
Submitted:
18 July 2025
Posted:
22 July 2025
You are already at the latest version
Abstract
Inflammatory bowel diseases (IBD), including Crohn's disease (CD) and ulcerative colitis (UC), are chronic conditions marked by dysregulated inflammation in the gastrointestinal tract. Although the pathophysiology of IBD remains incompletely understood, it involves complex interactions between genetic predisposition and environmental triggers, such as gut microbiota imbalances and immune dysfunction, leading to chronic inflammation and mucosal injury. IBD affects approximately 7 million individuals globally, with prev-alence increasing in Europe, North America and Oceania. This rise parallels the growing consumption of ultra-processed foods (UPF), which are typically rich in sugars, fats, and additives but low in fiber, vitamins, and other essential nutrients. High UPF consumption is associated with intestinal dysbiosis, marked by reduced microbial diversity, decreased short-chain fatty acid production, impaired barrier integrity, and mucus layer disruption. These alterations may promote immune-mediated diseases, including IBD, where dysbio-sis is often characterized by an overgrowth of pathogenic bacteria such as Clostridium and Enterococcus, ultimately triggering inflammatory responses in the host. Considering these associations, this review critically examines the influence of UPF consumption on gut mi-crobiota composition and function, and its potential link to IBD.
Keywords:
ultra-processed food
; gut microbiota
; Crohn's disease
; ulcerative colitis
; inflammatory bowel disease
1. Introduction
Over the years, various methods of obtaining, preserving, and preparing food have been developed to ensure survival in adverse environmental conditions. [1] The transition from nomadic societies to agricultural communities marked the beginning of a more direct relationship with food production, primarily based on minimally processed and seasonal ingredients. The Industrial Revolution brought about a profound transformation in production, distribution, and consumption of food. The emergence of preservation, refining, and industrialization technologies enabled large-scale production and the creation of products with extended shelf life, enhanced convenience, and increased palatability. However, these advancements have shifted dietary patterns away from traditional diets and increased reliance on highly processed foods. [1,2,3]
In recent decades, this phenomenon has intensified, particularly in developed countries and increasingly in developing nations, leading to the widespread adoption of Westernized dietary patterns. [3] These patterns are characterized by a high consumption of sugars, saturated fats, chemical additives, and ultra-processed foods (UPF). This significant dietary shift, coupled with an increasingly sedentary lifestyle, has been directly implicated in the escalating prevalence of non-communicable chronic diseases (NCDs), including type 2 diabetes, obesity, cardiovascular diseases, cancer, and various intestinal disorders. [4,5,6,7] Among these conditions, inflammatory bowel diseases (IBD) are particularly notable, with incidence rising alarmingly even in regions historically characterized by low prevalence, such as Latin America, Asia, and Africa. [8] The etiopathogenesis of IBD is multifactorial, involving genetic predisposition, immunological alterations, environmental factors, and, critically, gut microbiota dysbiosis. [3,9] The observed rise in IBD incidence is occurring concurrently with the increased intake of UPF across various countries. [10]
Considering this compelling evidence and the global impact of UPF consumption, this review aims to describe the role of UPF in the development of inflammatory bowel disease, explore its implications for gut microbiota composition and function, and discuss its potential associations with IBD risk.
2. Ultra-Processed Foods
According to the NOVA classification, UPF is an industrial formulation made from food-derived ingredients or synthesized in laboratories. UPF often contain additives such as emulsifiers, sweeteners, preservatives, and colorants, and may include components of natural foods, all of which are characterized by a high degree of processing. [11,12] (Figure 1)
UPF accounts for up to 58% of daily caloric intake in high-income countries and approximately 30% in emerging economies. This widespread availability and consumption, particularly since the 1970s, coincides with a rise in NCDs, including IBD. [13] These products, such as industrial sweets, snacks, breads, cereals, margarines, processed meats, soft drinks, boxed juices, and industrial alcoholic beverages, are nutritionally imbalanced, highly palatable, and easy to consume. Their high content of refined sugars, fats (especially n-6 polyunsaturated fatty acids), and low fiber content have been identified as risk factors for the development of IBD. [4,7] Another significant concern is the use of food additives. Substances like stabilizers, colorants, and emulsifiers are added to alter foods' physical, chemical, and sensory characteristics. Although approved by regulatory agencies, these compounds have been associated with alterations in the composition and function of the gut microbiota. [14]
While UPF are associated with microbial imbalances and increased intestinal inflammation, diets rich in fiber and minimally processed foods promote greater microbial diversity, playing a role in preventing chronic inflammatory diseases. Thus, the increasing consumption of UPF, paralleling the rising incidence of IBD in many countries, raises serious concerns about their impacts on gut health and systemic inflammation. [10]
3. Impact of Ultra-Processed Foods on Gut Microbiota and Intestinal Homeostasis
The gut microbiota plays a vital role in human health, constantly adapting to its intestinal environment. It provides numerous physiological benefits in a "healthy state", known as eubiosis or normobiosis. These processes include metabolizing dietary nutrients, fermenting prebiotic fibers, synthesizing vitamins B and K, secreting antimicrobial compounds, modulating immune responses, and maintaining the integrity of the intestinal epithelial barrier to prevent pathogen adhesion and colonization. [15,16] However, the consumption of UPF is consistently linked to significant alterations in gut microbiome composition and low-grade inflammation. Specifically, an increase in microbial taxa associated with obesity and metabolic disorders is observed, alongside a decline in beneficial groups like Bacteroides, Verrucomicrobia, Eubacterium rectale, Coprococcus coccoides, and Bifidobacterium. This dysbiotic shift often produces a proportional rise in pro-inflammatory phyla such as Bacillota and Pseudomonadota. This imbalance, coupled with elevated pro-inflammatory cytokines, including interleukin-1 (IL-1), interleukin-6 (IL-6), and Tumor Necrosis Factor (TNF-α)) and lipopolysaccharides (LPS), drives processes such as hyperinsulinemia, adipogenesis, and hepatic steatosis. [17,18]
The persistent low-grade inflammation driven by UPF consumption can also lead to a "leaky gut" or increased intestinal permeability. [5,19] This compromised barrier then facilitates the translocation of pathogenic bacteria and LPS into the bloodstream, significantly exacerbating the inflammatory response observed in IBD. LPS plays a central role by activating the innate immune system, triggering the recruitment of macrophages, neutrophils, and dendritic cells through Toll-Like Receptor (TLR) signaling, which in turn releases key pro-inflammatory mediators, such as IL-1α, IL-1β, TNF-α, and IL-6. [5,19,20] For instance, serum LPS levels are reportedly six times higher in patients with active Crohn's disease (CD) compared to healthy controls, highlighting its potential to intensify CD-related inflammation. [21] Some studies suggest that the IL-23/IL-17 axis, central to CD pathogenesis, may increase the intestinal epithelium's susceptibility to LPS exposure, further exacerbating inflammation. [22] While the evidence directly linking UPF to UC is less consistent—possibly due to its complex pathogenesis—circulating LPS may still act as a precursor in UC progression. [23]
Beyond LPS, flagellin levels have also been associated with CD inflammation and intestinal barrier integrity. [24] Although its exact mechanism is not fully elucidated, the hypothesis linking flagellin to CD pathogenesis suggests it may drive T-cell and B-cell immune responses, potentially arising from compromised transepithelial electrical resistance to pro-inflammatory cytokines after flagellin exposure, leading to reduced tight junction proteins. [25]
4. Impact of Food Additives on Gut Microbiota
Food additives, commonly found in UPF, are designed to enhance product characteristics and extend shelf life. However, a growing body of evidence indicates they can profoundly impact gut microbiota composition and function, thereby contributing to intestinal inflammation and dysbiosis. Studies in both humans and animals show that these additives can trigger pro-inflammatory changes in the microbiota, enhance LPS release, disrupt the production of beneficial metabolites like short-chain fatty acids (SCFAs), and impair the mucus layer and β-defensin expression. [26,27,28] Indeed, diets in which UPF constitutes over 20% of total daily caloric intake may compromise mucus barrier integrity, impair goblet cell function, and increase the risk of disease. [29] (Table 1).
4.1. Emulsifiers
In vitro study utilizing fecal samples from healthy individuals has investigated the effects of common emulsifiers dietary on the gut microbiome, including propylene glycol monostearate (PGMS) and sodium stearoyl lactylate (SSL). These latter two, frequently found in kinds of margarine, baked goods, and desserts, significantly reduced butyrate-producing families such as Clostridiaceae, Lachnospiraceae, and Ruminococcaceae, while increasing Bacteroidaceae and Enterobacteriaceae. SSL was also linked to elevated LPS and flagellin levels, possibly due to increased production by Escherichia. [39]
Specifically, carboxymethylcellulose (CMC) may promote bacterial adherence to the intestinal epithelium, facilitating bacterial proliferation and infiltration between intestinal villi. It can also alter the gut microbial composition, decreasing beneficial species such as Faecalibacterium prausnitzii and Ruminococcus spp. (both associated with SCFA production), while increasing Roseburia spp. and Lachnospiraceae bacterium. [31,36] Furthermore, consuming CMC and the emulsifier polysorbate 80 (P80) has been associated with elevated levels of Pseudomonadota, which promote mucosal inflammation. These changes can also increase bacterial adherence, penetration into the mucus layer, and migration toward intestinal crypts, amplifying the pro-inflammatory potential and promoting chronic inflammation. [40] P80 intake may even facilitate the translocation of bacteria, such as Escherichia coli, through M cells and Peyer's patches, thereby compromising intestinal barrier integrity. [41,42]
Xanthan Gum (INS 415), an extracellular polysaccharide from Xanthomonas campestris, is a common food additive. Recent studies suggest that human gut microbiota can partially degrade xanthan gum. [43] Individuals with Westernized gut microbiota often harbor specific bacterial species, particularly from the Ruminococcaceae family, that can metabolize xanthan gum into oligosaccharides, which are subsequently utilized by Bacteroides intestinalis, forming a microbial food chain. This degradation can lead to the production of SCFAs, including butyrate, which is known for its anti-inflammatory and gut-protective actions. [43] However, the effects are not uniformly beneficial. Other studies suggest that xanthan gum can induce alterations in the microbiota, promoting the proliferation of opportunistic bacteria, such as Ruminococcus gnavus and members of the Enterobacteriaceae family, both of which are associated with intestinal inflammation. [43] For instance, a study in Wistar rats administered xanthan gum found a significant increase in inflammatory cytokines (IL-1β and TNF-α) and harmful modulation of epithelial tight junction proteins, such as Claudin-2, suggesting that at high doses or in predisposed individuals, xanthan gum may compromise intestinal barrier integrity and foster an inflammatory environment. [35,43]
4.2. Non-Caloric Artificial Sweeteners
Non-caloric artificial sweeteners (NAS) are widely used for their calorie-free sweetness and have been implicated in adverse gut health outcomes. Suez et al. [30] observed that NAS intake impairs glucose tolerance in both humans and rodents, with changes directly linked to the gut microbiota, including an overgrowth of Bacteroides and a reduction in Bacteroidota, particularly Clostridiales species. Another study linked the consumption of artificial sweeteners, including sucralose, to alterations in gut microbiota and an increased inflammatory potential, suggesting intestinal barrier dysfunction and elevated inflammatory markers such as TNF-α and IL-6. [3]
4.3. Maltodextrin
Maltodextrin, a widely used polysaccharide additive, improves food texture, flavor, and stability. However, frequent consumption may be linked to changes in microbial phenotype and host antibacterial defenses. [44] A study by Laudisi et al. [32] observed that rats given a 5% maltodextrin solution in water for 45 days exhibited a reduction in intestinal mucin 2 (Muc-2) and an increase in epithelial adhesion of pathogenic bacteria, which could contribute to susceptibility to intestinal diseases. [32]
4.4. Carrageenan
Carrageenan (CGN) is a natural polysaccharide extracted from red algae, widely used in the industry for its gelling, thickening, and stabilizing properties. [45] However, its use has been questioned due to its potential breakdown in the gastrointestinal tract, leading to the formation of degraded carrageenans (dCGNs), which may pose health risks. [46] An in vitro study by Han et al. observed that the formation of dCGNs by the intestinal microbiota significantly increased the production of nitric oxide (NO2), cyclooxygenase-2 (COX-2), and pro-inflammatory cytokines like IL1-b, TNF-a, and IL-6, which suggests a possible link to the development of IBD. [38]
4.5. Synthetic Colorants (Azo Dyes)
Synthetic colorants have experienced a dramatic 500% increase in use over the past five decades, primarily to enhance product appearance. [47,48] These colorants, typically derived from petroleum, release reactive or toxic aromatic amines when metabolized in the gut. These metabolites possess carcinogenic potential and are associated with other chronic diseases. [48,49] Azo dyes (AZo), including Allura Red, Sunset Yellow, Amaranth, Tartrazine, Red 40, and Yellow 6, though permitted in food, have been linked to microbiota-related disease pathophysiology. [50] The Lachnospiraceae family, including species like Clostridium symbiosum and Hungatella effluvi, appears to be a primary group capable of reducing AZo dyes. [49] He et al. administered Red 40 and Yellow 6 to genetically susceptible rodents. They found that commensal bacteria like Bacteroides ovatus and Enterococcus faecalis metabolized these dyes to form sodium salt 1-amino-2-naphthol-6-sulfonate (ANSA-Na). [34] This metabolite was then implicated in colitis induction by activating pathogenic CD4+ T cells through the IL-23 axis, stimulating IFN-γ production, and intensifying intestinal inflammation in response to exposure to Red 40 and Yellow 6. [34]
4.6. Nanoparticles and Microparticles
Nanoparticles are increasingly used to enhance product characteristics such as color, texture, and appearance while improving stability and shelf life. [51] An in vitro study evaluating the absorption and biological impact of iron oxide (E172) nanoparticles subtypes (yellow, orange, red, and black) found that all variants interacted with intestinal barrier cells. However, none demonstrated cytotoxic effects at concentrations below 100μg Fe/mL after 24 hours of exposure. [37]
Commonly used microparticles include titanium dioxide (TiO₂ – E171), a whitening agent; aluminum silicate (AlSi – E559), an anti-caking agent; and silicon dioxide (E551). [52] In 2023, the European Food Safety Authority raised concerns about the genotoxic potential of TiO₂. Despite this, its use remains ‘authorized pending further review’ by Joint FAO/WHO Expert Committee on Food Additives (JECFA). [53] TiO₂ can alter the intestinal environment by reducing the expression of Muc2, a key protein in mucus layer formation, directly affecting goblet cell function, impairing mucus production and compromising intestinal barrier integrity. [33] In vitro research indicates that goblet cells efficiently absorb TiO₂, which may interfere with their ability to secrete mucus effectively. [33] Rodent studies show that TiO₂ ingestion leads to its uptake by intestinal cells, initiating a pro-inflammatory cascade. Prolonged exposure has been linked to the release of reactive oxygen species (ROS), resulting in altered gene transcription and promoting dysplasia. [26,52,54,55]
In this context, the harmful effects of UPF components, such as emulsifiers, sweeteners, synthetic colorants, and nanoparticles, are evident through their ability to induce inflammation, alter the gut microbiome, and reduce populations of beneficial microorganisms, thereby possibly contributing to the development of IBD.
5. Dysbiosis and Modulation of the Intestinal Microbiota in IBD
5.1. Microbial Dysbiosis in IBD
IBD is strongly linked to gut dysbiosis, a microbial imbalance characterized by an overgrowth of pathogenic bacteria such as Clostridium and Enterococcus. [9] This microbial alteration compromises mucosal integrity, increases intestinal permeability and facilitates bacterial translocation, ultimately leading to the intense activation of host inflammatory responses. [56] In individuals with IBD, a significant reduction in the phyla Bacillota (formerly Firmicutes) and Bacteroidota (formerly Bacteroidetes) is commonly observed. [57] This decrease impairs the modulation of inflammatory responses and the production of SCFAs. Conversely, opportunistic pathogens, including certain Pseudomonadota (formerly Proteobacteria) species, proliferate and can exacerbate underlying inflammatory conditions (Figure 2). [58,59] Studies, such as that by Santana et al., [60] confirm that individuals with IBD show a decreased abundance of beneficial bacteria and an increased presence of species that contribute to ongoing mucosal damage (Table 2 and Table 3).
SCFAs, particularly acetate, propionate, and butyrate, are microbial fermentation products with significant therapeutic relevance for human health. However, their levels are often altered in individuals with IBD. [61] Among these, butyrate is one of the most extensively studied in this population. It is produced by bacteria such as Roseburia spp., Eubacterium rectale, Faecalibacterium prausnitzii, and Clostridium leptum (phylum Bacillota), as well as Akkermansia muciniphila (phylum Verrucomicrobia). Butyrate serves as the main energy source for intestinal epithelial cells and supports cell signaling, proliferation, and differentiation. In addition, it has strong anti-inflammatory effects mediated through the activation of G-protein-coupled receptors. [62,63].
SCFAs also play a critical role in modulating immune responses by inhibiting the NF-κB (Fator Nuclear Kappa B) and histone deacetylases, thereby regulating innate and adaptive immunity. [64] These mechanisms support the restoration of tissue damaged by inflammation. [61] In patients with ulcerative colitis treated with infliximab, enzymes involved in butyrate oxidation (excluding SLC16A1) were found to be elevated in those who responded to biological therapy. These findings suggest that mucosal inflammation interferes with butyrate oxidation. [65] Consequently, the results indicate a correlation between inflammation and gene suppression in SCFA absorption and metabolism.
Obesity and diets high in trans and saturated fats, emulsifiers, processed foods, and alcohol contribute to elevated endotoxin levels, oxidative stress, and the expansion of pathogenic bacterial populations while reducing SCFA production. These changes promote inflammation and increase the risk of other comorbidities, including diabetes, metabolic dysfunction-associated steatohepatitis (MASLD), [66] and various cardiovascular and neurological conditions. [67,68,69]
5.2. Gut Microbiota Modulation in IBD
Modulating the gut microbiota has emerged as a key strategy in managing IBD9 and other chronic conditions. [70,71,72] Various factors, including long-term dietary patterns, physical activity, smoking, sleep quality, alcohol consumption, stress, medication use, and aging, influence the gut microbiota composition. [73] Several approaches are employed to promote intestinal microbiota modulation in individuals with IBD, described below. However, despite promising results in vitro and in experimental models, robust clinical evidence supporting their effectiveness in IBD patients remains limited.
5.3. Dietary Patterns
The Mediterranean Diet is a well-researched dietary pattern for its health-promoting effects. [74,75] Rich in fiber, vitamins, minerals, and bioactive compounds, it's associated with a lower risk of IBD development and improved inflammatory markers, alongside a protective effect on the intestinal mucosa. [76,77] Its benefits include reducing pro-inflammatory bacterial species (Escherichia coli, Ruminococcus gnavus), increased production of SCFAs, fecal secretory immunoglobulin A (sIgA), and enhanced microbiota diversity and intestinal barrier function. [78,79,80] Clinical studies support its efficacy. Dogan et al. [81] observed reduced inflammatory biomarkers in UC patients, and Chicco et al. [82] noted clinical and quality-of-life improvement. Conversely, high consumption of UPF is associated with impaired metabolic and intestinal health, leading to increased inflammation and dysbiosis. [83]
5.4. Prebiotics
Prebiotics are defined as "substrates that are selectively utilized by host microorganisms, providing a health benefit". [84]. Examples of such substrates include pectin, inulin, and resistant starch. [85] While the European Society for Clinical Nutrition and Metabolism (ESPEN) suggests potential as an adjunctive therapy in IBD, evidence remains limited. [86] A Cochrane systematic review found insufficient evidence to support their efficacy for induction and maintenance of remission in mild-to-moderate UC, likely due to a few studies. [87]
5.5. Probiotics
Probiotics are live microorganisms that confer health benefits to the host when administered adequately. [84,88] A 2020 Cochrane review analyzing probiotics for maintaining remission in UC found no significant differences compared to other interventions. [89] ESPEN advises against their use for inducing or maintaining remission in CD, as they are generally ineffective. [86] However, probiotics may be considered for mild to moderately active UC and are also recommended for preventing pouchitis. Despite these indications, current evidence does not support their routine use in clinical practice. [90]
5.6. Symbiotic
A symbiotic is defined as a mixture containing live microorganisms and substrates selectively utilized by host microorganisms, offering a health benefit. [91] A meta-analysis suggests that symbiotics may serve as an adjunctive therapy for moderate UC [92], but the limited number of studies necessitates further research. Preclinical studies show promise, but human studies have small sample sizes. [93,94]
5.7. Postbiotics
Postbiotics are preparations of inactivated microorganisms and/or their components confer a health benefit to the host. [95] Studies suggest postbiotics may aid immune modulation via SCFA production, improve intestinal barrier integrity, promote intestinal homeostasis, and inactivate inflammatory pathways. [96] However, most studies with IBD are preclinical or in vitro, with heterogeneous outcomes, underscoring the need for more robust clinical trials. [97]
5.8. Fecal Microbiota Transplantation (FMT)
Fecal Microbiota Transplantation (FMT) involves transferring stool from a healthy donor to a recipient's intestinal microbiota. [98] Primarily studied in recurrent Clostridium difficile infections and selected metabolic disorders, [97,99,100,101] a Cochrane review suggests FMT may be a promising therapeutic option for inducing remission in mild-to-moderate active UC. [102] Nevertheless, further clinical trials are essential before FMT can be routinely recommended for treating IBD. [90]
6. The Complex Relationship: UPF, Gut Microbiota, and IBD
The interplay between diet, gut microbiota, and IBD has been a significant research focus in recent decades. [103] Among various dietary factors, UPF has emerged as a critical contributor to gut dysbiosis and the exacerbation of inflammatory conditions, including CD and UC. [103]
Chronic and high intake of UPF is consistently associated with a pro-inflammatory and pro-oxidant metabolic profile [18] mediated through interconnected physiological pathways, including elevated ghrelin levels, reduced peptide YY (PYY) levels, endothelial dysfunction, altered glucose metabolism, hormonal imbalances, and insulin resistance. [104] Furthermore, a higher prevalence of pro-carcinogenic bacteria is observed in the gut microbiota, whose metabolites can induce genotoxic DNA damage. [105] These microbial alterations may also compromise the gut-brain axis, potentially impairing cognitive function and mental health. [105,106] Elevated UPF consumption, especially in older adults, has been associated with reduced antioxidant enzyme activity (e.g., catalase and superoxide dismutase) and increased levels of xanthine oxidase. [107] These findings indicate increased oxidative stress and diminished antioxidant defenses, accompanied by higher levels of pro-inflammatory cytokines, such as TNF-α, IL-6, and IL-15, and increased production of reactive oxygen species (ROS). [106] The heightened activity of myeloperoxidase (MPO), a pro-oxidant enzyme, further contributes to this pro-oxidative state. [107] These mechanisms may exacerbate endothelial dysfunction, thereby increasing the risk of intestinal ischemia, particularly in vulnerable populations. [104,107] Supporting these observations, a positive association between UPF consumption and IL-6 levels has been found in cohorts from diverse socioeconomic backgrounds. [35] Ultimately, UPF intake significantly influences IBD development and progression by promoting excessive ROS production and perpetuating oxidative stress in intestinal tissues. This oxidative environment directly contributes to cellular dysfunction and can induce necroptosis, thereby amplifying the inflammatory response and exacerbating the clinical course of IBD. [108,109]
Excessive UPF intake is linked to reduced beneficial bacteria, such as Faecalibacterium prausnitzii, and decreased production of butyrate, an SCFA essential for maintaining intestinal barrier integrity, as discussed previously. [36,40] A healthy gut microbiota is crucial for immune modulation and intestinal homeostasis, thereby preventing chronic inflammation and protecting against infections. In IBD, this homeostasis is frequently disrupted, leading to dysbiosis marked by reductions in beneficial commensals (e.g., Bacillota and Bacteroidota) and increases in intestinal pathogens (e.g., Escherichia coli and Pseudomonadota). [110] A disrupted microbiota in IBD is associated with elevated pro-inflammatory cytokines (including TNF-α and IL-6) and the activation of inflammatory signaling pathways like nuclear factor kappa B (NF-kB), contributing to sustained gastrointestinal inflammation. [110]
External influences, particularly diets high in UPF, exacerbate the pro-inflammatory state, which favors pathogenic bacteria growth and promotes excessive cytokine production. [17] The chronic inflammatory state stemming from UPF intake appears to be a key pathophysiological mechanism contributing to IBD development, particularly in CD. This disruption facilitates bacterial translocation and microbial imbalance, which activate an exaggerated immune response when combined with genetic predisposition, thereby increasing the risk of IBD onset. [26,111,112,113] (Figure 3).
A robust cohort study involving 245,112 participants in the United States identified 857 IBD cases. [99] High UPF intake was associated with an increased risk of developing CD, linked explicitly to ultra-processed bread, breakfast cereals, and ready-to-heat frozen meals. [99] This association may be attributed to the high levels of carrageenan, synthetic emulsifiers, and sodium chloride in these products, which disrupt the intestinal barrier by promoting the proliferation of pro-inflammatory bacteria within the Pseudomonadota phylum.. [99] While this process impairs epithelial integrity and reduces the mucus layer, favoring colitis development, the study found no statistically significant evidence for an association between UPF consumption and UC onset. [99]
Conversely, a 2021 study tracking over 150,000 participants across 31 countries found that higher UPF intake was associated with an increased risk of both CD and UC. However, gut microbiota composition was not assessed. [113] Consumption of UPF, particularly processed meats and soft drinks, was more prevalent in North America, Europe, and South America. [113] However, no food group or additive was independently linked to a higher risk of IBD, emphasizing that overall dietary patterns significantly influence disease pathogenesis. [113] Interestingly, a study in a Caucasian population from eight European countries (over 400 participants, 13.2-year follow-up) found no association between increased UPF consumption and a greater IBD risk. [114] Instead, whole and minimally processed foods appeared to protect against CD, with no significant association for UC incidence. [115]
Beyond incidence rates, research is increasingly focusing on the role of diet in disease progression. [99] Preda et al. (115) conducted a clinical trial of 168 participants with CD/UC. It showed sustained remission in 95.2% of a low-UPF intervention group versus 85.7% in a control group (p = 0.036), after one year, underscoring the role of lifestyle habits in IBD management. [115] Observational studies also suggest an inverse association between high fiber intake and CD risk, though UC results remain inconclusive. [115] Various dietary strategies, such as the CD Exclusion Diet (CDED), have been proposed to support remission in IBD. [115] CDED onsists of structured phases that remove components and additives harmful to the microbiota and intestinal barrier, while emphasizing high-quality proteins and foods that support microbial diversity. This diet is associated with reduced Pseudomonadota and increased Bacilliota. [116] A CDED study showed 76.7% remission by week 6, rising to 82.1% by week 12. [117] In a trial involving children with mild to moderate CD, CDED led to remission, reduced inflammatory markers (C-reactive protein and ESR), fewer bowel movements, and lower fecal calprotectin levels. Participants also reduced their UPF intake and improved their adherence to a Mediterranean diet. [118] The diet's maintenance phase is critical for developing and sustaining healthy eating habits, reinforcing the diet's importance in gut health and microbiome. [119]
7. Final Considerations
Excessive consumption of UPF can lead to various physiological alterations and impair gut microbiota function, contributing to the development of NCDs, including IBD. The impact of UPF on the gut microbiome and its association with the development and progression of IBD is a growing area of research. A high consumption of these foods promotes dysbiosis, increases intestinal permeability, and activates inflammatory pathways, which may exacerbate conditions such as CD and UC. Emerging evidence also supports the link between UPF consumption and increased IBD incidence, particularly CD. Conversely, diets rich in fresh, minimally processed foods promote gut health and may reduce inflammation, improving disease control in individuals with IBD. It is worth noting that few studies have evaluated the effect of UPF consumption on microbiota and the risk of IBD. Most studies have focused specifically on food additives, an area of knowledge essential for informing public policy and guiding regulatory measures. Modulating the gut microbiota through healthy dietary patterns represents a promising approach for preventing and managing IBD in at-risk populations. However, further research is necessary to clarify the underlying mechanisms and determine the most effective dietary interventions. Moreover, nutrition-based strategies aimed at modulating microbiota are expected to play an increasingly vital role in combating chronic diseases, enhancing patient quality of life, and reducing the broader economic and societal burdens associated with these conditions.
Author Contributions
S.A.L, C.B.G, N.E.J.Z, Y.R.N.Y, C.N.S, S.A, D.F. M.B, M. D.O, and S LY contributed equally to the conception and design of the article, writing, and editing of the manuscript and review of the literature. All the authors approved the final version of the article to be published. “Conceptualization, S.A.L.; methodology, S.A.L, C. B. G, Y., R.N.Y and C. N.S; software, S.A.L, C. B. G, Y., and R.N.Y..; writing—review and editing, S.A.L, C.B.G, N.E.J.Z, Y.R.N.Y, C.N.S.; supervision, , S.A, D.F.M.B, M. D.O, and S.L.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or non-profit sectors This research received no external funding.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
Spiller AL, Costa BG, Castelhano NS have received a Master scholarship grant from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Programa de Excelência Acadêmica (PROEX) – Finance Code 88887.807663/2023-00.
Abbreviations
The following abbreviations are used in this manuscript:
| CMC | Carboxymethylcellulose |
| CD | Crohn's disease |
| CDED | Crohn's disease Exclusion Diet |
| IBD | Inflammatory bowel diseases |
| IL-1 | Interleukin-1 |
| IL-6 | Interleukin-6 |
| IL-10 | Interleukin-10 |
| IL-18 | Interleukin-18 |
| LPS | Lipopolysaccharides |
| MASLD | Metabolic dysfunction-associated steatohepatitis |
| NAS | Non-caloric artificial sweeteners |
| NCDs | Non-communicable chronic diseases |
| NF-kB | Fator Nuclear Kappa B |
| SCFAs | Short-chain fatty acids |
| SFA | Saturated fatty acids |
| TIO2 | Titanium dioxide |
| TLR | Toll-like receptor |
| TNF-α | Tumor Necrosis Factor |
| UC | Ulcerative colitis |
| UPF | Ultra-processed foods |
References
- Ultra-processed foods and the nutrition transition: Global, regional and national trends, food systems transformations and political economy drivers - Baker - 2020 - Obesity Reviews - Wiley Online Library [Internet]. [citado 9 de julho de 2025]. Disponível em: https://onlinelibrary.wiley.com/doi/10.1111/obr.13126.
- Ultra-processed products are becoming dominant in the global food system - Monteiro - 2013 - Obesity Reviews - Wiley Online Library [Internet]. [citado 9 de julho de 2025]. Disponível em: https://onlinelibrary.wiley.com/doi/10.1111/obr.12107.
- Bernard Srour*, Melissa C Kordahi*, Erica Bonazzi*, Mélanie Deschasaux-Tanguy, Mathilde Touvier†, Benoit Chassaing. Ultra-processed foods and human health: from epidemiological evidence to mechanistic insights. Lancet Gastroenterol Hepatol. 8 de agosto de 2022;
- Jardim, M.Z.; Costa BVde, L.; Pessoa, M.C.; Duarte, C.K. Ultra-processed foods increase noncommunicable chronic disease risk. Nutrition Research. 1o de novembro de 2021, 95, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Babaei, A.; Pourmotabbed, A.; Talebi, S.; Mehrabani, S.; Bagheri, R.; Ghoreishy, S.M.; et al. The association of ultra-processed food consumption with adult inflammatory bowel disease risk: a systematic review and dose-response meta-analysis of 4 035 694 participants. Nutrition Reviews. 1o de julho de 2024, 82, 861–71. [Google Scholar] [CrossRef] [PubMed]
- Christensen, C.; Knudsen, A.; Arnesen, E.K.; Hatlebakk, J.G.; Sletten, I.S.; Fadnes, L.T. Diet, Food, and Nutritional Exposures and Inflammatory Bowel Disease or Progression of Disease: an Umbrella Review. Advances in Nutrition. 1o de maio de 2024, 15, 100219. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.M.; Gamage, E.; Du, S.; Ashtree, D.N.; McGuinness, A.J.; Gauci, S.; et al. Ultra-processed food exposure and adverse health outcomes: umbrella review of epidemiological meta-analyses. BMJ. 28 de fevereiro de 2024, 384, e077310. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies. The Lancet. 23 de dezembro de 2017, 390, 2769–78. [Google Scholar]
- Magro, D.O.; Rossoni, C.; Saad-Hossne, R.; Santos, A. INTERACTION BETWEEN FOOD PYRAMID AND GUT MICROBIOTA. A NEW NUTRITIONAL APPROACH. Arq Gastroenterol. 12 de maio de 2023, 60, 132–6. [Google Scholar] [CrossRef] [PubMed]
- Racine, A.; Carbonnel, F.; Chan, S.S.M.; Hart, A.R.; Bueno-de-Mesquita, H.B.; Oldenburg, B.; et al. Dietary Patterns and Risk of Inflammatory Bowel Disease in Europe: Results from the EPIC Study. Inflammatory Bowel Diseases. 1o de fevereiro de 2016, 22, 345–54. [Google Scholar] [CrossRef] [PubMed]
- Louzada, M.L.D.C.; Gabe, K.T. Classificação de alimentos Nova: uma contribuição da epidemiologia brasileira. Rev bras epidemiol [Internet]. 2025 [citado 9 de julho de 2025];28. Disponível em: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1415-790X2025000100201&tlng=pt.
- Monteiro, C.A.; Cannon, G.; Levy, R.B.; Moubarac, J.C.; Louzada, M.L.; Rauber, F.; et al. Ultra-processed foods: what they are and how to identify them. Public Health Nutr. abril de 2019, 22, 936–41. [Google Scholar] [CrossRef] [PubMed]
- Hracs, L.; Windsor, J.W.; Gorospe, J.; Cummings, M.; Coward, S.; Buie, M.J.; et al. Global evolution of inflammatory bowel disease across epidemiologic stages. Nature. junho de 2025, 642, 458–66. [Google Scholar] [CrossRef] [PubMed]
- Tabela de Aditivos - ANVISA.
- Llavero-Valero, M.; Martín, J.E.S.; Martínez-González, M.A.; Basterra-Gortari, F.J.; Fuente-Arrillaga Cde la Bes-Rastrollo, M. Ultra-processed foods and type-2 diabetes risk in the SUN project: A prospective cohort study. Clinical Nutrition. 1o de maio de 2021, 40, 2817–24. [Google Scholar] [CrossRef] [PubMed]
- LaFata, E.M.; Allison, K.C.; Audrain-McGovern, J.; Forman, E.M. Ultra-Processed Food Addiction: A Research Update. Curr Obes Rep. 1o de junho de 2024, 13, 214–23. [Google Scholar] [CrossRef] [PubMed]
- Kendig, M.D.; Hasebe, K.; McCague, R.; Lee, F.; Leigh, S.J.; Arnold, R.; et al. Adolescent exposure to a solid high-fat, high-sugar 'cafeteria' diet leads to more pronounced changes in metabolic measures and gut microbiome composition than liquid sugar in female rats. Appetite. 1o de maio de 2022, 172, 105973. [Google Scholar] [CrossRef] [PubMed]
- Mignogna, C.; Costanzo, S.; Castelnuovo, A.D.; Ruggiero, E.; Shivappa, N.; Hebert, J.R.; et al. The inflammatory potential of the diet as a link between food processing and low-grade inflammation: An analysis on 21,315 participants to the Moli-sani study. Clinical Nutrition. 1o de outubro de 2022, 41, 2226–34. [Google Scholar] [CrossRef] [PubMed]
- Bolte, L.A.; Vich Vila, A.; Imhann, F.; Collij, V.; Gacesa, R.; Peters, V.; et al. Long-term dietary patterns are associated with pro-inflammatory and anti-inflammatory features of the gut microbiome. Gut. julho de 2021, 70, 1287–98. [Google Scholar] [CrossRef] [PubMed]
- Martínez Leo, E.E.; Segura Campos, M.R. Effect of ultra-processed diet on gut microbiota and thus its role in neurodegenerative diseases. Nutrition. 1o de março de 2020, 71, 110609. [Google Scholar] [CrossRef] [PubMed]
- Magro, D.O.; Kotze, P.G.; Martinez, C.A.R.; Camargo, M.G.; Guadagnini, D.; Calixto, A.R.; et al. Changes in serum levels of lipopolysaccharides and CD26 in patients with Crohn's disease. Intest Res. julho de 2017, 15, 352–7. [Google Scholar] [CrossRef] [PubMed]
- Hölttä, V.; Klemetti, P.; Sipponen, T.; Westerholm-Ormio, M.; Kociubinski, G.; Salo, H.; et al. IL-23/IL-17 immunity as a hallmark of Crohn's disease. Inflamm Bowel Dis. setembro de 2008, 14, 1175–84. [Google Scholar] [CrossRef] [PubMed]
- Candelli, M.; Franza, L.; Pignataro, G.; Ojetti, V.; Covino, M.; Piccioni, A.; et al. Interaction between Lipopolysaccharide and Gut Microbiota in Inflammatory Bowel Diseases. Int J Mol Sci. 10 de junho de 2021, 22, 6242. [Google Scholar] [CrossRef] [PubMed]
- Bourgonje, A.R.; Andreu-Sánchez, S.; Vogl, T.; Hu, S.; Vila, A.V.; Gacesa, R.; et al. Phage-display immunoprecipitation sequencing of the antibody epitope repertoire in inflammatory bowel disease reveals distinct antibody signatures. Immunity. 13 de junho de 2023, 56, 1393–1409.e6. [Google Scholar] [CrossRef] [PubMed]
- Morgan, N.N.; Duck, L.W.; Wu, J.; Rujani, M.; Thomes, P.G.; Elson, C.O.; et al. Crohn's Disease Patients Uniquely Contain Inflammatory Responses to Flagellin in a CD4 Effector Memory Subset. Inflammatory Bowel Diseases. 1o de dezembro de 2022, 28, 1893–903. [Google Scholar] [CrossRef] [PubMed]
- Whelan, K.; Bancil, A.S.; Lindsay, J.O.; Chassaing, B. Ultra-processed foods and food additives in gut health and disease. Nat Rev Gastroenterol Hepatol. junho de 2024, 21, 406–27. [Google Scholar] [CrossRef] [PubMed]
- Bancil, A.S.; Sandall, A.M.; Rossi, M.; Chassaing, B.; Lindsay, J.O.; Whelan, K. Food Additive Emulsifiers and Their Impact on Gut Microbiome, Permeability, and Inflammation: Mechanistic Insights in Inflammatory Bowel Disease. Journal of Crohn’s and Colitis. 1o de junho de 2021, 15, 1068–79. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Sandall, A.; Smith, L.; Rossi, M.; Whelan, K. Food additive emulsifiers: a review of their role in foods, legislation and classifications, presence in food supply, dietary exposure, and safety assessment. Nutrition Reviews. 1o de junho de 2021, 79, 726–41. [Google Scholar] [CrossRef] [PubMed]
- Stolfi, C.; Pacifico, T.; Monteleone, G.; Laudisi, F. Impact of Western Diet and Ultra-Processed Food on the Intestinal Mucus Barrier. Biomedicines. 18 de julho de 2023, 11, 2015. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature. outubro de 2014, 514, 181–6. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.R.; Tomas, J.; Brenner, C.; Sansonetti, P.J. Impact of high-fat diet on the intestinal microbiota and small intestinal physiology before and after the onset of obesity. Biochimie. 1o de outubro de 2017, 141, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Laudisi, F.; Di Fusco, D.; Dinallo, V.; Stolfi, C.; Di Grazia, A.; Marafini, I.; et al. The Food Additive Maltodextrin Promotes Endoplasmic Reticulum Stress–Driven Mucus Depletion and Exacerbates Intestinal Inflammation. Cell Mol Gastroenterol Hepatol. 11 de setembro de 2018, 7, 457–73. [Google Scholar] [CrossRef] [PubMed]
- Pinget, G.; Tan, J.; Janac, B.; Kaakoush, N.O.; Angelatos, A.S.; O'Sullivan, J.; et al. Impact of the Food Additive Titanium Dioxide (E171) on Gut Microbiota-Host Interaction. Front Nutr. 14 de maio de 2019, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Chen, L.; Catalan-Dibene, J.; Bongers, G.; Faith, J.J.; Suebsuwong, C.; et al. Food colorants metabolized by commensal bacteria promote colitis in mice with dysregulated expression of interleukin-23. Cell Metab. 6 de julho de 2021, 33, 1358–1371.e5. [Google Scholar] [CrossRef] [PubMed]
- Santos FSdos Mintem, G.C.; Oliveira IOde Horta, B.L.; Ramos, E.; Lopes, C.; et al. Consumption of ultra-processed foods and IL-6 in two cohorts from high- and middle-income countries. British Journal of Nutrition. maio de 2023, 129, 1552–62. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Compher, C.; Bonhomme, B.; Liu, Q.; Tian, Y.; Walters, W.; et al. Randomized Controlled-Feeding Study of Dietary Emulsifier Carboxymethylcellulose Reveals Detrimental Impacts on the Gut Microbiota and Metabolome. Gastroenterology. 1o de março de 2022, 162, 743–56. [Google Scholar] [CrossRef] [PubMed]
- Sieg, H.; Schaar, C.; Fouquet, N.; Böhmert, L.; Thünemann, A.F.; Braeuning, A. Particulate iron oxide food colorants (E 172) during artificial digestion and their uptake and impact on intestinal cells. Toxicology in Vitro. 1o de abril de 2024, 96, 105772. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Guo, X.; Thanuphol, P.; Ji, R.; Zhu, Z.; Wu, Y.; et al. Gut Microbiota-Mediated Degradation of Food-Grade Lambda-Carrageenan by Bacteroides xylanisolvens and Its Role in Inflammation. J Agric Food Chem. 19 de fevereiro de 2025, 73, 4288–98. [Google Scholar] [CrossRef] [PubMed]
- Elmén, L.; Zlamal, J.E.; Scott, D.A.; Lee, R.B.; Chen, D.J.; Colas, A.R.; et al. Dietary Emulsifier Sodium Stearoyl Lactylate Alters Gut Microbiota in vitro and Inhibits Bacterial Butyrate Producers. Front Microbiol [Internet]. 15 de maio de 2020 [citado 9 de julho de 2025];11. Disponível em: https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2020.00892/full.
- Chassaing, B.; Koren, O.; Goodrich, J.; Poole, A.; Srinivasan, S.; Ley, R.E.; et al. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature. 5 de março de 2015, 519, 92–6. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.L.; Keita, Å.V.; Duncan, S.H.; O'Kennedy, N.; Söderholm, J.D.; Rhodes, J.M.; et al. Translocation of Crohn's disease Escherichia coli across M-cells: contrasting effects of soluble plant fibres and emulsifiers. Gut. outubro de 2010, 59, 1331–9. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Ung, V.; Sydora, B.C.; Loening-Baucke, V.; Doerffel, Y.; Verstraelen, H.; et al. Bacterial overgrowth and inflammation of small intestine after carboxymethylcellulose ingestion in genetically susceptible mice. Inflamm Bowel Dis. março de 2009, 15, 359–64. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Xu, X.; Qian, Z.; Cheng, H.; Shen, X.; Chen, S.; et al. Xanthan gum-assisted fabrication of stable emulsion-based oleogel structured with gelatin and proanthocyanidins. Food Hydrocolloids. 1o de junho de 2021, 115, 106596. [Google Scholar] [CrossRef]
- Thymann, T.; Møller, H.K.; Stoll, B.; Støy, A.C.F.; Buddington, R.K.; Bering, S.B.; et al. Carbohydrate maldigestion induces necrotizing enterocolitis in preterm pigs. Am J Physiol Gastrointest Liver Physiol. dezembro de 2009, 297, G1115–25. [Google Scholar] [CrossRef] [PubMed]
- Udo, T.; Mummaleti, G.; Mohan, A.; Singh, R.K.; Kong, F. Current and emerging applications of carrageenan in the food industry. Food Research International. 1o de novembro de 2023, 173, 113369. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Shani Levi, C.; Fahoum, L.; Ungar, Y.; Meyron-Holtz, E.G.; Shpigelman, A.; et al. Revisiting the carrageenan controversy: do we really understand the digestive fate and safety of carrageenan in our foods? Food Funct. 1o de março de 2018, 9, 1344–52. [Google Scholar] [CrossRef] [PubMed]
- Brito AKde, B.; Cardoso, K.G.M.; Soares, S.D.; Chisté, R.C. CORANTES ARTIFICIAIS PERMITIDOS NO BRASIL: PRINCIPAIS CARACTERÍSTICAS E EFEITOS TOXICOLÓGICOS. Em Editora Científica Digital; 2021 [citado 9 de julho de 2025]. p. 428–44. Disponível em: https://www.editoracientifica.com.br/artigos/corantes-artificiais-permitidos-no-brasil-principais-caracteristicas-e-efeitos-toxicologicos.
- Vojdani, A.; Vojdani, C. Immune reactivity to food coloring. Altern Ther Health Med. 2015, 21 Suppl 1, 52–62. [Google Scholar] [PubMed]
- Elder, R.; Vancuren, S.J.; Botschner, A.J.; Josephy, P.D.; Allen-Vercoe, E. Metabolism of azo food dyes by bacterial members of the human gut microbiome. Anaerobe. 1o de outubro de 2023, 83, 102783. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.; Jung, S. Food colors caught red-handed. Cell Metabolism. 6 de julho de 2021, 33, 1267–9. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, C.D. Desenvolvimento e caracterização de nanopartículas poliméricas para encapsulação de L-asparaginase /. UFRJ,; 2018.
- em 27/07/2023 15h33 P em 27/07/2023 12h49 A. Agência Nacional de Vigilância Sanitária - Anvisa. [citado 9 de julho de 2025]. Nota: avaliação do aditivo alimentar dióxido de titânio. Disponível em: https://www.gov.br/anvisa/pt-br/assuntos/noticias-anvisa/2023/nota-avaliacao-do-aditivo-alimentar-dioxido-de-titanio.
- Joint FAO/WHO Expert Committee on Food Additives risk assessment of titanium dioxide risk released – background information [Internet]. [citado 9 de julho de 2025]. Disponível em: https://www.who.int/publications/m/item/jecfa-risk-assessment-of-titanium-dioxide-risk-released-background-information.
- Bettini, S.; Boutet-Robinet, E.; Cartier, C.; Coméra, C.; Gaultier, E.; Dupuy, J.; et al. Food-grade TiO2 impairs intestinal and systemic immune homeostasis, initiates preneoplastic lesions and promotes aberrant crypt development in the rat colon. Sci Rep. 20 de janeiro de 2017, 7, 40373. [Google Scholar] [CrossRef] [PubMed]
- Proquin, H.; Rodríguez-Ibarra, C.; Moonen, C.G.J.; Urrutia Ortega, I.M.; Briedé, J.J.; de Kok, T.M.; et al. Titanium dioxide food additive (E171) induces ROS formation and genotoxicity: contribution of micro and nano-sized fractions. Mutagenesis. 1o de janeiro de 2017, 32, 139–49. [Google Scholar] [CrossRef] [PubMed]
- Quaglio, A.E.V.; Grillo, T.G.; Oliveira, E.C.S.D.; Stasi, L.C.D.; Sassaki, L.Y. Gut microbiota, inflammatory bowel disease and colorectal cancer. World Journal of Gastroenterology. 14 de agosto de 2022, 28, 4053–60. [Google Scholar] [CrossRef] [PubMed]
- Giambra, V.; Pagliari, D.; Rio, P.; Totti, B.; Di Nunzio, C.; Bosi, A.; et al. Gut Microbiota, Inflammatory Bowel Disease, and Cancer: The Role of Guardians of Innate Immunity. Cells. 19 de novembro de 2023, 12, 2654. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.T.; Amos, G.C.A.; Murphy, A.R.J.; Murch, S.; Wellington, E.M.H.; Arasaradnam, R.P. Microbial imbalance in inflammatory bowel disease patients at different taxonomic levels. Gut Pathog. 4 de janeiro de 2020, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; et al. Reduced Abundance of Butyrate-Producing Bacteria Species in the Fecal Microbial Community in Crohn's Disease. Digestion. 2016, 93, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Santana, P.T.; Rosas, S.L.B.; Ribeiro, B.E.; Marinho, Y.; de Souza, H.S.P. Dysbiosis in Inflammatory Bowel Disease: Pathogenic Role and Potential Therapeutic Targets. Int J Mol Sci. 23 de março de 2022, 23, 3464. [Google Scholar] [CrossRef] [PubMed]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; et al. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front Immunol. 11 de março de 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.; Han, S.; Kwon, J.; Ju, S.; Choi, T.G.; Kang, I.; et al. Roles of Short-Chain Fatty Acids in Inflammatory Bowel Disease. Nutrients. 21 de outubro de 2023, 15, 4466. [Google Scholar] [CrossRef] [PubMed]
- Hee Bvan der Wells, J.M. Microbial Regulation of Host Physiology by Short-chain Fatty Acids. Trends in Microbiology. 1o de agosto de 2021, 29, 700–12. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-Mediated Regulation of Skeletal Muscle Hypertrophy and Atrophy. Cells. 26 de agosto de 2020, 9, 1970. [Google Scholar] [CrossRef] [PubMed]
- De Preter, V.; Arijs, I.; Windey, K.; Vanhove, W.; Vermeire, S.; Schuit, F.; et al. Impaired butyrate oxidation in ulcerative colitis is due to decreased butyrate uptake and a defect in the oxidation pathway. Inflamm Bowel Dis. junho de 2012, 18, 1127–36. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Horn, P.; Wong, V.W.S.; Ratziu, V.; Bugianesi, E.; Francque, S.; et al. EASL–EASD–EASO Clinical Practice Guidelines on the management of metabolic dysfunction-associated steatotic liver disease (MASLD). Journal of Hepatology. 1o de setembro de 2024, 81, 492–542. [Google Scholar] [CrossRef] [PubMed]
- Rubino, F.; Cummings, D.E.; Eckel, R.H.; Cohen, R.V.; Wilding, J.P.H.; Brown, W.A.; et al. Definition and diagnostic criteria of clinical obesity. The Lancet Diabetes & Endocrinology. 1o de março de 2025, 13, 221–62. [Google Scholar]
- Ross, F.C.; Patangia, D.; Grimaud, G.; Lavelle, A.; Dempsey, E.M.; Ross, R.P.; et al. The interplay between diet and the gut microbiome: implications for health and disease. Nat Rev Microbiol. novembro de 2024, 22, 671–86. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Van Hul, M. Gut microbiota in overweight and obesity: crosstalk with adipose tissue. Nat Rev Gastroenterol Hepatol. março de 2024, 21, 164–83. [Google Scholar] [CrossRef] [PubMed]
- Nesci, A.; Carnuccio, C.; Ruggieri, V.; D’Alessandro, A.; Di Giorgio, A.; Santoro, L.; et al. Gut Microbiota and Cardiovascular Disease: Evidence on the Metabolic and Inflammatory Background of a Complex Relationship. Int J Mol Sci. 22 de maio de 2023, 24, 9087. [Google Scholar] [CrossRef] [PubMed]
- Góralczyk-Bińkowska, A.; Szmajda-Krygier, D.; Kozłowska, E. The Microbiota–Gut–Brain Axis in Psychiatric Disorders. Int J Mol Sci. 24 de setembro de 2022, 23, 11245. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.A.; Inniss, S.; Kumagai, T.; Rahman, F.Z.; Smith, A.M. The Role of Diet and Gut Microbiota in Regulating Gastrointestinal and Inflammatory Disease. Front Immunol. 5 de abril de 2022, 13, 866059. [Google Scholar] [CrossRef] [PubMed]
- Ferenc, K.; Sokal-Dembowska, A.; Helma, K.; Motyka, E.; Jarmakiewicz-Czaja, S.; Filip, R. Modulation of the Gut Microbiota by Nutrition and Its Relationship to Epigenetics. Int J Mol Sci. 19 de janeiro de 2024, 25, 1228. [Google Scholar] [CrossRef] [PubMed]
- Merra, G.; Noce, A.; Marrone, G.; Cintoni, M.; Tarsitano, M.G.; Capacci, A.; et al. Influence of Mediterranean Diet on Human Gut Microbiota. Nutrients. 22 de dezembro de 2020, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- García-Montero, C.; Fraile-Martínez, O.; Gómez-Lahoz, A.M.; Pekarek, L.; Castellanos, A.J.; Noguerales-Fraguas, F.; et al. Nutritional Components in Western Diet Versus Mediterranean Diet at the Gut Microbiota–Immune System Interplay. Implications for Health and Disease. Nutrients. fevereiro de 2021, 13, 699. [Google Scholar] [CrossRef] [PubMed]
- García-Gavilán, J.F.; Atzeni, A.; Babio, N.; Liang, L.; Belzer, C.; Vioque, J.; et al. Effect of 1-year lifestyle intervention with energy-reduced Mediterranean diet and physical activity promotion on the gut metabolome and microbiota: a randomized clinical trial. The American Journal of Clinical Nutrition. 1o de maio de 2024, 119, 1143–54. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; Laiola, M.; Roager, H.M.; De Filippis, F.; Roume, H.; Quinquis, B.; et al. Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut. julho de 2020, 69, 1258–68. [Google Scholar] [CrossRef] [PubMed]
- Haskey, N.; Estaki, M.; Ye, J.; Shim, R.K.; Singh, S.; Dieleman, L.A.; et al. A Mediterranean Diet Pattern Improves Intestinal Inflammation Concomitant with Reshaping of the Bacteriome in Ulcerative Colitis: A Randomised Controlled Trial. J Crohns Colitis. 24 de abril de 2023, 17, 1569–78. [Google Scholar] [CrossRef] [PubMed]
- Godny, L.; Elial-Fatal, S.; Arrouasse, J.; Fischler, T.S.; Reshef, L.; Kutukov, Y.; et al. Mechanistic Implications of the Mediterranean Diet in Patients With Newly Diagnosed Crohn's Disease: Multiomic Results From a Prospective Cohort. Gastroenterology. 1o de maio de 2025, 168, 952–964.e2. [Google Scholar] [CrossRef] [PubMed]
- Hashash, J.G.; Elkins, J.; Lewis, J.D.; Binion, D.G. AGA Clinical Practice Update on Diet and Nutritional Therapies in Patients With Inflammatory Bowel Disease: Expert Review. Gastroenterology. 1o de março de 2024, 166, 521–32. [Google Scholar] [CrossRef] [PubMed]
- Erol Doğan, Ö.; Karaca Çelik, K.E.; Baş, M.; Alan, E.H.; Çağın, Y.F. Effects of Mediterranean Diet, Curcumin, and Resveratrol on Mild-to-Moderate Active Ulcerative Colitis: A Multicenter Randomized Clinical Trial. Nutrients. 16 de maio de 2024, 16, 1504. [Google Scholar] [CrossRef] [PubMed]
- Chicco, F.; Magrì, S.; Cingolani, A.; Paduano, D.; Pesenti, M.; Zara, F.; et al. Multidimensional Impact of Mediterranean Diet on IBD Patients. Inflammatory Bowel Diseases. 1o de janeiro de 2021, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Randeni, N.; Bordiga, M.; Xu, B. A Comprehensive Review of the Triangular Relationship among Diet–Gut Microbiota–Inflammation. Int J Mol Sci. 29 de agosto de 2024, 25, 9366. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol. agosto de 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes. 6 de fevereiro de 2017, 8, 172–84. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Escher, J.; Hébuterne, X.; Kłęk, S.; Krznaric, Z.; Schneider, S.; et al. ESPEN practical guideline: Clinical Nutrition in inflammatory bowel disease. Clinical Nutrition. 1o de março de 2020, 39, 632–53. [Google Scholar] [CrossRef] [PubMed]
- Sinopoulou, V.; Gordon, M.; Gregory, V.; Saadeh, A.; Akobeng, A.K. Prebiotics for induction and maintenance of remission in ulcerative colitis - Sinopoulou, V - 2024 | Cochrane Library. [citado 9 de julho de 2025]; Disponível em: https://www.cochranelibrary.com/cdsr/doi/10.1002/14651858.CD015084.pub2/full.
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. agosto de 2014, 11, 506–14. [Google Scholar] [CrossRef] [PubMed]
- Iheozor-Ejiofor, Z.; Kaur, L.; Gordon, M.; Baines, P.A.; Sinopoulou, V.; Akobeng, A.K. Probiotics for maintenance of remission in ulcerative colitis - Iheozor-Ejiofor, Z - 2020 | Cochrane Library. [citado 9 de julho de 2025]; Disponível em: https://www.cochranelibrary.com/cdsr/doi/10.1002/14651858.CD007443.pub3/full.
- Bischoff, S.C.; Bager, P.; Escher, J.; Forbes, A.; Hébuterne, X.; Hvas, C.L.; et al. ESPEN guideline on Clinical Nutrition in inflammatory bowel disease. Clinical Nutrition. março de 2023, 42, 352–79. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat Rev Gastroenterol Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Rufino, M.N.; da Costa, A.L.; Jorge, E.N.; Paiano, V.F.; Camparoto, M.L.; Keller, R.; et al. Synbiotics improve clinical indicators of ulcerative colitis: systematic review with meta-analysis. Nutrition Reviews. 1o de fevereiro de 2022, 80, 157–64. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Dhaneshwar, S. Role of prebiotics, probiotics, and synbiotics in management of inflammatory bowel disease: Current perspectives. World Journal of Gastroenterology. 14 de abril de 2023, 29, 2078–100. [Google Scholar] [CrossRef] [PubMed]
- Lê, A.; Mantel, M.; Marchix, J.; Bodinier, M.; Jan, G.; Rolli-Derkinderen, M. Inflammatory bowel disease therapeutic strategies by modulation of the microbiota: how and when to introduce pre-, pro-, syn-, or postbiotics? American Journal of Physiology-Gastrointestinal and Liver Physiology. dezembro de 2022, 323, G523–53. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol. 2021, 18, 649–67. [Google Scholar] [CrossRef] [PubMed]
- Kavita Om, H.; Chand, U.; Kushawaha, P.K. Postbiotics: An alternative and innovative intervention for the therapy of inflammatory bowel disease. Microbiological Research. 1o de fevereiro de 2024, 279, 127550. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, P.; Wang, D.; Shen, S.; Wang, S.; Li, Y.; et al. Postbiotics in inflammatory bowel disease: efficacy, mechanism, and therapeutic implications. Journal of the Science of Food and Agriculture. 2025, 105, 721–34. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, G.; Ianiro, G.; Tilg, H.; Rajilić-Stojanović, M.; Kump, P.; Satokari, R.; et al. European consensus conference on faecal microbiota transplantation in clinical practice. Gut. abril de 2017, 66, 569–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wellens, J.; Kalla, R.; Fu, T.; Deng, M.; Zhang, H.; et al. Intake of Ultra-processed Foods Is Associated with an Increased Risk of Crohn's Disease: A Cross-sectional and Prospective Analysis of 187 154 Participants in the UK Biobank. J Crohns Colitis. 28 de outubro de 2022, 17, 535–52. [Google Scholar] [CrossRef] [PubMed]
- Peery, A.F.; Kelly, C.R.; Kao, D.; Vaughn, B.P.; Lebwohl, B.; Singh, S.; et al. AGA Clinical Practice Guideline on Fecal Microbiota–Based Therapies for Select Gastrointestinal Diseases. Gastroenterology. 1o de março de 2024, 166, 409–34. [Google Scholar] [CrossRef] [PubMed]
- Halkjær, S.I.; Lo, B.; Cold, F.; Højer Christensen, A.; Holster, S.; König, J.; et al. Fecal microbiota transplantation for the treatment of irritable bowel syndrome: A systematic review and meta-analysis. World J Gastroenterol. 28 de maio de 2023, 29, 3185–202. [Google Scholar] [CrossRef] [PubMed]
- Imdad, A.; Pandit, N.G.; Zaman, M.; Minkoff, N.Z.; Tanner-Smith, E.E.; Gomez-Duarte, O.G.; et al. Fecal transplantation for treatment of inflammatory bowel disease - Imdad, A - 2023 | Cochrane Library. [citado 9 de julho de 2025]; Disponível em: https://www.cochranelibrary.com/cdsr/doi/10.1002/14651858.CD012774.pub3/full.
- Yan, J.; Wang, L.; Gu, Y.; Hou, H.; Liu, T.; Ding, Y.; et al. Dietary Patterns and Gut Microbiota Changes in Inflammatory Bowel Disease: Current Insights and Future Challenges. Nutrients. 27 de setembro de 2022, 14, 4003. [Google Scholar] [CrossRef] [PubMed]
- Juul, F.; Vaidean, G.; Parekh, N. Ultra-processed Foods and Cardiovascular Diseases: Potential Mechanisms of Action. Advances in Nutrition. 1o de setembro de 2021, 12, 1673–80. [Google Scholar] [CrossRef] [PubMed]
- Rondinella, D.; Raoul, P.C.; Valeriani, E.; Venturini, I.; Cintoni, M.; Severino, A.; et al. The Detrimental Impact of Ultra-Processed Foods on the Human Gut Microbiome and Gut Barrier. Nutrients. 28 de fevereiro de 2025, 17, 859. [Google Scholar] [CrossRef]
- Vissers, E.; Wellens, J.; Sabino, J. Ultra-processed foods as a possible culprit for the rising prevalence of inflammatory bowel diseases. Front Med (Lausanne). 7 de novembro de 2022, 9, 1058373. [Google Scholar] [CrossRef] [PubMed]
- Quetglas-Llabrés, M.M.; Monserrat-Mesquida, M.; Bouzas, C.; Mateos, D.; Ugarriza, L.; Gómez, C.; et al. Oxidative Stress and Inflammatory Biomarkers Are Related to High Intake of Ultra-Processed Food in Old Adults with Metabolic Syndrome. Antioxidants (Basel). 31 de julho de 2023, 12, 1532. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, G.; Lou, P.; Zhang, M.; Yao, K.; Xiao, J.; et al. Excessive nucleic acid R-loops induce mitochondria-dependent epithelial cell necroptosis and drive spontaneous intestinal inflammation. Proc Natl Acad Sci U S A. 121, e2307395120.
- Dang, P.M.C.; Rolas, L.; El-Benna, J. The Dual Role of Reactive Oxygen Species-Generating Nicotinamide Adenine Dinucleotide Phosphate Oxidases in Gastrointestinal Inflammation and Therapeutic Perspectives. Antioxidants & Redox Signaling. 10 de agosto de 2020, 33, 354–73. [Google Scholar]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; et al. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens. 13 de agosto de 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Tristan Asensi, M.; Napoletano, A.; Sofi, F.; Dinu, M. Low-Grade Inflammation and Ultra-Processed Foods Consumption: A Review. Nutrients. 22 de março de 2023, 15, 1546. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Song, R.; Liu, Y.; Wu, Z.; Zhang, X. Effects of ultra-processed foods on the microbiota-gut-brain axis: The bread-and-butter issue. Food Research International. 1o de maio de 2023, 167, 112730. [Google Scholar] [CrossRef] [PubMed]
- Narula, N.; Wong, E.C.L.; Dehghan, M.; Mente, A.; Rangarajan, S.; Lanas, F.; et al. Association of ultra-processed food intake with risk of inflammatory bowel disease: prospective cohort study. BMJ. 15 de julho de 2021, 374, n1554. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Dong, C.; Casagrande, C.; Chan, S.S.M.; Huybrechts, I.; Nicolas, G.; et al. Food Processing and Risk of Crohn's Disease and Ulcerative Colitis: A European Prospective Cohort Study. Clinical Gastroenterology and Hepatology. 1o de junho de 2023, 21, 1607–1616.e6. [Google Scholar] [CrossRef] [PubMed]
- Preda, C.M.; Istratescu, D.; Nitescu, M.; Manuc, T.; Manuc, M.; Stroie, T.; et al. Diet Optimization in Inflammatory Bowel Disease: Impact on Disease Relapse and Inflammatory Markers. A 1-year Prospective Trial. Journal of Gastrointestinal and Liver Diseases. 29 de junho de 2024, 33, 184–93. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, Y.; Li, F.; Zhang, D. Dietary fiber intake reduces risk of inflammatory bowel disease: result from a meta-analysis. Nutrition Research. 1o de setembro de 2015, 35, 753–8. [Google Scholar] [CrossRef] [PubMed]
- Sigall Boneh, R.; Westoby, C.; Oseran, I.; Sarbagili-Shabat, C.; Albenberg, L.G.; Lionetti, P.; et al. The Crohn's Disease Exclusion Diet: A Comprehensive Review of Evidence, Implementation Strategies, Practical Guidance, and Future Directions. Inflamm Bowel Dis. 18 de novembro de 2023, 30, 1888–902. [Google Scholar] [CrossRef] [PubMed]
- Strisciuglio, C.; Cenni, S.; Serra, M.R.; Dolce, P.; Martinelli, M.; Staiano, A.; et al. Effectiveness of Mediterranean Diet's Adherence in Children with Inflammatory Bowel Diseases. Nutrients. 20 de outubro de 2020, 12, 3206. [Google Scholar] [CrossRef] [PubMed]
- Martín-Masot, R.; Herrador-López, M.; Navas-López, V.M. Dietary Habit Modifications in Paediatric Patients after One Year of Treatment with the Crohn's Disease Exclusion Diet. Nutrients. 20 de janeiro de 2023, 15, 554. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
NOVA classification of foods. [11].
Figure 1.
NOVA classification of foods. [11].
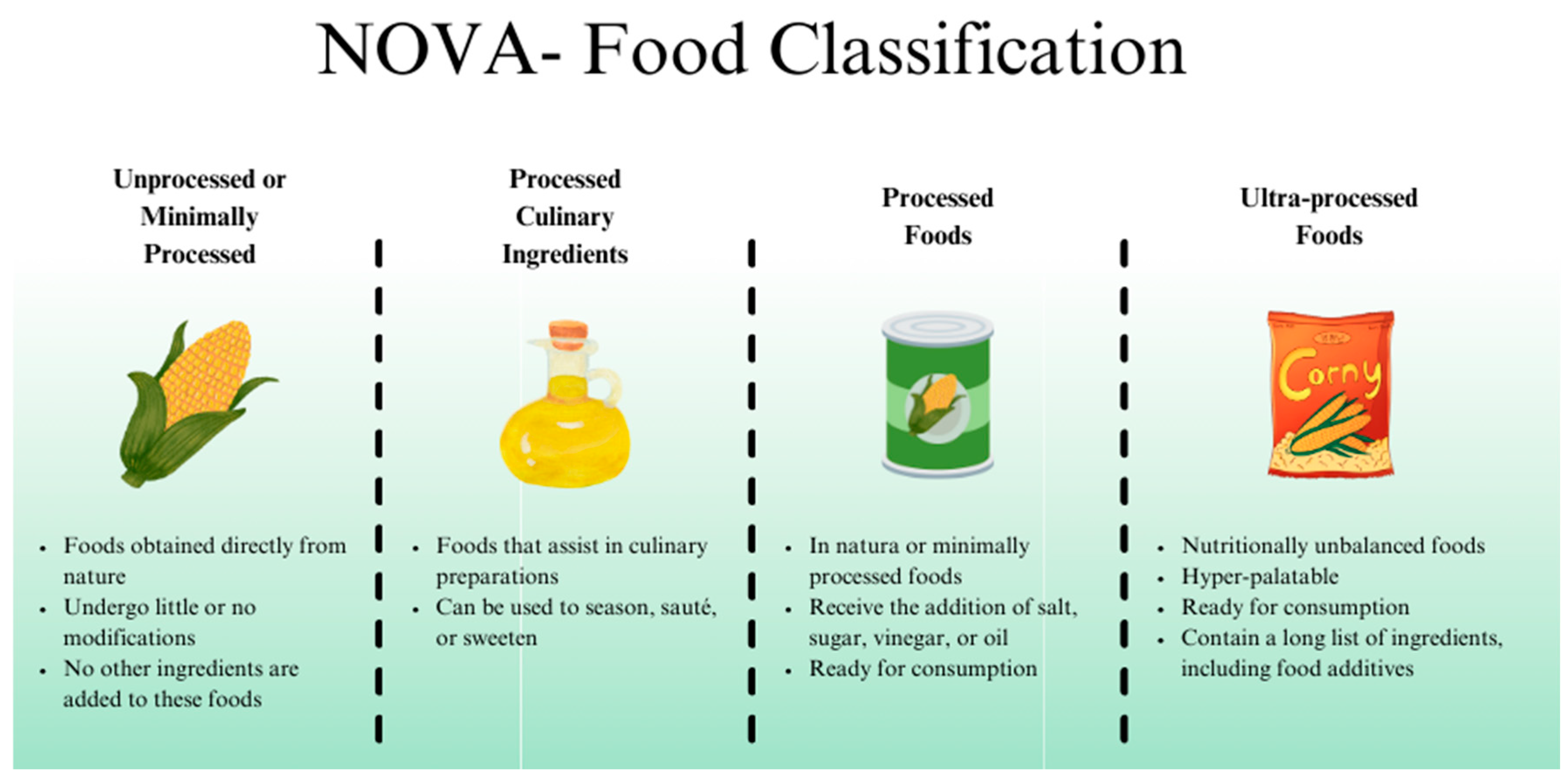
Figure 2.
The consumption of whole and/or minimally processed foods is associated with the promotion of an eubiotic environment characterized by increased production of short-chain fatty acids (SCFAs) and mucin, reduced intestinal permeability, lower levels of opportunistic bacteria and inflammatory factors, and elevated anti-inflammatory markers. In contrast, consuming ultra-processed foods contributes to dysbiosis, characterized by an increase in pathogenic bacteria, pro-inflammatory mediators, and metabolic disturbances. LPS: Lipopolysaccharides; NF-kB: Nuclear Factor Kappa B; IL-6: Interleukin-6; IL-10: Interleukin-10; IL-18: Interleukin-18; SFA: Saturated fatty acids; ꞷ-3: omega-3 fatty acids.
Figure 2.
The consumption of whole and/or minimally processed foods is associated with the promotion of an eubiotic environment characterized by increased production of short-chain fatty acids (SCFAs) and mucin, reduced intestinal permeability, lower levels of opportunistic bacteria and inflammatory factors, and elevated anti-inflammatory markers. In contrast, consuming ultra-processed foods contributes to dysbiosis, characterized by an increase in pathogenic bacteria, pro-inflammatory mediators, and metabolic disturbances. LPS: Lipopolysaccharides; NF-kB: Nuclear Factor Kappa B; IL-6: Interleukin-6; IL-10: Interleukin-10; IL-18: Interleukin-18; SFA: Saturated fatty acids; ꞷ-3: omega-3 fatty acids.
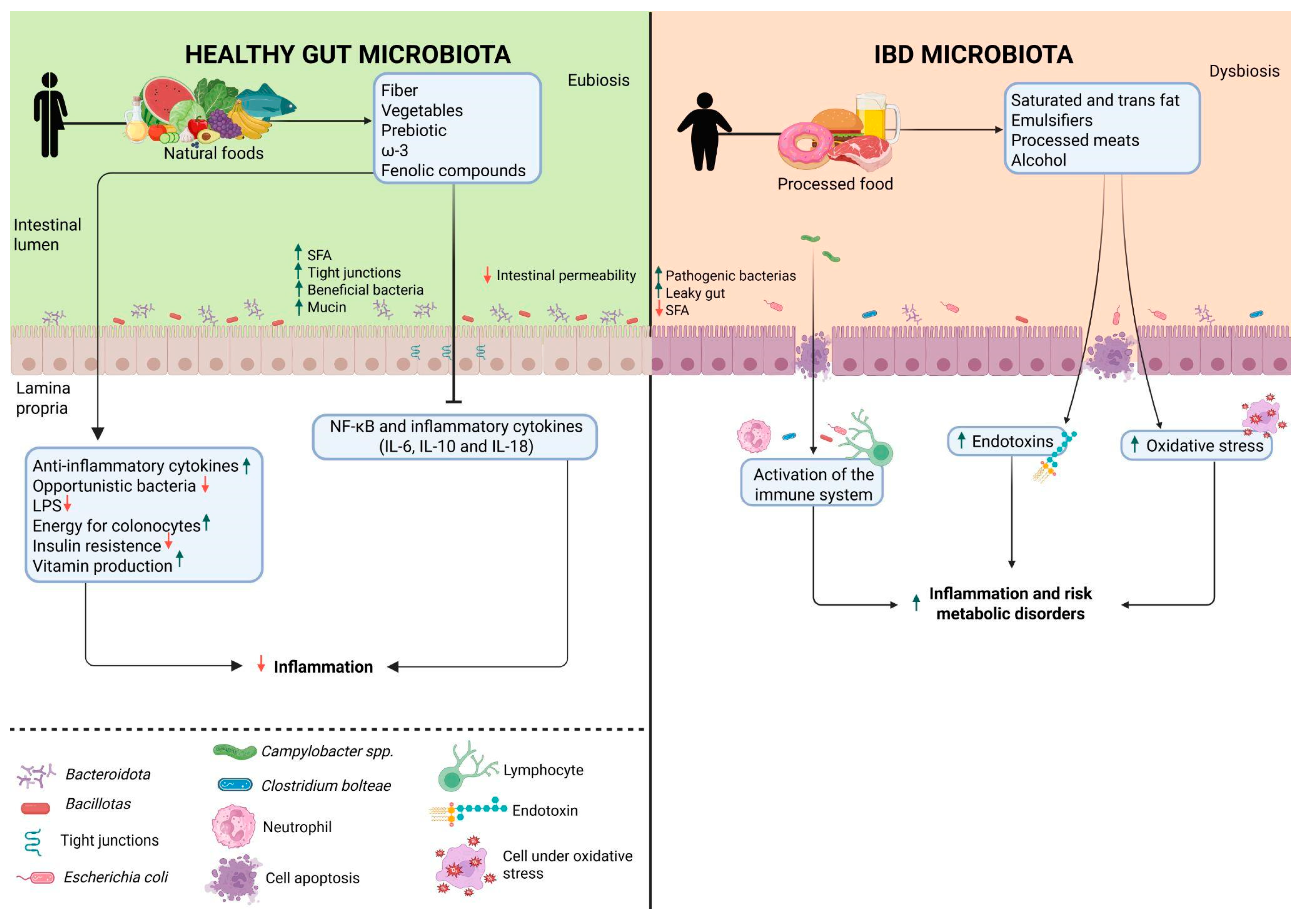
Figure 3.
The Western diet's impact on promoting dysbiosis, increasing intestinal permeability, and initiating the inflammatory cascade is a contributing risk factor for developing inflammatory bowel diseases (IBD). IL10: Interleukin-10; IL4: Interleukin-4; IL13: Interleukin-13; TGF-β: Transforming Growth Factor beta; IL6: Interleukin-6; IL1: Interleukin-1; IFN-γ: interferon gama; TNF-α: Fator de Necrose Tumoral alfa.
Figure 3.
The Western diet's impact on promoting dysbiosis, increasing intestinal permeability, and initiating the inflammatory cascade is a contributing risk factor for developing inflammatory bowel diseases (IBD). IL10: Interleukin-10; IL4: Interleukin-4; IL13: Interleukin-13; TGF-β: Transforming Growth Factor beta; IL6: Interleukin-6; IL1: Interleukin-1; IFN-γ: interferon gama; TNF-α: Fator de Necrose Tumoral alfa.
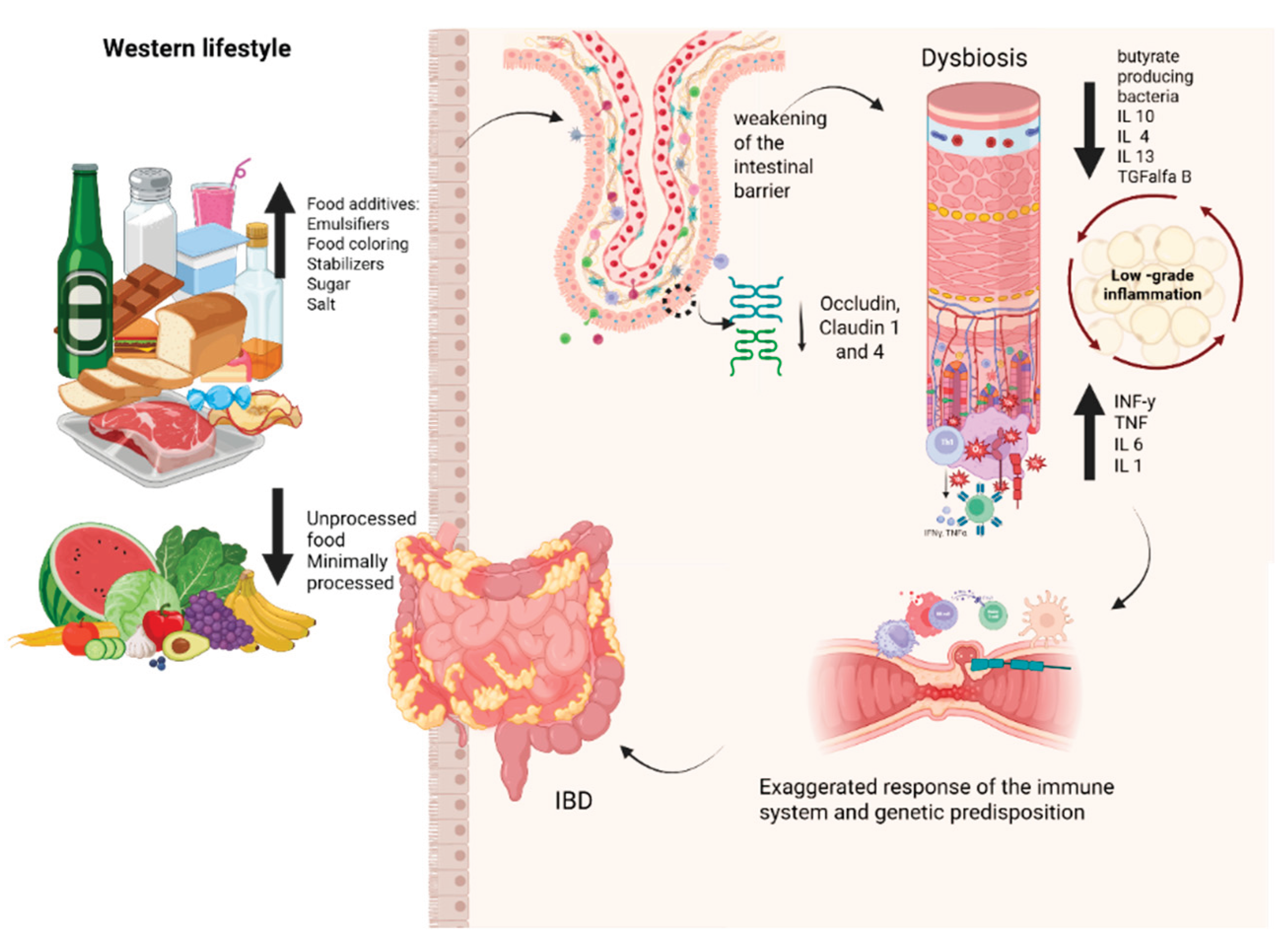

Table 1.
Summary of studies on food additives and the risk of gut disease.
| Study (year) | Model | Additive(s) | Key Findings Related to Gut Microbiota/Health |
|---|---|---|---|
| Suez et al., 2014 [30] | Germ-free mice | Non-caloric artificial sweeteners (NAS) | Excessive consumption may promote glucose intolerance, dysbiosis, and metabolic alteration. |
| Araújo, 2017 [31] | Humans, ages 18-60 years | Carboxymethylcellulose (CMC) | Increased bacterial proliferation and infiltration, with an increase in Roseburia spp. and Lachnospiraceae bacterium species. |
| Laudisi, 2018 [32] | Mice | Maltodextrin | Decreased Muc-2 results in greater adhesion of pathogenic bacteria. |
| Pinget et al., 2019 [33] | Mice | Titanium Dioxide (TiO₂) | TiO₂ may impair intestinal homeostasis, increase inflammatory cytokine expression, and decrease crypt length. |
| He et al., 2021 [34] | Mice | Colorants Red 40 and Yellow 6 | It can intensify intestinal inflammation and induce colitis. |
| Silva, 2022 [35] | Wistar rats | Xanthan gum | Continuous consumption increases pro-inflammatory cytokines (TNF-α, IL-6, and IL-10) and alters intestinal barrier integrity. |
| Chassaing et al., 2022 [36] | Humans, 16 adults | Carboxymethylcellulose (CMC) | Alteration in gut microbiota composition and reduction of metabolites like SCFAs. |
| Sieg et al., 2024 [37] | In vitro | Iron oxide food colorants (E 172) | E 172 showed strong interaction with intestinal cells, though no toxic effects were observed. |
| Han et al., 2025 [38] | In vitro | Carrageenan | Degraded carrageenan generates pro-inflammatory cytokines, such as IL1-b and TNF-a, which are related to the development of IBD. |
Table 2.
Main Phyla affected patients with Inflammatory Bowel Disease.
| Phylum | Description | Reference |
|---|---|---|
| Bacteroidota and Bacillotas | It comprises 90% of the gut microbiota and is often reduced, potentially impairing the inflammatory response and short-chain fatty acid production. | Giambra et al. [57]; Santana et al. [60] |
| Proteobacteria | It typically increases, including opportunistic pathogens, such as Enterobacteriaceae and Burkholderiaceae, that can exacerbate inflammation. | Alam et al. [58] |
| Actinobacteria | In patients with Crohn's disease, they are increase, which influences dysbiosis and intestinal inflammation. | Takahashi et al. [59] |
Table 3.
Altered bacterial profiles in Inflammatory Bowel Disease contribute to inflammation and exacerbation.[60,61,62,63].
| Increased in IBD | Decrease in IBD | ||
|---|---|---|---|
| Phylum | Species | Phylum | Species |
|
Proteobacteria |
E. coli Campylobacter spp. H. parainfluenzae E. corrodens |
Verrucomicrobia | A. muciniphila |
|
Bacteroidota |
B. fragilis |
Bacillota |
F. prausnitzii R. albus Eubacterium spp |
| Bacillota |
R. torques Ruminococcus spp. C. hathewayi C. bolteae R. gnavus |
||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



