera del Paraná / P. Iguazú | -25.61703, -54.59288 | October 2017 | 5 | ||
| 12 | Barrio Alto Paraná / P. Iguazú | -25.63943, -54.58304 | October 2017 | 2 | ||
| 13 | Barrio Belén / P. Iguazú | -25.61548, -54.58147 | October 2017 | 3 | ||
| 14 | Barrio Obrero / P. Iguazú | -25.60469, -54.58248 | October 2017 | 1 | ||
| 15 | Barrio Santa Rosa / P. Iguazú | -25.62087, -54.59100 | October 2017 | 6 | ||
| 16 | Barrio San Lucas / P. Iguazú | -25.61771, -54.58587 | October 2017 | 3 | ||
| 17 | Barrio San Cayetano / P. Libertad | -25.82811, -54.53633 | August 2017 | 4 | ||
| 18 | Barrio Selva 2 / Andrecito | -25.61210, -54.57252 | August 2017 | 3 | ||
| 19 | Caburé-i | -25.68127, -54.14172 | November 2017 | 6 | ||
Table 2.
GenBank accession numbers for 35 cyt-b Plasmodium vivax sequences used in the phylogenetic analysis.
Table 2.
GenBank accession numbers for 35 cyt-b Plasmodium vivax sequences used in the phylogenetic analysis.
| Country | Accession Numbers |
|---|---|
| Argentina | PP658217 |
| Belize | KF668402 |
| Brazil | AJ276847, AY791531, AY791535, AY791539, KC330561, KC330564, KC330598 |
| Colombia | AY282971, AY791547, KC330371, KC330378, KC330383, KC330374, KC330376, KC330379 |
| Ecuador | AY282957 |
| Mexico | KF668390, KF668368 |
| Papua New Guinea | KY923393, KY923401, KY923412, KY923384 |
| Peru | KC330635, KC330650, KC330652, KC330641, KC330640, KC330634, KC330645 |
| Venezuela | KC330665, KC330664, KC330666, KC330669, KC330671, KC330674, KC330673 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



