
Submitted:
07 August 2025
Posted:
08 August 2025
You are already at the latest version
Abstract
Electromagnetic field stimulation (EMFS) has emerged as a promising modality for regulating the structural dynamics of biological macromolecules implicated in aging and neurodegenerative disorders. In particular, EMFS can alter the conformation of abnor-mally folded proteins, such as amyloid-β (Aβ) peptides and their aggregates, thereby mitigating pathological protein aggregation and delaying neurodegeneration. The molecular interactions at play are governed by quantum mechanical forces acting on interfacial water molecules adjacent to hydrophilic protein surfaces. These interactions modulate the structural and functional dynamics of key proteins implicated in aging and neurodegeneration, including Heat Shock Factor 1 (HSF1) and amyloid-beta (Aβ).
This study advances the mechanistic understanding of water-mediated quantum effects, analogous to those observed in enzyme catalysis, highlighting the role of proton tunneling within extensive hydrogen-bond networks. These phenomena are especially prevalent in intrinsically disordered proteins (IDPs), which are central to proteostasis and highly susceptible to structural perturbations that lead to post-translational modifications associated with aging and neurodegeneration.
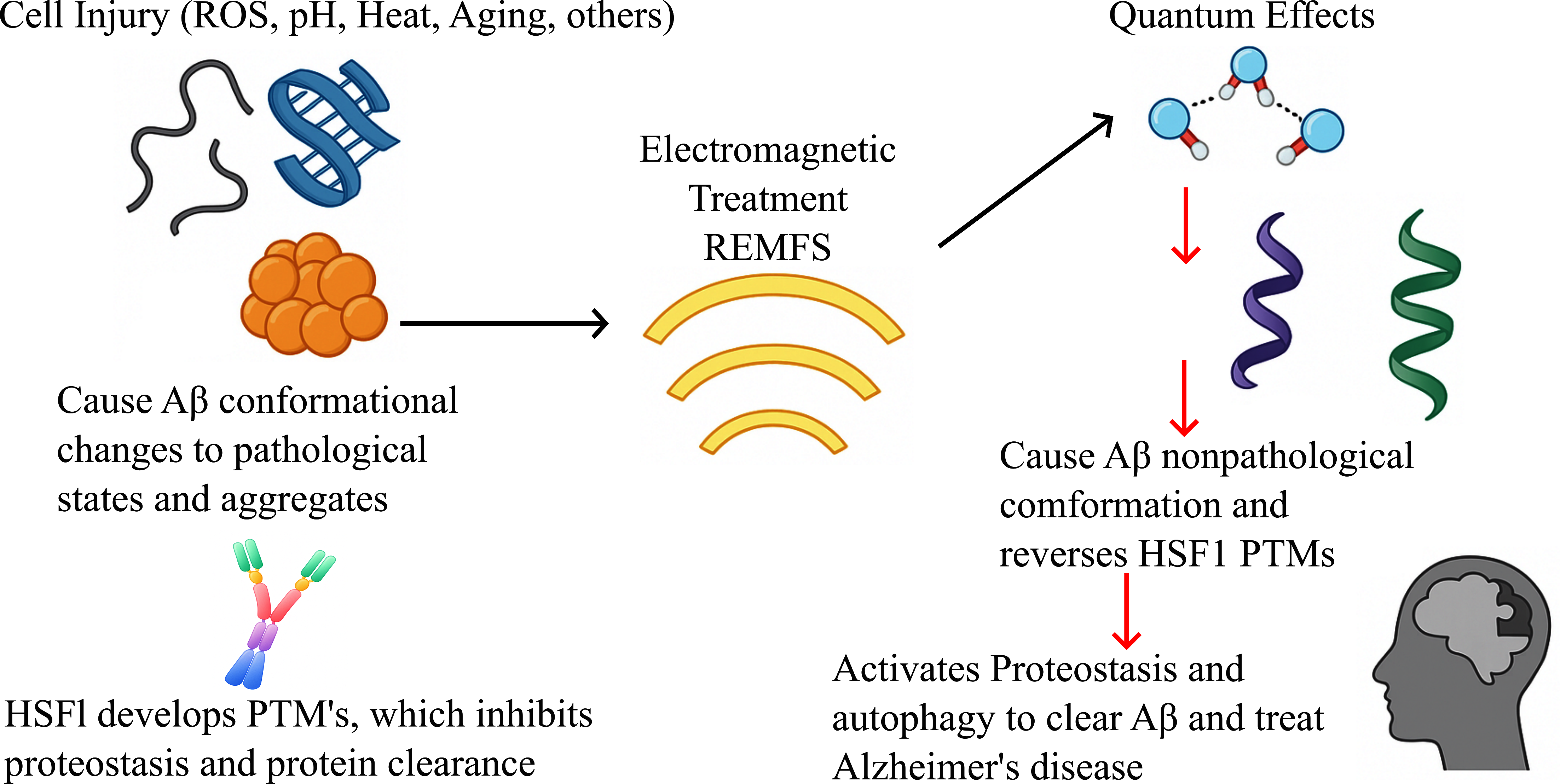
We propose repeated electromagnetic field stimulation (REMFS) as a novel thera-peutic strategy to restore the native conformational states of critical IDPs, including Aβ, α-synuclein, and Tau. Furthermore, REMFS appears to reverse the age-related post-translational modifications of key proteostasis regulators, such as HSF1 and other heat shock proteins. Through quantum-mechanical modulation, REMFS influences multiple Alzheimer’s disease (AD) pathways, demonstrating the potential to prevent or reverse pathological changes in cellular and organismal models. These findings support the further development of REMFS as an anti-amyloid and anti-aggregation therapy for AD and related neurodegenerative conditions.
Keywords:
aging
; Alzheimer’s disease
; treatment
; electromagnetic fields stimulation
; intrinsically disordered proteins
; quantum mechanics
; proteostasis
1. Introduction
Quantum EMF radiation involves photon particles as the fundamental carriers of discrete energy. The radiation can be described using quantum mechanics rather than Maxwell’s equations because the field’s energy is quantized into distinct photon units. Although photons are massless, they act as carriers of EM forces that interact with human tissues. Studying EM quantum and quantum biology may help us understand how energy transfers via photons in water-rich human tissues [1]. Quantum technology is already used in MRI machines, which interact more effectively with soft tissues to produce clearer images. This technology can potentially be expanded to address brain cell disorders such as Alzheimer's disease, serving as a future application. Research on quantum EM in the human brain could lead to new approaches for treating Alzheimer's [2]. It might also provide new insights into the role of amyloid fibrils, which are linked to the cause of Alzheimer’s disease [3].
The primary focus of the EMF quantum effects is on interfacial water, which was traditionally seen as a passive background for biochemical reactions. It is now increasingly understood as an active participant in biological processes, especially at the quantum level [4]. By emphasizing this parallel, we suggest that interfacial water regions have a catalytic role in cellular energy dynamics, facilitating proton transfer similar to enzyme active sites [5]. The key difference is that water’s “active site" isn’t a single pocket in a protein but a dynamic, quasi-two-dimensional sheet surrounding membranes, proteins, and nucleic acids. Protons might move through tunneling across this sheet from one site to another.
Proteins are not merely static structures; they are deeply entwined with their hydration shells. This relationship is especially pronounced in intrinsically disordered proteins (IDPs) [6]. Unlike folded proteins, IDPs lack a stable three-dimensional conformation and instead exist as dynamic ensembles of conformers. This structural plasticity is fundamental to their biological roles as molecular hubs in signaling networks, stress response, and proteome surveillance. Crucially, these roles are modulated not only by molecular interactions but also by the physical and quantum properties of the surrounding water. Structured water, particularly the interfacial form, exhibits semi-crystalline ordering [7], altered charge distribution, and capacity for electromagnetic coherence [8]. Recent advances in quantum biology suggest that interfacial water supports proton tunneling, energy resonance, and field-sensitive organization—all properties that profoundly impact protein-water interactions.
This paper examines the dynamic hydration of IDPs, considering recent developments in quantum mechanics. By integrating spectroscopy, molecular dynamics, and quantum electrodynamics studies, we propose a model in which hydration shells function as quantum-responsive buffers that regulate protein conformation, proteostasis, signaling, neurodegeneration, and longevity. We argue that oxidative stress during aging damages protein surfaces, disrupting hydrophilic zones and displacing ordered water. This causes the breakdown of these hydration structures, leading to post-translational modifications (PTM) during stress processes, which affect transcriptional factors and result in proteostasis collapse and protein deposition diseases such as AD. In this study, we also discuss potential therapeutic strategies such as repeated electromagnetic field stimulation (REFMS) to repair and reactivate proteins involved in proteostasis and autophagy for the treatment of age-related protein deposition diseases, including Alzheimer’s, frontotemporal disease, and Lewy body disease.
2. Intrinsically Disordered Proteins
IDPs are abundant in all cells and increase with the increase in the complexity of the organism [9]. These proteins are found more frequently in signaling proteins and proteins controlling gene activity, such as transcription factors [10]. Because of their amino acid sequences with high net charge and low hydrophobicity, IDPs do not follow conformational rules for ordered proteins with domains that lose structure and biological function at various environmental conditions, such as temperatures, pH, or EMFs; they gain ordered structure because of the enhancement of the hydrophobic interaction [11]. IDPs have a low content of bulky amino acids and a high proportion of polar and charged amino acids, which promotes hydrogen bonding between the polar atoms of charged residues and water molecules, improving interactions with interfacial water [12]. A lower amino acid alphabet characterizes IDPs compared to the alphabet utilized in the amino acid sequences of ordered domains and proteins [13] , and a higher sequence space than that of ordered proteins, which increases the vast structural complexity of these proteins and regions [13]. Also, because of their inability to fold into unique structures in isolation and their globally reduced structural content, parts of an IDP are disordered or ordered at different times and stimuli [14].
Therefore, IDPs are a complex mixture of foldable, partially foldable, potentially foldable, or non-foldable segments [15]. IDPs can have four different conformations depending on the environmental conditions, including folded or ordered, molten globule, pre-molten globule, and coil-like [16]. IDPs’ conformational plasticity predisposes them to their extreme sensitivity to changes in the environment and interaction with a multitude of proteins, and consequently to fold and bind in many different ways [17], that gives them unique properties such as control, regulate, interact with, as well as be controlled and regulated by multiple types of proteins and RNAs [18]. A multi-funnel structure characterizes IDPs’ free energy landscape, meaning it has various competing low-energy conformations rather than a single, stable, folded structure [19], which reflects the existence of numerous conformations that constitute its dynamic conformation, possessing a very disordered nature [20].
The shape of IDP energy is a multi-funnel-like landscape and can be easily altered by minimal variations in environmental factors, in contrast to the funnel-like energy landscape of an ordered protein, where the hilly plateau energy landscape of the IDP determines its conformational plasticity and its ability to fold differently depending on environmental factors. For this reason, IDPs can easily participate in cell regulation, signaling, and control, thereby serving as crucial regulators of various cellular processes [21]. All these characteristics predispose IDPs to develop posttranslational modifications (PTMs), which are chemical changes affecting proteins after molecular synthesis. PTMs can be covalently added to amino acids. IDPs use multiple mechanisms to modulate cell function, structural properties, and location of these proteins, including controlled degradation and posttranslational modifications [22].
PTMs induce many changes to IDPs’ structure. PTMs can covalently add chemical groups, carbohydrates, lipids, proteins, or nucleic acids to amino acid side chains of the IDPs [23]. IDPs can also undergo enzymatic cleavage of peptide bonds or removal of various chemical groups; some of these cleavage reactions produce toxic fragments, including aggregation-prone species, while others result in modified proteins with novel functions [24] and can give different functions to the same protein [25]. PTMs are readily reversible because of modifying and demodifying enzymes, and these modifications include phosphorylation [26], acetylation [27], glycosylation [28], lipidation [29], methylation [30], nitration [31], and protonation [32] , all of which give conformational variability of IDPs to control protein function [32]. A conspicuous member of IDPs is the peptide amyloid-β (Aβ) that aggregates into metal-enriched amyloid-forming plaques in Alzheimer’s disease [33]. Furthermore, genetics, oxidative and nitrative stress, as well as mitochondrial impairment, impact the structural flexibility of the unstructured IDPs to produce disease mechanisms [34]. In addition, simulations indicate that extended conformations are more likely to appear when hydrogen bonds are not stable, rather than when they are stable [35]. For example, IDPs like HSF1 and alpha-synuclein gain a more ordered structure with temperature rise and are more disordered at low temperatures [36]; they also fold in highly acidic or alkaline environments, folding into a more ordered conformation. This structural instability of IDPs/IDPRs is further supported by their well-known exceptional sensitivity to proteolytic degradation [37]. The poor regulation of IDPs is associated with various pathologies, including aging, cancer, neurodegeneration, cardiac disease, and diabetes [38].
3. Quantum Mechanisms in Structured Water
Cells are formed of 70% water, which is mostly bulk-like; however, a significant portion of intracellular water is interfacial, specifically a few nanometers of proteins, membranes, RNA, and DNA surfaces [39]. Structured water at biological interfaces (also known as exclusion zone (EZ) water) has demonstrated unusual behaviors such as long-range exclusion of solutes and charge separation [40]. Gerald Pollack and colleagues showed that hydrophilic surfaces can induce a several-hundred-micron-thick EZ region in water that is negatively charged and relatively ordered, expelling microspheres and solutes [41]. Notably, EZ water formation is accompanied by the release of protons into the surrounding bulk water, effectively creating a charge separation akin to a battery [40]. The behavior of this interfacial water cannot be explained by classical physics alone; for instance, infrared light dramatically expands the EZ, suggesting that electromagnetic fields directly interact with water’s structuring [42]. These observations have catalyzed new theoretical approaches, such as quantum electrodynamics (QED) models, that posit that water can form large-scale coherent domains with electromagnetic properties [43].
Experimental data obtained through ATR-IR spectroscopy, optical microscopy, and electrokinetic measurements confirm that water adjacent to hydrophilic interfaces organizes into highly ordered regions known as exclusion zones (EZs) or coherence domains. These zones exhibit semi-crystalline molecular alignment, a strong negative charge, and proton exclusion into the adjacent bulk water, creating a substantial electric potential of up to -200 mV across the domain[44]. This charge separation establishes an internal electric field that persists over micrometer distances, behaving similarly to a microscopic capacitor. The stability of these structures requires a continual energy input, primarily from infrared radiation, which has been shown experimentally to expand the size and stability of EZs[45]. The underlying electrostatic potential field can be modeled by the classical Poisson equation:
where φ is the electrostatic potential, ρ is the local charge density, and ε is the permittivity of the medium. This classical field description accurately captures the observed potential distribution within and around the EZ. Further experimental evidence supports that low-frequency electromagnetic fields (e.g., 50-75 Hz) can alter EZ structure in a non-thermal manner, consistent with resonant coupling to the hydrogen bond network [46]. Beyond classical descriptions, Del Giudice and Preparata proposed that coherence domains in water reflect a mesoscopic quantum electrodynamics (QED) phase, wherein water molecules oscillate in phase with a self-trapped electromagnetic field [47]. Although this theoretical model elegantly explains certain long-range ordering phenomena, it remains a theoretical extension beyond the directly observed experimental evidence. Thus, for biological integration, structured water’s experimentally validated electrostatic properties provide a sufficient foundation. However, experimental data obtained by ATR-IR spectroscopy are fully compatible with Quantum Electrodynamics (QED) predictions that induce a coherent state in the water [48]. It has shown that electromagnetic fields facilitate the interactions among non-polar molecules, and a density threshold with a critical temperature, also induce the appearance of a coherent state of the matter in that particles oscillate coherently between two quantum states giving rise to a long-range attraction among atoms or molecules that adds up to their electrostatic properties with net negative charge, excluding protons into the bulk. This charge separation creates an internal electric field and provides a medium for quantum coherence [49].
∇²φ = -ρ/ε
Furthermore, another study demonstrated that liquid water must be a two-phase system, that according to QED, part of the molecules are organized in collective coherent vibrational domains [50]. Their energy range corresponds to the spectral distance between the two energy levels at which the electron of each water molecule oscillates. The second phase comprises molecules set out of tune by thermal fluctuations that push components out of the coherent state. In addition, a meta-analysis showed a new order parameter characteristic for water molecule assembly, which implies quantum coherence and entanglement. This indicates water molecule assembly shows electromagnetic and electronic collective states that contain “quantum imprints or molds” because water molecules are ordered in a partially distorted tetrahedral geometry, which yields a specific network structure [51]. This water quantum coherence is stabilized by interaction with biomolecule surfaces [52] and posits that at room temperature, about 40% of cell water is in a coherent phase. Thus, every cell protein, DNA strand, or membrane is likely wrapped in a sheath of coherent water, where the water molecules are aligned and organized in a specific way rather than being randomly arranged like in regular water [53]. In RNA, the first layer of the interfacial water is considered a part of the nucleic acid structure because it defines structure, folding, and intramolecular interactions [54]. This water is a modulator of biological activity; many of the interesting properties of water arise from the hydrogen bonding network [55]. The key properties of this water in the cell include cooperative H-bonds, proton currents, osmosis, hydrostatic pressure, density variations, and the selective exclusion of ions. These changes make stronger and shorter H-bonds in the interfacial water with higher heat capacity than bulk water because more energy is necessary to break its H-bonds [56]. When considering the cell as a network of biomolecules, we must include their water shells; we have a network of articulate water domains pervading the cell and connecting at interface boundaries. The organization of the interfacial water depends on the type of bio-surface electrostatic forces [56]. The first layer of the interfacial water is in special quantum confinement since the polar and charged groups of the amino acids and nucleic acid interact with the surrounding H bonds [57] from the interfacial water. This interaction actively affects the structure, function, and H bonds of biomolecules [58]. Such polar and charged groups are sources of electric fields [59], which are very important for proton transfer [60]. This electrokinetic confined or trapped water exhibits a quantum tunneling behavior [61]. In structured hydration shells, similar tunneling might facilitate rapid proton translocation and charge redistribution [62], which is essential for signaling and energy transfer. Proton tunneling in this lattice mirrors the quantum mechanical effects seen in enzyme catalysis, where tunneling accelerates reaction rates by orders of magnitude [63].
Moreover, H-bonds oscillate collectively when these frequencies align with external EMFs, and coherent resonance is possible, enhancing water’s ability to store and transmit information [64]. This coherent water can profoundly affect biomolecular dynamics, particularly in intrinsically disordered proteins (IDPs) such as amyloid-β, α-synuclein, tau, and heat shock factor (HSF1). Because IDPs are highly reliant on stable, tightly bound hydration shells to maintain their flexible, functional ensembles, the disruption of the structured hydration layer can expose hydrophobic residues, facilitating pathogenic aggregation. Experimental work shows that around tau amyloid fibers, hydration water becomes significantly more mobile compared to soluble tau, favoring aggregation through an entropy-driven mechanism [65]. Similarly, simulations reveal that the expulsion of structured water upon protein misfolding contributes thermodynamically to amyloid assembly [66]. Thus, coherent, charge-separated water at the protein interface acts as a stabilizing matrix, suppressing aggregation and preserving proteostasis. In the context of aging and disease, loss of hydration coherence might tip this delicate balance, accelerating misfolding and dysfunction in critical IDPs such as amyloid-β, tau, α-synuclein, and HSF1 [67].
4. Water, Electromagnetic Fields, and Proton Tunneling
It is challenging to perform experiments with water under EMF because the picosecond lifetimes of H-bonds are too short for experimental techniques such as nuclear magnetic resonance (NMR) and dielectric spectroscopy time window [68]. However, in an EMF study, applying an intense THz pulse (peak electric field strength of 14.9 MV/cm) to liquid water increased H-bond stretching and bending vibrations, inducing proton tunneling [69]. Quantum tunneling is a phenomenon where particles pass through energy barriers that they would not be able to pass through classically. This occurs because of the probabilistic nature of quantum mechanics [70]. In the context of water molecules, quantum tunneling can influence the dynamics of hydrogen bonding and molecular rearrangements. Proton tunneling is a quantum mechanism highly relevant here. A proton in a hydrogen bond can tunnel through the energy barrier between two equilibrium positions (for example, between a donor and acceptor oxygen in adjacent water molecules). In bulk water, this underlies the Grotthuss mechanism, giving rise to anomalously high proton mobility (proton conductivity) [71]. In structured water at interfaces, several factors can enhance tunneling probability. First, the strong ordering and shorter hydrogen-bond distances in EZ water (with tightly aligned molecules) lower the proton transfer barriers [72]. External perturbations – such as an oscillating electromagnetic field – can further modulate the hydrogen bond length, potential energy landscape, and quantum tunneling [73].
Similar evidence that polarized EMF radiation causes its biological effects comes from the fact that it induces the dissociation of water into its constituent elements [74]. Rao examined distilled water under a polarized 2.45 GHz exposure. The Raman spectra of the treated water showed significant changes in the O–H stretch bond, which predisposes to proton tunneling and protonation of the surrounding molecules. Interestingly, despite different experimental conditions such as frequency (THz versus Hz), most of the conclusions are consistent with the fact that very different EMFs can produce water dissociation [75]. Furthermore, EMF exposures strengthen and shorten the H-bonds on the surface of biomolecules. For example, when hemoglobin and bovine serum albumin in water solutions were exposed to 50 Hz, samples revealed a significant increase in the absorbance signal of the Amide II band and an up-shift toward the high energies after exposure. These results suggested that EMF exposures strengthened the H-bonds of the secondary structures of these proteins [76]. Although EMF exposure increases the rotational kinetic energy of bulk water, the effects on the water of the first layer surrounding biomolecules are most prominent. The water of the first layer in the vicinity of biomolecules has a forced orientation and cannot rotate easily. However, under EMF, it can undergo large-amplitude librational motions [77]. EMF oscillations at the molecular level produce rotation of water molecules within the first layer of the interfacial water as they try to "flip" their polar directions to match the polarity of the radio wave radiation. As a result, the oscillating electric field from the EMF forces the water dipole moments to reorient themselves, which affects the H-bond that connects the first layer of the interfacial water [78].
This effect can be observed in biological tissues, where all polar molecules, such as water, are forced to oscillate in phase with the field and on planes parallel to its polarization [79]This is one of the most important factors for the quantum effects of REMFS: the man-made polarization of the excitatory oscillation of REMFS on contact with the interfacial water of the bio-system. These oscillations have a lower frequency relative to the exposed quantum system. They will change the frequency of the system to the excitatory frequency [80] like a driven harmonic oscillator[81]. The system is driven by energy imparted upon the harmonic oscillator continuously by an external force [82]. If the excitatory frequencies are slower, the oscillator frequency is pulled towards the excitatory frequency [82]. EMF decreases the water H-bond length and increases the H-bond angles as a function of the large amplitude motions [83]. These large amplitude vibrations decrease the H-bond length of the first layer of the interfacial water to the oxygen of the nucleic acid (O−H···O) to values below 1.85 Å [84], which are ideal lengths for raising the probability of proton tunneling [85]. This effect depends on the spatial location of the molecule concerning the field [86], affecting the H-Bond Network [87] in an anisotropic manner. Thus, the new configuration changes the hydrogen-bonding energy and distance as the front water molecule is rotated [88] and hydrogen pairs with a net dipole moment. This water reorientation is important in multiple quantum processes, including proton transfer [89], proton transport [90], and hydration of RNA or IDPs for their function [91].
The protonation state of amino acids, particularly those with titratable side chains (like histidine, lysine, cysteine, and aspartic acid), can change with EMF exposures. This alters the protein's overall charge and can affect its stability and interactions with other molecules, causing posttranslational modifications [92], as charge is a major driving force for many molecular interactions. Protonation can lead to conformational changes in IDPs because of the impact of electrostatic interactions on their dynamics, affecting their function [93]. The dynamic nature of IDPs means that they can exist in multiple tautomeric forms, which are structural isomers that differ by the position of a proton and a double bond [94]. In addition, several studies have found that EMF causes proton tunneling to produce tautomers in nucleic acids. Cerón-Carrasco found that electric fields induce proton transfer, which produces tautomers in nucleic acids using Quantum Mechanical (QM) calculations [95]. The electric field decreased the potential barrier leading to the tautomer by 20–55 kJ mol−1. The study concluded that in the presence of EFs, only Guanine-Cytosine fit the necessary kinetic criteria to be considered a viable route to the formation of tautomers. In a more recent study, Cerón-Carrasco investigated a more accurate DNA fragment in a simulation. The study found that at higher electric fields, tautomers are more stable than canonical bases [96]. In a classical molecular dynamics study, Cerón-Carrasco found that a continuous electric field exposure produces conformational changes in nucleic acids in 10 picoseconds [97]. These tautomers [98] are used by multiple RNAs to produce their functions [99]. Tautomer inter-conversions can adopt various secondary structures responsible for a variety of functions during biological processes like RNA conformational changes, DNA replication, packaging, and transcription [100,101]. Often, such conformational changes promote binding to activating factors that in turn affect transcription and translation of proteins.
5. IDPs and Hydration Water Dynamics
Water dynamics around IDPs play key roles in protein function, including enzyme activity and allostery. IDPs are characterized by high conformational entropy and a preponderance of charged and polar residues. IDPs show higher hydration water density as compared to the globular protein, and the water molecules are tetrahedrally ordered (more local structural order) around disordered regions/proteins as compared to ordered regions/globular protein because of higher mean net charge, promoting stronger water-protein interactions in disordered regions/proteins [102]. This composition attracts extensive hydration shells, resulting in bound water layers that exhibit restricted dynamics, at least in part due to the greater presence of charged groups in disordered regions [103]. A large fraction of IDPs contains positively and negatively charged residues, depending on the content of charged residues. This is reflected in a hilly energy landscape, where the highs are barriers to forbidden conformations [104]. The distribution of charges and their sequence likely determine the response to environmental changes, so disordered parts respond differently to similar stimuli [105]. These disordered parts play vital roles in protein aggregation, participate in protein-protein interactions, and undergo binding-induced folding [106]. Another crucial factor for the biological activity of proteins is the internal dynamics, which provides some degree of conformational flexibility and structural dynamics required for catalytic activity [106]. These structured hydration layers are not passive—they interact with both endogenous and external electromagnetic fields, modulating coherence within and across protein complexes [107].
These hydration shells stabilize disordered regions and act as mediators of protein-protein interactions, allostery, and phase separation. Reid et al. showed that negatively charged residues within IDRs form long-lived hydrogen bonds with water molecules, reducing water mobility compared to structured protein surfaces [6]. This behavior suggests a strong coupling between electrostatics and hydration dynamics. IDPs’ function depends on their interactions with other proteins; it needs tight coupling of experimental and computational methods because they do not have a single conformation, and their models require ensembles of conformations representing a distribution of states that the protein adopts in solution [108]. Studies using molecular dynamics simulations and dielectric spectroscopy indicate that water molecules around IDPs exhibit more heterogeneous rotational and translational dynamics than water around structured proteins [109]. This means that water molecules around IDPs are, on average, more restricted in their movement than water around structured proteins. This restricted motion is attributed to a more significant fraction of tightly bound water molecules in the first hydration shell around IDPs. This electrokinetic confined or trapped water exhibits a quantum tunneling behavior [110] linked to signaling and regulatory processes [111]. We propose a conceptual model in which intrinsically disordered proteins (IDPs), structured water, and quantum coherence operate as a highly integrated signaling system. IDPs are enveloped by hydration shells composed of ordered, polarized water that exhibit hydrogen bond resonance and facilitate proton tunneling. IDPs are also involved in stress granules, nucleoli, and other condensates that undergo hydration-modulated transitions from fluid to gel to solid states—progressions associated with aging and neurodegeneration [38]. These hydrogels [112] are composed of uniformly polymerized amyloid-like fibers [113].
6. Age-Related Breakdown of Hydration Coherence in IDPs
Intrinsically disordered proteins (IDPs) are characterized by high conformational variability, which gives them flexible hubs involved in the signaling and regulation of many biological processes, including the aging process [114]. A recent protein-protein interaction network analysis showed that IDPs are found in different clusters associated with several aging hallmarks [115], including genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, and stem cell exhaustion [116]. This underscores their critical involvement in both healthy aging and the pathology of neurodegenerative diseases, including Alzheimer's, Parkinson's, Huntington's, ALS, frontotemporal dementia, as well as cancer, diabetes, and cardiovascular disease [115]. IDPs contribute to disease through several mechanisms, including misfolding and aggregation, altered signaling, and disrupted protein interactions [117]. Some examples of IDPs in disease are Amyloid-beta, which is a key protein involved in Alzheimer's disease, Tau protein, which is also implicated in Alzheimer's and other neurodegenerative disorders, α-Synuclein is a protein associated with Parkinson's disease and Lewy body dementia, and p53 and BRCA1 associated with cancer [118]. Quantum coherence might play a role in the folding process of IDPs, allowing them to explore multiple conformations simultaneously before settling into a stable structure [119]. While an intuitive picture for classical coherence is a recurring pattern, quantum mechanical coherence is exemplified by superposition states [120], which are generated by the interaction of protons and heteronuclei [121]. A study with 1H–15N heteronuclear single quantum coherence (HSQC) spectrum of the intrinsically disordered C-terminal domain of a virus nucleoprotein showed low signal dispersion in the 1H dimension, that would normally result in an increased spectral overlap; however, this low signal dispersion is to some extent counterbalanced by the narrowed line widths of the NMR signals of IDPs because of their inherent dynamic nature [122]. As coherent water structures decay, so does the efficiency of proton tunneling, electron transfer, and protein dynamics [123]. IDPs, heavily reliant on hydration-mediated interactions, lose quantum coherence and begin to misfold and aggregate [124]. In the unfolding and refolding of the IDPs, coherence could be a component of the protein's ability to follow “ a pathway” in a directed process, countering perturbing actions by molecules not involved in the process [124].
Quantum coherence drives us to untangle mechanisms and dynamics controlled by order and synchronization at a quantum mechanical level to enhance functions or develop new molecular systems [125]. For example, a-synuclein structures occur at 30–42% in the trimeric and tetrameric forms and 58–70% for the monomeric species by Bayesian analysis because of rapid interconversions, suggesting unfolding and refolding of monomeric and multimeric species on timescales faster than tens of microseconds. The monomeric structures predominate in the NMR spectrum because of their enhanced flexibility compared to their multimeric counterparts [122]. This is supported by a study using 1H–15N heteronuclear single quantum coherence (HSQC) spectrum in that the proposed tetrameric species superimposes within the experimental error on the spectrum of monomeric a-synuclein from E. coli [126].
Also, oxidative stress during aging damages protein surfaces, disrupting interfacial water and IDPs quantum coherence, diminishing the cell’s ability to sustain structured hydration [127]. Coherence here refers to the degree to which an IDP's different functions or interactions are interconnected or depend on each other. For example, an IDP might have multiple binding sites or be involved in different signaling pathways that are linked or regulated together [128]. A study found that all the proteins associated with oxidative stress-induced JNK signaling and cell death were IDPs with molecular recognition features (MoRFs), posttranslational modification (PTM) sites, and short linear motifs (SLiMs) that were associated with the disordered regions [129]. Furthermore, cytoskeletal breakdown alters intracellular architecture, compromising the geometric stability of hydration domains [130].
An important IDP involved in the aging process and proteostasis is the transcriptional factor HSF1, which is a key regulator of heat shock proteins, contains extensive IDRs that respond to hydration state, temperature, and multiple PTMs, including redox status [131]. Its dysfunction under stress and age, because of PTMs, correlates with proteostasis collapse in neurodegeneration [132]. Other IDPs involved in neurodegeneration are Aβ, Tau, alpha-synuclein, and TDP-43 [133].
7. Expanded Insights into HSF1 Regulation via Hydration-Sensitive Mechanisms
PTMs enable proteins to rapidly switch between different structural and functional states in response to internal and external stimuli. These modifications occur more frequently in IDPs because they provide access to modifying and disorganizing enzymes and facilitate interactions with protein modules [135] that specifically recognize and bind to modified residues [136], which is a result of the abundance of disordered regions in proteomes. PTMs in IDPs happen more often than three to four modifications per protein [137]. Additionally, RNA-binding proteins (RBPs) like HSF1 are significantly more enriched in disorder compared to the human proteome [138].
HSF1, a master regulator of proteostasis, is profoundly influenced by hydration dynamics. Its activation involves trimerization, nuclear translocation, and DNA binding [139]—each modulated by IDRs sensitive to redox shifts, temperature, and water structuring. HSF1 undergoes phase separation upon heat shock, and its activity is regulated by post-translational modifications (PTMs), the molecular details underlying HSF1 phase separation, temperature sensing, and PTM regulation. HSF1 surfaces at hydrophobic regions restrict water molecules, thereby limiting the degrees of freedom and decreasing entropy. When proteins undergo phase separation, some associated water molecules are released from the protein because of the decrease in protein surface area, resulting in positive entropy gain [140]. Recent findings reveal that Heat Shock RNA-1 (HSR1), a long non-coding RNA, facilitates HSF1 activation by promoting this trimerization [141]. Because RNA structure and interaction affinity are hydration-dependent, HSR1 adds a hydration-sensitive layer to HSF1 regulation. PTM sites that modulate HSF1 activity are not primarily located in leucine zipper (LZ) regions but are instead concentrated in the disordered regulatory domain (RD) [132,142]. HSF1’s heptad repeats (HR-A/B/C) are IDR-rich and contain disorder-to-order transitions triggered by thermal stress [111]. HR-C unfolds at elevated temperatures, enabling trimerization via HR-A. These conformational shifts are fine-tuned by post-translational modifications (PTMs) such as acetylation and phosphorylation, which occur predominantly at hydration-exposed sites that are sensitive to redox state and surface exposure, particularly within intrinsically disordered regions [143,144]. Oxidative stress accelerates PTMs like CK2-mediated phosphorylation and SIRT1 inhibition, destabilizing HSF1 [145]. SIRT1 inhibition destabilizes HSF1 by impairing its DNA binding ability, leading to a reduced heat shock response and potentially impacting cellular stress responses and aging [146]. This dysregulation is linked to age-related diseases. For instance, Huntington’s disease involves hyperacetylation of HSF1 and impaired mitochondrial function [146]. HSF1 normally activates PGC1α, a coactivator promoting mitochondrial biogenesis and thermogenesis [147]. As SIRT1 and PGC1α levels decline with age, hydration coherence erodes, further weakening HSF1 functionality [148]. Altogether, this positions HSF1 as a hydration-sensitive quantum responder. It integrates redox signals, thermal fluctuations, and water structuring to maintain proteostasis—an ability compromised during aging and neurodegeneration. However, with aging, these hydration-based networks deteriorate, impairing both classical signaling pathways and quantum coherence mechanisms, ultimately contributing to the loss of proteostasis and cellular resilience.
8. EMF Studies in Cell Cultures and Animals
8.1. Studies in Cell Cultures
8.1.1. Studies in Animal and Human Immortalized Cell Cultures
A literature review [149] investigated the relationship between EMF and AD at the cellular level. It showed that some have a beneficial effect of EMF exposure on AD manifestations at the cellular level. On the contrary, some studies found no relationship with AD. These discrepancies in the results may be due to differences in the frequency, power deposition, exposure period of EMF, differences in the animal model, cell type [150], and specific absorption rate (SAR). Regulatory agencies recommend using SAR measurements for safety and RF biological effects[151]. For example, short-term EMF exposures activated the Aβ clearance pathway, such as the chaperone-mediated autophagy pathway in human neuroblastoma SH-SY5Y cells, with maximum effect between 30-60 minutes of EMF exposure[152]. The ubiquitin-proteasome pathway was activated in primary hippocampal rat neurons exposed to 100 mT for 15 minutes[153]. In another study, HT22 mouse hippocampal neuronal cells and SH-SY5Y human neuroblastoma cells were exposed to 1950 MHz [154] with a high-power specific absorption rate (SAR) of 6 W/kg for two hours per day for 3 days, the levels of APP, Aβ precursor protein cleaving enzyme 1, disintegrin metalloproteinase 10, and Presenilin-1 were not significantly different between EMF exposed culture and controls exposures. Remarkably, this researcher previously found that a SAR of 5 W/kg was beneficial for AD pathology in rats, suggesting an upper limit for the beneficial effects on AD pathology not higher than a SAR of 5 W//kg [155].
On the other hand, when IMR-32 neuroblastoma cells were exposed to 60 Hz at 50, 100, and 200 μT for four hours [156], there were no changes in the expression of APP695, an isoform of APP. In a study to assess the expression of proteins involved in the AD pathology (α3, α5, and α7 nicotinic acetylcholine receptors) in SHSY5Y human neuroblastoma cells exposed to EMF, there was no change in expression after exposure to extremely-low frequency EMF [157]. In addition, 50 Hz at 3.1 mT for 18 hours induced Aβ 1–42 secretion[158], and another study showed increased production of prostaglandin E2 and decreased phagocytosis of fibrillary Aβ 42[159]. All these studies demonstrate the potential harmful effects of EMF during prolonged exposure (>2 hours).
8.1.2. Primary Human Cultures
In our previous study, our lab utilized primary culture because it is directly extracted from human tissue and grown in a laboratory, maintaining its natural characteristics. On the other hand, an immortalized cell line is a genetically modified cell population that can divide indefinitely in culture, often derived from a tumor and therefore different from the natural cell function[160,161]. Previously, we found that repeated electromagnetic field stimulation (REFMS) at 50 MHz, exposure times of 5, 15, 30, 60, and 120 min, power of 0.5 W, and a SAR of 0.6 W/kg activated the HSF[162] (master regulator of proteostasis[161] and the autophagy proteins ATG5 and ATG12[152,163]) in primary human fibroblasts. Given that the age-related attenuation of HSF1[139,164] plays a central role in the process of abnormal autophagy[165] that occurs during aging, it is suggested that EMF interventions to push HSF1 toward its activated state are essential for the activation of autophagy and the clearance of abnormal proteins such as Aβ in age-related diseases[166,167]. This prompted us to examine REMFS effects on the Aβ levels in primary human brain cultures.
To determine an appropriate EMF dose (dosimetry) [168], we examined the positive and negative effects of REMFS treatments on memory and Alzheimer's disease (AD) pathology across multiple studies. We used the inverted U-shaped dose-effect curve (IUSDEC) for this purpose [169]. Initially, we reviewed literature [170,171,172] from cell culture [152,162,173,174,175,176,177,178], animal [179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194], and human [195,196,197,198,199] studies before conducting our human brain culture experiments. We found that RF power deposition resulting in a SAR between 0.25 and 5 W/kg improves AD pathology and memory. Conversely, SAR levels below 0.25 W/kg or above 5 W/kg [154,156,159,200,201,202,203,204,205,206,207,208,209,210,211] either had no effect or were harmful to AD pathology and cognition, indicating an IUSDEC. Two human studies support this optimal SAR range: one observed impaired cognitive tasks [212]peed at SAR values below 0.2 W/kg and above 5 W/kg, while accuracy improved within this range [213]. Additionally, longer exposures (5 hours) caused demyelination in mouse neurons [214], whereas shorter exposures (< 30 minutes) had no effect [150,152], suggesting a dose- and time-dependent relationship. Following ICNIRP [151] and IEEE [215] safety guidelines (2 W/kg local head exposure), and considering differences in size, tissue properties, exposure duration, and thermal physiology of animals- as well as the fact that neurons can be harmed even when global SAR stays within safety limits [151] we adjusted the SAR limits for our human brain culture experiments to a maximum of 0. 9 W/kg and a minimum of 0. 4 W/kg. We then exposed primary human mixed brain cultures (PHB) to various EMF frequencies, exposure durations, daily schedules, and SARs [173] to assess the safety and effectiveness of REMFS on human neurons. REMFS treatment reduced Aβ-40 and Aβ-42 levels without signs of toxicity. Starting treatment on day 7 in vitro (DIV 7), we measured Aβ-40 over 14 days. Initially, we used a frequency of 64 MHz with a SAR of 0. 6 W/kg for one hour daily; this resulted in a 46% reduction in Aβ-40 compared to untreated cultures. Later, we demonstrated that REMFS at 64 MHz with a SAR of 0. 4 W/kg over 14 days produced comparable reductions in Aβ-40 and Aβ-42 levels. Increasing exposure duration from 1 to 2 hours yielded a similar decrease in Aβ levels, establishing the maximum effective exposure time for REMFS. We also identified 0.4 W/kg as the minimum SAR needed to lower Aβ levels, suggesting that the SAR range of 0.4 to 0.9 W/kg could be both effective and safe for human studies.
8.2. Preliminary Findings from Animal Studies
Numerous studies investigated the relationship between EMF and AD in animal models; most of these are listed in recent reviews [170,171,216]. These take into account the various molecular biological mechanisms of AD that have been studied in various animal models, including transgenic AD mouse models and C. elegans. These models are explained in detail in the Ribeiro et al. paper [217]. For example, an AD mouse model exposed to REMFS at 918 MHz with a SAR of 0.25 w/kg for two hours a day for 6 months of treatment showed an improvement in Aβ deposition and cognitive function [182]. Also, REMFS at 918 MHz in the AD mouse model produces an improvement in brain mitochondrial function and an increase in soluble Aβ1-40 following a daily two-hour exposure for a month[181]. An EMF study at 50 Hz, 10 mT for 14 days in AD rats showed improved learning and memory performance [191]. A REMFS at 1950 MHz and SAR of 5W/kg for 2 h/day in 5xFAD mice and controls showed a significant reduction in Aβ plaques, APP, and CTFs in the hippocampus and entorhinal cortex [155]. A long-term REMFS study reduced hyperactivity and anxiety symptoms while improving memory and increased glucose metabolism[218]. Another REMFS at 918 MHz study in human and primary rat astrocytes decreased Aβ levels, reactive oxygen species (ROS), H2O2–induced phosphorylation of p38MAPK and ERK1/2, and mitochondrial ROS, while increasing MMP[174]. REMFS at 1950 MHz [219] and a SAR of 5 W/kg repressed microgliosis genes (Csf1r, CD68, and Ccl6), pro-inflammatory cytokine interleukin-1β. and microglial function genes, including Trem2, Fcgr1a, Ctss, and Spi1 in in 5×FAD mice, suggesting that REMFS has beneficial effects in AD pathology and cognition in AD models. Several more recent REMFS studies [220,221,222,223,224,225,226,227] and systematic reviews [228,229,230] in rodents confirmed the beneficial effects in AD.
On the contrary, prolonged exposure to continuous electromagnetic pulse at 100 Hz and a very high electric field (50 kV/m), for 8 months in Sprague Dawley led to cognitive and memory impairment, increased Aβ level, increased expression of Aβ oligomer and APP, and increased expression of Tau, suggesting a continuous high electric field exposure can cause harmful effects and increase AD pathology [202]. Also, another study at 100 Hz and a very high electric field (50 kV/m), for 8 months in Sprague Dawley, showed an increase in Aβ, BACE1, Tau, and APP in the hippocampus, and cognitive impairment [231]. Another study at 50 Hz and a low magnetic field of 100 μT in Sprague Dawley rats [232] for 12 weeks produced no effects on cognitive function and Aβ level changes, suggesting that a low magnetic field does not have biological effects. Another 15-minute REMFS exposure at 900 MHz and a higher SAR of 6 W/kg exposure showed that RF-EMF did not affect cognition in AD or control rats[233]. REMFS at 1950 MHz and SAR 5 W/kg 2 h/d for 3 months did not improve cognition in AD mice [234], suggesting that a treatment course of 3 months does not affect AD pathology, but longer courses improve cognition and AD pathology [188,219].
9. Discussion and Implications for Quantum Biology and Therapeutics
Here we present a hypothesis that reveals a novel domain of quantum biology: the control of the IDPs conformation via coherent water under electromagnetic fields. Through its influence on structured hydration shells, electromagnetic field exposure may promote quantum coherence [235] and trigger hormetic effects that delay aging and neurodegeneration [236]. EMF exposure can cause protonation [33], acetylation [237], phosphorylation [238], decrease reactive oxygen species [239], and potentially reverse PTMs in IDPs. In support of this, EMF exposure induces HSF1 activation in senescent human and rodent cells [162] and delays and reverses cellular senescence [162]. This indicates EMF exposures reverse the HSF1 PTMs that cause the age-related attenuation of the heat shock response and the proteostasis [240], which prevents binding to DNA in senescent cells. EMF exposures also lower Aβ protein aggregation in old rodents [179], supporting that EMF is also effective in older organisms. Furthermore, in a recent EMF study, AML12 and HEK293 cells were exposed to 10 µT for three hours, these exposures increased of HSP70 and HSP90, that was followed by an enhanced of HSP70 and HSP90 for HSP70/HSP90-organizing protein (HOP/STIP1) [237], suggesting that EMF enhances proteostasis because the acetylation of these HSPs reduced protein aggregates and increased cell viability and protein folding [241].
Various studies have examined the effects of EMFs on ROS, which are an important cause of PTMs in IDPs that affect aging and age-related diseases. A study found that EMF exposure at 7 MHz decreased ROS concentrations. They proposed that the ROS production in metabolic processes occurs through singlet-triplet modulation of semiquinone flavin (FADH•) enzymes and superoxide spin-correlated radical pairs. Spin-radical pair products are modulated by the 7 MHz RF magnetic fields that presumably decouple flavin hyperfine interactions during spin coherence [242]. A study found that oscillating magnetic fields alter relative yields of cellular superoxide and hydrogen peroxide, indicating coherent singlet-triplet mixing at the point of ROS formation [127]. This model reframes water not as a background solvent, but as an active participant in cellular information processing. It is noteworthy that the induction of ROS and signs of oxidative stress appear to be more common in neurons exposed to higher SAR and more to ELF-MF than to RF-EMF [243,244,245,246,247]. On the other hand, EMFs and pathological IDPs like Aβ and Tau have a complex relationship, especially in the context of Alzheimer's disease (AD) [248]. Several studies have shown that EMF exposure affects Aβ conformation, disrupts existing Aβ fibrils and aggregates, and prevents neuronal dysfunction. EMF at 0.25 V nm-1 causes a conformational shift of Aβ from a pathological β to a non-pathological non-β shape[249], implying that Aβ can become non-toxic with specific therapeutic strategies. Another EMF exposure at 900 MHz for 3 hours led to a significant reduction in Aβ fibrils in vitro, confirming the simulation results and supporting the idea that EMFs influence Aβ conformation changes [250]. The use of static electric field strengths ranging from 0 V/nm to 0.5 V/nm on an Aβ1-42 supramolecular assembly consisting of a pentamer and a tetramer [251]suggests that these fields induce a permanent or temporary dipole. This dipole causes a conformational change in the protein structure due to the mechanical stress exerted on the charged peptides.
A study investigated the stability of the minimal toxic species, i.e., β-amyloid dimers, in the presence of an oscillating electric field [252]. The simulation results provide evidence that an external electric field (oscillating at 1 GHz) can disrupt amyloid oligomers, which suggests a possible therapeutic strategy for AD [253]. An EMF study at 70 mV/nm-1GHz found that the diffusion of Aβ oligomers in the membrane decreased, and their interaction with the membrane was also reduced during exposure. Furthermore, at the cellular level, repeated electromagnetic field stimulation (REMFS) lowers Aβ levels and prevents the formation of oligomers in primary human brain cultures [173]. The proposed mechanism for these effects appears to be proton tunneling caused by the forced vibration from these exposures [150]. A recent study found that magnetoelectric dissociation of Alzheimer’s β-amyloid aggregates occurs in ex vivo brain tissues of an AD mouse model [254]. At the organismal level, EMF inhibits Aβ amyloid fibril formation both in vitro and in vivo, alleviating cognitive impairment in Alzheimer’s disease mice [225]. A recent study using 40 Hz gamma frequency showed a significant decrease in Aβ secretion, phosphorylation of AKT, mTOR, and tau, and Aβ aggregation in SH-SY5Y cells [255]. In an interesting study of Tau protein, a neuroblastoma neuronal cell line was exposed to 4 h of zinc intoxication, which causes Tau-microtubule dissociation. Cel cultures were previously treated with ELF-EMF at 40 Hz and 1 G (with multiple exposure schedules), the showed enhanced microtubule dynamics and increased Tau-microtubule interaction in the face of zinc toxicity, it showed Tau accentuated phosphorylation and reduction in beta tubulin isotypes, depending on electromagnetic frequencies, most pronounced at 3.9 Hz, suggesting that ELF-EMF modulation on the microtubule cytoskeleton essential for microtubule repair [256].
On the contrary, some molecular dynamics simulations suggest that EMF exposures can shift Aβ from a helical to a beta-sheet conformation under nonionizing radiation of varying EMFs [257]. It was shown that the EMFs induce peptide conformations dependent on the field frequency and strength, suggesting that specific ranges of EM field parameters produce well-ordered peptide conformations and modulate the formation of amyloid fibrils. An externally applied electric field (EF) of varying strengths (0.1-1 V/nm) using molecular dynamics (MD) caused Aβ aggregation, showing that the EF favors the switch of Aβ-peptides from helical to beta-sheet conformation [258]. These results suggest that EMF parameters can be selected as a rational design strategy for AD treatment. All the above suggest that targeting hydration dynamics—via EM field [249], we can modulate IDPs such as HSF1, Aβ, Tau, and other proteins to restore coherence to a non-pathological conformation, consequently delaying proteostasis collapse.
Another dimension worth exploring is the potential impact of electromagnetic harmonics, integer multiples of a fundamental EM frequency, on IDPs. Harmonic components naturally arise in non-linear biological environments and resonate with specific vibrational or electronic modes within IDPs, potentially enhancing or disrupting folding pathways through quantum resonance or interference effects [259,260]. Such interactions may alter tunneling probabilities or coherence lifetimes, thereby influencing the conformational landscape of these proteins. Understanding the biological sensitivity to not only primary EM frequencies but also their harmonic content could lead to more precise therapeutic targeting and tuning of REMFS protocols [261].
Additionally, a crucial factor is the type of EMF polarization [80], which can be either linear or circular. Circularly polarized electromagnetic fields rotate their electric field vector as they propagate, either right-handed or left-handed (chirality). This EMF chirality interacts especially with chiral molecules like proteins and DNA [262,263,264], which could be adjusted to left or right-handed polarization depending on the chirality of the biological target.
10. Conclusion
IDPs exhibit exceptional sensitivity to their hydration environments, which, under structured conditions, may exhibit quantum-coherent properties. Aging and environmental stressors—such as oxidative damage and posttranslational modifications (PTMs)—can destabilize this finely tuned protein-water interface. Such disruptions may contribute to hallmark features of aging and neurodegeneration by impairing the dynamic conformational landscape of IDPs. This shift can lead to a loss of PTM-mediated functional plasticity, promote pathological aggregation, or hinder essential protein-DNA interactions, as observed in age-related dysfunction of HSF1 and increased receptor binding of Aβ.
In this paper, we propose a quantum biological framework for a novel therapeutic strategy (REMFS) that can influence structured water, proton tunneling, and charge separation. This strategy aims to restore the conformational flexibility of IDPs to a physiologically stable, non-aggregated conformation in multiple proteins, such as Aβ, tau, α-synuclein, and Tau, which are central to neurodegenerative disorders. Furthermore, REMFS reverses PTMs that impair the normal function of IDPs, such as HSF1 and autophagy proteins, resulting in improved protein degradation, delayed aging [167], and potentially serving as an anti-amyloid therapy for human AD and neurodegenerative diseases [265]. By stabilizing hydration shells and reestablishing quantum coherence, EMFs may act as biophysical modulators of proteostasis. This insight introduces a paradigm shift: hydration shells are not passive solvents but active quantum interfaces that govern protein behavior. A recently developed human REMFS (repeated electromagnetic field stimulation) device offers a translational pathway for clinical application. Future iterations should incorporate harmonic frequency tuning and spatial field shaping to enable targeted modulation of IDPs based on their unique biophysical signatures. Such precision could enhance therapeutic efficacy while minimizing off-target effects.
Ultimately, this approach may enable fine-tuned control over protein conformation and signaling pathways by targeting structured water scaffolds. A novel design and manufacture of an appropriate human REMFS device was recently introduced as a potential pathway for clinical translation [265]. Building on this foundation, future iterations should consider the integration of harmonic frequency controls and field shaping capabilities, enabling targeted stimulation based on the specific biophysical properties of IDPs. This could enhance selectivity and therapeutic efficacy while minimizing off-target effects [266,267]. This may someday fine-tune protein conformation and molecular pathway activation by targeting the structured water scaffolds that will make it possible to prevent and reverse AD, and other neurodegenerative diseases associated with protein aggregation, such as Parkinson’s, Lewy body, Frontotemporal, and Multisystem Atrophy.
Author Contributions
Conceptualization, F.P., J.B., H.K., J.J., and M.R.; methodology, F.P.; software, H.K.; writing—original draft preparation, F.P., J.B., H.K., J.J., and M.R.; writing—review and editing, H.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the reported results are available on reasonable request to the first author.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kinsey, L.J.; Beane, W.S.; Tseng, K.A.-S. Accelerating an integrative view of quantum biology. Front. Physiol. 2024, 14, 1349013. [Google Scholar] [CrossRef]
- Patwa, H.; Babcock, N.S.; Kurian, P. Quantum-enhanced photoprotection in neuroprotein architectures emerges from collective light-matter interactions. Front. Phys. 2024, 12, 1387271. [Google Scholar] [CrossRef]
- M. Otten et al. arXiv:2406.18744, 2024.
- C. Messori, S. V. C. Messori, S. V. Prinzera, and F. B. di Bardone, "Deep into the water: exploring the hydro-electromagnetic and quantum-electrodynamic properties of interfacial water in living systems," Open Access Library Journal, vol. 6, no. 05, p. 1, 2019.
- Chen, L.; Zhang, S.; Liu, X.; Ge, X. Recent advances in water-mediated multiphase catalysis. Curr. Opin. Colloid Interface Sci. 2023, 65. [Google Scholar] [CrossRef]
- Reid, K.M.; Singh, A.K.; Bikash, C.R.; Wei, J.; Tal-Gan, Y.; Vinh, N.Q.; Leitner, D.M. The origin and impact of bound water around intrinsically disordered proteins. Biophys. J. 2022, 121, 540–551. [Google Scholar] [CrossRef]
- Assaker, K.; Carteret, C.; Lebeau, B.; Marichal, C.; Vidal, L.; Stébé, M.-J.; Blin, J.-L. Water-Catalyzed Low-Temperature Transformation from Amorphous to Semi-Crystalline Phase of Ordered Mesoporous Titania Framework. ACS Sustain. Chem. Eng. 2013, 2, 120–125. [Google Scholar] [CrossRef]
- Del Giudice, E.; Tedeschi, A.; Vitiello, G.; Voeikov, V. Coherent structures in liquid water close to hydrophilic surfaces.CONFERENCE NAME, LOCATION OF CONFERENCE, COUNTRYDATE OF CONFERENCE; p. 012028.
- Peng, Z.; Yan, J.; Fan, X.; Mizianty, M.J.; Xue, B.; Wang, K.; Hu, G.; Uversky, V.N.; Kurgan, L. Exceptionally abundant exceptions: comprehensive characterization of intrinsic disorder in all domains of life. Cell Mol. Life Sci. 2014, 72, 137–151. [Google Scholar] [CrossRef]
- Stavropoulos, I.; Khaldi, N.; Davey, N.E.; O’brien, K.; Martin, F.; Shields, D.C.; Kim, P.M. Protein Disorder and Short Conserved Motifs in Disordered Regions Are Enriched near the Cytoplasmic Side of Single-Pass Transmembrane Proteins. PLOS ONE 2012, 7, e44389. [Google Scholar] [CrossRef] [PubMed]
- V. N. Uversky, "Intrinsically disordered proteins and their environment: effects of strong denaturants, temperature, pH, counter ions, membranes, binding partners, osmolytes, and macromolecular crowding," The protein journal, vol. 28, pp. 305-325, 2009.
- V. N. Uversky, "Intrinsically disordered proteins from A to Z," The international journal of biochemistry & cell biology, vol. 43, no. 8, pp. 1090-1103, 2011.
- Uversky, V.N.; Li, J.; Fink, A.L. Evidence for a Partially Folded Intermediate in α-Synuclein Fibril Formation. J. Biol. Chem. 2001, 276, 10737–10744. [Google Scholar] [CrossRef]
- Ruff, K.M.; Roberts, S.; Chilkoti, A.; Pappu, R.V. Advances in Understanding Stimulus-Responsive Phase Behavior of Intrinsically Disordered Protein Polymers. J. Mol. Biol. 2018, 430, 4619–4635. [Google Scholar] [CrossRef]
- Fonin, A.V.; Darling, A.L.; Kuznetsova, I.M.; Turoverov, K.K.; Uversky, V.N. Intrinsically disordered proteins in crowded milieu: when chaos prevails within the cellular gumbo. Cell. Mol. Life Sci. 2018, 75, 3907–3929. [Google Scholar] [CrossRef]
- Turoverov, K.K.; Kuznetsova, I.M.; Uversky, V.N. The protein kingdom extended: Ordered and intrinsically disordered proteins, their folding, supramolecular complex formation, and aggregation. Prog. Biophys. Mol. Biol. 2010, 102, 73–84. [Google Scholar] [CrossRef]
- Dogan, J.; Gianni, S.; Jemth, P. The binding mechanisms of intrinsically disordered proteins. Phys. Chem. Chem. Phys. 2013, 16, 6323–6331. [Google Scholar] [CrossRef]
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 2014, 16, 18–29. [Google Scholar] [CrossRef]
- Fisher, C.K.; Stultz, C.M. Constructing ensembles for intrinsically disordered proteins. Curr. Opin. Struct. Biol. 2011, 21, 426–431. [Google Scholar] [CrossRef] [PubMed]
- V. N. Uversky, "Unusual biophysics of intrinsically disordered proteins," Biochimica et Biophysica Acta (BBA)-Proteins and Proteomics, vol. 1834, no. 5, pp. 932-951, 2013.
- R. Van Der Lee et al., "Classification of intrinsically disordered regions and proteins," Chemical reviews, vol. 114, no. 13, pp. 6589-6631, 2014.
- Darling, A.L.; Uversky, V.N. Intrinsic Disorder and Posttranslational Modifications: The Darker Side of the Biological Dark Matter. Front. Genet. 2018, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Mann, M.; Jensen, O.N. Proteomic analysis of post-translational modifications. Nat. Biotechnol. 2003, 21, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- P. R. Jungblut, H. G. P. R. Jungblut, H. G. Holzhütter, R. Apweiler, and H. Schlüter, "The speciation of the proteome," Chemistry Central Journal, vol. 2, pp. 1-10, 2008.
- Kulkarni, P.; Jolly, M.K.; Jia, D.; Mooney, S.M.; Bhargava, A.; Kagohara, L.T.; Chen, Y.; Hao, P.; He, Y.; Veltri, R.W.; et al. Phosphorylation-induced conformational dynamics in an intrinsically disordered protein and potential role in phenotypic heterogeneity. Proc. Natl. Acad. Sci. 2017, 114, E2644–E2653. [Google Scholar] [CrossRef]
- Saito, M.; Hess, D.; Eglinger, J.; Fritsch, A.W.; Kreysing, M.; Weinert, B.T.; Choudhary, C.; Matthias, P. Acetylation of intrinsically disordered regions regulates phase separation. Nat. Chem. Biol. 2018, 15, 51–61. [Google Scholar] [CrossRef]
- Rahman, M.M.; Zamakhaeva, S.; Rush, J.S.; Chaton, C.T.; Kenner, C.W.; Hla, Y.M.; Tsui, H.-C.T.; Uversky, V.N.; Winkler, M.E.; Korotkov, K.V.; et al. Glycosylation of serine/threonine-rich intrinsically disordered regions of membrane-associated proteins in streptococci. Nat. Commun. 2025, 16, 1–20. [Google Scholar] [CrossRef]
- Zhang, Z.; Ji, J.; Hossain, S.; Bailey, B.; Nangia, S.; Mozhdehi, D. Lipidation alters the phase-separation of resilin-like polypeptides. Soft Matter 2024, 20, 4007–4014. [Google Scholar] [CrossRef]
- Bauer, V.; Schmidtgall, B.; Gógl, G.; Dolenc, J.; Osz, J.; Nominé, Y.; Kostmann, C.; Cousido-Siah, A.; Mitschler, A.; Rochel, N.; et al. Conformational editing of intrinsically disordered protein by α-methylation. [CrossRef]
- Uversky, V.N.; Yamin, G.; Munishkina, L.A.; Karymov, M.A.; Millett, I.S.; Doniach, S.; Lyubchenko, Y.L.; Fink, A.L. Effects of nitration on the structure and aggregation of α-synuclein. Mol. Brain Res. 2005, 134, 84–102. [Google Scholar] [CrossRef]
- Geist, L.; Henen, M.A.; Haiderer, S.; Schwarz, T.C.; Kurzbach, D.; Zawadzka-Kazimierczuk, A.; Saxena, S.; Żerko, S.; Koźmiński, W.; Hinderberger, D.; et al. Protonation-dependent conformational variability of intrinsically disordered proteins. Protein Sci. 2013, 22, 1196–1205. [Google Scholar] [CrossRef]
- Feng, J.; She, Y.; Li, C.; Shen, L. Metal ion mediated aggregation of Alzheimer's disease peptides and proteins in solutions and at surfaces. Adv. Colloid Interface Sci. 2023, 320, 103009. [Google Scholar] [CrossRef]
- Wise-Scira, O.; Dunn, A.; Aloglu, A.K.; Sakallioglu, I.T.; Coskuner, O. Structures of the E46K Mutant-Type α-Synuclein Protein and Impact of E46K Mutation on the Structures of the Wild-Type α-Synuclein Protein. ACS Chem. Neurosci. 2013, 4, 498–508. [Google Scholar] [CrossRef]
- Levine, Z.A.; Larini, L.; LaPointe, N.E.; Feinstein, S.C.; Shea, J.-E. Regulation and aggregation of intrinsically disordered peptides. Proc. Natl. Acad. Sci. 2015, 112, 2758–2763. [Google Scholar] [CrossRef] [PubMed]
- M. Kjaergaard, A. B. M. Kjaergaard, A. B. Nørholm, R. Hendus‒Altenburger, S. F. Pedersen, F. M. Poulsen, and B. B. Kragelund, "Temperature-dependent structural changes in intrinsically disordered proteins: Formation of α‒helices or loss of polyproline II?," Protein Science, vol. 19, no. 8, pp. 1555-1564, 2010.
- M. Vidović and S. Komić Milić, "Regulation of proteolysis of intrinsically disordered proteins: Physiological consequences," A closer look at proteolysis, pp. 1-46, 2021.
- Uversky, V.N.; Oldfield, C.J.; Dunker, A.K. Intrinsically Disordered Proteins in Human Diseases: Introducing the D2Concept. Annu. Rev. Biophys. 2008, 37, 215–246. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Morita, S.; Hayashi, T. Role of interfacial water in determining the interactions of proteins and cells with hydrated materials. Colloids Surfaces B: Biointerfaces 2021, 198, 111449. [Google Scholar] [CrossRef]
- Seneff, S.; Kyriakopoulos, A.M. Taurine prevents mitochondrial dysfunction and protects mitochondria from reactive oxygen species and deuterium toxicity. Amino Acids 2025, 57, 1–15. [Google Scholar] [CrossRef]
- Figueroa, X.A.; Pollack, G.H. Exclusion-zone formation from discontinuous nafion surfaces. Int. J. Des. Nat. Ecodynamics 2011, 6, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Elton, D.C.; Spencer, P.D.; Riches, J.D.; Williams, E.D. Exclusion Zone Phenomena in Water—A Critical Review of Experimental Findings and Theories. Int. J. Mol. Sci. 2020, 21, 5041. [Google Scholar] [CrossRef]
- L. Lin, W. L. Lin, W. Jiang, X. Xu, and P. Xu, "A critical review of the application of electromagnetic fields for scaling control in water systems: mechanisms, characterization, and operation," NPJ Clean Water, vol. 3, no. 1, p. 25, 2020.
- Chai, B.; Mahtani, A.G.; Pollack, G.H. UNEXPECTED PRESENCE OF SOLUTE-FREE ZONES AT METAL-WATER INTERFACES. Contemp. Mater. 2012, 1, 1–12. [Google Scholar] [CrossRef]
- Chai, B.; Yoo, H.; Pollack, G.H. Effect of Radiant Energy on Near-Surface Water. J. Phys. Chem. B 2009, 113, 13953–13958. [Google Scholar] [CrossRef]
- Rad, I.; Stahlberg, R.; Kung, K.; Pollack, G.H.; Chin, W.-C. Low frequency weak electric fields can induce structural changes in water. PLOS ONE 2021, 16, e0260967. [Google Scholar] [CrossRef]
- Del Giudice, E.; Preparata, G.; Vitiello, G. Water as a Free Electric Dipole Laser. Phys. Rev. Lett. 1988, 61, 1085–1088. [Google Scholar] [CrossRef] [PubMed]
- A. De Ninno, E. A. De Ninno, E. Del Giudice, L. Gamberale, and A. C. I: Castellano, "The structure of liquid water emerging from the vibrational spectroscopy; arXiv:1310.0635, 2013.
- Pollack, G. The Fourth Phase of Water: A role in fascia? J. Bodyw. Mov. Ther. 2013, 17, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Taschin, A.; Bartolini, P.; Eramo, R.; Righini, R.; Torre, R. Evidence of two distinct local structures of water from ambient to supercooled conditions. Nat. Commun. 2013, 4, 2401. [Google Scholar] [CrossRef] [PubMed]
- H. J. Geesink, I. H. J. Geesink, I. Jerman, and D. K. Meijer, "Water, the cradle of life via its coherent quantum frequencies," Water, vol. 11, pp. 78-108, 2020.
- Ho, M.-W. Illuminating water and life: Emilio Del Giudice. Electromagn. Biol. Med. 2015, 34, 113–122. [Google Scholar] [CrossRef]
- M. C. Wahl and M. Sundaralingam, "C H… O hydrogen bonding in biology," Trends in biochemical sciences, vol. 22, no. 3, pp. 97-102, 1997.
- G. Otting, E. G. Otting, E. Liepinsh, and K. Wuthrich, "Protein hydration in aqueous solution," Science, vol. 254, no. 5034, pp. 974-980, 1991.
- D. E. Moilanen, I. R. D. E. Moilanen, I. R. Piletic, and M. D. Fayer, "Water dynamics in Nafion fuel cell membranes: The effects of confinement and structural changes on the hydrogen bond network," The Journal of Physical Chemistry C, vol. 111, no. 25, pp. 8884-8891, 2007.
- Mentré, P. Interfacial water: a modulator of biological activity. J. Biol. Phys. Chem. 2004, 4, 115–123. [Google Scholar] [CrossRef]
- Arunan, E.; Desiraju, G.R.; Klein, R.A.; Sadlej, J.; Scheiner, S.; Alkorta, I.; Clary, D.C.; Crabtree, R.H.; Dannenberg, J.J.; Hobza, P.; et al. Defining the hydrogen bond: An account (IUPAC Technical Report). Pure Appl. Chem. 2011, 83, 1619–1636. [Google Scholar] [CrossRef]
- Ball, P. Water as an Active Constituent in Cell Biology. Chem. Rev. 2007, 108, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.K.; Zewail, A.H. Dynamics of Water in Biological Recognition. Chem. Rev. 2004, 104, 2099–2124. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Zhang, C.; Weichselbaum, E.; Knyazev, D.G.; Pohl, P.; Carloni, P.; Johnson, C. Interfacial water molecules at biological membranes: Structural features and role for lateral proton diffusion. PLOS ONE 2018, 13, e0193454. [Google Scholar] [CrossRef] [PubMed]
- Bothma, J.P.; Gilmore, J.B.; McKenzie, R.H. The role of quantum effects in proton transfer reactions in enzymes: quantum tunneling in a noisy environment? New J. Phys. 2010, 12. [Google Scholar] [CrossRef]
- Meng, X.; Guo, J.; Peng, J.; Chen, J.; Wang, Z.; Shi, J.-R.; Li, X.-Z.; Wang, E.-G.; Jiang, Y. Direct visualization of concerted proton tunnelling in a water nanocluster. Nat. Phys. 2015, 11, 235–239. [Google Scholar] [CrossRef]
- R. K. Allemann and N. S. Scrutton, Quantum tunnelling in enzyme-catalysed reactions. Royal Society of Chemistry, 2009.
- del Giudice, E.; Doglia, S.; Milani, M.; Vitiello, G. Electromagnetic field and spontaneous symmetry breaking in biological matter. Nucl. Phys. B 1986, 275, 185–199. [Google Scholar] [CrossRef]
- Fichou, Y.; Schirò, G.; Gallat, F.-X.; Laguri, C.; Moulin, M.; Combet, J.; Zamponi, M.; Härtlein, M.; Picart, C.; Mossou, E.; et al. Hydration water mobility is enhanced around tau amyloid fibers. Proc. Natl. Acad. Sci. 2015, 112, 6365–6370. [Google Scholar] [CrossRef]
- Camino, J.D.; Gracia, P.; Cremades, N. The role of water in the primary nucleation of protein amyloid aggregation. Biophys. Chem. 2021, 269, 106520. [Google Scholar] [CrossRef]
- Bellissent-Funel, M.-C.; Hassanali, A.; Havenith, M.; Henchman, R.; Pohl, P.; Sterpone, F.; van der Spoel, D.; Xu, Y.; E Garcia, A. Water Determines the Structure and Dynamics of Proteins. Chem. Rev. 2016, 116, 7673–7697. [Google Scholar] [CrossRef]
- Elgabarty, H.; Kaliannan, N.K.; Kühne, T.D. Enhancement of the local asymmetry in the hydrogen bond network of liquid water by an ultrafast electric field pulse. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Zhao, H.; Tan, Y.; Zhang, L.; Zhang, R.; Shalaby, M.; Zhang, C.; Zhao, Y.; Zhang, X.-C. Ultrafast hydrogen bond dynamics of liquid water revealed by terahertz-induced transient birefringence. Light. Sci. Appl. 2020, 9, 1–10. [Google Scholar] [CrossRef]
- Faupin, J.; Fröhlich, J.; Schubnel, B. On the Probabilistic Nature of Quantum Mechanics and the Notion of Closed Systems. Ann. Henri Poincare 2015, 17, 689–731. [Google Scholar] [CrossRef]
- Day, T.J.F.; Schmitt, U.W.; Voth, G.A. The Mechanism of Hydrated Proton Transport in Water. J. Am. Chem. Soc. 2000, 122, 12027–12028. [Google Scholar] [CrossRef]
- B. J. Siwick and H. J. Bakker, "On the role of water in intermolecular proton-transfer reactions," Journal of the American Chemical Society, vol. 129, no. 44, pp. 13412-13420, 2007.
- J. Odutola, T. J. Odutola, T. Hu, D. Prinslow, S. O’dell, and T. Dyke, "Water dimer tunneling states with K= 0," The Journal of chemical physics, vol. 88, no. 9, pp. 5352-5361, 1988.
- M. L. Rao, S. R. M. L. Rao, S. R. Sedlmayr, R. Roy, and J. Kanzius, "Polarized microwave and RF radiation effects on the structure and stability of liquid water," Curr. Sci, vol. 98, no. 11, pp. 1500-1504, 2010.
- Vallée, P.; Lafait, J.; Legrand, L.; Mentré, P.; Monod, M.-O.; Thomas, Y. Effects of Pulsed Low-Frequency Electromagnetic Fields on Water Characterized by Light Scattering Techniques: Role of Bubbles. Langmuir 2005, 21, 2293–2299. [Google Scholar] [CrossRef]
- Calabrò, E.; Magazù, S. Response of hydrogen bonding to low-intensity 50 Hz electromagnetic field in typical proteins in bi-distilled water solution. Spectrosc. Lett. 2017, 50, 330–335. [Google Scholar] [CrossRef]
- James, D.; Armishaw, R. Structure of aqueous solutions: Infrared spectra of the water librational mode in solutions of monovalent halides. Aust. J. Chem. 1975, 28, 1179–1186. [Google Scholar] [CrossRef]
- English, N.J.; MacElroy, J.M.D. Molecular dynamics simulations of microwave heating of water. J. Chem. Phys. 2003, 118, 1589–1592. [Google Scholar] [CrossRef]
- Panagopoulos, D.J.; Messini, N.; Karabarbounis, A.; Philippetis, A.L.; Margaritis, L.H. A Mechanism for Action of Oscillating Electric Fields on Cells. Biochem. Biophys. Res. Commun. 2000, 272, 634–640. [Google Scholar] [CrossRef]
- Panagopoulos, D.J.; Johansson, O.; Carlo, G.L. Polarization: A Key Difference between Man-made and Natural Electromagnetic Fields, in regard to Biological Activity. Sci. Rep. 2015, 5, 14914. [Google Scholar] [CrossRef]
- Philbin, T.G. Quantum dynamics of the damped harmonic oscillator. New J. Phys. 2012, 14. [Google Scholar] [CrossRef]
- A. Pikovsky, M. A. Pikovsky, M. Rosenblum, and J. Kurths, "Synchronization—A unified approach to nonlinear science," ed: Cambridge University Press, Cambridge, 2001.
- S. Samdal, "The effect of large amplitude motion on the comparison of bond distances from ab initio calculations and experimentally determined bond distances, and on root-mean-square amplitudes of vibration, shrinkage, asymmetry constants, symmetry constraints, and inclusion of rotational constants using the electron diffraction method," Journal of molecular structure, vol. 318, pp. 133-141, 1994.
- Kirillova, S.; Carugo, O. Hydration sites of unpaired RNA bases: a statistical analysis of the PDB structures. BMC Struct. Biol. 2011, 11, 41–41. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, M.; Hänggi, P. Driven quantum tunneling. Phys. Rep. 1998, 304, 229–354. [Google Scholar] [CrossRef]
- S. Shun-Ping et al., "Geometrical structures, vibrational frequencies, force constants and dissociation energies of isotopic water molecules (H2O, HDO, D2O, HTO, DTO, and T2O) under dipole electric field," Chinese Physics B, vol. 20, no. 6, p. 063102, 2011.
- Zong, D.; Hu, H.; Duan, Y.; Sun, Y. Viscosity of Water under Electric Field: Anisotropy Induced by Redistribution of Hydrogen Bonds. J. Phys. Chem. B 2016, 120, 4818–4827. [Google Scholar] [CrossRef]
- Baranyai, A.; Bartók, A.; Chialvo, A.A. Computer simulation of the 13 crystalline phases of ice. J. Chem. Phys. 2005, 123, 054502. [Google Scholar] [CrossRef] [PubMed]
- Laage, D.; Hynes, J.T. A Molecular Jump Mechanism of Water Reorientation. Science 2006, 311, 832–835. [Google Scholar] [CrossRef]
- Marx, D.; Tuckerman, M.E.; Hutter, J.; Parrinello, M. The nature of the hydrated excess proton in water. Nature 1999, 397, 601–604. [Google Scholar] [CrossRef]
- Bagchi, B. Water Dynamics in the Hydration Layer around Proteins and Micelles. Chem. Rev. 2005, 105, 3197–3219. [Google Scholar] [CrossRef]
- Schönichen, A.; Webb, B.A.; Jacobson, M.P.; Barber, D.L. Considering Protonation as a Posttranslational Modification Regulating Protein Structure and Function. Annu. Rev. Biophys. 2013, 42, 289–314. [Google Scholar] [CrossRef]
- Fossat, M.J. MEDOC: A Fast, Scalable, and Mathematically Exact Algorithm for the Site-Specific Prediction of the Protonation Degree in Large Disordered Proteins. J. Chem. Inf. Model. 2025, 65, 873–881. [Google Scholar] [CrossRef]
- Pelton, J.; Torchia, D.; Meadow, N.; Roseman, S. Tautomeric states of the active-site histidines of phosphorylated and unphosphorylated IIIGlc, a signal-transducing protein from escherichia coli, using two-dimensional heteronuclear NMR techniques. Protein Sci. 1993, 2, 543–558. [Google Scholar] [CrossRef]
- Cerón-Carrasco, J.P.; Jacquemin, D. Electric-field induced mutation of DNA: a theoretical investigation of the GC base pair. Phys. Chem. Chem. Phys. 2013, 15, 4548–4553. [Google Scholar] [CrossRef] [PubMed]
- Cerón-Carrasco, J.P.; Jacquemin, D. Electric field induced DNA damage: an open door for selective mutations. Chem. Commun. 2013, 49, 7578–7580. [Google Scholar] [CrossRef]
- Cerón-Carrasco, J.P.; Cerezo, J.; Jacquemin, D. How DNA is damaged by external electric fields: selective mutation vs. random degradation. Phys. Chem. Chem. Phys. 2013, 16, 8243–8246. [Google Scholar] [CrossRef]
- L. Antonov, Tautomerism: Concepts and Applications in Science and Technology. John Wiley & Sons, 2016.
- Singh, V.; Fedeles, B.I.; Essigmann, J.M. Role of tautomerism in RNA biochemistry. RNA 2014, 21, 1–13. [Google Scholar] [CrossRef]
- Abou-Zied, O.K.; Jimenez, R.; Romesberg, F.E. Tautomerization Dynamics of a Model Base Pair in DNA. J. Am. Chem. Soc. 2001, 123, 4613–4614. [Google Scholar] [CrossRef]
- Peng, C.S.; Jones, K.C.; Tokmakoff, A. Anharmonic Vibrational Modes of Nucleic Acid Bases Revealed by 2D IR Spectroscopy. J. Am. Chem. Soc. 2011, 133, 15650–15660. [Google Scholar] [CrossRef]
- Aggarwal, L.; Biswas, P. Hydration Water Distribution around Intrinsically Disordered Proteins. J. Phys. Chem. B 2018, 122, 4206–4218. [Google Scholar] [CrossRef]
- Grubmüller, H.; Heller, H.; Windemuth, A.; Schulten, K. Generalized Verlet Algorithm for Efficient Molecular Dynamics Simulations with Long-range Interactions. Mol. Simul. 1991, 6, 121–142. [Google Scholar] [CrossRef]
- Maity, H.; Baidya, L.; Reddy, G. Salt-Induced Transitions in the Conformational Ensembles of Intrinsically Disordered Proteins. J. Phys. Chem. B 2022, 126, 5959–5971. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.; Bhattacharya, S.; Achuthan, S.; Behal, A.; Jolly, M.K.; Kotnala, S.; Mohanty, A.; Rangarajan, G.; Salgia, R.; Uversky, V. Intrinsically Disordered Proteins: Critical Components of the Wetware. Chem. Rev. 2022, 122, 6614–6633. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Faraggi, E.; Li, Z.; Zhou, Y. Intrinsically Semi-disordered State and Its Role in Induced Folding and Protein Aggregation. Cell Biochem. Biophys. 2013, 67, 1193–1205. [Google Scholar] [CrossRef]
- Nandi, P.K.; Futera, Z.; English, N.J. Perturbation of hydration layer in solvated proteins by external electric and electromagnetic fields: Insights from non-equilibrium molecular dynamics. J. Chem. Phys. 2016, 145, 205101. [Google Scholar] [CrossRef]
- Upadhyay, A.; Ekenna, C. A New Tool to Study the Binding Behavior of Intrinsically Disordered Proteins. Int. J. Mol. Sci. 2023, 24, 11785. [Google Scholar] [CrossRef] [PubMed]
- Abyzov, A.; Blackledge, M.; Zweckstetter, M. Conformational Dynamics of Intrinsically Disordered Proteins Regulate Biomolecular Condensate Chemistry. Chem. Rev. 2022, 122, 6719–6748. [Google Scholar] [CrossRef]
- Kolesnikov, A.I.; Reiter, G.F.; Choudhury, N.; Prisk, T.R.; Mamontov, E.; Podlesnyak, A.; Ehlers, G.; Seel, A.G.; Wesolowski, D.J.; Anovitz, L.M. Quantum Tunneling of Water in Beryl: A New State of the Water Molecule. Phys. Rev. Lett. 2016, 116, 167802–167802. [Google Scholar] [CrossRef]
- Mao, A.H.; Lyle, N.; Pappu, R.V. Describing sequence–ensemble relationships for intrinsically disordered proteins. Biochem. J. 2012, 449, 307–318. [Google Scholar] [CrossRef]
- Mitrea, D.M.; Kriwacki, R.W. Phase separation in biology; functional organization of a higher order. Cell Commun. Signal. 2016, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Han, T.W.; Xie, S.; Shi, K.; Du, X.; Wu, L.C.; Mirzaei, H.; Goldsmith, E.J.; Longgood, J.; Pei, J.; et al. Cell-free Formation of RNA Granules: Low Complexity Sequence Domains Form Dynamic Fibers within Hydrogels. Cell 2012, 149, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Dormann, D.; Lemke, E.A. Adding intrinsically disordered proteins to biological ageing clocks. Nat. Cell Biol. 2024, 26, 851–858. [Google Scholar] [CrossRef]
- Manyilov, V.D.; Ilyinsky, N.S.; Nesterov, S.V.; Saqr, B.M.G.A.; Dayhoff, G.W.; Zinovev, E.V.; Matrenok, S.S.; Fonin, A.V.; Kuznetsova, I.M.; Turoverov, K.K.; et al. Chaotic aging: intrinsically disordered proteins in aging-related processes. Cell. Mol. Life Sci. 2023, 80, 1–29. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Martinelli, A.H.S.; Lopes, F.C.; John, E.B.O.; Carlini, C.R.; Ligabue-Braun, R. Modulation of Disordered Proteins with a Focus on Neurodegenerative Diseases and Other Pathologies. Int. J. Mol. Sci. 2019, 20, 1322. [Google Scholar] [CrossRef]
- Anbo, H.; Sato, M.; Okoshi, A.; Fukuchi, S. Functional Segments on Intrinsically Disordered Regions in Disease-Related Proteins. Biomolecules 2019, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Scholes, G.D.; Fleming, G.R.; Chen, L.X.; Aspuru-Guzik, A.; Buchleitner, A.; Coker, D.F.; Engel, G.; Van Grondelle, R.; Ishizaki, A.; Jonas, D.; et al. Using coherence to enhance function in chemical and biophysical systems. Nature 2017, 543, 647–656. [Google Scholar] [CrossRef]
- C. Bennett, "S teck JE, B ehrm an E C," Quantum information and computation, vol. 404, no. 3, pp. 247-255, 2000.
- J. Cavanagh, W. J. J. Cavanagh, W. J. Fairbrother, A. G. Palmer, M. Rance, and N. J. Skelton, "CHAPTER 7 - HETERONUCLEAR NMR EXPERIMENTS," in Protein NMR Spectroscopy (Second Edition), J. Cavanagh, W. J. Fairbrother, A. G. Palmer, M. Rance, and N. J. Skelton Eds. Burlington: Academic Press, 2007, pp. 533-678.
- Kragelj, J.; Ozenne, V.; Blackledge, M.; Jensen, M.R. Conformational Propensities of Intrinsically Disordered Proteins from NMR Chemical Shifts. Chemphyschem 2013, 14, 3034–3045. [Google Scholar] [CrossRef]
- Narth, C.; Gillet, N.; Cailliez, F.; Lévy, B.; de la Lande, A. Electron Transfer, Decoherence, and Protein Dynamics: Insights from Atomistic Simulations. Accounts Chem. Res. 2015, 48, 1090–1097. [Google Scholar] [CrossRef] [PubMed]
- Tenenbaum, A. Kinetic coherence underlies the dynamics of disordered proteins. RSC Adv. 2021, 11, 36242–36249. [Google Scholar] [CrossRef] [PubMed]
- Rather, S.R.; Scholes, G.D.; Chen, L.X. From Coherence to Function: Exploring the Connection in Chemical Systems. Accounts Chem. Res. 2024, 57, 2620–2630. [Google Scholar] [CrossRef]
- Binolfi, A.; Theillet, F.-X.; Selenko, P. Bacterial in-cell NMR of human α-synuclein: a disordered monomer by nature? Biochem. Soc. Trans. 2012, 40, 950–954. [Google Scholar] [CrossRef]
- Usselman, R.J.; Chavarriaga, C.; Castello, P.R.; Procopio, M.; Ritz, T.; Dratz, E.A.; Singel, D.J.; Martino, C.F. The Quantum Biology of Reactive Oxygen Species Partitioning Impacts Cellular Bioenergetics. Sci. Rep. 2016, 6, 38543. [Google Scholar] [CrossRef]
- Pesce, F.; Bremer, A.; Tesei, G.; Hopkins, J.B.; Grace, C.R.; Mittag, T.; Lindorff-Larsen, K. Design of intrinsically disordered protein variants with diverse structural properties. Sci. Adv. 2024, 10, eadm9926. [Google Scholar] [CrossRef]
- Gehi, B.R.; Gadhave, K.; Uversky, V.N.; Giri, R. Intrinsic disorder in proteins associated with oxidative stress-induced JNK signaling. Cell. Mol. Life Sci. 2022, 79, 1–32. [Google Scholar] [CrossRef]
- J. Leterrier, "Water and the cytoskeleton," Cellular and molecular biology (Noisy-le-Grand, France), vol. 47, no. 5, pp. 901-923, 2001.
- Westerheide, S.D.; Morimoto, R.I. Heat Shock Response Modulators as Therapeutic Tools for Diseases of Protein Conformation. J. Biol. Chem. 2005, 280, 33097–33100. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Pastor, R.; Burchfiel, E.T.; Thiele, D.J. Regulation of heat shock transcription factors and their roles in physiology and disease. Nat. Rev. Mol. Cell Biol. 2017, 19, 4–19. [Google Scholar] [CrossRef] [PubMed]
- U. Sengupta and R. Kayed, "Amyloid β, Tau, and α-Synuclein aggregates in the pathogenesis, prognosis, and therapeutics for neurodegenerative diseases," Progress in neurobiology, vol. 214, p. 102270, 2022.
- Firman, T.; Ghosh, K. Sequence charge decoration dictates coil-globule transition in intrinsically disordered proteins. J. Chem. Phys. 2017, 148, 123305. [Google Scholar] [CrossRef] [PubMed]
- Theillet, F.-X.; Binolfi, A.; Frembgen-Kesner, T.; Hingorani, K.; Sarkar, M.; Kyne, C.; Li, C.; Crowley, P.B.; Gierasch, L.; Pielak, G.J.; et al. Physicochemical Properties of Cells and Their Effects on Intrinsically Disordered Proteins (IDPs). Chem. Rev. 2014, 114, 6661–6714. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, D. CORRELATION BETWEEN POSTTRANSLATIONAL MODIFICATION AND INTRINSIC DISORDER IN PROTEIN. Proceedings of the Pacific Symposium. LOCATION OF CONFERENCE, United StatesDATE OF CONFERENCE; pp. 94–103.
- Khoury, G.A.; Baliban, R.C.; Floudas, C.A. Proteome-wide post-translational modification statistics: frequency analysis and curation of the swiss-prot database. Sci. Rep. 2011, 1, 90. [Google Scholar] [CrossRef]
- Zhao, B.; Katuwawala, A.; Oldfield, C.J.; Hu, G.; Wu, Z.; Uversky, V.N.; Kurgan, L. Intrinsic Disorder in Human RNA-Binding Proteins. J. Mol. Biol. 2021, 433, 167229. [Google Scholar] [CrossRef]
- A. Y. Liu, C. A. A. Y. Liu, C. A. Minetti, D. P. Remeta, K. J. Breslauer, and K. Y. Chen, "HSF1, aging, and neurodegeneration," Cell Biology and Translational Medicine, Volume 18: Tissue Differentiation, Repair in Health and Disease, pp. 23-49, 2022.
- Ren, Q.; Li, L.; Liu, L.; Li, J.; Shi, C.; Sun, Y.; Yao, X.; Hou, Z.; Xiang, S. The molecular mechanism of temperature-dependent phase separation of heat shock factor 1. Nat. Chem. Biol. 2025, 21, 831–842. [Google Scholar] [CrossRef]
- Shamovsky, I.; Ivannikov, M.; Kandel, E.S.; Gershon, D.; Nudler, E. RNA-mediated response to heat shock in mammalian cells. Nature 2006, 440, 556–560. [Google Scholar] [CrossRef]
- Zhang, H.; Shao, S.; Zeng, Y.; Wang, X.; Qin, Y.; Ren, Q.; Xiang, S.; Wang, Y.; Xiao, J.; Sun, Y. Reversible phase separation of HSF1 is required for an acute transcriptional response during heat shock. Nat. Cell Biol. 2022, 24, 340–352. [Google Scholar] [CrossRef]
- Anckar, J.; Sistonen, L. Regulation of HSF1 Function in the Heat Stress Response: Implications in Aging and Disease. Annu. Rev. Biochem. 2011, 80, 1089–1115. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wu, J.; Xu, L.; Wang, J.; Chen, Z.; Yang, R. Regulation of HSF1 protein stabilization: An updated review. Eur. J. Pharmacol. 2018, 822, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Westerheide, S.D.; Anckar, J.; Stevens, S.M.; Sistonen, L.; Morimoto, R.I. Stress-Inducible Regulation of Heat Shock Factor 1 by the Deacetylase SIRT1. Science 2009, 323, 1063–1066. [Google Scholar] [CrossRef] [PubMed]
- Raynes, R.; Brunquell, J.; Westerheide, S.D. Stress Inducibility of SIRT1 and Its Role in Cytoprotection and Cancer. Genes Cancer 2013, 4, 172–182. [Google Scholar] [CrossRef]
- Ma, X.; Xu, L.; Alberobello, A.T.; Gavrilova, O.; Bagattin, A.; Skarulis, M.; Liu, J.; Finkel, T.; Mueller, E. Celastrol Protects against Obesity and Metabolic Dysfunction through Activation of a HSF1-PGC1α Transcriptional Axis. Cell Metab. 2015, 22, 695–708. [Google Scholar] [CrossRef]
- Zelin, E.; Freeman, B.C. Lysine Deacetylases Regulate the Heat Shock Response Including the Age-Associated Impairment of HSF1. J. Mol. Biol. 2015, 427, 1644–1654. [Google Scholar] [CrossRef]
- O. Dasdag, N. O. Dasdag, N. Adalier, and S. Dasdag, "Electromagnetic radiation and Alzheimer’s disease," Biotechnology & Biotechnological Equipment, vol. 34, no. 1, pp. 1087-1094, 2020.
- F. P. Perez, J. P. F. P. Perez, J. P. Bandeira, C. N. Perez Chumbiauca, D. K. Lahiri, J. Morisaki, and M. Rizkalla, "Multidimensional insights into the repeated electromagnetic field stimulation and biosystems interaction in aging and age-related diseases," Journal of Biomedical Science, vol. 29, no. 1, pp. 1-22, 2022.
- I. C. o. N.-I. R. Protection, "Guidelines for limiting exposure to electromagnetic fields (100 kHz to 300 GHz)," Health physics, vol. 118, no. 5, pp. 483-524, 2020.
- Marchesi, N.; Osera, C.; Fassina, L.; Amadio, M.; Angeletti, F.; Morini, M.; Magenes, G.; Venturini, L.; Biggiogera, M.; Ricevuti, G.; et al. Autophagy Is Modulated in Human Neuroblastoma Cells Through Direct Exposition to Low Frequency Electromagnetic Fields. J. Cell. Physiol. 2014, 229, 1776–1786. [Google Scholar] [CrossRef]
- Hirai, T.; Taniura, H.; Goto, Y.; Ogura, M.; Sng, J.C.G.; Yoneda, Y. Stimulation of ubiquitin–proteasome pathway through the expression of amidohydrolase for N-terminal asparagine (Ntan1) in cultured rat hippocampal neurons exposed to static magnetism. J. Neurochem. 2006, 96, 1519–1530. [Google Scholar] [CrossRef]
- Park, J.; Kwon, J.H.; Kim, N.; Song, K. Effects of 1950 MHz radiofrequency electromagnetic fields on Aβ processing in human neuroblastoma and mouse hippocampal neuronal cells. J. Radiat. Res. 2017, 59, 18–26. [Google Scholar] [CrossRef]
- Y. J. Jeong et al., "1950 MHz Electromagnetic Fields Ameliorate Abeta Pathology in Alzheimer's Disease Mice," Current Alzheimer research, vol. 12, no. 5, pp. 481-92, 2015. Online.. Available: http://www.ncbi.nlm.nih.gov/pubmed/26017559.
- Rao, R.R.; Halper, J.; Kisaalita, W.S. Effects of 60 Hz electromagnetic field exposure on APP695 transcription levels in differentiating human neuroblastoma cells. Bioelectrochemistry 2002, 57, 9–15. [Google Scholar] [CrossRef]
- R. A. Antonini et al., "Extremely low-frequency electromagnetic field (ELF-EMF) does not affect the expression of α3, α5 and α7 nicotinic receptor subunit genes in SH-SY5Y neuroblastoma cell line," Toxicology letters, vol. 164, no. 3, pp. 268-277, 2006.
- Del Giudice, E.; Facchinetti, F.; Nofrate, V.; Boccaccio, P.; Minelli, T.; Dam, M.; Leon, A.; Moschini, G. Fifty Hertz electromagnetic field exposure stimulates secretion of β-amyloid peptide in cultured human neuroglioma. Neurosci. Lett. 2007, 418, 9–12. [Google Scholar] [CrossRef]
- He, G.-L.; Luo, Z.; Shen, T.-T.; Li, P.; Yang, J.; Luo, X.; Chen, C.-H.; Gao, P.; Yang, X.-S. Inhibition of STAT3- and MAPK-dependent PGE2 synthesis ameliorates phagocytosis of fibrillar β-amyloid peptide (1-42) via EP2 receptor in EMF-stimulated N9 microglial cells. J. Neuroinflammation 2016, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Newton, T.M.; Duce, J.A.; Bayle, E.D. The proteostasis network provides targets for neurodegeneration. Br. J. Pharmacol. 2019, 176, 3508–3514. [Google Scholar] [CrossRef] [PubMed]
- Kovács, D.; Sigmond, T.; Hotzi, B.; Bohár, B.; Fazekas, D.; Deák, V.; Vellai, T.; Barna, J. HSF1Base: A Comprehensive Database of HSF1 (Heat Shock Factor 1) Target Genes. Int. J. Mol. Sci. 2019, 20, 5815. [Google Scholar] [CrossRef] [PubMed]
- Perez, F.P.; Zhou, X.; Morisaki, J.; Jurivich, D. Electromagnetic field therapy delays cellular senescence and death by enhancement of the heat shock response. Exp. Gerontol. 2008, 43, 307–316. [Google Scholar] [CrossRef] [PubMed]
- F. Manai et al., "A Low-Frequency Electromagnetic (LF-EMF) Exposure Scheme Induces Autophagy Activation to Counteract in Vitro Aβ-Amyloid Neurotoxicity," Atti del, p. 7.
- Trivedi, R.; Knopf, B.; Rakoczy, S.; Manocha, G.D.; Brown-Borg, H.; Jurivich, D.A. Disrupted HSF1 regulation in normal and exceptional brain aging. Biogerontology 2023, 25, 147–160. [Google Scholar] [CrossRef]
- Watanabe, Y.; Taguchi, K.; Tanaka, M. Roles of Stress Response in Autophagy Processes and Aging-Related Diseases. Int. J. Mol. Sci. 2023, 24, 13804. [Google Scholar] [CrossRef]
- Perez, F.P.; Moinuddin, S.S.; Shamim, Q.U.A.; Joseph, D.J.; Morisaki, J.; Zhou, X. Longevity Pathways: HSF1 and FoxO Pathways, a New Therapeutic Target to Prevent Age-Related Diseases. Curr. Aging Sci. 2012, 5, 87–95. [Google Scholar] [CrossRef]
- Perez, F.P.; Zhou, X.; Morisaki, J.; Ilie, J.; James, T.; Jurivich, D.A. Engineered Repeated Electromagnetic Field Shock Therapy for Cellular Senescence and Age-Related Diseases. Rejuvenation Res. 2008, 11, 1049–1058. [Google Scholar] [CrossRef]
- Chou, C.; Bassen, H.; Osepchuk, J.; Balzano, Q.; Petersen, R.; Meltz, M.; Cleveland, R.; Lin, J.; Heynick, L. Radio frequency electromagnetic exposure: Tutorial review on experimental dosimetry. Bioelectromagnetics 1996, 17, 195–208. [Google Scholar] [CrossRef]
- Baldi, E.; Bucherelli, C. The Inverted “U-Shaped” Dose-Effect Relationships in Learning and Memory: Modulation of Arousal and Consolidation. Nonlinearity Biol. Toxicol. Med. 2005, 3, 9–21. [Google Scholar] [CrossRef]
- Ahmad, R.H.M.A.; Fakhoury, M.; Lawand, N. Electromagnetic Field in Alzheimer's Disease: A Literature Review of Recent Preclinical and Clinical Studies. Curr. Alzheimer Res. 2020, 17, 1001–1012. [Google Scholar] [CrossRef]
- Shirbandi, K.; Khalafi, M.; Bevelacqua, J.J.; Sadeghian, N.; Adiban, S.; Zarandi, F.B.; Mortazavi, S.M.J.; Welsh, J.J. Exposure to Low Levels of Radiofrequency Electromagnetic Fields Emitted from Cell-phones as a Promising Treatment of Alzheimer’s Disease: A Scoping Review Study. J. Biomed. Phys. Eng. 2023, 13, 3–16. [Google Scholar] [CrossRef]
- BioInitiative 2012: A Rationale for Biologically-based Exposure Standards for Low-Intensity Electromagnetic Radiation. 2022 Updated Research Summaries.
- Perez, F.P.; Maloney, B.; Chopra, N.; Morisaki, J.J.; Lahiri, D.K. Repeated electromagnetic field stimulation lowers amyloid-β peptide levels in primary human mixed brain tissue cultures. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Tsoy, A.; Saliev, T.; Abzhanova, E.; Turgambayeva, A.; Kaiyrlykyzy, A.; Akishev, M.; Saparbayev, S.; Umbayev, B.; Askarova, S. The Effects of Mobile Phone Radiofrequency Electromagnetic Fields on β-Amyloid-Induced Oxidative Stress in Human and Rat Primary Astrocytes. Neuroscience 2019, 408, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T.; Taniura, H.; Goto, Y.; Ogura, M.; Sng, J.C.G.; Yoneda, Y. Stimulation of ubiquitin–proteasome pathway through the expression of amidohydrolase for N-terminal asparagine (Ntan1) in cultured rat hippocampal neurons exposed to static magnetism. J. Neurochem. 2006, 96, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Osera, C.; Amadio, M.; Falone, S.; Fassina, L.; Magenes, G.; Amicarelli, F.; Ricevuti, G.; Govoni, S.; Pascale, A. Pre-exposure of neuroblastoma cell line to pulsed electromagnetic field prevents H2O2-induced ROS production by increasing MnSOD activity. Bioelectromagnetics 2015, 36, 219–232. [Google Scholar] [CrossRef]
- Osera, C.; Fassina, L.; Amadio, M.; Venturini, L.; Buoso, E.; Magenes, G.; Govoni, S.; Ricevuti, G.; Pascale, A. Cytoprotective Response Induced by Electromagnetic Stimulation on SH-SY5Y Human Neuroblastoma Cell Line. Tissue Eng. Part A 2011, 17, 2573–2582. [Google Scholar] [CrossRef]
- D. Leszczynski, S. D. Leszczynski, S. Joenväärä, J. Reivinen, and R. Kuokka, "Non-thermal activation of the hsp27/p38MAPK stress pathway by mobile phone radiation in human endothelial cells: molecular mechanism for cancer-and blood-brain barrier-related effects," Differentiation, vol. 70, no. 2-3, pp. 120-129, 2002.
- Arendash, G.W.; Mori, T.; Dorsey, M.; Gonzalez, R.; Tajiri, N.; Borlongan, C.; Skoulakis, E.M.C. Electromagnetic Treatment to Old Alzheimer's Mice Reverses β-Amyloid Deposition, Modifies Cerebral Blood Flow, and Provides Selected Cognitive Benefit. PLOS ONE 2012, 7, e35751. [Google Scholar] [CrossRef]
- Arendash, G.W. Transcranial Electromagnetic Treatment Against Alzheimer's Disease: Why it has the Potential to Trump Alzheimer's Disease Drug Development. J. Alzheimer's Dis. 2012, 32, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Dragicevic, N.; Bradshaw, P.; Mamcarz, M.; Lin, X.; Wang, L.; Cao, C.; Arendash, G. Long-term electromagnetic field treatment enhances brain mitochondrial function of both Alzheimer's transgenic mice and normal mice: a mechanism for electromagnetic field-induced cognitive benefit? Neuroscience 2011, 185, 135–149. [Google Scholar] [CrossRef]
- Arendash, G.W.; Sanchez-Ramos, J.; Mori, T.; Mamcarz, M.; Lin, X.; Runfeldt, M.; Wang, L.; Zhang, G.; Sava, V.; Tan, J.; et al. Electromagnetic Field Treatment Protects Against and Reverses Cognitive Impairment in Alzheimer's Disease Mice. J. Alzheimer's Dis. 2010, 19, 191–210. [Google Scholar] [CrossRef]
- Jeong, H.; Andersson, J.; Hess, A.; Jezzard, P. Effect of subject-specific head morphometry on specific absorption rate estimates in parallel-transmit MRI at 7 T. Magn. Reson. Med. 2023, 89, 2376–2390. [Google Scholar] [CrossRef]
- Banaceur, S.; Banasr, S.; Sakly, M.; Abdelmelek, H. Whole body exposure to 2.4GHz WIFI signals: Effects on cognitive impairment in adult triple transgenic mouse models of Alzheimer's disease (3xTg-AD). Behav. Brain Res. 2013, 240, 197–201. [Google Scholar] [CrossRef]
- Y. J. Jeong et al., "Behavioral changes and gene profile alterations after chronic 1,950-MHz radiofrequency exposure: An observation in C57BL/6 mice," Brain and Behavior, vol. 10, no. 11, p. e01815, 2020.
- Kumlin, T.; Iivonen, H.; Miettinen, P.; Juvonen, A.; van Groen, T.; Puranen, L.; Pitkäaho, R.; Juutilainen, J.; Tanila, H. Mobile Phone Radiation and the Developing Brain: Behavioral and Morphological Effects in Juvenile Rats. Radiat. Res. 2007, 168, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Lu, J.-M.; Xing, Z.-H.; Zhao, Q.-R.; Hu, L.-Q.; Xue, L.; Zhang, J.; Mei, Y.-A. Effect of 1.8 GHz radiofrequency electromagnetic radiation on novel object associative recognition memory in mice. Sci. Rep. 2017, 7, srep44521. [Google Scholar] [CrossRef]
- Son, Y.; Kim, J.S.; Jeong, Y.J.; Jeong, Y.K.; Kwon, J.H.; Choi, H.-D.; Pack, J.-K.; Kim, N.; Lee, Y.-S.; Lee, H.-J. Long-term RF exposure on behavior and cerebral glucose metabolism in 5xFAD mice. Neurosci. Lett. 2018, 666, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Lai, J.; Wan, B.; Liu, X.; Zhang, Y.; Zhang, J.; Sun, D.; Ruan, G.; Liu, E.; Liu, G.-P.; et al. Long-term exposure to ELF-MF ameliorates cognitive deficits and attenuates tau hyperphosphorylation in 3xTg AD mice. NeuroToxicology 2016, 53, 290–300. [Google Scholar] [CrossRef]
- Liu, X.; Zuo, H.; Wang, D.; Peng, R.; Song, T.; Wang, S.; Xu, X.; Gao, Y.; Li, Y.; Wang, S.; et al. Improvement of Spatial Memory Disorder and Hippocampal Damage by Exposure to Electromagnetic Fields in an Alzheimer’s Disease Rat Model. PLOS ONE 2015, 10, e0126963–e0126963. [Google Scholar] [CrossRef]
- Akbarnejad, Z.; Esmaeilpour, K.; Shabani, M.; Asadi-Shekaari, M.; Goraghani, M.S.; Ahmadi-Zeidabadi, M. Spatial memory recovery in Alzheimer's rat model by electromagnetic field exposure. Int. J. Neurosci. 2017, 128, 691–696. [Google Scholar] [CrossRef]
- D. de Pomerai et al., "Non-thermal heat-shock response to microwaves," Nature, vol. 405, no. 6785, pp. 417-418, 2000.
- Shallom, J.M.; Di Carlo, A.L.; Ko, D.; Penafiel, L.M.; Nakai, A.; Litovitz, T.A. Microwave exposure induces Hsp70 and confers protection against hypoxia in chick embryos. J. Cell. Biochem. 2002, 86, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Weisbrot, D.; Lin, H.; Ye, L.; Blank, M.; Goodman, R. Effects of mobile phone radiation on reproduction and development in Drosophila melanogaster. J. Cell. Biochem. 2003, 89, 48–55. [Google Scholar] [CrossRef]
- G. Arendash et al., "A Clinical Trial of Transcranial Electromagnetic Treatment in Alzheimer’s Disease: Cognitive Enhancement and Associated Changes in Cerebrospinal Fluid, Blood, and Brain Imaging," Journal of Alzheimer's Disease, no. Preprint, pp. 1-26, 2019.
- Arendash, G.; Abulaban, H.; Steen, S.; Andel, R.; Wang, Y.; Bai, Y.; Baranowski, R.; McGarity, J.; Scritsmier, L.; Lin, X.; et al. Transcranial Electromagnetic Treatment Stops Alzheimer’s Disease Cognitive Decline over a 2½-Year Period: A Pilot Study. Medicines 2022, 9, 42. [Google Scholar] [CrossRef]
- Cao, C.; Abulaban, H.; Baranowski, R.; Wang, Y.; Bai, Y.; Lin, X.; Shen, N.; Zhang, X.; Arendash, G.W. Transcranial Electromagnetic Treatment “Rebalances” Blood and Brain Cytokine Levels in Alzheimer’s Patients: A New Mechanism for Reversal of Their Cognitive Impairment. Front. Aging Neurosci. 2022, 14, 829049. [Google Scholar] [CrossRef]
- F. Söderqvist, L. F. Söderqvist, L. Hardell, M. Carlberg, and K. H. Mild, "Radiofrequency fields, transthyretin, and Alzheimer's disease," Journal of Alzheimer's Disease, vol. 20, no. 2, pp. 599-606, 2010.
- Sandyk, R. Alzheimer's Disease: Improvement of Visual Memory and Visuoconstructive Performance by Treatment with Picotesla Range Magnetic Fields. Int. J. Neurosci. 1994, 76, 185–225. [Google Scholar] [CrossRef]
- H. Gen-Lin et al., "Inhibition of STAT3-and MAPK-dependent PGE2 synthesis ameliorates phagocytosis of fibrillar Beta-amyloid peptide (1-42) via EP2 receptor in EMF-stimulated N9 microglial cells," Journal of Neuroinflammation, vol. 13, 2016.
- Barthelemy, A.; Mouchard, A.; Villegier, A.-S. Glial markers and emotional memory in rats following cerebral radiofrequency exposures. 2016 IEEE Radio and Antenna Days of the Indian Ocean (RADIO). LOCATION OF CONFERENCE, FranceDATE OF CONFERENCE; pp. 1–2.
- Jiang, D.-P.; Zhang, J.; Xu, S.-L.; Kuang, F.; Lang, H.-Y.; Wang, Y.-F.; An, G.-Z.; Li, J.-H.; Guo, G.-Z. Electromagnetic Pulse Exposure Induces Overexpression of Beta Amyloid Protein in Rats. Arch. Med Res. 2013, 44, 178–184. [Google Scholar] [CrossRef]
- Jiang, D.-P.; Li, J.-H.; Zhang, J.; Xu, S.-L.; Kuang, F.; Lang, H.-Y.; Wang, Y.-F.; An, G.-Z.; Guo, G.-Z. Long-term electromagnetic pulse exposure induces Abeta deposition and cognitive dysfunction through oxidative stress and overexpression of APP and BACE1. Brain Res. 2016, 1642, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.; Peng, R.; Yan, H.; Gao, Y.; Wang, C.; Wang, S.; Zou, Y.; Xu, X.; Zhao, L.; Dong, J.; et al. Reduction of Phosphorylated Synapsin I (Ser-553) Leads to Spatial Memory Impairment by Attenuating GABA Release after Microwave Exposure in Wistar Rats. PLOS ONE 2014, 9, e95503. [Google Scholar] [CrossRef]
- Prochnow, N.; Gebing, T.; Ladage, K.; Krause-Finkeldey, D.; El Ouardi, A.; Bitz, A.; Streckert, J.; Hansen, V.; Dermietzel, R.; Meuth, S.G. Electromagnetic Field Effect or Simply Stress? Effects of UMTS Exposure on Hippocampal Longterm Plasticity in the Context of Procedure Related Hormone Release. PLOS ONE 2011, 6, e19437. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Zhou, H.; Wang, S.; Gao, Y.; Wang, L.; Yong, Z.; Zuo, H.; Zhao, L.; Dong, J.; et al. Impairment of long-term potentiation induction is essential for the disruption of spatial memory after microwave exposure. Int. J. Radiat. Biol. 2013, 89, 1100–1107. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Zhao, L.; Wang, S.; Gao, Y.; Wang, L.; Zuo, H.; Dong, J.; Xu, X.; Zhou, H.; et al. The relationship between NMDA receptors and microwave-induced learning and memory impairment: A long-term observation on Wistar rats. Int. J. Radiat. Biol. 2015, 91, 262–269. [Google Scholar] [CrossRef]
- Wang, H.; Tan, S.; Xu, X.; Zhao, L.; Zhang, J.; Yao, B.; Gao, Y.; Zhou, H.; Peng, R. Long term impairment of cognitive functions and alterations of NMDAR subunits after continuous microwave exposure. Physiol. Behav. 2017, 181, 1–9. [Google Scholar] [CrossRef]
- E. Foroozandeh, M. S. E. Foroozandeh, M. S. Naeini, H. Ahadi, and J. Foroozandeh, "Effects of 90min Exposure to 8mT Electromagnetic Fields on Memory in Mice," Journal of American Science, vol. 7, no. 7, 2011.
- Yang, X.; He, G.; Hao, Y.; Chen, C.; Li, M.; Wang, Y.; Zhang, G.; Yu, Z. The role of the JAK2-STAT3 pathway in pro-inflammatory responses of EMF-stimulated N9 microglial cells. J. Neuroinflammation 2010, 7, 54–54. [Google Scholar] [CrossRef]
- Cleary, S.F.; Cao, G.; Liu, L.-M.; Egle, P.M.; Shelton, K.R. Stress proteins are not induced in mammalian cells exposed to radiofrequency or microwave radiation. Bioelectromagnetics 1997, 18, 499–505. [Google Scholar] [CrossRef]
- S. J. Regel et al., "Pulsed radio-frequency electromagnetic fields: dose-dependent effects on sleep, the sleep EEG and cognitive performance," Journal of sleep research, vol. 16, no. 3, pp. 253-258, 2007.
- S. J. Regel et al., "Pulsed radio frequency radiation affects cognitive performance and the waking electroencephalogram," Neuroreport, vol. 18, no. 8, pp. 803-807, 2007.
- J. H. Kim, D.-H. J. H. Kim, D.-H. Yu, Y. H. Huh, E. H. Lee, H.-G. Kim, and H. R. Kim, "Long-term exposure to 835 MHz RF-EMF induces hyperactivity, autophagy and demyelination in the cortical neurons of mice," Scientific reports, vol. 7, no. 1, p. 41129, 2017.
- Safety Levels with Respect to Human Exposure to Electric, Magnetic, and Electromagnetic Fields, 0 Hz to 300 GHz, (2019). I. S. C95.1:2019, 2019.
- Lanni, I.; Chiacchierini, G.; Papagno, C.; Santangelo, V.; Campolongo, P. Treating Alzheimer’s disease with brain stimulation: From preclinical models to non-invasive stimulation in humans. Neurosci. Biobehav. Rev. 2024, 165, 105831. [Google Scholar] [CrossRef]
- F. M. Ribeiro, E. R. d. S. F. M. Ribeiro, E. R. d. S. Camargos, L. C. d. Souza, and A. L. Teixeira, "Animal models of neurodegenerative diseases," Revista brasileira de psiquiatria, vol. 35, no. Suppl 2, pp. S82-S91, 2013.
- Guerriero, F.; Botarelli, E.; Mele, G.; Polo, L.; Zoncu, D.; Renati, P.; Sgarlata, C.; Rollone, M.; Ricevuti, G.; Maurizi, N.; et al. An innovative intervention for the treatment of cognitive impairment–Emisymmetric bilateral stimulation improves cognitive functions in Alzheimer’s disease and mild cognitive impairment: an open-label study. Neuropsychiatr. Dis. Treat. 2015, 11, 2391–2404. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Park, H.-J.; Jeong, Y.J.; Choi, H.-D.; Kim, N.; Lee, H.-J. Long-term radiofrequency electromagnetic fields exposure attenuates cognitive dysfunction in 5×FAD mice by regulating microglial function. Neural Regen. Res. 2023, 18, 2497–2503. [Google Scholar] [CrossRef]
- Zhi, W.; Zou, Y.; Ma, L.; He, S.; Guo, Z.; Zhao, X.; Hu, X.; Wang, L. 900 MHZ electromagnetic field exposure relieved AD-like symptoms on APP/PS1 mice: A potential non-invasive strategy for AD treatment. Biochem. Biophys. Res. Commun. 2023, 658, 97–106. [Google Scholar] [CrossRef]
- Komaki, A.; Salehi, I.; Keymoradzadeh, A.; Azandaryani, M.T.; Golipoor, Z. Effect of Long-term Exposure to Extremely Low-frequency Electromagnetic Fields on β-amyloid Deposition and Microglia Cells in an Alzheimer Model in Rats. J. Guilan Univ. Med Sci. 2021, 30, 218–229. [Google Scholar] [CrossRef]
- S. Zhang et al., "Effects of 2.4 GHz radiofrequency electromagnetic field exposure on hippocampal proteins in APP/PS1 mice," 2024.
- Teranishi, M.; Ito, M.; Huang, Z.; Nishiyama, Y.; Masuda, A.; Mino, H.; Tachibana, M.; Inada, T.; Ohno, K. Extremely Low-Frequency Electromagnetic Field (ELF-EMF) Increases Mitochondrial Electron Transport Chain Activities and Ameliorates Depressive Behaviors in Mice. Int. J. Mol. Sci. 2024, 25, 11315. [Google Scholar] [CrossRef]
- Kim, J.H.; Yu, D.-H.; Kim, H.-J.; Huh, Y.H.; Cho, S.-W.; Lee, J.-K.; Kim, H.-G.; Kim, H.R. Exposure to 835 MHz radiofrequency electromagnetic field induces autophagy in hippocampus but not in brain stem of mice. Toxicol. Ind. Heal. 2017, 34, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.-W.; Xie, W.-J.; Yu, B.; Song, C.; Ji, X.-M.; Wang, X.-Y.; Zhang, M.; Zhang, X. Rotating magnetic field inhibits Aβ protein aggregation and alleviates cognitive impairment in Alzheimer’s disease mice. Zool. Res. 2024, 45, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Y.; Zhao, Y.; Wang, P.; Ding, H.; Liu, C.; Lyu, J.; Le, W. Terahertz Irradiation Improves Cognitive Impairments and Attenuates Alzheimer’s Neuropathology in the APPSWE/PS1DE9 Mouse: A Novel Therapeutic Intervention for Alzheimer’s Disease. Neurosci. Bull. 2023, 40, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Moya-Gómez, A.; Font, L.P.; Burlacu, A.; Alpizar, Y.A.; Cardonne, M.M.; Brône, B.; Bronckaers, A. Extremely Low-Frequency Electromagnetic Stimulation (ELF-EMS) Improves Neurological Outcome and Reduces Microglial Reactivity in a Rodent Model of Global Transient Stroke. Int. J. Mol. Sci. 2023, 24, 11117. [Google Scholar] [CrossRef]
- Abkhezr, H.; Mohaddes, G.; Nikniaz, Z.; Farhangi, M.A.; Heydari, H.; Nikniaz, L. The effect of Extremely Low Frequency Electromagnetic Field on spatial memory of mice and rats: A systematic review. Learn. Motiv. 2023, 81. [Google Scholar] [CrossRef]
- Eskandani, R.; Zibaii, M.I. Unveiling the biological effects of radio-frequency and extremely-low frequency electromagnetic fields on the central nervous system performance. BioImpacts 2023, 14, 30064–30064. [Google Scholar] [CrossRef]
- Perez, F.P.; Morisaki, J.; Kanakri, H.; Rizkalla, M. Electromagnetic Field Stimulation Therapy for Alzheimer’s Disease. 2024, 3. 3.
- Jiang, D.-P.; Li, J.-H.; Zhang, J.; Xu, S.-L.; Kuang, F.; Lang, H.-Y.; Wang, Y.-F.; An, G.-Z.; Guo, G.-Z. Long-term electromagnetic pulse exposure induces Abeta deposition and cognitive dysfunction through oxidative stress and overexpression of APP and BACE1. Brain Res. 2016, 1642, 10–19. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Zhang, J.; Li, N. Short-term effects of extremely low frequency electromagnetic fields exposure on Alzheimer's disease in rats. Int. J. Radiat. Biol. 2014, 91, 28–34. [Google Scholar] [CrossRef]
- Bouji, M.; Lecomte, A.; Gamez, C.; Blazy, K.; Villégier, A.-S. Impact of Cerebral Radiofrequency Exposures on Oxidative Stress and Corticosterone in a Rat Model of Alzheimer’s Disease. J. Alzheimer's Dis. 2019, 73, 467–476. [Google Scholar] [CrossRef]
- Son, Y.; Jeong, Y.J.; Kwon, J.H.; Choi, H.; Pack, J.; Kim, N.; Lee, Y.; Lee, H. 1950 MHz radiofrequency electromagnetic fields do not aggravate memory deficits in 5xFAD mice. Bioelectromagnetics 2016, 37, 391–399. [Google Scholar] [CrossRef]
- C. Messori, S. V. C. Messori, S. V. Prinzera, and F. B. di Bardone, "The super-coherent state of biological water," Open Access Library Journal, vol. 6, no. 02, p. 1, 2019.
- Madl, P.; Renati, P. Quantum Electrodynamics Coherence and Hormesis: Foundations of Quantum Biology. Int. J. Mol. Sci. 2023, 24, 14003. [Google Scholar] [CrossRef]
- Huang, Z.; Ito, M.; Zhang, S.; Toda, T.; Takeda, J.-I.; Ogi, T.; Ohno, K. Extremely low-frequency electromagnetic field induces acetylation of heat shock proteins and enhances protein folding. Ecotoxicol. Environ. Saf. 2023, 264, 115482. [Google Scholar] [CrossRef] [PubMed]
- Morotomi-Yano, K.; Oyadomari, S.; Akiyama, H.; Yano, K.-I. Nanosecond pulsed electric fields act as a novel cellular stress that induces translational suppression accompanied by eIF2α phosphorylation and 4E-BP1 dephosphorylation. Exp. Cell Res. 2012, 318, 1733–1744. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lee, Y.S.; Kim, N.; Choi, H.-D.; Lim, K.-M. 5G Electromagnetic Radiation Attenuates Skin Melanogenesis In Vitro by Suppressing ROS Generation. Antioxidants 2022, 11, 1449. [Google Scholar] [CrossRef] [PubMed]
- Lang, B.J.; Guerrero, M.E.; Prince, T.L.; Okusha, Y.; Bonorino, C.; Calderwood, S.K. The functions and regulation of heat shock proteins; key orchestrators of proteostasis and the heat shock response. Arch. Toxicol. 2021, 95, 1943–1970. [Google Scholar] [CrossRef] [PubMed]
- C. Hu et al., "Heat shock proteins: Biological functions, pathological roles, and therapeutic opportunities," MedComm, vol. 3, no. 3, p. e161, 2022.
- Usselman, R.J.; Hill, I.; Singel, D.J.; Martino, C.F.; Langowski, J. Spin Biochemistry Modulates Reactive Oxygen Species (ROS) Production by Radio Frequency Magnetic Fields. PLOS ONE 2014, 9, e93065. [Google Scholar] [CrossRef]
- Zeng, Y.; Shen, Y.; Hong, L.; Chen, Y.; Shi, X.; Zeng, Q.; Yu, P. Effects of Single and Repeated Exposure to a 50-Hz 2-mT Electromagnetic Field on Primary Cultured Hippocampal Neurons. Neurosci. Bull. 2017, 33, 299–306. [Google Scholar] [CrossRef]
- Benassi, B.; Filomeni, G.; Montagna, C.; Merla, C.; Lopresto, V.; Pinto, R.; Marino, C.; Consales, C. Extremely Low Frequency Magnetic Field (ELF-MF) Exposure Sensitizes SH-SY5Y Cells to the Pro-Parkinson’s Disease Toxin MPP+. Mol. Neurobiol. 2015, 53, 4247–4260. [Google Scholar] [CrossRef]
- Schuermann, D.; Mevissen, M. Manmade Electromagnetic Fields and Oxidative Stress—Biological Effects and Consequences for Health. Int. J. Mol. Sci. 2021, 22, 3772. [Google Scholar] [CrossRef]
- Kang, K.A.; Lee, H.C.; Hong, M.-N.; Park, M.-J.; Lee, Y.-S.; Choi, H.-D.; Kim, N.; Ko, Y.-G.; Lee, J.-S. Effects of combined radiofrequency radiation exposure on levels of reactive oxygen species in neuronal cells. J. Radiat. Res. 2013, 55, 265–276. [Google Scholar] [CrossRef]
- de Gannes, F.P.; Haro, E.; Hurtier, A.; Taxile, M.; Ruffié, G.; Billaudel, B.; Veyret, B.; Lagroye, I. Effect of Exposure to the Edge Signal on Oxidative Stress in Brain Cell Models. Radiat. Res. 2011, 175, 225–230. [Google Scholar] [CrossRef]
- Alejandro, M.-M.; Herlinda, B.-J.; Aparicio-Bautista, D.I.; Mondragón-Rodríguez, S.; Overduin, M.; Basurto-Islas, G. Molecular Mechanisms Associated with the Interaction of External Electromagnetic Fields in Protein Dynamics and Aggregation: A Focus on Amyloid-β Peptide. Prog. Biomed. Eng. 2025, 7. [Google Scholar] [CrossRef]
- Saikia, J.; Pandey, G.; Sashidharan, S.; Antony, F.; Nemade, H.B.; Kumar, S.; Chaudhary, N.; Ramakrishnan, V. Electric Field Disruption of Amyloid Aggregation: Potential Noninvasive Therapy for Alzheimer’s Disease. ACS Chem. Neurosci. 2019, 10, 2250–2262. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Choi, J.; Park, S.-J.; Shim, K.-H.; Baek, C.; Han, H.-B.; Hong, H.-C.; Yun, S.; Cha, M.; Shah, Y.H.; et al. In Vitro Treatment of Alzheimer’s disease by Disintegrating Amyloid-β Using Electromagnetic Waves. IEEE Trans. Antennas Propag. 2025, PP, 1–1. [Google Scholar] [CrossRef]
- Muscat, S.; Stojceski, F.; Danani, A. Elucidating the Effect of Static Electric Field on Amyloid Beta 1–42 Supramolecular Assembly. J. Mol. Graph. Model. 2020, 96, 107535. [Google Scholar] [CrossRef]
- Vargas-Rosales, P.A.; D’aDdio, A.; Zhang, Y.; Caflisch, A. Disrupting Dimeric β-Amyloid by Electric Fields. ACS Phys. Chem. Au 2023, 3, 456–466. [Google Scholar] [CrossRef]
- Salehi, N.; Lohrasebi, A.; Bordbar, A. Preventing the amyloid-beta peptides accumulation on the cell membrane by applying GHz electric fields: A molecular dynamic simulation. J. Mol. Graph. Model. 2023, 123, 108516. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.M.; Scanlan, J.M.; Gama-Chonlon, L. Bilateral rTMS Shows No Advantage in Depression nor in Comorbid Depression and Anxiety: A Naturalistic Study. Psychiatr. Q. 2023, 95, 107–120. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Hsieh, S.-W.; Chang, H.-W.; Sung, J.-L.; Chuu, C.-P.; Yen, C.-W.; Hour, T.-C. Gamma Frequency Inhibits the Secretion and Aggregation of Amyloid-β and Decreases the Phosphorylation of mTOR and Tau Proteins in vitro. J. Alzheimer's Dis. 2022, 90, 917–928. [Google Scholar] [CrossRef]
- Lobyntseva, A.; Ganaiem, M.; Ivashko-Pachima, Y.; Barnstable, C.J.; Weisinger, B.; Parabucki, A.; Segal, Y.; Shohami, E.; Gozes, I. Extremely Low-Frequency and Low-Intensity Electromagnetic Field Technology (ELF-EMF) Sculpts Microtubules. Eur. J. Neurosci. 2025, 61, e70023. [Google Scholar] [CrossRef]
- Todorova, N.; Bentvelzen, A.; Yarovsky, I. Electromagnetic field modulates aggregation propensity of amyloid peptides. J. Chem. Phys. 2020, 152, 035104. [Google Scholar] [CrossRef]
- Toschi, F.; Lugli, F.; Biscarini, F.; Zerbetto, F. Effects of Electric Field Stress on a β-Amyloid Peptide. J. Phys. Chem. B 2008, 113, 369–376. [Google Scholar] [CrossRef]
- Lin, J.C. Electromagnetic Fields in Biological Systems; Taylor & Francis: London, United Kingdom, 2016. [Google Scholar]
- N. Lambert, Y.-N. N. Lambert, Y.-N. Chen, Y.-C. Cheng, C.-M. Li, G.-Y. Chen, and F. Nori, "Quantum biology," Nature Physics, vol. 9, no. 1, pp. 10-18, 2013.
- MARINO, A.; BECKER, R. BIOLOGICAL EFFECTS OF EXTREMELY LOW-FREQUENCY ELECTRIC AND MAGNETIC-FIELDS - REVIEW. 1977, 9, 131–147.
- Semchenko, I.V.; Mikhalka, I.S.; Khakhomov, S.A.; Samofalov, A.L.; Balmakou, A.P. DNA-like Helices as Nanosized Polarizers of Electromagnetic Waves. Front. Nanotechnol. 2022, 4. [Google Scholar] [CrossRef]
- Li, M.; Liu, M.; Sha, Y. Induced and Inversed Circularly Polarized Luminescence of Achiral Thioflavin T Assembled on Peptide Fibril. Small 2021, 18, 2106130. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, F.; Shi, W.; Zhang, T.; Chang, S. Terahertz circular polarization sensing for protein denaturation based on a twisted dual-layer metasurface. Biomed. Opt. Express 2021, 13, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Perez, F.; Morisaki, J.; Kanakri, H.; Rizkalla, M.; Abdalla, A. A Novel Design of a Portable Birdcage via Meander Line Antenna (MLA) to Lower Beta Amyloid (Aβ) in Alzheimer’s Disease. IEEE J. Transl. Eng. Heal. Med. 2025, 13, 158–173. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Gas, P. Electromagnetic Fields and Neurodegenerative Diseases. Przeglad Elektrotechniczny 2019, 1, 131–135. [Google Scholar] [CrossRef]
- A. L. Fymat, "Electromagnetic Therapy for Neurological and Neurodegenerative Diseases: II. Deep Brain Stimulation," Open Access Journal of Neurology and Neurosurgery, vol. 13, no. 1, pp. 1-17, 2020.
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



