Submitted:
25 July 2025
Posted:
28 July 2025
You are already at the latest version
Abstract
Major Depressive Disorder (MDD) is a highly prevalent psychiatric condition characterized by complex neurobiological mechanisms, including oxidative stress and neuroinflammation, with microglial activation playing a key role in its pathophysiology. Conventional antidepressants, though widely used, often fail to achieve remission due to limited efficacy, adverse effects, and poor patient adherence. In this context, nanotechnology-based drug delivery systems have emerged as promising strategies to overcome pharmacological limitations, enhance blood–brain barrier penetration, and target neuroinflammatory pathways. This narrative review explores the role of microglia as both mediators of neuroinflammation and potential therapeutic targets in MDD. We examine different nanocarriers — such as polymeric nanoparticles, solid lipid nanoparticles, nanostructured lipid carriers, dendrimers, magnetic nanoparticles, and liposomes — and their ability to modulate microglial activation, promote a shift from a pro-inflammatory (M1) to an anti-inflammatory (M2) phenotype, and enhance antidepressant efficacy. Preclinical studies have demonstrated that nanoparticle-based systems not only improve drug bioavailability and brain targeting but also potentiate neuroprotective effects by reducing oxidative stress, promoting neurogenesis, and restoring synaptic plasticity. These findings highlight the potential of nanotechnology as a novel approach to precision neuropsychopharmacology. This review aims to provide an integrative perspective on how nanocarrier-based strategies targeting microglia could redefine future therapeutic paradigms for MDD.
Keywords:
drugs delivery
; neurological diseases
; neuroinflammation
; immunomodulation
1. Introduction
Depression is one of the leading causes of morbidity worldwide, with a high prevalence and a profound impact on quality of life. Prolonged stress is a major precipitating factor for severe depression and neurocognitive deficits, while affected individuals frequently develop comorbidities such as cardiovascular disease, diabetes, cancer, and obesity. Recent studies indicate that increased metabolic stress and accelerated cellular aging further compromise physical health, with oxidative stress — a key component of metabolic stress — being strongly implicated in the pathophysiology of depression [1,2].
Building on this clinical burden, mood disorders are among the most prevalent and severe psychiatric illnesses, encompassing unipolar depression (characterized by fluctuations between euthymia and depression) and bipolar disorder (which involves episodes of hypomania, mania, euthymia, and depression). According to the Diagnostic and Statistical Manual of Mental Disorders (DSM-5, 2013), major depressive episodes are associated with complex symptoms, including alterations in sleep, appetite, psychomotor activity, cognition, and mood [3].There are several risk factors associated with the development of depression, including In addition to their clinical heterogeneity, depressive disorders arise from a multifactorial interplay between genetic, environmental, and medical factors. Risk factors include a family history of depression (with approximately 35% of the risk being hereditary), childhood abuse and neglect, female sex, and recent exposure to stressors. Moreover, metabolic and autoimmune conditions significantly increase the risk of developing depressive symptoms [4].
Given this complexity, advances in neurobiology have been crucial in uncovering the underlying mechanisms of Major Depressive Disorder (MDD). Beyond classical monoaminergic hypotheses, robust evidence highlights the role of neuroinflammation, particularly microglial activation, as a key driver of persistent and treatment-resistant depression [5,6,7]. Microglia, the primary immune cells of the central nervous system (CNS), maintain brain homeostasis through immune surveillance, clearance of cellular debris, and modulation of synaptic activity. However, under chronic stress or systemic inflammation, microglia adopt a prolonged pro-inflammatory (M1) phenotype, releasing cytokines such as interleukin-1β (IL-1β), tumor necrosis factor-alpha (TNF-α), and interleukin-6 (IL-6), along with reactive oxygen species (ROS) [8]. These inflammatory mediators lead to neurochemical and neuroendocrine alterations, including glutamatergic dysfunction, neurotoxicity, reduced hippocampal neurogenesis, and impaired synaptic plasticity, all of which contribute to the persistence of treatment-resistant symptoms [9].
Supporting this inflammatory hypothesis, clinical and preclinical studies have consistently reported elevated levels of inflammatory markers, including C-reactive protein (CRP), IL-6, and TNF-α, in patients with treatment-resistant depression. This reinforces the urgent need for therapeutic approaches capable of modulating microglia-mediated neuroinflammation [7,10,11]. In this context, novel controlled drug delivery systems have emerged as a promising strategy to address the limitations of conventional antidepressant therapies, which are often associated with suboptimal efficacy, significant side effects, and poor patient adherence. Controlled and targeted drug delivery platforms have the potential to enhance therapeutic precision, reduce systemic toxicity, and improve clinical outcomes [12,13].
Furthermore, advanced delivery systems can be engineered for sustained or pulsatile drug release, decreasing dosing frequency and promoting better adherence. They also enable the co-delivery of multiple agents (e.g., antidepressants combined with anti-inflammatory or antioxidant drugs), supporting multimodal approaches to address the complex pathophysiology of treatment-resistant depression [14]. Among these innovative approaches, nanocarriers — such as polymeric nanoparticles, lipid-based systems, dendrimers, and exosomes — have gained attention for their ability to enhance bioavailability, achieve targeted brain delivery, and modulate the pharmacokinetics of therapeutic agents. These systems can cross the blood–brain barrier (BBB) via passive diffusion, carrier-mediated transport, or receptor-mediated transcytosis, and can be functionalized to respond to pathological cues such as pH changes, oxidative stress, or inflammation [15]. Importantly, in the context of MDD, nanotechnology-based strategies hold promise not only for optimizing drug delivery but also for directly modulating microglial activity, a critical element in the neuroinflammatory pathways of depression [16]. Therefore, this review aims to explore the therapeutic potential of targeting microglia via nanotechnology as an innovative strategy to overcome neuroinflammation and advance the treatment of Major Depressive Disorder.
1.1. Morphofunction Aspects of Microglia
In the central nervous system (CNS), the maintenance of cerebral homeostasis depends on a complex balance, primarily regulated by glial cells, especially microglia. Among the various functions attributed to these cells, a key role is their ability to detect changes in the neural environment and respond to threats to tissue integrity through receptors collectively known as the microglial sensome [17]. When homeostasis is disrupted, as in cases of infections, injuries, or chronic stress, glial cells undergo a phenotypic transformation, shifting from a quiescent state to a reactive profile. This transformation initiates a localized inflammatory response, a phenomenon referred to as neuroinflammation [18].
Microglial activation is triggered by danger signals such as pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), which are recognized by specialized receptors, notably the Toll-like receptors (TLRs). Upon activation, microglia undertake a range of essential functions, including the clearance of cellular debris, phagocytosis of dysfunctional synapses, removal of neurotoxic proteins, and active participation in synaptic pruning, a critical process during neural development and in synaptic remodeling throughout life [19].
Morphological and functional alterations of microglia in key brain regions—such as the prefrontal cortex (PFC), hippocampus (HIP), anterior cingulate cortex (ACC), and amygdala—have been consistently associated with the onset and persistence of major depressive disorder (MDD). Exacerbated microglial activation contributes to sustained states of neuroinflammation [20]. Microglial activation alternates between two functional states: M1 and M2. The M1 state is characterized by a pro-inflammatory response, whereas the M2 state is associated with anti-inflammatory and tissue-repair functions [21]. During M1 activation, microglial cells increase in size, acquire an amoeboid morphology, and release pro-inflammatory cytokines to neutralize potential threats. This state is typically induced by stimuli such as interferon-gamma (IFN-γ), lipopolysaccharide (LPS), and beta-amyloid (Aβ). As a result, there is an elevated secretion of pro-inflammatory mediators, including tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), reactive oxygen species (ROS), and nitric oxide (NO). Conversely, the M2 phenotype is activated by anti-inflammatory cytokines such as interleukin-4 (IL-4), interleukin-10 (IL-10), and interleukin-13 (IL-13). This state is characterized by the release of trophic factors, notably brain-derived neurotrophic factor (BDNF) and transforming growth factor-beta (TGF-β). The M2 profile is associated with the resolution of inflammation, tissue repair, and the promotion of neuroplasticity [22,23] (Figure 1).
1.2. Molecular Mechanisms of Neuroinflammation in MDD
Accumulating evidence indicates that Major Depressive Disorder (MDD) is closely linked to the activation of inflammatory pathways and increased oxidative stress within the central nervous system. Elevated levels of inflammation are associated with an increased risk of developing recent-onset depressive episodes; however, the causal directionality between inflammation and depression remains incompletely understood. Current literature supports a bidirectional relationship whereby depression may induce an inflammatory state, while systemic inflammatory processes can precipitate depressive symptoms [24].
Astrocytes, beyond their traditional role in neural support, have emerged as significant sources of cytokine and inflammatory mediator production, contributing to the neurochemical and behavioral dysfunction observed in depression [25]. This exacerbated production can impair synaptic plasticity and diminish levels of neurotrophic factors, such as brain-derived neurotrophic factor (BDNF), thereby adversely affecting neurogenesis and neuronal regeneration processes [26].
Alterations in the levels of pro-inflammatory cytokines, such as interleukin-1 beta (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-α), have been consistently associated with Major Depressive Disorder (MDD) [27]. Activation of inflammatory signaling pathways is observed involving both peripheral immune cells and components of the central nervous system, particularly astrocytes and microglia. Elevated levels of cytokines such as IL-1β, IL-6, and TNF-α are consistently detected in patients with depression, reinforcing the role of an activated immune response as a biological marker of the disorder [28] (Figure 2).
Despite the presence of endogenous antioxidant mechanisms such as glutathione, glutathione peroxidase, heme oxygenase-1, and thioredoxin reductase, these systems exhibit limitations when confronted with sustained oxidative overload. Astrocytes, the primary producers of glutathione in the central nervous system via the xc– antiporter system (cystine/glutamate), play a central role in neuronal protection. Nevertheless, neurons, which rely on this glutathione for redox defense, become highly vulnerable in contexts of glial dysfunction [29].
Simultaneously, oxidative stress (OS), characterized by the accumulation of reactive oxygen species (ROS), emerge as a key component in the pathophysiology of Major Depressive Disorder (MDD). It has been identified as a precipitating factor for the inflammatory response, intensifying cytokine production and glial activation. The brain, due to its high metabolic rate, elevated oxygen consumption, and abundance of unsaturated fatty acids, is particularly vulnerable to oxidative damage. In this context, excessive ROS can induce cellular injury through lipid peroxidation of proteins and nucleic acids, as well as trigger intracellular inflammatory mechanisms such as inflammasome activation in microglia, which further amplifies the release of inflammatory cytokines, including IL-1β, TNF-α, and IFN-γ [30].
In preclinical studies, a widely employed experimental model to investigate this relationship is the systemic administration of lipopolysaccharide (LPS), which induces a depressive-like phenotype in animals. LPS promotes astrocyte activation, increasing the expression of inflammatory markers such as inducible nitric oxide synthase (iNOS), TNF-α, IL-1β, and glial fibrillary acidic protein (GFAP) in the hippocampus and cerebral cortex regions [31].
Furthermore, neurotrophic factors, particularly brain-derived neurotrophic factor (BDNF), have emerged as central elements in understanding the pathophysiology of Major Depressive Disorder (MDD) and in mediating response to antidepressant treatment. BDNF is a key regulator of neuronal survival, growth, and differentiation, essential for maintaining synaptic plasticity and hippocampal neurogenesis—processes frequently impaired in individuals with depression [32].
Studies indicate that patients with Major Depressive Disorder (MDD) often exhibit reduced levels of BDNF and other neurotrophic factors, such as glial cell line-derived neurotrophic factor (GDNF), particularly in critical brain regions like the hippocampus. The decreased expression of these factors is associated with impaired adult neurogenesis and synaptic dysfunction, directly contributing to the cognitive and emotional deficits observed in depression. The BDNF receptor, TrkB (tropomyosin receptor kinase B), also plays a pivotal role in transducing neuroprotective signals. Therefore, activation of the BDNF-TrkB pathway is considered a key mechanism underlying the therapeutic effects of various antidepressants [32].
Although BDNF has traditionally been attributed to neurons, recent studies have confirmed that astrocytes are also significant sources of this factor. In primary cultures and murine models, astrocytic overexpression of BDNF in the hippocampus was sufficient to promote neurogenesis and induce anxiolytic-like behaviors, representing a promising area of research for depression treatment [33].
Furthermore, BDNF released by astrocytes in response to chronic antidepressant treatment contributes to synaptic plasticity by regulating neurotransmitter release, vesicular coupling, and the expression of synaptic proteins, thereby promoting enduring changes in neural connectivity [33].
Another relevant growth factor is vascular endothelial growth factor (VEGF), traditionally recognized for its role in angiogenesis. However, recent findings demonstrate that VEGF also influences neural cell function, exerting direct effects on hippocampal neurogenesis and neuroprotection. Activation of tyrosine kinase receptors via VEGF signaling pathways suggests a potential modulatory role of this factor in stress response and functional restoration of the limbic circuitry under depressive conditions [34].
Among the most relevant components involved in neurotrophic factors is the activation of the NLRP3 inflammasome — an intracellular inflammatory platform composed of the proteins NLRP3 (nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin domain-containing protein 3), pro-caspase-1, and ASC (apoptosis-associated speck-like protein containing a CARD).
Activation of this inflammasome plays a critical role in mediating neuroinflammation associated with Major Depressive Disorder (MDD). Evidence suggests that sensitization of the NLRP3 inflammasome is directly linked to the suppression of BDNF synthesis, contributing to neurotrophic dysfunction and, consequently, to the manifestation of depressive symptoms [35].
Furthermore, activation of the NLRP3 inflammasome in astrocytes induces a pro-inflammatory response characterized by increased production of the cytokines IL-1β and IL-18, thereby amplifying the inflammatory cascade and reinforcing the pathological cycle linking neuroinflammation, synaptic dysfunction, and behavioral alterations. This pathological axis positions astrocytes as key elements not only in maintaining cerebral homeostasis but also in actively mediating chronic neuroinflammation and its repercussions on affective behavior [34].
1.3. Conventional Therapeutic Approaches and Their Limitations
Major Depressive Disorder (MDD) is a highly prevalent psychiatric condition associated with significant morbidity. It is estimated that more than 280 million people worldwide are affected by this disorder, which is characterized by depressed mood, anhedonia, sleep disturbances, appetite changes, and impaired ability to perform daily activities. Although not the only treatment modality, pharmacotherapy plays a central role in managing moderate to severe cases. Over the past 50 years, significant progress has been made in the development of antidepressants, with advances in both selectivity and safety profiles [36,37].
Pharmacological treatment of Major Depressive Disorder (MDD) is complex and multifactorial, requiring clinicians to have a detailed understanding of antidepressants’ mechanisms of action, pharmacokinetics, and safety profiles. Drug selection must be individualized, considering comorbidities, prior treatment response history, and risk of adverse effects. Studies such as those by Cipriani et al. [38] and Cui et al. [34] which comparatively analyze the efficacy and acceptability of various antidepressants, remain valuable tools for guiding evidence-based clinical practice.
Antidepressants exhibit a broad pharmacokinetic spectrum, influencing both clinical selection and dosing regimens. Hepatic metabolism via cytochrome P450 isoenzymes is a critical factor, as it affects drug interactions and individual therapeutic response. Pharmacogenetics, particularly involving CYP2D6, has also gained prominence in treatment personalization [39,40].
In this regard, selective serotonin reuptake inhibitors (SSRIs) are considered first-line drugs in the treatment of Major Depressive Disorder (MDD) due to their efficacy and lower side effect profile compared to older antidepressants. The most used drugs include fluoxetine, sertraline, paroxetine, citalopram, and escitalopram. SSRIs selectively block the serotonin transporter (SERT) in the synaptic cleft, inhibiting reuptake of this neurotransmitter and increasing its postsynaptic availability [41].
Generally, they exhibit good oral absorption, high plasma protein binding, and hepatic metabolism via the cytochrome P450 family, especially CYP2D6 and CYP3A4. Fluoxetine stands out due to its long half-life (1 to 3 days for fluoxetine and 7 to 15 days for its active metabolite, norfluoxetine), which supports once-daily dosing. However, sexual dysfunction, nausea, insomnia, weight gain, and, in some cases, serotonin syndrome when combined with other serotonergic drugs, are examples of adverse reactions that may hinder treatment adherence [42].
Another drug class used is serotonin-norepinephrine reuptake inhibitors (SNRIs), with venlafaxine, duloxetine, and desvenlafaxine being the main representatives. Their efficacy is related to the simultaneous action on serotonin (SERT) and norepinephrine (NET) transporters, increasing synaptic levels of both neurotransmitters. Venlafaxine exhibits low protein binding and is primarily metabolized by hepatic CYP2D6, whereas duloxetine has a half-life of approximately 12 hours and is metabolized by CYP1A2 and CYP2D6. Adverse reactions observed include dose-dependent hypertension (mainly with venlafaxine), nausea, dry mouth, sweating, and insomnia. Sexual dysfunction may also occur [43].
Tricyclic antidepressants (TCAs), represented by amitriptyline, nortriptyline, and imipramine, were the first antidepressants developed and, although effective, are currently less used due to their less favorable safety profile. Their mechanism of action is based on the non-selective inhibition of serotonin and norepinephrine reuptake, as well as blocking cholinergic, histaminergic, and adrenergic receptors. Thus, the nonspecific reuptake blockade and interference with cholinergic, histaminergic, and adrenergic pathways result in more intense adverse reactions. Such reactions include sedation, orthostatic hypotension, constipation, urinary retention, blurred vision, and an increased risk of cardiac toxicity in cases of overdose. Regarding pharmacokinetic characteristics, TCAs have high bioavailability, extensive plasma protein binding, and hepatic metabolism via CYP2D6. Half-life ranges from 12 to 36 hours depending on the substance [44].
Although less commonly used today, monoamine oxidase inhibitors (MAOIs) are indicated for treatment-resistant cases, with phenelzine, tranylcypromine, and moclobemide being the main representatives. These agents act by inhibiting the monoamine oxidase (MAO) enzyme, which is responsible for the degradation of monoaminergic neurotransmitters (serotonin, norepinephrine, and dopamine), thereby increasing their synaptic availability. Phenelzine and tranylcypromine have short half-lives but prolonged therapeutic effects due to irreversible MAO inhibition. Moclobemide, in contrast, is a reversible MAOI with a lower risk of dietary interactions. Excess neurotransmitter levels in the synaptic cleft are responsible for common adverse effects, including orthostatic hypotension, weight gain, insomnia, and the risk of hypertensive crisis when combined with tyramine-rich foods (the "tyramine effect"), as tyramine is also metabolized by MAO [45].
Despite the broad therapeutic arsenal currently employed for depression treatment, symptom persistence and failure to achieve remission remain common. Furthermore, antidepressants present several drawbacks. Modern agents such as selective serotonin reuptake inhibitors (SSRIs) are associated with side effects such as nausea, headaches, insomnia, dizziness, sexual dysfunction, and weight gain. These symptoms frequently contribute to early treatment discontinuation by patients [46].
Recent studies have questioned the efficacy of antidepressants in mild to moderate depression, suggesting that the benefits may be limited and not superior to non-pharmacological interventions such as psychotherapy and lifestyle changes. Additionally, antidepressant discontinuation can be challenging due to withdrawal symptoms, which include dizziness, flu-like symptoms, sleep disturbances, and sensory alterations [47].
In addition, studies have shown that individuals with these disorders frequently exhibit elevated levels of pro-inflammatory cytokines, such as interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α), suggesting a link between systemic inflammation and depressive symptoms. Inflammation, or the inflammatory response, results from immune system activation and is typically a localized reaction to irritation, injury, or infection. This response is characterized by heat, redness, swelling, and pain, and may also cause fever. It is essential for eliminating harmful agents and restoring tissue homeostasis. Various immune cells and mechanisms are involved in this process; however, dysregulation of inflammatory responses often contributes to disease development, with increasing evidence implicating inflammation in psychiatric disorders such as depression [48].
The innate immune system represents the first line of defense and comprises cells such as macrophages/monocytes, dendritic cells, and innate lymphoid cells, which mount rapid responses. Receptors on these cells detect damage-associated molecular patterns (DAMPs) and pathogen-associated molecular patterns (PAMPs). DAMPs, also known as alarmins, are endogenous molecules released by damaged cells (e.g., heat shock proteins), whereas PAMPs are molecular structures derived from infectious microorganisms. Upon detection, macrophages and dendritic cells become activated and release pro-inflammatory cytokines, which recruit other immune cells to the site of injury or infection. As part of the inflammatory response, dendritic cells also initiate the adaptive immune reaction by presenting antigens to T and B lymphocytes, which is why they are referred to as antigen-presenting cells (APCs) [49].
Similar to what occurs in peripheral tissues, the brain contains specialized immune cells known as microglia, which constitute approximately 5%–10% of the total brain cell population and perform functions analogous to those of macrophages. Recent findings have revealed that microglia are critically involved in synaptic modulation, including neurogenesis, and are activated in various neurodegenerative and neuropsychiatric disorders, contributing to the development of pathology by promoting neuroinflammation [6].
Microglial cells, the resident immune cells of the central nervous system (CNS), are the primary mediators of neuroinflammation. Under physiological conditions, they play a key role in maintaining neural homeostasis through the clearance of cellular debris and immune surveillance. However, in response to pathological stimuli such as neuronal injury or chronic stress, microglia become activated and release a range of pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6, as well as signaling molecules such as nitric oxide (NO) and prostaglandins. These inflammatory mediators induce changes in the neuronal microenvironment, disrupting synaptic function, promoting neuronal dysfunction, and potentially leading to cell death [50].
Beyond microgliocytes, astrocytes play a crucial role in neuroinflammation. Under chronic inflammatory conditions, astrocytes may become activated, leading to excessive glutamate release, which can result in neuronal excitotoxicity. This dysregulated glutamate release increases neuronal excitability and may induce cell death, particularly in brain regions involved in cognitive and emotional processing, such as the hippocampus and prefrontal cortex [51].
Another important mechanism related to neuroinflammation is the alteration of neuronal energy metabolism. Activation of the inflammatory pathway leads to reduced mitochondrial function, impairing ATP production and contributing to oxidative stress. This stress can trigger a vicious cycle of inflammation and cell death, which has been associated with diseases such as Alzheimer’s and Parkinson’s, as well as psychiatric disorders like depression and schizophrenia [51].
It has been documented that pro-inflammatory cytokines and acute-phase proteins are elevated in patients with Major Depressive Disorder (MDD), with increased levels of IL-6, IL-10, IL-12, IL-13, IL-18, TNF, and C-reactive protein (CRP) observed in the blood of depressed individuals compared to healthy subjects [52,53]. These cytokines may promote a cascade of neurochemical changes, leading to altered brain connectivity and greater vulnerability to stress. Notably, inflammatory markers have been found to correlate with symptom severity, suggesting that neuroinflammation may serve both as a cause and a consequence of these conditions [54].
In depression, the inflammatory process is believed to affect key brain regions such as the prefrontal cortex, hippocampus, and amygdala. These regions are involved in emotional regulation, memory, and stress response. Chronic inflammation in these areas has been shown to impair neuroplasticity, disrupt neurotransmitter systems (such as serotonin, dopamine, and glutamate), and increase neuronal excitability, all of which are implicated in mood disorders [55].
Given the presence of neuroinflammation, it has been questioned whether successful conventional treatment should consider reducing the inflammatory process. Although no currently approved antidepressant treatment was originally designed to modulate the immune response, there is evidence that conventional antidepressants exert significant anti-inflammatory effects. For example, selective serotonin reuptake inhibitors (SSRIs) reduce levels of IL-1β, IL-6, and TNF (Wang et al., 2019). Conversely, other studies have shown that antidepressants such as serotonin-norepinephrine reuptake inhibitors (SNRIs) may induce the production of IL-6 and TNF [56,57,58].
These findings suggest that the effects of conventional antidepressants on cytokine regulation remain underexplored, although they are generally believed to shift the immune balance toward an anti-inflammatory profile. On the other hand, elevated levels of pro-inflammatory cytokines—particularly TNF—have been observed in treatment-resistant depressed patients, suggesting a negative correlation between therapeutic response and pro-inflammatory cytokine levels [5].
The interaction between neuroinflammation and psychiatric symptoms highlights the therapeutic potential of targeting inflammation in the treatment of depression. Anti-inflammatory agents, whether pharmacological interventions or lifestyle-based approaches (e.g., diet and exercise)—are being explored as adjuncts to conventional antidepressant treatments, aiming to improve patient outcomes by addressing the underlying inflammatory processes. In this sense, neuroinflammation represents a promising avenue for understanding the biological basis of depression. As research continues to elucidate the mechanisms linking inflammation to mood regulation, there is growing hope for the development of more effective treatments targeting these pathways, offering relief to individuals affected by these widespread mental health conditions [59].
Considering this evidence, it becomes essential that future treatment strategies for persistent depression include agents with specific actions on central nervous system inflammatory mechanisms, particularly those that modulate microglial activity, such as pro-inflammatory cytokine inhibitors (e.g., anti-TNF-α), antioxidants, and glutamate receptor (NMDA) modulators. This represents a paradigm shift, integrating neuroimmunology into clinical psychiatry and opening new perspectives for managing treatment-resistant depressive disorders [9,60].
2. Materials and Methods
The present study conducted a comprehensive narrative review of scientific literature sourced from reputable databases, including Google Scholar, ScienceDirect, and PubMed. The search strategy employed a combination of relevant keywords such as "Major Depressive Disorder," "Neuroinflammation," "Microglia," and "Nanotechnology" to ensure a broad yet focused examination of existing research. This narrative review aimed to explore how nanotechnological approaches can modulate microglial activity as a means to alleviate neuroinflammation associated with major depressive disorder (MDD). Selected studies were critically analyzed to identify current advances in nanocarrier systems—such polymeric nanoparticles, solid lipid nanoparticle, magnetic nanoparticle, dendrimers, and liposome that demonstrate potential in crossing the blood-brain barrier and selectively targeting activated microglia. The review further assessed how these nanotechnological strategies influence inflammatory signaling pathways. The delivery routes and transport mechanisms explored include the use of nanocarriers capable of crossing the blood–brain barrier (BBB) via receptor-mediated transcytosis (RMT), adsorptive-mediated transcytosis (AMT), and carrier-mediated transport (CMT). Additionally, approaches involving the pharmacological or genetic modulation of efflux transporters, such as P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP), were considered to enhance brain bioavailability. Several delivery platforms were analyzed, including polymeric nanoparticles, solid lipid nanoparticles (SLNs), nanostructured lipid carriers (NLCs), dendrimers, magnetic nanoparticles, liposomes, exosomes, nanoemulsions, self-emulsifying drug delivery systems, and thermosensitive hydrogels, some of which are suitable for intranasal administration. These technologies were evaluated for their ability to overcome the BBB, modulate inflammatory pathways—such as the NLRP3 inflammasome and Toll-like receptors (TLRs)—and promote microglial polarization from the pro-inflammatory (M1) to the anti-inflammatory (M2) phenotype.—highlighting their therapeutic implications in restoring neuroimmune homeostasis in MDD. Therefore, this study conducted an in-depth analysis of the mechanisms by which nanoparticle-based delivery systems interact with neuroimmune targets to reduce microglial-mediated neuroinflammation.
3. Results
3.1. Brain-Targeted Drug Delivery Systems
The delivery of pharmacological agents to the central nervous system (CNS) remains one of the most critical challenges in the treatment of neuropsychiatric disorders, particularly major depressive disorder (MDD). Despite the growing understanding of the neuroinflammatory mechanisms involved in depression, therapeutic interventions are frequently limited by the restricted accessibility of systemically administered drugs to the brain (Wu et al., 2023). This limitation is primarily attributed to the properties of the blood-brain barrier (BBB), a highly specialized interface that regulates molecular exchange between the bloodstream and the brain parenchyma. While the BBB serves a vital homeostatic and neuroprotective role, it also severely limits the brain bioavailability of therapeutic compounds, rendering many potentially effective agents clinically ineffective in CNS disorders, including treatment-resistant depression [61].
Structurally, the BBB is composed of a continuous monolayer of brain microvascular endothelial cells (BMECs), sealed together by tight junctions formed by transmembrane proteins such as claudin-5, occludin, and junctional adhesion molecules, supported intracellularly by scaffolding proteins like ZO-1 [62]. These junctions drastically restrict the paracellular movement of hydrophilic or high-molecular-weight molecules. In contrast to peripheral endothelial cells, BMECs exhibit minimal pinocytosis, complete absence of fenestrae, a high density of mitochondria, and a unique expression pattern of transporters and enzymes, making them uniquely selective [63,64].
The BBB is supported by a complex neurovascular unit composed not only of BMECs but also pericytes, astrocytic end-feet, microglia, the extracellular matrix, and perivascular macrophages. Pericytes, which share the basement membrane with endothelial cells, are particularly abundant in the CNS and play critical roles in microvascular stability, regulation of cerebral blood flow, angiogenesis, and immune surveillance through phagocytic properties. Astrocytes, the most abundant glial cells in the brain, cover approximately 99% of the capillary surface with their end-feet, secreting factors that regulate BBB permeability and participating actively in the control of cerebral metabolism [65,66]. Perivascular macrophages and resident microglia constitute the primary immunological defense at the barrier, capable of responding to systemic or local insults by releasing pro-inflammatory cytokines and reactive oxygen species. In pathological states such as chronic stress or systemic inflammation, excessive activation of these immune components can degrade tight junction proteins, increase BBB permeability, and compromise its selective function [67,68].
The selective permeability of the BBB is governed by both physical and biochemical mechanisms. A major biochemical limitation is the presence of ATP-binding cassette (ABC) efflux transporters, including P-glycoprotein (P-gp/ABCB1) and breast cancer resistance protein (BCRP/ABCG2), which are abundantly expressed on the luminal surface of BMECs. These proteins actively extrude a broad spectrum of structurally diverse drugs from the endothelial cytoplasm back into the bloodstream, limiting intracerebral drug accumulation [69]. Structurally, P-gp is composed of two transmembrane domains and two nucleotide-binding domains that hydrolyze ATP to power the efflux of substrates—primarily lipophilic, neutral, or weakly cationic molecules [70]. BCRP plays a complementary role, particularly in conditions where P-gp is inhibited or saturated, and is more selective for polar and amphiphilic compounds [71].
Importantly, many antidepressants, including fluoxetine, sertraline, citalopram, and venlafaxine, are substrates of these efflux systems. For instance, venlafaxine’s CNS penetration is influenced by P-gp activity, and interindividual variability in treatment response has been linked to genetic polymorphisms in the ABCB1 gene or to drug interactions that modulate transporter expression [72]. This contributes to the heterogeneous clinical outcomes observed in MDD patients undergoing conventional pharmacotherapy [73].
In addition to efflux systems, the BBB also functions as a metabolic barrier equipped with a variety of enzymes that further limit the bioavailability of therapeutics intended for the CNS. Among these enzymes, cytochrome P450 isoforms (e.g., CYP3A4 and CYP2D6), monoamine oxidases (MAO-A and MAO-B), esterases, and peptidases are prominently expressed in BMECs [74,75]. These enzymes catalyze the biotransformation of xenobiotics and endogenous substrates during their transendothelial passage, often resulting in the premature degradation or inactivation of therapeutic agents.
The ability of a drug to cross the BBB is also influenced by its physicochemical properties. Lipophilicity is a major determinant, as lipophilic molecules readily diffuse through the lipid bilayers of endothelial cell membranes. However, other features—such as molecular weight, charge, and hydrogen bonding capacity—also play critical roles. According to Lipinski’s Rule of 5, compounds with high molecular weight (>500 Da), more than five hydrogen bond donors, or more than ten hydrogen bond acceptors, tend to exhibit poor membrane permeability. While originally devised for oral bioavailability, this rule has also been applied, with caveats, to BBB permeability [76,77]. Notably, some antidepressants fit within the favorable physicochemical window (e.g., low molecular weight, moderate lipophilicity, low hydrogen bonding potential), promoting passive diffusion across the BBB. In contrast, drugs classified as “beyond the Rule of 5” (bRo5) may require chemical modifications or carrier-based delivery systems to enhance CNS penetration [78,79]. Carrier-mediated transport (CMT) also plays a vital role in the supply of essential nutrients to the brain. Specific membrane-bound proteins facilitate the transport of glucose (via GLUT1), amino acids, nucleotides, vitamins, and small peptides across BMECs. These transporters operate either by facilitated diffusion or by active transport, using ATP or ionic gradients to mediate substrate influx [80].
Critically, BBB function is dynamic and susceptible to pathological modulation. In MDD, chronic stress, systemic inflammation, and dysregulated cytokine signaling have been implicated in BBB disruption. Preclinical studies report downregulation of claudin-5, increased paracellular permeability, and upregulation of endothelial adhesion molecules such as VCAM-1 and ICAM-1, facilitating peripheral leukocyte infiltration and promoting neuroinflammatory cascades [81,82]. Although this dysfunction may transiently increase drug penetration, it is typically associated with neuronal damage, microglial overactivation, and worsening of depressive pathology, rendering it therapeutically undesirable.
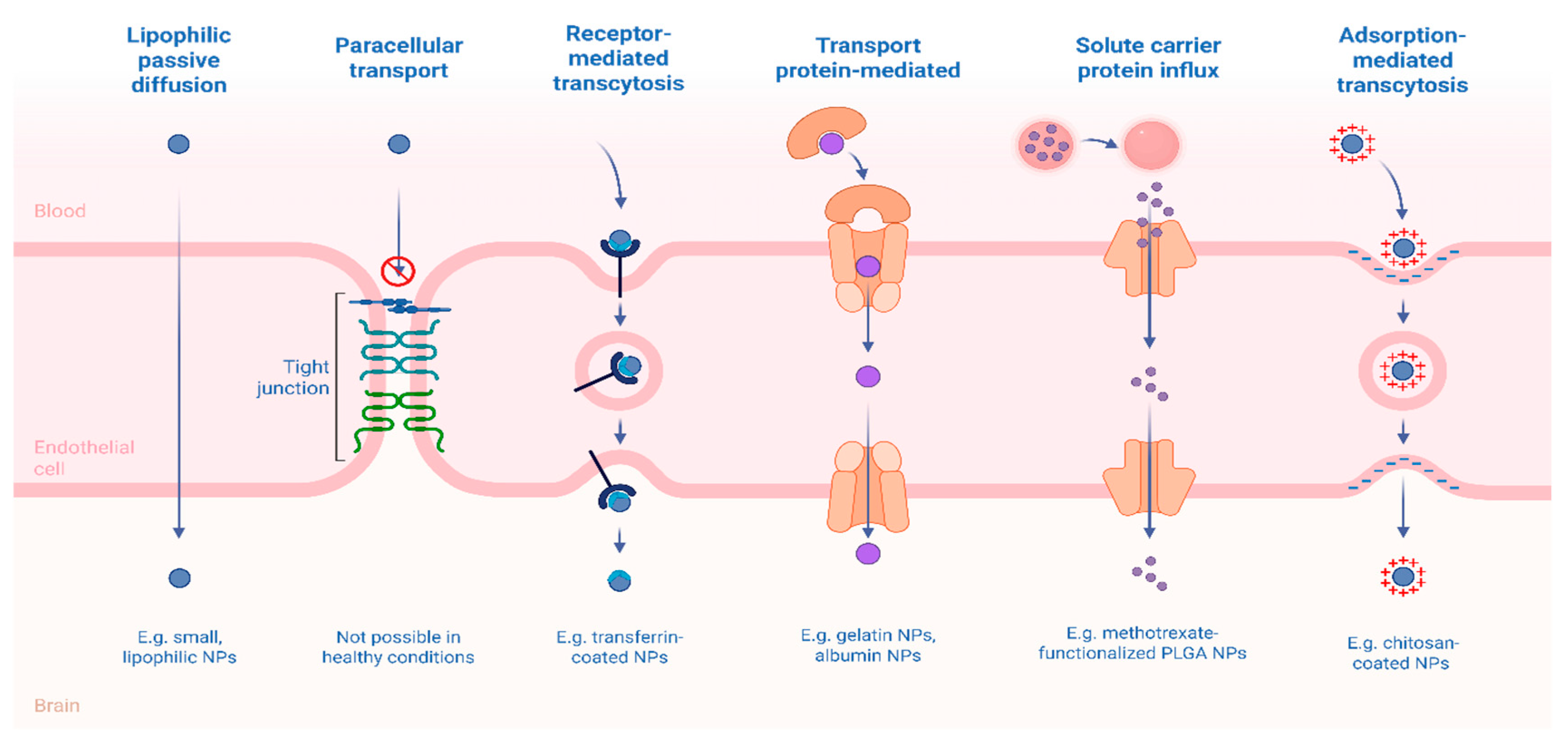
These challenges have encouraged the development of innovative strategies designed to bypass or modulate the BBB. Among these, receptor-mediated transcytosis (RMT) has emerged as a leading approach, whereby nanocarriers are functionalized with ligands targeting overexpressed endothelial receptors, triggering endocytosis and enabling transcellular transport of drug-loaded nanoparticles into the brain parenchyma [83]. Adsorptive-mediated transcytosis (AMT), which leverages electrostatic interactions between cationic surfaces of nanocarriers and negatively charged endothelial membranes, offers a less specific but still viable route of drug delivery [84]. Furthermore, the pharmacological, genetic, or technological modulation of BBB efflux transporters presents a promising complementary strategy, enabling the transient and controlled enhancement of CNS drug delivery (Figure 3).
3.2. Controlled Release Systems
In the pursuit of improving therapeutic outcomes for central nervous system (CNS) disorders such as major depressive disorder (MDD), controlled release systems have emerged as transformative tools in neuropharmacology. Unlike conventional formulations that often suffer from rapid clearance, erratic absorption, and non-specific biodistribution, controlled release systems are engineered to deliver therapeutic agents at a predetermined rate, location, and duration [85]. This pharmacokinetic control is particularly relevant in the context of CNS drug delivery, where crossing the blood-brain barrier (BBB) is a fundamental obstacle and where fluctuations in drug levels can exacerbate neurotoxicity or diminish efficacy [86].
Among the most promising technologies in this domain are nanocarriers—engineered systems typically ranging from 1 to 1000 nm—that can be tailored in terms of size, surface properties, composition, and degradation kinetics to optimize delivery to the CNS. Nanoparticles, particularly those based on polymeric and lipidic materials, have shown considerable potential in this regard [87]. Controlled release from these systems is often achieved through diffusion, degradation, swelling, or environmental responsiveness to stimuli such as pH, redox potential, or enzymatic activity. This versatility allows for synchronization of drug release with pathological cues within the CNS microenvironment [88].
In addition to size, nanoparticle morphology plays a crucial role. Rod-shaped nanoparticles demonstrate enhanced adhesion to endothelial surfaces and increased circulation time compared to spherical particles. Particles with sizes ranging between 50 and 100 nm have shown superior ability to cross the BBB [89]. Surface charge is also critical moderately or highly negative charges (from -1 to -45 mV) are commonly associated with successful BBB permeation, whereas strongly positive nanoparticles may induce BBB toxicity. However, in some cases, cationic surfaces facilitate transport due to electrostatic interactions with the negatively charged glycoproteins present on the BBB [89].
The advantages of controlled release in brain-targeted therapies lie not only in the protection of drugs by encapsulation but also in the capacity of these systems to maintain therapeutic concentrations within the brain over extended periods while minimizing peripheral exposure [85]. This sustained presence is essential not only for drugs with short half-lives or narrow therapeutic indices but also for modulating chronic neuroinflammatory pathways, which are increasingly recognized as contributors to the pathogenesis of MDD [61]. By modulating release kinetics, nanosystems can reduce dosing frequency, enhance patient adherence, and allow for precise spatial-temporal drug targeting. Furthermore, many of these platforms are adaptable to surface functionalization, enabling integration with brain-targeting ligands that exploit receptor-mediated transcytosis [90].
Several ligands have been explored to enhance targeting efficiency, including glycoproteins (e.g., transferrin, lactoferrin), peptides (e.g., glutathione, Angiopeps, cell-penetrating peptides), vitamins (e.g., folate, thiamine), and carbohydrates (e.g., mannose) [91,92]. Surface modification with such ligands not only enhances BBB penetration but also promotes drug accumulation at the target site, reducing off-target toxicity, minimizing dosing, and limiting the development of drug resistance [93].
Among the most widely investigated nanosystems for controlled drug release are polymeric nanoparticles, lipid-based systems, and dendrimers. Each platform possesses unique structural and functional properties that can be strategically exploited to optimize brain-targeted therapy. Polymeric nanoparticles, for instance, are colloidal systems typically ranging from 100 to 500 nm, composed of natural or synthetic biodegradable polymers such as poly(lactic-co-glycolic acid) (PLGA) and chitosan [94]. PLGA is FDA-approved, highly biocompatible, and degrades hydrolytically into lactic and glycolic acid, which are naturally metabolized in the Krebs cycle. Its degradation rate—and therefore drug release profile—can be precisely tuned by altering the polymer’s molecular weight and the lactic-to-glycolic acid ratio [95]. This tunability makes PLGA nanoparticles particularly attractive for sustained CNS drug delivery, enabling controlled drug exposure over hours to weeks, depending on formulation parameters.
Chitosan, on the other hand, is a natural polysaccharide derived from chitin. It is mucoadhesive, positively charged under physiological conditions, and biodegradable. Chitosan-based nanoparticles are especially useful for intranasal delivery routes targeting the brain, as they can adhere to mucosal surfaces, prolong residence time, and facilitate transport via the olfactory or trigeminal nerves [96]. Furthermore, their amine-rich surface can be easily functionalized with targeting ligands or polyethylene glycol (PEG) to enhance BBB penetration and systemic circulation stability [97].
Lipid-based nanocarriers cover a broad category, but solid lipid nanoparticles (SLNs) and nanostructured lipid carriers (NLCs) have garnered particular attention for brain-targeted controlled release. SLNs consist of a solid lipid core stabilized by surfactants, in which the drug is either dispersed or embedded, while NLCs represent a second-generation advancement over SLNs, being composed of a mixture of solid and liquid lipids, creating a less ordered lipid matrix that improves drug loading capacity and reduces drug expulsion during storage [98]. NLCs exhibit superior control over release kinetics due to their semi-crystalline structure and can be optimized to avoid burst release (Graván et al., 2023). These lipid particles are particularly suitable for brain delivery because they can be administered via parenteral or intranasal routes, and their lipid composition favors interaction with cell membranes and potential uptake via endocytosis mechanisms [99].
Dendrimers are nanoscale, highly branched polymers with a globular, tree-like structure. They consist of a central core, an internal branching architecture (called generations), and numerous surface functional groups. These surface termini can be modified with hydrophilic polymers (such as PEG), targeting moieties, or drugs themselves. Internally, dendrimers can encapsulate hydrophobic drugs within their void spaces via hydrophobic or electrostatic interactions, while simultaneously carrying hydrophilic drugs or ligands on their surface. This dual-loading capacity allows for multifunctional drug delivery, combining therapeutic, targeting, and imaging functions in a single system [100].
One of the most studied dendrimer types is poly(amidoamine) (PAMAM) dendrimers, which have well-defined molecular weights and surface chemistries. They offer predictable pharmacokinetics and controlled release based on their generation size and surface modifications [101]. In brain delivery, dendrimers can be designed to cross the BBB through surface conjugation with ligands that exploit receptor-mediated transport pathways. Once in the brain, release is often governed by hydrolysis of cleavable bonds (e.g., ester or disulfide linkages) or by passive diffusion from the dendrimer core [101,102].
In preclinical models, the theoretical advantages of controlled release systems have translated into significant improvements in brain bioavailability and behavioral outcomes. For example, Dadkhah et al. (2024) demonstrated that fluoxetine-loaded PEGylated chitosan nanoparticles enhanced cognitive performance, increased hippocampal brain-derived neurotrophic factor levels, and reduced demyelination in a rat model of hippocampal injury, outperforming conventional fluoxetine therapy [103]. Similarly, Neves et al. (2021) developed transferrin-functionalized solid lipid nanoparticles and nanostructured lipid carriers loaded with curcumin, which significantly enhanced its permeability across an in vitro human BBB model [104]. The functionalized systems demonstrated 1.5-fold higher brain endothelial transport compared to non-functionalized nanoparticles and free curcumin, highlighting their potential for targeted and controlled neuroprotective delivery.
Likewise, Eldeeb et al. (2024) reported that co-administration of chitosan and curcumin nanoparticles not only reversed hydroxyapatite-induced neurotoxicity in rats but also elevated neurotransmitter and antioxidant levels beyond control values: dopamine by 13% and norepinephrine by 24% [105]. While inflammatory markers remained slightly elevated, mitochondrial biogenesis genes were upregulated up to 27% over control, and histological analysis confirmed substantial neuroprotection, underscoring the synergistic potential of these nanosystems in neuroinflammatory conditions.
In summary, controlled-release nanosystems offer a multifaceted solution to the challenges of CNS drug delivery by simultaneously addressing BBB permeability, pharmacokinetic stability, and site-specific release. Depending on the nanocarrier design, these systems can be tailored to overcome key physiological barriers and achieve effective brain targeting. With advancing research and continued clinical validation, the integration of these nanoparticles into therapeutic protocols for MDD and other neurological disorders may substantially improve efficacy and safety profiles compared to conventional drug formulations.
Figure 4.
Mechanisms of Transport and Delivery of Nanoparticles Across the BBB. Created by Biorrender (2025).
Figure 4.
Mechanisms of Transport and Delivery of Nanoparticles Across the BBB. Created by Biorrender (2025).
3.3. Drug Delivery Strategies for Microglial Modulation
Microglia are important targets for therapeutic interventions; however, their use as targets presents challenges, such as effectively regulating their states to achieve the desired therapeutic effect. Drug delivery systems can deliver specific modulators to microglia, improving the precision and efficacy of interventions [16]. Different nanocarriers show potential for targeted drug delivery for microglial modulation in the treatment of depression, such as polymeric nanoparticles, solid lipid nanoparticles, magnetic nanoparticles, nanostructured lipid carriers, hydrogels, self-emulsifying drug delivery system, nanoemulsion. Table 1 presents a summary of the information found in the literature about the nanosystems used in the modulation of microglia in the treatment of MDD.
Legend: DA stands for dopamine; 5-HT, serotonin; PDI, polydispersity index; PZ, zeta potential; EE, encapsulation efficiency; NMDA, N-methyl-D-aspartate; Res, resveratrol; THS, thermosensitive hydrogel system; MDA, malondialdehyde (an oxidative stress marker); GSH, reduced glutathione (a cellular antioxidant); NLC C, curcumin-loaded lipid nanocapsules; Aβ₂₅₋₃₅, beta-amyloid peptide 25-35; TCA, open field test; TUR, curcuma extract; NE, nanoemulsion; C₅₀, half maximal inhibitory concentration; ABTS•⁺, ABTS cation radical; and CU, curcumin.
3.4. Polymeric Nanoparticles
A variety of nanosystems have been reported to modulate microglial activity. Recent studies have demonstrated that polymeric nanoparticles (NPs) can both inhibit and reprogram the phenotype of these cells, depending on their composition, making them applicable to a range of neurodegenerative diseases, central nervous system injuries, and major depressive disorder [124]. Polymeric nanoparticles (PNs) are solid colloidal particles ranging in size from 1 to 1000 nm, obtained from natural, synthetic or semi-synthetic polymers that can be biodegradable or non-biodegradable. They can be classified as nanosphere or nanocapsule. Nanospheres are matrix particles in which the active pharmaceutical ingredient (API) is adsorbed on the particle surface or dispersed in the particle. On the other hand, nanocapsules are vesicular systems in which the API is confined to a water/oil liquid core surrounded by a polymeric membrane [125,126,127]. Polymeric nanoparticles can provide controlled and targeted drug release, enhanced drug bioavailability and increased therapeutic index [128] (Figure 5).
Jiang et al. (2025) designed memantine-loaded nanoparticles incorporating polydopamine, a self-adhesive biomimetic polymer, coated with BV2 microglial cell membranes (PDA-Mem@M) [109]. Memantine is an NMDA receptor antagonist that reduces brain excitotoxicity, potentially modulating microglial activity. The PDA-Mem@M formulation exhibited a size of 163.5 ± 1.5 nm and a zeta potential of -54.3 ± 2.2 mV, with rapid memantine release under acidic conditions (48% release within 30 min at pH 6.8), relevant to the inflammatory brain microenvironment. In vitro assays showed no significant cytotoxicity in SH-SY5Y (human neuroblastoma) and bEnd.3 (murine brain endothelial) cells, maintaining over 80% cell viability at high concentrations (200 µg/mL). Additionally, PDA-Mem@M significantly reduced intracellular ROS levels in LPS-induced cellular models and polarized microglia from a pro-inflammatory M1 phenotype (reduction in CD86, TNF-α, and IL-2) to an anti-inflammatory M2 phenotype (increase in CD206, TGF-β, and IL-10) in BV2 cells.
In vivo studies over 21 days in mice subjected to a chronic unpredictable stress (CUS) model demonstrated that PDA-Mem@M exhibited enhanced brain and hippocampal targeting (1.3-fold higher fluorescence than PDA) after 4 hours post-injection. PDA-Mem@M outperformed memantine monotherapy at lower and less frequent doses, indicating a faster onset of action and greater efficacy, with minimal observed toxicity. Furthermore, the nanoformulation reduced hippocampal ROS and pro-inflammatory cytokines (IL-1β and IL-2), while increasing anti-inflammatory cytokines (IL-10), stimulating neurogenesis, and exerting neuroprotective effects by reducing neuroinflammation and restoring synaptic plasticity [109].
Beyond synthetic polymeric systems, endogenous carriers like exosomes offer another promising approach to targeted microglial modulation due to their inherent biocompatibility and natural ability to cross the BBB. They have been employed in nanoformulations to deliver molecules like celastrol and minocycline (anti-inflammatory and neuroprotective agents) across the BBB to resident brain cells [129]. Lv et al. (2024) developed PLGA nanoparticles loaded with exosomes, celastrol, and minocycline (CMC-EXPL), exhibiting a size of 132 nm and a zeta potential of -35.5 mV [106]. In BV2 microglial cells, the nanoformulation achieved 65.8% uptake within 270 minutes. Cocultures of SH-SY5Y neurons and LPS-stimulated BV2 microglia (induced to an M1 phenotype) revealed that CMC-EXPL significantly suppressed M1 polarization (reduction in CD80 and iNOS) and promoted M2 polarization (increase in CD206 and Arg1), showing superior anti-inflammatory therapeutic potential compared to other tested formulations.
The animal model of post-stroke depression (POSD), induced by middle cerebral artery occlusion (MCAO) followed by chronic unpredictable mild stress (CUMS), was used for CMC-EXPL administration. The nanoformulation significantly attenuated weight loss and reversed depressive-like behaviors. M1 microglial markers (iNOS and CD86) were elevated in MCAO and POSD groups, while M2 markers (Arg-1 and CD206) were reduced. Treatment with CMC-EXPL significantly downregulated M1 markers and upregulated M2 markers, indicating enhanced microglial polarization toward the M2 phenotype in POSD rats, contributing to its antidepressant efficacy [106]. This strategy not only enhanced brain delivery but also effectively reprogramed microglia in complex models, suggesting broader applicability to comorbid neuropsychiatric conditions.
In the study by Nagpal, Singh, and Mishra (2013), the safety and efficacy of chitosan-based polymeric nanoparticles encapsulating minocycline (MHPNs) were evaluated. In vivo results demonstrated that MHNPs exhibited antidepressant effects and were safe, with no observed toxicological effects on the brain, heart, spleen, liver, or kidneys [130].
Similarly, Yusuf et al. (2016) incorporated curcumin into polymeric nanoparticles (CUR-NPs), which showed enhanced antidepressant activity in the Forced Swim Test (FST) and Tail Suspension Test (TST), even at a low dose (5 mg/kg) [107]. CUR-PNs increased brain bioavailability by 1.6 times compared to free curcumin. The in vitro release profile was biphasic, with an initial release of 69.43 ± 6.56% at 24 hours and sustained release reaching 95.56 ± 4.67% at 144 hours, following the Higuchi model (r² = 0.95). CUR-PNs also significantly increased the activity of antioxidant enzymes SOD and catalase, and exhibited strong interaction with MAO-B (ΔG = –7.79 kcal/mol), suggesting an antidepressant effect through multiple mechanisms. These findings highlight the potential of CUR-NPs for controlled release, improved brain penetration, and enhanced therapeutic efficacy.
Yang et al. (2022) investigated the effects of selenium nanoparticles (SePNs) in alleviating depression-like behavior induced by fluoride (F) exposure in mice, focusing on the JAK2–STAT3 signaling pathway [95]. In vivo results showed that SePNs restored dopamine and norepinephrine levels in the cortex, reduced microglial activation and interleukin-1β secretion, inhibited the nuclear translocation of p-STAT3, increased the number of surviving neurons, and reduced cortical vacuolization. These outcomes indicate that SeNPs mitigated fluoride-induced neurotoxicity and depressive-like behaviors, possibly by inhibiting the JAK2–STAT3 pathway.
Zhu et al. (2023) evaluated the antidepressant effects of polydopamine nanoparticles (PDA-PNs, 2.5 mg/kg) in C57BL/6 mice with LPS-induced depression (1 mg/kg, i.p.). PDA-NPs reversed depressive and anxious behaviors, reducing immobility in the TST from 170.8 ± 10.6 s to 95.2 ± 11.6 s and increasing center time in the open field test from 38.7 ± 4.3 s to 68.7 ± 6.2 s. Additionally, PDA-PNs significantly lowered TNF-α levels from 235.8 ± 20.4 to 136.5 ± 9.4 pg/mg and IL-1β from 143.2 ± 17.3 to 83.9 ± 5.9 pg/mg. The PN also reduced splenomegaly, inhibited microglial activation (Iba1), downregulated IL-1β and iNOS expression, upregulated Arg-1, crossed the blood-brain barrier, and reduced brain oxidative stress, thereby demonstrating a neuroprotective effect [131].
3.5. Solid Lipid Nanoparticle
Solid lipid nanoparticles (SLN) are particles with a size of <1000 nm, which have a solid hydrophobic core of biocompatible and biodegradable lipids such as triglycerides, partial glycerides, waxes, steroids, and fatty acids that are solid at normal room temperature. These lipids are stabilized by different biocompatible surfactants that can be ionic or nonionic. SLN offers a viable method for incorporation of hydrophilic or lipophilic drugs and peptides, enhanced drug targeting and bioavailability, controlled drug release, enhanced drug stability, and increased drug payload [132].
Similarly, lipid-based nanosystems such as NLCs have been widely studied for brain delivery due to their favorable size, biocompatibility, and ability to traverse biological membranes [116]. Thus, NLCs loaded with curcumin (CUR-NLCs) were investigated for their neuroprotective effects in a rat model of LPS-induced depression and anxiety. CUR-NLCs exhibited a particle size of 147.8 ± 10.4 nm, surface charge of -32.8 ± 1.4 mV, and an encapsulation efficiency of 91.0 ± 4.6%. In vivo studies indicated an indirect microglial effect, as CUR-NLCs markedly attenuated LPS-induced neurodegeneration, restored tissue architecture and cellular integrity, and increased neuronal survival. Additionally, CUR-NLCs suppressed brain expression of pro-inflammatory markers such as p-NF-κB, TNF-α, and COX-2, thus reducing neuroinflammation [115]. Although the microglial effects were indirect, these results underscore the anti-inflammatory potential of natural compounds like curcumin when properly encapsulated for CNS delivery.
Building on these findings, other antidepressants such as agomelatine have also been encapsulated into NLCs to enhance therapeutic response through inflammatory pathway modulation. For instance, Gul et al. (2022) developed agomelatine-loaded NLCs (AGM-NLCs) with an average size of 99.8 ± 2.6 nm and a charge of 23.2 ± 1.2 mV to enhance in vivo antidepressant potential. Agomelatine supports neuronal integrity and exerts an indirect effect on microglia by reducing inflammation. AGM-NLCs effectively suppressed LPS-induced neuroinflammation, reducing both expression and concentration of TNF-α and COX-2 in the brain. [117]
Rubab et al. (2021) produced curcumin encapsulated nanostructured lipid carriers (CUR-NLCs) with particle size of 147.8 ± 10.4 nm, PDI of 0.27 ± 0.02, zeta potential of –32.8 ± 1.4 mV, and incorporation efficiency of 91.0 ± 4.6%. In vitro release study demonstrated ~27% drug release within 2 h followed by sustained release with ~54% and ~73% curcumin release within 12 h and 24 h, respectively. The results of in vivo studies in rats subjected to a lipopolysaccharide-induced model of depression and anxiety revealed a significant increase in fighting time (95.3 ± 5.1 s) and a reduction in immobility time (87.9 ± 4.3 s) in the forced swimming and tail suspension tests. In addition, there was a substantial increase in the time spent (65.0 ± 2.7 s) and in the number of entries into the illuminated and open compartments (14.7 ± 0.7 s) in the light-dark box and elevated plus maze tests, indicating antidepressant and anxiolytic effects. Histological and immunohistochemical analyses revealed a reduction in brain inflammation with suppression of the expression of inflammatory markers such as p-NFκB, TNF-α and COX-2 in brain tissue [115].
3.6. Magnetic Nanoparticle
Magnetic nanoparticles (MNPs) are nanometer-sized materials that respond to external magnetic fields. They are typically composed of iron oxides such as magnetite (Fe₃O₄) or maghemite (γ-Fe₂O₃), pure metals like Fe, Co, or Ni, or a combination of metals and polymers. These particles may be coated with polymers, lipids, or silica to enhance their stability, biocompatibility, and functionalization. The magnetic behavior of these materials can be tuned not only by external magnetic fields but also by their size, shape, and morphology [133].
Khadrawy et al. (2021) developed iron oxide nanoparticles coated with curcumin (Cur-IONPs) with an average size of 15 ± 3 nm and a zeta potential of −25.4 mV, indicating good stability. In a reserpine-induced rat model of depression, treatment with Cur-IONPs (5 mg/kg for 14 days) significantly reduced immobility time in the forced swim test, restoring active swimming time to near-control levels, suggesting behavioral improvement. Markers of oxidative stress, which were elevated in the cortex (MDA +48.9%, NO +55%, GST +61%, GSH −29%) and hippocampus (MDA +36.1%, NO +233%, GST +29.8%, GSH −16.9%), were normalized after treatment, indicating an antioxidant effect. Enzymatic activities altered in the cortex (AchE +144.7%, Na⁺/K⁺-ATPase +18.2%, MAO +56.6%) and hippocampus (AchE +52.1%, Na⁺/K⁺-ATPase +24.2%, MAO +92.5%) were also restored to values close to those of the control group, suggesting improved neuronal function and neurotransmitter regulation. Regarding neurotransmitter levels, reductions observed in the cortex (5-HT −37.8%, NE −65.9%, DA −49.6%) and hippocampus (5-HT −46.6%, NE −40.7%, DA −32%) were partially reversed, with noradrenaline and dopamine levels restored in the cortex and serotonin in the hippocampus, corroborating the antidepressant effects of the treatment [113].
In the reserpine-induced depression model studied by Fahmy et al. (2023), there was a significant reduction in behaviors such as square crossing frequency (−100%), rearing (−100%), and grooming (−84.75%), along with a 663.3% increase in immobility time and a 91% reduction in struggle time. Treatment with Zn(cur)O and ZnO nanoparticles, as well as free curcumin, attenuated these effects. Grooming behavior increased by up to 90.25% with curcumin and 59.73% with ZnO PNs; immobility time was reduced to 228.9% (Zn(cur)O NPs) and 50% (ZnO PNs). MDA levels were restored to control values, and GSH levels in the cortex, hippocampus, and striatum significantly increased (up to 384.7% with Zn(cur)O NPs). Levels of the neurotransmitters 5-HT and NE were significantly elevated, with hippocampal 5-HT increasing by 146.17% following treatment with Zn(cur)O PNs, indicating a strong antidepressant effect of these compounds [114].
Fahmy et al. (2023) further investigated the antidepressant activity of zinc oxide nanoparticles conjugated with curcumin (ZnO PNs) in adult male Wistar rats with reserpine-induced depression. The nanoparticles had a particle size of 342 ± 22.3 nm and a zeta potential of −25.6 ± 4.61 mV. In vivo results showed improved motor activity and reduced immobility time. Additionally, a decrease in malondialdehyde (MDA) levels and an increase in reduced glutathione (GSH) and catalase (CAT) levels were observed, along with elevated concentrations of serotonin (5-HT) and norepinephrine (NE) [114].
3.7. Dendrimers
Dendrimers are hyperbranched, radially symmetric macromolecules with a precisely tailored architecture composed of surface functional groups. These surface functional groups can be modified to alter their physicochemical and biological properties [134]. In another study, poly(amidoamine) (PAMAM) dendrimers labeled with Cyanine5 (D-Cy5) were used to investigate microglial migration dynamics and dendrimer interactions in the presence of neuroinflammation. Using organotypic whole-hemisphere brain slice cultures from newborn rabbits, PAMAM uptake (4 nm in size) by microglia was faster and more pronounced in slices from rabbits with cerebral palsy (CP) than in healthy controls (1.6-fold increase after 4 hours, with 80% of microglia containing dendrimers). In vivo studies demonstrated selective accumulation of PAMAM dendrimers in activated microglia in the brains of newborn rabbits. When conjugated with N-acetylcysteine (NAC), the formulation significantly improved motor function and attenuated neuroinflammation. These findings suggest that increased dendrimer uptake by impaired microglia can be exploited for targeted drug delivery and functional modulation (Zhang et al., 2016) [123].
3.8. Liposome
Liposomes are spherical lipid bilayer vesicles composed primarily of phospholipids. Their structures result from the self-assembly of amphiphiles in an aqueous medium forming single or multiple concentric bilayers, where the polar head groups are in contact with the aqueous medium and the fatty acids form the hydrophobic core of the bilayers that are protected from water. They present several advantages, such as biocompatibility, biodegradability, low toxicity, non-immunogenicity, improved drug solubility, the possibility of co-encapsulation and surface modifications for targeted, controlled and sustained release of payload materials [127,135].
Interestingly, even existing drugs not originally developed for depression have shown antidepressant effects when incorporated in nanocarrier systems, particularly via microglial activation. For example, Gao et al. (2019) investigated the antidepressant potential of commercially available liposomal amphotericin B (L-AmB), an antifungal. Male mice (6–8 weeks old) subjected to CUS or LPS-induced depression were treated with either free AmB (1, 3, or 5 mg/kg, i.p.) or liposomal AmB (0.5, 1, or 3 mg/kg, i.p.). In CUS-exposed mice, L-AmB (1 and 3 mg/kg) dose-dependently reversed depressive-like behaviors—decreased immobility in the Tail Suspension Test (TST) and Forced Swim Test (FST), increased sucrose consumption in the Sucrose Preference Test (SPT), and increased center time in the Open Field Test (OFT)—effects not observed with free AmB. Regarding microglial activation, pretreatment with minocycline (a microglial inhibitor) suppressed the antidepressant effect of L-AmB in CUS-stressed mice, with reduced IL-1β and IL-6 levels. Similarly, PLX3397, a microglial depletor, abolished both the antidepressant and pro-inflammatory effects of L-AmB in treated groups. These findings confirm that modulating microglial activation, rather than simply suppressing it, may be essential to the antidepressant action of certain compounds [120].
4. Conclusions
MDD remains a major global health challenge, largely due to the multifactorial nature of its pathophysiology and the limited efficacy of current pharmacological treatments. Emerging evidence strongly supports the role of microglial activation and neuroinflammation as key drivers of treatment resistance, highlighting the need for therapeutic strategies that go beyond monoaminergic modulation. Nanotechnology offers a unique opportunity to address these challenges by enabling targeted drug delivery, controlled release, and co-delivery of multimodal agents capable of modulating microglial function and inflammatory signaling pathways. Preclinical studies have shown that nanocarriers can improve antidepressant bioavailability, enhance brain penetration, and promote neuroprotective effects through antioxidant and anti-inflammatory actions. While clinical translation remains in its early stages, the integration of nanotechnology with precision medicine approaches holds promise for the development of next-generation therapies for depression. Targeting microglia via nanotechnology emerges as a cutting-edge strategy with the potential to transform the therapeutic landscape of MDD, offering more effective and personalized interventions.
Author Contributions
Conceptualization, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V.; Methodology, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V.; Validation, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V.; Investigation, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V.; Resources, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V.; Data Curation, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V.; Writing – Original Draft, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V.; Writing – Review & Editing, Da Silva, P. R.; Scotti, L.; Da Silva Leite, J. M.; Alves, L. P.; De Andrade, J. C.; Xavier-Júnior, F. H.; Bezerra Felipe, C. F.; Formiga, A. L. D.; Uchôa, A. F. C.; Neri, L. C. D.; De Castro, R. D.; Barbosa, N. M. M. V. Authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) through the postdoctoral fellowship granted for the period 2023–2025.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A Review on the Oxidative and Nitrosative Stress (O&NS) Pathways in Major Depression and Their Possible Contribution to the (Neuro)Degenerative Processes in That Illness. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2011, 35, 676–692. [Google Scholar] [CrossRef]
- Black, C.N.; Bot, M.; Scheffer, P.G.; Cuijpers, P.; Penninx, B.W.J.H. Is Depression Associated with Increased Oxidative Stress? A Systematic Review and Meta-Analysis. Psychoneuroendocrinology 2015, 51, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Kiani, A.K.; Maltese, P.E.; Dautaj, A.; Paolacci, S.; Kurti, D.; Picotti, P.M.; Bertelli, M. Neurobiological Basis of Chiropractic Manipulative Treatment of the Spine in the Care of Major Depression. Acta Bio Medica Atenei Parmensis 2020, 91, e2020006. [Google Scholar] [CrossRef]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef] [PubMed]
- Worthen, R.J.; Beurel, E. Inflammatory and Neurodegenerative Pathophysiology Implicated in Postpartum Depression. Neurobiology of Disease 2022, 165, 105646. [Google Scholar] [CrossRef]
- Yirmiya, R.; Rimmerman, N.; Reshef, R. Depression as a Microglial Disease. Trends in Neurosciences 2015, 38, 637–658. [Google Scholar] [CrossRef]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and Depression: A Review. Eur J of Neuroscience 2021, 53, 151–171. [Google Scholar] [CrossRef]
- Miller, A.H.; Raison, C.L. The Role of Inflammation in Depression: From Evolutionary Imperative to Modern Treatment Target. Nat Rev Immunol 2016, 16, 22–34. [Google Scholar] [CrossRef]
- Miller, A.H.; Haroon, E.; Felger, J.C. Therapeutic Implications of Brain–Immune Interactions: Treatment in Translation. Neuropsychopharmacol 2017, 42, 334–359. [Google Scholar] [CrossRef]
- Zhao, S.S.; Jones, G.T.; Hughes, D.M.; Moots, R.J.; Goodson, N.J. Depression and Anxiety Symptoms at TNF Inhibitor Initiation Are Associated with Impaired Treatment Response in Axial Spondyloarthritis. Rheumatology 2021, 60, 5734–5742. [Google Scholar] [CrossRef]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. The Semantics of Microglia Activation: Neuroinflammation, Homeostasis, and Stress. J Neuroinflammation 2021, 18. [Google Scholar] [CrossRef]
- Zorkina, Y.; Abramova, O.; Ushakova, V.; Morozova, A.; Zubkov, E.; Valikhov, M.; Melnikov, P.; Majouga, A.; Chekhonin, V. Nano Carrier Drug Delivery Systems for the Treatment of Neuropsychiatric Disorders: Advantages and Limitations. Molecules 2020, 25, 5294. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Chen, M.; Zhang, J.; Maincent, P.; Xia, X.; Wu, W. Updated Progress of Nanocarrier-Based Intranasal Drug Delivery Systems for Treatment of Brain Diseases. Crit Rev Ther Drug Carrier Syst 2018, 35, 433–467. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, C.; Silva, S.; Bicker, J.; Falcão, A.; Fortuna, A. Antidepressants and Nose-to-Brain Delivery: Drivers, Restraints, Opportunities and Challenges. Drug Discovery Today 2019, 24, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, T.; Dong, J.; Lu, Y. The Blood–Brain Barriers: Novel Nanocarriers for Central Nervous System Diseases. J Nanobiotechnol 2025, 23. [Google Scholar] [CrossRef]
- Li, H.; Guan, M.; Zhang, N.-N.; Wang, Y.; Liang, T.; Wu, H.; Wang, C.; Sun, T.; Liu, S. Harnessing Nanomedicine for Modulating Microglial States in the Central Nervous System Disorders: Challenges and Opportunities. Biomedicine & Pharmacotherapy 2024, 177, 117011. [Google Scholar] [CrossRef]
- Hickman, S.E.; Kingery, N.D.; Ohsumi, T.K.; Borowsky, M.L.; Wang, L.; Means, T.K.; El Khoury, J. The Microglial Sensome Revealed by Direct RNA Sequencing. Nat Neurosci 2013, 16, 1896–1905. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Alterations in Immune Cells and Mediators in the Brain: It’s Not Always Neuroinflammation! Brain Pathology 2014, 24, 623–630. [Google Scholar] [CrossRef]
- Escoubas, C.C.; Molofsky, A.V. Microglia as Integrators of Brain-Associated Molecular Patterns. Trends in Immunology 2024, 45, 358–370. [Google Scholar] [CrossRef]
- Wang, H.; He, Y.; Sun, Z.; Ren, S.; Liu, M.; Wang, G.; Yang, J. Microglia in Depression: An Overview of Microglia in the Pathogenesis and Treatment of Depression. J Neuroinflammation 2022, 19, 132. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.-Q.; Ma, X.-T.; Hu, Z.-W.; Yang, S.; Chen, M.; Bosco, D.B.; Wu, L.-J.; Tian, D.-S. Dual Functions of Microglia in Ischemic Stroke. Neurosci. Bull. 2019, 35, 921–933. [Google Scholar] [CrossRef]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. The Semantics of Microglia Activation: Neuroinflammation, Homeostasis, and Stress. J Neuroinflammation 2021, 18, 258. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biological Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Li, S.; Wang, S.; Wu, X.; Liu, Y.; Yu, W.; Wang, Y.; Tang, Y.; Xia, M.; Li, B. Major Depressive Disorder: Hypothesis, Mechanism, Prevention and Treatment. Sig Transduct Target Ther 2024, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, M.; Magistretti, P.J. The Role of Astroglia in Neuroprotection. Dialogues in Clinical Neuroscience 2009, 11, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, M.; Magistretti, P.J. The Role of Astroglia in Neuroprotection. Dialogues in Clinical Neuroscience 2009, 11, 281–295. [Google Scholar] [CrossRef]
- Early, J.O.; Menon, D.; Wyse, C.A.; Cervantes-Silva, M.P.; Zaslona, Z.; Carroll, R.G.; Palsson-McDermott, E.M.; Angiari, S.; Ryan, D.G.; Corcoran, S.E.; et al. Circadian Clock Protein BMAL1 Regulates IL-1β in Macrophages via NRF2. Proc. Natl. Acad. Sci. U.S.A. 2018, 115. [Google Scholar] [CrossRef]
- Brown, S.J.; Huang, X.-F.; Newell, K.A. The Kynurenine Pathway in Major Depression: What We Know and Where to Next. Neuroscience & Biobehavioral Reviews 2021, 127, 917–927. [Google Scholar] [CrossRef]
- Maes, M.; Landucci Bonifacio, K.; Morelli, N.R.; Vargas, H.O.; Barbosa, D.S.; Carvalho, A.F.; Nunes, S.O.V. Major Differences in Neurooxidative and Neuronitrosative Stress Pathways Between Major Depressive Disorder and Types I and II Bipolar Disorder. Mol Neurobiol 2019, 56, 141–156. [Google Scholar] [CrossRef]
- Greaney, J.L.; Surachman, A.; Saunders, E.F.H.; Alexander, L.M.; Almeida, D.M. Greater Daily Psychosocial Stress Exposure Is Associated With Increased Norepinephrine-Induced Vasoconstriction in Young Adults. JAHA 2020, 9. [Google Scholar] [CrossRef]
- Wang, Y.; Ni, J.; Zhai, L.; Gao, C.; Xie, L.; Zhao, L.; Yin, X. Inhibition of Activated Astrocyte Ameliorates Lipopolysaccharide- Induced Depressive-like Behaviors. Journal of Affective Disorders 2019, 242, 52–59. [Google Scholar] [CrossRef]
- Kennis, M.; Gerritsen, L.; Van Dalen, M.; Williams, A.; Cuijpers, P.; Bockting, C. Prospective Biomarkers of Major Depressive Disorder: A Systematic Review and Meta-Analysis. Mol Psychiatry 2020, 25, 321–338. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Q.; Zheng, Z.; Sun, Y.; Niu, Y.; Li, D.; Wang, S.; Meng, W. BDNF Enhances Electrophysiological Activity and Excitatory Synaptic Transmission of RA Projection Neurons in Adult Male Zebra Finches. Brain Research 2023, 1801, 148208. [Google Scholar] [CrossRef]
- Cui, L.; Li, S.; Wang, S.; Wu, X.; Liu, Y.; Yu, W.; Wang, Y.; Tang, Y.; Xia, M.; Li, B. Major Depressive Disorder: Hypothesis, Mechanism, Prevention and Treatment. Sig Transduct Target Ther 2024, 9. [Google Scholar] [CrossRef] [PubMed]
- Alcocer-Gómez, E.; Cordero, M.D. NLRP3 Inflammasome: A New Target in Major Depressive Disorder. CNS Neurosci Ther 2014, 20, 294–295. [Google Scholar] [CrossRef]
- Malhi, G.S.; Mann, J.J. Depression. The Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-W.; Suzuki, K.; Kavalali, E.T.; Monteggia, L.M. Ketamine: Mechanisms and Relevance to Treatment of Depression. Annu. Rev. Med. 2024, 75, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, A.; Furukawa, T.A.; Salanti, G.; Chaimani, A.; Atkinson, L.Z.; Ogawa, Y.; Leucht, S.; Ruhe, H.G.; Turner, E.H.; Higgins, J.P.T.; et al. Comparative Efficacy and Acceptability of 21 Antidepressant Drugs for the Acute Treatment of Adults with Major Depressive Disorder: A Systematic Review and Network Meta-Analysis. The Lancet 2018, 391, 1357–1366. [Google Scholar] [CrossRef]
- Hiemke, C.; Bergemann, N.; Clement, H.; Conca, A.; Deckert, J.; Domschke, K.; Eckermann, G.; Egberts, K.; Gerlach, M.; Greiner, C.; et al. Consensus Guidelines for Therapeutic Drug Monitoring in Neuropsychopharmacology: Update 2017. Pharmacopsychiatry 2018, 51, 9–62. [Google Scholar] [CrossRef]
- Sousa, R.D.D.; Rodrigues, A.M.; Gregório, M.J.; Branco, J.D.C.; Gouveia, M.J.; Canhão, H.; Dias, S.S. Anxiety and Depression in the Portuguese Older Adults: Prevalence and Associated Factors. Front. Med. 2017, 4. [Google Scholar] [CrossRef]
- Zakaria, F.H.; Samhani, I.; Mustafa, M.Z.; Shafin, N. Pathophysiology of Depression: Stingless Bee Honey Promising as an Antidepressant. Molecules 2022, 27, 5091. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Guski, L.S.; Freund, N.; Gøtzsche, P.C. Suicidality and Aggression during Antidepressant Treatment: Systematic Review and Meta-Analyses Based on Clinical Study Reports. BMJ 2016, i65. [Google Scholar] [CrossRef] [PubMed]
- Hall, S.; Parr, B.-A.; Hussey, S.; Anoopkumar-Dukie, S.; Arora, D.; Grant, G.D. The Neurodegenerative Hypothesis of Depression and the Influence of Antidepressant Medications. European Journal of Pharmacology 2024, 983, 176967. [Google Scholar] [CrossRef] [PubMed]
- Richelson, E. Multi-Modality: A New Approach for the Treatment of Major Depressive Disorder. International Journal of Neuropsychopharmacology 2013, 16, 1433–1442. [Google Scholar] [CrossRef]
- Severe, J.; Greden, J.F.; Reddy, P. Consequences of Recurrence of Major Depressive Disorder: Is Stopping Effective Antidepressant Medications Ever Safe? FOC 2020, 18, 120–128. [Google Scholar] [CrossRef]
- Cuijpers, P.; Miguel, C.; Ciharova, M.; Ebert, D.; Harrer, M.; Karyotaki, E. Transdiagnostic Treatment of Depression and Anxiety: A Meta-Analysis. Psychol. Med. 2023, 53, 6535–6546. [Google Scholar] [CrossRef]
- Nikolova, V.L.; Cleare, A.J.; Young, A.H.; Stone, J.M. Acceptability, Tolerability, and Estimates of Putative Treatment Effects of Probiotics as Adjunctive Treatment in Patients With Depression: A Randomized Clinical Trial. JAMA Psychiatry 2023, 80, 842. [Google Scholar] [CrossRef]
- Ji, C.; Tang, Y.; Zhang, Y.; Li, C.; Liang, H.; Ding, L.; Xia, X.; Xiong, L.; Qi, X.-R.; Zheng, J.C. Microglial Glutaminase 1 Deficiency Mitigates Neuroinflammation Associated Depression. Brain, Behavior, and Immunity 2022, 99, 231–245. [Google Scholar] [CrossRef]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-Sensing Receptors in Sterile Inflammation and Inflammatory Diseases. Nat Rev Immunol 2020, 20, 95–112. [Google Scholar] [CrossRef]
- Suneson, K.; Lindahl, J.; Chamli Hårsmar, S.; Söderberg, G.; Lindqvist, D. Inflammatory Depression—Mechanisms and Non-Pharmacological Interventions. IJMS 2021, 22, 1640. [Google Scholar] [CrossRef]
- Stephan, A.H.; Barres, B.A.; Stevens, B. The Complement System: An Unexpected Role in Synaptic Pruning During Development and Disease. Annu. Rev. Neurosci. 2012, 35, 369–389. [Google Scholar] [CrossRef]
- Kiraly, D.D.; Horn, S.R.; Van Dam, N.T.; Costi, S.; Schwartz, J.; Kim-Schulze, S.; Patel, M.; Hodes, G.E.; Russo, S.J.; Merad, M.; et al. Altered Peripheral Immune Profiles in Treatment-Resistant Depression: Response to Ketamine and Prediction of Treatment Outcome. Transl Psychiatry 2017, 7, e1065–e1065. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.A.; Beurel, E.; Loewenstein, D.A.; Lowell, J.A.; Craighead, W.E.; Dunlop, B.W.; Mayberg, H.S.; Dhabhar, F.; Dietrich, W.D.; Keane, R.W.; et al. Defective Inflammatory Pathways in Never-Treated Depressed Patients Are Associated with Poor Treatment Response. Neuron 2018, 99, 914–924.e3. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Wan, X.; Zhuang, P.; Jia, W.; Ao, Y.; Liu, X.; Tian, Y.; Zhu, L.; Huang, Y.; Yao, J.; et al. High Fried Food Consumption Impacts Anxiety and Depression Due to Lipid Metabolism Disturbance and Neuroinflammation. Proc. Natl. Acad. Sci. U.S.A. 2023, 120. [Google Scholar] [CrossRef] [PubMed]
- Westfall, S.; Caracci, F.; Estill, M.; Frolinger, T.; Shen, L.; Pasinetti, G.M. Chronic Stress-Induced Depression and Anxiety Priming Modulated by Gut-Brain-Axis Immunity. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Hannestad, J.; DellaGioia, N.; Bloch, M. The Effect of Antidepressant Medication Treatment on Serum Levels of Inflammatory Cytokines: A Meta-Analysis. Neuropsychopharmacol 2011, 36, 2452–2459. [Google Scholar] [CrossRef]
- Warner-Schmidt, J.L.; Vanover, K.E.; Chen, E.Y.; Marshall, J.J.; Greengard, P. Antidepressant Effects of Selective Serotonin Reuptake Inhibitors (SSRIs) Are Attenuated by Antiinflammatory Drugs in Mice and Humans. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 9262–9267. [Google Scholar] [CrossRef]
- Piletz, J.E.; Halaris, A.; Iqbal, O.; Hoppensteadt, D.; Fareed, J.; Zhu, H.; Sinacore, J.; DeVane, C.L. Pro-Inflammatory Biomakers in Depression: Treatment with Venlafaxine. The World Journal of Biological Psychiatry 2009, 10, 313–323. [Google Scholar] [CrossRef]
- Guo, B.; Zhang, M.; Hao, W.; Wang, Y.; Zhang, T.; Liu, C. Neuroinflammation Mechanisms of Neuromodulation Therapies for Anxiety and Depression. Transl Psychiatry 2023, 13. [Google Scholar] [CrossRef]
- Zazula, R.; Husain, M.I.; Mohebbi, M.; Walker, A.J.; Chaudhry, I.B.; Khoso, A.B.; Ashton, M.M.; Agustini, B.; Husain, N.; Deakin, J.; et al. Minocycline as Adjunctive Treatment for Major Depressive Disorder: Pooled Data from Two Randomized Controlled Trials. Aust N Z J Psychiatry 2021, 55, 784–798. [Google Scholar] [CrossRef]
- Shi, J.; Tan, C.; Ge, X.; Qin, Z.; Xiong, H. Recent Advances in Stimuli-Responsive Controlled Release Systems for Neuromodulation. J. Mater. Chem. B 2024, 12, 5769–5786. [Google Scholar] [CrossRef] [PubMed]
- Kadry, H.; Noorani, B.; Cucullo, L. A Blood–Brain Barrier Overview on Structure, Function, Impairment, and Biomarkers of Integrity. Fluids Barriers CNS 2020, 17. [Google Scholar] [CrossRef] [PubMed]
- Lochhead, J.J.; Yang, J.; Ronaldson, P.T.; Davis, T.P. Structure, Function, and Regulation of the Blood-Brain Barrier Tight Junction in Central Nervous System Disorders. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Liebner, S.; Czupalla, C.J.; Wolburg, H. Current Concepts of Blood-Brain Barrier Development. Int. J. Dev. Biol. 2011, 55, 467–476. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, Y.; Fan, X. Microvascular Pericytes in Brain-Associated Vascular Disease. Biomedicine & Pharmacotherapy 2020, 121, 109633. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Ayyadurai, S.; Zlokovic, B.V. Pericytes of the Neurovascular Unit: Key Functions and Signaling Pathways. Nat Neurosci 2016, 19, 771–783. [Google Scholar] [CrossRef]
- Alsbrook, D.L.; Di Napoli, M.; Bhatia, K.; Biller, J.; Andalib, S.; Hinduja, A.; Rodrigues, R.; Rodriguez, M.; Sabbagh, S.Y.; Selim, M.; et al. Neuroinflammation in Acute Ischemic and Hemorrhagic Stroke. Curr Neurol Neurosci Rep 2023, 23, 407–431. [Google Scholar] [CrossRef]
- Varatharaj, A.; Galea, I. The Blood-Brain Barrier in Systemic Inflammation. Brain, Behavior, and Immunity 2017, 60, 1–12. [Google Scholar] [CrossRef]
- Lye, P.; Bloise, E.; Imperio, G.E.; Chitayat, D.; Matthews, S.G. Functional Expression of Multidrug-Resistance (MDR) Transporters in Developing Human Fetal Brain Endothelial Cells. Cells 2022, 11, 2259. [Google Scholar] [CrossRef]
- Loscher, W.; Potschka, H. Blood-Brain Barrier Active Efflux Transporters: ATP-Binding Cassette Gene Family. 2005, 2.
- Taskar, K.S.; Yang, X.; Neuhoff, S.; Patel, M.; Yoshida, K.; Paine, M.F.; Brouwer, K.L.R.; Chu, X.; Sugiyama, Y.; Cook, J.; et al. Clinical Relevance of Hepatic and Renal P-gp/BCRP Inhibition of Drugs: An International Transporter Consortium Perspective. Clin Pharma and Therapeutics 2022, 112, 573–592. [Google Scholar] [CrossRef]
- Radosavljevic, M.; Svob Strac, D.; Jancic, J.; Samardzic, J. The Role of Pharmacogenetics in Personalizing the Antidepressant and Anxiolytic Therapy. Genes 2023, 14, 1095. [Google Scholar] [CrossRef]
- O’Brien, F.E.; Dinan, T.G.; Griffin, B.T.; Cryan, J.F. Interactions between Antidepressants and P-glycoprotein at the Blood–Brain Barrier: Clinical Significance of in Vitro and in Vivo Findings. British J Pharmacology 2012, 165, 289–312. [Google Scholar] [CrossRef]
- Kuban, W.; Daniel, W.A. Cytochrome P450 Expression and Regulation in the Brain. Drug Metabolism Reviews 2021, 53, 1–29. [Google Scholar] [CrossRef]
- Manzoor, S.; Hoda, N. A Comprehensive Review of Monoamine Oxidase Inhibitors as Anti-Alzheimer’s Disease Agents: A Review. European Journal of Medicinal Chemistry 2020, 206, 112787. [Google Scholar] [CrossRef]
- Boado, R.J.; Tsukamoto, H.; Pardridge, W.M. Drug Delivery of Antisense Molecules to the Brain for Treatment of Alzheimer’s Disease and Cerebral AIDS. Journal of Pharmaceutical Sciences 1998, 87, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Rhea, E.M.; Reed, M.J.; Erickson, M.A. The Penetration of Therapeutics across the Blood-Brain Barrier: Classic Case Studies and Clinical Implications. Cell Reports Medicine 2024, 5, 101760. [Google Scholar] [CrossRef] [PubMed]
- Dichiara, M.; Amata, B.; Turnaturi, R.; Marrazzo, A.; Amata, E. Tuning Properties for Blood–Brain Barrier Permeation: A Statistics-Based Analysis. ACS Chem. Neurosci. 2020, 11, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Huddart, R.; Hicks, J.K.; Ramsey, L.B.; Strawn, J.R.; Smith, D.M.; Bobonis Babilonia, M.; Altman, R.B.; Klein, T.E. PharmGKB Summary: Sertraline Pathway, Pharmacokinetics. Pharmacogenetics and Genomics 2020, 30, 26–33. [Google Scholar] [CrossRef]
- Koepsell, H. Glucose Transporters in Brain in Health and Disease. Pflugers Arch - Eur J Physiol 2020, 472, 1299–1343. [Google Scholar] [CrossRef]
- Sun, Z.-W.; Wang, X.; Zhao, Y.; Sun, Z.-X.; Wu, Y.-H.; Hu, H.; Zhang, L.; Wang, S.-D.; Li, F.; Wei, A.-J.; et al. Blood-Brain Barrier Dysfunction Mediated by the EZH2-Claudin-5 Axis Drives Stress-Induced TNF-α Infiltration and Depression-like Behaviors. Brain, Behavior, and Immunity 2024, 115, 143–156. [Google Scholar] [CrossRef]
- Meixensberger, S.; Kuzior, H.; Fiebich, B.L.; Süß, P.; Runge, K.; Berger, B.; Nickel, K.; Denzel, D.; Schiele, M.A.; Michel, M.; et al. Upregulation of sICAM-1 and sVCAM-1 Levels in the Cerebrospinal Fluid of Patients with Schizophrenia Spectrum Disorders. Diagnostics 2021, 11, 1134. [Google Scholar] [CrossRef] [PubMed]
- Haqqani, A.S.; Bélanger, K.; Stanimirovic, D.B. Receptor-Mediated Transcytosis for Brain Delivery of Therapeutics: Receptor Classes and Criteria. Front. Drug Deliv. 2024, 4. [Google Scholar] [CrossRef]
- Ding, S.; Khan, A.I.; Cai, X.; Song, Y.; Lyu, Z.; Du, D.; Dutta, P.; Lin, Y. Overcoming Blood–Brain Barrier Transport: Advances in Nanoparticle-Based Drug Delivery Strategies. Materials Today 2020, 37, 112–125. [Google Scholar] [CrossRef]
- Mubarak, N.; Waqar, M.A.; Khan, A.M.; Asif, Z.; Alvi, A.S.; Virk, A.A.; Amir, S. A Comprehensive Insight of Innovations and Recent Advancements in Nanocarriers for Nose-to-Brain Drug Targeting. Designed Monomers and Polymers 2025, 28, 7–29. [Google Scholar] [CrossRef]
- Jiao, Y.; Yang, L.; Wang, R.; Song, G.; Fu, J.; Wang, J.; Gao, N.; Wang, H. Drug Delivery Across the Blood–Brain Barrier: A New Strategy for the Treatment of Neurological Diseases. Pharmaceutics 2024, 16, 1611. [Google Scholar] [CrossRef]
- Adepu, S.; Ramakrishna, S. Controlled Drug Delivery Systems: Current Status and Future Directions. Molecules 2021, 26, 5905. [Google Scholar] [CrossRef]
- Martín-Morales, C.; Caspani, S.; Desco, M.; Tavares De Sousa, C.; Gómez-Gaviro, M.V. Controlled Drug Release Systems for Cerebrovascular Diseases. Advanced Therapeutics 2025, 8. [Google Scholar] [CrossRef]
- Song, Y.H.; De, R.; Lee, K.T. Emerging Strategies to Fabricate Polymeric Nanocarriers for Enhanced Drug Delivery across Blood-Brain Barrier: An Overview. Advances in Colloid and Interface Science 2023, 320, 103008. [Google Scholar] [CrossRef]
- Tang, K.; Tang, Z.; Niu, M.; Kuang, Z.; Xue, W.; Wang, X.; Liu, X.; Yu, Y.; Jeong, S.; Ma, Y.; et al. Allosteric Targeted Drug Delivery for Enhanced Blood-Brain Barrier Penetration via Mimicking Transmembrane Domain Interactions. Nat Commun 2025, 16. [Google Scholar] [CrossRef]
- Gao, X.; Xu, J.; Yao, T.; Liu, X.; Zhang, H.; Zhan, C. Peptide-Decorated Nanocarriers Penetrating the Blood-Brain Barrier for Imaging and Therapy of Brain Diseases. Advanced Drug Delivery Reviews 2022, 187, 114362. [Google Scholar] [CrossRef]
- Teixeira, M.I.; Lopes, C.M.; Amaral, M.H.; Costa, P.C. Surface-Modified Lipid Nanocarriers for Crossing the Blood-Brain Barrier (BBB): A Current Overview of Active Targeting in Brain Diseases. Colloids and Surfaces B: Biointerfaces 2023, 221, 112999. [Google Scholar] [CrossRef]
- Rafati, N.; Zarepour, A.; Bigham, A.; Khosravi, A.; Naderi-Manesh, H.; Iravani, S.; Zarrabi, A. Nanosystems for Targeted Drug Delivery: Innovations and Challenges in Overcoming the Blood-Brain Barrier for Neurodegenerative Disease and Cancer Therapy. International Journal of Pharmaceutics 2024, 666, 124800. [Google Scholar] [CrossRef] [PubMed]
- Toader, C.; Dumitru, A.V.; Eva, L.; Serban, M.; Covache-Busuioc, R.-A.; Ciurea, A.V. Nanoparticle Strategies for Treating CNS Disorders: A Comprehensive Review of Drug Delivery and Theranostic Applications. IJMS 2024, 25, 13302. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zeng, H.; Luo, Y.; Chen, Y.; Wang, M.; Wu, C.; Hu, P. Recent Applications of PLGA in Drug Delivery Systems. Polymers 2024, 16, 2606. [Google Scholar] [CrossRef] [PubMed]
- Gabold, B.; Adams, F.; Brameyer, S.; Jung, K.; Ried, C.L.; Merdan, T.; Merkel, O.M. Transferrin-Modified Chitosan Nanoparticles for Targeted Nose-to-Brain Delivery of Proteins. Drug Deliv. and Transl. Res. 2023, 13, 822–838. [Google Scholar] [CrossRef]
- Malhotra, M.; Tomaro-Duchesneau, C.; Saha, S.; Prakash, S. Intranasal, siRNA Delivery to the Brain by TAT/MGF Tagged PEGylated Chitosan Nanoparticles. Journal of Pharmaceutics 2013, 2013, 1–10. [Google Scholar] [CrossRef]
- Graván, P.; Aguilera-Garrido, A.; Marchal, J.A.; Navarro-Marchal, S.A.; Galisteo-González, F. Lipid-Core Nanoparticles: Classification, Preparation Methods, Routes of Administration and Recent Advances in Cancer Treatment. Advances in Colloid and Interface Science 2023, 314, 102871. [Google Scholar] [CrossRef]
- Eleraky, N.; M. Omar, M.; A. Mahmoud, H.; A. Abou-Taleb, H. Nanostructured Lipid Carriers to Mediate Brain Delivery of Temazepam: Design and In Vivo Study. Pharmaceutics 2020, 12, 451. [Google Scholar] [CrossRef]
- Wang, H.; He, Y.; Sun, Z.; Ren, S.; Liu, M.; Wang, G.; Yang, J. Microglia in Depression: An Overview of Microglia in the Pathogenesis and Treatment of Depression. J Neuroinflammation 2022, 19. [Google Scholar] [CrossRef]
- Florendo, M.; Figacz, A.; Srinageshwar, B.; Sharma, A.; Swanson, D.; Dunbar, G.L.; Rossignol, J. Use of Polyamidoamine Dendrimers in Brain Diseases. Molecules 2018, 23, 2238. [Google Scholar] [CrossRef]
- Gaitsch, H.; Hersh, A.M.; Alomari, S.; Tyler, B.M. Dendrimer Technology in Glioma: Functional Design and Potential Applications. Cancers 2023, 15, 1075. [Google Scholar] [CrossRef]
- Dadkhah, M.; Afshari, S.; Samizadegan, T.; Shirmard, L.R.; Barin, S. Pegylated Chitosan Nanoparticles of Fluoxetine Enhance Cognitive Performance and Hippocampal Brain Derived Neurotrophic Factor Levels in a Rat Model of Local Demyelination. Experimental Gerontology 2024, 195, 112533. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Van Der Putten, L.; Queiroz, J.F.; Pinheiro, M.; Reis, S. Transferrin-Functionalized Lipid Nanoparticles for Curcumin Brain Delivery. Journal of Biotechnology 2021, 331, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Eldeeb, G.M.; Yousef, M.I.; Helmy, Y.M.; Aboudeya, H.M.; Mahmoud, S.A.; Kamel, M.A. The Protective Effects of Chitosan and Curcumin Nanoparticles against the Hydroxyapatite Nanoparticles-Induced Neurotoxicity in Rats. Sci Rep 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Zhao, C.; Wu, X.; Chen, Y.; Zheng, C.; Zhang, X.; Xu, Y.; Zhu, L.; Wang, H.; Xie, G.; et al. Facile Engineered Macrophages-Derived Exosomes-Functionalized PLGA Nanocarrier for Targeted Delivery of Dual Drug Formulation against Neuroinflammation by Modulation of Microglial Polarization in a Post-Stroke Depression Rat Model. Biomedicine & Pharmacotherapy 2024, 179, 117263. [Google Scholar] [CrossRef]
- Yusuf, M.; Khan, M.; Khan, R.A.; Maghrabi, I.A.; Ahmed, B. Polysorbate-80-Coated, Polymeric Curcumin Nanoparticles for in Vivo Anti-Depressant Activity across BBB and Envisaged Biomolecular Mechanism of Action through a Proposed Pharmacophore Model. Journal of Microencapsulation 2016, 33, 646–655. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, Y.; Jiang, Z.; He, C.; Wang, B.; Wang, Q.; Wang, Z.; Wu, T.; Chen, X.; Deng, Z.; et al. Bioinspired Polydopamine Nanoparticles as Efficient Antioxidative and Anti-Inflammatory Enhancers against UV-Induced Skin Damage. J Nanobiotechnol 2023, 21. [Google Scholar] [CrossRef]
- Jiang, C.; Yang, X.; Huang, Q.; Lei, T.; Luo, H.; Wu, D.; Yang, Z.; Xu, Y.; Dou, Y.; Ma, X.; et al. Microglial-Biomimetic Memantine-Loaded Polydopamine Nanomedicines for Alleviating Depression. Advanced Materials 2025, 37. [Google Scholar] [CrossRef]
- Fidelis, E.M.; Savall, A.S.P.; Da Luz Abreu, E.; Carvalho, F.; Teixeira, F.E.G.; Haas, S.E.; Bazanella Sampaio, T.; Pinton, S. Curcumin-Loaded Nanocapsules Reverses the Depressant-Like Behavior and Oxidative Stress Induced by β-Amyloid in Mice. Neuroscience 2019, 423, 122–130. [Google Scholar] [CrossRef]
- He, X.; Yang, L.; Wang, M.; Zhuang, X.; Huang, R.; Zhu, R.; Wang, S. Targeting the Endocannabinoid/CB1 Receptor System For Treating Major Depression Through Antidepressant Activities of Curcumin and Dexanabinol-Loaded Solid Lipid Nanoparticles. Cell Physiol Biochem 2017, 42, 2281–2294. [Google Scholar] [CrossRef]
- He, X.; Zhu, Y.; Wang, M.; Jing, G.; Zhu, R.; Wang, S. Antidepressant Effects of Curcumin and HU-211 Coencapsulated Solid Lipid Nanoparticles against Corticosterone-Induced Cellular and Animal Models of Major Depression. IJN 2016, Volume 11, 4975–4990. [Google Scholar] [CrossRef]
- Khadrawy, Y.A.; Hosny, E.N.; Magdy, M.; Mohammed, H.S. Antidepressant Effects of Curcumin-Coated Iron Oxide Nanoparticles in a Rat Model of Depression. European Journal of Pharmacology 2021, 908, 174384. [Google Scholar] [CrossRef]
- Fahmy, H.M.; Aboalasaad, F.A.; Mohamed, A.S.; Elhusseiny, F.A.; Khadrawy, Y.A.; Elmekawy, A. Evaluation of the Therapeutic Effect of Curcumin-Conjugated Zinc Oxide Nanoparticles on Reserpine-Induced Depression in Wistar Rats. Biol Trace Elem Res 2024, 202, 2630–2644. [Google Scholar] [CrossRef] [PubMed]
- Rubab, S.; Naeem, K.; Rana, I.; Khan, N.; Afridi, M.; Ullah, I.; Shah, F.A.; Sarwar, S.; Din, F.U.; Choi, H.-I.; et al. Enhanced Neuroprotective and Antidepressant Activity of Curcumin-Loaded Nanostructured Lipid Carriers in Lipopolysaccharide-Induced Depression and Anxiety Rat Model. International Journal of Pharmaceutics 2021, 603, 120670. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A.; Rana, I.; Choi, H.-I.; Lee, C.-H.; Baek, S.-W.; Lim, C.-W.; Khan, N.; Arif, S.T.; Sahar, N.U.; Alvi, A.M.; et al. Potential and Applications of Nanocarriers for Efficient Delivery of Biopharmaceuticals. Pharmaceutics 2020, 12, 1184. [Google Scholar] [CrossRef] [PubMed]
- Gul, M.; Shah, F.A.; Sahar, N.; Malik, I.; Din, F.U.; Khan, S.A.; Aman, W.; Choi, H.-I.; Lim, C.-W.; Noh, H.-Y.; et al. Formulation Optimization, in Vitro and in Vivo Evaluation of Agomelatine-Loaded Nanostructured Lipid Carriers for Augmented Antidepressant Effects. Colloids and Surfaces B: Biointerfaces 2022, 216, 112537. [Google Scholar] [CrossRef]
- Zhou, J.-F.; Duan, L.; Wang, Y.-X.; Wang, C.-L.; Tian, M.-L.; Qi, X.-J.; Qiu, F. Design, Characterization of Resveratrol-Thermosensitive Hydrogel System (Res-THS) and Evaluation of Its Anti-Depressant Effect via Intranasal Administration. Materials & Design 2022, 216, 110597. [Google Scholar] [CrossRef]
- Qi, X.-J.; Liu, X.-Y.; Tang, L.-M.-Y.; Li, P.-F.; Qiu, F.; Yang, A.-H. Anti-Depressant Effect of Curcumin-Loaded Guanidine-Chitosan Thermo-Sensitive Hydrogel by Nasal Delivery. Pharmaceutical Development and Technology 2020, 25, 316–325. [Google Scholar] [CrossRef]
- Gao, M.; Hu, P.; Cai, Z.; Wu, Y.; Wang, D.; Hu, W.; Xu, X.; Zhang, Y.; Lu, X.; Chen, D.; et al. Identification of a Microglial Activation-Dependent Antidepressant Effect of Amphotericin B Liposome. Neuropharmacology 2019, 151, 33–44. [Google Scholar] [CrossRef]
- Sheng, L.; Wei, Y.; Pi, C.; Cheng, J.; Su, Z.; Wang, Y.; Chen, T.; Wen, J.; Wei, Y.; Ma, J.; et al. Preparation and Evaluation of Curcumin Derivatives Nanoemulsion Based on Turmeric Extract and Its Antidepressant Effect. IJN 2023, Volume 18, 7965–7983. [Google Scholar] [CrossRef]
- Suchiwa, P.-O.; Nudmamud-Thanoi, S.; Phoungpetchara, I.; Tiyaboonchai, W. Comparison of Curcumin-Based Nano-Formulations for Antidepressant Effects in an Animal Model. Journal of Drug Delivery Science and Technology 2023, 88, 104901. [Google Scholar] [CrossRef]
- Zhang, F.; Nance, E.; Alnasser, Y.; Kannan, R.; Kannan, S. Microglial Migration and Interactions with Dendrimer Nanoparticles Are Altered in the Presence of Neuroinflammation. J Neuroinflammation 2016, 13. [Google Scholar] [CrossRef]
- Ahmad, S.; Ahmad, S.; Ali, S.; Esa, M.; Khan, A.; Yan, H. Recent Advancements and Unexplored Biomedical Applications of Green Synthesized Ag and Au Nanoparticles: A Review. IJN 2024, Volume 19, 3187–3215. [Google Scholar] [CrossRef]
- Leite, J.M.D.S.; Santana, L.M.D.; Nadvorny, D.; Abreu, B.O.D.; Rebouças, J.D.S.; Formiga, F.R.; Soares, M.F.D.L.R.; Soares-Sobrinho, J.L. Nanoparticle Design for Hydrophilic Drugs: Isoniazid Biopolymeric Nanostructure. Journal of Drug Delivery Science and Technology 2023, 87, 104754. [Google Scholar] [CrossRef]
- Leite, J.M.D.S.; Oliveira, A.C.D.J.; Dourado, D.; Santana, L.M.D.; Medeiros, T.S.; Nadvorny, D.; Silva, M.L.R.; Rolim-Neto, P.J.; Moreira, D.R.M.; Formiga, F.R.; et al. Rifampicin-Loaded Phthalated Cashew Gum Nano-Embedded Microparticles Intended for Pulmonary Administration. International Journal of Biological Macromolecules 2025, 303, 140693. [Google Scholar] [CrossRef]
- Da Silva Leite, J.M.; Patriota, Y.B.G.; De La Roca, M.F.; Soares-Sobrinho, J.L. New Perspectives in Drug Delivery Systems for the Treatment ofTuberculosis. CMC 2022, 29, 1936–1958. [Google Scholar] [CrossRef]
- Hnamte, M.; Pulikkal, A.K. Biocompatible Polymeric Nanoparticles as Carriers for Anticancer Phytochemicals. European Polymer Journal 2024, 202, 112637. [Google Scholar] [CrossRef]
- Zheng, Y.; He, R.; Wang, P.; Shi, Y.; Zhao, L.; Liang, J. Exosomes from LPS-Stimulated Macrophages Induce Neuroprotection and Functional Improvement after Ischemic Stroke by Modulating Microglial Polarization. Biomater. Sci. 2019, 7, 2037–2049. [Google Scholar] [CrossRef]
- Nagpal, K.; Singh, S.K.; Mishra, D. Evaluation of Safety and Efficacy of Brain Targeted Chitosan Nanoparticles of Minocycline. International Journal of Biological Macromolecules 2013, 59, 20–28. [Google Scholar] [CrossRef]
- Zhu, R.-R.; Cheng, L.-M.; He, X.-L.; Yang, L.; Wang, Z.-J.; Huang, R.-Q. Solid Lipid Nanoparticles Loading with Curcumin and Dexanabinol to Treat Major Depressive Disorder. Neural Regen Res 2021, 16, 537. [Google Scholar] [CrossRef]
- Choudhary, A.J.; Mahajan, S.S.; Majumdar, A.S. Nose to Brain Delivery of Flurbiprofen from a Solid Lipid Nanoparticles-Based Thermosensitive in-Situ Gel. Neuroscience Applied 2024, 3, 104062. [Google Scholar] [CrossRef]
- Zarezadeh, A.; Alizadeh, P.; Yourdkhani, A.; Motesadi Zarandi, F. Synthesis of Copper Ferrite-Decorated Bioglass Nanoparticles: Investigation of Magnetic Properties, Biocompatibility, and Drug Release Behaviour. Applied Surface Science 2025, 708, 163713. [Google Scholar] [CrossRef]
- Omolo, C.A.; Ismail, E.A.; Amuhaya, E.K.; Fasiku, V.; Nwabuife, J.C.; Mohamed, M.; Gafar, M.A.; Kalhapure, R.; Govender, T. Revolutionizing Drug Delivery: The Promise of Surface Functionalized Dendrimers in Overcoming Conventional Limitations. European Polymer Journal 2025, 235, 114079. [Google Scholar] [CrossRef]
- Yu, Y.; Yuan, M.; Qin, W.; Bai, H.; Liu, H.; Che, J. A Novel Strategy for Liposomal Drug Separation in Plasma by TiO2 Microspheres and Application in Pharmacokinetics. IJN 2023, Volume 18, 1321–1334. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of microglial activation and functional polarization into M1 (pro-inflammatory) and M2 (anti-inflammatory) phenotypes. Resting microglia are activated by danger signals such as PAMPs and DAMPs. Classical activation induces the M1 phenotype, associated with pro-inflammatory cytokine release and neurotoxic functions. Alternative activation promotes the M2 phenotype, characterized by anti-inflammatory and neuroprotective functions. The plasticity between M1 and M2 states highlights the complexity of the immune response in the central nervous system. Created by Biorrender (2025).
Figure 1.
Schematic representation of microglial activation and functional polarization into M1 (pro-inflammatory) and M2 (anti-inflammatory) phenotypes. Resting microglia are activated by danger signals such as PAMPs and DAMPs. Classical activation induces the M1 phenotype, associated with pro-inflammatory cytokine release and neurotoxic functions. Alternative activation promotes the M2 phenotype, characterized by anti-inflammatory and neuroprotective functions. The plasticity between M1 and M2 states highlights the complexity of the immune response in the central nervous system. Created by Biorrender (2025).
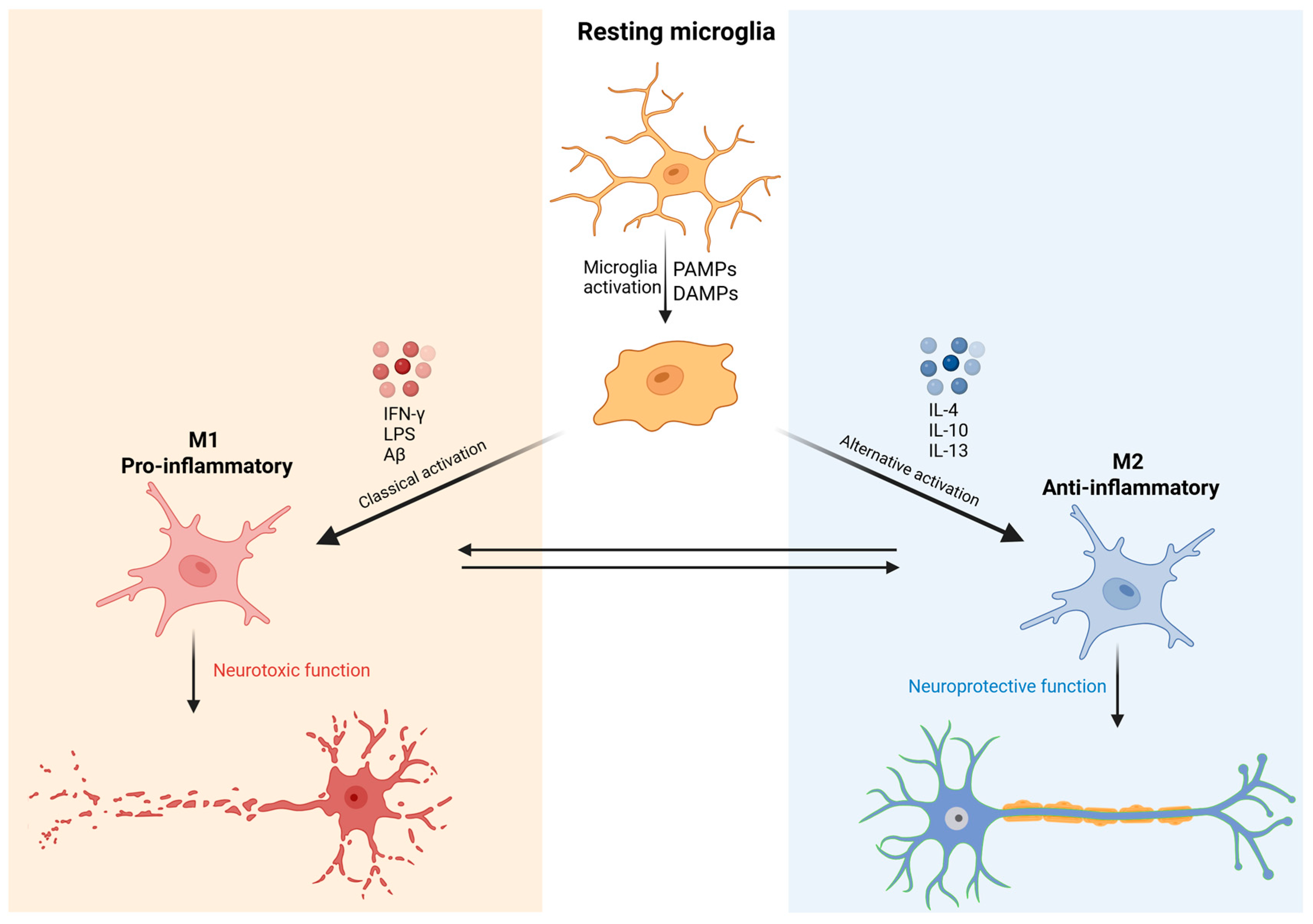
Figure 2.
Schematic representation of microglial activation and its association with neuroinflammation and major depressive disorder (MDD). Under physiological conditions, microglia remain in a quiescent state, constantly surveying the neural environment. When cerebral homeostasis is disrupted, danger signals like pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) activate specialized receptors, particularly Toll-like receptors (TLRs), leading to microglial phenotypic transformation. This activation contributes to the recruitment and polarization of M0 macrophages into pro-inflammatory M1 macrophages, which release inflammatory cytokines (TNF-α, IFN-γ, IL-1β, IL-6, IL-18) and reactive oxygen species (ROS), perpetuating a neuroinflammatory state and depressive symptoms. This pro-inflammatory environment contributes to the activation of the NLRP3 inflammasome and a reduction in neuroprotective factors such as BDNF and glutathione. The resulting chronic neuroinflammation is implicated in the pathophysiology of MDD, as illustrated by the association between brain inflammation and depressive symptoms. Created by Biorrender (2025).
Figure 2.
Schematic representation of microglial activation and its association with neuroinflammation and major depressive disorder (MDD). Under physiological conditions, microglia remain in a quiescent state, constantly surveying the neural environment. When cerebral homeostasis is disrupted, danger signals like pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) activate specialized receptors, particularly Toll-like receptors (TLRs), leading to microglial phenotypic transformation. This activation contributes to the recruitment and polarization of M0 macrophages into pro-inflammatory M1 macrophages, which release inflammatory cytokines (TNF-α, IFN-γ, IL-1β, IL-6, IL-18) and reactive oxygen species (ROS), perpetuating a neuroinflammatory state and depressive symptoms. This pro-inflammatory environment contributes to the activation of the NLRP3 inflammasome and a reduction in neuroprotective factors such as BDNF and glutathione. The resulting chronic neuroinflammation is implicated in the pathophysiology of MDD, as illustrated by the association between brain inflammation and depressive symptoms. Created by Biorrender (2025).
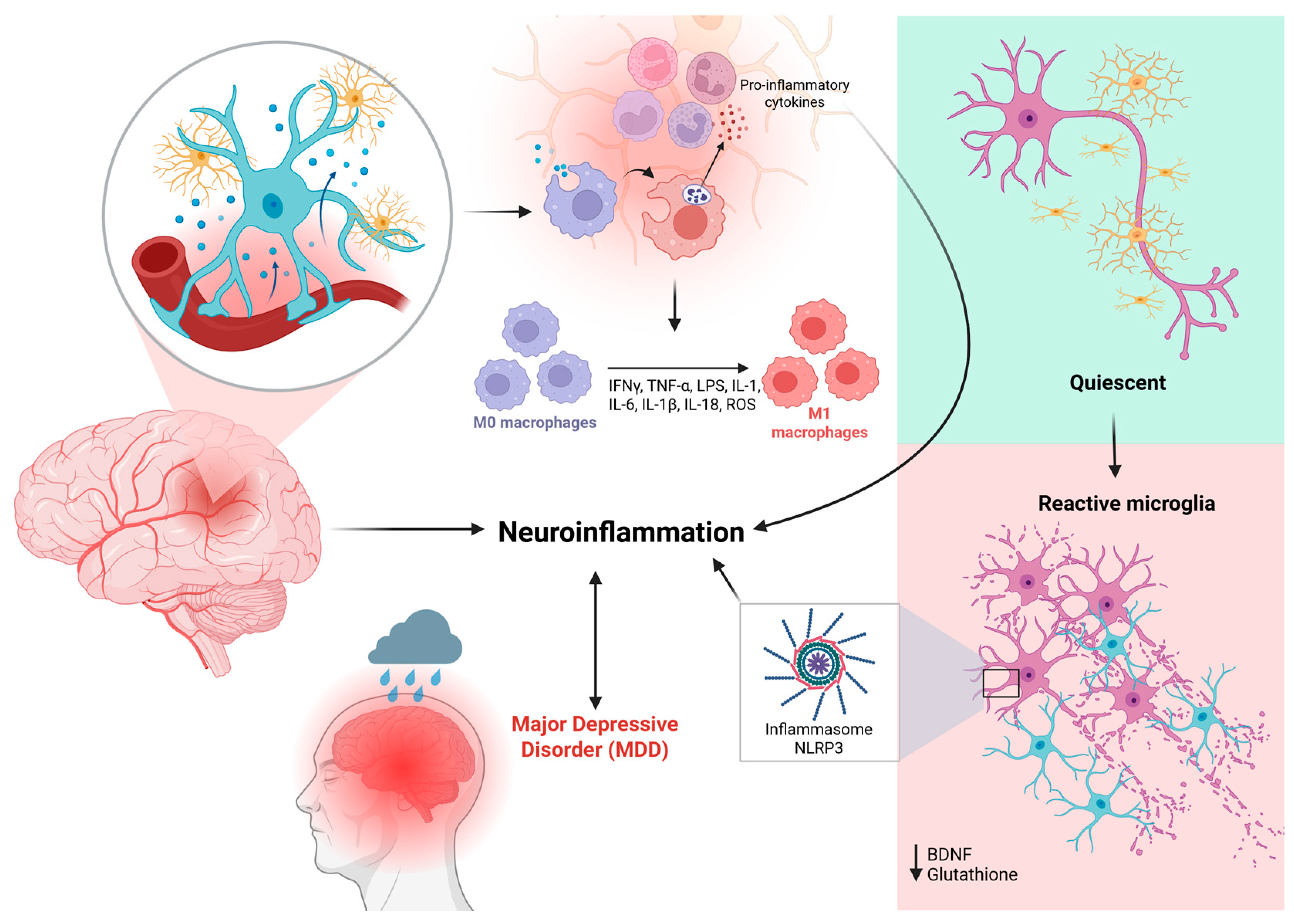
Figure 3.
Blood-brain barrier and drug delivery strategies to the CNS. (A) Anatomical illustration of the brain and a schematic view of the blood-brain barrier (BBB), highlighting its selective function that restricts drug entry into the brain parenchyma. (B) Left panel: Functional BBB structure showing tight junctions composed of proteins such as occludin, claudin-5, and ZO-1, along with efflux transporters (e.g., P-gp) and selective channels. Right panel: BBB dysfunction under inflammatory conditions, leading to junctional disruption, increased paracellular permeability, immune cell infiltration, and compromised barrier selectivity. (C) Drug delivery strategies using nanocarriers. Created by Biorrender (2025).
Figure 3.
Blood-brain barrier and drug delivery strategies to the CNS. (A) Anatomical illustration of the brain and a schematic view of the blood-brain barrier (BBB), highlighting its selective function that restricts drug entry into the brain parenchyma. (B) Left panel: Functional BBB structure showing tight junctions composed of proteins such as occludin, claudin-5, and ZO-1, along with efflux transporters (e.g., P-gp) and selective channels. Right panel: BBB dysfunction under inflammatory conditions, leading to junctional disruption, increased paracellular permeability, immune cell infiltration, and compromised barrier selectivity. (C) Drug delivery strategies using nanocarriers. Created by Biorrender (2025).
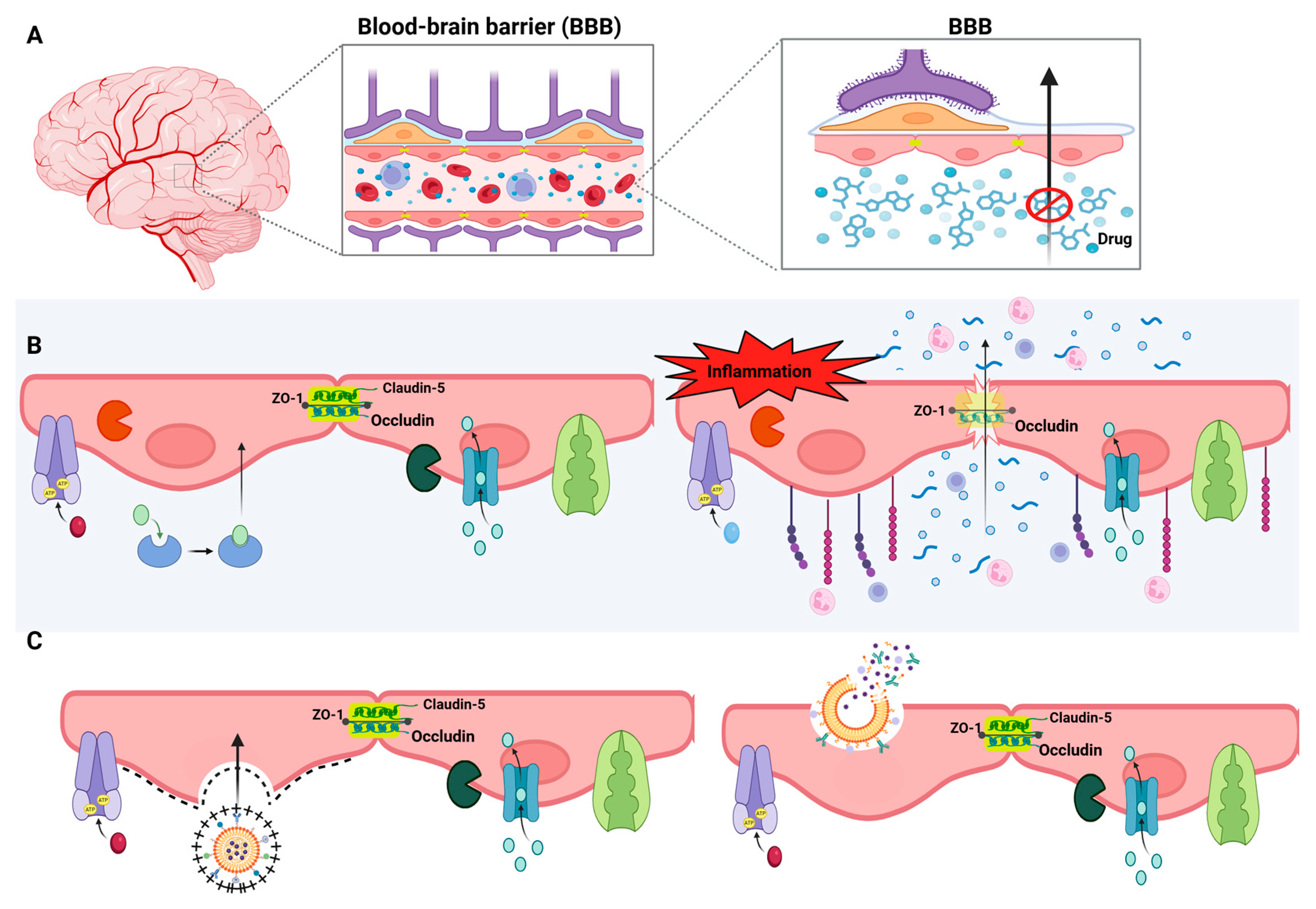
Figure 5.
Structure and drug loading approaches in polymeric nanoparticles. Schematic representation of the main types of polymeric nanoparticles used in controlled drug delivery systems: nanocapsules and nanospheres. Nanocapsules consist of an oily or aqueous core surrounded by a polymeric shell, allowing the drug to be either encapsulated within the core or adsorbed onto the particle surface. Nanospheres are solid matrix systems in which the drug can be homogeneously dispersed throughout the polymeric matrix or adsorbed externally. Created by Biorrender (2025).
Figure 5.
Structure and drug loading approaches in polymeric nanoparticles. Schematic representation of the main types of polymeric nanoparticles used in controlled drug delivery systems: nanocapsules and nanospheres. Nanocapsules consist of an oily or aqueous core surrounded by a polymeric shell, allowing the drug to be either encapsulated within the core or adsorbed onto the particle surface. Nanospheres are solid matrix systems in which the drug can be homogeneously dispersed throughout the polymeric matrix or adsorbed externally. Created by Biorrender (2025).
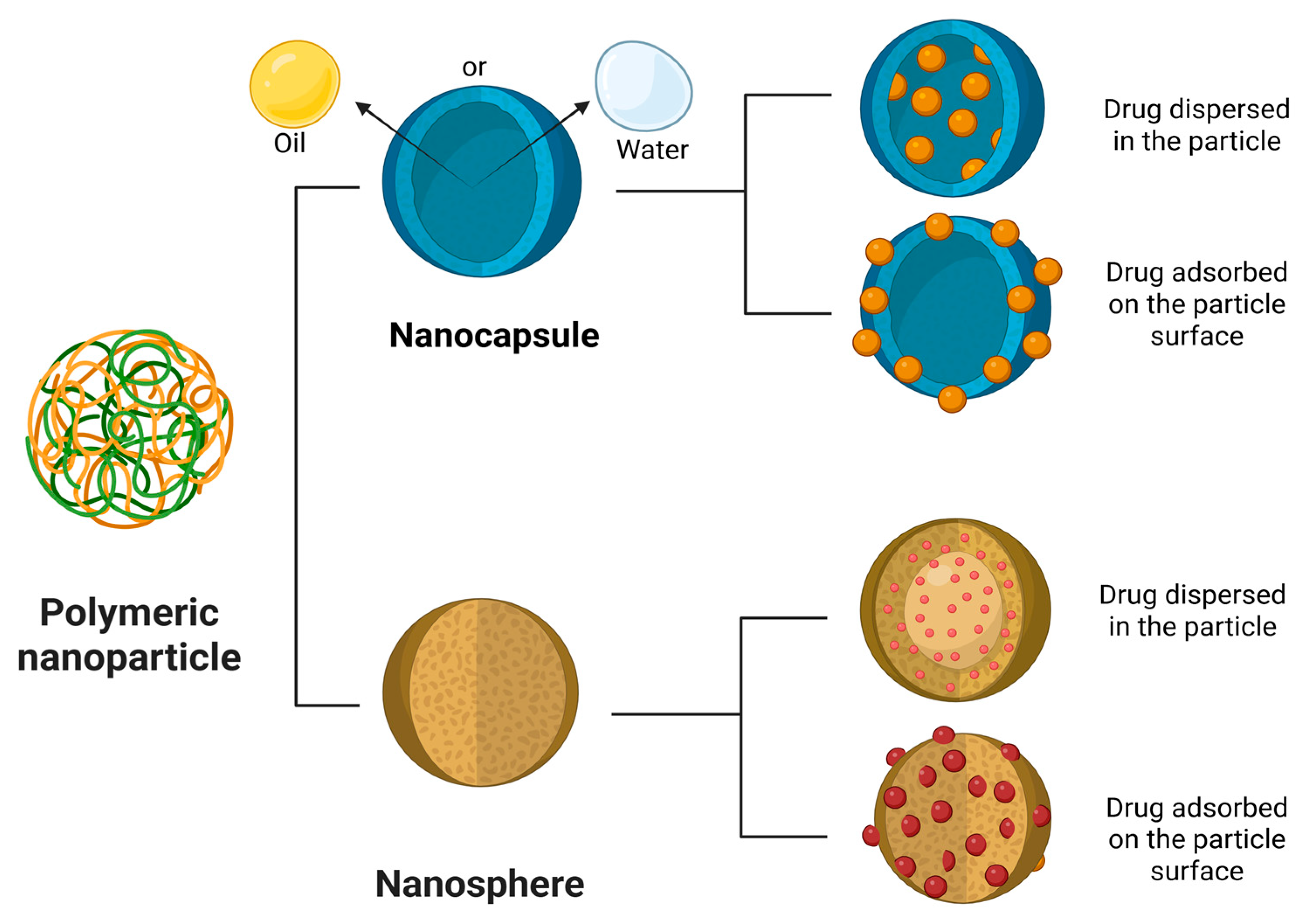

Table 1.
Nanosystems used for modulation of microglia in the treatment of MDD.
| Drug delivery systems | Compound used | Physico-chemical properties | In vitro release | In vitro activity | In vivo activity | Reference |
|---|---|---|---|---|---|---|
| Polymeric nanoparticle | Celastrol and minocycline | Size: 132 nm PZ: 35,5 mV |
- | - | Decrease in iNOS and CD86; increase in Arg-1 and CD206. Enhanced antidepressant activities and functional capacity. Alleviated brain inflammation in rats. | Lv et al (2024) [106] |
| Polymeric nanoparticle | Curcumin | Size: 194,3±14,8 nm PZ: 19,5±2,6 mV PDI: 0,359±0,014 EE: 63,9±7,2% |
83,78 ± 7,43 a 95,56 ± 4,67% em 144h | - | CUR-NP: decreased immobility (FST/TST), effective at 5 mg/kg, increased bioavailability (1.6x), increased SOD/catalase activity | Yusuf (2016) [107] |
| Polymeric nanoparticle | Dopamine hydrochloride | Size: 244 nm PDI: 0,119 PZ:-48 mV |
- | Polydopamine nanoparticles (NPs) eliminated >90% of DPPH radicals at 20 μg/mL (no significant difference compared to ascorbic acid) and significantly reduced intracellular ROS levels in LPS-treated PC12 cells | - | Zhang (2023) [108] |
| Polymeric nanoparticle | Memantine | Size: 163.5 ± 1.5 nm; PZ: -54.3 ± 2.2 mV |
Safety: No significant cytotoxicity (over 80% viability) in human neuroblastoma and mouse brain cells at high concentrations. Anti-inflammatory Action: Significantly reduced intracellular ROS and polarized microglia from M1 (pro-inflammatory) to M2 (anti-inflammatory) phenotype, decreasing CD86, TNF-α, IL-2, and increasing CD206, TGF-β, IL-10. |
Biodistribution and Efficacy: Showed higher accumulation in the brain and hippocampus (1.3x > PDA), confirming improved targeting. Outperformed Memantine monotherapy with lower, less frequent doses and greater efficacy. Safety: Demonstrated minimal observed toxicity. Inflammatory Modulation: Reduced hippocampal ROS and pro-inflammatory cytokines (IL-1β, IL-2); increased anti-inflammatory cytokines (IL-10). Neuroplasticity and Neuroprotection: Stimulated neurogenesis, exhibited neuroprotective effects, and restored synaptic plasticity (leading to decreased neuroinflammation and improved neuronal function). |
Jiang et al., 2025 [109] | |
| Polymeric nanoparticle | Celastrol and minocycline | 132 nm; PZ: -35.5 mV |
- | Microglial Modulation: CMC-EXPL significantly suppressed M1 (pro-inflammatory) microglial polarization (reducing CD80 and iNOS) and promoted M2 (anti-inflammatory) polarization (increasing CD206 and Arg1) in LPS-stimulated BV-2 cells. Therapeutic Potential: Showed superior anti-inflammatory therapeutic potential compared to other nanoformulations tested. |
Antidepressant Behavior: The nanoformulation controlled weight loss and reversed induced depressive behaviors in POSD rats. Microglial Modulation: Significantly reduced M1 markers (iNOS and CD86) and increased M2 markers (Arg-1 and CD206), confirming the M1 to M2 microglial polarization in the brains of POSD rats, contributing to its antidepressant efficacy. |
Lv et al., 2024 [106] |
| Solid lipid nanoparticle | Curcumin | 291 a 312 nm PZ: 22-36 mV |
- | - | NLC C reversed the effects of Aβ₂₅₋₃₅ (↑663.3% immobility in TST/FST), normalized SOD/CAT levels, and did not alter OFT performance | Fidelis et al., (2019) [110] |
| Solid lipid nanoparticle | Curcumin and dexanabinol | PZ: -22,6 ± 0,9 mV; EE: 19,12% ± 1,43% e 0,81% ± 0,04% |
Increase in DA/5-HT; reduction in cellular apoptosis | Increase in DA/5-HT levels and mRNA expression of CB1, p-MEK1, and p-ERK1/2 in the hippocampus and striatum | He et al 2017 [111] | |
| Solid lipid nanoparticles | HU-211 and curcumin | PZ: -21,7 ± 0,4 mV | 77% in 7 days | Increased expression of CB1, p-MEK1, and p-ERK1/2; cellular uptake: 99% | Increased dopamine levels | He et al 2016 [112] |
| Magnetic nanoparticles | Curcumin | 15 ± 3 nm PZ: -25 mV |
- | - | Increase in Na⁺, K⁺-ATPase activity and elevated levels of monoamine neurotransmitters. Prevention of excitotoxicity mediated by NMDA receptor overactivation. | Khadrawy et al., 2021 [113] |
| Magnetic nanoparticles | Curcumin | 342 ± 22,3 PDI: 1 PZ: − 25,6 ± 4,61 mV |
- | - | Decrease in malondialdehyde (MDA) levels, increase in reduced glutathione (GSH) and catalase (CAT) levels, and elevated concentrations of serotonin (5-HT) and norepinephrine (NE) | Fahmy et al. (2023) [114] |
| Nanostructured lipid carriers | Curcumin | 147,8 ± 10,4 nm; PDI: 0,27 ± 0,02; PZ: − 32,8 ± 1,4 mV; EE: 91,0 ± 4,6% | ~54% and ~73% at 12 h and 24 h, respectively | - | Suppression of p-NF-κB, TNF-α, and COX-2 expression | Rubab et al. (2021) [115] |
| Nanostructured lipid carriers | Curcumin | 147.8 ± 10.4 nm; PZ: -32.8 ± 1.4 mV |
- | - | Indirect Microglial Neuroprotection: CUR-NLCs markedly attenuated LPS-induced neurodegenerative damage and restored tissue architecture and cellular integrity. Anti-inflammatory Action: The nanoparticles suppressed the expression of pro-inflammatory markers (p-NF-κB, TNF-α, and COX-2) induced by LPS in the brain, reducing neuroinflammation. |
Zeb et al., 2020 [116] |
| Nanostructured lipid carriers | Agomelatine | 99.8 ± 2.6 nm; PZ: 23.2 ± 1.2 mV |
- | - | Enhanced Anti-inflammatory Effect: AGM-NLCs effectively suppressed LPS-induced neuroinflammation, reducing both the expression and concentrations of TNF-α and COX-2 in the brain. Indirect Microglial Action: Encapsulated agomelatine exerts an indirect effect on microglia by reducing inflammation, contributing to neuronal defense and integrity maintenance. |
Gul et al., 2022 [117] |
| Hydrogel | Resveratrol |
Porous structure characterized by dense and irregularly sized voids |
Res ~35% and Res-THS ~59% in 10 hours | - | Immobility time in the FST (~180 s → ~95 s); Corticosterone (CORT) levels (~160 → ~100 ng/mL); 5-HT, DA, and NA levels in the brain. |
Zhou et al. (2022) [118] |
| Hydrogel | Curcumin | Viscosity at 15–25 °C. | Thermosensitive GCS hydrogel containing 12.75% curcumin and curcumin/HP-β-CD complex achieved 55.54% release in 10 hours. | - | Significant reduction in immobility time in the forced swim test (FST) and tail suspension test (TST); reversal of ptosis and hypothermia (except for hydrogel at 58.4 μg/kg); increased levels of 5-HT, NE, and DA in the brain; effect comparable to fluoxetine | Qi et al., 2020 [119] |
| Liposome | Amphotericin B | - | - | - | Antidepressant Efficacy: In mice with Chronic Unpredictable Stress (CUS)-induced depression, AmB liposomal (at 1 and 3 mg/kg doses) markedly reversed depressive behaviors in a dose-dependent manner. This effect was not observed with isolated Amphotericin B. Microglial Mechanism of Action: Microglial activation is essential for the antidepressant effect of AmB liposomal. Pre-treatment with minocycline (a microglial inhibitor) inhibited the antidepressant effect and reduced IL-1β and IL-6. PLX3397 (a microglial depletor) abolished both the antidepressant effect and the pro-inflammatory and depressive effects. |
Gao et al., 2019 [120] |
| Nanoemulsion | Curcumin | 116,0 ± 0,305 nm PDI: 0,121 ± 0,007 PZ: −11,6 ± 1,23 mV EE: 98,45 ± 0,31% |
TUR-NE showed 36.51 ± 3.24% release in HCl and 44.90 ± 2.47% release in PBS over 48 hours | TUR exhibited the highest antioxidant activity (ABTS•⁺), followed by CU and Vitamin C, with IC₅₀ values of 17.9, 29.1, and 57 µg/mL, respectively | TUR-NE increased sucrose preference (77.5% vs. 54.0% in CUMS), decreased immobility time in the forced swim test (FST) (P < 0.01), reduced latency to feed in the novelty-suppressed feeding test (NSFT), and elevated plasma serotonin levels (21.7 vs. 16.3 ng/mL) as well as brain serotonin levels (46.9 vs. 44.0 ng/mL); MAO activity showed no significant changes. | Sheng et al. (2023) [121] |
| Self-emulsifying system | Curcumin | 150 nm | 100% release within 5 minutes | 7.4 ± 0.2 mL (sucrose) | 103.4 ± 5.8 s (FST) | 247 ± 3.1 g (weight) | Partial neuroprotective and hepatoprotective effects | Suchiwa Pan et al. (2023) [122] | |
| Dendrimers | Polyamidoamine | 4 nm | - | Increased Microglial Uptake: Uptake of PAMAM dendrimers by microglial cells was faster and more extensive in cerebral slices from rabbits with Cerebral Palsy (CP) compared to healthy controls. It was 1.6 times higher after 4 hours of treatment, with 80% of cells containing dendrimers in CP slices. | Selective Accumulation: PAMAM dendrimers selectively accumulated in activated microglia within the brains of newborn rabbits. Therapeutic Outcome: When conjugated with N-acetylcysteine (NAC), this accumulation led to a dramatic improvement in motor function and attenuated neuroinflammation. |
Zhang et al., 2016 [123] |
Legend: DA stands for dopamine; 5-HT, serotonin; PDI, polydispersity index; PZ, zeta potential; EE, encapsulation efficiency; NMDA, N-methyl-D-aspartate; Res, resveratrol; THS, thermosensitive hydrogel system; MDA, malondialdehyde (an oxidative stress marker); GSH, reduced glutathione (a cellular antioxidant); NLC C, curcumin-loaded lipid nanocapsules; Aβ₂₅₋₃₅, beta-amyloid peptide 25-35; TCA, open field test; TUR, curcuma extract; NE, nanoemulsion; C₅₀, half maximal inhibitory concentration; ABTS•⁺, ABTS cation radical; and CU, curcumin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



